NINJA connects the co-repressor TOPLESS to Jasmonate signallingsignalling Laurens Pauwels1,2*, Gemma...
111
NINJA connects the co-repressor TOPLESS to Jasmonate signalling Laurens Pauwels1,2*, Gemma Ferna´ndez Barbero3*, Jan Geerinck1,2*, Sofie Tilleman1,2, Wim Grunewald1,2,5, Amparo Cue´llar Pe´rez1,2, Jose´ Manuel Chico3, Robin Vanden Bossche1,2, Jared Sewell6, Eduardo Gil4, Gloria Garcı´a-Casado4, Erwin Witters7,8, Dirk Inze´1,2, Jeff A. Long6, Geert De Jaeger1,2, Roberto Solano3,4 & Alain Goossens1,2 Vol 464| 1 April 2010| /nature 1Department of Plant Systems Biology, Flanders Institute for Biotechnology (VIB), Technologiepark 927, B-9052 Gent, Belgium. 2Department of Plant Biotechnology and Genetics, Ghent University, Technologiepark 927, B-9052 Gent, Belgium. 3Departamento de Gene´tica Molecular de Plantas, 4Genomics Unit, Centro Nacional de Biotecnologı´a-CSIC, Campus Universidad Auto´noma, 28049 Madrid, Spain. 5Department of Molecular Biotechnology, Faculty of Bioscience Engineering, Ghent University, Coupure links 653, B-9000 Gent, Belgium. 6Plant Biology Laboratory, The Salk Institute for Biological Studies, 10010 North Torrey Pines Road, La Jolla, California 92037, USA. 7Department of Biology, EBT- CEPROMA, University of Antwerp, Groenenborgerlaan 171, B-2020 Antwerpen, Belgium. 8Flemish Institute for Technological Research VITO-MANT, Boeretang 200, B-2400 Mol, Belgium. *These authors contributed equally to this work. In the Name of God 1
-
Upload
beryl-phelps -
Category
Documents
-
view
214 -
download
0
Transcript of NINJA connects the co-repressor TOPLESS to Jasmonate signallingsignalling Laurens Pauwels1,2*, Gemma...

NINJA connects the co-repressor TOPLESS to Jasmonate signalling
Laurens Pauwels1,2*, Gemma Ferna´ndez Barbero3*, Jan Geerinck1,2*, Sofie Tilleman1,2, Wim Grunewald1,2,5,
Amparo Cue´llar Pe´rez1,2, Jose´ Manuel Chico3, Robin Vanden Bossche1,2, Jared Sewell6, Eduardo Gil4,
Gloria Garcı´a-Casado4, Erwin Witters7,8, Dirk Inze´1,2, Jeff A. Long6, Geert De Jaeger1,2, Roberto Solano3,4
& Alain Goossens1,2
Vol 464| 1 April 2010| /nature1Department of Plant Systems Biology, Flanders Institute for Biotechnology (VIB), Technologiepark 927, B-9052
Gent, Belgium. 2Department of Plant Biotechnology and Genetics,Ghent University, Technologiepark 927, B-9052 Gent, Belgium. 3Departamento de Gene´tica Molecular de
Plantas, 4Genomics Unit, Centro Nacional de Biotecnologı´a-CSIC, CampusUniversidad Auto´noma, 28049 Madrid, Spain. 5Department of Molecular Biotechnology, Faculty of Bioscience
Engineering, Ghent University, Coupure links 653, B-9000 Gent,Belgium. 6Plant Biology Laboratory, The Salk Institute for Biological Studies, 10010 North Torrey Pines Road, La
Jolla, California 92037, USA. 7Department of Biology, EBT-CEPROMA,University of Antwerp, Groenenborgerlaan 171, B-2020 Antwerpen, Belgium. 8Flemish Institute for
Technological Research VITO-MANT, Boeretang 200, B-2400 Mol, Belgium.*These authors contributed equally to this work.
In the Name of God

2

Objectives
• By Yeast two-hybrid system, tandem affinity purification and mass spectrometry approaches,
• the authors showed that JA signalling repressors (JAZs) interact with the novel interactor of JAZ (NINJA) and TPL protein complex in order to repress JA-induced genes.

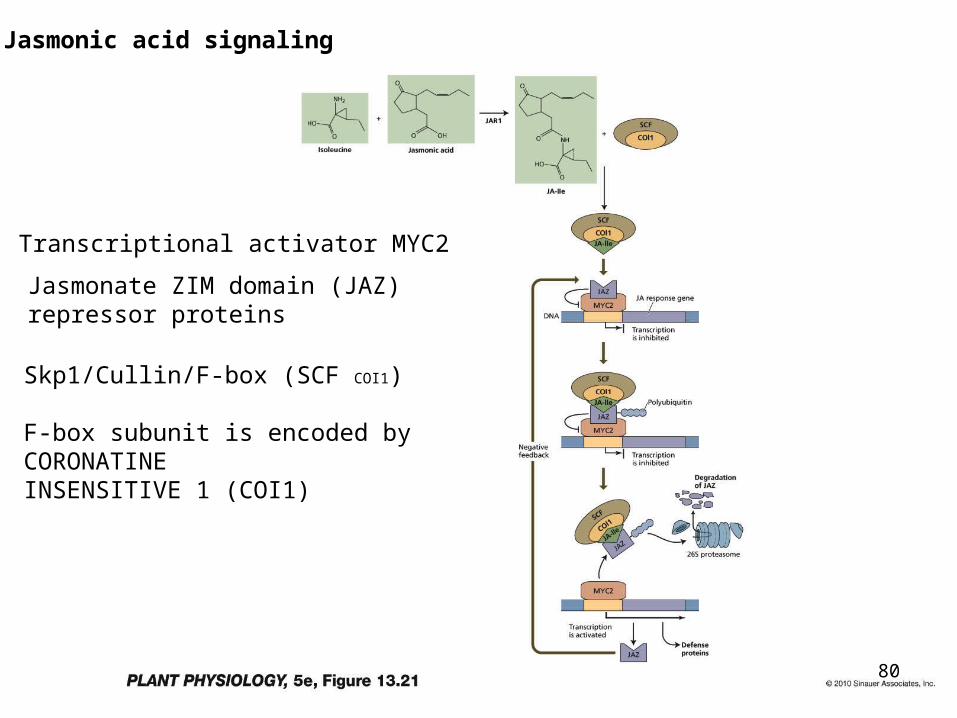
JA-Ile signal transduction
• JA-Ile signal transduction is controlled by a Skp1/Cullin/F-box (SCF)E3 ubiquitin ligase whose F-box subunit is encoded by CORONATINE INSENSITIVE 1 (COI1).
• The JAZ proteins have been identified as the targets of SCFCOI1.
• In the presence of (1)-7-iso-JA-L-Ile, the endogenous bioactive hormone, JAZ proteins interact with COI1 and are subsequently degraded by the 26S proteasome.

JA-Ile signal transduction
• In an unelicited state, JAZ proteins bind the transcription factor MYC2 and inactivate its function.
• Hormone-triggered removal of JAZ proteins releases MYC2, enabling the transcriptional activator to regulate the expression of early jasmonate-responsive genes, including the JAZ genes.

JA-Ile signal transduction
• Tandem affinity purification (TAP) was applied to isolate the core jasmonate signalling module and to find new JAZ interactors
• We used a recently established TAP echnology platform that allows the efficient isolation and identification of protein complexes from Arabidopsis cells
• For the simultaneous visualization of the dynamics of JAZ complex assembly, TAP was performed with cell cultures elicited or not with jasmonic acid (JA).
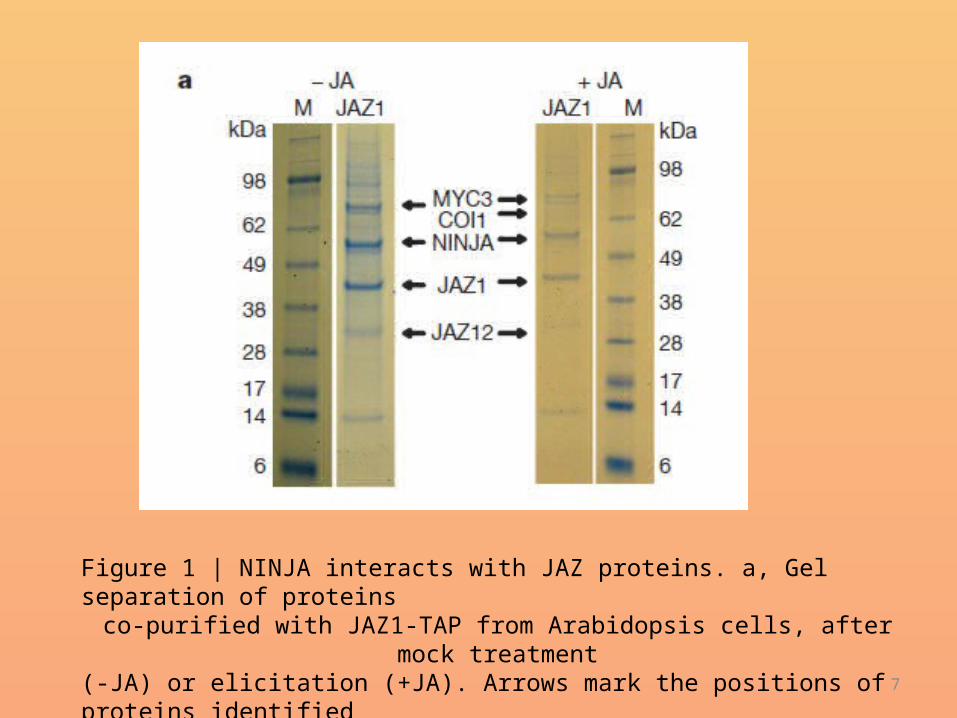
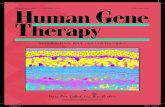
Figure 1 | NINJA interacts with JAZ proteins. a, Gel separation of proteinsco-purified with JAZ1-TAP from Arabidopsis cells, after mock treatment
(-JA) or elicitation (+JA). Arrows mark the positions of proteins identifiedby MS. LaneMshows molecular mass markers.

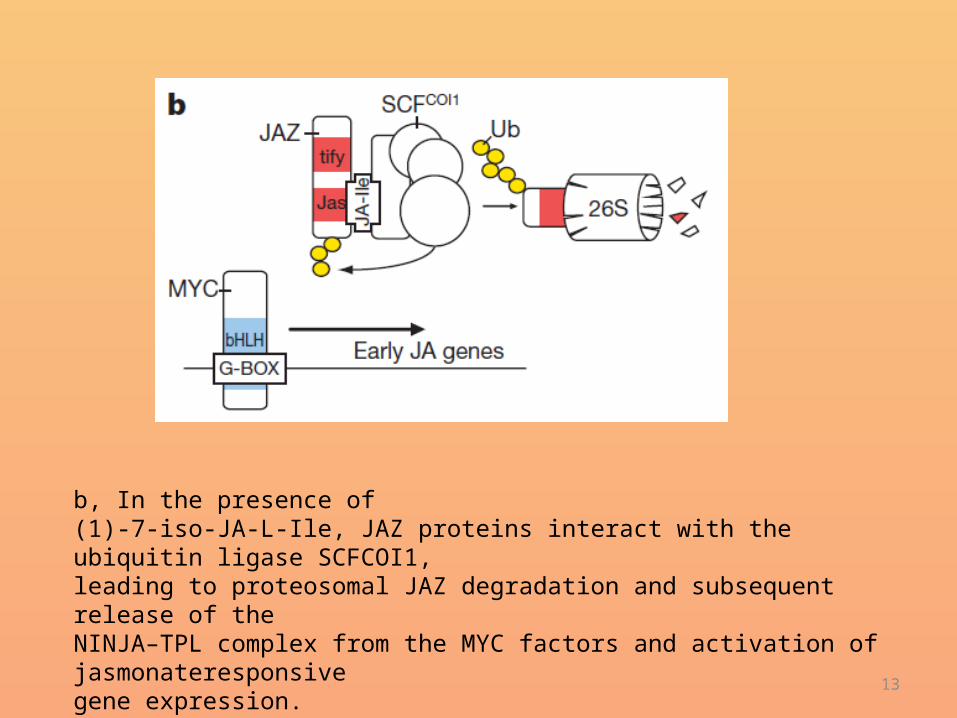
b, c, NINJA-interacting and JAZ-interacting protein domains in Y2H assays. Drawings represent JAZ1, NINJA and derivatives. Numbers indicate terminal amino-acid residues.Transformed yeasts were spotted in 10-fold and 100-fold dilutions oncontrol medium (-2) or selective medium (-3). Controls are empty vectors. AD, activation domain. b, Interaction of NINJA with the TIFY motif of JAZ1. c, Interaction of JAZ1 with the C domain of NINJA.DBD: DNA binding domain(residues 128–133; Dtify)

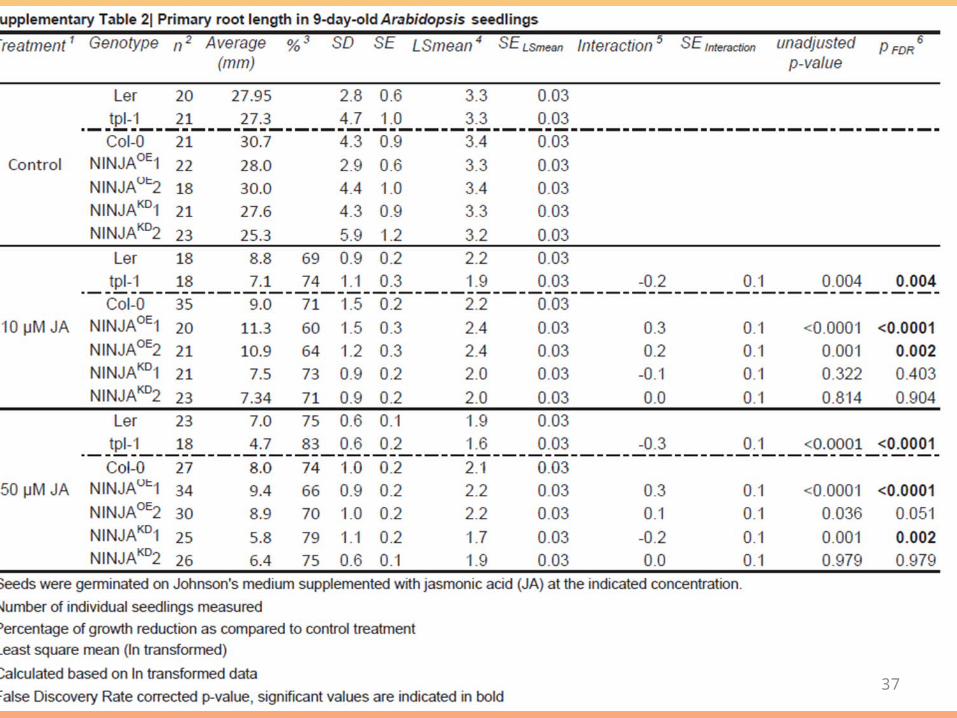
Figure 2 | NINJA negatively regulates JA signalling. a, Phenotypes oftransgenic NINJA overexpression (NINJAOE) and NINJA knockdown(NINJAKD) Arabidopsis lines. Average primary root lengths of 9-day-oldseedlings germinated on control medium, or on 10 or 50 mM JA are shown.Wildtype Col-0 seedlingsSchmidt, L., Hummel, G.M., Schoettner, M., Schurr, U. and Walter, A. 2010. Jasmonic acid does not mediate root growth responses to wounding in Arabidopsis thaliana. Plant, Cell & Environment, 33: 104-116.

b,Upregulation Of MeJA-responsive genes in NINJA knockdown plants. Heat maps, generatedby Expression Browser software (http://bbc.botany.utoronto.ca), representMeJA-modulated expression over time (in hours) of genes significantlyinduced in NINJA knockdown plants. The scale bar indicates log fold change.

Figure 3 | NINJA interacts with TPL and functions as a transcriptionalrepressor. a, NINJA-interacting and TPL-interacting protein domains in aY2H assay. Drawings represent TPL, NINJA and derivatives. Y2H analysiswas performed as in Fig. 1.
Groucho/Tup1-type corepressor TOPLESS (TPL)

Figure 4 | Model for a general function of TPL proteins in plant hormonesignalling. a, In the absence of jasmonates, basic helix–loop–helix (bHLH)MYC factors interact with the Jas domain of JAZ proteins that interactthrough their TIFY motif with domainCof NINJA. TheEAR motif of NINJAis essential for interaction with the TPL co-repressors.
b, In the presence of(1)-7-iso-JA-L-Ile, JAZ proteins interact with the ubiquitin ligase SCFCOI1,leading to proteosomal JAZ degradation and subsequent release of theNINJA–TPL complex from the MYC factors and activation of jasmonateresponsivegene expression.

c, Jasmonate and auxin pathways are built onsimilar signalling modules.

d, NINJA interacts with other group II TIFYproteins, which might be recruited by as yet unknown transcription factors.

e, Interaction of the NINJA-related AFP proteins with ABI5 and TPL toregulate responses to ABA.

THE END
17

18
19

20
THE END

21
Fig 3
b, JA phenotype of tpl-1. Root growth analysiswas performed as in Fig. 2a. Ler, Landsberg erecta.

22
Fig 3
c, d, Transactivation activity in tobacco protoplasts transfected with a PUAS–fLUC reporter construct, effector constructs fused or not with GAL4DBD, and a P35SrLUCnormalization construct. Data in c and d are means6s.e.m. relative tothe reference (black bar) for n58 (two asterisks, P#0.01; three asterisks,P#0.001; analysis of variance, Dunnett’s post-hoc test).c, Functionality of the NINJA EAR repression motif.

23
d, MYC2 activity is repressed by NINJA.

24

Supplementary Figure 1| Induction of rapid JAZ1 degradation by JA. a, Schematicoverview of the T-DNA constructed by multisite Gateway cloning with the destination vectorpKCTAP. LB and RB, T-DNA borders; B1, B2, B3 and B4, Gateway recombination sites; eGFP,enhanced Green Fluorescent Protein under control of the rolD promoter; KmR, kanamycinresistance marker under control of the nos promoter; fLUC, firefly luciferase. b, Cycloheximide(CHX) chase experiment. Stably transformed cell cultures expressing fLUC or a JAZ1-fLUCfusion were treated with 200 μg/mL CHX and 100 μM JA or CHX alone and sampled up to 5 minafter treatments. Average normalized fLUC activities ± s.d. are plotted. Full lines are best-fitlines.

Supplementary Figure 2| Induction of NINJA expression by MeJA, but not by ABA. a,Microarray analysis of AFP and NINJA gene expression response to MeJA and ABA in 7-dayoldArabidopsis seedlings38. Mean expression values relative to the untreated control areplotted. Asterisks indicate a significantly differential expression (FDR corrected p-value ≤ 0.05).

b, Real-time qPCR analysis of MYC2, JAZ1 and NINJA expression over time in response to 25μM MeJA (dashed lines) or mock treatment (solid lines). Fold changes relative to the zero timepoint are plotted in function of time (hours).



Supplementary Figure 5| Stability and subcellular localization of NINJA-GFP. Laserscanning confocal microscopy images of Arabidopsis seedling roots expressing JAZ1-GFP (a)or NINJA-GFP (b). Roots were treated with 25 μM MeJA for the time indicated (minutes) andstained with propidium iodide to visualize cell borders. GFP fluorescence was observed andimaged with a Zeiss LSM 510 laser scanning confocal microscope. Scale bars, 50 μm.

Supplementary Figure 6| Expression of NINJA in transgenic gain-of-function or loss-offunctionArabidopsis plants. a, RNA gel blot analysis of NINJA expression in 3-week-old wildtype(Col-0) and two NINJA overexpressing (OE) lines. Total RNA (8 μg) was loaded. b, RealtimeqPCR analysis of NINJA expression in 7-day-old wild-type (Col-0) and two NINJAknockdown (KD) lines. Data are means ± s.e.m.


Supplementary Figure 8| Upregulation of early MeJA responsive genes in elicited NINJAKD plants. Shown in heat maps is the MeJA modulated expression in wild-type 7-day-oldseedlings (0.5, 1 and 3 h after treatment) of genes significantly induced in NINJA KD plants(versus wild-type) treated with 0.5 μM coronatine for 6 h. Data were obtained with theExpression Browser software (http://bar.toronto.ca). The scale bar indicates the log fold change.Only genes early induced by MeJA (97 out of 169) are included. A complete list of the genesthat are differentially regulated in coronatine-treated NINJA KD plants is given in SupplementalTable 4. Many of the genes in this list are also regulated by MeJA at later times (e.g. PDFgenes).






JA-Ile signal transduction
• Because JAZ degradation on treatment with JA has been reported to be very rapid15, we first investigated JAZ degradation kinetics in our model system
• A cycloheximide chase experiment revealed an estimated half-life of 1.4 min for a JAZ1–firefly luciferase (fLUC) fusion protein on treatment with JA.
• Next, JAZ1 was carboxy-terminally fused to a TAP tag and expressed under a cauliflower mosaic virus (CaMV) 35S promoter in Arabidopsis cells.

• To avoid full degradation of the tagged bait, cell cultures producing JAZ1–TAP were harvested 1 min after treatment with JA.
• Several proteins already shown to interact with JAZ1 were identified (Fig. 1a), validating the approach (for the massspectrometry-driven identification of candidate interactors.
• First, JAZ1 formed a complex with JAZ12, matching the reported dimerization of JAZ proteins
• Second, interaction of JAZ1 with MYC3, a close relative to MYC2, was observed.
• Last, COI1 occurred in a complex with JAZ1, but only in the presence of JA, showing that TAP is applicable for the identification of ligand-mediated protein–protein interactions

JA-Ile signal transduction
• We focused on an uncharacterized protein isolated in the TAP analysis (Fig. 1a), hereafter designated NINJA, and encoded by the locus At4g28910.
• NINJA is related to the ABI-FIVE BINDING PROTEIN (AFP) family, which consists of four members in Arabidopsis.
• Whereas abscisic acid (ABA) induces the expression of AFP genes in Arabidopsis seedlings18, NINJA expression is induced by methyl jasmonate (MeJA) within 1 h and for at least 12 h after elicitation (Supplementary Fig. 2).

JA-Ile signal transduction
• This pattern corresponds to that of the previously identified NINJA homologue of tobacco (Nicotiana tabacum) in a screen for MeJA-inducible transcripts.

yeast two-hybrid• In parallel, we performed yeast two-hybrid (Y2H) library screens with
JAZ2 and JAZ3 as baits, and both identified NINJA as a direct interactor.• Systematic Y2H analysis of the specificity of NINJA for JAZ proteins
revealed interactions with most JAZ proteins; the exceptions were JAZ7 and JAZ8 (Supplementary Fig. 3a).
• Furthermore, other ZIM-domain proteins, containing the TIFY (TIF[F/Y]XG) motif and belonging to the group II TIFY proteins20, such as PEAPOD 1 (PPD1), PPD2 and TIFY8, interacted with NINJA (Supplementary Fig. 4a).
• Additionally, we checked whether AFP proteins could bind JAZ proteins, but neither AFP2 nor AFP3 interacted with JAZ1 (Supplementary Fig. 4b), suggesting the specificity of JAZ proteins for NINJA.

JA-Ile signal transduction
• Besides the TIFY motif, JAZ proteins have a C-terminal conserved Jas domain, which is involved in the interaction with both MYC2 and COI1 (refs 6, 11, 21).
• Y2H with a JAZ1 deletion series revealed that NINJA was only capable of binding JAZ1 fragments containing the TIFY motif.
• Moreover, a 39-amino-acid fragment holding the conserved TIFY motif was sufficient for the binding of NINJA (Fig. 1b and Supplementary Fig. 4c).
• Consistently, NINJA did not interact with a JAZ1 protein fusion lacking the TIFYAG sequence (residues
• 128–133; Dtify) (Fig. 1b).

JA-Ile signal transduction
• Pull-down experiments with JAZ– altose binding protein (JAZ–MBP) fusion proteins and extracts of transgenic plants expressing NINJA fused to green fluorescent protein (NINJA–GFP) confirmed the Y2H observations.
• NINJA was pulled down by all JAZ–MBP fusion proteins except JAZ7, JAZ11 and JAZ12 (Supplementary Fig. 3b).
• Moreover, by using JAZ3 derivatives, only the fragment containing the TIFY motif (JAZ3DC) was able to pull down NINJA–GFP (Supplementary Fig. 3b).
• NINJA–GFP did not interact with NINJA–MBP, COI1–MBP or MYC2–MBP in these pull-down experiments (Supplementary Fig. 3b).

JA-Ile signal transduction
• NINJA and all AFP proteins are characterized by three conserved protein domains designated A, B and C.
• Functionality has been described only for the C domain,which has been shown to be necessary and sufficient for binding of the AFP proteins with the bZIP transcription factor ABSCISIC ACID INSENSITIVE 5 (ABI5)18.
• A deletion series of NINJA was constructed (Fig. 1c) and tested for JAZ1 interaction, showing that the C domain of NINJA was responsible and sufficient for JAZ protein interaction.
• Finally, the 39-residue protein fragment harbouring the TIFY motif of JAZ1 interacted directly with the C domain of NINJA (Supplementary Fig. 4d).

JA-Ile signal transduction
• Analysis of seedlings producing a C-terminal GFP fusion with NINJA revealed a clear nuclear localization for NINJA (Supplementary Fig. 5), as had already been shown for both AFP and JAZ proteins6,7,15,18.
• Furthermore, NINJA–GFP levels remained constant for 3 h after treatment with jasmonate, whereas a JAZ1–GFP fusion was nearly undetectable only 5 min after the application of jasmonate (Supplementary Fig. 5), suggesting that, unlike JAZ proteins, the stability of NINJA is unaffected by jasmonates.
• Knockout T-DNA insertion lines for NINJA were not available from the Arabidopsis database (http://www.arabidopsis.org).
• We therefore generated transgenic Arabidopsis lines expressing fulllength NINJA or NINJA hairpin RNA-mediated interference (RNAi) constructs, both under control of the CaMV 35S promoter (Supplementary Fig. 6a, b).

JA-Ile signal transduction
• NINJA overexpression significantly decreased JA sensitivity, as reflected by an impaired inhibition of root growth in the transgenic plants (Fig. 2a and Supplementary Table 2).
• An in vitro degradation experiment with extracts from NINJA overexpressionlines showed that this phenotype could not be explained by protectionof JAZ proteins from degradation (Supplementary Fig. 7), whichcould account for the phenotype caused by truncated JAZ3 proteinsin the jai3 mutant6.
• Conversely, NINJA knockdown lines showed a derepressed response to JA.

JA-Ile signal transduction
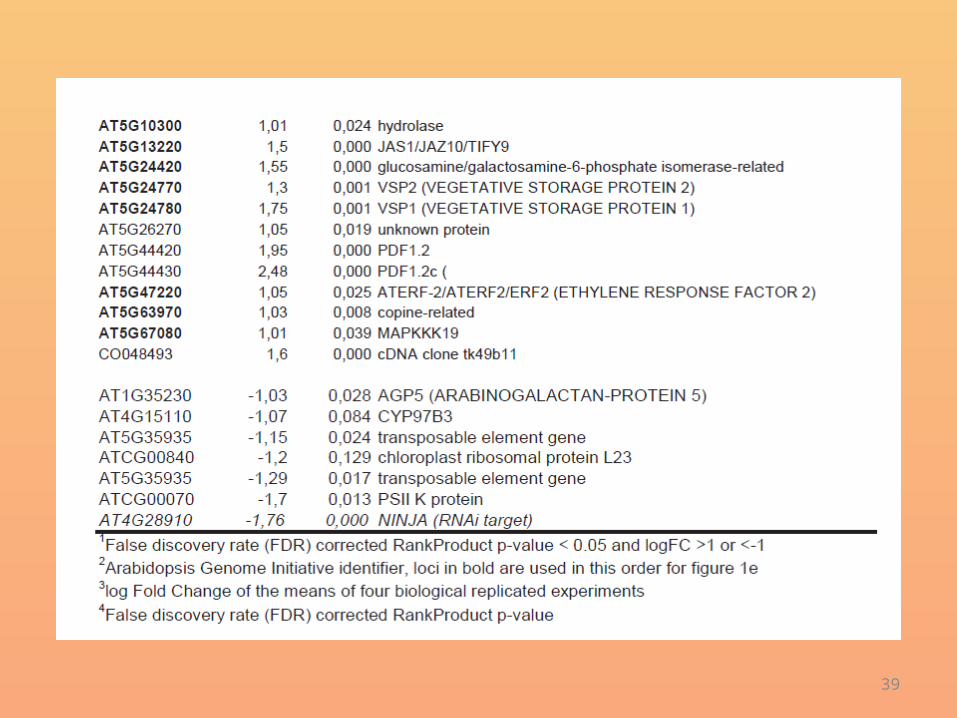
• Microarray analysis revealed that, even without exogenous hormone treatment, NINJA knockdown lines overexpressed many known early jasmonate-responsive genes (Fig. 2band Supplementary Table 3).
• This effect was even more prominent when plants were treated with the JA-Ile mimic coronatine (Supplementary Fig. 8 and Supplementary Table 4).
• Consistently, a slight increase in JA-mediated inhibition of root growth was observed inNINJA knockdown lines (Fig. 2a and Supplementary Table 2).
• Taken together, these results indicate that NINJA functions as a negative regulator of jasmonate signalling.

JA-Ile signal transduction
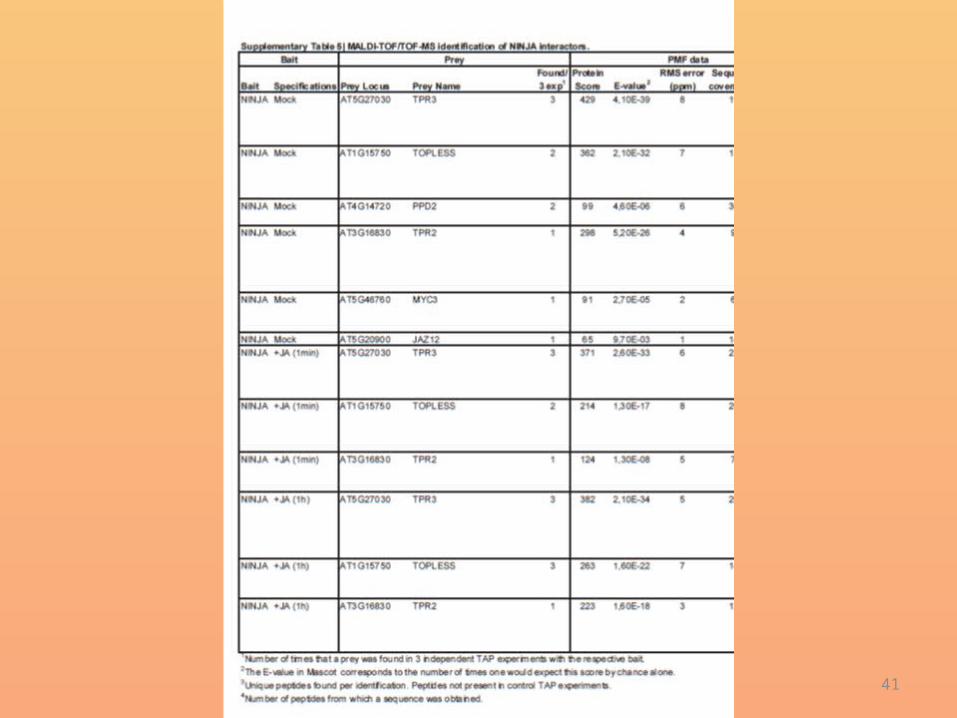
• To investigate NINJA function in JA signalling, we set up a new TAP experiment with NINJA as bait.
• NINJA was present in a complex withthe Groucho/Tup1-type co-repressor TPL and its homologues TPR2and TPR3, independently of JA elicitation (Supplementary Table 5).
• Furthermore, MYC3 and the group II TIFY proteins JAZ12 and PPD2were also identified, confirming the previous interaction data (Fig. 1aand Supplementary Figs 3 and 4).
• TPL interacts directly with the amino-terminal ethylene-responsive element binding factor-associate damphiphilic repression (EAR)motif of the AUX/IAA protein IAA12 (ref. 8).
• The EAR motif is a hallmark of transcriptional repressors and is encountered in most AUX/IAAproteins22.

JA-Ile signal transduction• Correspondingly, scanning of the NINJA amino-acid sequence revealed an EAR
motif in domain A (Supplementary Fig. 9),suggesting a direct interaction between the EAR protein NINJA and TPL, a hypothesis that was confirmed by Y2H and bimolecular fluorescence complementation (Fig. 3a and Supplementary Fig. 10).
• Removal of domain A did not affect the interaction with JAZ1(Fig. 1c) but abolished that with TPL; this also occurred when three Leu residues were mutated to Ala in the NINJA EAR motif (mEAR;Fig. 3a).
• Moreover, domainnA of NINJA was sufficient for the interaction• With TPL(Fig. 3a). • The association of TPL with the JAZ protein complex was further supported by
phenotypic analysis of the tpl-1 mutant23,which showed hypersensitivity to JA (Fig. 3b and Supplementary Table 2).
• Taken together, these results indicate that NINJA connects the JAZ proteins with the TPL co-repressors.

NINJA as a transcriptionalrepressor of jasmonate responses
• Considering the association with TPL, we assessed the transcriptional• repressor capacity of NINJA. • First, a protein fusion of NINJA with the DNA-binding domain (DBD) of
GAL4 could repress the basal activity of a promoter containing upstream activation sequence(PUAS) elements in tobacco protoplasts (Fig. 3c).
• Second, a NINJA fragment containing the EAR motif, but lacking a JAZ interaction domain, was sufficient for repression.
• Conversely, deletion of thedomain A or mutation of three conserved Leu residues in the EAR motif abolished the repression capacity of NINJA (mEAR; Fig. 3c).
• Third, the transcriptional activation of the PUAS by a DBD–MYC2fusion protein could be counteracted by co-expression with NINJA(Fig. 3d).
• Taken together, these results establish NINJA as a transcriptional repressor of jasmonate responses.

JA-Ile signal transduction• Our findings expand the parallelism between the auxin and jasmonate• signalling pathways: the hormones (indole-3-acetic acid and (1)-• 7-iso-JA-L-Ile, respectively) are perceived by F-box proteins (TIR1 and• COI1, respectively) that target repressor proteins (AUX/IAA and JAZ,• respectively) for rapid degradation by the 26S proteasome, which• finally releases the transcription factors (ARFs andMYCs, respectively)• that activate thedownstreamhormone responses24,25 (Fig. 4).Weshow• that transcriptional repression in jasmonate signalling is mediated as• in auxin signalling, by a protein containing an EAR repression motif• that recruits the TPL co-repressors to the complex8 (Fig. 4). However,• in jasmonate signalling the EAR motif is not present on the SCF degradation• targets but on a separate, stable protein, namely NINJA. In this• manner, extra features might have been added to the complex, allowing• crosstalk with other hormone signalling pathways in the cell and• fine-tuning of the jasmonate response.

JA-Ile signal transduction
• The TIFY motif of JAZ proteins is necessary and sufficient for binding to the NINJA–TPL repressor complex, corresponding to the essential role of this domain in the repression of jasmonate signalling by JAZ10.4 (ref. 17).
• The TIFY motif is also necessary for the dimerization of JAZ proteins16,17.
• Future research should reveal whether binding of NINJA and JAZ proteins to the TIFY motif can occur simultaneously and how this dual functionality shapes the repressor activity of the JAZ proteins.

JA-Ile signal transduction
• The TIFY motif is also present in ZIM-domain proteins, such as PPD1, PPD2 and TIFY8, that are not inducible by jasmonates but that nonetheless interact with NINJA (Supplementary Fig. 4a).
• Because the phenotypes associated with the PPD proteins do not seem to be related to jasmonatesignalling26, NINJA might have more functions than mediating jasmonate-responsive gene expression (Fig. 4d).
• The EAR motif of NINJA, as well as that of the AUX/IAA proteins, is responsible for the interaction with TPL proteins.

JA-Ile signal transduction
• This motif is also conserved among all NINJA-related AFP proteins, involved in ABA signalling18 (Fig. 4e), which indicates a possible implication of TPL proteins in multiple signalling pathways, a hypothesis supported by Y2H analysis that demonstrated the direct interaction of AFP proteins and TPL (Supplementary Fig. 4e).
• Thus, TPL proteins emerge as general co-repressors in plants that are recruited to different signalling pathways by specific adaptor proteins, such as AUX/IAA proteins (for auxin), AFP proteins (for ABA) and the complex NINJA–JAZ (for jasmonate).

THE END

Signal transductionSignaling pathways:
• Salicylic acid (SA)• Jasmonic acid (JA)• Ethylene (ET)
• Here, we report on recent advances, updating current knowledge on classical defense hormones SA, JA, and ET, and the roles of auxin, abscisic acid (ABA), cytokinins (CKs), and brassinosteroids in molding plant-pathogen interactions.
61

Signal transduction
Vlot et al., 2008 62

Talking trees?63

Or listening trees?
Airborne Signaling
64

Signal transductionFigure 2Genetically defined relationships betweensalicylic acid (SA)-dependent andethylene–jasmonic acid (ET–JA)-dependentdefense responses in Arabidopsis.The SA-dependent response is deployedagainst a biotrophic pathogen that obtainsnutrients from living cells, whereasthe ET–JA response is activated bynecrotrophic pathogens that kill plant tissue.Resistance to necrotrophs (but notthe biotroph) is compromised in anArabidopsis mutant (coi1) that does notrespond to JA. Expression of a transgene(NahG) that degrades SA compromisesresistance to the biotroph but not thenecrotrophs. These pathways appear tobe mutually inhibitory.

SA and JA induction of plant defensive gene (anti-microbial protein) expression
SA
NPR1
JA ET
EIN2
Defense response(PR genes)
Defense response(?)
Defense response( PDF1.2, Thi2.1,HEL, CHIB)
Erisyphesp.P.syringaeP.parasiticaX.campestris
(?)
wounding
Pythium sp.A.brassicicolaB.cinerea
E.carotovora
JAR1
Biotropic pathogen Necrotropic pathogen
Courtesy of Ji-Young Lee
66

Pieterse et al (2002) Plant Biology 4:535-544 67

68

Jasmonic Acid Signaling

Jasmonic acid
Jasmonic acid (JA)Isolated from plants and fungi
Inhibits:seed and pollen germinationroot growth
Stimulates:plant defenses against microbial and insect pathogenswound responses
ripeningexogenous application decreases expression of genesassociated with photosynthesis
70

First isolation of JAME→Jasminum grandiflorum
Jasmonates occurring : algae, mosses, fungi, gym-nosperms and angiosperms.
Remarkably high in fungiFusarium oxysporum : more than 20 jasmonatesAspergillus niger : 25 jasmonate compounds
Fungi the biological function of various jasmonates is unknown
Structural requirements for biological activityBased on different assays, such as alkaloid formation (Blechert et al., 1995), tendril coiling (Blechert et al., 1999), tuber formation (Koda, 1992), or gene expression (Miersch et al., 1999b), the follow-ing structural requirements have been found
Diversity among naturally occurring jasmonates and structure–activity relationships
71

ACTION OF JASMONATES
• Jasmonates as novel anti-cancer agents
• Jasmonates as signals in biotic and abiotic stresses
Solanaceae
Arabidopsis
• Jasmonates in development
Root growth inhibition
Tuber formation
Tendril coiling and touch
Flower development
Senescense
72

History
73

In 1962 : Jasmonic acid methyl ester was isolated for the first time from the essential oil of Jasminum grandiflorum.
In 1980 : Described a senescence-promoting effect of JA,
In 1981 : Growth inhibitorsroot growth became the most prominent assay for screening of mutants affected in respect of JA signalling
In 1984 : The biosynthesis of JA was elucidated
In 1987 : The synthesis of abundantly accumulating proteins, so-called jasmonatein-duced proteins (JIPs) and degradation of housekeeping proteins such as Rubisco were described
In 1990 : Another dramatic alteration of gene expression by JAME was observed in tomato plants proteinase (inhibitor (PIN2) was found to accumulate upon wounding (such as that caused by herbivore attack) or by treatment with airborne JAME)
In 1992 : JA-induced alteration of gene expression and metabolite patterns became the focus of studies examining accumulation of vegetative storage proteins
74

In 1993 : cloning of a JA biosynthetic enzyme, allene oxide synthase (AOS)
In 1994 : isolation of the first JA-insensitive mutant ( coronatineinsensitive1, Coi1)
In 1998 : COI1 codes for an F-box protein, which is part of the ubiquitin- ediated protea-some pathway The corresponding F-box protein in auxin signaling TIR1 was cloned.
In 2001 : The identification of the first transcription factors regulating JA-responsive gene expression
In 2002 : Other important steps in understanding the action of jasmonates were the identification of mutants affected in JA-biosynthesisThe cloning of JA amino acid conjugate synthase (JAR1)
In 2005 : The proof that fatty acid b-oxidation acts in JA biosynthesis
A number of reviews emphasizing different aspects of JA physiology have ap-peared in recent years. The following review will cover some recent results in biosynthesis, metabo-lism, signal transduction and action of jasmonates. I apologize for references not cited due to space limitations.
75

Biosynthesis of jasmonic acid
Linolenic acid (18:3) is released from membrane lipids and then converted into JA
JA induces transcription of host genes involved in plant defense. Example:Madagascar periwinkle (Cathranthus roseus) - transcription factor identified that responds to JA by activating expressions of genes encoding alkaloid biosynthetic genes - alkaloids: anticancer activity
76

Conversion of linolenic acid to jasmonic acid
77

Conversion of linolenic acid to jasmonic acid
78

Jasmonic acid is involved in defense against insects and
microbes
79
Jasmonic acid signaling
80
Skp1/Cullin/F-box (SCF COI1)
F-box subunit is encoded by CORONATINEINSENSITIVE 1 (COI1)
Jasmonate ZIM domain (JAZ) repressor proteins
Transcriptional activator MYC2

81

82

Tandem Affinity Purification (“TAP Tagging”)
Uses two tags, and sequential purifications to enhance specificity.
83

TAP Tagging• Specificity is the major challenge in pull-down assays.• Specificity may be enhanced using sequential tag-captures
and specific elutions.
•In the first round, protein complexes are captured via the first affinity tag (e.g., protein A and IgG Sepharose).
BAIT
PREYsepharose
84

TAP Tagging • These first-round captures are eluted by cleavage of a
specific protease tag.
BAIT
PREYsepharose
85
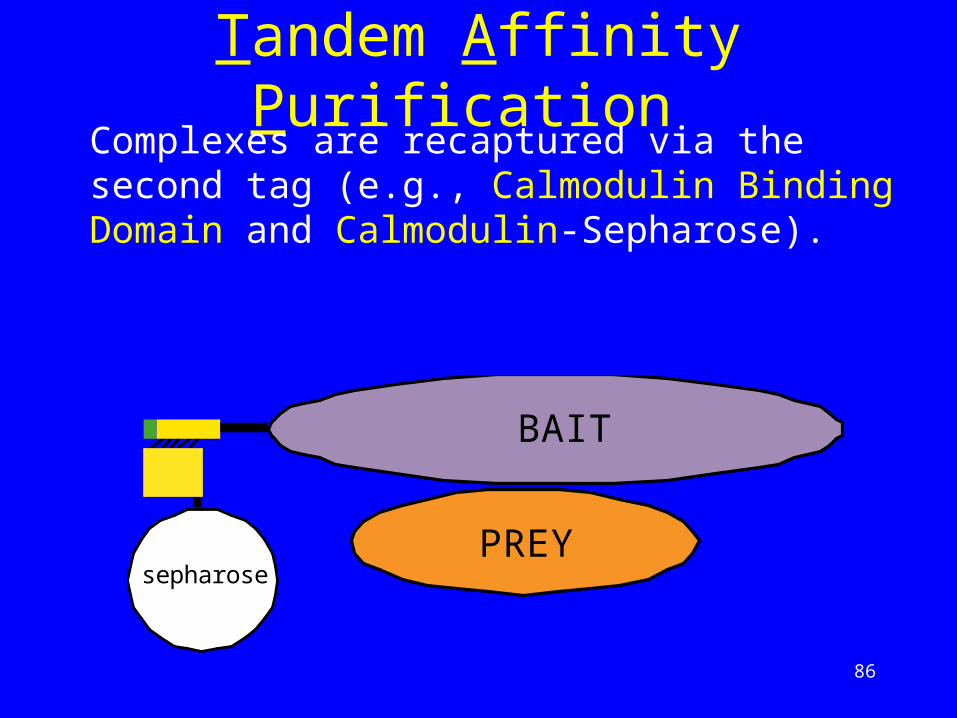
Tandem Affinity Purification Complexes are recaptured via the second tag (e.g., Calmodulin Binding Domain and Calmodulin-Sepharose).
sepharose
BAIT
PREY
86

Tandem Affinity Purification The second-round complex is then specifically eluted (e.g., by chelation of calcium to disrupt the CBD / Cm interaction).
sepharose
BAIT
PREY
87

Rationale for Using TAP/MS Method
1.Fast purification with high yield
2.In vivo
3.Over-expression of proteins not required
4.Prior knowledge of protein complex not required
5.Purified complex can be used in a variety of studies
88

The Tandem Affinity Purification Tag
target protein
target protein
89

TAP Purification Strategy
90

TAP Purification Strategy
91

TAP Purification Strategy
92

TAP Purification Strategy … in detail
1. PCR of the TAP cassette
2. Transformation of yeast cells (homologous recombination)
3. Selection of positive clones
4. Large-scale cultivation
5. Cell lysis Tandem affinity purification
6. One-dimensional SDS-PAGE
7. MALDI-TOF protein identification
8. Bioinformatic data interpretation
93

94
Yeast two-hybrid
• If the two proteins don't interact, the reporter gene remains inactive and the cells can't grow on -His plates:
• If the two proteins interact, the reporter gene (here: HIS3) is switched on and the diploids can grow on -His plates:

Animation 1
Animation 2
95

Molecular Mass Spectroscopy
96

Introduction
• In Mass Spectroscopy (MS), atomic and molecular weights are generally expressed in terms of atomic mass units (amu). The atomic mass unit is based on upon a relative scale in which the reference is the carbon isotope 126C, which is assigned a mass of exactly 12 amu. Thus the amu is defined as 1/12 of the mass of one neutral carbon atom.
97

Mass Spectroscopy
• Mass spectroscopy is perhaps one of the most widely applicable of all the analytical tools available to the analytical chemist in the sense that this technique is capable of providing information about
98

Technique is capable of providing information about
• the qualitative and quantitative composition of both organic and inorganic analytes in complex mixtures
• this instrument measures compounds with molecular masses up to 200, 000 Daltons.
• the structures of a wide variety of complex molecular species
• isotopic ratios of atoms in samples and• the structure and composition of solid
surfaces.
99

The four main components of a molecular mass spectrometer
100

The Sample Inlet System
• Batch Inlet Systems:
• These systems are the simplest and simply involve the volatilization of the sample externally and then the gradual leakage of the volatilized sample into the evacuated ionization chamber. For gases, the sample is introduced into the metering volume container and then expanded into the reservoir flask where it is then leaked into the ionization chamber. For liquids, a small quantity of sample is introduced into the reservoir and the pressure of the system is reduced to about 10-5 torr. The inlet system is lined with glass to avoid losses of polar analytes by adsorption.
• The Direct Probe Inlet:
• These systems are used for solids and non-volatile liquids and in these systems the sample is introduced into the ionization region by means of a sample holder, or probe, which is inserted through a vacuum lock. Probes are also used when the amount of the sample to be analyzed is small. With a probe, the sample is generally held on the surface of a glass or aluminum capillary tube and positioned within a few meters of the ionization source.
101

Magnetic Sector Analyzers• Magnetic sector analyzers employ a permanent magnet or
electromagnet to cause the beam from the ion source to travel in a circular path of 180, 90, or 60 degrees. Here, ions are formed by electron impact.
• The translational energy of an ion of mass m and charge z upon exciting slit B is given by
•K. = Zev = ½ mv2 Equation 1
• where V is the voltage between A and B, v is the velocity of the ion after acceleration, and e is the charge of the ion.
• The path in the sector described by the ions of a given mass and charge represents a balance between two forces acting upon them. These two forces are the centripetal force and the magnetic force and equating these two forces yields:
• Bzev = mv2/rwhich rearranges to v = Bzer/m• Substituting the above equation into (1) gives m/z =
B2r2e/2V• The last equation shows how mass spectra can be obtained
by varying one of the three variables B, V, or r. 102

Double Focusing Instruments
• These type of instruments, unlike single-focusing which simply minimize directional errors, are designed to limit both the errors introduced because ions are initially moving in different directions and also the errors introduced due to the fact that ions of the same mass-to-charge ratio may have different translational energies. 103

A schematic of a double-focusing instrument is shown
104

Time-of-Flight Analyzers
• In time-of-flight instruments, positive ions are produced periodically by bombardment of the sample with brief pulses of electrons, secondary ions or laser generated photons. The ions produced are then accelerated by an electric field and then made to pass into a field-free drift tube about a meter long. Because all ions entering the tube ideally have the same kinetic energies, their velocities in the tube must vary inversely with their masses, with the lighter particles arriving at the detector earlier than the heavier ones.
105

Isotopes
• By definition isotopes are atoms having the same atomic number but different mass numbers this technique is advantageous because in mass spectroscopy isotopes are easily differentiated. As depicted below: a mixture which contains isotopes are differentiated and shown in a spectrum.
106

Gas-Phase Sources • Gas-phase sources require volatilization of the
sample before ionization and thus are limited to thermally stable compounds that have boiling points less than about 500°C.
• Electron Impact Source : In the sources, electrons emitted from a filament are accelerated by a potential of about 70 V and made to collide with gaseous atoms or molecules of the sample causing ionization. Electron-impact ionization is not very efficient and only about one molecule in a million undergoes the primary reaction
M + e- = M.+ + 2e-• Electron Impact spectra are very complex due to
the high energies possessed by the accelerated electrons which collide with the sample and lead to fragmentation. These complex spectra are very useful for compound identification.
107

Advantages of Electron Impact sources
• They are convenient and produce high ion currents.
• Extensive fragmentation can lead to unambiguous identification of analytes.
108

Disadvantages of Electron Impact sources
• The need to volatilize the sample limits this method since it excludes analysis of thermally unstable compounds.
• Excessive fragmentation can lead to the disappearance of the molecular ion peak therefore preventing the molecular mass of the analyte to be determined.
109

Identification of Pure Compounds by Mass
Spectroscopy • Mass spectroscopy can be used to determine the molecular
weight of a compound but this involves an identification of a molecular peak and a comprehensive study of a spectrum.
• TANDEM MASS SPECTROSCOPY: This type of spectroscopy simply involves the coupling of one mass spectrometer to another and this hyphenated technique has resulted in dramatic progress in the analysis of complex mixtures.
•SECONDARY ION MASS SPECTROSCOPY: This is one of the most highly developed of the mass spectrometric surface methods, with several manufacturers offering instruments for this technique. It involves the bombarding of a surface with a beam of ions formed in an ion gun. The ions generated from the surface layer are then drawn into a spectrometer for mass analysis.
110

References
• TSRI Mass Spectrometry Home Page: http://masspec.scripps.edu/hist.html
• American Chemical Society:http://www.acs.org• Chemical Abstracts Service:http://www.cas.org• Chemical Center Home Page:
http://www.chemcenter/org• Science Magazine: http://www.sciencemag.org• Journal of Chemistry and Spectroscopy:• http://www.kerouac.pharm.uky.edu/asrg/wave/wavehp.html• http://www.uis.edu/trammell/che425/ms-2• http://www.usf.edu/~sl/mass _spec/MS_instrumentation.html• http://minyos.its.rmit.edu.au/~rcmfa/mstheory.html• http://www.cem.msu.edu/~reusch/VirtualText/Spectrpy/Mas
sSpec/masspec1.htm• http://www.abrf.org/ABRFNews/1996/September1996/sep96i
ontrap.html 111