
Neurotransmitters and neuromodulators during early human development
17
Review article Neurotransmitters and neuromodulators during early human development Eric Herlenius, Hugo Lagercrantz* Neonatal Unit, Department of Women and Child Health, Astrid Lindgren Children’s Hospital, Karolinska Institutet, S-171 76 Stockholm, Sweden Received 22 March 2001; received in revised form 4 June 2001; accepted 5 June 2001 Abstract Background: Neurotransmitters such as monoamines appear in the embryo before the neurones are differentiated. They may have other functions than neurotransmission during embryogenesis such as differentiation and neuronal growth. For example, serotonin may act as a morphogen. A number of neuropeptides are expressed during ontogenesis, but their function has been difficult to establish. Maybe some of them remain as evolutionary residues. Fast-switching neurotransmitters like the excitatory amino acids and the more ionotropic receptors dominate in the human brain, but appear probably later during evolution as well as during ontogeny. Methods: The distribution of catecholamines during development has been analysed with a fluorescense method, while most of the other neuortransmitters have been mapped with immunohistochemical methods. The classical method to determine the physiological role of a neurotransmitter or modulator is to study the physiological effect of its antagonist, blocking the endogenous activity. By transgenic technique, the genes encoding for enzymes involved in the synthesis of neurotransmitters can be knocked-out. Major findings: Pharmacological blocking of endogenous activity has, for example, demonstrated that adenosine suppresses fetal respiration. Knocking out the dopamine beta-hydroxylase gene results in fetal death, suggesting that noradrenaline is essential for survival. Some neuro- transmitters change their effect during embryogenesis, e.g. GABA which is excitatory in the embryo, but inhibitory after birth due to a switch from a high to low chloride content in the nerve cells. It is possible that this is of importance for the wiring of neuronal network in early life. NMDA receptors dominate in the foetus, while kainate and AMPA receptors appear later. At birth, there is a surge of neurotransmitters such as catecholamines, which may be of importance for the neonatal adaptation. Conclusions: Neurotransmitters and modulators are not only important for the neural trafficking in the embryo, but also for the development of the neuronal circuits. Prenatal or 0378-3782/01/$ – see front matter D 2001 Elsevier Science Ireland Ltd. All rights reserved. PII:S0378-3782(01)00189-X * Corresponding author. Tel.: +46-8-5177-4700; fax: +46-8-5177-5121. E-mail address: [email protected] (H. Lagercrantz). www.elsevier.com/locate/earlhumdev Early Human Development 65 (2001) 21 – 37
-
Upload
eric-herlenius -
Category
Documents
-
view
212 -
download
0
Transcript of Neurotransmitters and neuromodulators during early human development

Review article
Neurotransmitters and neuromodulators during early
human development
Eric Herlenius, Hugo Lagercrantz*
Neonatal Unit, Department of Women and Child Health, Astrid Lindgren Children’s Hospital,
Karolinska Institutet, S-171 76 Stockholm, Sweden
Received 22 March 2001; received in revised form 4 June 2001; accepted 5 June 2001
Abstract
Background: Neurotransmitters such as monoamines appear in the embryo before the neurones
are differentiated. They may have other functions than neurotransmission during embryogenesis
such as differentiation and neuronal growth. For example, serotonin may act as a morphogen. A
number of neuropeptides are expressed during ontogenesis, but their function has been difficult to
establish. Maybe some of them remain as evolutionary residues. Fast-switching neurotransmitters
like the excitatory amino acids and the more ionotropic receptors dominate in the human brain, but
appear probably later during evolution as well as during ontogeny. Methods: The distribution of
catecholamines during development has been analysed with a fluorescense method, while most of
the other neuortransmitters have been mapped with immunohistochemical methods. The classical
method to determine the physiological role of a neurotransmitter or modulator is to study the
physiological effect of its antagonist, blocking the endogenous activity. By transgenic technique,
the genes encoding for enzymes involved in the synthesis of neurotransmitters can be knocked-out.
Major findings: Pharmacological blocking of endogenous activity has, for example, demonstrated
that adenosine suppresses fetal respiration. Knocking out the dopamine beta-hydroxylase gene
results in fetal death, suggesting that noradrenaline is essential for survival. Some neuro-
transmitters change their effect during embryogenesis, e.g. GABA which is excitatory in the
embryo, but inhibitory after birth due to a switch from a high to low chloride content in the nerve
cells. It is possible that this is of importance for the wiring of neuronal network in early life.
NMDA receptors dominate in the foetus, while kainate and AMPA receptors appear later. At birth,
there is a surge of neurotransmitters such as catecholamines, which may be of importance for the
neonatal adaptation. Conclusions: Neurotransmitters and modulators are not only important for the
neural trafficking in the embryo, but also for the development of the neuronal circuits. Prenatal or
0378-3782/01/$ – see front matter D 2001 Elsevier Science Ireland Ltd. All rights reserved.
PII: S0378 -3782 (01 )00189 -X
* Corresponding author. Tel.: +46-8-5177-4700; fax: +46-8-5177-5121.
E-mail address: [email protected] (H. Lagercrantz).
www.elsevier.com/locate/earlhumdev
Early Human Development 65 (2001) 21–37

neonatal stress (hypoxia), as well as various drugs, may disturb the wiring and cause long-term
behavioural effects (fetal and neonatal programming). D 2001 Elsevier Science Ireland Ltd. All
rights reserved.
Keywords: Neurotransmitters; Neuromodulators; Early human development
1. Introduction
Although genes mainly determine the development of the scaffold of the CNS, the
detailed wiring of the neuronal circuits is to a large degree self-generated dependent on
the action of neurotransmitters and neuromodulators. They can promote, amplify,
block, inhibit or attenuate the micro-electric signals which are passed on to neurones.
Thereby, they give rise to the signalling patterns between myriads of neuronal
networks providing the physical networks of cerebral neurones. Neurotransmitters such
as the catecholamines appear in the embryos of vertebrate and invertebrate animals
even before neurones are differentiated [1]. Some of the cells in the neuronal crest
contain noradrenaline from the outset, but become cholinergic due to environmental
influences [2].
Many neuroactive molecules change their functional role in the CNS during develop-
ment. The same molecule may be crucial for differentiation, neuronal growth and
establishment of neuronal networks in the immature CNS while switching to a more
modulatory role of the ongoing traffic in the mature CNS. Receptor subunits may
exchange during development, i.e. the NMDA receptors, whose subunits allow longer
open channel time during early development then switch to a shorter, more stable adult
subunit composition. This is of importance for the plasticity of the immature brain and
subsequently, for memory storage and preservation in the adult brain [3].
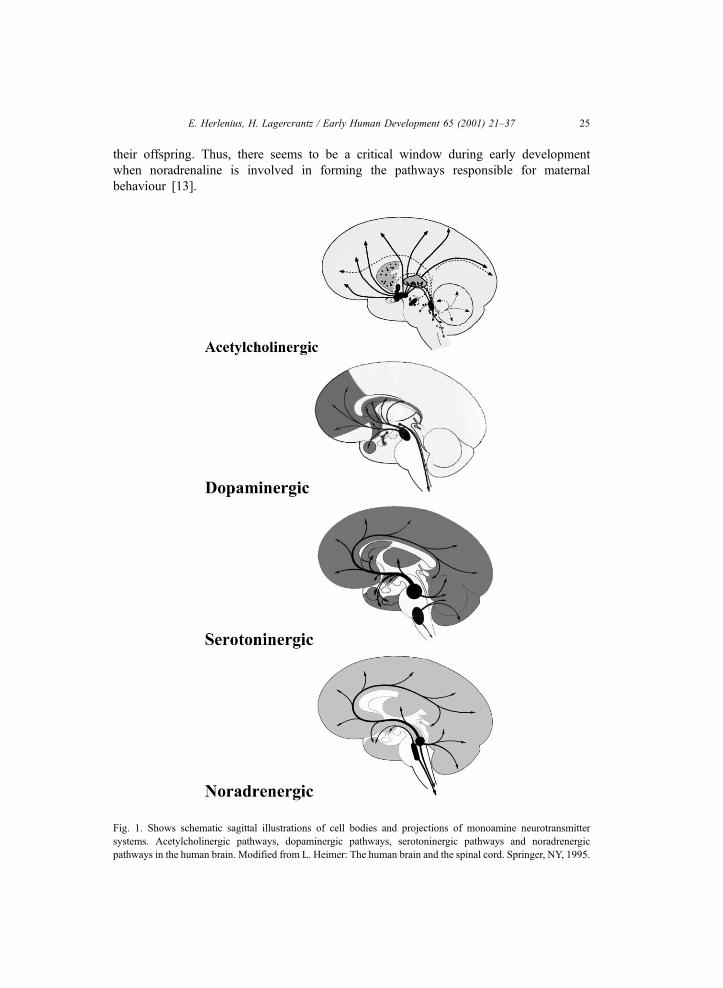
Noradrenaline and acetylcholine are regarded as classical neurotransmitters and
dominate in the peripheral nervous system (Fig. 1). They appear at an early stage
during both phylogenesis and ontogenesis. Many of the neuropeptides were first
identified in the gastrointestinal tract and probably appear early during CNS develop-
ment. They act slowly since they have to be synthesised and packaged in the cell soma
and carried to the terminals before they can be released. The recently evolved and more
sophisticated mammalian brain requires more fast-switching neurotransmitters acting
directly on ion-channels. Therefore, excitatory and inhibitory amino acids seem to
dominate in the mature CNS, where the monoamines and neuropeptides may act more as
neuromodulators (see Ref. [4]).
The distinction between a transmitter and a modulator is far from clear, since several of
the neuroactive agents described to date change their role during brain development or
have different actions depending on brain region or innervated neurones. Furthermore, a
given transmitter may have different effects depending on brain region, postsynaptic
receptor configuration, G-protein coupling and second messenger system.
A neuroactive agent can be expressed in high amounts during certain stages of
development, but then persists in only a few synapses [5]. It is possible that this agent
E. Herlenius, H. Lagercrantz / Early Human Development 65 (2001) 21–3722

either has only a transitory role in a critical window in development or that it remains
mainly as an evolutionary residue, with minor functions in, e.g. mammals. If the
synthesis of some of these neurotransmitters/modulators is blocked pharmacologically or
knocked-out by transgenic techniques, it does not seem to affect survival or even
important physiological functions. This illustrates the plasticity of the brain during early
development. Other neuroactive agents seem to be able to take over. Markers for
neurotransmitters and neuromodulators during CNS development generally appear first
in the caudal and phylogenetically older part of the brain probably due to earlier
neurogenesis (see Ref. [6]).
Classification of the main neurotransmitters and modulators according to princi-
pal biochemical differences and tentative ontogenetic appearance is depicted in
Table 1.
2. Receptors
The neurotransmitters or modulators can act on either metabotropic or ionotropic
receptors (see reviews by, e.g. Ref. [7]). The action of the metabotropic receptors is based
on their effects on G- or N-proteins in the lipid bilayer of the membrane to affect their
enzymes and channels. This effect is slower (tens of milliseconds) than for the ionotropic
receptors. Metabotropic receptors are probably expressed at an earlier stage during
ontogeny and play a more modulatory role in the mature CNS.
The ionotropic receptors respond rapidly and are also termed class I receptors. They act
on ion gates, which they can open or close in less than a millisecond. The ion channels
consist of transmembrane proteins, which can be selective for cations (activatory
receptors) or anions (inhibitory). The ionotropic nicotinic acetylcholine receptor (nAcR),
the GABAA-R and glycine receptor GlyR are members of the same evolutionary super
Table 1
Major neurotransmitters and neuromodulators presented in presumed order of appearance during ontogeny
Purines
Adenosine, ATP
Monoamines
Serotonin (indoleamines)
Dopamine, noradrenaline, adrenaline
Neuropeptides
Opioids: enkephalins, endorphin, dynorphin
Tachykinins: substance P, neurokinin
Glucagon-related: vasoactive intestinal polypeptide (VIP), pituitary adenylate cyclase activating
peptide (PACAP)
Neuropeptide Y (NPY)
Somatostatin
Neurotensin, calcitonin gene-related peptide (CGRP)
Acetylcholine
Amino acids
Glycine, GABA, glutamate, aspartate
E. Herlenius, H. Lagercrantz / Early Human Development 65 (2001) 21–37 23

family and have a similar structure. A fetal subunit of the acetylcholine receptor (gamma-
AchR) is replaced by an adult type (epsilon-AchR) in the muscle end-plate to increase the
conductance [8].
3. Ontogeny of neurotransmitter systems
The choice of neurotransmitter of a precursor neuron depends on the environment. In a
series of remarkable experiments, Le Douarin [9] demonstrated that when the sympathetic
trunk crest from a quail was transplanted into the vagal region of a chick host, the nerves
became cholinergic. Conversely, when vagal neurones were transplanted into the sym-
pathetic trunk, the nerves became adrenergic. The expression of neurotransmitter type
seemed to be dependent on a tissue factor. When sympathetic ganglia cells were cultivated
in a medium from a heart cell culture, the adrenergic neurones became cholinergic [2]. The
choice of transmitter could also be affected by corticosteroids. Thus, environmental factors
are important for the differentiation and which neuroactive agent will be used in
communication with the surrounding cells.
4. Catecholamines
Catecholamines can be found in protozoa as well as in the very early embryo.
The synthesising enzyme tyrosine hydroxylase has been detected the first day of/after
incubation of the chicken; dopamine the second day, and noradrenaline and adrena-
line the third day. High concentrations of catecholamines have been found in
Hensen’s node, corresponding to the notochord of the mammalian embryo (see
Ref. [1]).
Noradrenaline is essential for normal brain development. The noradrenergic system
regulates the development of the Cajal-Retzius cells, which are the first neurones to be
born in the cortex and proposed to be instrumental in neuronal migration and laminar
formation [10]. Administration of 6-OH-dopamine prevents the natural-programmed cell
death of these neurones and delays the formation of cortical layers. Lesioning the
noradrenergic projections or blocking neurotransmission with receptor antagonist prevents
astrogliosis and glial cell proliferation. Depleting noradrenaline during the perinatal period
results in subtle dendritic changes and possibly also alterations in cortical differentiation
(see Ref. [11]).
The role of noradrenaline has been investigated by targeted disruption of the
dopamine b-hydroxylase (DBH) gene. The DBH locus of the DBH proximal promoter
and the first exon were replaced with a neomycin-resistance cassette [12]. This resulted
in fetal death, probably due to cardiovascular failure. Only about 5% of the
homozygotic mice survived until adulthood, presumably due to some placental transfer
of noradrenaline. Most of the mice could be rescued to birth by providing them with
dihydroxyphenylserine (DDPS), a precursor that can be converted to noradrenaline in
the absence of DBH. These mice had a reduced ability of acquisition and retention for
some tasks. Interestingly, female mice seemed to have deficient ability to take care of
E. Herlenius, H. Lagercrantz / Early Human Development 65 (2001) 21–3724
their offspring. Thus, there seems to be a critical window during early development
when noradrenaline is involved in forming the pathways responsible for maternal
behaviour [13].
Fig. 1. Shows schematic sagittal illustrations of cell bodies and projections of monoamine neurotransmitter
systems. Acetylcholinergic pathways, dopaminergic pathways, serotoninergic pathways and noradrenergic
pathways in the human brain. Modified from L. Heimer: The human brain and the spinal cord. Springer, NY, 1995.
E. Herlenius, H. Lagercrantz / Early Human Development 65 (2001) 21–37 25

Dopamine plays a very important role in motor and cognitive programmes. Dopami-
nergic neurones appear early during development 6–8 weeks in human [14] (Fig. 2),
earlier in females than in males. The dopamine turnover is relatively high during the
perinatal period compared to adults. Extremely high levels of D1 receptors have been
reported in the pallidum during the perinatal period [15]. D1 receptor stimulation regulates
transcription of other genes, and it is possible that abnormal perinatal stimulation can
result in long-term consequences (see below).
Disturbances of the development of the dopaminergic system may lead to
dyskinesia, dystonia, tics, obsessive-compulsive disorders and abnormal eye move-
ments. This has been observed in DA-depleted rats after 6-hydroxydopamine treat-
ments but with preserved noradrenaline effect. Tyrosine–hydroxylase gene-deleted
mice were hypoactive and suffered from adipsia and aphagia, which could be treated
with L-dopa [16].
5. Serotonin
Serotonin can already be detected in the fertilised egg and is involved in early
morphogenesis of the heart, the craniofacial epithelia and other structures. If embryos
are cultured in the presence of serotonin uptake inhibitors or receptor ligands,
specific craniofacial malformations occur. Serotoninergic cells in the raphe are among
the earliest to be generated in the brain (about E11 to E15 in the rat and between
weeks 5 and 12 in the human foetus) (Fig. 2). These cells send axons to the
forebrain and may be of importance in the differentiation of neuronal progenitors
[17]. Excess of serotonin prevents the normal development of the somatosensory
cortex, which has been demonstrated in monoamine oxidase knockout mice [18]. At
birth, serotonergic neurones penetrate all cortical layers, but then decline markedly
after about 3 weeks. Depletion of serotonin after birth seems to have little effect on
cortical development.
There are some indirect evidences that reduced serotonergic innervation of the cortex
results in impaired behaviour in juvenile rats. Autism has been suggested to be related
to hyperserotonism.
Fig. 2. Arbitrary levels of monoamines and acetylcholine in man versus age (10-logarithmic scale). Data from
Refs. [14,58,59].
E. Herlenius, H. Lagercrantz / Early Human Development 65 (2001) 21–3726

6. Drugs affecting monoaminergic activity
Cocaine is probably the most well-known drug interacting with the catecholaminergic
systems in the brain during development [19]. It inhibits the presynaptic transport
mechanisms, removing and terminating the action of dopamine and noradrenaline. While
cocaine potentiates the catecholamine effects in the adult, it inhibits the activity during the
immediate postnatal period in most brain regions. Prenatal cocaine exposure results in
disturbance of neuronal migration and consequently leads to severe neurobehavioural
disturbances. Prenatal cocaine exposure in humans causes abnormal motor behaviour
immediately after birth and abnormal behaviour is apparent at 2 and 3 years follow-up,
probably mainly due to disturbance of the dopaminergic system. Neuroleptic drugs
administrated during pregnancy can block dopamine receptors and cause long-lasting
effects (see Ref. [15]).
7. Acetylcholine
Acetylcholine is one of the major neurotransmitters in the brain of importance for
cortical activation, memory and learning. It has a major role in the control motor tone and
movement and probably counterbalances the effect of dopamine (see Ref. [20]). The
cholinergic innervation of the cortex occurs later than the monoaminergic around week 20
in the human foetus. Mature levels in rodents are not reached until after 8 weeks postnatally
(see Ref. [11]). The concentrations of ACh reach about 20% of the adult levels at E15 in the
whole brain of the rat and about 40% at day P7 (Fig. 2). The levels of choline
acetyltrasferase (ChAT) are much lower (1% and 8%) at the corresponding ages, indicating
low firing rates of the cholinergic neurones. Conversely, the receptors reach adult levels
earlier. The cholinergic markers appear sooner in the pons-medulla, probably due to earlier
neurogenesis in the caudal and phylogenetically older part of the brain (see Ref. [6]).
The cholinergic afferents seem to have an important role in the differentiation of the
cortex. After a considerable reduction of the cholinergic innervation, a delay of cortical
cytodifferentiation was revealed (see Ref. [11]). The cholinergic innervation has been
found to be disturbed in Down’s syndrome, lead and ethanol toxicity and asphyxia.
Perinatal manipulations of the cholinergic system result in major changes of
cortical structure. These changes can be correlated to cognitive deficits but do not affect
motor behaviour.
8. Amino acid transmitters
The amino acids are involved in the wiring of neuronal networks and building CNS
cytoarchitecture [21] (Fig. 3). Amino acid transmitters are the most abundant transmitters
in the central nervous system. However, they were recognised as neurotransmitters in the
mammalian brain much later than the monoamines and acetylcholine. This was probably
due to the fact that they are involved in intermediate metabolism and constitute important
building blocks in the proteins.
E. Herlenius, H. Lagercrantz / Early Human Development 65 (2001) 21–37 27
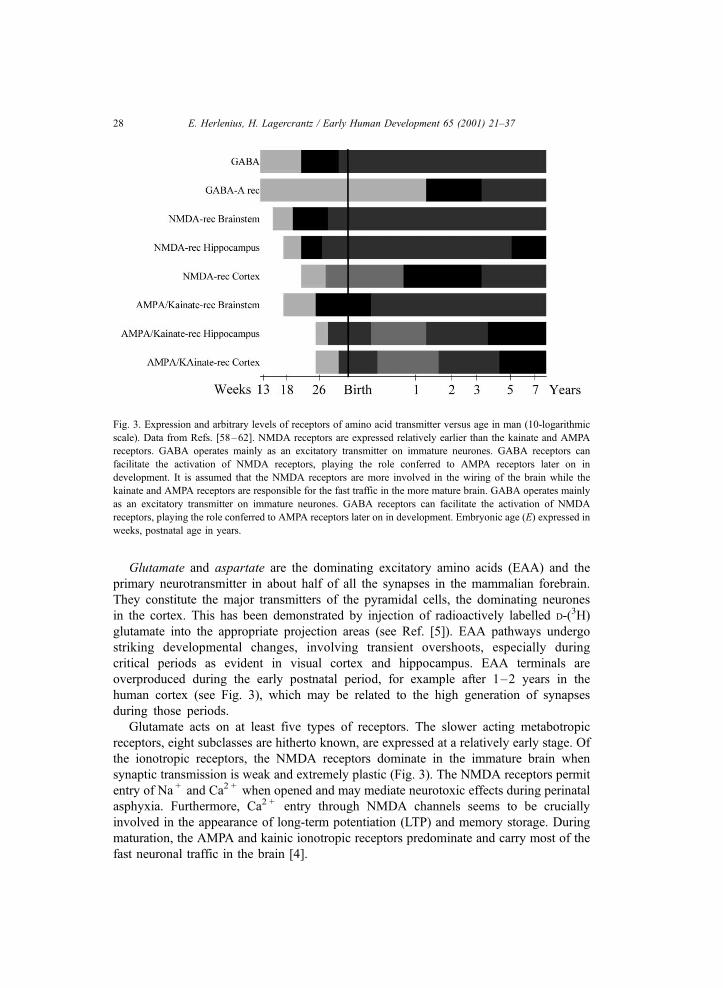
Glutamate and aspartate are the dominating excitatory amino acids (EAA) and the
primary neurotransmitter in about half of all the synapses in the mammalian forebrain.
They constitute the major transmitters of the pyramidal cells, the dominating neurones
in the cortex. This has been demonstrated by injection of radioactively labelled D-(3H)
glutamate into the appropriate projection areas (see Ref. [5]). EAA pathways undergo
striking developmental changes, involving transient overshoots, especially during
critical periods as evident in visual cortex and hippocampus. EAA terminals are
overproduced during the early postnatal period, for example after 1–2 years in the
human cortex (see Fig. 3), which may be related to the high generation of synapses
during those periods.
Glutamate acts on at least five types of receptors. The slower acting metabotropic
receptors, eight subclasses are hitherto known, are expressed at a relatively early stage. Of
the ionotropic receptors, the NMDA receptors dominate in the immature brain when
synaptic transmission is weak and extremely plastic (Fig. 3). The NMDA receptors permit
entry of Na + and Ca2 + when opened and may mediate neurotoxic effects during perinatal
asphyxia. Furthermore, Ca2 + entry through NMDA channels seems to be crucially
involved in the appearance of long-term potentiation (LTP) and memory storage. During
maturation, the AMPA and kainic ionotropic receptors predominate and carry most of the
fast neuronal traffic in the brain [4].
Fig. 3. Expression and arbitrary levels of receptors of amino acid transmitter versus age in man (10-logarithmic
scale). Data from Refs. [58–62]. NMDA receptors are expressed relatively earlier than the kainate and AMPA
receptors. GABA operates mainly as an excitatory transmitter on immature neurones. GABA receptors can
facilitate the activation of NMDA receptors, playing the role conferred to AMPA receptors later on in
development. It is assumed that the NMDA receptors are more involved in the wiring of the brain while the
kainate and AMPA receptors are responsible for the fast traffic in the more mature brain. GABA operates mainly
as an excitatory transmitter on immature neurones. GABA receptors can facilitate the activation of NMDA
receptors, playing the role conferred to AMPA receptors later on in development. Embryonic age (E) expressed in
weeks, postnatal age in years.
E. Herlenius, H. Lagercrantz / Early Human Development 65 (2001) 21–3728

Dark rearing or blocking the activity with tetrodotoxin results in preservation of the
NMDA receptors in the visual cortex. Dark rearing also preserves the immature form of
the NMDA receptors containing NR2B subunit and the expression of NR2A is delayed.
This subunit switch is essential for development of rapid synaptic transmission [3].
Fetal rats exposed to NMDA antagonists were found to have excessive apoptosis in the
same way as the asphyxiated perinatal brain. NMDA receptor stimulation by excessive
glutamate release leads to Ca+ + influx, which may induce subsequent neuronal apoptosis.
‘‘Thus, either too much or too little NMDA receptor activity can be life-threatening to the
developing neurones’’ [22].
Gamma-aminobutyric acid (GABA) is the dominating neurotransmitter in the non-
pyramidal cells, as demonstrated by uptake of (3H)-GABA and immunochemical labelling
of the GABA-synthesising enzyme glutamic acid (GAD). Perhaps 25–40% of all nerve
terminals contain GABA. GABA is regarded as the main inhibitory transmitter in the
mature animal, but has a different role during early development.
There are two types of GABA receptors: GABAA and GABAB. The GABAA-receptor
(GABAA-R) is an ionotropic receptor that gates a chloride channel. It is a transmembrane
protein built of several subunits where, for example benzodiazepines, barbiturates and
ethanol, can bind to specific sites and modulate the opening properties of the chloride
channel. The GABAB-R is coupled to a G-protein, is present in lower levels in the CNS
than the GABAA-receptor and starts to function late in CNS development (postnatal life
in rodents).
During early development, the Cl � concentration is high in the nerve cells. When
GABA opens the Cl � channels, a depolarisation (i.e. excitation) occurs. During matura-
tion, the Cl � concentration decreases which results in an opposite effect of GABA, i.e.
Cl � ions are pumped out and the cell becomes hyperpolarised. In this way, GABA
switches from an excitatory to an inhibitory neurotransmitter [23]. This switch occurs
around birth in the rat [23] but it is not known when it occurs in the human.
Thus, GABA operates mainly as an excitatory transmitter on immature neurones. As
described above, glutaminergic synapses initially lack functional AMPA receptors and
the NMDA channels are blocked by Mg2 + at resting membrane potentials. GABA
depolarises immature neurones, which may result in Ca2 + influx by removing the Mg2 +
blockage of NMDA channels. Thus, GABAA-receptors play the role conferred to AMPA
receptors in the more matured CNS [21,24]. An increase in the intracellular Ca2 +
concentration activates a wide range of intracellular cascades and is involved in neuronal
growth and differentiation. Furthermore, GABA excitation and Ca2 + influx may act as
triggers for plasticity of synaptic connections and for establishing and patterning of
neural networks.
The GABAA-receptors have a strong affinity for benzodiazepines. Several anxiolytic
and anticonvulsant drugs increase the ability for GABA to open chloride channels. In
neonatal neurones, GABA currents are potentiated by barbiturates but are insensitive to
benzodiazepines [25].
As GABA has a trophic role during early brain development, interference with the
function of GABAergic transmission during this period may affect the development of
neuronal wiring, plasticity of neuronal network, and also have a profound influence on
neural organisation.
E. Herlenius, H. Lagercrantz / Early Human Development 65 (2001) 21–37 29

In fact, ethanol, abused by some mothers during pregnancy, interacts with the
GABAA-receptor. The sensitive time window in rat cerebral cortex for ethanol
exposure is situated between P3 and P10. It is worth noting that GABA, during
this same period, seem to have mainly depolarising and trophic effects on developing
cortical neurones through effects on cell proliferation and migration [26]. In humans,
the intellectual deficits produced by abnormalities of brain growth are the most
important components of fetal alcohol syndrome [27]. Craniofacial abnormalities in
human foetuses related to first trimester alcohol exposure are similar to the facial
defects seen in GABAA subunit receptor knockout mice [28]. Thus, ethanol inter-
action with the GABAA-receptor could be implicated in the pathogenesis of fetal
alcohol syndrome.
Glycine has both excitatory and inhibitory actions and can be regarded as the
phylogenetically older inhibitory transmitter restricted to the brain stem and spinal cord
in the adult. A similar switch as regarding the GABAA-receptors from excitatory to
inhibitory effects seems to occur with maturation [23].
The NMDA receptor has a modulatory site where glycine in submicromolar concen-
trations increases the frequency of NMDA-receptor channel opening. Conditions that
alter the extracellular concentration of glycine can dramatically alter NMDA-receptor-
mediated responses.
9. Neuropeptides
More than 50 neuropeptides have been identified. In contrast to most of the other
neurotransmitters/modulators, the neuropeptides are synthesised and packaged in large
dense-cored vesicles in the cell soma and are carried to the nerve terminals by axonal
transport at a rate of 1.5 mm/h. It is obvious that by this relatively slow process, the
neuropeptides cannot act as fast-switching neurotransmitters. Rather, they have a
neuromodulatory role. They are often stored together with other neurotransmitters,
i.e. monoamines or EAA, and it is possible that they play a role in setting of the
sensitivity [29]. Some of them are probably of less physiological importance and occur
in the body mainly as evolutionary residues [30]. Still they are of great neuro-
pharmacological interest and their analogues or antagonists can be used as drugs.
The most well-known examples are the opioids and naloxone.
b-Endorphin is expressed early during fetal brain development and seems to be
involved in fetal growth and postnatal development at least in the rat [31]. m-Receptor binding sites are present during mid-fetal time and have a high density in
cardiorespiratory-related brainstem nuclei, whereas the d-opioid receptors primarily
appear during the postnatal period in rats [32]. Although, opioid binding sites
progressively increase in the developing brain (Fig. 4), the effect of opioids appears
to be dependent on the status of neuronal maturation. In addition, many neuronal
populations exhibit transient expression of one or the other opioid genes but the
physiological role of this is not clear. Opioid agonists inhibit mitosis and DNA
synthesis in the developing brain and endogenous opioids exert potent regulatory
effects on brain development and morphogenesis, as demonstrated by the admin-
E. Herlenius, H. Lagercrantz / Early Human Development 65 (2001) 21–3730

istration of exogenous opioid agonist and antagonist during the fetal period [33].
Human neonates who have been exposed in utero to opioids, such as heroine, have a
smaller head circumference and reduced body weight due to a decrease in cell
number [27].
Substance P is a primary sensory transmitter mediating pain sensations via the thin
C-fibres. Substance P is also involved in the transmission of chemoreceptor and
barometric input from the carotid and aortic chemo- and baro-receptors. In humans,
there is an increase towards birth and then a levelling off during the first 6 months [34]
(Fig. 4).
Substance P may play a role in neurogenesis. It seems to counteract damage induced
by neurotoxins and accelerates regeneration of cortical catecholamine fibres [35].
Increased expression of mRNA coding for pre-protachykinin A, the substance P
precursor, has been recorded in respiratory-related nuclei in both the rabbit (see Ref.
[36]) and the rat (see Ref. [37]). Increased expression of the substance P precursor has
also been detected in patches in the nucleus caudatus and putamen of the human
newborn brain [38]. Thus, there are suggestions that substance P is involved in the
resetting and adaption of the organism to extrauterine life.
Vasoactive intestitinal polypeptide (VIP) appears to have important trophic effects on
the development of the cerebral cortex. Radio-labelled VIP has been demonstrated to
pass the placental barrier and stimulate growth of the brain in the mouse. VIP promotes
astrocytogenesis in the germinative zone [39].
Calcitonin gene-related peptide (CGRP) is an example of a neuropeptide with trophic
effects, which might be involved in synaptogenesis. It has been demonstrated to
stimulate the synthesis of ACh receptors in cultured muscle cells from the chick. The
expression of CGRP is most prominent from E11 to E19, when the motor end-plates are
formed [40] (see also Ref. [29]).
Somatostatin is mainly an inhibitory neuromodulator co-existing with GABA.
Somatostatin-related immunoflourescence (SRIF) neurones are transiently expressed to
a high degree between E14 and E21 in the fetal rat. The peak is at E17 [5]. Whether this
high expression is crucial for the formation of the cortex or just reflects an evolutionary
residue is not really known [41].
Fig. 4. Expression and arbitrary levels of some major neuropeptides versus age in man (10-logarithmic scale).
Data from Refs. [34,38,63,64].
E. Herlenius, H. Lagercrantz / Early Human Development 65 (2001) 21–37 31

10. Purines
Purines are fundamental components in the energy turnover of all cells but
also modulate neuronal activity through synaptic or non-synaptic release and
interaction with specific receptors. The purinergic receptors are divided into
type-1 receptors (P1) sensitive to adenosine and AMP, and type-2 (P2) sensitive
for ATP and ADP. The action of purines is related as a rapid breakdown of ATP
increases the levels of adenosine. The purine nucleotide ATP is the main energy
source of cells, but is also stored in synaptic vesicles and released together with
classical transmitters such as noradrenaline and acetylcholine. The ratio between
ATP and catecholamines in chromaffin granules has been found to be higher
during early life than later, suggesting that it is a phylogenetic and ontogenetic
old signalling substance [42].
Adenosine is a constituent of all body fluids, including the extracellular space
of the central nervous system. It has multiple effects on organs and cells of the
body. Thus, its levels are tightly regulated by a series of enzymatic steps [43].
Adenosine can be regarded more as a neuromodulator such that it does not seem
to be stored in vesicles with a regulated release from nerve terminals.
During basal conditions, adenosine levels are 30–300 nM and can rise
following stimuli that cause an imbalance between ATP synthesis and ATP
breakdown. Thus, the levels during ischemia or hypoxia can rise 100-fold
[43,44]. The extracellular concentrations of adenosine might be higher in the fetal
brain than postnatally, since fetal PaO2 can decrease below the level (30 mm Hg)
when a significant increase in extracellular adenosine can be expected [44].
Overall, adenosine decreases oxygen consumption and has neuroprotective effects
[43]. However, hypoxia also induces a decrease in neonatal respiration. Theophyl-
line and caffeine are adenosine antagonists that cause ventilation to increase and
decrease the incidence of neonatal apnoeas when given systemically, mainly due to
the antagonistic effect of theophylline on adenosine A1-receptors in the medulla
oblongata (see Ref. [45]).
11. Transition at birth
The levels of most neurotransmitters and neuromodulators increase concomitantly
with synapse formation. Some of them surge during the perinatal period (such as
glutamate, catecholamines and some neuropeptides) and then level off. The
interesting question is to what extent the expression of neuroactive agents is
related to the functional state of the foetus and the newborn. On one hand, there
is an intense firing and wiring in the fetal brain, particularly during active sleep.
Therefore, an inhibitory neurotransmitter, such as GABA, seems to be mainly
excitatory in the fetal period (see above). Amino acid transmitters also act via
NMDA receptors, which are important for the wiring and plasticity of the
immature brain, while the main excitatory fast-switching receptors (AMPA) are
expressed later.
E. Herlenius, H. Lagercrantz / Early Human Development 65 (2001) 21–3732

Activities such as respiratory movements are suppressed. The foetus seldom or never
becomes aroused or wakes up. The sympathetic tonus is low. Furthermore, the foetus is
adapted to the low oxygen level in the womb. If it is challenged by asphyxia, it is not
excited as an adult responding with a flight or fight reaction, but rather becomes
immobilised, stops breathing and becomes bradycardic (see Ref. [36]). This paralytic
state of the foetus can be caused by inhibition of the chemical neurotransmission or
lack of expression of excitatory neuroactive agents. Adenosine is such a neuro-
modulator, which might be involved in this suppression of the fetal brain. It has a
general sedatory effect. Its concentration increases during energy failure and hypoxia
and it has been suggested that it can act as a modulator to cope with the hypoxic
situation [46]. Adenosine A1-R activation depresses breathing substantially in the foetus
and the neonate by inhibiting synaptic transmission and hyperpolarising certain neuro-
nes [47].
Neuropeptides which might be involved in the suppression of fetal activity are NPY,
somatostatin and endogenous opioids. The levels of NPY are relatively high in the fetal
brain and decline after birth. Plasma levels of endorphins and enkephalins are increased
in the umbilical cord at birth. Blocking endogenous opioids with naloxone increases
breathing in the newborn rabbit.
The healthy newborn baby is aroused and awake the first 2 h after birth and starts
continuous breathing movements. Factors like squeezing and squashing of the foetus,
increased sensory input and cooling are probably of importance. We can hypothesise that
there is a surge of excitatory neurotransmitters and down regulation of inhibitory ones in
the brain.
The increased neuronal activity is indicated by the increased expression of immediate
early genes [48]. The arousal and vigilance of the newborn can probably partially be due
to activation of the noradrenergic system in the brain, particularly locus coeruleus from
where noradrenergic neurones are distributed in the whole brain (see above). The
noradrenaline turnover as indicated by the ratio of the metabolite MHPG and NA was
increased 2–3-fold in the newborn rat (see Ref. [36]). There are indirect indications that
there is also a noradrenaline surge in the human brain, by the finding of high level of
plasma catecholamines after birth.
The most important driving mechanism of respiration, coupling the ventilation to
metabolic demand is the CO2-drive. This sensitivity seems to be mediated by a
cholinergic mechanism [49]. The CO2-drive seems to be strongly upregulated at birth.
An increased expression of immediate early genes (c-fos) has been recorded at the
ventral surface of the newborn rat [37]. Whether this could trigger an upregulation of the
cholinergic neurotransmission is not yet known.
A neuropeptide involved in respiratory control at birth has been found to be
substantially upregulated at birth. mRNA encoding for the substance P precursor pre-
protachykinin A is increased about 4-fold in respiratory nuclei but not in others during
the first day after birth in rabbit pups (see Ref. [36]).
A rapid decrease of the inhibitory neuromodulator adenosine in the brain occurs as
partial pressure of oxygen in arterial blood rapidly increases after birth. In addition, a
decreased sensitivity during the first postnatal days for adenosine seems to contribute to
the maintenance of continuous breathing [45,47].
E. Herlenius, H. Lagercrantz / Early Human Development 65 (2001) 21–37 33

12. Pre and perinatal programming
The concept of fetal and neonatal programming first described by David Barker (see
Ref. [50]) also applies to the ontogeny of neurotransmitters and neuromodulators, i.e. an
early stimulus or insult at a critical period can result in long-term changes in the structure
and the function of the organism. For example, it can be postulated that prenatal or
perinatal stress can disturb the timetable of the expression of neurotransmitters and
neuromodulators and their receptors. Hydrocortisone given to neonatal rats has been
found to enhance the maturation of the monoaminergic systems in the brain [51].
Administration of extra glucocorticosteroids to the rat foetus induces alterations of
dopamine receptor responses, which affects the spontanous motor-control both in short-
and long-term perspectives [52]. Chronic prenatal hypoxia alters the monoamine turnover
in the locus coeruleus and nucleus tractus solitarius in the adolescent rat [53]. This was
related to disturbed control of respiratory behaviour. Human handling of newborn rats for
15 min during the first weeks of life appeared to affect ascending serotonergic projections
into the hipoocampus and long-lasting increase in glucocorticoid receptors (see Ref. [54]).
There are also clinical studies indicating that prenatal stress is associated with attention
deficit disorders in children [55], possibly due to disturbance of monoaminergic turnover
and subsequent wrong programming. Schizophrenic patients seem to have experienced
more pregnancy and birth complications than their healthy siblings [56]. For example,
mothers of schizophrenic patients suffered more often from severe infections during
pregnancy, possibly affecting cytokines and indirectly, the development of monoaminergic
circuits in the fetal brain [57].
13. Conclusions
Monoamines are expressed in the very early embryo, at which stage the notochord
already contains high noradrenaline levels. They may have an important role for
neurotransmission in the foetus. Purines and neuropeptides are probably also expressed
at an early stage, in a similar way as they occur early during phylogenesis. In the adult
mammal, the fast-switching excitatory amino acids dominate. However, they also seem
to be important for the wiring of the brain and the plasticity before birth. NMDA
receptors that might mediate these effects dominate and are then substituted by AMPA
receptors. The main inhibitory amino acids GABA and glycine seem to be excitatory
before birth, which could be of major importance for the wiring of neuronal circuits
during development. Prenatal or neonatal stress, for example hypoxia, can affect the
programming of neurotransmitter and receptor expression, which can lead to long-term
behavioural effects.
Acknowledgements
This article is a shorter version of a chapter entitled ‘Neurotransmitters and and
neuromodulators’ published in: The Newborn Brain. Neuroscience and Clinical
E. Herlenius, H. Lagercrantz / Early Human Development 65 (2001) 21–3734

applications (Eds. H. Lagercrantz, M. Hanson, P. Evrard, C. Rodeck), Cambridge Univ.
Press, 2001.
Supported by the Swedish Medical Research Council (5234), The Axelson-Johnson
Foundation, Marta and Gunnar Philipson Foundation.
References
[1] Pendleton RG, Rasheed A, Roychowdhury R, Hillman RA. New role for catecholamines: ontogenesis.
Trends Pharmacol Sci 1998;19:248–51.
[2] Pattersson PH, Chun ILY. The induction of acetylcholine synthesis in primary cultures of dissociated rat
sympathetic neurons: I. Effects of conditioned medium. Dev Biol 1977;56:263–80.
[3] Fox K, Henley J, Isaac J. Experience-dependent development of NMDA receptor transmission. Nat Neuro-
sci 1999;2(4):297–9.
[4] Cooper JR, Bloom FE, Roth RH. The Biochemical Basis of Neuropharmacology. 7th edn. New York:
Oxford Univ. Press; 1996.
[5] Cavanagh ME, Parnavelas JG. Neurotransmitter differentiation in cortical neurons. In: Parnavelas JG,
Stern CD, Stirling RV, editors. The Making of the Nervous System. London: Oxford Univ. Press; 1988. p.
435–53.
[6] Semba K. Development of central cholinergic neurons. In: Bjorklund A, Hokfelt T, Tohyama M, editors.
Ontogeny of Transmitters and Petides in the CNS. Amsterdam: Elsevier; 1992. p. 33–62.
[7] Bertrand D, Changeux J-P. Nicotinic receptor: an allosteric protein specialized for intercellular communi-
cation. Semin Neurosci 1995;7:75–90.
[8] Herlitze S, Villaroel A, Witzemann VMK, Sakmann B. Structural determinants of channel conductance in
fetal and adult rat muscle acetylcholine receptors. J Physiol 1996;492:775–87.
[9] Le Douarin NM. Plasticity in the development of the peripheral nervous system. Ciba Symp 1981;83:19–46.
[10] Naqui SZH, Harris BS, Thomaidou D, Parnavelas JG. The noradrenergic system influences in fate of Cajal-
Retzius cells in the developing cerebral cortex. Dev Brain Res 1999;113:75–82.
[11] Berger-Sweeney J, Hohmann CF. Behavioral consequences of abnormal cortical development: insights into
developmental disabilities. Behav Brain Res 1997;86(2):121–42.
[12] Thomas SA, Matsumoto AM, Palmiter RD. Noradrenaline is essential for mouse fetal development. Nature
1995;374:643–6.
[13] Thomas SA, Palmiter RD. Impaired maternal behavior in mice lacking norepinephrine and epinephrine.
Cell 1997;91:583–92.
[14] Sundstrom E, Kolare S, Souverbie F, Samuelsson EB, Pschera H, Lunell NO, et al. Neurochemical differ-
entiation of human bulbospinal monoaminergic neurons during the first trimester. Brain Res Dev Brain Res
1993;75(1):1–12.
[15] Boyson SJ, Adams CE. D1 and D2 dopamine receptors in perinatal and adult basal ganglia. Pediatr Res
1997;41(6):822–31.
[16] Zhou Q-YO, Quaife CJ, Palmiter RD. Targeted disruption of the tyrosine hydroxylas gene reveals that
catecholamines are required for mouse fetal development. Nature 1995;374:640–6.
[17] Lauder JM, Moiseiwitsch J, Liu J, Wilkie MB. Serotonin in development and pathophysiology. In: Lou HC,
Greisen G, Falck-Larsen J, editors. Brain Lesions in the Newborn. Alfred Benzon Symposium, vol. 37.
Copenhagen: Munksgaard; 1994. p. 60–72.
[18] Cases O, Lebrand C, Giros B, Vitalis T, De Maeyer E, Caron MG, et al. Plasma membrane transporters of
serotonin, dopamine, and norepinephrine mediate serotonin accumulation in atypical locations in the devel-
oping brain of monoamine oxidase A knock-outs. J Neurosci 1998;18(17):6914–27.
[19] Seidler FJ, Temple SW, McCook EC, Slotkin TA. Cocaine inhibits central noradrenergic and dopaminergic
activity during the critical developmental period in which catecholamines influence cell development. Brain
Res Dev Brain Res 1995;85(1):48–53.
[20] Johnston MV, Silverstein FS. Development of neurotransmitters. In: Polin RA, Fox WW, editors. Fetal and
Neonatal Physiology. Philadelphia: Saunders; 1998. p. 2116–7.
E. Herlenius, H. Lagercrantz / Early Human Development 65 (2001) 21–37 35

[21] Ben-Ari Y, Khazipov R, Leinekugel X, Caillard O, Gaiarsa J-L. GABA-A, NMDA and AMPA receptors: a
developmentally regulated ‘‘menage a trois’’. Trends Neurosci 1997;20:523–9.
[22] Lipton SA, Nakanishi N. Shakespeare in love—with NMDA receptors? Nat Med 1999;5(3):270–1.
[23] Miles R. A homeostatic switch. Nature 1999;397:215–6.
[24] Onimaru H, Herlenius E, Homma I. GABA-dependent responses of respiratory neurons in the fetal rat
medulla. Neurosci Res Suppl 1999;23:77.
[25] Cherubini E, Gaiarsa JL, Ben-Ari Y. GABA: an excitatory transmitter in early postnatal life. Trends Neuro-
sci 1991;14(12):515–9.
[26] Belhage B, Hansen GH, Elster L, Schousboe A. Effects of gamma-aminobutyric acid (GABA) on synapto-
genesis and synaptic function. Perspect Dev Neurobiol 1998;5(2–3):235–46.
[27] Kopecky EA, Koren G. Maternal drug abuse: effects on the fetus and neonate. In: Polin RA, Fox WW,
editors. Fetal and Neonatal Physiology. Philadelphia, PA: Saunders; 1998. p. 203–20.
[28] Condie BG, Bain G, Gottlieb DI, Capecchi MR. Cleft palate in mice with a targeted mutation in the gamma-
aminobutyric acid-producing enzyme glutamic acid decarboxylase 67. Proc Natl Acad Sci U S A 1997;
94(21):11451–5.
[29] Hokfelt T. Neuropeptides in perspective: the last ten years. Neuron 1991;7(6):867–79.
[30] Bowers CW. Superfluous neurotransmitters? Trends Neurosci 1994;17(8):315–20.
[31] Wang Y-Q, Li J-S, Li H-M, Tohyama M. Ontogeny of pro-opiomelanocortin (POMC)-derived peptides in
the brain and pituitary. In: Bjorklund A, Hokfelt T, Tohyama M, editors. Ontogeny of Transmiutters and
Peptides in the CNS. Amsterdam: Elsevier; 1992. p. 297–323.
[32] Befort K, Kieffer BL. Structure-activity relationships in the delta opioid receptor. Pain Rev 1997;4:
100–21.
[33] Lichtensteiger W. Developmental neuropharmacology. In: Polin RA, Fox WW, editors. Fetal and Neonatal
Physiology. Philadelphia: Saunders; 1998. p. 226–39.
[34] Bergstrom L, Lagercrantz H, Terenius L. Post-mortem analyses of neuropeptides in brains from sudden
infant death victims. Brain Res 1984;323(2):279–85.
[35] Sakanaka M. Development of neuronal elements with substance P-like immunoreactivity in the central
nervous system. In: Bjorklund A, Hokfelt T, Tohyama M, editors. Ontogeny of Transmitters and Peptides
in the CNS. Amsterdam: Elsevier; 1992. p. 197–250.
[36] Lagercrantz H. Stress, arousal and gene activation at birth. News Physiol Sci 1996;11:214–8.
[37] Wickstrom HR, Holgert H, Hokfelt T, Lagercrantz H. Birth-related expression of c-fos, c-Jun and substance
P mRNAs in the rat brainstem and pia mater: possible relationship to changes in central chemosensitivity.
Brain Res Dev Brain Res 1999;112(2):255–66.
[38] Brana C, Charron G, Aubert I, Carles D, Martin-Negrier ML, Trouette H, et al. Ontogeny of the striatal
neurons expressing neuropeptide genes in the human fetus and neonate. J Comp Neurol 1995;360(3):
488–505.
[39] Gressens P, Paindaveine B, Hill JM, Brenneman DE, Evrard P. Growth factor properties of VIP during early
brain development. Whole embryo culture and in vivo studies. Ann NY Acad Sci 1997;814:152–60.
[40] Laufer R, Fontaine B, Klarsfeld A, Cartaud J, Changeux JP. Regulation of acetylcholine receptor biosyn-
thesisi during motor endplate morphogenesis. News Physiol Sci 1989;4:5–9.
[41] Shiosaka S. Ontogeny of the central somatostatinergic system. In: Bjorklund A, Hokfelt T, Tohyama M,
editors. Ontogeny of Transmitters and Petides in the CNS. Amsterdam: Elsevier; 1992. p. 369–94.
[42] O’Brien RA, Da Prada M, Pletscher A. The ontogenesis of catecholamines and adenosine-50-triphosphate in
the adrenal medulla. Life Sci [I] 1972;11(16):749–59.
[43] Fredholm BB. Adenosine and neuroprotection. Int Rev Neurobiol 1997;40:259–80.
[44] Winn HR, Rubio R, Berne RM. Brain adenosine concentration during hypoxia in rats. Am J Physiol 1981;
241:235–42.
[45] Herlenius E. Respiratory Activity in Medulla Oblongata and its Modulation by Adenosine and Opioids.
Stockholm: Karolinska Institutet; 1998. http://diss.kib.ki.se/1998/91-628-3240-9.
[46] Berne RM. Adenosine: an important physiological regulator. Trends Neurosci 1986;1:163–7.
[47] Herlenius E, Lagercrantz H. Adenosinergic modulation of respiratory neurones in the neonatal rat brainstem
in vitro. J Physiol (London) 1999;518(1):159–72.
[48] Ringstedt T, Tang LQ, Persson H, Lendahl U, Lagercrantz H. Expression of c-fos, tyrosine hydroxylase, and
E. Herlenius, H. Lagercrantz / Early Human Development 65 (2001) 21–3736

neuropeptide mRNA in the rat brain around birth: effects of hypoxia and hypothermia. Pediatr Res 1995;
37(1):15–20.
[49] Nattie EE, Wood J, Mega A, Goritski W. Rostral ventrolateral medulla muscarinic receptor involvement in
central ventilatory chemosensitivity. J Appl Physiol 1989;66(3):1462–70.
[50] Sayer AA, Cooper C, Barker DJ. Is lifespan determined in utero? Arch Dis Child Fetal Neonat Ed 1997;
77(3):F162–4 [editorial].
[51] Kurosawa A, Kageyama H, John TM, Hirota R, Itoh S. Effect of neonatal hydrocortisone treatment on brain
monoamines in developing rats. Jpn J Pharmacol 1980;30(2):213–20.
[52] Diaz R, Fuxe K, Ogren SO. Prenatal corticosterone treatment induces long-term changes in spontaneous and
apomorphine-mediated motor activity in male and female rats. Neuroscience 1997;81(1):129–40.
[53] Peyronnet J, Roux JC, Geloen A, Tang LQ, Pequignot JM, Lagercrantz H, et al. Prenatal hypoxia impairs
the postnatal development of neural and functional chemoafferent pathway in rat. J Physiol (London) 2000;
524(Pt 2):525–37.
[54] Sapolsky RM. The importance of a well-groomed child. Science 1997;277(5332):1620–1 [comment].
[55] Weinstock M. Does prenatal stress impair coping and regulation of hypothalamic–pituitary–adrenal axis?
Neurosci Biobehav Rev 1997;21(1):1–10.
[56] Stefan MD, Murray RM. Schizophrenia: developmental disturbance of brain and mind? Acta Paediatr
Scand, Suppl 1997;422:112–6.
[57] Jarskog LF, Xiao H, Wilkie MB, Lauder JM, Gilmore JH. Cytokine regulation of embryonic rat dopamine
and serotonin neuronal survival in vitro. Int J Dev Neurosci 1997;15(6):711–6.
[58] Herschkowitz N, Kagan J, Zilles K. Neurobiological bases of behavioral development in the first year.
Neuropediatrics 1997;28(6):296–306.
[59] Hagberg H, Bona E, Gilland E, Puka-Sundvall M. Hypoxia– ischaemia model in the 7-day-old rat: possi-
bilities and shortcomings. Acta Paediatr Suppl 1997;422:85–8.
[60] Kalb RG, Fox AJ. Synchronized overproduction of AMPA, kainate, and NMDA glutamate receptors during
human spinal cord development. J Comp Neurol 1997;384(2):200–10.
[61] Court JA, Perry EK, Johnson M, Piggott MA, Kerwin JA, Perry RH, et al. Regional patterns of cholinergic
and glutamate activity in the developing and aging human brain. Brain Res Dev Brain Res 1993;74(1):
73–82.
[62] Andersen DL, Tannenberg AE, Burke CJ, Dodd PR. Regional development of glutamate-N-methyl-D-
aspartate receptor sites in asphyxiated newborn infants. J Child Neurol 1998;13(4):149–57.
[63] Uylings HB, Delalle I. Morphology of neuropeptide Y-immunoreactive neurons and fibers in human
prefrontal cortex during prenatal and postnatal development. J Comp Neurol 1997;379(4):523–40.
[64] Ding YQ, Zheng HX, Wang DS, Xu JQ, Gong LW, Lu Y, et al. The distribution of substance P receptor
(NK1)-like immunoreactive neurons in the newborn and adult human spinal cord. Neurosci Lett 1999;
266(2):133–6.
E. Herlenius, H. Lagercrantz / Early Human Development 65 (2001) 21–37 37


















