
Nitric Oxide Mediates the Hormonal Control of … · Nitric Oxide Mediates the Hormonal Control of...
15
Nitric Oxide Mediates the Hormonal Control of Crassulacean Acid Metabolism Expression in Young Pineapple Plants 1[W][OA] Luciano Freschi, Maria Aurineide Rodrigues, Douglas Silva Domingues, Eduardo Purgatto, Marie-Anne Van Sluys, Jose Ronaldo Magalhaes, Werner M. Kaiser, and Helenice Mercier* Departamento de Bota ˆnica, Instituto de Biocie ˆncias, Universidade de Sa ˜o Paulo, CEP 05508–900, Sao Paulo, Brazil (L.F., M.A.R., D.S.D., M.-A.V.S., H.M.); Departamento de Alimentos e Nutric ¸a ˜o Experimental, Faculdade de Cie ˆncias Farmace ˆuticas, Universidade de Sa ˜o Paulo, CEP 05422–970, Sao Paulo, Brazil (E.P.); Empresa Brasileira de Pesquisa Agropecua ´ria, CEP 36038–330, Juiz de Fora, Brazil (J.R.M.); and Julius-von-Sachs-Institut fu ¨ r Biowissenschaften, Lehrstuhl fu ¨ r Molekulare Pflanzenphysiologie und Biophysik, D–97082 Wuerzburg, Germany (W.M.K.) Genotypic, developmental, and environmental factors converge to determine the degree of Crassulacean acid metabolism (CAM) expression. To characterize the signaling events controlling CAM expression in young pineapple (Ananas comosus) plants, this photosynthetic pathway was modulated through manipulations in water availability. Rapid, intense, and completely reversible up-regulation in CAM expression was triggered by water deficit, as indicated by the rise in nocturnal malate accumulation and in the expression and activity of important CAM enzymes. During both up- and down-regulation of CAM, the degree of CAM expression was positively and negatively correlated with the endogenous levels of abscisic acid (ABA) and cytokinins, respectively. When exogenously applied, ABA stimulated and cytokinins repressed the expression of CAM. However, inhibition of water deficit-induced ABA accumulation did not block the up-regulation of CAM, suggesting that a parallel, non-ABA-dependent signaling route was also operating. Moreover, strong evidence revealed that nitric oxide (NO) may fulfill an important role during CAM signaling. Up-regulation of CAM was clearly observed in NO-treated plants, and a conspicuous temporal and spatial correlation was also evident between NO production and CAM expression. Removal of NO from the tissues either by adding NO scavenger or by inhibiting NO production significantly impaired ABA-induced up-regulation of CAM, indicating that NO likely acts as a key downstream component in the ABA-dependent signaling pathway. Finally, tungstate or glutamine inhibition of the NO-generating enzyme nitrate reductase completely blocked NO production during ABA-induced up-regulation of CAM, characterizing this enzyme as responsible for NO synthesis during CAM signaling in pineapple plants. Crassulacean acid metabolism (CAM) is present in more than 6% of vascular plant species and represents one of the most complex plant adaptations to limited water availability (Cushman, 2001). By allowing CO 2 uptake at night when the evapotranspiration rates are lower, the CAM pathway brings about substantial water conservation and represents a considerable adaptive advantage for plants living in habitats char- acterized by seasonal or intermittent restrictions in water supply (Lu ¨ ttge, 2004; Winter et al., 2005). The extent to which the biochemical and physiolog- ical traits of CAM are expressed can greatly vary both within and between species (Lu ¨ ttge, 2004). The so- called C 3 -CAM facultative plants are probably the best examples of intraspecific plasticity in the expression of CAM, since these rare plants are able to perform either C 3 or CAM photosynthesis depending on the environ- mental conditions (Cushman, 2001; Cushman and Borland, 2002). In contrast, the vast majority of CAM species are classified as constitutive CAM plants, exhibiting a more limited flexibility in the proportion of CO 2 uptake over the day and night (Winter et al., 2008). Regardless of these differences in classification, the degree of CAM expression in both facultative and constitutive CAM species has been shown to be strongly dependent on the photosynthetic maturity of the tissues (Winter et al., 2008). As a result, even well-documented constitutive CAM species may per- form C 3 photosynthesis during their initial stages of development (Jones, 1975; Holthe et al., 1987; Winter et al., 2008); therefore, a progression from C 3 to CAM is believed to occur along their ontogeny. However, 1 This work was supported by the Fundac ¸a ˜o de Amparo a ` Pesquisa do Estado de Sa ˜o Paulo. * Corresponding author; e-mail [email protected]. The author responsible for distribution of materials integral to the findings presented in this article in accordance with the policy described in the Instructions for Authors (www.plantphysiol.org) is: Helenice Mercier ([email protected]). [W] The online version of this article contains Web-only data. [OA] Open Access articles can be viewed online without a sub- scription. www.plantphysiol.org/cgi/doi/10.1104/pp.109.151613 Plant Physiology Ò , April 2010, Vol. 152, pp. 1971–1985, www.plantphysiol.org Ó 2010 American Society of Plant Biologists 1971 www.plantphysiol.org on July 12, 2018 - Published by Downloaded from Copyright © 2010 American Society of Plant Biologists. All rights reserved.
-
Upload
nguyencong -
Category
Documents
-
view
220 -
download
0
Transcript of Nitric Oxide Mediates the Hormonal Control of … · Nitric Oxide Mediates the Hormonal Control of...

Nitric Oxide Mediates the Hormonal Control ofCrassulacean Acid Metabolism Expression in YoungPineapple Plants1[W][OA]
Luciano Freschi, Maria Aurineide Rodrigues, Douglas Silva Domingues, Eduardo Purgatto,Marie-Anne Van Sluys, Jose Ronaldo Magalhaes, Werner M. Kaiser, and Helenice Mercier*
Departamento de Botanica, Instituto de Biociencias, Universidade de Sao Paulo, CEP 05508–900, Sao Paulo,Brazil (L.F., M.A.R., D.S.D., M.-A.V.S., H.M.); Departamento de Alimentos e Nutricao Experimental,Faculdade de Ciencias Farmaceuticas, Universidade de Sao Paulo, CEP 05422–970, Sao Paulo, Brazil(E.P.); Empresa Brasileira de Pesquisa Agropecuaria, CEP 36038–330, Juiz de Fora, Brazil (J.R.M.); andJulius-von-Sachs-Institut fur Biowissenschaften, Lehrstuhl fur Molekulare Pflanzenphysiologie undBiophysik, D–97082 Wuerzburg, Germany (W.M.K.)
Genotypic, developmental, and environmental factors converge to determine the degree of Crassulacean acid metabolism(CAM) expression. To characterize the signaling events controlling CAM expression in young pineapple (Ananas comosus)plants, this photosynthetic pathway was modulated through manipulations in water availability. Rapid, intense, andcompletely reversible up-regulation in CAM expression was triggered by water deficit, as indicated by the rise in nocturnalmalate accumulation and in the expression and activity of important CAM enzymes. During both up- and down-regulation ofCAM, the degree of CAM expression was positively and negatively correlated with the endogenous levels of abscisic acid(ABA) and cytokinins, respectively. When exogenously applied, ABA stimulated and cytokinins repressed the expression ofCAM. However, inhibition of water deficit-induced ABA accumulation did not block the up-regulation of CAM, suggestingthat a parallel, non-ABA-dependent signaling route was also operating. Moreover, strong evidence revealed that nitric oxide(NO) may fulfill an important role during CAM signaling. Up-regulation of CAM was clearly observed in NO-treated plants,and a conspicuous temporal and spatial correlation was also evident between NO production and CAM expression. Removalof NO from the tissues either by adding NO scavenger or by inhibiting NO production significantly impaired ABA-inducedup-regulation of CAM, indicating that NO likely acts as a key downstream component in the ABA-dependent signalingpathway. Finally, tungstate or glutamine inhibition of the NO-generating enzyme nitrate reductase completely blocked NOproduction during ABA-induced up-regulation of CAM, characterizing this enzyme as responsible for NO synthesis duringCAM signaling in pineapple plants.
Crassulacean acid metabolism (CAM) is present inmore than 6% of vascular plant species and representsone of the most complex plant adaptations to limitedwater availability (Cushman, 2001). By allowing CO2uptake at night when the evapotranspiration rates arelower, the CAM pathway brings about substantialwater conservation and represents a considerableadaptive advantage for plants living in habitats char-acterized by seasonal or intermittent restrictions inwater supply (Luttge, 2004; Winter et al., 2005).
The extent to which the biochemical and physiolog-ical traits of CAM are expressed can greatly vary bothwithin and between species (Luttge, 2004). The so-called C3-CAM facultative plants are probably the bestexamples of intraspecific plasticity in the expression ofCAM, since these rare plants are able to perform eitherC3 or CAM photosynthesis depending on the environ-mental conditions (Cushman, 2001; Cushman andBorland, 2002). In contrast, the vast majority of CAMspecies are classified as constitutive CAM plants,exhibiting a more limited flexibility in the proportionof CO2 uptake over the day and night (Winter et al.,2008). Regardless of these differences in classification,the degree of CAM expression in both facultative andconstitutive CAM species has been shown to bestrongly dependent on the photosynthetic maturityof the tissues (Winter et al., 2008). As a result, evenwell-documented constitutive CAM species may per-form C3 photosynthesis during their initial stages ofdevelopment (Jones, 1975; Holthe et al., 1987; Winteret al., 2008); therefore, a progression fromC3 to CAM isbelieved to occur along their ontogeny. However,
1 This work was supported by the Fundacao de Amparo aPesquisa do Estado de Sao Paulo.
* Corresponding author; e-mail [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policydescribed in the Instructions for Authors (www.plantphysiol.org) is:Helenice Mercier ([email protected]).
[W] The online version of this article contains Web-only data.[OA] Open Access articles can be viewed online without a sub-
scription.www.plantphysiol.org/cgi/doi/10.1104/pp.109.151613
Plant Physiology�, April 2010, Vol. 152, pp. 1971–1985, www.plantphysiol.org � 2010 American Society of Plant Biologists 1971 www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

despite the large number of constitutive CAM species,little is known about the environmental and en-dogenous control of CAM expression in such plants(Cushman and Borland, 2002; Winter et al., 2008).
In fact, much of our current understanding of theregulation of CAM expression has derived from stud-ies on C3-CAM facultative species (Cushman andBorland, 2002; Taybi et al., 2002). In these plants,environmental factors such as light intensity, temper-ature, salinity, photoperiod, and especially wateravailability have long been recognized to affect themagnitude of CAM expression (Cushman andBorland, 2002; Taybi et al., 2002), and some signalingevents underlying the conversion of these environ-mental signals into changes in the expression andoperation of the CAM have already been identified(Taybi et al., 2002).
In the facultative CAM species Mesembryanthemumcrystallinum, the most intensively studied CAMmodel,elevations in endogenous abscisic acid (ABA) areusually observed during the CAM induction by saltor water stress (Thomas et al., 1992; Taybi et al., 1995;Taybi and Cushman, 2002), while exogenous ABA caninduce the expression and activity of several CAMenzymes even in the absence of environmental con-straints (Chu et al., 1990; Taybi et al., 1995). Althoughthese observations strongly suggest that ABA is in-volved in CAM induction, the inhibition of ABAsynthesis did not block the salt- or drought-inducedCAM expression inM. crystallinum, suggesting that anABA-independent signaling pathway also mediatesthe C3-CAM transition process in this plant (Thomaset al., 1992; Taybi and Cushman, 2002).
Cytokinins (CKs) are also known to participate in thecontrol of CAM expression inM. crystallinum; however,contradictory effects have been obtained depending onthe organ treated with these hormones (Thomas et al.,1992; Peters et al., 1997). CKs applied to roots trigger anincrement in phosphoenolpyruvate carboxylase (PEPC)transcript abundance, while foliar application stronglyinhibits the induction of PEPC expression by salt ordrought stress (Schmitt and Piepenbrock, 1992; Thomaset al., 1992; Dai et al., 1994; Peters et al., 1997). However,since a reduction in endogenous CKs was observedduring the C3-to-CAM transition in wilting leaves ofM.crystallinum, these hormones have been suggested to actas negative regulators of CAM expression (Peters et al.,1997). Comparatively fewer studies have analyzed theinvolvement of other hormonal classes in the signaltransduction chain leading to the establishment ofCAM (Taybi et al., 2002). Based on these limited data,auxins and ethylene (ET) seem to have no effect onCAM induction (Dai et al., 1994; Hurst et al., 2004),whereas GA3 andmethyl jasmonatewere found to havea limited influence on CAM expression, acting aspositive and negative regulators, respectively (Taybiet al., 2002).
In addition to plant hormones, some other signalingfactors have also been shown to participate in thesignaling cascades that underpin the expression of
CAM in M. crystallinum (Taybi and Cushman, 1999;Taybi et al., 2002). Current data demonstrate thatcalcium (Ca2+) mobilization from both intracellularand extracellular pools represents a crucial step duringCAM induction by drought, salinity, osmotic stress, orABA (Taybi and Cushman, 1999; Taybi et al., 2002). Onthe other hand, inositol 1,4,5-triphosphate mediatesspecifically the ABA-dependent signaling pathwaythat leads to CAM, not affecting the induction of thisphotosynthetic behavior by dehydration, salinity, orosmotic stress (Taybi and Cushman, 2002). Moreover,hydrogen peroxide (H2O2) has also been shown toinduce CAM expression when exogenously applied atrelatively high concentrations, and increases in theendogenous levels of this reactive oxygen species werefound to precede the establishment of CAM in re-sponse to salt stress (Slesak et al., 2003, 2008). Fur-thermore, protein synthesis, phosphorylation, anddephosphorylation are clearly involved in the controlof CAM expression in M. crystallinum (Taybi andCushman, 1999, 2002), although the exact targets ofthese regulatory mechanisms remain to be established.
Given the multitude of environmental cues impli-cated in the establishment and operation of the CAMpathway, the involvement of other signaling com-pounds in the regulation of CAM induction seemsinevitable. Based on the premise that CAM expressionis particularly affected by abiotic constraints, signalingmolecules associated with plant stress responses rep-resent a logical target for new studies on the signalingmechanisms that induce and/or maintain the CAMexpression. Nitric oxide (NO), for instance, has re-cently emerged as a mandatory participant in severalplant adaptive responses to biotic and abiotic stresses(Gould et al., 2003; Arasimowicz and Floryszak-Wieczorek, 2007; Corpas et al., 2008). In fact, accumu-lating evidence indicates that this free radical, whichwas first identified as a second messenger in animals,plays important roles in a myriad of physiologicalprocesses in plants, including defense responses topathogens (Delledonne et al., 1998) and environ-mental challenges such as drought (Garcia-Mata andLamattina, 2001, 2002; Neill et al., 2008), salinity (Zhaoet al., 2004), heat (Gould et al., 2003), cold (Corpas et al.,2008; Zhao et al., 2009), UV-B radiation (Tossi et al.,2009), ozone (Ederli et al., 2006), and heavy metaltoxicity (Rodriguez-Serrano et al., 2009).
In addition, increased NO production by plant tis-sues has also been reported to occur in response toseveral abiotic stresses such as drought, salinity, os-motic stress, and low temperature (Gould et al., 2003;Arasimowicz and Floryszak-Wieczorek, 2007; Corpaset al., 2008). Under these stressful conditions, nitratereductase (NR) and/or a nitric oxide synthase (NOS)-like activity have been suggested as themain enzymaticsources responsible for the increasedNO production byplant tissues (Zhao et al., 2001, 2009; Hao et al., 2008,Neill et al., 2008; Sang et al., 2008). Current evidencealso indicates that NO closely interacts with severalsignaling molecules usually involved in plant adaptive
Freschi et al.
1972 Plant Physiol. Vol. 152, 2010 www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

stress responses, including ABA (Garcia-Mata andLamattina, 2002; Bright et al., 2006), cytosolic Ca2+
(Courtois et al., 2008), and H2O2 (Neill et al. 2008).Taking into consideration that these samemolecules arealso known to participate in the signaling of CAMinduction and the fact that many environmental chal-lenges that stimulate NO production also trigger the C3-to-CAM transition in facultative CAM plants (Taybiet al., 2002; Slesak et al., 2003, 2008), it seems plausiblethat NO could also have a role in the signal transduc-tion chain leading to CAM expression. However, to ourknowledge, no study has evaluated the role of NO inthe induction or up-regulation of CAM either in facul-tative or constitutive CAM plants.Among CAM plants, cultivated pineapple (Ananas
comosus) is by far the most valuable commercial spe-cies (Cushman, 2001). Adult plants of pineapple areconsidered a classical example of a constitutive CAMplant (Martin, 1994; Zhu et al., 1997), but little isknown about the photosynthetic behavior of youngindividuals of this species. In a previous study, wehave demonstrated that young (3-month-old) pineap-ple plants exhibited C3 photosynthesis when culti-vated under constant diurnal temperature, whereas atypical CAM cycle was induced by thermoperiodictreatment (Nievola et al., 2005; Freschi et al., 2009).However, the influence of several environmental fac-tors and their downstream signaling networks on thecontrol of CAM expression in this agronomically im-portant species has remained largely unexplored.In this study, the roles and interrelationships of
water stress, plant hormones, and NO in the regula-tion of CAM expression in young pineapple plantswere investigated through a combination of pharma-cological, biochemical, and physiological approaches.By changing water availability, we have demonstratedthat young individuals of this bromeliad can displayrapid and completely reversible changes in the levelsof CAM expression, which were closely associatedwith marked variations in the endogenous hormonalbalance. Evidence was obtained that water deficit-induced up-regulation of CAM in pineapple mayinvolve both ABA-dependent and -independent path-ways, whereas CKs seem to represent a potent repres-sor of CAM induction by water stress. Finally, we alsohave shown, to our knowledge for the first time, thatNO plays an important role during the signalingevents controlling CAM expression in young pineap-ple plants, acting as an essential downstream compo-nent in the ABA-dependent signaling pathway.
RESULTS
CAM Expression in Young Pineapple Plants IsInfluenced by Water Availability
Under control conditions, young (3-month-old)pineapple plants exhibited a low background level ofCAM, as indicated by the reduced transcript accumu-
lation and activities of the CAM enzymes PEPC,malate dehydrogenase (MDH), and phosphoenolpyr-uvate carboxykinase (PEPCK), associated with onlybasal values of nocturnal accumulation of malate (Dmalate; Fig. 1). On the other hand, when water deficitwas imposed by incubating the plants in a growthmedium containing 30% polyethylene glycol (PEG), aprogressive rise in the expression of these CAM en-zymes was observed after 5 d of treatment (Fig. 1, A, C,and E), revealing a relatively rapid up-regulation ofCAM in response to this abiotic stress. After 7 d ofwater deficit, the activities of these same enzymesincreased in an exponential fashion (Fig. 1, B, D, and F)with a parallel accumulation of malate during thenight (Fig. 1G). This intensification of CAM activitycoincided with a significant reduction in the relativewater content (RWC) of the pineapple leaf tissues (Fig.1H; from 78% on day 4 to 52% on day 7). In addition,the highest activities of the CAM enzymes and Dmalate were observed when RWC was already at theminimum values (Fig. 1).
In contrast, when water deficit-treated plants weresubjected to a recovery period in the absence of PEG, agradual decrease in transcript abundance and activi-ties of CAM enzymes was observed (Fig. 2, A–F),which was associated with a corresponding reductionin the D malate (Fig. 2G). This attenuation in CAMexpression was paralleled by a progressive increase inthe leaf RWC (Fig. 2H). Overall, the water deficit-induced increases in D malate and in the activities ofthe CAM enzymes were fully reverted between 15 and20 d after returning the plants to a PEG-free medium(Fig. 2, A–G), coinciding, therefore, with the reestab-lishment of the original water content in the pineappleleaf tissues (Fig. 2H). Altogether, these data revealedthat young pineapple plants exhibit a remarkablephotosynthetic flexibility in response to changes inwater availability, allowing a detailed investigation ofthe signaling events involved in the control of CAMexpression in this species.
ABA and CKs Modulate CAM Expression in YoungPineapple Plants
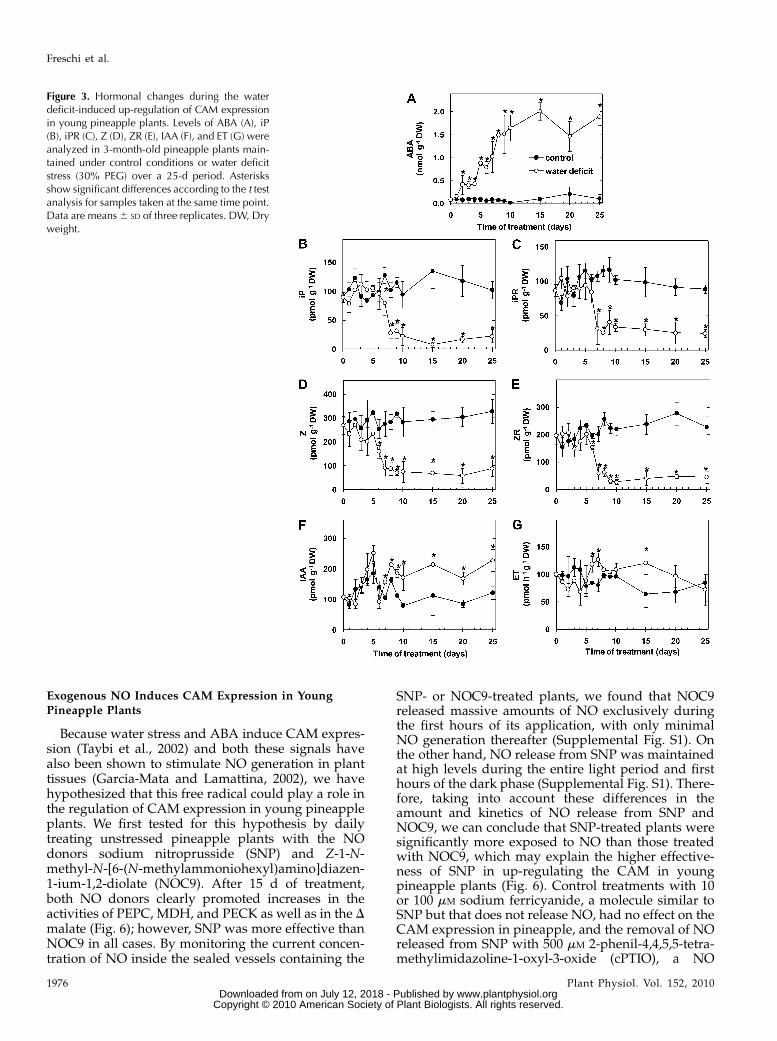
To investigate a possible role played by endogenoushormones in mediating the water deficit-inducedchanges in pineapple CAM expression, the levels ofABA, isopentenyladenine (iP), isopentenyladenine ri-boside (iPR), zeatin (Z), zeatin riboside (ZR), indolil-acetic acid (IAA), and ET were analyzed during boththe intensification and attenuation of CAM activity inresponse to changes in water availability. As shown inFigure 3A, young pineapple plants exposed to waterdeficit responded with a rapid (within 2 d) increase inthe leaf content of ABA, which preceded the up-regulation of CAM expression in this bromeliad (Fig.1, A–G). In general, the water deficit-induced rise inendogenous ABA (Fig. 3A) was inversely proportionalto the decrease in the RWC of the pineapple leaves(Fig. 1H). Within 8 d of treatment, the leaf ABA content
ABA and NO Interact during CAM Signaling
Plant Physiol. Vol. 152, 2010 1973 www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.
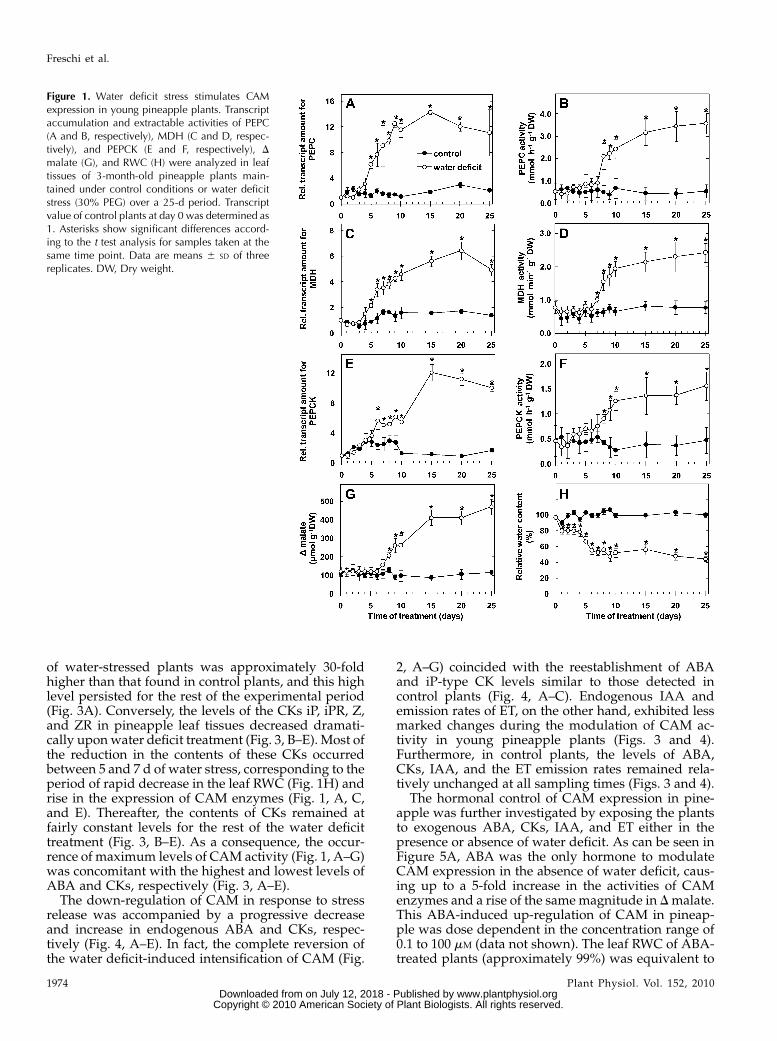
of water-stressed plants was approximately 30-foldhigher than that found in control plants, and this highlevel persisted for the rest of the experimental period(Fig. 3A). Conversely, the levels of the CKs iP, iPR, Z,and ZR in pineapple leaf tissues decreased dramati-cally uponwater deficit treatment (Fig. 3, B–E). Most ofthe reduction in the contents of these CKs occurredbetween 5 and 7 d of water stress, corresponding to theperiod of rapid decrease in the leaf RWC (Fig. 1H) andrise in the expression of CAM enzymes (Fig. 1, A, C,and E). Thereafter, the contents of CKs remained atfairly constant levels for the rest of the water deficittreatment (Fig. 3, B–E). As a consequence, the occur-rence of maximum levels of CAM activity (Fig. 1, A–G)was concomitant with the highest and lowest levels ofABA and CKs, respectively (Fig. 3, A–E).
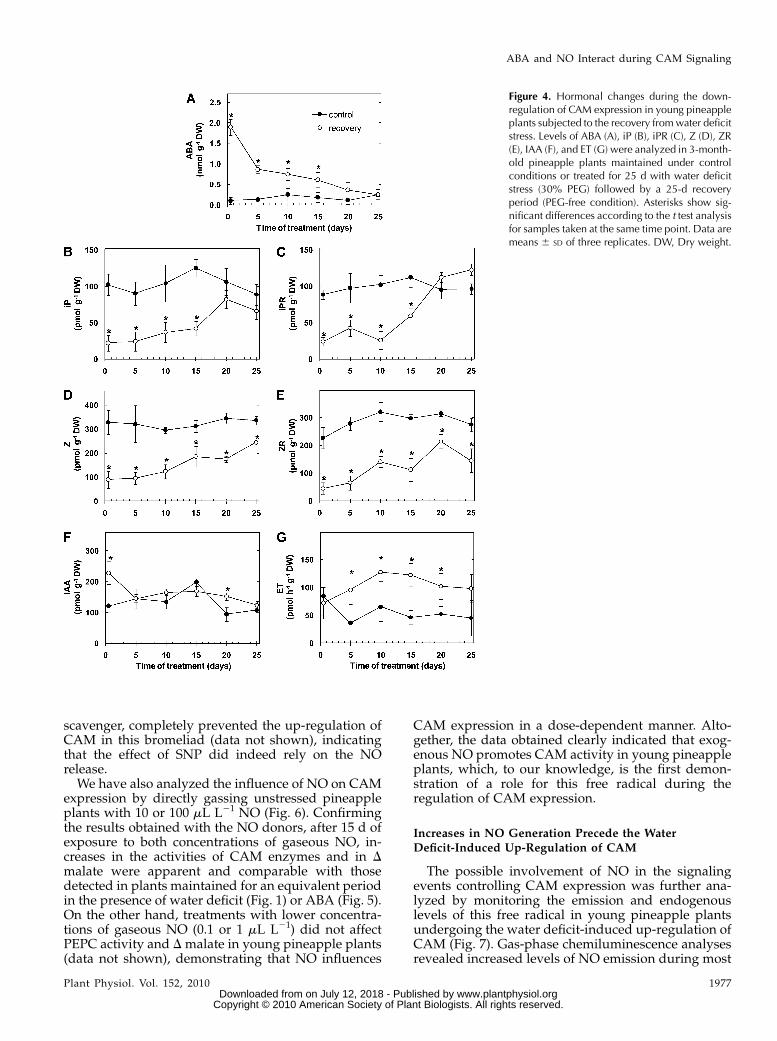
The down-regulation of CAM in response to stressrelease was accompanied by a progressive decreaseand increase in endogenous ABA and CKs, respec-tively (Fig. 4, A–E). In fact, the complete reversion ofthe water deficit-induced intensification of CAM (Fig.
2, A–G) coincided with the reestablishment of ABAand iP-type CK levels similar to those detected incontrol plants (Fig. 4, A–C). Endogenous IAA andemission rates of ET, on the other hand, exhibited lessmarked changes during the modulation of CAM ac-tivity in young pineapple plants (Figs. 3 and 4).Furthermore, in control plants, the levels of ABA,CKs, IAA, and the ET emission rates remained rela-tively unchanged at all sampling times (Figs. 3 and 4).
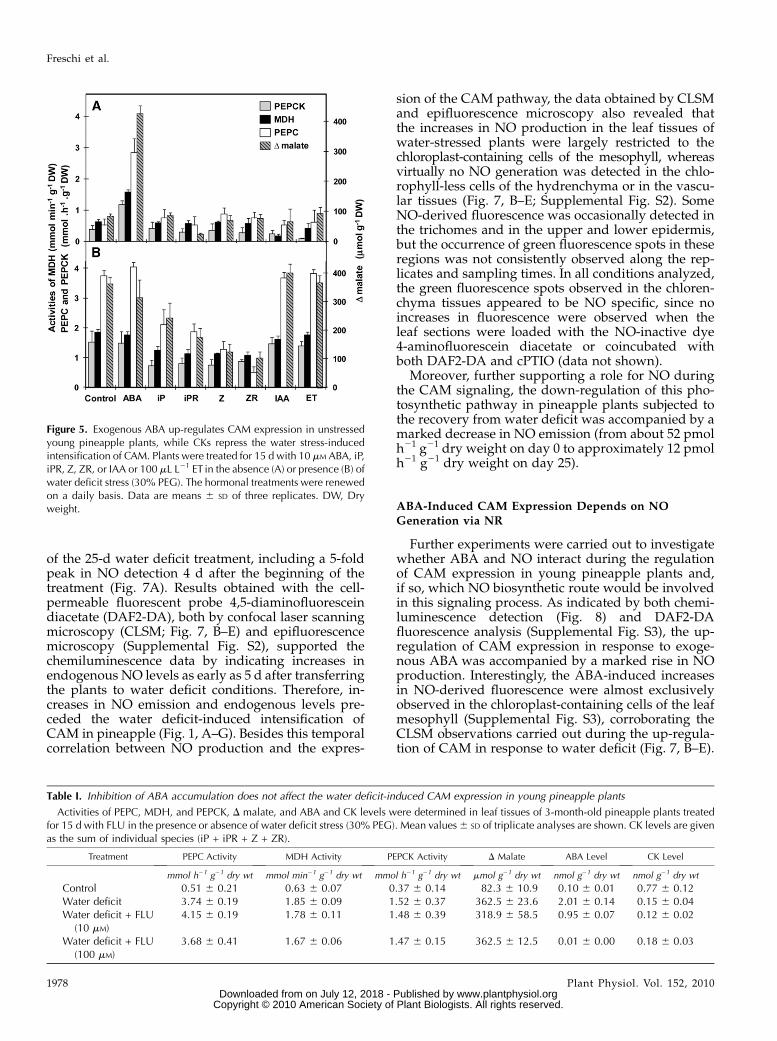
The hormonal control of CAM expression in pine-apple was further investigated by exposing the plantsto exogenous ABA, CKs, IAA, and ET either in thepresence or absence of water deficit. As can be seen inFigure 5A, ABA was the only hormone to modulateCAM expression in the absence of water deficit, caus-ing up to a 5-fold increase in the activities of CAMenzymes and a rise of the samemagnitude in Dmalate.This ABA-induced up-regulation of CAM in pineap-ple was dose dependent in the concentration range of0.1 to 100 mM (data not shown). The leaf RWC of ABA-treated plants (approximately 99%) was equivalent to
Figure 1. Water deficit stress stimulates CAMexpression in young pineapple plants. Transcriptaccumulation and extractable activities of PEPC(A and B, respectively), MDH (C and D, respec-tively), and PEPCK (E and F, respectively), D
malate (G), and RWC (H) were analyzed in leaftissues of 3-month-old pineapple plants main-tained under control conditions or water deficitstress (30% PEG) over a 25-d period. Transcriptvalue of control plants at day 0 was determined as1. Asterisks show significant differences accord-ing to the t test analysis for samples taken at thesame time point. Data are means 6 SD of threereplicates. DW, Dry weight.
Freschi et al.
1974 Plant Physiol. Vol. 152, 2010 www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.
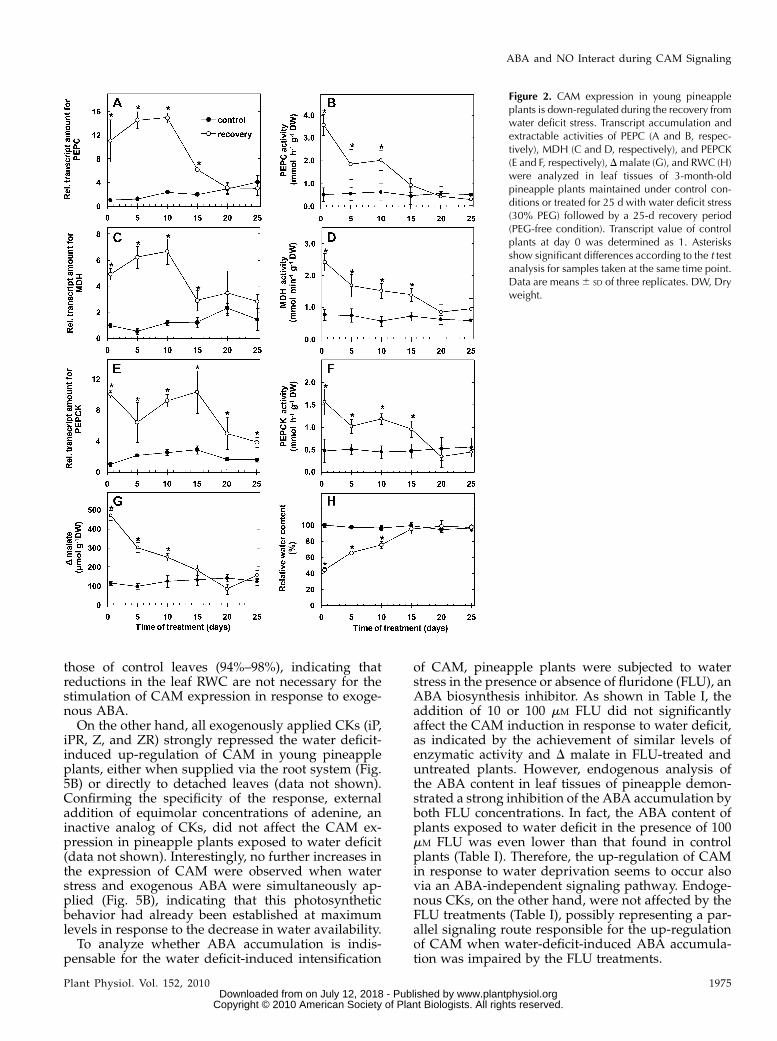
those of control leaves (94%–98%), indicating thatreductions in the leaf RWC are not necessary for thestimulation of CAM expression in response to exoge-nous ABA.On the other hand, all exogenously applied CKs (iP,
iPR, Z, and ZR) strongly repressed the water deficit-induced up-regulation of CAM in young pineappleplants, either when supplied via the root system (Fig.5B) or directly to detached leaves (data not shown).Confirming the specificity of the response, externaladdition of equimolar concentrations of adenine, aninactive analog of CKs, did not affect the CAM ex-pression in pineapple plants exposed to water deficit(data not shown). Interestingly, no further increases inthe expression of CAM were observed when waterstress and exogenous ABA were simultaneously ap-plied (Fig. 5B), indicating that this photosyntheticbehavior had already been established at maximumlevels in response to the decrease in water availability.To analyze whether ABA accumulation is indis-
pensable for the water deficit-induced intensification
of CAM, pineapple plants were subjected to waterstress in the presence or absence of fluridone (FLU), anABA biosynthesis inhibitor. As shown in Table I, theaddition of 10 or 100 mM FLU did not significantlyaffect the CAM induction in response to water deficit,as indicated by the achievement of similar levels ofenzymatic activity and D malate in FLU-treated anduntreated plants. However, endogenous analysis ofthe ABA content in leaf tissues of pineapple demon-strated a strong inhibition of the ABA accumulation byboth FLU concentrations. In fact, the ABA content ofplants exposed to water deficit in the presence of 100mM FLU was even lower than that found in controlplants (Table I). Therefore, the up-regulation of CAMin response to water deprivation seems to occur alsovia an ABA-independent signaling pathway. Endoge-nous CKs, on the other hand, were not affected by theFLU treatments (Table I), possibly representing a par-allel signaling route responsible for the up-regulationof CAM when water-deficit-induced ABA accumula-tion was impaired by the FLU treatments.
Figure 2. CAM expression in young pineappleplants is down-regulated during the recovery fromwater deficit stress. Transcript accumulation andextractable activities of PEPC (A and B, respec-tively), MDH (C and D, respectively), and PEPCK(E and F, respectively), Dmalate (G), and RWC (H)were analyzed in leaf tissues of 3-month-oldpineapple plants maintained under control con-ditions or treated for 25 d with water deficit stress(30% PEG) followed by a 25-d recovery period(PEG-free condition). Transcript value of controlplants at day 0 was determined as 1. Asterisksshow significant differences according to the t testanalysis for samples taken at the same time point.Data are means6 SD of three replicates. DW, Dryweight.
ABA and NO Interact during CAM Signaling
Plant Physiol. Vol. 152, 2010 1975 www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.
Exogenous NO Induces CAM Expression in YoungPineapple Plants
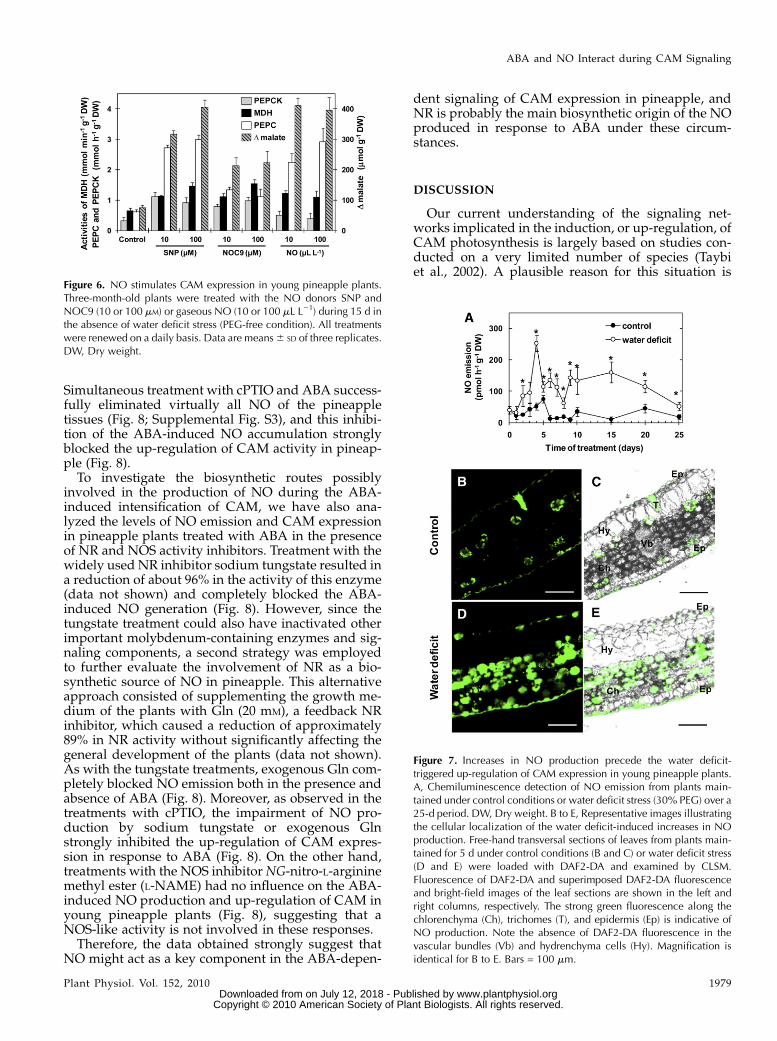
Because water stress and ABA induce CAM expres-sion (Taybi et al., 2002) and both these signals havealso been shown to stimulate NO generation in planttissues (Garcia-Mata and Lamattina, 2002), we havehypothesized that this free radical could play a role inthe regulation of CAM expression in young pineappleplants. We first tested for this hypothesis by dailytreating unstressed pineapple plants with the NOdonors sodium nitroprusside (SNP) and Z-1-N-methyl-N-[6-(N-methylammoniohexyl)amino]diazen-1-ium-1,2-diolate (NOC9). After 15 d of treatment,both NO donors clearly promoted increases in theactivities of PEPC, MDH, and PECK as well as in the Dmalate (Fig. 6); however, SNP was more effective thanNOC9 in all cases. By monitoring the current concen-tration of NO inside the sealed vessels containing the
SNP- or NOC9-treated plants, we found that NOC9released massive amounts of NO exclusively duringthe first hours of its application, with only minimalNO generation thereafter (Supplemental Fig. S1). Onthe other hand, NO release from SNP was maintainedat high levels during the entire light period and firsthours of the dark phase (Supplemental Fig. S1). There-fore, taking into account these differences in theamount and kinetics of NO release from SNP andNOC9, we can conclude that SNP-treated plants weresignificantly more exposed to NO than those treatedwith NOC9, which may explain the higher effective-ness of SNP in up-regulating the CAM in youngpineapple plants (Fig. 6). Control treatments with 10or 100 mM sodium ferricyanide, a molecule similar toSNP but that does not release NO, had no effect on theCAM expression in pineapple, and the removal of NOreleased from SNP with 500 mM 2-phenil-4,4,5,5-tetra-methylimidazoline-1-oxyl-3-oxide (cPTIO), a NO
Figure 3. Hormonal changes during the waterdeficit-induced up-regulation of CAM expressionin young pineapple plants. Levels of ABA (A), iP(B), iPR (C), Z (D), ZR (E), IAA (F), and ET (G) wereanalyzed in 3-month-old pineapple plants main-tained under control conditions or water deficitstress (30% PEG) over a 25-d period. Asterisksshow significant differences according to the t testanalysis for samples taken at the same time point.Data are means6 SD of three replicates. DW, Dryweight.
Freschi et al.
1976 Plant Physiol. Vol. 152, 2010 www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.
scavenger, completely prevented the up-regulation ofCAM in this bromeliad (data not shown), indicatingthat the effect of SNP did indeed rely on the NOrelease.We have also analyzed the influence of NO on CAM
expression by directly gassing unstressed pineappleplants with 10 or 100 mL L21 NO (Fig. 6). Confirmingthe results obtained with the NO donors, after 15 d ofexposure to both concentrations of gaseous NO, in-creases in the activities of CAM enzymes and in Dmalate were apparent and comparable with thosedetected in plants maintained for an equivalent periodin the presence of water deficit (Fig. 1) or ABA (Fig. 5).On the other hand, treatments with lower concentra-tions of gaseous NO (0.1 or 1 mL L21) did not affectPEPC activity and Dmalate in young pineapple plants(data not shown), demonstrating that NO influences
CAM expression in a dose-dependent manner. Alto-gether, the data obtained clearly indicated that exog-enous NO promotes CAM activity in young pineappleplants, which, to our knowledge, is the first demon-stration of a role for this free radical during theregulation of CAM expression.
Increases in NO Generation Precede the WaterDeficit-Induced Up-Regulation of CAM
The possible involvement of NO in the signalingevents controlling CAM expression was further ana-lyzed by monitoring the emission and endogenouslevels of this free radical in young pineapple plantsundergoing the water deficit-induced up-regulation ofCAM (Fig. 7). Gas-phase chemiluminescence analysesrevealed increased levels of NO emission during most
Figure 4. Hormonal changes during the down-regulation of CAM expression in young pineappleplants subjected to the recovery fromwater deficitstress. Levels of ABA (A), iP (B), iPR (C), Z (D), ZR(E), IAA (F), and ET (G) were analyzed in 3-month-old pineapple plants maintained under controlconditions or treated for 25 d with water deficitstress (30% PEG) followed by a 25-d recoveryperiod (PEG-free condition). Asterisks show sig-nificant differences according to the t test analysisfor samples taken at the same time point. Data aremeans 6 SD of three replicates. DW, Dry weight.
ABA and NO Interact during CAM Signaling
Plant Physiol. Vol. 152, 2010 1977 www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.
of the 25-d water deficit treatment, including a 5-foldpeak in NO detection 4 d after the beginning of thetreatment (Fig. 7A). Results obtained with the cell-permeable fluorescent probe 4,5-diaminofluoresceindiacetate (DAF2-DA), both by confocal laser scanningmicroscopy (CLSM; Fig. 7, B–E) and epifluorescencemicroscopy (Supplemental Fig. S2), supported thechemiluminescence data by indicating increases inendogenous NO levels as early as 5 d after transferringthe plants to water deficit conditions. Therefore, in-creases in NO emission and endogenous levels pre-ceded the water deficit-induced intensification ofCAM in pineapple (Fig. 1, A–G). Besides this temporalcorrelation between NO production and the expres-
sion of the CAM pathway, the data obtained by CLSMand epifluorescence microscopy also revealed thatthe increases in NO production in the leaf tissues ofwater-stressed plants were largely restricted to thechloroplast-containing cells of the mesophyll, whereasvirtually no NO generation was detected in the chlo-rophyll-less cells of the hydrenchyma or in the vascu-lar tissues (Fig. 7, B–E; Supplemental Fig. S2). SomeNO-derived fluorescence was occasionally detected inthe trichomes and in the upper and lower epidermis,but the occurrence of green fluorescence spots in theseregions was not consistently observed along the rep-licates and sampling times. In all conditions analyzed,the green fluorescence spots observed in the chloren-chyma tissues appeared to be NO specific, since noincreases in fluorescence were observed when theleaf sections were loaded with the NO-inactive dye4-aminofluorescein diacetate or coincubated withboth DAF2-DA and cPTIO (data not shown).
Moreover, further supporting a role for NO duringthe CAM signaling, the down-regulation of this pho-tosynthetic pathway in pineapple plants subjected tothe recovery from water deficit was accompanied by amarked decrease in NO emission (from about 52 pmolh21 g21 dry weight on day 0 to approximately 12 pmolh21 g21 dry weight on day 25).
ABA-Induced CAM Expression Depends on NOGeneration via NR
Further experiments were carried out to investigatewhether ABA and NO interact during the regulationof CAM expression in young pineapple plants and,if so, which NO biosynthetic route would be involvedin this signaling process. As indicated by both chemi-luminescence detection (Fig. 8) and DAF2-DAfluorescence analysis (Supplemental Fig. S3), the up-regulation of CAM expression in response to exoge-nous ABA was accompanied by a marked rise in NOproduction. Interestingly, the ABA-induced increasesin NO-derived fluorescence were almost exclusivelyobserved in the chloroplast-containing cells of the leafmesophyll (Supplemental Fig. S3), corroborating theCLSM observations carried out during the up-regula-tion of CAM in response to water deficit (Fig. 7, B–E).
Figure 5. Exogenous ABA up-regulates CAM expression in unstressedyoung pineapple plants, while CKs repress the water stress-inducedintensification of CAM. Plants were treated for 15 d with 10 mM ABA, iP,iPR, Z, ZR, or IAA or 100 mL L21 ET in the absence (A) or presence (B) ofwater deficit stress (30% PEG). The hormonal treatments were renewedon a daily basis. Data are means 6 SD of three replicates. DW, Dryweight.
Table I. Inhibition of ABA accumulation does not affect the water deficit-induced CAM expression in young pineapple plants
Activities of PEPC, MDH, and PEPCK, D malate, and ABA and CK levels were determined in leaf tissues of 3-month-old pineapple plants treatedfor 15 d with FLU in the presence or absence of water deficit stress (30% PEG). Mean values6 SD of triplicate analyses are shown. CK levels are givenas the sum of individual species (iP + iPR + Z + ZR).
Treatment PEPC Activity MDH Activity PEPCK Activity D Malate ABA Level CK Level
mmol h21 g21 dry wt mmol min21 g21 dry wt mmol h21 g21 dry wt mmol g21 dry wt nmol g21 dry wt nmol g21 dry wt
Control 0.51 6 0.21 0.63 6 0.07 0.37 6 0.14 82.3 6 10.9 0.10 6 0.01 0.77 6 0.12Water deficit 3.74 6 0.19 1.85 6 0.09 1.52 6 0.37 362.5 6 23.6 2.01 6 0.14 0.15 6 0.04Water deficit + FLU
(10 mM)4.15 6 0.19 1.78 6 0.11 1.48 6 0.39 318.9 6 58.5 0.95 6 0.07 0.12 6 0.02
Water deficit + FLU(100 mM)
3.68 6 0.41 1.67 6 0.06 1.47 6 0.15 362.5 6 12.5 0.01 6 0.00 0.18 6 0.03
Freschi et al.
1978 Plant Physiol. Vol. 152, 2010 www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.
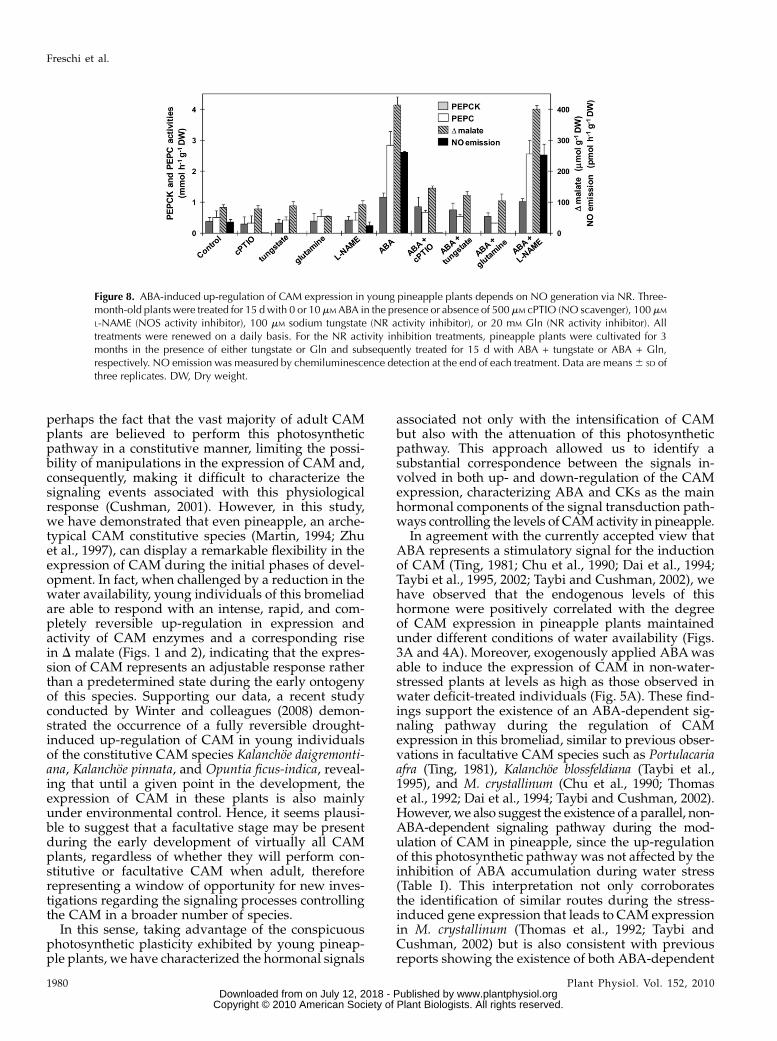
Simultaneous treatment with cPTIO and ABA success-fully eliminated virtually all NO of the pineappletissues (Fig. 8; Supplemental Fig. S3), and this inhibi-tion of the ABA-induced NO accumulation stronglyblocked the up-regulation of CAM activity in pineap-ple (Fig. 8).To investigate the biosynthetic routes possibly
involved in the production of NO during the ABA-induced intensification of CAM, we have also ana-lyzed the levels of NO emission and CAM expressionin pineapple plants treated with ABA in the presenceof NR and NOS activity inhibitors. Treatment with thewidely used NR inhibitor sodium tungstate resulted ina reduction of about 96% in the activity of this enzyme(data not shown) and completely blocked the ABA-induced NO generation (Fig. 8). However, since thetungstate treatment could also have inactivated otherimportant molybdenum-containing enzymes and sig-naling components, a second strategy was employedto further evaluate the involvement of NR as a bio-synthetic source of NO in pineapple. This alternativeapproach consisted of supplementing the growth me-dium of the plants with Gln (20 mM), a feedback NRinhibitor, which caused a reduction of approximately89% in NR activity without significantly affecting thegeneral development of the plants (data not shown).As with the tungstate treatments, exogenous Gln com-pletely blocked NO emission both in the presence andabsence of ABA (Fig. 8). Moreover, as observed in thetreatments with cPTIO, the impairment of NO pro-duction by sodium tungstate or exogenous Glnstrongly inhibited the up-regulation of CAM expres-sion in response to ABA (Fig. 8). On the other hand,treatments with the NOS inhibitorNG-nitro-L-argininemethyl ester (L-NAME) had no influence on the ABA-induced NO production and up-regulation of CAM inyoung pineapple plants (Fig. 8), suggesting that aNOS-like activity is not involved in these responses.Therefore, the data obtained strongly suggest that
NO might act as a key component in the ABA-depen-
dent signaling of CAM expression in pineapple, andNR is probably the main biosynthetic origin of the NOproduced in response to ABA under these circum-stances.
DISCUSSION
Our current understanding of the signaling net-works implicated in the induction, or up-regulation, ofCAM photosynthesis is largely based on studies con-ducted on a very limited number of species (Taybiet al., 2002). A plausible reason for this situation is
Figure 6. NO stimulates CAM expression in young pineapple plants.Three-month-old plants were treated with the NO donors SNP andNOC9 (10 or 100 mM) or gaseous NO (10 or 100 mL L21) during 15 d inthe absence of water deficit stress (PEG-free condition). All treatmentswere renewed on a daily basis. Data are means6 SD of three replicates.DW, Dry weight.
Figure 7. Increases in NO production precede the water deficit-triggered up-regulation of CAM expression in young pineapple plants.A, Chemiluminescence detection of NO emission from plants main-tained under control conditions or water deficit stress (30% PEG) over a25-d period. DW, Dry weight. B to E, Representative images illustratingthe cellular localization of the water deficit-induced increases in NOproduction. Free-hand transversal sections of leaves from plants main-tained for 5 d under control conditions (B and C) or water deficit stress(D and E) were loaded with DAF2-DA and examined by CLSM.Fluorescence of DAF2-DA and superimposed DAF2-DA fluorescenceand bright-field images of the leaf sections are shown in the left andright columns, respectively. The strong green fluorescence along thechlorenchyma (Ch), trichomes (T), and epidermis (Ep) is indicative ofNO production. Note the absence of DAF2-DA fluorescence in thevascular bundles (Vb) and hydrenchyma cells (Hy). Magnification isidentical for B to E. Bars = 100 mm.
ABA and NO Interact during CAM Signaling
Plant Physiol. Vol. 152, 2010 1979 www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.
perhaps the fact that the vast majority of adult CAMplants are believed to perform this photosyntheticpathway in a constitutive manner, limiting the possi-bility of manipulations in the expression of CAM and,consequently, making it difficult to characterize thesignaling events associated with this physiologicalresponse (Cushman, 2001). However, in this study,we have demonstrated that even pineapple, an arche-typical CAM constitutive species (Martin, 1994; Zhuet al., 1997), can display a remarkable flexibility in theexpression of CAM during the initial phases of devel-opment. In fact, when challenged by a reduction in thewater availability, young individuals of this bromeliadare able to respond with an intense, rapid, and com-pletely reversible up-regulation in expression andactivity of CAM enzymes and a corresponding risein D malate (Figs. 1 and 2), indicating that the expres-sion of CAM represents an adjustable response ratherthan a predetermined state during the early ontogenyof this species. Supporting our data, a recent studyconducted by Winter and colleagues (2008) demon-strated the occurrence of a fully reversible drought-induced up-regulation of CAM in young individualsof the constitutive CAM species Kalanchoe daigremonti-ana, Kalanchoe pinnata, and Opuntia ficus-indica, reveal-ing that until a given point in the development, theexpression of CAM in these plants is also mainlyunder environmental control. Hence, it seems plausi-ble to suggest that a facultative stage may be presentduring the early development of virtually all CAMplants, regardless of whether they will perform con-stitutive or facultative CAM when adult, thereforerepresenting a window of opportunity for new inves-tigations regarding the signaling processes controllingthe CAM in a broader number of species.
In this sense, taking advantage of the conspicuousphotosynthetic plasticity exhibited by young pineap-ple plants, we have characterized the hormonal signals
associated not only with the intensification of CAMbut also with the attenuation of this photosyntheticpathway. This approach allowed us to identify asubstantial correspondence between the signals in-volved in both up- and down-regulation of the CAMexpression, characterizing ABA and CKs as the mainhormonal components of the signal transduction path-ways controlling the levels of CAM activity in pineapple.
In agreement with the currently accepted view thatABA represents a stimulatory signal for the inductionof CAM (Ting, 1981; Chu et al., 1990; Dai et al., 1994;Taybi et al., 1995, 2002; Taybi and Cushman, 2002), wehave observed that the endogenous levels of thishormone were positively correlated with the degreeof CAM expression in pineapple plants maintainedunder different conditions of water availability (Figs.3A and 4A). Moreover, exogenously applied ABAwasable to induce the expression of CAM in non-water-stressed plants at levels as high as those observed inwater deficit-treated individuals (Fig. 5A). These find-ings support the existence of an ABA-dependent sig-naling pathway during the regulation of CAMexpression in this bromeliad, similar to previous obser-vations in facultative CAM species such as Portulacariaafra (Ting, 1981), Kalanchoe blossfeldiana (Taybi et al.,1995), and M. crystallinum (Chu et al., 1990; Thomaset al., 1992; Dai et al., 1994; Taybi and Cushman, 2002).However, we also suggest the existence of a parallel, non-ABA-dependent signaling pathway during the mod-ulation of CAM in pineapple, since the up-regulationof this photosynthetic pathway was not affected by theinhibition of ABA accumulation during water stress(Table I). This interpretation not only corroboratesthe identification of similar routes during the stress-induced gene expression that leads to CAM expressionin M. crystallinum (Thomas et al., 1992; Taybi andCushman, 2002) but is also consistent with previousreports showing the existence of both ABA-dependent
Figure 8. ABA-induced up-regulation of CAM expression in young pineapple plants depends on NO generation via NR. Three-month-old plants were treated for 15 dwith 0 or 10 mM ABA in the presence or absence of 500 mM cPTIO (NO scavenger), 100 mM
L-NAME (NOS activity inhibitor), 100 mM sodium tungstate (NR activity inhibitor), or 20 mM Gln (NR activity inhibitor). Alltreatments were renewed on a daily basis. For the NR activity inhibition treatments, pineapple plants were cultivated for 3months in the presence of either tungstate or Gln and subsequently treated for 15 d with ABA + tungstate or ABA + Gln,respectively. NO emission was measured by chemiluminescence detection at the end of each treatment. Data are means6 SD ofthree replicates. DW, Dry weight.
Freschi et al.
1980 Plant Physiol. Vol. 152, 2010 www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

and -independent pathways during the general sig-naling transduction of environmental stresses such ascold, drought, and salinity (Ishitani et al., 1997; Bellaireet al., 2000).CKs were also shown to have an important and very
clear influence on the modulation of CAM in pineap-ple; however, contrary to the endogenous patternobserved for ABA, the leaf content of CKs in plantsexposed to different conditions of water availabilitywas negatively correlated with the degree of CAMexpression (Figs. 3 and 4), suggesting that the internalpool of these hormones could contribute to maintainlow levels of CAM in young individuals of this bro-meliad. In agreement with this suggestion, our dataalso showed that, under water stress, the up-regulationof genes encoding key enzymes of the CAM path-way coincided specifically with the period of majordecrease in the leaf CK concentration (Fig. 3, B–E).Therefore, the reduction in the leaf content of CKstriggered by limitations in water availability mayrepresent an important signal for derepressing CAMexpression in young pineapple plants. This view wasalso supported by the fact that exogenous CKs wereable to strongly repress the water deficit-inducedCAM expression either when supplied to the roots(Fig. 5B) or to detached leaves of this bromeliad.Likewise, a negative regulatory role of CKs has alsobeen demonstrated to occur during both salt- anddrought-induced expression of CAM in M. crystalli-num (Schmitt and Piepenbrock, 1992; Peters et al.,1997); however, in this species, exogenous CKs onlyacted as repressors of CAM when applied directly tothe leaves, having an opposite effect when suppliedvia the root system (Schmitt and Piepenbrock, 1992;Dai et al., 1994; Peters et al., 1997). Based on thisscenario, a gradual decrease in endogenous CKsthroughout leaf development could possibly accountfor a higher responsiveness of mature leaf tissues toenvironmental signals that stimulate CAM expression;however, whether this relationship occurs in faculta-tive or constitutive species is unknown at present.Moreover, the mechanisms by which developmentallyor environmentally triggered changes in hormonalbalance orchestrate the complex array of biochemicaland physiological modifications required for theC3-CAM transition also remain largely unresolved.Although some second messengers responsible for
transducing the hormonal stimuli into changes inCAM expression have already been described for M.crystallinum (Taybi and Cushman, 1999, 2002), ourunderstanding of these downstream signaling eventsis still considerably fragmented (Taybi et al., 2002). Infact, cytosolic Ca2+, inositol 1,4,5-triphosphate, H2O2,and protein kinases and phosphatases are, to the bestof our knowledge, the only secondmessengers alreadyidentified during the signal transduction cascadesleading to CAM expression (Taybi and Cushman,1999, 2002; Taybi et al., 2002). Here, we present toour knowledge the first evidence showing that the freeradical NO may also play an important role in the
CAM modulation, representing a positive signal forthe establishment of this specialized mode of photo-synthesis. Exogenous NO, applied as donors or gas-eous mixtures, was shown to be sufficient to induceincreases in the activities of CAM enzymes and Dmalate in non-water-stressed pineapple plants (Fig. 6),these responses being at comparable levels to thosedetected in plants subjected to water deficit or ABAtreatment (Fig. 5). In agreement with these pharmaco-logical data, we have also demonstrated that the up-regulation of CAM expression in pineapple plantsunder water stress was accompanied by increases inboth NO emission and endogenous levels (Fig. 7;Supplemental Fig. S2), while the down-regulation ofthis photosynthetic pathway coincided with a corre-sponding decrease in NO emission. Therefore, theseresults seem to indicate that increased NO productionand/or accumulation in young pineapple plants couldrepresent a signal to induce and, subsequently, main-tain high levels of CAM expression under water stressconditions.
Supporting our findings, water deficit has beenshown to increase NO synthesis and/or accumulationin tissues of numerous plant species (Leshem andHaramaty, 1996; Zhao et al., 2001; Garcia-Mata andLamattina, 2002; Gould et al., 2003; Hao et al., 2008).Moreover, NO seems to be implicated in the inductionof important protective responses to cope with waterstress, including the reduction in the transpirationalwater loss due to stomatal closure (Garcia-Mata andLamattina, 2001, 2002; Neill et al., 2002; Bright et al.,2006) and the activation of antioxidant defenses tocombat oxidative stress (Zhang et al., 2007; Hao et al.,2008). In agreement, NO supplementation has consis-tently been shown to confer drought tolerance to C3andC4 species, favoring themaintenance of higher RWCand minimizing oxidative damage in leaf tissues ofwater-stressed plants (Garcia-Mata and Lamattina, 2002;Bright et al., 2006; Tian and Lei, 2006; Hao et al., 2008).
In fact, a close relationship between NO and oxida-tive stress has been consistently reported in the liter-ature (Neill et al., 2002, 2008; Hao et al., 2008). Forinstance, reactive oxygen species such as superoxideanions can readily react with NO and potentiallydecrease the concentration of NO available for signal-ing, whereas other reactive oxygen species such asH2O2 may lead to increased NO levels by stimulatingthe activity of NO-generating enzymes (Neill et al.,2002, 2008). Therefore, it is interesting to contextualizeour results in the face of previous studies that haveindicated a possible signaling role for oxidative stresson the C3-CAM transition in M. crystallinum. In one ofthese studies, Slesak et al. (2003) demonstrated thatconditions that typically result in an oxidative burdenin plant cells, such as excessive light, photosyntheticelectron transport inhibitors, and exogenous H2O2, canstimulate CAM expression in this species. Moreover,treatments with ozone also resulted in the up-regulationof important metabolic components of the CAM path-way in M. crystallinum (Niewiadomska et al., 2002;
ABA and NO Interact during CAM Signaling
Plant Physiol. Vol. 152, 2010 1981 www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

Borland et al., 2006), even though this gaseous oxidantfailed to induce functional CAM expression, as evi-denced by the lack of nocturnal acidity accumulationin ozone-treated individuals of this species (Borlandet al., 2006). Although a discrete up-regulation ofCAM has been observed in H2O2-treated young pine-apple plants (data not shown), to date, NO was theonly redox-related molecule that, when exogenouslyapplied, induced full CAM expression in non-water-stressed individuals, thereby suggesting a specificsignaling role for this nitrogen-reactive species onthe control of CAM expression in pineapple.
Accumulating evidence has also demonstrated thatNO and ABA closely interact during the signalingprocesses that underpin the induction of plant adap-tive responses to environmental stresses such asdrought (Zhao et al., 2001; Garcia-Mata and Lamattina,2002; Xing et al., 2004; Bright et al., 2006; Neill et al.,2008; Hao et al., 2008), salinity (Ruan et al., 2004),temperature (Song et al., 2008), and UV-B radiation(Tossi et al., 2009). Although some influence of NO onthe endogenous ABA levels has been reported (Zhaoet al., 2001; Xing et al., 2004), most of the availableinformation suggests that this free radical acts as adownstreammessenger in the ABA signaling cascades(Garcia-Mata and Lamattina, 2001; Bright et al., 2006;Hao et al., 2008; Song et al., 2008; Tossi et al., 2009).Interestingly, data obtained in this study firmly indi-cated that NO is also involved in the ABA-induced up-regulation of CAM in pineapple, probably acting as asecond messenger of this hormone. As evidence of thisinteraction, increases in both emission and endoge-nous levels of NO were clearly observed during theup-regulation of CAM in non-water-stressed plantstreated with exogenous ABA (Fig. 8; SupplementalFig. S3). Moreover, removal of NO by cPTIO almostcompletely blocked the ABA-triggered CAM expres-sion in this bromeliad (Fig. 8).
Further supporting the idea that NO acts down-stream of ABA to trigger CAM expression in youngpineapple plants, ABA-induced up-regulation ofCAM was strongly reduced when NO generationwas impaired due to the inhibition of NR activityeither by tungstate or Gln treatment (Fig. 8). Actually,NR was identified in this work as the most likelybiosynthetic origin of NO during the up-regulation ofCAM expression triggered by ABA (Fig. 8) or waterstress (data not shown), since the inhibition of thisenzymatic activity completely abolished NO produc-tion in all analyzed conditions. On the other hand, noevidence was obtained in support of a role for a NOS-like activity as a source of NO during the signaling ofCAM up-regulation in pineapple (Fig. 8). In compar-ison, plants under water stress have been shown toproduce NO via NR, NOS-like activity, or a combina-tion of these two biosynthetic routes (Bright et al.,2006; Hao et al., 2008; Neill et al., 2008; Sang et al.,2008).
Finally, although the CAM induction involves sub-stantial changes in the general metabolism of the plant,
most of the physiological, biochemical, and gene ex-pression reprogramming required for the establish-ment of the CAM pathway is believed to be restrictedto the chlorophyll-containing cells of the leaf meso-phyll (i.e. chlorenchyma). In fact, it is in these chloro-plast-containing cells that the gene expression,transduction, and activity of most regulatory proteinsof the CAM, special transporters, enzymes responsiblefor the nocturnal fixation of CO2 (PEPC), synthesis oforganic acids (MDH and enolases), and daytime de-carboxylation of these acids (PEPCK and malic en-zyme) occur (Springer and Outlaw, 1988; Borlandet al., 1998; Cushman and Bohnert, 1999). Therefore,since the increases in NO generation triggered eitherby water deficit (Fig. 7; Supplemental Fig. S2) orexogenous ABA (Supplemental Fig. S3) were virtuallyconfined to the chlorenchyma of pineapple leaves, wecan conclude that there was also a spatial coincidencebetween the cellular localization of NO and the ex-pression of CAM within the leaf mesophyll of youngpineapple plants, thereby reinforcing a possible role ofthis free radical in the CAM signaling processes.Moreover, accumulating evidence has indicated thatNO can also interact with thiol groups present inproteins to form S-nitrosothiols (Lindermayr et al.,2005), which are currently believed to play a role asstable NO carriers for both short- and long-distancetransport in plants (Arasimowicz and Floryszak-Wieczorek, 2007). Therefore, it would be interestingto determine whether the NO detected inside thechlorenchyma cells of water-stressed pineapple plantswas synthesized directly in these cells or resulted fromthe decomposition of NO derivatives (i.e. S-nitroso-thiols) translocated from other plant tissues even morerapidly affected by the water stress stimulus, such asthe roots.
In conclusion, here we have provided evidence thatABA, CKs, and NO may represent important pieces ofthe complex signaling puzzle regulating CAM expres-sion in pineapple plants. Two parallel signaling routesbased on hormones with opposite effects on the reg-ulation of CAM (i.e. ABA and CKs) seem to provide aneffective fine-tuning between the expression of thisphotosynthetic behavior and the environmental con-ditions endured by young individuals of this brome-liad. Moreover, we have shown, to our knowledge forthe first time, that NO plays an important role duringthe signaling of CAM expression, acting as a keydownstream component in the ABA-dependent sig-naling pathway. Finally, our data clearly indicated thatNR is primarily responsible for NO formation inpineapple plants undergoing the up-regulation ofCAM triggered either by ABA or water deficit.
MATERIALS AND METHODS
Plant Material and Growth Conditions
Pineapple (Ananas comosus ‘Smooth Cayenne’) plants were obtained by in
vitro vegetative multiplication according to Nievola et al. (2005). Throughout
Freschi et al.
1982 Plant Physiol. Vol. 152, 2010 www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

the experiments, and for the preceding 2 weeks of acclimation, pineapple
plants were maintained in a growth chamber at 250 mmol m22 s21 with a 12-h
photoperiod and air temperature of 25�C 6 2�C.
Water Deficit and Recovery Treatments
To impose water deficit, 3-month-old pineapple plants (for growth stage
characterization, see Supplemental Fig. S4) were transferred to liquidMurashige
and Skoog (MS) medium (Murashige and Skoog, 1962) containing 30% (w/v)
PEG 6000 for 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, and 25 d. Recovery treatments
were performed by returning plants that had been exposed to PEG for 25 d to a
newMS basal medium for a further 0, 5, 10, 15, 20, and 25 d. Control plants for
both treatments were transferred to fresh MS basal medium for the same time
intervals. After each treatment, leaf samples were collected 1 h before the
onset of illumination, weighed, frozen in liquid nitrogen, and stored at280�Cuntil used in the biochemical and gene expression analyses. For the determi-
nation of Dmalate, additional leaf samples were collected 1 h before the end of
the light period. Each experimental treatment was performed in three repli-
cates. For each replicate, fully expanded leaves of approximately 100 plants
were harvested, cut into small pieces, and thoroughly mixed before taking the
aliquots for the analytical procedures.
Hormonal and NO Treatments
To assess the effects of plant hormones on CAM expression, pineapple
plants were transferred to a newMS liquid growth medium in the presence of
ABA (10 mM), CKs (Z, ZR, iP, or iPR; 10 mM), IAA (10 mM), ET (100 mL L21), or
FLU (10 or 100 mM) either with or without 30% PEG. In parallel, the influence
of exogenous NO on the expression of CAM was investigated by treating the
plants with the NO donors SNP or NOC9 (10 or 100 mM) or gaseous NO (10 or
100 mL L21). As a control for the NO donor treatments, 500 mM cPTIO was
added together with SNP or NOC9. Sodium ferrocyanide (10 or 100 mM) was
used as an additional control for SNP. Treatments with ABA (10 mM) combined
with cPTIO (500 mM), L-NAME (100 mM), Gln (20 mM), or sodium tungstate (100
mM) were also carried out. In all cases, the chemicals and the growth medium
were renewed on a daily basis for a 15-d period. Control plants were
transferred to a fresh growth medium for the same time intervals. All
treatments were carried out with 3-month-old plants grown in MS medium,
except for the ABA plus tungstate or ABA plus Gln treatments, which were
performed with plants cultivated for 3 months in a modified MS medium
containing tungstate (100 mM) or Glu (20 mM), respectively. After each
treatment, leaf samples were collected 1 h before the onset of illumination
and 1 h before the end of the light period, weighed, frozen in liquid nitrogen,
and stored at280�C until used in the biochemical analyses. Each experimental
treatment was performed at least in three replicates. For each replicate, fully
expanded leaves of approximately 100 plants were harvested, cut into small
pieces, and thoroughly mixed before taking the aliquots for the analytical
procedures.
RWC Determination
RWC, expressed as a percentage, was determined in pineapple leaves
according to the following formula: RWC (%) = 100 3 [(fresh mass – dry
mass)/(saturated mass – dry mass)]. Fresh mass was measured just after the
end of the experiments, and saturated mass was determined after incubation
of the leaves in distilled water for 6 h at room temperature. Dry mass was
measured after drying the samples at 60�C for at least 72 h.
Malate Determination
Concentrations of L-malate in pineapple leaf samples were determined
spectrophotometrically according to the method of Hohorst (1965), with the
modifications described by Freschi et al. (2009). The recovery of malate
through this procedure was over 75%.
Enzyme Activities
PEPC (EC 4.1.1.31) and MDH (EC 1.1.1.37) extraction and assay were
carried out as described by Freschi et al. (2010). PEPCK (EC 4.1.1.38) activity
was determined according to the procedure of Borland et al. (1998), with
modifications. Frozen leaf samples were ground to a fine powder, extracted in
5 volumes (v/w) of buffer containing 500 mM Bicine (pH 10.4), 1% (w/v) PEG
6000, and 50mM dithiothreitol, and centrifuged at 15,000g for 5min at 4�C. Thecarboxylation reaction of PEPCK was assayed in a 2-mL reaction medium
containing 50 mM HEPES-KOH (pH 7.3), 5 mM phosphoenolpyruvate, 0.2 mM
NADH, 5mM dithiothreitol, 50 mMNaHCO3, 5 mMMnCl2, 0.5 mMADP, and 10
units of MDH. The reaction was initiated by the addition of 400 mL of extract,
and the changes in the A340 were continuously measured for 4 min at 25�C.
RNA Isolation and Hybridization
Total RNA was extracted from 100 mg of powdered plant material using
the Plant RNeasy kit according to the manufacturer’s instructions (Qiagen).
RNA-blot hybridization was performed with specific 32P-labeled DNA probes
against PEPC, MDH, and PEPCK generated by PCR using specific primer
pairs based on pineapple expressed sequences (Supplemental Table S1). RNA
blotting, membrane hybridization, washing conditions, and PCR procedures
for probe amplification were undertaken as described by Freschi et al. (2009).
To normalize the loading quantity of RNA, a DNA probe from pineapple
small RNA subunit 18S genes (18S rRNA; Supplemental Table S1) was used in
subsequent rehybridizations. Transcript values are mean values obtained
from at least three repetitions.
Hormonal Analysis
Endogenous levels of the CKs iP, iPR, Z, and ZR in frozen leaf samples of
pineapple were analyzed by HPLC associated with ELISA as described by
Freschi et al. (2009). The recovery of CKs through this method was over 60%.
ABA and IAA were purified by HPLC and quantified by gas chromatog-
raphy-tandem mass spectrometry-selected ion monitoring based on Santos
et al. (2004), with somemodifications. For ABA and IAA quantification, frozen
leaf samples were ground in liquid nitrogen, extracted in 5 volumes (v/w) of
cold 80% (v/v) methanol containing 146 mM butylhydroxytoluene, and stirred
for 60 h at 4�C. During this first stage of sample preparation, 1 mg of [13C6]IAA
(Cambridge Isotopes) and 0.5 mg of [2H6]ABA (Olchemin) were added to each
sample as internal standards. IAA and ABA were purified by HPLC using a
C18 column (Prep Nova-Pack HR; 6 mm, 7.8 3 300 mm; Waters), which was
eluted with a gradient of methanol and 0.2% formic acid at a flow rate of 3 mL
min21. The fractions corresponding to ABA and IAA were collected, evapo-
rated, and treated with diazomethane. The methylated samples were nitrogen
dried, resuspended in 50 mL of ethyl acetate, and analyzed in a gas chromato-
graph (Hewlett-Packard 6890) connected to a mass selective detector (model
5973). The column used for separation was an HP-1701 (30 m, i.d. 0.25 mm, 0.5
mm), and helium was the carrier gas at a flow rate of 1 mL min21. Ratios of the
heavy isotope ABA ion (194) compared with endogenous ABA ion (190) and
of the heavy isotope IAA ion (130) compared with native IAA ion (136) were
used to calculate the endogenous concentrations of these hormones in the
pineapple leaf tissues. The recoveries of [13C6]IAA and [2H6]ABA were over
55% and 65%, respectively.
For the ET emission analyses, intact plants of pineapple (typically 30
individuals) were placed into a 0.5-L sealed glass vessel, flushed with ET-free
air (3 L min21) for 5 min, and incubated for 6 h (second half of the light period)
under the experimental conditions. After incubation, 10-mL gas samples were
withdrawn with a gas-tight syringe and injected into a Trace GC Ultra gas
chromatograph (Thermo Electron) fitted with a flame-ionization detector and
a RT-alumina Plot column (Restek). Nitrogen was used as the carrier gas at a
flow rate of 4 mL min21, and commercial standard mixtures of ETwere used
for the calibration curves. Column, injector, and detector temperatures were
40�C, 250�C, and 250�C, respectively.
Measurement of NO by Chemiluminescence
NO emission from pineapple plants was measured by chemiluminescence
detection (Planchet and Kaiser, 2006). About 30 plants were placed in a 0.5-L
sealed glass vessel, and a constant flow of NO-free nitrogen of approximately
0.3 L min21 was pulled through the container and subsequently through a
chemiluminescence detector (CLD 88ep; Eco-Physics). Pressurized nitrogen
was passed through a custom-made charcoal column (1 m long, 5 cm i.d.,
particle size 2 mm) in order to provide a continuous source of NO-free
measuring gas. The detector was adjusted to 20-s time resolution. Calibration
was routinely carried out with NO-free nitrogen and with a commercial
ABA and NO Interact during CAM Signaling
Plant Physiol. Vol. 152, 2010 1983 www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

standard mixture of 120 nL L21 NO in nitrogen (Air Liquid). In all cases, NO
emission values were determined 3 h before the end of the light period.
NO Detection by CLSM
Endogenous NO formation was visualized using the NO-sensitive fluo-
rescent probe DAF2-DA (Sigma-Aldrich) and CLSM. Free-hand transversal
leaf sections from control andwater deficit-treated plants were stained with 10
mMDAF2-DA dissolved in 10mM Tris-HCl buffer (pH 7.4) for 30min at 25�C in
the dark. Thereafter, the leaf sections were thoroughly washed to remove
excess fluorophore and immediately analyzed using a CLSM apparatus (LSM
510; Zeiss). The sections were excited at 488 nm, and emission signals between
505 and 530 nm were collected. Images were processed and analyzed using
Zeiss LSM 510 software. Identical confocal settings were maintained during
the course of all experiments. To verify if the increases in green epifluo-
rescence were attributable to NO production, additional leaf sections were
loaded for 30 min with the NO-inactive probe 4-aminofluorescein diacetate
(10 mM) rather than DAF2-DA or coincubated with both DAF2-DA (10 mM) and
cPTIO (500 mM), thoroughly washed, and then visualized by CLSM. All
images shown represent typical results from observation of at least five
replicate leaves from each experiment, with every experiment repeated at
least two times.
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure S1. Time-course analysis of NO release by NO
donors in the experimental conditions.
Supplemental Figure S2. Epifluorescence microscopy analysis of endog-
enous NO in water deficit-treated pineapple plants.
Supplemental Figure S3. Epifluorescence microscopy analysis of endog-
enous NO in ABA-treated pineapple plants.
Supplemental Figure S4. In vitro multiplication of pineapple plants and
characterization of the growth stage of the plants used in the experi-
ments.
Supplemental Table S1. List of primers used for the amplification of
selected genes by PCR in pineapple.
Received November 25, 2009; accepted February 7, 2010; published February
10, 2010.
LITERATURE CITED
Arasimowicz M, Floryszak-Wieczorek J (2007) Nitric oxide as a bioactive
signalling molecule in plant stress responses. Plant Sci 172: 876–887
Bellaire BA, Carmody J, Braud J, Gossett DR, Banks SW, Lucas MC,
Fowler TE (2000) Involvement of abscisic acid-dependent and -inde-
pendent pathways in the upregulation of antioxidant enzyme activity
during NaCl stress in cotton callus tissue. Free Radic Res 33: 531–545
Borland A, Elliott S, Patterson S, Taybi T, Cushman J, Pater B, Barnes J
(2006) Are the metabolic components of Crassulacean acid metabolism
up-regulated in response to an increase in oxidative burden? J Exp Bot
57: 319–328
Borland AM, Tecsi LI, Leegood RC, Walker RP (1998) Inducibility of
Crassulacean acid metabolism (CAM) in Clusia species: physiological/
biochemical characterisation and intercellular localization of carboxyl-
ation and decarboxylation processes in three species which exhibit
different degrees of CAM. Planta 205: 342–351
Bright J, Desikan R, Hancock JT, Weir IS, Neill SJ (2006) ABA-induced
NO generation and stomatal closure in Arabidopsis are dependent on
H2O2 synthesis. Plant J 45: 113–122
Chu C, Dai ZY, Ku MSB, Edwards GE (1990) Induction of Crassulacean
acid metabolism in the facultative halophyte Mesembryanthemum crys-
tallinum by abscisic acid. Plant Physiol 93: 1253–1260
Corpas FJ, Chaki M, Fernandez-Ocana A, Valderrama R, Palma JM,
Carreras A, Begara-Morales JC, Airaki M, del Rio LA, Barroso JB
(2008) Metabolism of reactive nitrogen species in pea plants under
abiotic stress conditions. Plant Cell Physiol 49: 1711–1722
Courtois C, Besson A, Dahan J, Bourque S, Dobrowolska G, Pugin A,
Wendehenne D (2008) Nitric oxide signalling in plants: interplays with
Ca2+ and protein kinases. J Exp Bot 59: 155–163
Cushman JC (2001) Crassulacean acid metabolism: a plastic photosynthetic
adaptation to arid environments. Plant Physiol 127: 1439–1448
Cushman JC, Bohnert HJ (1999) Crassulacean acid metabolism: molecular
genetics. Annu Rev Plant Physiol Plant Mol Biol 50: 305–332
Cushman JC, Borland AM (2002) Induction of Crassulacean acid metab-
olism by water limitation. Plant Cell Environ 25: 295–310
Dai Z, Ku MSB, Zhang DZ, Edwards GE (1994) Effects of growth
regulators on the induction of Crassulacean acid metabolism in the
facultative halophyte Mesembryanthemum crystallinum L. Planta 192:
287–294
Delledonne M, Xia YJ, Dixon RA, Lamb C (1998) Nitric oxide functions as
a signal in plant disease resistance. Nature 394: 585–588
Ederli L, Morettini R, Borgogni A, Wasternack C, Miersch O, Reale L,
Ferranti F, Tosti N, Pasqualini S (2006) Interaction between nitric oxide
and ethylene in the induction of alternative oxidase in ozone-treated
tobacco plants. Plant Physiol 142: 595–608
Freschi L, Nievola CC, Rodrigues MA, Domingues DS, Van Sluys MA,
Mercier H (2009) Thermoperiod affects the diurnal cycle of nitrate
reductase expression and activity in pineapple plants by modulating the
endogenous levels of cytokinins. Physiol Plant 137: 201–212
Freschi L, Takahashi CA, Cambui CA, Semprebom TR, Cruz AB, Mioto
PT, Versieux LM, Calvente A, Latansio-Aidar SR, Aidar MPM, et al
(2010) Specific leaf areas of the tank bromeliad Guzmania monostachia
perform distinct functions in response to water shortage. J Plant Physiol
(in press)
Garcia-Mata C, Lamattina L (2001) Nitric oxide induces stomatal closure
and enhances the adaptive plant responses against drought stress. Plant
Physiol 126: 1196–1204
Garcia-Mata C, Lamattina L (2002) Nitric oxide and abscisic acid cross talk
in guard cells. Plant Physiol 128: 790–792
Gould KS, Lamotte O, Klinguer A, Pugin A, Wendehenne D (2003) Nitric
oxide production in tobacco leaf cells: a generalized stress response?
Plant Cell Environ 26: 1851–1862
Hao GP, Xing Y, Zhang JH (2008) Role of nitric oxide dependence on nitric
oxide synthase-like activity in the water stress signaling of maize
seedling. J Integr Plant Biol 50: 435–442
Hohorst HJ (1965) L-(-)-Malate: determination with malic dehydrogenase
and DPN. In HU Bergmeyer, ed, Methods of Enzymatic Analysis.
Academic Press, New York, pp 328–332
Holthe PA, Sternberg LDL, Ting IP (1987) Developmental control of CAM
in Peperomia scandens. Plant Physiol 84: 743–747
Hurst AC, Grams TEE, Ratajczak R (2004) Effects of salinity, high irradi-
ance, ozone, and ethylene on mode of photosynthesis, oxidative stress
and oxidative damage in the C-3/CAM intermediate plant Mesembry-
anthemum crystallinum L. Plant Cell Environ 27: 187–197
Ishitani M, Xiong LM, Stevenson B, Zhu JK (1997) Genetic analysis of osmotic
and cold stress signal transduction in Arabidopsis: interactions and conver-
gence of abscisic acid-dependent and abscisic acid-independent pathways.
Plant Cell 9: 1935–1949
Jones MB (1975) Effect of leaf age on leaf resistance and CO2 exchange of
CAM plant Bryophyllum fedtschenkoi. Planta 123: 91–96
Leshem YY, Haramaty E (1996) The characterization and contrasting effects
of the nitric oxide free radical in vegetative stress and senescence of
Pisum sativum Linn foliage. J Plant Physiol 148: 258–263
Lindermayr C, Saalbach G, Durner J (2005) Proteomic identification of
S-nitrosylated proteins in Arabidopsis. Plant Physiol 137: 921–930
Luttge U (2004) Ecophysiology of Crassulacean acid metabolism (CAM).
Ann Bot (Lond) 93: 629–652
Martin CE (1994) Physiological ecology of the Bromeliaceae. Bot Rev
60: 1–82
Murashige T, Skoog F (1962) A revised medium for rapid growth and
bioassays with tobacco tissue cultures. Physiol Plant 15: 473–497
Neill S, Barros R, Bright J, Desikan R, Hancock J, Harrison J, Morris P,
Ribeiro D, Wilson I (2008) Nitric oxide, stomatal closure, and abiotic
stress. J Exp Bot 59: 165–176
Neill SJ, Desikan R, Clarke A, Hurst RD, Hancock JT (2002) Hydrogen
peroxide and nitric oxide as signalling molecules in plants. J Exp Bot 53:
1237–1247
Nievola CC, Kraus JE, Freschi L, Souza BM,Mercier H (2005) Temperature
determines the occurrence of CAM or C3 photosynthesis in pineapple
plantlets grown in vitro. In Vitro Cell Dev Biol Plant 41: 832–837
Freschi et al.
1984 Plant Physiol. Vol. 152, 2010 www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.

Niewiadomska E, Pater B, Miszalski Z (2002) Does ozone induce a
C3-CAM transition in Mesembryanthemum crystallinum leaves? Phyton
42: 69–78
Peters W, Beck E, Piepenbrock M, Lenz B, Schmitt JM (1997) Cytokinin as
a negative effector of phosphoenolpyruvate carboxylase induction in
Mesembryanthemum crystallinum. J Plant Physiol 151: 362–367
Planchet E, Kaiser WM (2006) Nitric oxide (NO) detection by DAF
fluorescence and chemiluminescence: a comparison using abiotic and
biotic NO sources. J Exp Bot 57: 3043–3055
Rodriguez-Serrano M, Romero-Puertas MC, Pazmino DM, Testillano PS,
Risueno MC, del Rio LA, Sandalio LM (2009) Cellular response of pea
plants to cadmium toxicity: cross talk between reactive oxygen species,
nitric oxide, and calcium. Plant Physiol 150: 229–243
Ruan HH, ShenWB, Xu LL (2004) Nitric oxide involved in the abscisic acid
induced proline accumulation in wheat seedling leaves under salt
stress. Acta Bot Sin 46: 1307–1315
Sang JR, Jiang MY, Lin F, Xu SC, Zhang A, Tan MP (2008) Nitric oxide
reduces hydrogen peroxide accumulation involved in water stress-
induced subcellular anti-oxidant defense in maize plants. J Integr Plant
Biol 50: 231–243
Santos HP, Purgatto E, Mercier H, Buckeridge MS (2004) The control of
storage xyloglucan mobilization in cotyledons of Hymenaea courbaril.
Plant Physiol 135: 287–299
Schmitt JM, Piepenbrock M (1992) Regulation of phosphoenolpyruvate
carboxylase and Crassulacean acid metabolism induction in Mesembry-
anthemum crystallinum L by cytokinin: modulation of leaf gene expres-
sion by roots. Plant Physiol 99: 1664–1669
Slesak I, Karpinska B, Surowka E, Miszalski Z, Karpinski S (2003) Redox
changes in the chloroplast and hydrogen peroxide are essential for
regulation of C3-CAM transition and photooxidative stress responses in
the facultative CAM plant Mesembryanthemum crystallinum L. Plant Cell
Physiol 44: 573–581
Slesak I, Libik M, Miszalski Z (2008) The foliar concentration of hydrogen
peroxide during salt-induced C3-CAM transition in Mesembryanthemum
crystallinum L. Plant Sci 174: 221–226
Song LL, Ding W, Shen J, Zhang ZG, Bi YR, Zhang LX (2008) Nitric oxide
mediates abscisic acid induced thermotolerance in the calluses from two
ecotypes of reed under heat stress. Plant Sci 175: 826–832
Springer SA, Outlaw WH (1988) Histochemical compartmentation of
photosynthesis in the Crassulacean acid metabolism plant Crassula
falcata. Plant Physiol 88: 633–638
Taybi T, Cushman JC (1999) Signaling events leading to Crassulacean acid
metabolism induction in the common ice plant. Plant Physiol 121: 545–555
Taybi T, Cushman JC (2002) Abscisic acid signaling and protein synthesis
requirements for phosphoenolpyruvate carboxylase transcript induc-
tion in the common ice plant. J Plant Physiol 159: 1235–1243
Taybi T, Cushman JC, Borland AM (2002) Environmental, hormonal and
circadian regulation of Crassulacean acid metabolism expression. Funct
Plant Biol 29: 669–678
Taybi T, Sotta B, Gehrig H, Guclu S, KlugeM, Brulfert J (1995) Differential
effects of abscisic acid on phosphoenolpyruvate carboxylase and CAM
operation in Kalanchoe blossfeldiana. Bot Acta 108: 240–246
Thomas JC, McElwain EF, Bohnert HJ (1992) Convergent induction of
osmotic stress-responses: abscisic acid, cytokinin, and the effects of
NaCl. Plant Physiol 100: 416–423
Tian X, Lei Y (2006) Nitric oxide treatment alleviates drought stress in
wheat seedlings. Biol Plant 50: 775–778
Ting IP (1981) Effects of abscisic acid on CAM in Portulacaria afra. Photo-
synth Res 2: 39–48
Tossi V, Lamattina L, Cassia R (2009) An increase in the concentration of
abscisic acid is critical for nitric oxide-mediated plant adaptive re-
sponses to UV-B irradiation. New Phytol 181: 871–879
Winter K, Aranda J, Holtum JAM (2005) Carbon isotope composition and
water-use efficiency in plants with Crassulacean acid metabolism. Funct
Plant Biol 32: 381–388
Winter K, Garcia M, Holtum JAM (2008) On the nature of facultative and
constitutive CAM: environmental and developmental control of CAM
expression during early growth of Clusia, Kalanchoe, and Opuntia. J Exp
Bot 59: 1829–1840
Xing H, Tan LL, An LH, Zhao ZG,Wang SM, Zhang CL (2004) Evidence for
the involvement of nitric oxide and reactive oxygen species in osmotic
stress tolerance of wheat seedlings: inverse correlation between leaf
abscisic acid accumulation and leaf water loss. Plant Growth Regul 42:
61–68
Zhang AY, Jiang MY, Zhang JH, Ding HD, Xu SC, Hu XL, Tan MP (2007)
Nitric oxide induced by hydrogen peroxide mediates abscisic acid-
induced activation of the mitogen-activated protein kinase cascade
involved in antioxidant defense in maize leaves. New Phytol 175:
36–50
Zhao LQ, Zhang F, Guo JK, Yang YL, Li BB, Zhang LX (2004) Nitric oxide
functions as a signal in salt resistance in the calluses from two ecotypes
of reed. Plant Physiol 134: 849–857
ZhaoMG,ChenL, ZhangLL, ZhangWH (2009)Nitric reductase-dependent
nitric oxide production is involved in cold acclimation and freezing
tolerance in Arabidopsis. Plant Physiol 151: 755–767
Zhao ZG, Chen GC, Zhang CL (2001) Interaction between reactive oxygen
species and nitric oxide in drought-induced abscisic acid synthesis in
root tips of wheat seedlings. Aust J Plant Physiol 28: 1055–1061
Zhu J, Bartholomew DP, Goldstein G (1997) Effect of elevated carbon
dioxide on the growth and physiological responses of pineapple, a
species with Crassulacean acid metabolism. J Am Soc Hortic Sci 122:
233–237
ABA and NO Interact during CAM Signaling
Plant Physiol. Vol. 152, 2010 1985 www.plantphysiol.orgon July 12, 2018 - Published by Downloaded from
Copyright © 2010 American Society of Plant Biologists. All rights reserved.


















