![Isolated Testicular Tuberculosis Mimicking Testicular ... involvement, but testicular involvement is an unusual clinical condition [3]. In this report, a case with isolated testicular](https://static.fdocuments.in/doc/165x107/5f3d57bf74280d66ef795ba2/isolated-testicular-tuberculosis-mimicking-testicular-involvement-but-testicular.jpg)
Nonylphenol-induced oxidative stress and cytotoxicity in testicular Sertoli cells
8
Reproductive Toxicology 22 (2006) 623–630 Nonylphenol-induced oxidative stress and cytotoxicity in testicular Sertoli cells Yi Gong, Xiao D. Han ∗ Immunology and Reproductive Biology Laboratory, Medical School, Nanjing University, Nanjing, Jiangsu 210093, PR China Received 19 May 2005; received in revised form 9 April 2006; accepted 14 April 2006 Available online 13 June 2006 Abstract Nonylphenol (NP) as an environment contaminant has been demonstrated to adversely affect male reproduction. The main objective of this study was to evaluate NP-induced oxidative stress and toxicity in testicular Sertoli cells. Sertoli cells were exposed to 10–40 M NP for 24 h. Cell death and growth inhibition were observed by flow cytometric analysis and 3-[4,5-dimethylthiazol-2yl]-2,5-diphenyltetrazolium bromide (MTT) assay, while fluorescein diacetate (FDA) and propidium iodide (PI) staining was used to examine the morphological changes following NP exposure. Subsequently, we found that short-term treatment (2 h) of NP caused intracellular accumulation of reactive oxygen species (ROS), which was evaluated by loading of 2 ,7 -dichlorodihydrofluorescein diacetate (DCFH-DA) without visible morphological changes. Loss of mitochondrial membrane potential (MMP) was detected following 12 and 24 h treatment of NP and assessment by Rhodamine 123 (Rh123) staining. In addition, incubation with NP for 12 h also increased lipid peroxidation of Sertoli cells. These results indicated that low micromolar concentrations of NP induce an adverse oxidative stress in rat Sertoli cells. © 2006 Elsevier Inc. All rights reserved. Keywords: Nonylphenol; Sertoli cells; Reactive oxygen species; Mitochondrial membrane potential; Lipid peroxidation; Cell death 1. Introduction Nonionic surfactants possess specific physico-chemical char- acteristics, including relative ionic insensitivity and sorptive behavior, which lead to their extensive application in indus- try, processing technology and science, and usage in detergents. Alkylphenol ethoxylate (APEO), consisting of approximately 80% nonylphenol ethoxylate (NPEO), is a major group of non- ionic surfactants [1,2]. The primary degradation product of NPEO, nonylphenol (NP) has been documented to appear in aquatic environment and sediment, where the concentration has approached mg/l and mg/kg level, respectively [3–5]. NP has weak estrogenic activity [6,7]. It has been demon- strated that NP could interfere with reproduction in fish, reptiles and mammals, and induce the cell death in gonads and changes to other reproductive parameters [8–10]. Previously, we found that male reproduction and testicular structure was disrupted in rats exposed to NP [11]; however, the precise mechanism of ∗ Corresponding author. Tel.: +86 25 83686497; fax: +86 25 83686559. E-mail address: [email protected] (X.D. Han). action is not clear. Some research has emphasized the estrogenic effect of NP, such as inducing the expression of estrogen receptor (ER) and inhibiting estrogen binding to ER, which might cause endocrine disruption [12,13]. Others studies, however, focused on cytotoxity. For example, NP could induce testicular cell death by inhibiting the activity of endoplasmic reticulum Ca 2+ pumps [14]. Various environmental contaminants can induce oxidative stress by generating reactive oxygen species (ROS) such as hydrogen peroxide (H 2 O 2 ) and superoxide anion (O 2 − ). ROS may initiate sequelae of reactions that damage cellular compo- nents culminating in cell death [15,16]. NP has been shown to produce oxidative stress, enhancing ROS generation in human blood neutrophils [17]. Furthermore, NP administration increased ROS level and lipid peroxidation and depressed the activity of antioxidant enzymes such as superoxide dismutase and glutathione reductase in rat testis [18]. We have taken the hypothesis that oxidative stress may par- tially be responsible for the reproductive toxicity following NP exposure. Sertoli cells are primary supporting cells for sper- matogenesis and they contribute to the construction of the blood–testis barrier [19]. Thus, damage to Sertoli cells may 0890-6238/$ – see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.reprotox.2006.04.019
Transcript of Nonylphenol-induced oxidative stress and cytotoxicity in testicular Sertoli cells

A
sC(ewmii©
K
1
abtA8iNaa
satti
0d
Reproductive Toxicology 22 (2006) 623–630
Nonylphenol-induced oxidative stress andcytotoxicity in testicular Sertoli cells
Yi Gong, Xiao D. Han ∗Immunology and Reproductive Biology Laboratory, Medical School, Nanjing University, Nanjing, Jiangsu 210093, PR China
Received 19 May 2005; received in revised form 9 April 2006; accepted 14 April 2006Available online 13 June 2006
bstract
Nonylphenol (NP) as an environment contaminant has been demonstrated to adversely affect male reproduction. The main objective of thistudy was to evaluate NP-induced oxidative stress and toxicity in testicular Sertoli cells. Sertoli cells were exposed to 10–40 �M NP for 24 h.ell death and growth inhibition were observed by flow cytometric analysis and 3-[4,5-dimethylthiazol-2yl]-2,5-diphenyltetrazolium bromide
MTT) assay, while fluorescein diacetate (FDA) and propidium iodide (PI) staining was used to examine the morphological changes following NPxposure. Subsequently, we found that short-term treatment (2 h) of NP caused intracellular accumulation of reactive oxygen species (ROS), whichas evaluated by loading of 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA) without visible morphological changes. Loss of mitochondrial
embrane potential (MMP) was detected following 12 and 24 h treatment of NP and assessment by Rhodamine 123 (Rh123) staining. In addition,ncubation with NP for 12 h also increased lipid peroxidation of Sertoli cells. These results indicated that low micromolar concentrations of NPnduce an adverse oxidative stress in rat Sertoli cells.
2006 Elsevier Inc. All rights reserved.
mem
ae(eob[
shmnthi
eywords: Nonylphenol; Sertoli cells; Reactive oxygen species; Mitochondrial
. Introduction
Nonionic surfactants possess specific physico-chemical char-cteristics, including relative ionic insensitivity and sorptiveehavior, which lead to their extensive application in indus-ry, processing technology and science, and usage in detergents.lkylphenol ethoxylate (APEO), consisting of approximately0% nonylphenol ethoxylate (NPEO), is a major group of non-onic surfactants [1,2]. The primary degradation product ofPEO, nonylphenol (NP) has been documented to appear in
quatic environment and sediment, where the concentration haspproached mg/l and mg/kg level, respectively [3–5].
NP has weak estrogenic activity [6,7]. It has been demon-trated that NP could interfere with reproduction in fish, reptilesnd mammals, and induce the cell death in gonads and changes
o other reproductive parameters [8–10]. Previously, we foundhat male reproduction and testicular structure was disruptedn rats exposed to NP [11]; however, the precise mechanism of∗ Corresponding author. Tel.: +86 25 83686497; fax: +86 25 83686559.E-mail address: [email protected] (X.D. Han).
aa
temb
890-6238/$ – see front matter © 2006 Elsevier Inc. All rights reserved.oi:10.1016/j.reprotox.2006.04.019
brane potential; Lipid peroxidation; Cell death
ction is not clear. Some research has emphasized the estrogenicffect of NP, such as inducing the expression of estrogen receptorER) and inhibiting estrogen binding to ER, which might causendocrine disruption [12,13]. Others studies, however, focusedn cytotoxity. For example, NP could induce testicular cell deathy inhibiting the activity of endoplasmic reticulum Ca2+ pumps14].
Various environmental contaminants can induce oxidativetress by generating reactive oxygen species (ROS) such asydrogen peroxide (H2O2) and superoxide anion (O2
−). ROSay initiate sequelae of reactions that damage cellular compo-
ents culminating in cell death [15,16]. NP has been showno produce oxidative stress, enhancing ROS generation inuman blood neutrophils [17]. Furthermore, NP administrationncreased ROS level and lipid peroxidation and depressed thectivity of antioxidant enzymes such as superoxide dismutasend glutathione reductase in rat testis [18].
We have taken the hypothesis that oxidative stress may par-
ially be responsible for the reproductive toxicity following NPxposure. Sertoli cells are primary supporting cells for sper-atogenesis and they contribute to the construction of thelood–testis barrier [19]. Thus, damage to Sertoli cells may
6 tive T
lwo
2
2
Kmgip(bVoC
gst
2
kSom0tcswcobc
2
9bwsw
3r
2
htflfddsopou
2
tSNfi2wcg
2
cflFfcr
2
Fm
24 Y. Gong, X.D. Han / Reproduc
ead to impairment of male reproduction. For these reasons,e selected primary rat Sertoli cells cultures to investigate thexidative stress and cytotoxicity caused by NP.
. Material and methods
.1. Chemicals and reagents
NP (4-nonylphenol) with 98% analytical standard was from Tokyo Kaseiogyo Co. (Tokyo, Japan). Dulbecco’s modified Eagle’s medium-Ham’s F-12edium (DMEM-F12 medium), penicillin, streptomycin sulfate, trypsin, colla-
enase I, dimethyl sulphoxide (DMSO), fluorescein diacetate (FDA), propidiumodide (PI) and 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA) wereurchased from Sigma–Aldrich Inc. (St. Louis, MO, USA). C8H17N2O4SNaHEPES sodium salt) and 3-(4,5-dimethylthiazol-2yl)-2,5-diphenyl tetrazoliumromide (MTT) were obtained from Amresco Inc. (Solon, OH, USA). AnnexinApoptosis Kit, Cat. No. Sc-4252 was purchased from Santa Cruz Biotechnol-
gy, Inc. (Santa Cruz, CA, USA). Rhodamine 123 (Rh123) was bought fromalbiochem Inc. (La Jolla, CA, USA).
The culture medium used was DMEM-F12 (1:1) supplemented with 4 mMlutamine, 15 mM Hepes, 6 mM l-(1)-lactic hemicalcium salt hydrate, 1 mModium pyruvate, antibiotics (final concentrations: penicillin, 100 IU/ml; strep-omycin sulfate, 100 �g/ml) and 5% fetal bovine serum.
.2. Isolation and culture of Sertoli cells
Sprague–Dawley Rats were purchased from Nanjing Medical University andept in accordance with NIH Guide for the Care and Use of Laboratory Animals.ertoli cells were isolated from rats at the age of 20 days according to the methodf Steinberger with some modifications [20]. Briefly, testes were decapsulated,inced and rinsed twice in phosphate-buffered saline (PBS), then digested with
.25% trypsin at 37 ◦C for 30 min. The isolated testicular fragments were cen-rifuged at 500 rpm and washed twice in PBS before further digestion in 0.1%ollagenase for 30 min. Digested fragments were filtrated through a 100-meshtainless steel filter. Cells were collected by centrifugation and then washed twiceith PBS before being resuspended in DMEM-F12 medium. Finally, dispersed
ells were seeded into culture flasks and maintained in a humidified atmospheref 95% air, 5% CO2 at 34 ◦C. Two days later, when Sertoli cells attached to theottom of culture flasks, the supernatant was discarded. Then the purified Sertoliells could quickly spread to form a monolayer in new medium.
.3. Cell viability assay
Cell viability was evaluated by the MTT test. Briefly, cells were plated on a
6-well culture plate at 2 × 104 cells/well in 100 �l culture medium. After incu-ation for 24 h, NP at various concentrations was added to each well. Then cellsere cultivated for another 12 or 24 h, followed by the addition of 25 �l MTTolution (5 mg/ml) to each well and further incubation for 4 h. The supernatantsere removed before adding 100 �l DMSO to dissolve the formazan crystal at
pDtwA
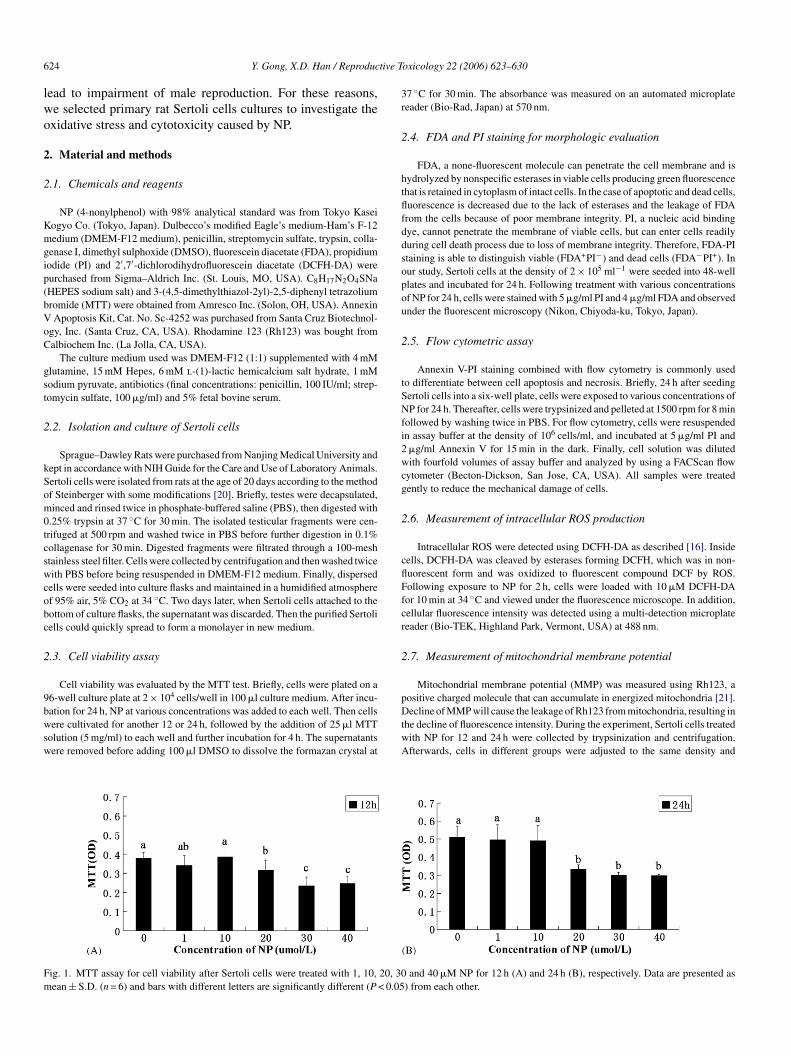
ig. 1. MTT assay for cell viability after Sertoli cells were treated with 1, 10, 20, 30ean ± S.D. (n = 6) and bars with different letters are significantly different (P < 0.05
oxicology 22 (2006) 623–630
7 ◦C for 30 min. The absorbance was measured on an automated microplateeader (Bio-Rad, Japan) at 570 nm.
.4. FDA and PI staining for morphologic evaluation
FDA, a none-fluorescent molecule can penetrate the cell membrane and isydrolyzed by nonspecific esterases in viable cells producing green fluorescencehat is retained in cytoplasm of intact cells. In the case of apoptotic and dead cells,uorescence is decreased due to the lack of esterases and the leakage of FDArom the cells because of poor membrane integrity. PI, a nucleic acid bindingye, cannot penetrate the membrane of viable cells, but can enter cells readilyuring cell death process due to loss of membrane integrity. Therefore, FDA-PItaining is able to distinguish viable (FDA+PI−) and dead cells (FDA−PI+). Inur study, Sertoli cells at the density of 2 × 105 ml−1 were seeded into 48-welllates and incubated for 24 h. Following treatment with various concentrationsf NP for 24 h, cells were stained with 5 �g/ml PI and 4 �g/ml FDA and observednder the fluorescent microscopy (Nikon, Chiyoda-ku, Tokyo, Japan).
.5. Flow cytometric assay
Annexin V-PI staining combined with flow cytometry is commonly usedo differentiate between cell apoptosis and necrosis. Briefly, 24 h after seedingertoli cells into a six-well plate, cells were exposed to various concentrations ofP for 24 h. Thereafter, cells were trypsinized and pelleted at 1500 rpm for 8 min
ollowed by washing twice in PBS. For flow cytometry, cells were resuspendedn assay buffer at the density of 106 cells/ml, and incubated at 5 �g/ml PI and�g/ml Annexin V for 15 min in the dark. Finally, cell solution was dilutedith fourfold volumes of assay buffer and analyzed by using a FACScan flow
ytometer (Becton-Dickson, San Jose, CA, USA). All samples were treatedently to reduce the mechanical damage of cells.
.6. Measurement of intracellular ROS production
Intracellular ROS were detected using DCFH-DA as described [16]. Insideells, DCFH-DA was cleaved by esterases forming DCFH, which was in non-uorescent form and was oxidized to fluorescent compound DCF by ROS.ollowing exposure to NP for 2 h, cells were loaded with 10 �M DCFH-DAor 10 min at 34 ◦C and viewed under the fluorescence microscope. In addition,ellular fluorescence intensity was detected using a multi-detection microplateeader (Bio-TEK, Highland Park, Vermont, USA) at 488 nm.
.7. Measurement of mitochondrial membrane potential
Mitochondrial membrane potential (MMP) was measured using Rh123, a
ositive charged molecule that can accumulate in energized mitochondria [21].ecline of MMP will cause the leakage of Rh123 from mitochondria, resulting inhe decline of fluorescence intensity. During the experiment, Sertoli cells treatedith NP for 12 and 24 h were collected by trypsinization and centrifugation.fterwards, cells in different groups were adjusted to the same density and
and 40 �M NP for 12 h (A) and 24 h (B), respectively. Data are presented as) from each other.

tive T
sirIwfl
2
pw
Tiwtotr
F1as
Y. Gong, X.D. Han / Reproduc
tained with 10 �g/ml Rh123 at 34 ◦C in the dark for 30 min followed by rinsingn PBS. Fluorescence intensity was assessed using a multi-detection microplateeader at excitation wavelength of 488 nm and emission wavelength of 530 nm.n addition, Sertoli cells transplanted into 96-well plate at the same densityere treated with NP for 12 h, stained with Rh123 and photographed under theuorescent microscopy.
.8. Evaluation of lipid peroxidation
Intracellular ROS can initiate a sequence of reactions resulting in theroduction of peroxidation compounds, such as malonic aldehyde (MDA),hich can react with thiobarbituric acid (TBA) and form red products.
2
p
ig. 2. Fluorescent pictures of Sertoli cells with FDA-PI staining. Cells exposed to�M, (C) 10 �M, (D) 20 �M, (E) 30 �M, (F) 40 �M. (G) is higher-magnification picrrow 2 show PI staining within dead cells. Sertoli cells in the control show all viablhow orange within dead cells. Bar = 100 �m.
oxicology 22 (2006) 623–630 625
herefore, thiobarbituric acid-reactive substance (TBARS) assay is a methodn common use to evaluate lipid peroxidation. Sertoli cells were treatedith various concentrations of NP for 12 h, then trypsinized and cen-
rifuged. The pellets triturated by ultrasonic were used to evaluate lipid per-xidation following the protocol of TBARS assay kit. The absorbance ofhe final supernatant was read at 532 nm on a multi-detection microplateeader.
.9. Statistical analysis
Results were shown as mean ± S.D. Statistical evaluation of the data waserformed by using one-way analysis of variance (ANOVA) in SPSS followed
NP for 24 h were stained with FDA (green) and PI (orange): (A) control, (B)tures of Sertoli cells with FDA-PI staining. Arrow1 show FDA staining, whilee cells, and an appreciable number of cells incubated in 20, 30 and 40 �M NP

6 tive T
bl
3
3
tvtwt
3
pfl1oiawms
3
uinlo1e
3
rswwgbw
F1
26 Y. Gong, X.D. Han / Reproduc
y Student–Newman–Keuls post-test. P < 0.05 was accepted as the minimumevel of significance.
. Result
.1. MTT assay for cell viability
Cell viability was assessed by MTT assay following exposureo NP for 12 and 24 h (Fig. 1). NP significantly reduced celliability at concentrations of 20 �M and higher; furthermore,he suppression increased with time. No significant differenceas observed at low concentrations of NP (1 and 10 �M) relative
o the control group.
.2. Sertoli cell morphology
FDA-PI staining was used to evaluate NP-induced mor-hologic changes on Sertoli cells. Fig. 2 shows representativeuorescent micrographs at each concentration of NP (Control,, 10, 20, 30 and 40 �M). NP treatment caused featuresf necrosis such as cell shrinkage and loss of membranentegrity as evidenced by the increase of PI staining (orange)
s well as the decrease of FDA staining (green). This effectas strong at 20 �M and higher. Fig. 2G shows higheragnification, which more explicitly displays the FDA-PItaining.
3
s
ig. 3. NP-induced death of Sertoli cells as determined by flow cytometric assay. Ce0, 20, 30 and 40 �M for 24 h. Numbers (%) in up-left quadrant represent the propor
oxicology 22 (2006) 623–630
.3. Flow cytometric detection of dead cells
Annexin V-PI staining combined with flow cytometry wassed to study the effect of NP on Sertoli cell death. As shownn Fig. 3, higher concentration of NP yielded larger number ofecrotic Sertoli cells, which were present as number (%) in up-eft quadrant, and the effect was significant at the concentrationsf 20 �M and higher. At lower concentrations of NP (1 and0 �M), the percentage of dead cells also increased, but theffect was not as evident.
.4. DCF assay for ROS
The effect of NP on ROS generation, expressed as DCF fluo-escence intensity, was determined in Sertoli cells. After expo-ure to NP for 2 h, although no distinct morphological changesere observed (Fig. 4), ROS increased at 40 �M NP. Thereere no significant differences between 0, 1, 10, 20, 30 �Mroups, although a trend of increasing fluorescence intensity cane observed (Fig. 5). The experiment was repeated four timesith similar results.
.5. Effect of NP on mitochondrial membrane potential
Loss of mitochondrial membrane potential (MMP), as mea-ured by Rh123 staining, is shown in Fig. 6. The results
lls transplanted into six-well plate were exposed to NP at concentrations of 1,tion of dead cells.

Y. Gong, X.D. Han / Reproductive Toxicology 22 (2006) 623–630 627
F re exp2 ed u
rat(
Fesw
ig. 4. DCF fluorescent images of Sertoli cells upon exposure to NP. Cells we0 �M, (E) 30 �M and (F) 40 �M for 2 h, then loaded with DCFH-DA and view
evealed a suppressing effect of NP from 20 �M for 12 h,nd 10 �M for 24 h. Fluorescence images of Sertoli cellsreated with NP for 12 h also revealed the same trendFig. 7).
ig. 5. ROS levels expressed as DCF fluorescence intensity in Sertoli cells uponxposure to NP. Cells exposed to different concentrations of NP for 2 h weretained with DCFH-DA. Data are presented as mean ± S.D. (n = 6) and barsith different letters are significantly different (P < 0.05) from each other.
3
Tpp
4
ltcmdrcmc
i
osed to different concentrations of NP: (A) 0 �M, (B) 1 �M, (C) 10 �M, (D)nder fluorescence microscope. Bar = 50 �m.
.6. NP-induced lipid peroxidation of Sertoli cells
Lipid peroxidation of Sertoli cells was investigated byBARS assay. Both 30 and 40 �M NP markedly induced lipideroxidation (Fig. 8); however, no significant increase of lipideroxidation was detected at other concentrations of NP.
. Discussion
NP is a widespread aquatic contaminant that can accumu-ate in fish [22,23], which increases the latent risk of exposureo higher vertebrates via the food chain. Many reports havelassified NP as hazardous to the health of human and ani-als, especially to male reproduction [11,24]. The present study
emonstrated that NP was toxic to rat Sertoli cells. NP treatmentesulted in the decrease of Sertoli cell viability and increasedell death. Furthermore, ROS generation, loss of mitochondrial
embrane potential and increased lipid peroxidation were indi-ators of oxidative stress detected following NP exposure.It was well documented that exposure to NP exhibited inhib-
ted cellular proliferation and led many kinds of cells to death

628 Y. Gong, X.D. Han / Reproductive Toxicology 22 (2006) 623–630
F al mema measa
[1miuop
tpTa
F2
ig. 6. NP-induced decline of mitochondrial membrane potential. Mitochondrifter incubation in various concentrations of NP for 12 h (A) and 24 h (B) werere significantly different (P < 0.05) from each other.
25,26]. Our study found that concentrations of NP (above0 �M) were active toward Sertoli cell death; however, theechanisms are still unknown. Some researchers hold the opin-
on that NP induces cell death by inhibiting endoplasmic retic-
lum Ca2+ pumps [14]. Others consider NP as the regulatorf cellular cycle [25]. In our study, a slight increase of ROSroduction in Sertoli cells was observed after short-term (2 h)Tlm
ig. 7. Fluorescent images of NP-treated Sertoli cells with Rh123 staining. Sertoli0 �M (D), 30 �M (E) and 40 �M (F) NP for 12 h and stained with Rh123 as mentio
brane potential (indicated by fluorescence intensity of Rh123) of Sertoli cellsured. Data are presented as mean ± S.D. (n = 6) and bars with different letters
reatment with NP but at the time of ROS determination no mor-hologic alteration or decrease of cell viability was observed.his suggested the increased ROS did not result from cell dam-ge but rather from the proximate reaction of NP treatment.
his finding is similar to other studies. For example, NP simu-ated the generation of ROS and impeded cell growth of E. coliutant cells [27]. Another study reported NP-stimulated pro-
cells at the same density were treated with 0 �M (A), 1 �M (B), 10 �M (C),ned in Section 2. Bar = 50 �m.

Y. Gong, X.D. Han / Reproductive T
Fig. 8. Effect of NP on lipid peroxidation of Sertoli cells. Values representcontents of MDA, the product of lipid peroxidation in each NP-treated group,wa(
dip
mpbctnricpwec
oortwibep
HrdgcNoss
coocNso
ataicftce
A
ewta
R
Magni AM, et al. Temporary impairment of reproduction in freshwater
hich were expressed in the form of percentage of control. Data are presenteds mean ± S.D. (n = 6) and bars with different letters are significantly differentP < 0.05) from each other.
uction of ROS in rat epididymal sperm [28]. Therefore, bothn prokaryotic cells and eukaryotic cells, NP can enhance ROSroduction.
Recently, several studies demonstrated that ROS was a com-on inducer of cell death. Increased ROS may induce the
eroxidation of cardiolipin in the mitochondrial inner mem-rane, which triggers dissociation of cytochrome c [29]. Onceytochrome c is released from mitochondria the cell is commit-ed to die by activation of the apoptotic caspase cascade anducleic DNA fragmentation, or necrosis due to collapse of theespiratory function, resulting from over-production of ROS andnsufficient supply of ATP [30]. In this regard, the effect of higheroncentrations of NP on Sertoli cell to dissipate mitochondrialolarization is consistent with other studies [31]. Moreover, itas reported that NP inhibited cellular respiration and proton
xtrusion, which were highly related to the function of mito-hondria [32].
The mitochondrial respiratory chain is a major cellular sitef ROS production [33]. ROS-induced attack to the componentsf mitochondria might in turn cause an increase of ROS level,esulting in the peroxidation of organellar membranes. Fromhe outcomes of lipid peroxidation test following NP treatment,e demonstrated that high concentrations of NP (30–40 �M)
ncreased lipid peroxidation. Lipid peroxidation may alter mem-rane characteristics and functions, leading to cell death. How-ver, it is difficult to prove the causal relationship between lipideroxidation and cell death, as they can affect each other.
Many other chemicals shared the oxidative effect on cells.owever, their concentrations used were very high. It was
eported that 250 �M daidzein could induce great oxidativeamage in human sperm and lymphocytes, while 5 mM halo-enated methane caused hazardous oxidative stress in hepato-ytes [33,34]. In comparison with these doses, the highest dose of
P chosen in our study was relatively small. The oxidative effectf low-dose fumonisin B1 induced a slight increase of oxidativetress; however, LDH release also increased [35]. This suggests alight increase of oxidative stress can induce cytotoxicity, which[
oxicology 22 (2006) 623–630 629
an also support the results of our research. The present study notnly focused on the oxidative stress but also on the confirmationf the dose range that NP was functioning; furthermore, signifi-ance of the present study would be decreased if higher doses ofP were used, as NP level in the environment cannot approach
o high. For all these reasons, only appropriate concentrationsf NP were used in our study.
In summary, direct exposure to NP can induce ROS gener-tion in rat Sertoli cells and trigger a sequence of changes ofhe characteristics and functions of cellular components, suchs loss of mitochondrial membrane potential and lipid perox-dation, subsequently depressing cellular viability and leadingells to death. It is likely that multiple mechanisms are at work,or only a slight increase of ROS was observed following NP-reatment. If all the effects were attributed to ROS, Sertoliells should not have been so dramatically damaged by directxposure.
cknowledgements
The work was supported by the program of National Sci-nce Foundation of China (20577019, 20377022). The authorsish to thank Lingying Niu, Dongmei Li and Guanju Ren for
heir valuable advice in our experiment and Dakai Mu for hisnalytical assistance.
eferences
[1] de Voogt P, de Beer K, der Wielen F. Determination of alkylphenolethoxylates in industrial and environmental samples. Trends Anal Chem1997;16(10):584–95.
[2] Hawrelak M, Bennett E, Metcalfe C. The environmental fate of the primarydegradation products of alkylphenol ethoxylate surfactant in recycled papersludge. Chemosphere 1999;39(5):745–52.
[3] Naylor CG, Mieure JP, Adam WJ, Weeks JA, Castaldi FJ, Ogle LD, etal. Alkylphenol ethoxylates in the environment. J Am Oil Chem Soc1992;69:695–703.
[4] Mark J, Guardia L, Robert C. Alkylphenol ethoxylate degradation prod-ucts in land-applied sewage sludge (biosolids). Environ Sci Technol2001;35:4798–804.
[5] Shao B, Hu J, Yang M, An W, Tao S. Nonylphenol and nonylphe-nol ethoxylates in river water, drinking water, and fish tissues in thearea of Chongqing, China. Arch Environ Contam Toxicol 2005;48:467–73.
[6] Schwaiger J, Mallow U, Ferling H, Knoerr S, Braunbeck Th, Kalbfus W,et al. How estrogenic is nonylphenol? A transgenerational study usingrainbow trout (Oncorhynchus mykiss) as a test organism. Aquat Toxicol2002;59:177–89.
[7] Van den Belt K, Berckmans P, Vangenechten C, Verheyen R, Witters H.Comparative study on the in vitro/in vivo estrogenic potencies of 17-estradiol, estrone, 17-ethynylestradiol and nonylphenol. Aquat Toxicol2004;66:183–95.
[8] Weber PL, Kiparissis Y, Hwang GS, Niimi AJ, Janz DM, Metcalfe CD.Increased cellular apoptosis after chronic aqueous exposure nonylphenoland quercetin in adult medaka (Oryzias latipes). Comp Biochem PhysiolC 2002;131:51–9.
[9] Cardinali M, Maradonna F, Olivotto I, Bortoluzzi G, Mosconi G, Polzonetti-
teleost exposed to nonylphenol. Reprod Toxicol 2004;18:597–604.10] Nagao T, Wada K, Marumo H, Yoshimura S, Ono H. Reproductive effects
of nonylphenol in rats after gavage administration: a two-generation study.Reprod Toxicol 2001;15:293–315.

6 tive T
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
30 Y. Gong, X.D. Han / Reproduc
11] Han XD, Tu ZG, Gong Y, Shen SN, Wang XY, Kang LN, et al. The toxiceffects of nonylphenol on the reproductive system of male rats. ReprodToxicol 2004;19:215–21.
12] Fekadu Y, Augustine A, Anders G, Rune M. Induction of hepatic estrogenreceptor in juvenile Atlantic salmon in vivo by the environmental estrogen,4-nonylphenol. Sci Total Environ 1999;233(1–3):201–10.
13] Kwack SJ, Kwon O, Kim HS, Kim SS, Kim SH, Sohn KH, et al.Comparative evaluation of alkylphenol compounds on estrogenic activ-ity in vitro and in vitro. Toxicol Environ Health A 2002;65(5–6):419–31.
14] Hughes PJ, Mclellan H, Lowes DA, Zafar Khan S, Bilmen JG, ToveySC, et al. Estrogenic alkylphenol inducing cell death by inhibiting testisendoplasmic reticulum Ca2+ Pumps. Biochem Biophys Res Commun2000;277:568–74.
15] Chitra KC, Latchoumycandane C, Mathur PP. Induction of oxidativestress by bisphenol A in the epididymal sperm of rats. Toxicology2003;185:119–27.
16] Wang HT, Yang XL, Zhang ZH, Xu HB. Both calcium and ROS as com-mon signals mediate Na2SeO3-induced apoptosis in SW480 human coloniccarcinoma cells. J Inorg Biochem 2003;97:221–30.
17] Okai Y, Sato EF, Higashi-Okai K, Inoue M. Enhancing effect of theendocrine disruptor para-nonylphenol on the generation of reactiveoxygen species in human blood neutrophils. Environ Health Perspect2004;112(5):553–6.
18] Chitra KC, Mather PP. Vitamin E prevent nonylphenol-induced oxidativestress in testis of rats. Indian J Exp Biol 2004;42(2):220–3.
19] Buzzard JJ, Wreford NG, Morrison JR. Marked extension of proliferationof rat Sertoli cells in culture using recombinant human FSH. Reproduction2002;124:633–41.
20] Steinberger A, Heindel JJ, Lindsey JN, Elkington JS, Sanborn BM, stein-berger E. Isolation and culture of FSH responsive Sertoli cells. Endocr ResCommun 1975;2:261–72.
21] Hong H, Liu GQ. Protection against hydrogen peroxide-induced cytotoxi-city in PC12 cells by scutellarin. Life Sci 2004;74:2959–73.
22] Coldham NG, Sivapathasundaram S, Dave M, Ashfield LA, Pottinger TG,
Goodall C, et al. Biotransformation, tissue distribution, and persistenceof4-nonylphenol residues in juvenile rainbow trout (Oncorhynchus mykiss).DrugMetab Dispos 1998;26:347–53.23] Lewis KS, Lech JJ. In vivo estrogenic activity of nonylphenol in rainbowtrout. Fund Appl Toxicol 1996;30:229–32.
[
oxicology 22 (2006) 623–630
24] Hossaini A, Dalgaard M, Vinggard AM, Frandsen H, Larsen J. Inutero reproductive study in rats exposed to nylphenol. Reprod Toxicol2001;15:537–43.
25] Kudo C, Wada K, Masuda T, Yonemura T, Shibuya A, Fujimoto Y, etal. Nonylphenol induces he death of neural stem cells due to activationof the caspase cascade and regulation of the cell cycle. J Neurochem2004;88(6):1416–23.
26] Aokia M, Kurasaki M, Saito T, Seki S, Hosokawa T, Takahashi Y, et al.Nonylphenol enhances apoptosis induced by serum deprivation in PC12cell. Life Sci 2004;74:2301–12.
27] Okai Y, Sato EF, Higashi-Okai K, Inoue M. Effect of endocrine disrup-tor para-nonylphenol on the cell growth and oxygen radical generation inEscherichia coli mutant cells deficient in catalase and superoxide dismu-tase. Free Radic Biol Med 2004;37(9):1412–8.
28] Chitra KC, Latchoumycandane C, Mathur PP. Effect of nonylphenolon the antiooxidant system in epididymal sperm of rats. Arch Toxicol2002;76(9):545–51.
29] Imai H, Nakagawa Y. Biological significance of phospholipid hydroper-oxide glutathione peroxidase (PHGPx, GPx4) in mammalian cells. FreeRadic Biol Med 2003;34:145–69.
30] Kroemer G, Dallaporta B, Resche RM. The mitochondrial death/life regu-lator in apoptosis and necrosis. A Rev Physiol 1998;60:619–42.
31] Bragadin M, Perin G, Iero A, Manente S, Rizzoli V, Scutari G. An invitro study on the toxic effects of nonylphenol (NP) in microchondria.Chemosphere 1999;38(9):1997–2001.
32] Okai Y, Higashi-Okai K, Machida K, Nakamura H, Nakayama K, FijitaK, et al. Protective effects of �-tocopherol and �-carotene on para-nonylphenol-induced inhibition of cell growth, cellular respiration andglucose-induced proton extrusion of bacteria. FEMS Microbiol Lett2000;187:161–5.
33] Anderson D, Schmid TE, Baumgartner A, Cemeli-Carratala E, BrinkworthMH, Wood JM. Oestrogenic compounds and oxidative stress (in humansperm and lymphocytes in the Comet assay). Mutat Res 2003;544:173–8.
34] Geissa KT, Frazierb JM. QSAR modeling of oxidative stress in vitro fol-
lowing hepatocyte exposures to halogenated methanes. Toxicol In Vitro2001;15:557–63.35] Able S, Gelderblom WCA. Oxidative damage and fumonisin B1-inducedtoxicity in primary rat hepatocytes and rat liver in vivo. Toxicology1998;131:121–31.