
Nicotiana plumbaginifolia hlg mutants have a mutation in a PHYB-type phytochrome gene: they have...
11
The Plant Journal (1997) 12(5), 1091-1101 Nicotiana plumbaginifolia hlg mutants have a mutation in a PHYB-type phytochrome gene: they have elongated hypocotyls in red light, but are not elongated as adult plants Matthew Hudson 1,*, Paul R.H. Robson 1, Yvan Kraepiel 2,t, Michel Caboche 2,~ and Harry Smith 1 7Department of Botany, University of Leicester, Leicester LE1 7RH, UK, and 2Biologie Cellulaire, INRA Versailles, F-78026 Versailles Cedex, France Summary Two new allelic mutants of Nicotiana plumbaginifolia have been isolated which d~splay a hypocotyl which is long (hlg) when seedlings are grown in continuous white light (W). This can be accounted for by the decreased response to red light (R) of the hypocotyl elongation rate in these mutants. Responses to other wavelengths are unaffected in the mutants. When grown in white light, mature hlg mutants are not elongated with respect to the wild- type; they also bolt and flower later. The shade-avoidance responses to red/far red ratio (R:FR) are intact in these mutants. Both mutants are deficient in a phyB-like poly- peptide that is immunodetectable in the wild-type; both have wild-type levels of a phyA-like polypeptide. These alleles are inherited in a partially dominant manner, and correspond to single-base missense mutations in a gene highly homologous to IV. tabacum PHYB, which codes for a phytochrome B-type photoreceptor. One allele, hlg-1, has an introduced amino acid substitution; this may define a residue essential for phytochrome protein stability. The other allele, big-2, has a stop codon introduced C-terminal to the chromophore binding domain. As these phyB mutants are unaffected in shade-avoidance responses, but deficient in perception of R, it is concluded that the phyB absent in these mutants is responsible for R perception in the N. plumbaginifolia seedling, but is not a R:FR sensor in light-grown plants. Introduction Phytochromes are a family of biliproteins which exist in two forms (Pr and Pfr), interconvertible by red (R) and Received 14April 1997;revised 20 June 1997; accepted 3 July 1997. *For correspondence (fax+44 116252 2791;[email protected]). tCurrent address:Physiologie du D~veloppement des Plantes, Tour 53, Universit~ Pierre et Marie Curie, 4 Place Jussieu, 75252 Paris Cedex 05, France. tCurrentaddress: Plant Homeostasis Group, RIKEN, 2-1Hirosawa, Wako-shi, Saitama 351-01,Japan. far-red (FR) light. They control numerous morphogenic and biochemical processes in plants, many of which are associated with the process of de-etiolation. Two classes of phytochrome have been recognized. Light-labile or type I phytochromes are represented by phytochrome A from oat, Arabidopsis and other plants. Other phytochrome genes have been shown to code for light-stable or type II phytochromes, including the phytochrome B gene (PHYB) in Arabidopsis, and closely homologous genes isolated from numerous other plants (for in-depth reviews, see Koornneef and Kendrick, 1994; Quail, 1994; Whitelam and Harberd, 1994). The functions of the phytochromes have been investi- gated with the aid of several mutants of different species. Some clearly have a lesion in one of the genes encoding the phytochrome apoproteins, while others appear to be deficient in the biosynthesis of the tetrapyrrole chromo- phore (Whitelam and Harberd, 1994). Phytochrome apo- protein mutants are characterized by deficiencies in photomorphogenic responses and a partial dominance of the mutant allele (Whitelam and Harberd, 1994), which is almost certainly due to an effect of gene copy number on phytochrome level (Wester et al., 1994). Mutants in the Arabidopsis PHYB gene have been extensively charac- terised (Koornneef etal., 1980; McCormac etal., 1993; Reed et aL, 1994; Somerset aL, 1991) and were until recently the only light-stable phytochrome mutants in which lesions within the native apoprotein gene had been identified and sequenced (Bradley et aL, 1996; Reed et aL, 1993). Mutants displaying a deficiency in a gel-blot band detectable with monoclonal antibodies to a phytochrome-B-like apoprotein have been isolated in cucumber (L6pez-Juez et aL, 1992), Brassica rapa (Devlin et aL, 1992), pea (Weller et aL, 1995) and Sorghum bicolor (Childs et aL, 1992; Foster et aL, 1994). Very recently, the Sorghum Ma3 R and Brassica rapa ein mutants have been confirmed as phyB mutants (Childs et aL, 1997; Devlin et aL, 1997). All the above mutants characteristically show a decreased sensitivity to continu- ous R as seedlings, which manifests itself in white light (W) as an elongated phenotype. Phytochrome B (phyB) has also been overexpressed in Arabidopsis, under the control of a viral promoter (Wagner et al., 1991). The light-dependent short hypocotyl pheno- type of these transformants is R-specific. In addition, these transgenic lines were shown to have short hypocotyls relative to the wild-type at all R:FR ratios, and their sensitiv- 1091
-
Upload
matthew-hudson -
Category
Documents
-
view
212 -
download
0
Transcript of Nicotiana plumbaginifolia hlg mutants have a mutation in a PHYB-type phytochrome gene: they have...

The Plant Journal (1997) 12(5), 1091-1101
Nicotiana plumbaginifolia hlg mutants have a mutation in a PHYB-type phytochrome gene: they have elongated hypocotyls in red light, but are not elongated as adult plants
Matthew Hudson 1,*, Paul R.H. Robson 1, Yvan Kraepiel 2,t, Michel Caboche 2,~ and Harry Smith 1 7Department of Botany, University of Leicester, Leicester LE1 7RH, UK, and 2Biologie Cellulaire, INRA Versailles, F-78026 Versailles Cedex, France
Summary
Two new allelic mutants of Nicotiana plumbaginifolia have been isolated which d~splay a hypocotyl which is long (hlg) when seedlings are grown in continuous white light (W). This can be accounted for by the decreased response to red light (R) of the hypocotyl elongation rate in these mutants. Responses to other wavelengths are unaffected in the mutants. When grown in white light, mature hlg mutants are not elongated with respect to the wild- type; they also bolt and flower later. The shade-avoidance responses to red/far red ratio (R:FR) are intact in these mutants. Both mutants are deficient in a phyB-like poly- peptide that is immunodetectable in the wild-type; both have wild-type levels of a phyA-like polypeptide. These alleles are inherited in a partially dominant manner, and correspond to single-base missense mutations in a gene highly homologous to IV. tabacum PHYB, which codes for a phytochrome B-type photoreceptor. One allele, hlg-1, has an introduced amino acid substitution; this may define a residue essential for phytochrome protein stability. The other allele, big-2, has a stop codon introduced C-terminal to the chromophore binding domain. As these phyB mutants are unaffected in shade-avoidance responses, but deficient in perception of R, it is concluded that the phyB absent in these mutants is responsible for R perception in the N. plumbaginifolia seedling, but is not a R:FR sensor in light-grown plants.
Introduction
Phytochromes are a family of biliproteins which exist in two forms (Pr and Pfr), interconvertible by red (R) and
Received 14 April 1997; revised 20 June 1997; accepted 3 July 1997. *For correspondence (fax +44 116 252 2791; e-mail [email protected]). tCurrent address: Physiologie du D~veloppement des Plantes, Tour 53, Universit~ Pierre et Marie Curie, 4 Place Jussieu, 75252 Paris Cedex 05, France. tCurrent address: Plant Homeostasis Group, RIKEN, 2-1 Hirosawa, Wako-shi, Saitama 351-01, Japan.
far-red (FR) light. They control numerous morphogenic and biochemical processes in plants, many of which are associated with the process of de-etiolation. Two classes of phytochrome have been recognized. Light-labile or type I phytochromes are represented by phytochrome A from oat, Arabidopsis and other plants. Other phytochrome genes have been shown to code for light-stable or type II phytochromes, including the phytochrome B gene (PHYB) in Arabidopsis, and closely homologous genes isolated from numerous other plants (for in-depth reviews, see Koornneef and Kendrick, 1994; Quail, 1994; Whitelam and Harberd, 1994).
The functions of the phytochromes have been investi- gated with the aid of several mutants of different species. Some clearly have a lesion in one of the genes encoding the phytochrome apoproteins, while others appear to be deficient in the biosynthesis of the tetrapyrrole chromo- phore (Whitelam and Harberd, 1994). Phytochrome apo- protein mutants are characterized by deficiencies in photomorphogenic responses and a partial dominance of the mutant allele (Whitelam and Harberd, 1994), which is almost certainly due to an effect of gene copy number on phytochrome level (Wester et al., 1994). Mutants in the Arabidopsis PHYB gene have been extensively charac- terised (Koornneef etal., 1980; McCormac etal., 1993; Reed et aL, 1994; Somerset aL, 1991) and were until recently the only light-stable phytochrome mutants in which lesions within the native apoprotein gene had been identified and sequenced (Bradley et aL, 1996; Reed et aL, 1993). Mutants displaying a deficiency in a gel-blot band detectable with monoclonal antibodies to a phytochrome-B-like apoprotein have been isolated in cucumber (L6pez-Juez et aL, 1992), Brassica rapa (Devlin et aL, 1992), pea (Weller et aL, 1995) and Sorghum bicolor (Childs et aL, 1992; Foster et aL, 1994). Very recently, the Sorghum Ma3 R and Brassica rapa ein mutants have been confirmed as phyB mutants (Childs et aL, 1997; Devlin et aL, 1997). All the above mutants characteristically show a decreased sensitivity to continu- ous R as seedlings, which manifests itself in white light (W) as an elongated phenotype.
Phytochrome B (phyB) has also been overexpressed in Arabidopsis, under the control of a viral promoter (Wagner et al., 1991). The light-dependent short hypocotyl pheno- type of these transformants is R-specific. In addition, these transgenic lines were shown to have short hypocotyls relative to the wild-type at all R:FR ratios, and their sensitiv-
1091

1092 Matthew Hudson et al.
ity to changes in R:FR was not enhanced (McCormac et al., 1993).
A general opinion has developed that phyB plays a dominant role in mediating shade-avoidance responses (e,g. Koornneef and Kendrick, 1994; Quail, 1994) This con- clusion is based on the superficial resemblance of phyB mutants in Arabidopsis to shade-avoiding plants. The shade-avoidance response represents a competitive phyto- chrome-mediated response in light-grown plants. It is char- acterized by early bolting and/or flowering, elongation of internodes and petioles, and an increase in the rate of overall elongation, in response to low R:FR (Smith, 1994). As light-grown adults, previously isolated mutants deficient in phyB-like phytochromes appear phenotypically similar when grown in high R:FR to wild-type plants grown in low R:FR. They typically show a reduced response to low R:FR, but the responses are not eliminated (Devlin et aL, 1992; Robson etal., 1993; Smith etaL, 1992; Whitelam and Smith, 1991; van Tuinen et aL, 1995) demonstrating that phyB is not essential for shade-avoidance.
The PHYB gene of Arabidopsis has an ortholog in the PHYD gene, which is thought to belong to a sub-family of genes encoding light-stable phyB-like photoreceptors (Clack et al., 1994). The sequences of PCR fragments of two PHYB-like genes from tomato are more closely related to each other than to the corresponding regions of the Arabidopsis genes PHYB and PHYD, and they have con- sequently been referred to as PHYB-1 and PHYB-2. It has been suggested on the basis of this evidence that the phylogeny of the phyB-like sub-family in the Solanaceae is at variance with that in Arabidopsis (Pratt et al., 1995). If this is the case, then any division of function between light-stable phytochromes in Arabidopsis may not hold true in other plant families.
A tomato mutant, tri, deficient in a phyB-type apoprotein, has aberrant PHYB-1 mRNAs (Kerckhoffs et aL, 1996; van Tuinen et aL, 1995). Although this mutant appears to show a deficiency in the response to R as a seedling, this deficiency is described as temporary. However, the tri mutants elongate more rapidly than wild-type plants when grown in W of high R:FR, although this phenotype is not as marked as in other phyB mutants (van Tuinen et aL, 1995). In contrast, potato plants in which PHYB gene expression has been ablated by antisense techniques show a considerably more severe phenotype. They develop very elongated internodes in W, and show lower levels of CAB 1 gene expression than that characteristic of wild-type plants. They also display aberrant photoperiodic responses (Jackson et al., 1996).
Here we present two allelic mutants of Nicotiana plumb- aginifolia which display an elongated hypocotyl as seedlings grown in W or R. We show that these mutants, in contrast to other mutants deficient in phyB-type phytochromes, do not show any component of the shade-
avoidance response when grown in high R:FR, and retain normal responses to reduced R:FR.
Results
Mutant isolation and prefiminary characterisation
The hlg mutant alleles were isolated at INRA (Versailles, France) from an ethyl methanesulphonate (EMS) muta- genised M2 population of N. plumbaginifolia background Pblhd. The mutants displayed elongated hypocotyls under fluorescent W with a 16 h photoperiod. Mutants isolated in this way were referred to as hlg mutants (hypocotyle long), a phenotype distinct from the partially etiolated pew mutants (Kraepiel et al., 1994). Two mutant lines, hlg-I and hlg-2, are investigated here. At Leicester, we confirmed that hlg-1 and hlg-2 were allelic by showing that no wild- type plants were observed in a population of approximately 500 seedlings of the hlg-I x hlg-2 F2 (data not shown).
PHYB mRNA is present at wild-type levels in the mutants hlg-1 and hlg-2, as is the phytochrome regulated CAB mRNA, and both hlg alleles are deficient in a phyB-type apoprotein
The level of phytochrome B transcript in the mutants was investigated by RNA gel blotting of mRNA extracted from light-grown leaf tissue of the mutant and wild-type plants at the rosette stage. A gene-specific radiolabelled probe was derived from an RT-PCR product amplified from wild- type N. plumbaginifolia cDNA using primers designed by homology to the published Nicotiana tabacum PHYB gene. It can be seen from the poly(A) mRNA blots in Figure l(a) that the level of PHYB transcript in the mutants, when compared with the ubiquitin loading control, is equivalent to that in the wild-type.
It can be seen from the total RNA blot also shown in Figure l(a) that the phytochrome regulated CAB 1 gene message (Castresana et al., 1987) is also the correct size and present at wild-type levels, in light-grown leaf tissue of both mutants at the mid-point of the light period.
Figure l(b) shows immunoblots of protein extracts from dark-adapted leaf tissue from the hlg mutants and the wild- type, which have either been maintained in darkness or exposed to continuous R for 24 h. It can be seen from the uppermost blot, probed with AS-32, a monoclonal antibody to phyA (Holdsworth, 1987), that the mutants have a high phyA apoprotein level in the dark but that the protein is light-labile, indicating that it is photochemically active. Figure l(b) also shows two blots which were probed with antibodies raised to N. tabacum phyB, one (centre) polyclonal (PMTB, supplied by T. Kunkel), the other (bottom) monoclonal (mAT-l, Lopez-Juez etal., 1992). Both blots show that a phyB co-immunoreactive protein is
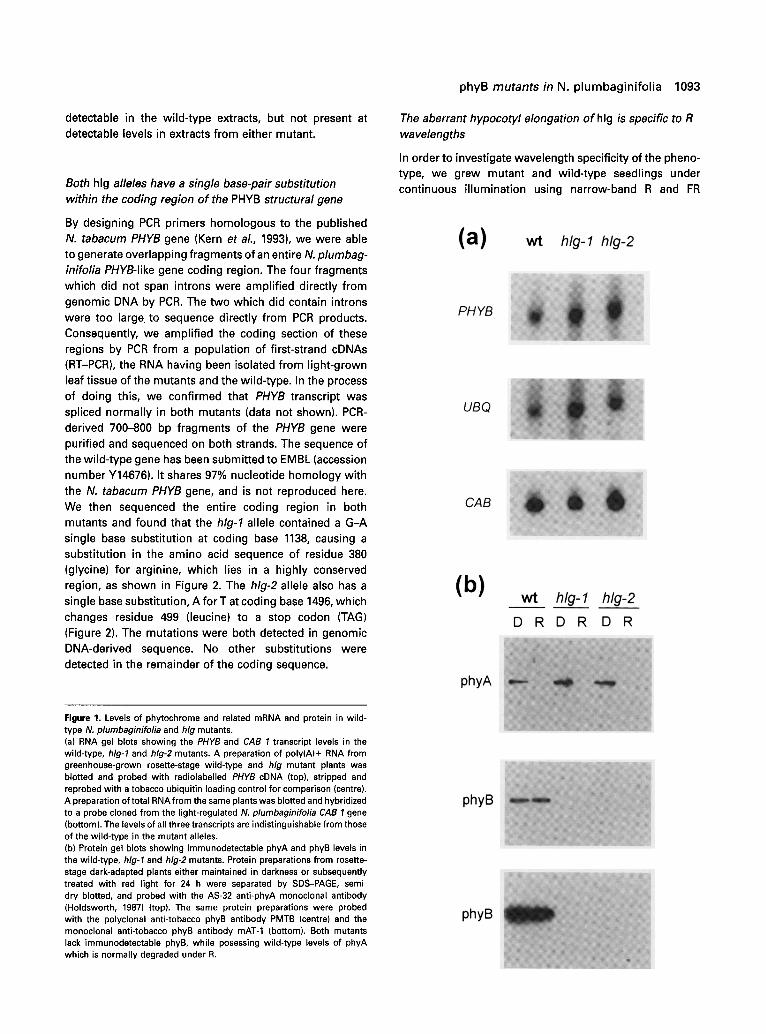
detectable in the wild-type extracts, but not present at detectable levels in extracts from either mutant.
Both hlg alleles have a single base-pair substitution within the coding region of the PHYB structural gene
By designing PCR primers homologous to the published N. tabacum PHYB gene (Kern et aL, 1993), we were able to generate overlapping fragments of an entire N. plumbag- inifolia PHYB-like gene coding region. The four fragments which did not span introns were amplified directly from genomic DNA by PCR. The two which did contain introns were too large, to sequence directly from PCR products. Consequently, we amplified the coding section of these regions by PCR from a population of first-strand cDNAs (RT-PCR), the RNA having been isolated from light-grown leaf tissue of the mutants and the wild-type. In the process of doing this, we confirmed that PHYB transcript was spliced normally in both mutants (data not shown). PCR- derived 700-800 bp fragments of the PHYB gene were purified and sequenced on both strands. The sequence of the wild-type gene has been submitted to EMBL (accession number Y14676). It shares 97% nucleotide homology with the N. tabacum PHYB gene, and is not reproduced here. We then sequenced the entire coding region in both mutants and found that the hlg-1 allele contained a G-A single base substitution at coding base 1138, causing a substitution in the amino acid sequence of residue 380 (glycine) for arginine, which lies in a highly conserved region, as shown in Figure 2. The hlg-2 allele also has a single base substitution, A for T at coding base 1496, which changes residue 499 (leucine) to a stop codon (TAG) (Figure 2). The mutations were both detected in genomic DNA-derived sequence. No other substitutions were detected in the remainder of the coding sequence.
phyB mutants in N. p lumbag in i fo l ia 1093
The aberrant hypocotyl elongation of hlg is specific to R wavelengths
In order to investigate wavelength specificity of the pheno- type, we grew mutant and wild-type seedlings under continuous il lumination using narrow-band R and FR
Figure 1. Levels of phytochrome and related mRNA and protein in wild- type IV. plumbaginifo/ia and hlg mutants. (a) RNA gel blots showing the PHYB and CAB 1 transcript levels in the wild-type, hlg-1 and hlg-2 mutants. A preparation of poly(A)+ RNA from greenhouse-grown rosette-stage wild-type and hlg mutant plants was blotted and probed with radiolabelled PHYB cDNA (top), stripped and reprobed with a tobacco ubiquitin loading control for comparison (centre). A preparation of total RNA from the same plants was blotted and hybridized to a probe cloned from the light-regulated N. plumbaginifolia CAB 1 gene (bottom). The levels of all three transcripts are indistinguishable from those of the wild-type in the mutant alleles. (b) Protein gel blots showing immunodetectable phyA and phyB levels in the wild-type, hlg-1 and hlg-2 mutants. Protein preparations from rosette- stage dark-adapted plants either maintained in darkness or subsequently treated with red light for 24 h were separated by SDS-PAGE, semi- dry blotted, and probed with the AS-32 anti-phyA monoclonal antibody (Holdsworth, 1987) (top). The same protein preparations were probed with the polyclonal anti-tobacco phyB antibody PMTB (centre) and the monoclonal anti-tobacco phyB antibody mAT-1 (bottom). Both mutants lack immunodetectable phyB, while posessing wild-type levels of phyA which is normally degraded under R.

1094 Matthew Hudson et al.
hlg mutants is visible in white light at the seedling stage, little difference is observed between the mutants and the wild-type at the rosette stage. The mutants may have a more highly branched root system. However, we have not observed the long root hairs seen in Arabidopsis phyB mutants by Reed et al. (1993) (data not shown).
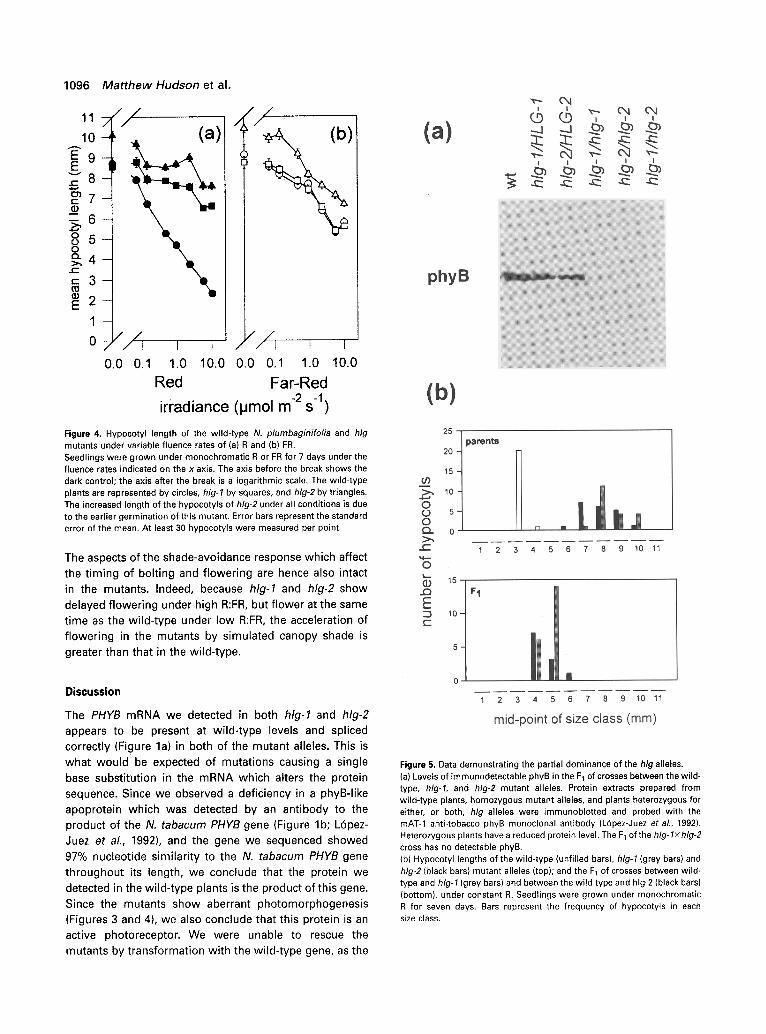
Red light causes separate responses to very-low-fluence, Iow-fluence and high-irradiance treatments (Mancinelli, 1994). It can be seen from Figure 4(a) that the irradiance- dependent inhibition of hypocotyl elongation by R in the mutants is substantially reduced, while the irradiance- dependent inhibition of hypocotyl elongation by FR appears to be normal (Figure 4b). At high fluence rates, an R response can still be seen in the mutant. The hlg-2 mutant again displays a longer hypocotyl under all conditions. This effect is due to a second site mutation in the hlg-2 line (data not shown).
Figure 2. Mutations in the phyB polypeptide of the hlg- 1 and hlg-2 mutants. The loci of the mutations hlg-1 and hlg-2were detected by DNA sequencing and comparison with the wild-type gene. The upper part of the figure shows the change in the deduced amino acid sequence of the phyB polypeptide in both alleles. The position of the truncation of the phyB polypeptide in the hlg-2 mutant is indicated. The missense hlg-I mutation is also shown together with a Clustal alignment of the conserved region in which this mutation lies, showing selected phytochrome sequences from disparate plant taxonomical groups. The phyA-109 mutation in the Arabidopsis PHYA gene, which also causes loss of immunodetectable protein (Xu et al, 1995) is also shown.
sources, a broad-band B source and fluorescent W, along with dark-grown controls.
Figure 3(a) shows that the hlg mutant hypocotyl pheno- type is only visible in W and R. The hypocotyl is longer, root development is inhibited and the cotyledons are smaller and relatively pale in both alleles of the mutant, in comparison with the wild-type. However, the mutants are by no means blind to R, with hypocotyl hook opening, cotyledon expansion and greening occurring in response to R. There is also some inhibition of hypocotyl elongation in R relative to the dark control. Normal photomorphogen- esis is seen in response to FR and B, and wild-type skotomorphogenesis in darkness. It is not possible to distinguish between the dark controls of the hlg-1 mutant and the wild-type hypocotyls. However, the hlg-2 mutant can be seen to have slightly longer hypocotyls than the wild-type under all conditions.
Figure 3(b) Shows that, although the phenotype of the
Both alleles display a partially dominant effect on both the phyB-like apoprotein level and the inhibition of hypocotyl elongation by R
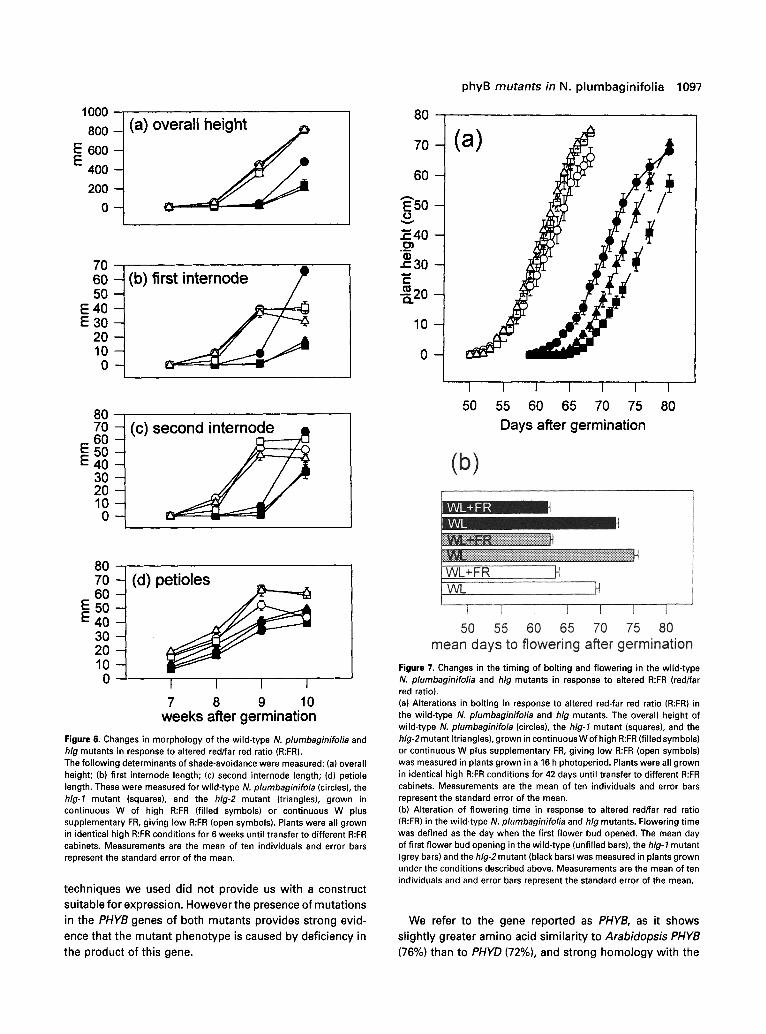
The effect of the hlg-HLG heterozygous condition can be seen in Figure 5(a), with the heterozygotes containing a noticeably reduced amount of immunodetectable phyB- type apoprotein. The heterozygotes of both alleles display a lessened response to R in terms of the inhibition of hypocotyl elongation, as shown by the hypocotyl length frequency plot in Figure 5(b). This is consistent with a PHYB gene dosage effect, as seen in Arabidopsis by Wester et aL (1994).
The hlg mutants show no aspect of shade-avoidance as adults grown in high R:FR, and show no deficiency in the response to low R:FR
When the rosette-stage hlg mutant alleles are grown in continuous high R:FR W together with wild-type N. plumb- aginifolia, as shown in Figure 6, both overall height and internode length in the mutants are shorter than in the wild-type plants, and petiole length appears identical in the wild-type and in the mutants. Under low R:FR W, the mutants display a similar internode length and overall height to the wild-type, and have longer petioles. The morphogenic aspects of the response to low R:FR (the shade-avoidance response; Smith, 1994) are therefore seen to a greater extent in the mutants than they are in the wild- type. When the mutants are grown in high R:FR W with a 16 h photoperiod, as shown in Figure 7, they are observed to be slightly later in bolting and flowering than the wild- type plants. This difference is lost when FR light is added to reduce the R:FR, and both the mutant alleles and the wild-type plants are substantially earlier to bolt, accelerated in bolt growth, and earlier to flower, under a lower R:FR.

p h y B m u t a n t s in N. p l u m b a g i n i f o l i a 1095
Figure 3. Gross phenotype of the wild-type N. plumbaginifolia and hlg mutants as seedlings and adults. (a) Seedling morphology of the wild-type and the hlg-1 and hlg-2 mutants under different light conditions. Seedlings were grown under fluorescent W, monochromatic B, R or FR, or in the dark for 7 days (see Experimental procedures). The long hypocotyl phenotype of the mutant alleles is clearly visible in R, and visible to some extent in W. The mutants show a wild-type response to B and FR. (b) Phenotype of the hlg mutants and the wild-type grown in W for 60 days post germination. The plants shown were grown in a greenhouse at high PAR and R:FR for 60 days before the roots were washed and the plants photographed. The mutants are not elongated, and display a more branched root system than the wild-type plants.
1096 Matthew Hudson et al.
11
10
E 9 E
~ 6 8 5 - o
~ 4 - ¢ -
c 3 -
E 2 -
1
q
/•/// 8
0 7 ~/I I I / ~Jl I I 0.0 0.1 1.0 10.0 0.0 0.1 1.0 10.0
Red Far-Red irradiance (pmol m -2 s -1)
Figure 4. Hypocotyl length of the wild-type N. plumbaginifolia and hlg mutants under variable fluence rates of (a) R and (b) FR. Seedlings were grown under monochromatic R or FR for 7 days under the fluence rates indicated on the x axis. The axis before the break shows the dark control; the axis after the break is a logarithmic scale, The wild-type plants are represented by circles, hlg-1 by squares, and hlg-2 by triangles. The increased length of the hypocotyls of hlg-2 under all conditions is due to the earlier germination of this mutant, Error bars represent the standard error of the mean. At least 30 hypocotyls were measured per point.
The aspects of the shade-avoidance response which affect the timing of bolting and flowering are hence also intact in the mutants. Indeed, because hlg-I and hlg-2 show delayed flowering under high R:FR, but flower at the same time as the wild-type under low R:FR, the acceleration of flowering in the mutants by simulated canopy shade is greater than that in the wild-type.
Discussion
The PHYB mRNA we detected in both hlg-1 and hlg-2 appears to be present at w i ld - type levels and spliced
correct ly (Figure la) in both of the mutant alleles. This is
what wou ld be expected of mutat ions causing a single
base subst i tut ion in the mRNA which alters the protein
sequence. Since we observed a def ic iency in a phyB-l ike
apoprote in which was detected by an ant ibody to the
product of the N. tabacum PHYB gene (Figure lb ; Lopez-
Juez et al., 1992), and the gene we sequenced showed
97% nucleot ide s imi lar i ty to the N. tabacum PHYB gene
th roughou t its length, we conclude that the protein we
detected in the wi ld- type plants is the product of this gene.
Since the mutants show aberrant pho tomorphogenes is
(Figures 3 and 4), we also conclude that this protein is an
active photoreceptor. We were unable to rescue the
mutants by t ransformat ion wi th the wi ld - type gene, as the
Figure 5. Data demonstrating the partial dominance of the hlg alleles. (a) Levels of immunodetectable phyB in the F1 of crosses between the wild- type, hlg-1, and hlg-2 mutant alleles. Protein extracts prepared from wild-type plants, homozygous mutant alleles, and plants heterozygous for either, or both, hlg alleles were immunoblotted and probed with the mAT-1 anti-tobacco phyB monoclonal antibody (L6pez-Juez et aL, 1992). Heterozygous plants have a reduced protein level. The F~ of the hlg-l×hlg-2 cross has no detectable phyB. (b) Hypocotyl lengths of the wild-type (unfilled bars), hlg-1 (grey bars) and hlg-2 (black bars) mutant alleles (top); and the F~ of crosses between wild- type and hlg-1 (grey bars) and between the wild-type and hlg-2 (black bars) (bottom), under constant R. Seedlings were grown under monochromatic R for seven days. Bars represent the frequency of hypocotyls in each size class.
p h y B mutan ts in N. p l u m b a g i n i f o l i a 1097
Figure 6. Changes in morphology of the wild-type N. plumbaginifo/ia and hlg mutants in response to altered red/far red ratio (R:FR). The following determinants of shade-avoidance were measured: (a) overall height; (b) first internode length; (c) second internode length; (d) petiole length. These were measured for wild-type N. p/umbaginifola (circles), the hlg-1 mutant (squares), and the h/g-2 mutant (triangles), grown in continuous W of high R:FR (filled symbols) or continuous W plus supplementary FR, giving low R:FR (open symbols). Plants were all grown in identical high R:FR conditions for 6 weeks until transfer to different R:FR cabinets. Measurements are the mean of ten individuals and error bars represent the standard error of the mean.
t e c h n i q u e s w e used d id no t p r o v i d e us w i t h a cons t r uc t
s u i t a b l e f o r e x p r e s s i o n . H o w e v e r the p r e s e n c e o f m u t a t i o n s
in the PHYB g e n e s o f bo th m u t a n t s p r o v i d e s s t r o n g ev id -
ence t ha t t he m u t a n t p h e n o t y p e is caused by d e f i c i e n c y in
the p r o d u c t o f th is gene .
Figure 7. Changes in the timing of bolting and flowering in the wild-type N. plumbaginifolia and hlg mutants in response to altered R:FR (red/far red ratio). (a) Alterations in bolting in response to altered red-far red ratio (R:FR) in the wild-type N. p/umbaginifolia and h/g mutants. The overall height of wild-type N. plumbaginifo/a (circles), the hlg-1 mutant (squares), and the h/g-2mutant (triangles), grown in continuous W of high R:FR (filled symbols) or continuous W plus supplementary FR, giving low R:FR (open symbols) was measured in plants grown in a 16 h photoperiod. Plants were all grown in identical high R:FR conditions for 42 days until transfer to different R:FR cabinets. Measurements are the mean of ten individuals and error bars represent the standard error of the mean. (b) Alteration of flowering time in response to altered red/far red ratio (R:FR) in the wild-type N. p/umbaginifo/ia and hlg mutants. Flowering time was defined as the day when the first flower bud opened. The mean day of first flower bud opening in the wild-type (unfilled bars), the h/g-1 mutant (grey bars) and the hlg-2 mutant (black bars) was measured in plants grown under the conditions described above. Measurements are the mean of ten individuals and and error bars represent the standard error of the mean.
We re fe r to the g e n e r e p o r t e d as PHYB, as it s h o w s
s l i gh t l y g r e a t e r a m i n o acid s im i l a r i t y to Arabidopsis PHYB (76%) t h a n to PHYD (72%), and s t r ong h o m o l o g y w i t h the

1098 Matthew Hudson et al.
N. tabacum PHYB gene. Despite N. plumbaginifolia PHYB showing 92% similarity to the published fragment of tomato PHYB-1 and 88% similarity to the PHYB-2fragment at the amino acid level (M.E. Hudson, unpublished results; Pratt et aL, 1995) we have not named it PHYB-1, as these results may be different when the full tomato sequences are known. However, this does not discount the possibility of the presence of additional PHYB-type loci in the genome of N. plumbaginifolia.
The hlg-2 allele is a missense mutation introducing a stop codon which truncates the open reading-frame in the PHYB gene to less than half its normal length, removing the C-terminus from the conceptually translated protein (Figure 2). The C-terminus of Avena phytochrome A must be present for it to be functional (Cherry et al., 1992), and the same is thought to be true of Arabidopsis phytochrome B (Quail et al., 1995). Many apparently null alleles of hy3 in Arabidopsis contain stop codons which truncate the C- terminus of phyB (Bradley et aL, 1996; Reed et al., 1993). The hlg-2 mutant plants are hence unlikely to produce any functional phytochrome from this gene, and can be regarded as null. We did not detect the truncated protein which might be expected from such a stop codon mutation, possibly because it was unstable or did not contain the epitopes to which the antibodies were raised.
The phyB apoprotein signal in the hlg-1 mutant is undetectably low (Figure lb), which is an uncommon result of mutations causing single amino-acid changes in phytochromes (Reed et aL, 1993; Quail et aL, 1995; Wagner and Quail, 1995; Xu et aL, 1995). It is possible that the signal from the monoclonal antibody mAT-1 is absent due to alteration of the epitope rather than the absence of protein. However, since this result is repeated with the polyclonal antibody PMTB, our conclusion is that hlg-1 is deficient in this protein. The hlg-1 allele is an amino-acid substitution in a region C-terminal to the chromophore-attachment site, shown in Figure 2, which is highly conserved throughout plant phytochromes. A mutation in this region in the Arabidopsis PHYA gene is known to cause decreased stability of the phyA apoprotein (phyA-109; Xu et aL, 1995). Furthermore, the N-terminal chromophore-bearing domain is known to regulate the differential light-lability of phytochromes (Wagner et al., 1996). The hlg-I mutation may define a motif which is involved in this process, or in otherwise regulating phytochrome stability.
The amino acid which is substituted in the hlg-1 allele, residue 380, is not conserved throughout the plant kingdom like many of the neighbouring residues in the conserved motif which contains it, as shown in Figure 2. This residue is a cysteine in phytochromes from ferns and mosses to Arabidopsis, but a glycine in Arabidopsis PHYE, the PHYB genes from potato, N. tabacum and N. plumbaginifolia, and tomato PHYB-1 and PHYB-2
(Figure 2). As we have shown that this residue plays an essential role in phyB stability, it is tempting to speculate that the substitution of glycine for cysteine has some functional significance.
The hlg mutant phenotype at the seedling level correlates well with other known and putative phyB- deficient mutants (Whitelam and Harberd, 1994; Smith, 1995). It displays a specific deficiency in R responses, while retaining some responsiveness to R and having a wild-type phenotype in other monochromatic wavelengths (Figures 3a and 4). It follows that what is thought to be the major role of phyB in dicotyledonous seedlings, de- etiolation in response to R, appears to be highly conserved.
The phenotype of the hlg mutants as adults, however, is inconsistent with the phenotypes of other known or suspected mutants deficient in phyB (Koornneef and Kendrick, 1994; Smith, 1995). An obvious characteristic of the Arabidopsis and Sorghum phyB mutants is that they are early-flowering. This is not seen in the hlg mutant alleles, which are late-flowering (Figure 7). Unlike phyB-deficient mutants of Arabidopsis (Goto et al., 1991; Whitelam and Smith 1991), these mutants do not display a lessened response of flowering time to changes in R:FR (Figure 7). Since N. plumbaginifolia is day-neutral, this may indicate that the aberrant flowering responses of phyB mutants in non-day-neutral backgrounds may be due to effects of the absence of phyB on photo- periodism rather than shade-avoidance.
The results presented here (Figures 3b, 6 and 7) show that the phyB affected by the hlg mutations is not responsible for R:FR perception in light grown adults. The hlg mutants appear to have responses to R:FR which are, if anything, more sensitive than those of the wild- type plants (Figures 6 and 7). However, the phyB in which these alleles are deficient is important in mediating the responses of the seedling to R, and is analogous in this respect, and in its reactivity to mAT-l, to the mutated photoreceptors in previously described phyB-deficient mutants. Unlike Arabidopsis, adult N. plumbaginifolia does not display obvious elongation when grown under low irradiance W or briefly transferred to darkness (M.E. Hudson, unpublished data). Consequently, the shade avoidance responses in the adult hlg mutants may not be masked by constitutive elongation caused by the loss of R perception, unlike the case in other phyB mutants. While the overexpression of phyB in Arabidopsis gives rise to plants which have a reduced sensitivity to small changes in R:FR (McCormac et al., 1993), these transgenic plants were shown to be much more sensitive to changes in R irradiance. In Arabidopsis too, therefore, there is no evidence that phyB behaves in the way expected of a R:FR sensor. We can therefore propose that the role of

phyB is l imi ted to the percept ion of R, and that it does
not act as the p r imary R:FR sensor in h igher plants.
Experimental procedures
Growth conditions
Seeds were germinated using gibberellic acid (GA3, Sigma, Poole, Dorset, UK) to break dormancy. Seeds were incubated in 500 tlg m1-1 GA 3 for 1 h before washing with three changes of deionised water. They were then surface-dried on filter paper before being sown onto half-strength MS medium (Sigma) with the omission of sucrose, solidified with 0.6% agarose. Seeds were allowed to imbibe water in the dark for 2 days before germination was synchronized by a 30 min pulse of fluorescent W. They were returned to darkness for 5 days to germinate before placement in monochromatic light treatments. The R and FR sources used have been described before by Robson and Smith (1996) and the B source was cool white fluorescent light, filtered through blue perspex and 1 M CuSO 4. Fluence rate measurements were made using an Analytical Devices personal spectrometer model PS2 00A (Analytical Spectral Devices, Boulder, CO). Where not stated in the figure, fluence rates were 8 l imol m -2 sec -1 for R and FR, 3.5 i~mol m -2 sec -1 for B, and 20 ~mol m -2 sec -1 for W. Monochromatic light treatments lasted for 7 days. The cabinets capable of delivering W together with FR have been described in detail by Keiller and Smith (1989). Seeds were germinated as before, and seedlings grown under identical conditions in W for 6 weeks to produce strong rosette-stage plants before transfer into W or W + FR. Fluence rates were 92 ~mol m -2 sec -1 PAR in both cabinets, with the W-only cabinet having an R:FR of 4.31 and the W + FR cabinet having an R:FR of 0.17. Plants for DNA, RNA and protein extractions and for sexual crosses were grown from seed treated with GA 3 as before in compost in a heated greenhouse kept at approximately 25°C.
RNA extraction and gel blotting
Leaf tissue from plants grown to the rosette stage (4-6 weeks) in the greenhouse was ground in liquid nitrogen and total RNA was extracted by the fol lowing method. Powdered frozen tissue (1 g) was transferred from a pestle and mortar into a 50 ml disposable polycarbonate centrifuge tube containing 5 ml CHCI3, 5 ml liquefied phenol washed in Tris buffer (Fisher, Loughborough, UK) and 10- ml extraction buffer (10 mM Tris pH 8, 1% w/v TNS (triisopropyl- naphthalenesulphonic acid, Kodak), 4% w/v PAS (p-amino salicylic acid, BDH, Poole, Dorset, UK) and 0.2% v/v 2-mercaptoethanol) using a spatula cooled in liquid nitrogen. The mixture was vor- texed, and the phases separated at 3000 rev min -1 for 10 min in a benchtop centrifuge. The aqueous phase was removed and RNA purified by sodium acetate-ethanol and lithium chloride precipitations. This RNA was used as 'total RNA' after quantifica- tion by A260. Poly(A)+ RNA was isolated using the polyAtract mRNA isolation kit (Promega). Denaturing formaldehyde gels and RNA blotting and hybridization were carried out by standard methods. The N. tabacum ubiquitin gene used as a loading control was provided by Dr Luis Mur of the Leicester Botany Department. The PHYB probe was the PCR product of primer pair 4 (Figure 2a). The CAB I gene was described by Castresana et al. (1987).
Phytochrome extraction and immunoblotting
Protein extracts were prepared from leaf tissue of rosette-stage greenhouse-grown plants, by the method of Somerset al. (1991).
p h y B mutants in N. p l u m b a g i n i f o l i a 1099
Extracted protein was separated by SDS-PAGE, with a 7.5% concentration of acrylamide in the resolving gel. Equal amounts of protein, determined by the Bio-Rad protein assay, were loaded into each lane. Protein was blotted onto Immobilon P PVDF membranes (Millipore) by the semi-dry method. Antibody binding and detection was carried out using the Boehringer Mannheim Chemiluminescence Western Blotting Kit.
Sequencing of the PHYB gene
The following primer pairs had the T3 primer sequence at the 5' end of the forward primer and the T7 primer sequence at the 5' end of the reverse primer. The specific parts of the primer sequences were: pair 1 forward (lf) = AGCTTACTACTCTGGTAGAACTTCCC pair 1 reverse (lr) = AAAGCAGGGTCCTCTGTTCTAGCAGG 2f = GCCTGAGATCCTCACTGTTGGAAC 2r = AAGCCCCACAGCCTCA'I-FGAAC 3f = GGCAAATATGGGGTCTAI-FGCGTC 3r = CCTCGGCATCCTTAAATGAATCTC 4f = TGGTGATGCAGTTTGTGGTATGG 4r = ACGCAAATATGGGTGGGATCAG 5f = GAAGAAGGCTGTTTTTGTGGTGG 5r = TCTTAGACATCTGCCTCTCACAAGC 6f = GCGACAGATTTGACAGAAAACCAG 6r = CCATACTTGCTCTC'I-I'GTGTAACCC Genomic DNA was isolated using the CTAB method (Doyle and Doyle, 1987). Total RNA isolated as described above was converted to first-strand cDNA using the Gibco BRL 3' RACE kit. The primers described above were used to amplify by PCR from 100 ng genomic DNA, or cDNA generated from the equivalent of 25 ng total RNA. Thermal cycling conditions were one cycle of 5 min at 94°C, 1 min at 51°C and 3 min at 72°C, fol lowed by 29 cycles of 1 min at 94°C, 1 min at 63°C and 2 min at 72°C. Products were run on a 1.2% agarose TAE gel.
Fragments from pairs 4 and 6 were generated from cDNA by RT-PCR as above, other fragments from genomic DNA. Four consecutive PCR reactions for each pair were performed and afterwards pooled, and the correct products purified by agarose gel electrophoresis and the Qiagen gel purification kit (Qiagen) with the protocol modified by a 15-min vacuum-drying step after the ethanol column wash. Purified fragments were sequenced directly using dye-labelled primers and terminators from the T3 and T7 primer sites and primers internal to the products on an ABI 377 automated sequencer (ABI, Warrington, Cheshire, UK).
Acknowledgements
We are grateful to Akira Nagatani for supplying the mAT-1 anti- body, to Tim Kunkel and Eberhard Sch~fer for supplying the PMTB antibody, and to Garry Whitelam for supplying the AS-32 antibody and for help and advice. The staff of the Protein and Nucleic Acid Chemistry Laboratory at Leicester University assisted with some of the automated sequencing procedures. We would also like to thank Murray Grant for helpful discussions concerning sequencing strategies. M.E.H. is the recipient of a postgraduate maintenance grant from the Natural Environment Research Council (UK).
References
Bradley, J.M., Murphy, G.P., Whitelam, G.C. and Harberd, N.P. (1996) Identification of phytochrome B amino acid residues

1100 Mat thew Hudson et al.
mutated in three new phyB mutants of Arabidopsis thaliana. J. Exp. Bot. 47, 1449-1455.
Castresana, C., Staneloni, R., Malik, V.S. and Cashmore, A.R. (1987) Molecular characterisation of two clusters of genes encoding the type I CAB polypeptide of PS II in Nicotiana plumbaginifolia. Plant Mol. Biol. 10, 117-126.
Cherry, J.R., Hondred, D., Walker, J.M. and Vierstra, R.D. (1992) Phytochrome requires the 6-kDa N-terminal domain for full biological activity. Proc. Natl Acad. ScL USA, 89, 5039-5043.
Childs, K.L., Cordonnier-Pratt, M.-M., Pratt, L.H. and Morgan, P.W. (1992) Genetic regulation of development in Sorghum bico/or. VII. ma3 R flowering mutant lacks a phytochrome that predominates in green tissue. Plant Physio/. 99, 765-770.
Childs, K.L., Miller, ER., Cordonnier-Pratt, M.M., Pratt, L.H., Morgan, P.W. and Mullet, J.E. (1997) The Sorghum photoperiod sensitivity gene, Ma 3, encodes a phytochrome B. Plant Physio/. 113, 611-619.
Clack, T., Mathews, S. and Sharrock, R.A. (1994) The phytochrome apoprotein family in Arabidopsis is encoded by five genes: the sequences and expression of PHYD and PHYE. Plant Mol, Biol. 25, 413-427.
Devlin, P.F., Rood, S.B., Somers, D.E., Quail, P.H. and Whitelam, G.C. (1992) Photophysiology of the elongated internode (ein) mutant of Brassica rapa. Plant Physiol. 100, 1442-1447.
Devlin, P.F., Somers, D.E., Quail, P.H. and Whitelam, G.C. (1997) The e/ongated internode gene of Brassica rapa encodes phytochrome B. Plant Mo/. Bio/. 34, 537-547.
Doyle, J.H. and Doyle, J.L. (1987) A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bu//. 19, 11-15.
Foster, K.R., Miller, F.R., Childs, K.L. and Morgan, P.W. (1994) Genetic regulation of development in Sorghum bico/or. VIII. Shoot growth, tillering, flowering, gibberellin biosynthesis and phytochrome levels are differentially affected by dosage of the ma3 R allele. P/ant Physio/. 105, 941-948.
Goto, N., Kumagai, T. and Koornneef, M. (1991) Flowering responses to light-breaks in photomorphogenic mutants of Arabidopsis thaliana, a long day plant. Physiol. Plant. 83, 209-215.
Holdsworth, M. (1987) Characterisation of Phytochrome using Monoclonal Antibodies. PhD thesis, University of Leicester.
Jackson, S.D., Heyer, A., Dietze, J. and Prat, S. (1996) Phytochrome B mediates the photoperiodic control of tuber formation in potato. Plant J. 9, 159-166.
Keiller, D. and Smith, H. (1989) Control of carbon partitioning by light quality mediated by phytochrome. Plant Sci. 63, 25-29.
Kerckhoffs, L.H.J., van Tuinen, A., Hauser, B.A., Cordonnier- Pratt, M.-M., Nagatani, A., Koornneef, M, Pratt, L.H. and Kendrick, R.E. (1996) Molecular analysis of tri mutant alleles in tomato indicates the Tri locus is the gene encoding the apoprotein of phytochrome BI. Planta, 199, 152-157.
Kern, R,, Gasch, A., Deak, M., Kay, S.A. and Chua, N.-H. (1993) PhyB of tobacco, a new member of the phytochrome family. Plant PhysioL 102, 1363-1364.
Koornneef, M. and Kendrick, R.E. (1994) Photomorphogenic mutants of higher plants, In Photomorphogenesis in Plants, 2nd Edn (Kendrick, R.E. and Kronenberg, G.H.M., eds). Dordrecht: Kluwer, pp. 601-628.
Koornneef, M., Rolff, E. and Spruit, C.J.P. (1980) Genetic control of light-inhibited hypocotyl elongation in Arabidopsis thaliana. Heynh, Z. PflanzenphysioL 100, 147-160.
Kraepiel, Y., Julien, M., Cordonnier-Pratt, M.-M and Pratt, L. (1994) Identification of two loci involved in phytochrome expression in Nicotiana plumbaginifolia and lethality of the corresponding double mutant. Mol, Gen. Genet. 242, 559-565.
Lopez-Juez, E., Nagatani, A., Tomizawa, K.-I., Deak, M., Kern, R., Kendrick, R.E. and Furuya, M. (1992) The cucumber long hypocotyl mutant lacks a light-stable PHYB-like phytochrome. Plant Cell, 4, 241-251.
McCormac, A.C., Wagner, D., Boylan, M.T., Quail, P.H., Smith, H. and Whitelam, G.C. (1993) Photoresponses of transgenic Arabidopsis seedlings expressing introduced phytochrome B- encoding cDNAs: evidence that phytochrome A and phytochrome B have distinct photoregulatory functions. Plant J. 4, 19-27.
Mancinelli, A.L. (1994) The physiology of phytochrome action. In Photomorphogenesis in Plants, 2nd Edn (Kendrick, R.E. and Kronenberg, G.H.M., eds). Dordrecht: Kluwer, pp. 211-266.
Pratt, L.H., Cordonnier-Pratt, M.-M., Hauser, B. and Caboche, M. (1995) Tomato contains two differentially expressed genes encoding B-type phytochromes, neither of which can be considered an orthologue of Arabidopsis phytochrome B. Planta, 197, 203-206.
Quail, P.H. (1994) Phytochrome genes and their expression. In Photomorphogenesis in Plants, 2nd Edn (Kendrick, R.E. and Kronenberg, G.H.M, eds.) Dordrecht: Kluwer, pp. 71-104.
Quail, P.H., Boylan, M.T., Parks, B.M., Short, T,W., Xu, Y. and Wagner, D. (1995) Phytochromes: photosensory perception and signal transduction. Science, 268, 675-680.
Reed, J.W., Nagpal, P., Poole, D.S., Furuya, M. and Chory, J. (1993) Mutations in the gene for the red/far red light receptor phytochrome B alter cell elongation and physiological responses throughout Arabidopsis development. Plant Cell, 5, 147-157.
Reed, J.W., Nagatani, A., Elich, T.D., Fagan, M, and Chory, J. (1994) Phytochrome A and phytochrome B have overlapping but distinct functions in Arabidopsis development. Plant Physiol 104, 1139-1149.
Robson, P.R.H. and Smith, H. (1996) Genetic and transgenic evidence that phytochromes A and B act to modulate the gravitropic orientation of Arabidopsis thaliana hypocotyls. Plant Physiol. 110, 211-216.
Robson, P.R.H., Whitelam, G.C. and Smith, H. (1993) Selected components of the shade-avoidance syndrome are displayed in a normal manner in mutants of Arabidopsis thaliana and Brassica rapa deficient in phytochrome B. Plant Physiol. 102, 1179-1184.
Smith, H. (1994) Sensing the light environment: the functions of the phytochrome family. In Photomorphogenesis in Plants, 2nd Edn. (Kendrick, R.E. and Kronenberg, G.H.M, eds). Dordrecht: Kluwer, pp. 377-414.
Smith, H. (1995) Physiological and ecological function within the phytochrome family. Annu. Rev. Plant Physiol. Plant MoL Biol. 46, 289-315.
Smith, H., Turnbull, M. and Kendrick, R.E. (1992) Light-grown plants of the cucumber long hypocotyl mutant exhibit both long-term and rapid elongation responses to irradiation with supplementary far-red light. Photochem. PhotobioL 56, 607-610.
Somers, D.E., Sharrock, R.A., Tepperman, J.M. and Quail, P.H. (1991) The hy3 long hypocotyl mutant of Arabidopsis is deficient in phytochrome B. Plant Cell, 3, 1263-1274.
van Tuinen, A., Kerckhoffs, L.H.J., Nagatani, A., Kendrick, R.E. and Koornneef, M. (1995) A temporarily red light insensitive mutant of tomato lacks a light stable, B-like phytochrome. Plant Physiol. 108, 939-947.
Wagner, D. and Quail, P.H. (1995) Mutational analysis of phytochrome-B identifies a small COOH-terminal domain critical for regulatory activity. Proc. Natl Acad. ScL USA, 92, 8596-8600.

phyB mutants in N. p lumbag in i fo l ia 1101
Wagner, D., Teppermen, J.M. and Quail, RH. (1991) Overexpression of phytochrome B induces a short hypocotyl phenotype in transgenic Arabidopsis. Plant Cell, 3, 1275-1288.
Wagner, D., Fairchild, C.D., Kuhn, R.M. and Quail, RH. (1996) Chromophore bearing NH 2 terminal domains of phytochrome- A and phytochrome-B determine their photosensory specificity and differential light lability. Proc. Natl Acad. ScL USA, 93, 4011-4015.
Weller, J.L., Nagateni, A., Kendrick, R.E., Muffet, I.A. and Reid, J.B. (1995) New Iv mutants of pea are deficient in phytochrome B. Plant Physiol. 108, 525-532.
Wester, L., Somers, D.E., Clack, T. and Sharrock, R.A. (1994) Transgenic complementation of the hy3 phytochrome B
mutation and response to PHYB gene copy number in Arabidopsis. Plant J. 5, 261-272.
Whitelam, G.C. and Harberd, N.P. (1994) Action and function of phytochrome family members revealed through the study of mutant and transgenic plants. Plant Cell Environ. 17, 615-625.
Whitelam, G.C. and Smith, H. (1991) Retention of phytochrome- mediated shade avoidance responses in phytochrome deficient mutants of Arabidopsis, cucumber and tomato. J. Plant PhysioL 139, 119-125.
Xu, Y., Parks, B.M., Short, T.W. and Quail, P.H. (1995) Missense mutations define a restricted segment in the C-terminal domain of phytochrome A critical to its regulatory activity. Plant Cell, 7, 1433-1443.
EMBL accession number for N. plumbaginifolia phyB gene: Y14676


















