
NONIDEAL STATISTICS AND POSITIVE CORRELATION IN …NONIDEAL STATISTICS AND POSITIVE CORRELATION IN...
19
Copyright 0 1983 by the Genetics Society of America NONIDEAL STATISTICS AND POSITIVE CORRELATION IN PHAGE RECOMBINATION: STUDIES WITH X TANDEM DUPLICATION PHAGES JOHN B. HAYS,’ JULIE B. WOLF‘ AND ROBERT J. ZAGURSKY’ Department of Chemistv, University o f Maryland Baltimore County, Catonsuille, Maryland 21228 Manuscript received July 1, 1983 Revised copy accepted August 24, 1983 ABSTRACT The question of nonideality in phage recombination, that is, the extent to which recombinant frequencies differ from those expected from the propor- tions of the two parental types in the mass culture, was addressed by experi- ments with X tandem duplication phages. Isolation and genotypic analysis of triplication-phage progeny, all of which must be the result of intermolecular recombination, yielded a value of about 0.5 for the nonideality parameter h, i.e., the frequency of unlike-parent matings was only about ‘/z the “ideal” value. This value was independent of multiplicity and about the same for the Rec or Red recombination systems. Similar analysis of single-copy phage progeny yielded estimates of k, the ratio of intramolecular to intermolecular recombi- nation of about ’/6 for the Rec system; no intramolecular events were detected in Red-mediated crosses. Consideration of known nonideality factors (finite input, limited number of intracellular sites for phage growth) suggests that the observed h values correspond to intracellular mixing efficiencies of 55 to loo%, depending on the number of intracellular phage growth sites assumed. Analysis of long-range positive correlation (negative interference) indicates that statisti- cal effects caused unlike-parent double crossovers to be three to four times as frequent as an independent-event calculation would predict. In addition, Rec- mediated crosses showed a 1.3-fold positive correlation for unlike-parent cross- overs (in a second interval) among the progeny of like-parent recombinations. INCE genetic recombination of bacteriophages involves interactions among S members of an intracellular pool of phage chromosomes, some sort of statistical theory is necessary if recombinant frequencies are to be interpreted in terms of chromosomal exchanges. A useful basis for phage-mapping func- tions has been the mating theory, which postulates random, sequential matings of pairs of phage chromosomes in the pool (VISCONTI and DELBRUCK 1953; STEINBERC and STAHL 1958; HERSHEY 1958). The elements of the theory and Abbreviations used MOI, multiplicity of infection; PFU, plaque-forming units; RCP, recombination correla- tion parameter; AAUPRCP, RCP for recombination between phage chromosomes synapsed in any (staggered or even) alignment; NSRCP, RCP for recombination among nonxlected phages; SALRCP, RCP for synapsis in staggered alignment between like parental chromosomes; SAURCP, RCP for synapsis in staggered alignment between unlike parental chromosomes. ’ To whom correspondence should be addressed. ’ Present address: GENEX Corporation, 16020 Industrial Drive, Gaithersburg, Maryland 20877. ’ Present address: National Cancer Institute-Frederick Cancer Research Facility, P.O. Box B, Frederick, Maryland 2 170 1. Genetics 105 781-799 December, 1983.
Transcript of NONIDEAL STATISTICS AND POSITIVE CORRELATION IN …NONIDEAL STATISTICS AND POSITIVE CORRELATION IN...

Copyright 0 1983 by the Genetics Society of America
NONIDEAL STATISTICS AND POSITIVE CORRELATION IN PHAGE RECOMBINATION: STUDIES WITH X TANDEM
DUPLICATION PHAGES
JOHN B. HAYS,’ JULIE B. WOLF‘ AND ROBERT J. ZAGURSKY’
Department of Chemistv, University of Maryland Baltimore County, Catonsuille, Maryland 21228
Manuscript received July 1, 1983 Revised copy accepted August 24, 1983
ABSTRACT
The question of nonideality in phage recombination, that is, the extent to which recombinant frequencies differ from those expected from the propor- tions of the two parental types in the mass culture, was addressed by experi- ments with X tandem duplication phages. Isolation and genotypic analysis of triplication-phage progeny, all of which must be the result of intermolecular recombination, yielded a value of about 0.5 for the nonideality parameter h, i.e., the frequency of unlike-parent matings was only about ‘/z the “ideal” value. This value was independent of multiplicity and about the same for the Rec or Red recombination systems. Similar analysis of single-copy phage progeny yielded estimates of k, the ratio of intramolecular to intermolecular recombi- nation of about ’/6 for the Rec system; no intramolecular events were detected in Red-mediated crosses. Consideration of known nonideality factors (finite input, limited number of intracellular sites for phage growth) suggests that the observed h values correspond to intracellular mixing efficiencies of 55 to loo%, depending on the number of intracellular phage growth sites assumed. Analysis of long-range positive correlation (negative interference) indicates that statisti- cal effects caused unlike-parent double crossovers to be three to four times as frequent as an independent-event calculation would predict. In addition, Rec- mediated crosses showed a 1.3-fold positive correlation for unlike-parent cross- overs (in a second interval) among the progeny of like-parent recombinations.
INCE genetic recombination of bacteriophages involves interactions among S members of an intracellular pool of phage chromosomes, some sort of statistical theory is necessary if recombinant frequencies are to be interpreted in terms of chromosomal exchanges. A useful basis for phage-mapping func- tions has been the mating theory, which postulates random, sequential matings of pairs of phage chromosomes in the pool (VISCONTI and DELBRUCK 1953; STEINBERC and STAHL 1958; HERSHEY 1958). The elements of the theory and
Abbreviations used MOI, multiplicity of infection; PFU, plaque-forming units; RCP, recombination correla- tion parameter; AAUPRCP, RCP for recombination between phage chromosomes synapsed in any (staggered or even) alignment; NSRCP, RCP for recombination among nonxlected phages; SALRCP, RCP for synapsis in staggered alignment between like parental chromosomes; SAURCP, RCP for synapsis in staggered alignment between unlike parental chromosomes. ’ To whom correspondence should be addressed. ’ Present address: GENEX Corporation, 16020 Industrial Drive, Gaithersburg, Maryland 20877. ’ Present address: National Cancer Institute-Frederick Cancer Research Facility, P.O. Box B, Frederick, Maryland 2 170 1 .
Genetics 105 781-799 December, 1983.

782 J. B. HAYS, J. B. WOLF AND R. J. ZAGURSKY
its application to recombination of phages X and T4 have recently been re- viewed and discussed in detail by STAHL (1979), in chapter 2 of the reference cited.
The relationship between parental phage frequencies in a mass culture and actual frequencies of intracellular matings between genetically dissimilar chro- mosomes remains an undetermined variable in mating theory. For “ideal” sta- tistics it is assumed that the fraction of matings that involve unlike chromo- somes equals the product of the two mass-culture parental frequencies; devia- tions can be described by some sort of nonideality parameter. Two of the factors that contribute to nonideality, finite input of phages into cells and limitations on the number of phages which can develop in a single cell, have been considered (LENNOX, LEVINTHAL and SMITH 1953). Another factor, fail- ure of clones of replicating chromosomes to be thoroughly mixed with one another, is less susceptible to analysis.
Direct experimental measurement of nonideality has not been possible in the past, since the frequency of genetically silent recombinations (matings be- tween like chromosomes) could not be determined. Here we have employed X tandem duplication phages, recombination of which yields progeny of three distinct buoyant densities: triplication, duplication and “single-copy” phages. All phages in purified triplication and single-copy populations must be the products of recombination (or replicas of such products). Genotypic analysis of the triplication population, thus, yields the ratio of like-parent to unlike-parent matings, which allows calculation of the apparent nonideality factor. Once this factor is known, analysis of the single-copy population permits estimation of the frequency of intramolecular recombination. This latter parameter has the- oretical significance in the formulation of recombination mechanisms and has some practical relevance in the design of in vitro recombination systems (HAYS and ZAGURSKY 1978; NASH 1974; SHULMAN and GOTTESMAN 1971). We find that the proportion of unlike matings is less than half the ideal value, and that intramolecular recombination is relatively infrequent.
We have also used the data from these crosses to analyze factors responsible for long-range positive correlation (“negative interference”) between exchanges in two genetic intervals. Here, as well, duplication-phage recombination affords certain advantages. First, recombination in the duplication region so as to produce triplication and single-copy phages (synapsis in “staggered alignment”) is isolated by nonhomology from recombination elsewhere in the chromosome. Second, comparison of the frequency of second recombinations among the triplication (and single-copy) products of like-parent recombination with that among unselected phages permits an estimation of positive correlation factors (“activation”) unrelated to distribution nonideality. Third, comparison of un- like-parent triplication progeny with like-parent triplications with respect to recombination in a second interval measures positive correlation due solely to distribution nonideality, independent of putative activation effects.
MATERIALS AND METHODS
Bncteria: The Escherichia coli K-12 strains used were: N99, Su- ret+ (MESELSON 28 via H. NASH); N100, as N99 but recA3 (from H . NASH); C600, SuII’ ret' (from F. STAHL); JH1048, S U P recA3

RECOMBINATION OF DUPLICATION PHAGES 783
(recA? transductant of C600); N2217, SuZZZ+ lig7(ts) (from M. GELLERT); RZ181, NlOO (pZ115). Plasmid pZ115 encodes X exo+bet+gam+ under lac PO control (ZAGURSKY and HAYS 1983).
Bacteriobhages: The duplication-deletion phage used for construction of all of the X phages employed in these experiments, X a106-12 b5?8 betam270 cZ857, was itself constructed from X a106-12 b221 betain270 cZ26 (EMMONS and THOMAS 1975) and X b538 betam270 cI857 U. HAYS, unpublished results) by standard techniques. The al06-12 duplication extends from genes I/ to L, 18.6 to 28.2% on the standard X map. X aI06-12 b538 chromosomes are 93.6% A+ length; chromosomes of triplication phages [X(a106-12)2 b5?8] are 103.2% A+ length, and those of single- copy phages (A b538) are 84%. X betam270 phages are "Red-" (lack the phage-coded general recombination system) in N99 but "Red'" in JH1048 and C600. Markers subsequently crossed into X 0106-12 b5?8 betam270 ~1857, their sources, and the product phages used for recombination experiments were as follows: Aam19 (from R. WEISBERG) to yield X a106-12 b538 Aaml9 betam270 rZ857; Anm?2 Bnml (from R. W. BALDWIN) and bet3 (from H. NASH) to yield X a106-12 b538 Ana32 Bainl bet3 ~1857; Jam6 (from R. WEISBERC), bet3 and cZ26 (from F. STAHL) to yield X a106- 12 b5?8 Jam6 bet? cI26; cZ26 to yield X 0106-12 b538 betam270 ~126.
Growth ofphages: The media and procedures have been described in detail elsewhere (HAYS and ZAGURSKY 1978; HAYS, KORBA and KONRAD 1980). Phages were grown in tryptone-maltose broth (TBMM) using C600 bacteria, purified by two rounds of equilibrium sedimentation in CsCI, and stored in TM-buffer.
For lytic infections (single cycles) of bacteria, cells were grown overnight in tryptone broth (TBM) and used to inoculate fresh TBMM cultures. These were incubated (with shaking) at 38", harvested by centrifugation (10,000 rpm for 10 min at 4") in a Sorvall SS-34 rotor at a density of 1 X 10' cells/ml and resuspended to 2 X 10' cells/ml in cold TM-buffer. Apparent multiplicity is the ratio between the titer of the phage stock (PFU/ml), determined using trypticase (TCMB) plates and C600 bacteria as indicator, and the density of the resuspended cells, determined from measured turbidity and a previous calibration [ l I O Klett units (red filter) corresponds to 1 X 10' cells/ml]. Phages and cells were mixed, and after adsorption for 20 min at O", mixtures (usually 0.2 ml) were diluted 50-fold with warm broth (TBM) and shaken for 100 min at 38". Cultures were shaken with chloroform (l.O%, v/v) and centrifuged briefly to remove debris (8000 rpm for 8 min at 4").
Equilibrium sedinzeiztatior~ in CsCI: Lysates were mixed with CsCl (0.76 g/ml of lysate) and sedi- mented in a Beckman Ti50 rotor at 29.000 rpm for 36 h at 4". Fractions (usually 0.15 ml) were collected from the bottoms of tubes into plastic microtiter dishes.
Scoring genotypes of individual plaques: Progeny emerging from crosses of Aam32 Baml bet3 cI857 with betom270 cZ26 phages were scored for all eight possible progeny genotypes (two parental and six recombinant). Lysates were fractionated by two rounds of equilibrium sedimentation in CsCl and several fractions each from triplication, duplication and single-copy pools selected for analysis. Dilutions were plated on TCMB plates (no more than 2 days old) using 0.2 ml of a fresh stationary phase culture of C600 bacteria, so as to yield about 40 plaques per plate.
All plaques, regardless of size, were stabbed with sterile toothpicks and transferred onto appro- priate bacterial lawns for analysis, except that no plaques closer than 5 mm to any other plaques or within 2 mm of the edge of the plate were stabbed. Lawns of the SuZIZ+ lig7(ts) strain N2217, used to distinguish bet? (no plaque) from betam270 phages, were poured using 0.5 ml of an N2217 culture grown overnight in TBM broth at 31", incubated at room temperature for 3-4 h, stabbed and then incubated 16-20 h at 33". Lawns of the Su- strain N99, used to distinguish Aam Bam (no plaque) from A+B+ phages, were poured using 0.3 ml of an N99 culture grown overnight in TBM broth at 31", incubated 2-3 h at 33", stabbed and then incubated at 33". Lawns of the SuZZ+ strain C600, used to distinguish cZ26 (clear plaque) from cZ857 (turbid plaque) phages, were poured using 0.2 ml of overnight culture, stabbed immediately and incubated at 33". The order of stabbing was N2217, N99, C600. About 80 plaques were transferred onto each plate.
Stocks of phages corresponding to all eight genotypes were used as standards. These were plated out every 5-6 days and stabbed onto N2217, N99 and C600 lawns along with the phage plaques to be analyzed. Usually, two plaques of each standard (one small, one large) were stabbed onto every fourth set of analytical plates. There was no correlation between the size of the plaque stabbed and the size of the resulting plaque on the analytical plate for any of the genotypes. Recoveries of recombinant genotypes, from a total of 22,212 plaques scored for two different

784 J. B. HAYS, J. B. WOLF AND R. J. ZAGURSKY
crosses (Table 2), were approximately equal for each of the three pairs of recombinants expected (AB-bet crossovers, bet-cl crossovers, double crossovers), i.e., the scoring procedure exerted no systematic bias against any of the eight genotypes.
RESULTS
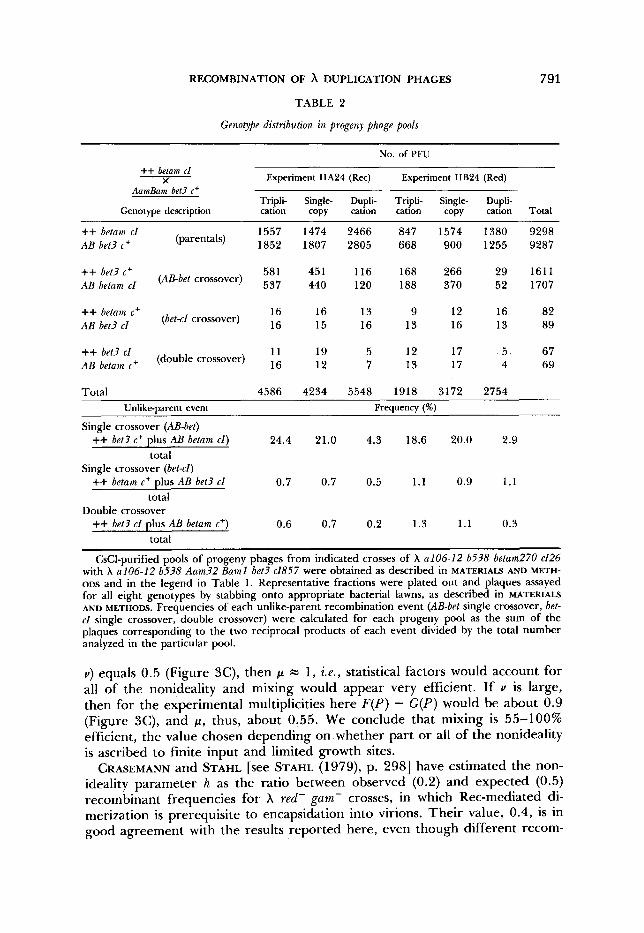
Determination of phage recombination nonideality parameter: Recombination of tandem duplication phages yields chromosomes of three different lengths (Fig- ure 1). The products of intermolecular crossovers involving duplication chro- mosomes in “even” alignment (“equal crossing-over”) are themselves duplica- tions; intermolecular crossovers in “staggered” alignment (“unequal crossing over”) result in triplication and single-copy progeny; intramolecular recombi- nation yields single-copy DNA molecules. As long as all progeny chromosomes can be packaged into phage particles with reasonable efficiency [lengths about 0.75 to 1.08 p1 units (FEISS et al. 1977; STERNBERG et al. 1979)], virions with three distinctive buoyant densities appear among the progeny. These are read- ily separated from one another by equilibrium sedimentation in CsCl (Figure 2A). Approximate frequencies of triplication and single-copy phages, estimated from peak areas, are given in Table 2, for crosses in which recombination was mediated by the E. coli Rec system or by the phage X Red system. In Rec- Red- crosses, no triplication or single-copy phages were observed.
b538
c l Y L Y L J A B i
Aam 1gl I I v bet 3 cl 857 I I I
I I I I I
, Jam6 betam270 cf26
Separation by CsCl Sedimentation I
Analysis of Genotypes FIGURE 1.-Top, Map of phage X genetic markers and tandem chromosomal duplication (V -
L) employed, approximately to scale. Bottom, Cartoon of analysis scheme.

RECOMBINATION OF DUPLICATION PHAGES 785
loo0 1
A. a- s
M I
Fraction Number Fractlon Number
t
+
c E
a
t L
Fraction Number
FIGURE 2.-Equilibrium sedimentation of phages in CsCI. Growth and infection of cells UH1048) with phages X a106-12 6538 Aam32 Baml bet-3 cI857 and X a106-12 6538 betam270 cI26 phages (12 each, cross IIB24) and CsCl equilibrium sedimentation of lysate were as described in MATERIALS AND METHODS. A, Initial equilibrium sedimentation of lysate. B, Resedimentation of peak fractions. Lefthand (triplication) and righthand (single-copy) fractions from initial sedimen- tation were subjected to a second equilibrium sedimentation. PFU per milliliter were determined by plating on C600. C, Frequency of recombinant phages. The frequency of A+B+ cI857 phages among all A+B+ phages was determined by scoring the frequency of turbid plaques when fractions were plated on the Su- strain N99.
Accurate analysis of the triplication and single-copy fractions requires their separation from contaminating duplication phages, most of which are nonre- combinant parental phages, by a second round of CsCl sedimentation (Figure 2B). Those triplication and single-copy fractions in which the frequency of nonparental genotypes (e.g., A+B' ~1857) was constant were judged essentially free from duplication phages (Figure 2C).

786 J. B. HAYS, J. B. WOLF AND R. J. ZAGURSKY
In the accompanying APPENDIX the fraction of all triplication phages that arise by recombination between unlike parents (genetically marked on each side of a chromosome duplication) is shown to be equal to ' / z the nonideality parameter h , when equal parental multiplicities are employed. Thus, when phage distribution statistics are ideal (h = l ) , the fraction of all matings that involve unlike parents is 0.5. The extent to which the unlike-parent fraction of triplication progeny decreases to less than 0.5 ( h < 1) is a measure of the effects upon phage recombination of finite input, limited number of intracel- lular phage growth sites and incomplete intracellular mixing of phage chro- mosomes.
The fraction of unlike-parent triplication progeny is approximately equal to the fraction that is genetically recombinant for two markers that flank the duplication. In the crosses described in Table 2 the flanking markers were either Aam and Jam (crosses I) or Aum Bum and cI (crosses 11), and the fraction of unlike-parent triplication progeny was taken as twice the frequency of the particular recombinant genotype scored. Determinations were made for recom- bination mediated by the E. coli Rec system alone and by the X Red system alone, either phage encoded or plasmid encoded; in some cases the MO1 was varied. Variations in multiplicity and source of Red function had no significant effect. Average values of the nonideality parameter h for Rec-mediated crosses were 0.46 (X Aam betam cI857 X X Jam bet3 cI26) and 0.42 (A Aam Bum bet3 cI857 x X betum cI26) . For Red-mediated crosses, the average value of h was 0.34.
The data in Table 3 were obtained by scoring, in separated triplication, single-copy and duplication pools, all eight genotypes arising from crosses of A+B+ betam cI26 with Aum Bum bet3 cI857 phages (crosses designated in Table 2 as IIA24 and IIB24). Although the number of recombinants scored was quite small in some cases, the expected reciprocal products of the various exchanges appeared with approximately equal frequencies, and no systematic bias against any particular genotype was apparent.
Because the bet locus is closer to the duplication than the c l locus scored in Table 2, the data of Table 3 should provide a slightly more precise experi- mental determination of the fraction of unlike-parent triplication progeny. Also, both products of exchanges in a particular interval were considered in Table 3. The fraction of unlike-parent triplication progeny, taken here as the frequency, among triplication phages, of unlike-parent crossovers in the AB-bet interval plus the frequency of unlike-parent double recombinants, was 0.249 for the Rec-mediated cross and 0.199 for the Red-mediated cross. The cor- responding values of the nonideality parameter h were 0.50 and 0.38. (The Table 3 values corresponding to the experimental definition of the fraction of unlike-parent triplication progeny used in Table 2, i . e . , twice the fraction of unlike-parent AB-cl recombinants, would be 0.250 and 0.198, respectively.)
In a third set of crosses, between Aam Bum K + and A+B+Kam duplication phages at total multiplicities of 5 , 10 and 20 phages/cell, the heterogeneity parameter was again found to be about 0.5 and independent of multiplicity, for Rec+Red-, Rec-Red+ and Rec+Red+ crosses (data not shown).

RECOMBINATION OF DUPLICATION PHAGES 787
Estimation of intramolecular recombination: Deviation of the fraction of unlike- parent single-copy progeny from the “ideal-statistics” value of 0.5, after cor- rection for nonideality effects, should be a measure of the relative frequency of intramolecular recombination. Thus, in the APPENDIX the fraction of unlike- parent single-copy progeny is shown to equal hR/[2(R + l)], where k is defined as the ratio of intramolecular to intermolecular recombination. Using this expression we obtain, from Tables 2 and 3, values for k of 0.45 (IA crosses) and 0.15 to 0.2 (IIA crosses). For Red-mediated crosses, the fraction of unlike- parent recombination progeny among single-copy phages was the same or- higher than that among triplications, i.e., no intramolecular recombination was detectable.
Positive correlation among recombination events: As the APPENDIX describes in more detail, the frequencies of the eight genotypes emerging from three-factor crosses in which two markers flank a chromosomal duplication can be used to construct various RCPs; certain ratios of these parameters (positive correlation ratios) provide information about factors that make recombination in a second interval more likely if a chromosome has already experienced an exchange in a first interval.
In the upper portion of Table 3 are presented frequencies of bet-cZ recom- bination among unselected progeny phages [correlated recombination fre- quency (C)], among phages that have already experienced an unlike-parent recombination in the AB-bet interval [frequency (A)] and among those that have experienced a like-parent recombination in AB-bet [frequency (B)]. These were calculated from the data of Table 2 for appropriate pools of triplication, single-copy and duplication phages.
Appropriate average values of these correlated recombinant frequencies cor- respond to the RCPs defined in the APPENDIX; these appear in the middle portion of Table 3. The RCP for nonselected phages (NSRCP) is a measure of bet-cZ recombination in the entire progeny phage population. The RCP for AB-bet recombination between unlike parents in any (staggered or even) align- ment (AAUPRCP) is a measure of bet-cZ recombination among all progeny (triplication, single-copy, duplication) that have experienced an unlike-parent recombination in AB-bet. The RCP for staggered alignment unlike-parent AB- bet recombinants (SAUPRCP) measures bet-d among triplication and single- copy phages formed by unlike-parent recombination in AB-bet, and the RCP for staggered-alignment AB-bet recombinations (SALPRCP) is the analogous parameter for triplication and single-copy products of like-parent crossovers.
In the APPENDIX it is argued in more detail that certain ratios between RCPs can be interpreted in terms of various sources of positive correlation in recom- bination. The ratio (AAUPRCP)/(NSRCP) compares phages that have already experienced a first unlike-parent recombination to unselected phages and, thus, corresponds to the classical definition of negative interference [see APPENDIX and, for instance, STAHL (1979), pp. 27, 121. Although based on a very small number of doubly recombinant plaques (Table 3) , the experimental values, about 7 for both Rec and Red, are in reasonable agreement with previous estimates by KAISER ( 1 955) and AMATI and MESELSON (1 964).

788 J. B. HAYS, J. B. WOLF AND R. J. ZAGURSKY
The ratio (SAUPRCP)/(NSRCP) compares phages that have already expe- rienced an unlike-parent recombination between duplicated segments staggered with respect to one another (Figure 1, leftmost and rightmost arrows) with unselected phages. The long-range positive correlation (negative interference) measured here should, thus, be free from one postulated source of apparent “excess” double recombinants, formation of extended heterozygous regions by branch migration (LICHTEN and FOX 1983; MAKIN, SZYBALSKI and BLATTNER 1982), since synapsis of duplication chromosomes in staggered alignment au- tomatically results in a substantial (10 kb) nonhomology to the right of the duplicated segments. The observed ratios, about 4 in both cases, were in fact somewhat lower than the “classical negative interference” values (6-7). Thus, it is possible that the 10-kb nonhomology has prevented the formation of extended heterozygous regions and, thus, removed or reduced one source of positive correlation. Although branch migration past nonhomologies extending for a few hundred base pairs during Red-mediated recombination has been demonstrated genetically in vivo (LICHTEN and FOX 1983), and inefficient but significant migration past nonhomologies of similar size mediated by the E. coli RecA protein has been directly observed in vitro (BIANCHI and RADDING 1983), the distance dependence reported by these investigators indicates that branch migration past nonhomologies as large as 10 kb is unlikely.
The ratio (SAUPRCP)/(SALPRCP) compares positive correlation of bet-cZ crossovers with unlike-parent recombination in the duplication region to cor- relation with like-parent recombination in the duplication region. This ratio should reflect only positive correlation due to distribution nonideality (finite input, limited number of growth sites, incomplete mixing), since all chromo- somes considered have experienced at least one recombination (unlike-parent or like-parent) in the AB-bet interval and putative activation effects should cancel out. The ratios, about 3 for Rec and 4 for Red, are slightly higher than the values of 2 and 2.5 that would be predicted on the basis of the respective nonideality parameters measured here (0.5, 0.4).
The ratio (SALPRCP)/(NSRCP), which compares the frequency of bet-cZ recombination among phages that have experienced as well a like-parent re- combination in the duplication region to the frequency among nonselected progeny, should be unity if the only factors contributing to positive correlation are nonideal distribution statistics. The value of 1.3 for Rec-mediated crosses, thus, suggests the involvement of another factor in positive correlation.
t
DISCUSSION
ilionideality in phage distribution statistics: The recombination studies described here exploit a useful property of X tandem duplication phages: their triplication and single-copy progeny, which are readily separated from duplication phages, constitute populations in which every phage chromosome is the result of one or more recombination events. The triplication progeny analyzed here have a genotypic distribution which, differing considerably from that expected for ideal statistics, corresponds to a nonideality parameter of 0.5 or slightly less. Thus, unlike-parent matings are only ‘/9 as frequent as like-parent matings, and

RECOMBINATION OF DUPLICATION PHAGES 789
conventional phage crosses only detect 1/4 of all recombination events. AMATI and MESELSON (1 964) estimated the mean number of unlike-parent exchange clusters per progeny phage to be 0.32 if each exchange cluster is considered to be the result of one crossover event; our data suggest that the average number of crossovers per chromosome is at least 1.2. The contributions to nonideality of three factors, finite phage input, limited intracellular sites, in- complete intracellular mixing, will be considered separately.
Since each bacterium is infected by a relatively small number of phages (finite input), some cells receive unequal numbers of the two parental types, even if parental frequencies in the mass culture are equal. The expected effect on recombinant frequency, assuming Poisson statistics, has been calculated by LENNOX, LEVINTHAL and SMITH (1953) in terms of a reduction factor F(P), where P is the average total multiplicity. Their result is reproduced in Figure 3 A .
The factor F ( P ) must itself be corrected by a second factor G(P, U) if there is a maximum number U of phages that can participate in intracellular devel- opment. This would be the case, for instance, if the number of replication sites or replication complexes were limited (DULBECCO 1949). It has been reported that as few as one to five X chromosomes make the transition from circle-circle to rolling-circle replication (MURIALDO 1974). Curves for G ( P , U) calculated from the LENNOX-LEVINTHAL-SMITH expression for various choices of v, are plotted in Figure 3B. For small values of v, and multiplicities large enough that most cells have more phages than "growth sites" (assuming a Poisson distribution), the correction is large. Since both F(P) and G(P, U) increase with P, the overall statistical factor F ( P ) - G(P, v) is essentially constant over a large range of multiplicities, especially for small values of U (Figure 3C). Thus, our experimentally observed independence of nonideality with respect to mul- tiplicity (Table l ) is consistent with these expressions. However, none of our
A.
Y a50 za75n 0.25 0 5 10 15 20
P
0.25 t[ P
FIGURE S.-Theoretical correction factor curves for phage recombination frequencies. A, The correction factor F(P), for the case of no limit on phage growth sites, calculated according to the equation of LENNOX, LEVINTHAL and SMITH (1953) and plotted as a function of average total multiplicity P . B, The secondary factor C(P, v), to correct F ( P ) when the number of phages participating in intracellular growth cannot exceed v, calculated according to the equation of LENNOX, LEVINTHAL and SMITH (1953) and plotted as a function of average total multiplicity P, for the fixed values of Y indicated. C, The total correction factor, F(P) - C(P, v), plotted as a function of P for fixed 1.

790 J. B. HAYS, J. B. WOLF AND R. J. ZAGURSKY
TABLE 1
Recombznant frequencies in tnplzcatzon and szngle-copy pools
Recombi-
p~~~~~ ~ ~ ~ ~ ~ ~ p , Recombinant frequency (XlO') (XlO')
Experi- nation Total Triplica- Single- Experimental genotypes ment system MO1 tion copy Triplication Single-copy
Parents Aam J+betam c+ A'Jam bet3 c l
Recombinant scoring [all A'J'I
total
Parents A+B+betam cI Aain Bain bet3 c+
Recombinant scoring [all A+B+C+]
[all A+B+]
1A8 IA24 ID8 ID24
IIA8 HA24 IIB24 IIC24 IID24
Rec" Rec" Noneb Noneb
Rec" Rec" Rec' Redd Noneb
8 24 8
24
8 24 24 24 24
ND ND ND' ND'
ND 2.9 2.1 ND N d
ND ND NW N d
ND 3.0 3.4 ND N d
22 f 1 24 f 3
ND ND
20 * 0.5 22 f 1 17 f 1 17 f 1
ND
17 f 1 1 6 f 1
ND ND
1 5 2 1 22 2 2 1 7 2 1 21 f 0.3
ND
Appropriate bacteria were infected with equal numbers of the indicated pairs of duplication phages, at the total multiplicities indicated, and the lysates purified by two rounds of equilibrium sedimentation in CsCI, as described in MATERIALS AND METHODS. Progeny pool sizes were estimated from profiles of the first CsCl gradient (see Figure 2A, for example); where necessary the tails of the central (duplication phase) peak were approximated by eye. Recombinant frequencies were scored as follows: experiments 1, 2 X (titer on strain N99)/(titer on strain C600), corrected for differences in plating efficiency of N99 and C600 bacteria (determined in parallel); experiments 11, frequency of turbids among all plaques on N99. Three central fractions from each purified pool were used for each assay (two to four times in experiments I, once in experiments 11). Parental phage employed: Aam betam e+, X A a m l 9 a106-12 betam270 cI857; A'Jam6 bet3 c l , X a106-12 b538 Jam bet3 c126; A+B+ betam cI, X a106-12 b538 betam270 cI26; Aam Bam be13 c', X a106-12 b538 Aam32 Baml bet3 ~ 1 8 5 7 . ND = not determined.
Crosses in N99. bCrosses in N100. 'Crosses in JH1048. dCrosses in RZ181 with IPTG induc- tion of exo+ bet+. 'Crosses in RZ181 without IPTG induction. 'No peaks of triplication or single- copy phages were apparent.
experimental multiplicities was extremely low, and this work does not consti- tute a test of the LENNOX-LEVINTHAL-SMITH expression.
If phage DNA molecules are not mixed with complete efficiency during replication, matings between sister chromosomes in replicating clones will be favored. Thus, the relative frequency of matings between unlike chromosomes will not be YZ, even in cells in which growth sites are occupied equally by the two parental types. From the definition of the nonideality parameter h in the APPENDIX, it follows that, if the only other factors contributing to statistical nonideality are finite input and limitation on intracellular sites, then the ap- parent mixing efficiency P can be set equal to the nonideality parameter, suitably corrected for finite input and limited growth site effects:
From Tables 2 and 3, we found h to be 0.4 to 0.5. We have no basis here for estimating the maximum number of growth sites. If v = 2, so that F ( P ) - G(P,
RECOMBINATION OF DUPLICATION PHAGES 791
TABLE 2
Genotype distribution in progeny phage pools
No. of PFU ++ betam d
AamBam bet3 c+
X Experiment IIA24 (Rec) Experiment IIB24 (Red)
Tripli- Single- Dupli- Tripli- Single- Dupli- Genotype description cation copy cation cation copy cation Total
++ betain c l 1557 1474 2466 847 1574 1380 9298 1852 1807 2805 668 900 1255 9287 AB bet? c + (parentals)
581 451 116 168 266 29 1611 537 440 120 188 370 52 1707
(AB-bet crossover) ++ bet? c+ AB betain c l
++ betain c+ 16 16 13 9 12 16 82 16 15 16 13 16 13 89 AB bet3 c l (bet-cl crossover)
1 1 19 5 12 17 5 67 16 12 7 13 17 4 69
++ (double crossover) AB betam c+
Total 4586 4234 5548 1918 3172 2754 Unlike-Darent event Frequency (%)
Single crossover (AB-bet) ++ bet? c+ plus AB betam c l ) 24.4 21.0 4.3 18.6 20.0 2.9
total Single crossover (bet-cl)
++ betam c+ DIUS AB bet3 c l 0.7 0.7 0.5 1 . 1 0.9 1 . 1 total
Double crossover ++ bet? cl plus AB betam c+) 0.6 0.7 0.2 1.3 1 . 1 0.3
CsC1-purified pools of progeny phages from indicated crosses of X a106-12 b538 betum270 c126 with X a106-12 b538 Aam32 Bum1 bet? c1857 were obtained as described in MATERIALS AND METH- ODS and in the legend in Table 1 . Representative fractions were plated out and plaques assayed for all eight genotypes by stabbing onto appropriate bacterial lawns, as described in MATERIALS AND METHODS. Frequencies of each unlike-parent recombination event (AB-bet single crossover, bet- cl single crossover, double crossover) were calculated for each progeny pool as the sum of the plaques corresponding to the two reciprocal products of each event divided by the total number analyzed in the particular pool.
v) equals 0.5 (Figure 3C), then p = 1, i.e., statistical factors would account for all of the nonideality and mixing would appear very efficient. If v is large, then for the experimental multiplicities here F ( P ) - G(P) would be about 0.9 (Figure 3C), and p, thus, about 0.55. We conclude that mixing is 55-100% efficient, the value chosen depending on whether part or all of the nonideality is ascribed to finite input and limited growth sites.
CRASEMANN and STAHL [see STAHL (1979), p. 2981 have estimated the non- ideality parameter h as the ratio between observed (0.2) and expected (0.5) recombinant frequencies for X red- gam- crosses, in which Rec-mediated di- merization is prerequisite to encapsidation into virions. Their value, 0.4, is in good agreement with the results reported here, even though different recom-
total

792 J. B. HAYS, J. B. WOLF AND R. J. ZAGURSKY
TABLE 3
Positive correlations among recombinant frequencies
Relative frequency (X 10py
++ betam cl X Experiment llA24 (Rec) Experiment IIB24 (Red)
AamBam be13 c+ Pools pools
Correlated recombinant Ex rimental Tripli- Single- Dupli- Tripli- Single- Dupli- frequency &nition cation copy cation cation copy cation
(A) bet-cl recombinants
among products of unlike-parent ex- changes in AB-bet in- terval
(B)
among products of like-parent exchanges in AB-bet interval
bet-rl recombinants
(C) bet-cl recombinants
among unselected progeny
AB betam c+
all AB betam A'B'bet3 c l
all A ' P bet3
Average
A'B+ betam c' all A+B+ betam AB bet3 c l all AB bet3 Average
All betam c+
all betam All bet3 cl
all bet3
2.9 2.7 5.5 6.5 4.4 7.1
1.9 4.0 4.1 6.7 6.0 14.7
2.4 3.3 4.8 6.6 5.2 10.9
1.0 1.1 NA 1.1 0.8 NA
0.9 0.8 NA 1.9 1.8 NA
0.9 0.9 NA 1.5 1.3 NA
NA NA 0.8 NA NA 1.4
NA NA 0.7 NA NA 1.4
-
Recombination correlation parameter Experimental definition Rec Red
Average NA NA 0.7 NA NA 1.4 Relative frequency ( ~ 1 0 ' )
NSRCP AAUPRCP SAUPRCP
SALPRCP
Frequency (C) (duplication)b 0.7 1.4 Frequency (A) (duplicationr 4.8 10.9 Frequency (A) (triplication-sin- 2.9 5.7
gle-copy average)'
de -cow averaeey Frequency (B) (triplication-sin- 0.9 1.4
Positive correlation ratio
Positive correlation effect' Experimental definition of ratio Rec Red
Classical negative interference
Strictly long-range positive corre-
Pure nonideality lation
Activation
AAUPRCP NSRCP
SAUPRCP NSRCP
SAUPRCP SAL P R C P SALPRCP
NSRCP
6.5 f 1.0 7.9 f 2.7
3.9 f 1.0 4.1 f 0.6
3.1 f 0.7 4.1 f 0.6
1.3 f 0.1 1.0 f 0.5
NA = not applicable. " Genotype distribution data were used to calculate, for appropriate progeny pools, the indicated
recombinant frequencies.

RECOMBINATION OF DUPLICATION PHAGES 793
bination systems were involved. [Since CRASEMANN and STAHL employed gam- phages, the E. coli RecBC pathway was most likely the predominant operative system (UNGER, ECHOLS and CLARK 1972). In our Rec+ crosses (red+ gam+ phage) the RecBC system would have been partially, but not totally, inhibited so that recombination would have been mediated by both the RecBC and RecF pathways (CLARK 1974; KORBA and HAYS 1980).]
Intramolecular recombination: The genotype distribution of single-copy phages reflects the same nonideality factors that affect triplication phages, plus intra- molecular recombination. The ratio k, of frequencies of intramolecular to in- termolecular conversion of duplication to single-copy chromosomes, was found here to be about '/7 to ' /5 for the most thoroughly analyzed Rec-mediated crosses. Although this value is subject to considerable experimental uncertainty, k was significantly less than 1.0 in three of four measurements and in previous U. B. HAYS and R. J. ZAGURSKY, unpublished results) experiments with Aam Bum and Kam phages. This estimate does not depend on assumptions about packaging efficiency of phage chromosomes of different lengths nor on as- sumed ideality of mating statistics. It is considerably lower than previous esti- mates of k for various X recombination processes, for which statistical ideality was assumed: 1-5 for general recombination of X duplication phages (HAYS and ZAGURSKY 1978); 2.5 for site-specific integrative recombination (NASH 1974); 4 for site-specific excisive recombination (SHULMAN and GOTTE~MAN 197 1).
If k is taken as a measure of the competition, for any given chromosome site, between the homologous site on the adjacent duplicated segment (intra- moIecular recombination) and homologous sites on other chromosomes (inter- molecular recombination), then the average effective concentration of phage DNA in the cell can be estimated. We first calculate, following the method of WANG and DAVIDSON (1 969), the probability density for homologous regions
The NSRCP, defined rigorously as the frequency of bet-cl recombinants among progeny neither selected for AB-bet recombination nor fractionated according to chromosome length, is here a p proximated by the frequency among nonselected duplication phages, since duplications comprise more than 90% of the unfractionated progeny (Table 1) and triplication and single-copy phages do not show exceptionally high bet-cl recombinant frequencies (Table 2).
' The AAUPRCP, defined rigorously as the frequency of bet-cl recombinants among progeny selected for unlike-parent AB-bet recombination but not fractionated according to chromosome length, is here approximated by the frequency among AB-bet recombinant (unlike-parent) dupli- cation phage, since unlike-parent triplication and single-copy phages only comprise about 1.2% of the total phage population (Table l), and their bct-cl recombinant frequencies are not exceptionally high (Table 2).
The SAUPRCP is defined as the frequency of bet-cl recombinants among triplication and single- copy phages formed by unlike-parent recombination in AB-bet.
e The SALPRCP, rigorously defined as the frequency of bet-cl recombinants among phages formed by like-parent intermolecular recombinants, is approximated here by an average over triplication and single-copy phages without correction for intramolecular recombination events, which were only about '/6 as frequent as intermolecular events. 1 Positive correlation ratios, ascribed to the effects indicated and defined experimentally as
indicated, were calculated from the RCP values in the middle portion of Table 3.

7 94 J. B. HAYS, J. B. WOLF AND R. J. ZAGURSKY
of the duplicated segments to be in the neighborhood of one another, by the formula of JACOBSON and STOCKMAYER (1950), j = [3/(2IILb)]"'. If the statis- tical segment length b is assumed to be 150 nm (HAYS, MAGAR and ZIMM 1969) and the contour length L of DNA between corresponding points in the duplicated segments is taken to be 1.56 pm [assuming A+ length of 16.2 pm (SANGER et al. 1982)], then j is 2.76 x 10" ~ m - ~ . In Rec-mediated crosses, intramolecular recombination is about l/6 as frequent as intermolecular; the effective concentration of phage chromosomes is, therefore, 6j, or about 16 X 10" molecules ~ m - ~ . If the volume of an E. coli cell is taken as 1.6 X lo-'' cm3 (WATSON 1976), this average effective concentration corresponds to about 26 phage chromosomes per cell.
For Red-mediated crosses the frequency of nonparental genotypes was slightly greater among single-copy phages than among triplications, so that formal calculations of intramolecular recombination yielded negative numbers. We cannot explain this observation. Perhaps, in some of the like-parent mat- ings, those between the daughter chromosomes of a replicating molecule for example, the mechanism that prevents Red-mediated events from yielding re- ciprocal pairs of recombinants [see STAHL (1979), p. 291 acts somewhat selec- tively against single-copy progeny.
Positive correlation between recombination events: The well-known negative inter- ference, or positive correlation, between multiple exchanges in X recombina- tion (KAISER 1955; AMATI and MESELSON 1964) is qualitatively consistent with the nonideality effects measured here. Recombinations that actually reassort genetic markers are more likely in cells in which nearly equal numbers of parental-type chromosomes participate in growth and there is efficient mixing of opposite types with one another. Thus, chromosomes that have successfully accomplished one exchange are more likely to be in a situation favoring an- other. We have shown here that the frequency of linkage alterations is only half or less of that expected for ideal statistics. This result implies a correlation factor of slightly more than 2.0 and is, thus, in reasonable agreement with the factor of 3.0 reported by KAISER and by AMATI and MESELSON, with the values of 2.2 (Rec-mediated) and 1.5 (Red-mediated) reported by LICHTEN and Fox (1983), who used widely separated genetic intervals, and with the value of about 2 (Rec+Red+) observed by MAKIN, SZYBALSKI and BLATTNER (1982) using intervals separated by nonhomologies. However, our own estimates of long-range positive correlation are 4-6 when all correlation effects are taken into account, and 3-4 when only statistical nonideality is considered. We can- not explain this discrepancy, which appears to exceed the (considerable) ex- perimental variations in our data, but do note that our experimental conditions (Red+Gam+RecA- and Red-Gam+RecA+) were different from those employed in the cited works.
The Rec crosses display an additional small positive correlation factor, not explainable in terms of phage distribution statistics. Triplication phages arising from like-parent crossovers in the AB-bet interval are about 1.3 times as likely to have experienced unlike-parent crossover in the bet-cl interval as unselected phages, as if one mating had slightly activated the chromosome for a second

RECOMBINATION OF DUPLICATION PHAGES 795
mating with a different partner. Perhaps, RecA protein remains bound after the first exchange is completed. Also, recombination (to form triplication chro- mosomes) might sometimes engender extra replication forks, as is the case with phage T4 (LUDER and MOSIG 1982), and these forks might be more recom- binogenic than replication initiating at the X ori locus. Red-mediated crosses did not display the apparent activation phenomenon.
This work was supported by an National Institutes of Health grant (GM22586) and by American Cancer Society grants, (NP-293A; NP293B) to J.B.H. A University of Maryland Graduate Fellow- ship to R.J.Z. is acknowledged. We thank SCOTT EMMONS for suggesting experimental approaches, CLAIRE WYMAN, TOM JORDAN, LAMBROS VIENNAS, CONSTANCE SUMMERS and GANESAN SRINIVASAN for technical assistance and GARY KNOTT, Division of Computer Research and Technology, Na- tional lnstitutes of Health, for computer summation of series.
LITERATURE CITED
Localized negative interference in bacteriophage A. Genetics
Heteroduplex DNA with large unpaired regions made by Rec A protein. J. Cell. Biochem. 7B (Suppl): 152.
Progress toward a metabolic interpretation of genetic recombination of Esch- erichia coli and bacteriophage lambda. Genetics 7 8 259-272.
The number of particles of bacteriophage T 2 that can participate in intra- cellular growth. Genetics 34 126-1 32.
Tandem genetic duplications in phage lambda. IV. The locations of spontaneously arising tandem duplications. J. Mol. Biol. 91: 147-1 52.
Packaging of the bacteriophage X chromosome length. Virology 77: 281-293.
Novel mutations of Escherichia coli that produce recombinogenic lesions in DNA. I. Identification and mapping of arl mutations. J. Mol. Biol. 139 455-472.
Persistence length of DNA. Biopolymers 8:
Generalized recombination in tandem duplications of
The production of recombinants in phage crosses. Cold Spring Harbor
lntramolecular reaction in polycondensations. I.
AMATI, P., and M. MESELSON, 1964 51: 369-379.
BIANCHI, M. E. and C. M. RADDING, 1983
CLARK, A. J., 1974
DULBECCO, R., 1949
EMMONS, S. E. and J. 0. THOMAS, 1975
FEISS, M., R. A. FISHER, M. A. CRAYTON and C. EGNER, 1977
HAYS, J. B., B. E. KORBA and E. B. KONRAD, 1980
HAYS, J. B., E. M. MAGAR and B. H. ZIMM, 1969
HAYS, J. B. and R. J. ZAGURSKY, 1978
HERSHEY, A. D., 1958
531-536.
bacteriophage lambda. Mol. Gen. Genet. 160: 325-330.
Symp. Quant. Biol. 23: 19-46.
The theory of linear systems. J. Chem. Phys. 18: 1600-1606. JACOBSON, H. and W. H. STOCKMAYER, 1950
KAISER, A. D., 1955
KORBA, B. E. and J. B. HAYS, 1980
A genetic study of the temperate coliphage lambda. Virology 1: 424-443.
Novel mutations of Escherichia coli that produce recombino- genic lesions in DNA. 11. Properties of recombinogenic X phages grown on bacteria carrying arl mutations. J. Mol. Biol. 139: 473-489.
LENNOX, E. S., C. LEVINTHAL and F. SMITH, 1953 The effect of finite input in reducing recom- binant frequency. Genetics 38: 508-51 1.
LICHTEN, M. and M. S. Fox, 1983 Effects of nonhomology on bacteriophage lambda recombi- nation. Genetics 103: 5-22
LUDER, A. and G. MOSIG, 1982 Proc. Natl. Acad. Sci. USA 7 9 1101-1105.

796
MAKIN, G. J. V., W. SZYBAISKI and F. R. BLATTNER, 1982 Asymmetric effects of deletions and
MURIALDO, H., 1974 Restriction in the number of infecting lambda phage genomes that can
NASH, H., 1974 Xatt B-att P , a X derivative containing both sites involved in integrative recom-
SANGER, F., A. P. COULSON, G. F. HONG, D. F. HILL and G. B. PETERSEN, 1982 Nucleotide
SHULMAN, M. and M. COTTESMAN, 1971
J. B. HAYS, J. B. WOLF AND R. J. ZAGURSKY
substitutions on high negative interference in coliphage lambda. Genetics 102: 299-3 17.
participate in intracellular growth. Virology 6 0 128-1 38.
bination. Virology 57: 207-216.
sequence of bacteriophage X DNA. J. Mol. Biol. 162: 729-773. Lambda att: a transducing phage capable of intramo-
lecular int-xis promoted recombination. pp. 477-488. In: The Bacteriophage Lambda, Edited by A. D. HERSHEY. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
STAHL, F. W., 1979 Genetic Recombanation: Thinking about It in Phage and Fungi. W. H. Freeman and Company, San Francisco.
STEINBERG, C. and F. STAHL, 1958 The theory of formal phage genetics. Cold Spring Harbor Symp. Quant. Biol. 23: 42-46.
STERNBERG, N., D. HAMILTON, L. ENQUIST and R. WEISBERG, 1979 A simple technique for the isolation of deletion mutants of phage lambda. Gene 8 35-51.
UNGER, R. C., H. ECHOIS and A. J. CLARK, 1972 Interaction of the recombination pathways of bacteriophage A and host Escherichia coli: effects on X recombination. J. Mol. Biol. 70: 531- 537.
The mechanism of genetic recombination in phage. Ge- netics 38: 5-33.
Cyclization of phage DNAs. Cold Spring Harbor Symp.
The Molecular Biology of the Gene. p. 61. W. A. Benjamin, Inc., Menlo Park,
Expression of the phage X recombination genes ex0 and
VISCONTI, N. and M. DELBRUCK, 1953
WANG, J. C. and N. DAVIDSON, 1969
WATSON, J. D., 1976
ZAGURSKY, R. J. and J. B HAYS, 1983
Quant. Biol. 33: 409-415.
California.
bet under lac PO control on a multi-copy plasmid. Gene 23: 277-292.
Corresponding editor: G. MOSIG
APPENDIX
DETERMINATION OF NONIDEALITY PARAMETER, INTRAMOLECULAR RECOMBINATION FREQUENCY AND POSITIVE CORRELATION FACTORS IN
PHAGE RECOMBINATION
John B. Hays Determination of nonideality parameter: In a biparental phage cross that conforms to ideal statistics
the only factor that determines the fraction of intracellular matings that involve phage chromo- somes of unlike genotype is the frequency f of a given parent in the mass-phage population. In this case the frequency of unlike-parent encounters among all matings is 2Al - J), and the observable recombinant frequency is proportional to this factor. However, there are at least three sources of nonideality that may reduce the probability of unlike-genotype matings and, thus, decrease the observable recombinant frequency.
First, the number of infecting phages is not the same for every cell but is randomly distributed, the finite input effect. The result is a lower probability of unlike-parent matings, hence, a lower recombinant frequency, in some cells. This effect has been analyzed theoretically by LENNOX, LEVINTHAL and SMITH (1953), who assumed perfect mixing of intracellular phage chromosomes

RECOMBINATION OF DUPLICATION PHAGES 797
and a Poisson distribution of phages among cells. They calculated the recombinant frequency as proportional to F ( P ) 2 [ f ( l -A], where the finite input correction factor F(P) depended only on the average total number of phages per cell P . From the plot of F ( P ) in Figure 3A, it can be seen that the finite input effect is negligible for total multiplicities in excess of 20 or so.
Second, the number of intracellular sites at which phage growth can take place may be limited (DULBECCO 1969). If so, statistics will be nonideal no matter how high the multiplicity. Assuming that there are only U sites per cell at which phage DNA replication can take place, and that only phage chromosomes occupying these sites (or their descendents) participate in recombination events, LENNOX, LEVINTHAL and SMITH ( 1 9 5 3 ) derived an expression G(P, v) to correct F ( P ) for finite site effects. This correction term and the combined factor [ F ( P ) - G(P, v)] are graphed in Figure 3, B and C, respectively.
Third, phage chromosomes within infected cells might not be distributed randomly. For ex- ample, the members of one intracellular clone of replicating chromosomes might be inefficiently mixed with members of other clones, so that like-genotype matings would be more probable than unlike. That the effect upon recombinant frequency of incomplete mixing might be represented by a single factor, independent of the multiplicity P and the growth site limit v, may be seen by examining the derivation by LENNOX et al. of the correction factor [ F ( P ) - G(P, 41. This factor was obtained by summing the terms corresponding to the probability of unlike-parent mating in a cell infected with m phage of one type and n of a second type over all values of m and n . (Each term was weighted by the corresponding Poisson distribution probability for an average total multiplicity of P phages per cell.) The fraction of matings that involve unlike parents, 2mn in the perfect-mixing case, would be 2pmn if the effective concentration of unlike chromosomes available to a given chromosome of the first type were proportional to pn rather than n. Therefore, a constant mixing parameter p can be factored out of each term in the sum over all m and n of the individual probabilities. The resulting total correction factor pc[F(P) - G(P, v)] may then be re- placed by a single nonideality parameter, h, so that the frequency of genetically distinguishable recombinants will be proportional to 2 h u l - f ) ] , where f and 1 - f a r e parental frequencies in the mass-phage population. The frequency of like-parent (genetically indistinguishable) recombi- nants will not be a simple function of h but will be 1 - 2 h m l - f ) ] .
Consider recombination between duplication phages genetically marked on each side of the duplication. Iff = 1 - f = 0 . 5 , or nearly so, and there are no nonideality effects, (h = l.O), half of the phages in the triplication pool will be the result of recombination between unlike parents (genetically distinguishable recombinants) and half will be products of like-parent recombination. If recombination statistics are not ideal, the corresponding proportions will be 0.5h and 1-0.5h. Thus, h is equal to twice the fraction of triplication phages with nonparental genotypes.
Estimation of intramolecular recombznatzon frequenczes: Following the notation of NASH (1974), let k equal the ratio [frequency of intramolecular recombination between duplication phage chromo- somes] to [frequency of intermolecular recombination of duplication chromosomes in staggered alignment (to yield triplication and single-copy progeny)]. If the two parental multiplicities are equal, then in the pool of single-copy phages the ratio (unlike-parent intermolecular recombinants) to (like-parent intermolecular recombinants) to (intramolecular recombinants) must be h / 2 : (1 - h / 2 ) : k . Thus, the ratio k can be obtained from the experimental fraction of unlike-parent single- copy recombinants, which must equal h/[2(k + l)].
Positme correlatzon of recombination events (negative znterference): The traditional definition of positive correlation (negative interference) is the ratio of the observed frequency of double recombinants (measured in three-factor crosses) to the predicted independent-event double-crossover frequency (product of the two corresponding single-crossover frequencies measured in two-factor crosses). The data presented in the accompanying report differ from traditional interference data in two respects. First, novel recombination parameters are measured. These parameters include frequen- cies, among the products of staggered-alignment recombination (unequal crossing over) in a given interval (t.e., triplication and single-copy phages), of phages that are recombinant (by genetic criteria) in the same and/or another interval. Clearly, the apparent frequency, among triplication or single-copy progeny, of genetically observable single crossovers in the interval containing the duplication is quite different from the recombinant frequency in that interval among unselected phages. Thus, positive correlation must be described in terms of parameters other than the classical

798 J. B. HAYS, J. B. WOLF AND R. J. ZAGURSKY
two-factor crossover frequencies traditionally used to define negative interference. Second, all eight genotypes arising from three-factor crosses are scored, providing the data used in these new definitions.
I define recombination correlation parameters (RCPs) to be frequencies of progeny recombinant in one interval among those already recombinant (by physical and/or genetic criteria) in another interval and will use various RCPs to define positive correlation ratios. Let the parental genotypes be X-Y-Z- and X+Y+Z+, where X and Yare markers that flank a tandem chromosomal duplication. We will be interested in the frequency of Y-Z crossovers (either direction) among various subpop- ulations of X-Y recombinants. These subpopulations are designated [O], [I], [2] and [3].
Subpopulation [ O ] consists of completely unselected progeny phages (nonrecombinant or recom- binant in the duplication-containing interval X - 9 . The corresponding correlation parameter for recombination in the second interval Y-Z, designated the “nonselected RCP” (NSRCP), is experi- mentally defined as the frequency of all Y+Z- phages among all Y+ (or equivalently, the frequency of all Y-Z+ among all r).
Subpopulation [ 11 is defined as all phages resulting from an unlike-parent recombination in the duplication-containing interval, in any alignment (staggered or even) and, thus, consists of dupli- cation, triplication and single-copy phages, all genetically recombinant in the X-Y interval. The corresponding correlation parameter for Y-Z recombination, designated the “any-alignment unlike- parent RCP” (AAUPRCP), may be experimentally defined as the frequency of X-Y+Z- phages among all X-Y+ (or of X+Y-Z+ among all X+Y-).
Subpopulation [2] consists of all phages resulting from recombination in the duplication region between unlike parents, in staggered alignment, i .e. , all triplication (or single-copy) phages that are genetically recombinant in X-Y. The corresponding Y-Z recombination parameter, designated the “staggered-alignment unlike-parent RCP” (SAUPRCP), is the frequency of X-Y+Z- among all X-Y+ triplications (or X+Y-Z+ among X’Y-).
Subpopulation [3] consists of all phages resulting from recombination in the duplication region between like parents in staggered alignment, i .e. , all triplication phages (or single-copy phages not produced by intramolecular recombination) of parental genotype at X and Y. The corresponding Y-Z recombination parameter, designated the “staggered-alignment like-parent RCP” (SALPRCP), is the frequency of X+Y+Z- among X+Y+ triplications (or X-Y-Z+ among X-Y-).
Certain ratios of the RCPs defined are of interest in the analysis of correlated recombination events. First, if recombination per se somehow activates a chromosome for a second event, then chromosomes that have already recombined in X-Y, even with an identical chromosome, are more likely to subsequently recombine in Y-Z with an unlike parent than chromosomes that have never recombined. If the probability that the next mating partner is of unlike genotype (which should depend only on multiplicity and distribution nonideality) is assumed to be the same for the chro- mosomes in subpopulation [3] (like-parent triplications) and [0] (unselected) than the ratio of the corresponding recombination correlation parameters (SALPRCP)/(NSRCP) can be considered a measure of activation.
Second, if subpopulation [2] (unlike-parent triplications) and [ 31 (like-parent triplications) are assumed to be activated by their X-Y recombinations to the same extent (if at ail), then the ratio (SAUPRCP/SAUPRCP) measures the greater probability of subsequent mating with an unlike parent (so as to yield Y-Z recombination) that would be expected for subpopulation [3] chromo- somes (which would most likely mate again with the same partners with which they accomplished the X-Y recombination). This “pure nonideality” ratio should be a function of the factors affecting the nonideality parameter h.
Third, subpopulation [2] should be more likely than subpopulation [3] to recombine in Y-Z, both because of activation and because of distribution nonideality. Thus, the corresponding cor- relation ratio, (SAUPRCP)/(NSRCP), should be a measure of all long-range positive correlation effects. The staggered-alignment synapsis that is presumably a necessary step in the formation of triplication phages imposes a nonhomology block on any branch migration to the right that might otherwise create a long heterozygous region, initiated in X-Y and extending into Y-2, and on other effects dependent on continuous homology. Extensive branch migration has been postulated to be a source of apparent “excess” double crossovers [“high” or “localized” negative interference; MAKIN, SZYBALSKI and BLATTNER 1982; LICHTEN and Fox (1983)l. Formally, in terms of the definitions,

RECOMBINATION OF DUPLICATION PHAGES 799
"strictly long-range positive correlation" is the product of the activation and pure nonideality factors.
Fourth, the ratio (AAUPRCP)/(NSRCP), which compares subpopulation [ 11 to subpopulation [O], reflects all sources of positive correlation: nonideality, activation, long heterozygous regions (or other effects dependent on continuous homology). It is easy to show that this ratio is equivalent to the classical definition of negative interference (frequency of Y-2 recombinants among X-Y recombinations divided by product of X-Y and Y-2 frequencies among unselected phages).


















