
Novel Synthetic Analogs of Sialyl Lewis X Can Inhibit Angiogenesisin Vitroandin Vivo
8
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 228, 716–723 (1996) ARTICLE NO. 1722 Novel Synthetic Analogs of Sialyl Lewis X Can Inhibit Angiogenesis in Vitro and in Vivo Mai Nguyen, M.D.,* Frederick R. Eilber, M.D.,* and Shawn Defrees, Ph.D.² *UCLA School of Medicine and ² Cytel Corporation, Los Angeles, California 90095 Received October 7, 1996 Selectins have been shown to play an important role in angiogenesis. We have set out to synthesize multiple analogs of the selectin ligand sialyl Lewis X. We show here that analogs of sialyl Lewis X at concentrations of 50 mg/ml or greater significantly inhibited angiogenesis in in vitro assays of endothelial cell migration and proliferation (p õ 0.05). These analogs also exerted significant inhibitory effects in in vivo angiogenesis assays using the chick chorioallantoic membrane at doses of 2 mg/chick or greater (p õ 0.05). In contrast, the related carbohydrate Lewis X did not inhibit angiogenesis in both in vitro and in vivo assays. These data indicate that angiogenesis can be inhibited specifically by synthetic analogs of sialyl Lewis X. These analogs may potentially be useful in the treatment of angiogenesis-related diseases. q 1996 Academic Press, Inc. Angiogenesis, the formation of new capillaries, is critical in many normal conditions as well as in several disease states (1). Angiogenesis is a complex multistep process which includes endothelial cell migration, proliferation and differentiation into tube-like structures. These steps involve multiple growth factors, proteases and adhesion molecules among endothe- lial cells as well as those with other supporting cells. Recently, the selectins have been suggested to be important candidates in the process of angiogenesis (2). We have found that bovine E- selectin is expressed constitutively at a low level in bovine capillary endothelial cells, and that this expression is increased upon capillary tube formation (3). Sialylated and fucosylated N- linked oligosaccharides, potential ligands for E-selectin, were also found to be preferentially synthesized in bovine capillary endothelial cells undergoing tube formation (4). Koch et al. has recently shown that soluble E-selectin is angiogenic (5). Kraling et al. demonstrated that normal newborn foreskin and placenta stained positive for E-selectin whereas adult skin did not, suggesting that E-selectin is preferentially expressed in angiogenic and/or proliferating tissues (6). P-selectin, another member of the selectin family, may also participate in the process of angiogenesis (7, 8). The role of P-selectin, however, is likely to be less important than that of E-selectin, since we have found that the expression of bovine P-selectin is constitutive in endothelial cells and does not change with capillary tube formation (3). If the interaction between the selectins and their ligands is indeed important in angiogenesis, compounds directed at disrupting this process may be useful in the treatment of angiogenesis- related diseases. We therefore proceeded to synthesize multiple analogs of sialyl Lewis X, a known ligand for the selectins, and to test whether these synthetic sialyl Lewis X analogs can inhibit angiogenesis in vitro and in vivo. MATERIALS AND METHODS Cell lines and reagents. Human umbilical vein endothelial cells (HUVEC) were purchased from Clonetics (San Diego, CA). Human aortic endothelial cells (HAEC) were obtained from Dr. Judith Berliner (Los Angeles, CA). The cells were plated on tissue culture flasks coated with 1.5% gelatin (Difco, Detroit, MI) in phosphate buffered saline (PBS, Mediatech, Washington DC). They were maintained in endothelial growth media which consists of media 199 0006-291X/96 $18.00 Copyright q 1996 by Academic Press, Inc. All rights of reproduction in any form reserved. 716
-
Upload
mai-nguyen -
Category
Documents
-
view
213 -
download
1
Transcript of Novel Synthetic Analogs of Sialyl Lewis X Can Inhibit Angiogenesisin Vitroandin Vivo

BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 228, 716–723 (1996)ARTICLE NO. 1722
Novel Synthetic Analogs of Sialyl Lewis X Can InhibitAngiogenesis in Vitro and in Vivo
Mai Nguyen, M.D.,* Frederick R. Eilber, M.D.,* and Shawn Defrees, Ph.D.†
*UCLA School of Medicine and †Cytel Corporation, Los Angeles, California 90095
Received October 7, 1996
Selectins have been shown to play an important role in angiogenesis. We have set out to synthesizemultiple analogs of the selectin ligand sialyl Lewis X. We show here that analogs of sialyl Lewis X atconcentrations of 50 mg/ml or greater significantly inhibited angiogenesis in in vitro assays of endothelialcell migration and proliferation (p õ 0.05). These analogs also exerted significant inhibitory effects in invivo angiogenesis assays using the chick chorioallantoic membrane at doses of 2 mg/chick or greater (p õ0.05). In contrast, the related carbohydrate Lewis X did not inhibit angiogenesis in both in vitro and invivo assays. These data indicate that angiogenesis can be inhibited specifically by synthetic analogs ofsialyl Lewis X. These analogs may potentially be useful in the treatment of angiogenesis-related diseases.q 1996 Academic Press, Inc.
Angiogenesis, the formation of new capillaries, is critical in many normal conditions aswell as in several disease states (1). Angiogenesis is a complex multistep process whichincludes endothelial cell migration, proliferation and differentiation into tube-like structures.These steps involve multiple growth factors, proteases and adhesion molecules among endothe-lial cells as well as those with other supporting cells. Recently, the selectins have been suggestedto be important candidates in the process of angiogenesis (2). We have found that bovine E-selectin is expressed constitutively at a low level in bovine capillary endothelial cells, and thatthis expression is increased upon capillary tube formation (3). Sialylated and fucosylated N-linked oligosaccharides, potential ligands for E-selectin, were also found to be preferentiallysynthesized in bovine capillary endothelial cells undergoing tube formation (4). Koch et al.has recently shown that soluble E-selectin is angiogenic (5). Kraling et al. demonstrated thatnormal newborn foreskin and placenta stained positive for E-selectin whereas adult skin didnot, suggesting that E-selectin is preferentially expressed in angiogenic and/or proliferatingtissues (6). P-selectin, another member of the selectin family, may also participate in theprocess of angiogenesis (7, 8). The role of P-selectin, however, is likely to be less important thanthat of E-selectin, since we have found that the expression of bovine P-selectin is constitutive inendothelial cells and does not change with capillary tube formation (3).
If the interaction between the selectins and their ligands is indeed important in angiogenesis,compounds directed at disrupting this process may be useful in the treatment of angiogenesis-related diseases. We therefore proceeded to synthesize multiple analogs of sialyl Lewis X, aknown ligand for the selectins, and to test whether these synthetic sialyl Lewis X analogs caninhibit angiogenesis in vitro and in vivo.
MATERIALS AND METHODS
Cell lines and reagents. Human umbilical vein endothelial cells (HUVEC) were purchased from Clonetics (SanDiego, CA). Human aortic endothelial cells (HAEC) were obtained from Dr. Judith Berliner (Los Angeles, CA). Thecells were plated on tissue culture flasks coated with 1.5% gelatin (Difco, Detroit, MI) in phosphate buffered saline(PBS, Mediatech, Washington DC). They were maintained in endothelial growth media which consists of media 199
0006-291X/96 $18.00Copyright q 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.
716
AID BBRC 5654 / 6911$$$341 11-05-96 08:00:18 bbrcas AP: BBRC

Vol. 228, No. 3, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
(Mediatech) with 10% heat-inactivated FCS (fetal calf serum, Gemini, Calabasas, CA), 1% antibiotic-antimycotic(Life Technologies, Grand Island, NY) and 25 mg/ml endothelial mitogen (Biomedical Technologies, Stoughton, MA).
A carbohydrate analog of sialyl Lewis X (CY-1503) and a non-carbohydrate analog of sialyl Lewis X (CY-31214)and the Lewis X carbohydrates were prepared at Cytel Corporation (San Diego, CA) by combined chemical andenzymatic synthesis (9, 10). ELISA assays were carried out according to the previously described procedure (11).Briefly, human soluble recombinant E-selectin or P-selectin (Cytel, San Diego, CA) was coated on plates, followedby addition of HL-60 cells (American Type Culture Collection, Rockville, MD) and analogs or carbohydrates. Afterincubation, the plates were rinsed and the adhesion determined by the cell lysis and myeloperoxidase method. IC 50
was the concentration that inhibited cell adhesion by 50%. This method gave consistent results with 10% deviation.The murine monoclonal antibodies to sialyl Lewis X (CSLEX-1) and to Lewis X (CLEX-1) were purchased from theUCLA Tissue Typing Laboratory. Basic fibroblast growth factor (bFGF) and vascular endothelial growth factor(VEGF) were obtained from R&D (Minneapolis, MN).
In vitro assays of angiogenesis. Both HUVECs and HAECs were used in migration assays. Migration was measuredwith the use of a 48-well chemotaxis chamber and nucleopore membrane filters with 8 um pores (Neuro Probe, CabinJohn, MD) coated with 6.7 mg/ml fibronectin (Chemicon, Temecula, CA). Either 10 ng/ml of the chemoattractantbFGF or 20 ng/ml of VEGF was added to the lower wells. The upper wells received 20,000 cells/well suspended inplain DMEM (Mediatech) alone. Test samples were added to the upper wells. The chamber was incubated at 377 Cin 5% CO2 for 4-5 hours. The cells that have migrated through the membrane onto the lower surface of the membranefilter were fixed in methanol and stained with haematoxylin. The number of cells within a rectangular grid werecounted at 4001 power with a microscope (Zeiss, Germany).
In proliferative assays, the cells were plated onto gelatinized 48-well culture plates at 10,000 cells/well and incubatedat 377 C in 5% CO2 for 24 hours in DMEM with 10% FCS. The media was then replaced with endothelial growthmedia and the test sample applied. In some assays, the media was replaced with media 199 containing either 10ng/ml bFGF or 20 ng/ml VEGF. After 72 hours, the cells were trypsinized and counted with a Coulter counter (CoulterElectronics, Hialeah, FL). All of the in vitro assays were performed in duplicates or triplicates. Statistical analysis ofin vitro assays was performed with the student’s t-test.
In vivo assay of angiogenesis. The chick chorioallantoic membrane (CAM) was used. Briefly, fertilized chickeneggs (McIntyre Poultry, Lakeside, CA) were cracked on day 3 and the embryos were individually placed into petridishes inside a tissue culture incubator at 377 C in 2% CO2. The test reagent was mixed with 0.5% methycellulose(Fisher, Fair Lawn, NJ) in order to form a clear pellet, which was then placed on top of the chorioallantoic membraneon day 6. Inhibition was observed as an avascular zone surrounding the test pellet on day 8.
RESULTS
In ELISA assays using E-selectin, the IC 50 was 0.15 mM for CY-31214, 0.9 mM for CY-1503,and ú 10 mM for the Lewis X carbohydrates. In ELISA assays using P-selectin, the IC 50 was0.02 mM for CY-31214, ú 10 mM for CY-1503, and ú 10 mM for the Lewis X carbohydrates.
In migration assays utilizing HUVECs, 400 mg/ml of the analogs of sialyl Lewis X inhibitedmigration induced by bFGF to 2% { 1% (CY-31214) and 24% { 9% (CY-1503) of control(Figure 1A). In contrast, the Lewis X carbohydrates at the same concentration of 400 ug/mlstill allowed a migration of 74% { 7% of control. Compared to the effect exerted by theLewis X carbohydrates, the inhibition seen with the analogs of sialyl Lewis X were highlysignificant (p Å 0.00003 for CY-31214 and p Å 0.0016 for CY-1503). The same phenomenonwas observed when HAECs were used (data not shown). The inhibition exerted by the analogsof sialyl Lewis X was dose-dependent (Figure 1B). The monoclonal antibodies to sialyl LewisX at 1:50 dilution inhibited migration to 31% { 5% of control whereas the antibodies to LewisX at the higher 1:25 dilution showed a migration of 75% { 11% of control (Figure 1A).Compared to the antibodies to Lewis X, the inhibition exerted by the antibodies to sialyl LewisX was significant (p Å 0.012).
In proliferation assays, thenon-carbohydrate analog of sialyl Lewis X, CY-31214, was ableto inhibit the growth of HUVECs to 33% { 3% of control (Figure 2A). Cells exposed to thesame concentration of 600 ug/ml of the carbohydrate analog of sialyl Lewis X, CY-1503,showed a proliferation index of 84% { 5% of control and those exposed to the Lewis Xcarbohydrates had a proliferation index of 114% { 3% of control. Compared to the Lewis Xcarbohydrates, the inhibitory effect seen with the analogs of sialyl Lewis X was significant
717
AID BBRC 5654 / 6911$$$341 11-05-96 08:00:18 bbrcas AP: BBRC
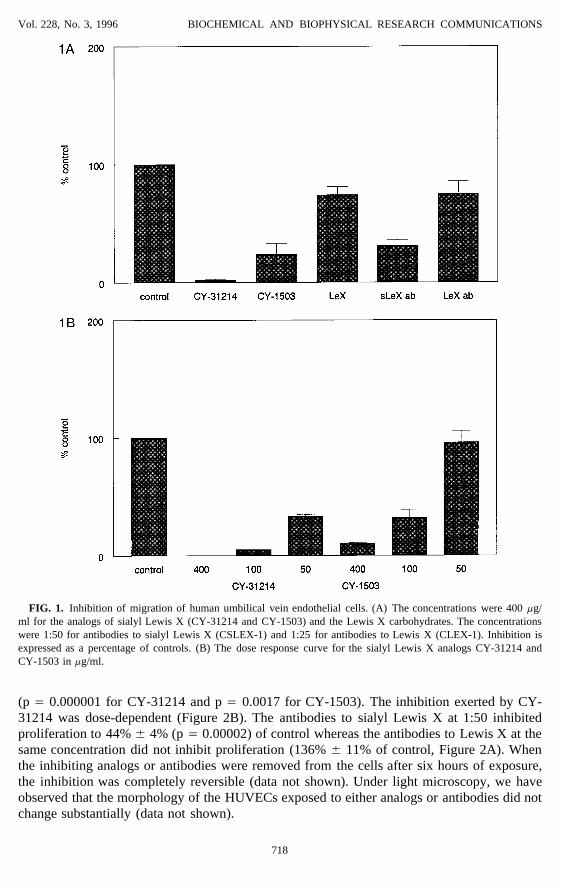
Vol. 228, No. 3, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 1. Inhibition of migration of human umbilical vein endothelial cells. (A) The concentrations were 400 mg/ml for the analogs of sialyl Lewis X (CY-31214 and CY-1503) and the Lewis X carbohydrates. The concentrationswere 1:50 for antibodies to sialyl Lewis X (CSLEX-1) and 1:25 for antibodies to Lewis X (CLEX-1). Inhibition isexpressed as a percentage of controls. (B) The dose response curve for the sialyl Lewis X analogs CY-31214 andCY-1503 in mg/ml.
(p Å 0.000001 for CY-31214 and p Å 0.0017 for CY-1503). The inhibition exerted by CY-31214 was dose-dependent (Figure 2B). The antibodies to sialyl Lewis X at 1:50 inhibitedproliferation to 44% { 4% (p Å 0.00002) of control whereas the antibodies to Lewis X at thesame concentration did not inhibit proliferation (136% { 11% of control, Figure 2A). Whenthe inhibiting analogs or antibodies were removed from the cells after six hours of exposure,the inhibition was completely reversible (data not shown). Under light microscopy, we haveobserved that the morphology of the HUVECs exposed to either analogs or antibodies did notchange substantially (data not shown).
718
AID BBRC 5654 / 6911$$$341 11-05-96 08:00:18 bbrcas AP: BBRC

Vol. 228, No. 3, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 2. Effect on human umbilical vein endothelial cell proliferation. (A) The concentrations were 600 mg/ml forthe analogs of sialyl Lewis X (CY-31214 and CY-1503) and the Lewis X carbohydrates and 1:25 dilution for themonoclonal antibodies to sialyl Lewis X (CSLEX-1) and Lewis X (CLEX-1). (B) The dose response curve for thesialyl Lewis X analog CY-31214 in mg/ml.
In in vivo assays of angiogenesis using the chick chorioallantoic membrane, we found thatthe analogs of sialyl Lewis X at doses of 2 mg or greater produced an avascular zone (Figure3). On the other hand, the Lewis X carbohydrates did not have any effect on the CAM.
We then tested VEGF as well as bFGF in migration and proliferation assays. CY-31214had a significantly more pronounced inhibition of migration mediated by bFGF than VEGF(6% { 6% versus 36% { 11%, p Å 0.034, Figure 4A). The same phenomenon was observedin proliferation assays (67% { 4% versus 128% { 4%, p Å 0.0001). On the other hand, theantibodies to sialyl Lewis X inhibited migration mediated by bFGF and VEGF equally (6%
719
AID BBRC 5654 / 6911$$$341 11-05-96 08:00:18 bbrcas AP: BBRC
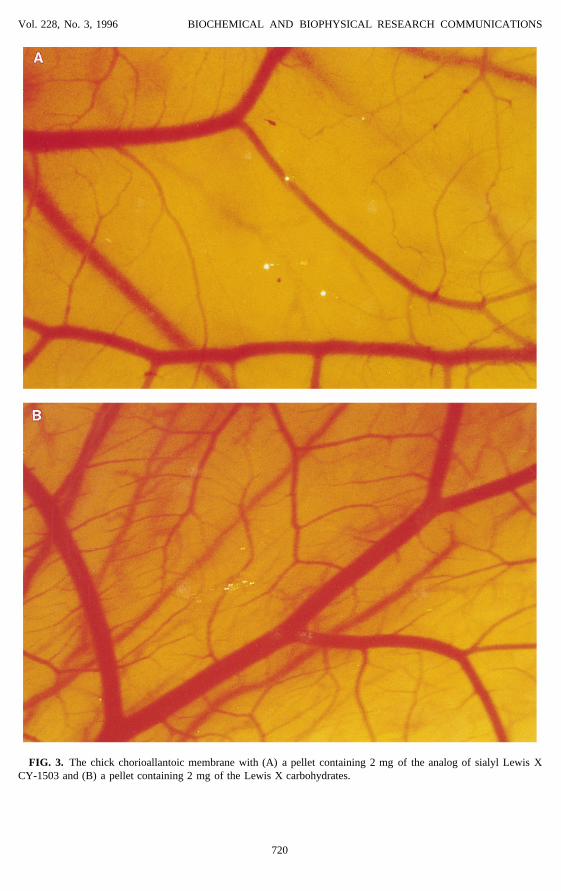
Vol. 228, No. 3, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 3. The chick chorioallantoic membrane with (A) a pellet containing 2 mg of the analog of sialyl Lewis XCY-1503 and (B) a pellet containing 2 mg of the Lewis X carbohydrates.
720
AID BBRC 5654 / 6911$$5654 11-05-96 08:00:18 bbrcas AP: BBRC
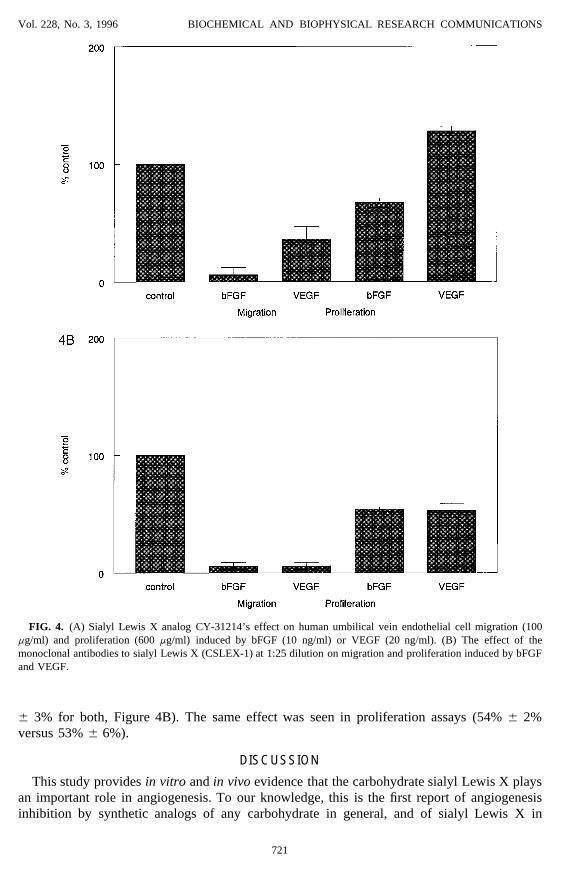
Vol. 228, No. 3, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 4. (A) Sialyl Lewis X analog CY-31214’s effect on human umbilical vein endothelial cell migration (100mg/ml) and proliferation (600 mg/ml) induced by bFGF (10 ng/ml) or VEGF (20 ng/ml). (B) The effect of themonoclonal antibodies to sialyl Lewis X (CSLEX-1) at 1:25 dilution on migration and proliferation induced by bFGFand VEGF.
{ 3% for both, Figure 4B). The same effect was seen in proliferation assays (54% { 2%versus 53% { 6%).
DISCUSSION
This study provides in vitro and in vivo evidence that the carbohydrate sialyl Lewis X playsan important role in angiogenesis. To our knowledge, this is the first report of angiogenesisinhibition by synthetic analogs of any carbohydrate in general, and of sialyl Lewis X in
721
AID BBRC 5654 / 6911$$$341 11-05-96 08:00:18 bbrcas AP: BBRC

Vol. 228, No. 3, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
particular. The inhibition of sialyl Lewis X function with either analogs or antibodies is quitespecific since interference with the function of the related Lewis X carbohydrate did not resultin angiogenesis inhibition.
The analogs and antibodies to sialyl Lewis X may exert their inhibitory effect by interferingwith the interaction between E-selectin and P-selectin and their carbohydrate ligands. In vitro,we have found by ELISA that HUVECs express a low level of E-selectin on the surface, andthis E-selectin may play a role in cell adhesion in the migration assay. Also, sparse HUVECsexpressed more E-selectin than confluent ones, and again this E-selectin may play a role inthe proliferation assay (data not shown). By immunohistochemistry, we have observed theexpression of both E-selectin and P-selectin in newly formed capillaries.
Another interesting finding was that the analogs of sialyl Lewis X inhibit bFGF-mediatedangiogenesis more effectively than that mediated by VEGF. The antibodies to sialyl Lewis Xexerted equal inhibitory effects on both bFGF and VEGF-mediated angiogenesis. The analogsmay bind differentially to the bFGF receptors than to the VEGF receptors. The bFGF receptorshave been described to be glycosylated to N-linked complex oligosaccharides which are neces-sary for receptor function (12). Recently, Friedlander et al. (13) has shown that bFGF andVEGF use different integrins in order to exert their angiogenic activity. Similarly, we speculatethat separate lectins may play a role in the angiogenesis induced by bFGF and VEGF. It ispossible that lectins other than E-selectin and P-selectin are also involved in angiogenesis,since it has been recently reported that endothelial cells from E-selectin-deficient mice canform capillary tubes in vitro (14).
Studies are under way in order to test whether these analogs of sialyl Lewis X can inhibitangiogenesis in animal models. Similar carbohydrate analogs have been found to be nontoxicin animals as well as in human subjects. They have been used effectively in situations involvingthe process of inflammation, where the interaction between E-selectin and its ligands has beenshown to be important (15-18). It is very possible that these analogs of sialyl Lewis X can inthe future contribute to the treatment of angiogenesis-related diseases.
ACKNOWLEDGMENTSThis work was supported by the American Cancer Society, the UCLA Jonsson Comprehensive Cancer Center, and
the Stein-Oppenheim Foundation. We thank Dr. James Paulson for providing us with the analogs and carbohydratesand for critical review of the manuscript and Ms. Geraldine Jackson for technical help.
REFERENCES1. Folkman, J. (1995) Nature Med. 1, 27–31.2. Polverini, P. J. (1996) Am. J. Pathol. 148, 1023–1029.3. Nguyen, M., Strubel, N. A., and Bischoff, J. (1993) Nature 365, 267 –269.4. Nguyen, M., Folkman, J., and Bischoff, J. (1992) J. Biol. Chem. 267, 26157–26165.5. Koch, A. E., Halloran, M. M., Haskell, C. J., Shah, M. R., and Polverini, P. J. (1995) Nature 376, 517 –519.6. Kraling, B. M., Razon, M. J., Boon, L. M., Zurakowski, D., Seachord, C., Darveau, R. P., Mulliken, J. B., Corless,
C. L., and Bischoff, J. (1996) Am. J. Pathol. 148, 1181–1191.7. Fox, S. B., Turner, D. H., Gatter, K. C., and Harris, A. L. (1995) J. Pathol. 177, 369 –376.8. Strubel, N. A., Nguyen, M., Kansas, G. S., Tedder, T. F., and Bischoff, J. (1993) Biochem. Biophys. Res. Commun.
192, 338 –344.9. Ichikawa, Y., Lin, Y. C., Dumas, D. P., Shen, G. Y., Garcia-Junceda, E., Williams, M. A., Bayer, R., Ketcham,
C., Walker, L. E., Paulson, J. C., and Wong, C. H. (1992) J. Am. Chem. Soc. 114, 9283–9298.10. Ball, G. E., O’Neill, R. A., Schultz, J. E., Lowe, J. B., Weston, B. W., Nagy, J. O., Brown, E. G., Hobbs, C. J.,
and Bednarski, M. D. (1992) J. Am. Chem. Soc. 114, 5449–5451.11. Defrees, S. A., Kosch, W., Way, W., Paulson, J. C., Sabesan, S., Halcomb, R. L., Huang, D. H., Ichikawa, Y.,
and Wong, C. H. (1995) J. Am. Chem. Soc. 117, 66–79.12. Feige, J. J., and Baird, A. (1988) J. Biol. Chem. 263, 14023–14029.13. Friedlander, M., Brooks, P. C., Shaffer, R. W., Kincaid, C. M., Varner, J. A., and Cheresh, D. A. (1995) Science
270, 1500–1502.
722
AID BBRC 5654 / 6911$$$341 11-05-96 08:00:18 bbrcas AP: BBRC

Vol. 228, No. 3, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
14. Gerritsen, M. E., Shen, C. P., Atkinson, W. J., Padgett, R. C., Gimbrone, M. A., and Milstone, D. S. (1996) Lab.Invest. 75, 175–184.
15. Paulson, J. C. (1994) Prog. Brain Res. 101, 179–184.16. Mulligan, M. S., Lowe, J. B., Larsen, R. D., Paulson, J., Zheng, Z., Defrees, S., Maemura, K., Fukuda, M., and
Ward, P. A. (1993) J. Exp. Med. 178, 623–631.17. Skurk, C., Buerke, M., Guo, J. P., Paulson, J., and Lefer, A. M. (1994) Am. J. Physiol. 267, H2124–H2131.18. Lefer, D. J., Flynn, D. M., Phillips, M. L., Ratcliffe, M., and Buda, A. J. (1994) Circulation 90, 2390–2401.
723
AID BBRC 5654 / 6911$$$341 11-05-96 08:00:18 bbrcas AP: BBRC


















