
Neuronal firing rates account for distractor effects on
13
B i o l C y b e r n ( 2 0 0 7 ) 9 6 : 4 0 7 4 1 9 D O I 1 0 . 1 0 0 7 / s 0 0 4 2 2 - 0 0 6 - 0 1 3 9 - 8 O R I G I N A L P A P E R Neuronal firing rates account for distractor effects on mnemonic accuracy in a visuo-spatial working memory task Julian Macoveanu · Torkel Klingberg · Jesper Tegnér R e c e i v e d : 3 J u n e 2 0 0 5 / A c c e p t e d : 2 N o v e m b e r 2 0 0 6 / P u b l i s h e d o n l i n e : 2 7 J a n u a r y 2 0 0 7 ' S p r i n g e r - V e r l a g 2 0 0 7 Abstract P e r s i s t e n t n e u r a l a c t i v i t y c o n s t i t u t e s o n e n e u r o n a l c o r r e l a t e o f w o r k i n g m e m o r y , t h e a b i l i t y t o h o l d a n d m a n i p u l a t e i n f o r m a t i o n a c r o s s t i m e , a p r e - r e q u i s i t e f o r c o g n i t i o n . Y e t , t h e u n d e r l y i n g n e u r o n a l mechanisms a re still elusive . Here , w e design a visuo- s p a t i a l d e l a y e d - r e s p o n s e t a s k t o i d e n t i f y t h e r e l a t i o n s h i p b e t w e e n t h e c u e - d i s t r a c t o r s p a t i a l d i s t a n c e a n d m n e - m o n i c a c c u r a c y . U s i n g a s h a r e d e x p e r i m e n t a l a n d c o m - p u t a t i o n a l t e s t p r o t o c o l , w e p r o b e h u m a n s u b j e c t s i n c o m p u t e r e x p e r i m e n t s , a n d s u b s e q u e n t l y w e e v a l u a t e d i f f e r e n t n e u r a l m e c h a n i s m s u n d e r l y i n g p e r s i s t e n t a c t i v - ity u sing an in silico p refrontal network model. F ive m o d e s o f a c t i o n o f t h e n e t w o r k w e r e t e s t e d : w e a k o r s t r o n g s y n a p t i c i n t e r a c t i o n s , w i d e s y n a p t i c a r b o r i z a t i o n , c e l l u l a r b i s t a b i l i t y a n d r e d u c e d s y n a p t i c N M D A c o m - p o n e n t . T h e v e n e u r a l m e c h a n i s m s a n d t h e h u m a n b e h a v i o r a l d a t a , a l l e x h i b i t e d a s i g n i c a n t d e t e r i o r a - t i o n o f t h e m n e m o n i c a c c u r a c y w i t h d e c r e a s e d s p a t i a l J . M a c o v e a n u · J . T e g n Ø r C o m p u t a t i o n a l B i o l o g y , D e p a r t m e n t o f P h y s i c s , L i n k p i n g U n i v e r s i t y o f T e c h n o l o g y , 5 8 1 8 3 L i n k p i n g , S w e d e n T . K l i n g b e r g D e p a r t m e n t o f W o m a n a n d C h i l d H e a l t h , M R c e n t e r , K a r o l i n s k a In s t i t u t e t , 1 7 1 7 6 S t o c k h o l m , S w e d e n J . T e g n Ø r C o m p u t a t i o n a l M e d i c i n e G r o u p , D e p a r t m e n t o f M e d i c i n e , K a r o l i n s k a In s t i t u t e t , 1 7 1 7 6 S t o c k h o l m , S w e d e n J . T e g n Ø r ( B ) D i v i s i o n f o r C o m p u t a t i o n a l B i o l o g y , D e p a r t m e n t o f P h y s i c s , I F M , L i n k p i n g U n i v e r s i t y o f T e c h n o l o g y , 5 8 1 8 3 L i n k p i n g , S w e d e n e-mail: j espert@ifm.liu.se d i s t a n c e b e t w e e n t h e d i s t r a c t o r a n d t h e c u e . A s u b s e - q u e n t c o m p u t a t i o n a l a n a l y s i s r e v e a l e d t h a t t h e r i n g r a t e a n d n o t t h e n e u r a l m e c h a n i s m p e r s e , a c c o u n t e d f o r t h e p o s i t i v e c o r r e l a t i o n b e t w e e n m n e m o n i c a c c u r a c y a n d s p a t i a l d i s t a n c e . Mo r e o v e r , t h e c o m p u t a t i o n a l m o d - e l i n g p r e d i c t s a n i n v e r s e c o r r e l a t i o n b e t w e e n a c c u r a c y a n d d i s t r a c t i b i l i t y . I n c o n c l u s i o n , a n y p h a r m a c o l o g i c a l m o d u l a t i o n , p a t h o l o g i c a l c o n d i t i o n o r m e m o r y t r a i n i n g p a r a d i g m t a r g e t i n g t h e u n d e r l y i n g n e u r a l c i r c u i t r y a n d a l t e r i n g t h e n e t p o p u l a t i o n r i n g r a t e d u r i n g t h e d e l a y i s p r e d i c t e d t o d e t e r m i n e t h e a m o u n t o f i n u e n c e o f a v i s u a l d i s t r a c t i o n . 1 Introduction W o r k i n g m e m o r y ( W M ) , t h e c a p a c i t y t o h o l d a n d m a n i p - u l a t e i n f o r m a t i o n d u r i n g a b r i e f p e r i o d o f t i m e , i s c e n t r a l f o r c o g n i t i v e p e r f o r m a n c e i n c l u d i n g l a n g u a g e , p r o b l e m solving and it also constitutes a basis for guiding future a c t i o n s ( N o r m a n 1 9 7 0 ; B a d d e l e y 1 9 8 6 ). T h e transient s t o r a g e g o v e r n e d b y W M i s a n a c t i v e a n d d y n a m i c p r o - c e s s , w h e r e p e r s i s t e n t a n d s t i m u l u s s e l e c t i v e e l e v a t e d n e u r a l a c t i v i t y h a s b e e n i d e n t i e d a s o n e n e u r a l c o r r e - late for v isuo-spatial WM ( G o l d m a n - R a k i c 1 9 9 5 ; W a n g 2 0 0 1 ) . P o s t e r i o r p a r i e t a l , i n f e r o t e m p o r a l a n d p r e f r o n t a l c o r t i c a l a r e a s a r e c l o s e l y a s s o c i a t e d w i t h W M , a s d e m o n - s t r a t e d b y l e s i o n , b r a i n i m a g i n g a n d e l e c t r o p h y s i o l o g i c a l s t u d i e s ( G o l d m a n - R a k i c 1 9 8 7 , 1 9 9 5 ; F u s t e r 1 9 9 5 ) . I f t h e persistent activity i s d isrupted by electrical stimulation o r b y d i s t r a c t i n g s t i m u l i d u r i n g t h e d e l a y p e r i o d , t h e cue-related i nformation in memory is lost ( F u n a h a s h i e t a l . 1 9 8 9 ; F u s t e r 1 9 9 7 ) . C o n s e q u e n t l y , n e u r a l c o n t r o l o f t h e s u s t a i n e d d e l a y a c t i v i t y a n d r o b u s t n e s s a g a i n s t d i s t r a c t o r s i s c r u c i a l f o r a n i n t a c t c o g n i t i v e a p p a r a t u s .
Transcript of Neuronal firing rates account for distractor effects on

Biol Cybern (2007) 96:407–419DOI 10.1007/s00422-006-0139-8
ORIGINAL PAPER
Neuronal firing rates account for distractor effects on mnemonicaccuracy in a visuo-spatial working memory task
Julian Macoveanu · Torkel Klingberg ·Jesper Tegnér
Received: 3 June 2005 / Accepted: 2 November 2006 / Published online: 27 January 2007© Springer-Verlag 2007
Abstract Persistent neural activity constitutes oneneuronal correlate of working memory, the ability tohold and manipulate information across time, a pre-requisite for cognition. Yet, the underlying neuronalmechanisms are still elusive. Here, we design a visuo-spatial delayed-response task to identify the relationshipbetween the cue-distractor spatial distance and mne-monic accuracy. Using a shared experimental and com-putational test protocol, we probe human subjects incomputer experiments, and subsequently we evaluatedifferent neural mechanisms underlying persistent activ-ity using an in silico prefrontal network model. Fivemodes of action of the network were tested: weak orstrong synaptic interactions, wide synaptic arborization,cellular bistability and reduced synaptic NMDA com-ponent. The �ve neural mechanisms and the humanbehavioral data, all exhibited a signi�cant deteriora-tion of the mnemonic accuracy with decreased spatial
J. Macoveanu· J. TegnérComputational Biology, Department of Physics,Linköping University of Technology,581 83 Linköping, Sweden
T. KlingbergDepartment of Woman and Child Health, MR center,Karolinska Institutet, 171 76 Stockholm, Sweden
J. TegnérComputational Medicine Group, Department of Medicine,Karolinska Institutet, 171 76 Stockholm, Sweden
J. Tegnér (B)Division for Computational Biology,Department of Physics, IFM,Linköping University of Technology,581 83 Linköping, Swedene-mail: [email protected]
distance between the distractor and the cue. A subse-quent computational analysis revealed that the �ringrate and not the neural mechanism per se, accountedfor the positive correlation between mnemonic accuracyand spatial distance. Moreover, the computational mod-eling predicts an inverse correlation between accuracyand distractibility. In conclusion, any pharmacologicalmodulation, pathological condition or memory trainingparadigm targeting the underlying neural circuitry andaltering the net population �ring rate during the delayis predicted to determine the amount of in�uence of avisual distraction.
1 Introduction
Working memory (WM), the capacity to hold and manip-ulate information during a brief period of time, is centralfor cognitive performance including language, problemsolving and it also constitutes a basis for guiding futureactions (Norman 1970; Baddeley 1986). The transientstorage governed by WM is an active and dynamic pro-cess, where persistent and stimulus selective elevatedneural activity has been identi�ed as one neural corre-late for visuo-spatial WM ( Goldman-Rakic 1995; Wang2001). Posterior parietal, inferotemporal and prefrontalcortical areas are closely associated with WM, as demon-strated by lesion, brain imaging and electrophysiologicalstudies (Goldman-Rakic 1987, 1995; Fuster 1995). If thepersistent activity is disrupted by electrical stimulationor by distracting stimuli during the delay period, thecue-related information in memory is lost ( Funahashiet al. 1989; Fuster 1997). Consequently, neural controlof the sustained delay activity and robustness againstdistractors is crucial for an intact cognitive apparatus.

408 Biol Cybern (2007) 96:407–419
Yet, the underlying neuronal mechanisms which governthe sustained activity and control the sensitivity againstdistractors have not yet been identi�ed. Computationalmodeling of cortical circuits is an important researchtool which facilitates an identi�cation of the contribu-tion from different synaptic and cellular factors ( Wang2001). Here, we use a biophysical network model oper-ating in different modes to produce persistent activitywhere our objective is to evaluate different cellular andsynaptic mechanisms which have been suggested to gen-erate persistent neuronal activity in connection with avisuo-spatial WM tasks. We used spatially distractingstimuli to investigate both the different regimes of thenetwork model and behavioral experiments performedon human subjects. Speci�cally, we investigated the rela-tionship between spatial distance to distractors and mne-monic accuracy in a visuo-spatial delayed-response task.Our integrated experimental and computational distrac-tor design enables us to correlate the spatial sensitivitybetween distractors and mnemonic accuracy for differ-ent neural mechanisms with human behavior.
For cortical networks in general, several broad clas-ses of neural mechanisms have been discussed. First,syn�re chains—chains of neuronal subgroups feedfor-ward connected—that permit a wave of synchronousactivity to be maintained through closed loops (Abeles1991; Diesmann et al. 1999; Gewaltig et al. 2001); syn-chronous oscillations as a basis for cortical dynamicssubserving perception, memory and cognition (Singer1993); networks operating with fast Hebbian plastic-ity ( Brunel 1996; Sandberg et al. 2003). However, themost accepted and biologically plausible mechanisticframework in the �eld of visuo-spatial or object workingmemory is a recurrent network model in which the per-sistent activity is represented as self-sustained dynami-cally stable states through reverberatory neural activity(Amari 1977; Hop�eld 1982 ; Amit 1995). This mecha-nism was �rst investigated in the 1930s by Lorente deNó and Hebb (1949) and refers to the �ow of neuralactivity due to the early observation that cerebral cor-tex has an abundance of recurrent synaptic connectionsbetween neurons. Several studies concerning the neu-ral mechanisms underlying WM activity have been per-formed using different neuronal and synaptic instancesof this basic recurrent neural network idea originat-ing from the work of Lorente de Nó and Hebb ( Seung1996; Amit and Brunel 1997; Fransen and Lansner 1998;Compte et al. 2000; Seung et al. 2000; Tegnér et al. 2002;Egorov et al. 2002; Koulakov et al. 2002; Goldman et al.2003; Loewenstein and Sompolinsky 2003; Wang et al.2004). These computational studies have revealed thateven though the mechanistic framework based on thereverberatory idea appears straightforward, there are
several non-trivial issues such as what controls the �r-ing rate and the stability of the neural activity duringthe delay-period. Thus, the major reason why there areseveral different versions of the original reverberatorycircuit is that those issues depend on the particularnature of the neuronal and synaptic mechanisms gov-erning the persistent activity. Our strategy here is toevaluate the major modes of operations by investigat-ing the consequences of a distractor in the different neu-ral modes of action for the network. The advantage isthat the outcome of those in silico experiments can bedirectly evaluated using the human behavioral experi-ments we have performed in the present study.
Due to the inherent stability problems with the recur-rent model several modi�cations have been suggestedsuch as an increased synaptic NMDA component andstronger or wider excitatory synaptic connections (Amitand Brunel 1997; Compte et al. 2000; Tegnér et al. 2002).Additionally, as it recently gained more experimentalsupport (Egorov et al. 2002), cellular bistability has beensuggested to stabilize the persistent activity (Koulakovet al. 2002). A bistable mode of action allows neuronsto jump between two stable activity states with low andhigh �ring rates. Goldman et al. (2003) reduced the largejumps in �ring rates commonly associated with bistabil-ity by incorporating multiple dendritic subunits. As aconsequence, the �ring rate of a neuron increases onlyincrementally as single dendritic compartments switchstate. To evaluate these different mechanistic proposals,we �rst implemented the biophysical network modelin a reference mode (A) from which we can modifythe model parameters to enable the following differentmodes of action: strong recurrent excitatory synapticconnections, referred to as (B) high E-E mode; widerexcitatory connections, referred to as (C) wide pro�lemode; a bistable mode of action, referred to as (D) bista-ble mode; and �nally a low NMDA synaptic component,referred to as (E) low NMDA mode.
2 Methods
2.1 Behavioral distractor trial
A visuo-spatial WM task using visual distractors dur-ing the delay period was performed by 31 participants(14 men and 17 women, average age 25.8± 4.6). Theywere asked to remember the spatial position of �veblue circles (cues) which appeared one at a time on a28.5× 21.5 cm computer monitor, 50 cm distance fromthe participant’s eyes. The cues and distractors couldappear at 16 possible locations on the perimeter of a

Biol Cybern (2007) 96:407–419 409
Fig. 1 Schematic diagram showing an example of the delayed-response task. The participants �xed their gaze on a central spotduring the entire task. After a �xation screen of 500 ms �ve cues(circles) and one distractor (square) were presented at differentlocations on the peripheral circle with 150 ms presentation timeand 750 ms inter-cue delay. Distractors followed the third or thefourth cue at �ve distances, 22.5◦, 45◦, 67.5◦, 90◦ and 180◦. Afterthe cue presentation the participants indicated in an orderly man-ner the location of each cue with the computer mouse
circle of 20.0 cm diameter which was visible during theentire task (Fig. 1).
The participants initiated the trial �xating a point for500 ms in the center of the screen and were asked tomaintain �xation during the entire trial. The �xationperiod was followed by a succession of �ve cues (bluecircles) and one intervening distractor (blue square),with 150 ms presentation time and 750 ms delay betweenthe cues. The last cue was followed by a delay periodof 750 ms after which the cursor of a computer mouseappeared and the participants had to respond by clickingon the remembered cue locations.
In every trial either the third or the fourth cue wasfollowed by a distractor that was to be ignored. Thedistractor could be presented at �ve spatial distancesrelative to the memorized cue, 22.5◦, 45◦, 67.5◦, 90◦ and180◦ angles with the vertex on the �xation point. Thus,360◦ represented the whole span of the visual �eld. Theshorter the distance the higher was the expected inter-ference of the distractor with the memory of the cue.A total of 40 trials were performed in random order byevery participant.
The distractor effect was evaluated by means of accu-racy, de�ned as the spatial distance (measured indegrees) from the presented cue to the location indi-cated by the participant at the end of the trial ( ∼4 s aftercue onset).
The participants responded by clicking on the screen(continuous range of locations) for all �ve memorizedcues in the order of appearance. If the distance from thecue location to the location pointed out by the partici-pant (accuracy) exceeded a prede�ned threshold of 20◦the cue location was considered forgotten and the trial
was discarded. Only the accuracy of the cue followed bydistractor and its corresponding control (no distraction)were taken into consideration.
The signi�cance of the behavioral results was assessedusing one-way repeated-measure ANOVA and two-sided paired t-test.
2.2 Recurrent network model
The neural network model used for the simulations waspreviously studied byTegnér et al.(2002) and representsa local cortical circuit capable of upholding states of per-sistent neuronal activity accounting for the memory stor-age in WM (Amit and Brunel 1997; Wang 1999; Compteet al. 2000;Tegnér et al. 2002). The model included 2 neu-ronal populations, 512 excitatory (E) and 128 inhibitory(I) cells (Fig.2a). The neurons were spatially distributedin a ring in accordance with the preferred visual angleof the stimulus location. Thus, the neurons respondedselectively to visual stimuli depending on their spatiallocation in the visual �eld ( Ben-Yishai et al. 1995),resembling previous oculomotor delayed-response taskexperiments (Funahashi et al. 1989; Goldman-Rakic1995). The neurons were parameterized by the angleθ (0◦–360◦) that speci�ed their preferred visual angle.Figure 2b shows a schematic diagram of the connectivitystructure. The architecture of the model and its recurrentconnectivity resembles the columnar organization of theprefrontal cortex ( Levitt et al. 1993; Goldman-Rakic1995; Kritzer and Goldman-Rakic 1995; Mountcastle1997). The neuron models used followed the Hodg-kin–Huxley formalism with action potential pro�le andneuronal input–output relation calibrated using cortical-slice data (McCormick et al. 1985; Markram et al. 1997)and the postsynaptic current gating kinetics of AMPA,NMDA ( Hestrin et al. 1990; Jahr and Stevens 1990;Spruston et al. 1995) and GABA A receptors (Amari1977; Salin and Prince 1996; Xiang et al. 2002).
Excitatory neurons were of pyramidal type with threecompartments, representing soma, proximal and distaldendrites. The somatic compartment containsINa andIK currents, a high-threshold calcium current ICa, anda slow calcium-dependent cationic current ICAN . Theproximal dendritic compartment has a persistent sodiumcurrent INaP and a slowly inactivating potassium currentIKS. The distal dendritic compartment has an ICa and atransient A-type potassium current IA . Additionally, allneuronal compartments included a leak current IL . Themembrane equations for the somatic voltageVs, prox-imal dendritic voltage Vd1, and distal dendritic voltageVd2 were

410 Biol Cybern (2007) 96:407–419
Fig. 2 Schematic representation of the model connectivity.a Twopopulations of neurons 512 excitatory (E) and 128 Inhibitory ( I)cells with all-to-all connections both within and between the pop-ulations. b The strength of the recurrent connections betweenexcitatory cells depended on their proximity, represented by theangle between them. Inhibitory projections, which extend to allcells, are evenly distributed across the pyramidal and inhibitoryunits of the network. See text for more details
CmdVs
dt= −INa − IK − ICa − IL − ICAN
−gc1(Vs − Vd1)
p1 − Isyn
CmdVd1
dt= −INaP − IKS − IL − gc1(Vd1 − Vs)
p2
−gc2(Vd1 − Vd2)
p2 − Isyn
CmdVd2
dt= −IA − ICa − IL − gc2(Vd2 − Vd1)
(1 − p1 − p2) − Isyn.
The inhibitory neurons were of fast-spiking type andwere represented by one compartment containing onlyspike generating sodium and potassium currents. Themembrane equation was
CmdVdt
= −INa − IK − ICa − IL .
Synaptic currents were modeled according to Isyn =gsyns(V − Esyn), where gsyn represents the maximal syn-
aptic conductance,Esyn the synaptic reversal potentialand s is the gating variable which decides the fraction ofopen synaptic ion channels. A complete description ofneuron models, with details of ion channel kinetics andconductance parameters, can be found in (Tegnér et al.2002).
The synaptic connection strength between excitatoryE neurons decreases with the angle between the pre-ferred cues of two neurons following a smooth Gauss-ian connectivity drop-off. The connectivity W betweenneuron i and j is described by (Compte et al. 2000)
W(θi − θj) = J+ + (J− − J+)e−(θi−θj)2/2σ2
.
W(θi − θj) is the normalized connectivity. TheJ− parameter represents the strength of the weak distantconnections, J+ the strength of the stronger adjacentconnections andσ the footprint of the connectivity. Theexcitatory connection strength from E to I as well asall inhibitory connection from I neurons is indepen-dent on the inter-neuronal distance and is evenly dis-tributed across the E and I units of the network. Therecurrent excitatory synaptic conductances of the modelwere mediated by NMDA and AMPA receptor chan-nels. Previous theoretical studies (Amari 1977; Lismanet al. 1998; Wang 1999; Compte et al. 2000; Tegnér et al.2002) revealed that a dominant slow excitation medi-ated by NMDA receptors stabilizes the �ring rate of thepyramidal neurons generating the sustained activity.
2.2.1 Reference parameters
Conductances (mS/cm2). NMDA channels: gEE = 1.67,gEI = 1.19; AMPA channels:gEE = 0,gEI = 0; GABAreceptors:gIE = 1.29,gII = 0.65.
Connectivity. J+ = 5.25,σ = 0.05.Overall external excitation: uncorrelated Poisson spike
trains of 1,000 Hz per cell mediated by AMPA recep-tors; conductances (mS/cm2): gext,E = 0.235,gext,I =0.042.
2.2.2 Modeled neural mechanisms
We studied �ve distinct network instances, all based onexcitatory reverberation, but with potentially alterna-tive neural mechanisms: reference mode (A), high E–Emode (B), wide pro�le mode (C), bistable mode (D)and low NMDA mode (E). The distinction was assessedby simulations of all network instances using a commonsimulation protocol with one cue and one distractor.
The reference mode (A), which used the unchangedreference parameters, was characterized by low connec-tivity between the excitatory cells. The rest of the net-work modes contained a single modi�ed characteristic

Biol Cybern (2007) 96:407–419 411
compared to the reference mode. For the high excit-atory connectivity mechanism employed in the high E–Emode (B), the peak conductance of the NMDA chan-nels was increased,gEE = 1.82 mS/cm2. The wide pro-�le mode (C) used a wide footprint of the excitatoryconnectivity. This was achieved by simultaneouslydecreasing the strength of the adjacent E–E connec-tion (J+ = 4) and increasing the connectivity footprint(σ = 0.07).
The bistable mode (D) used excitatory units with a bi-stable regime in their �ring pattern. Cellular bistabilityhave been attributed to voltage-dependent componentsthat can exhibit self-sustained activation, such as theNMDA channel and voltage-sensitive Ca2+ channels(Lee and Heckman 1998; Schiller and Schiller 2001).Our model incorporated a Ca2+-activated non-selectivecationic (CAN) current into the membrane of the distaldendritic compartment of the pyramidal cells ( Tegnéret al. 2002). This slow calcium-dependent current hasbeen identi�ed in rat prefrontal neurons ( Haj-Dahmaneand Andrade 1998). The bistable behavior of the neuro-nal units allows them to switch between a low state anda high state of activity depending on the temporary pro-gress of the current magnitude. The size of the bistableregime is proportional to the magnitude of the ICANchannel conductance (gCAN ). We used a gCAN valueof 0.2 mS/cm2 to assure a signi�cant proportion of theactive units in the high mode of �ring. One observedadvantage of networks composed of bistable units istheir ability to resist parameter modi�cation to a higherdegree. The mechanisms underlying bistability are notthought to in�uence this attribute ( Koulakov et al. 2002).Nonetheless, the biological substrate for bistable neu-rons in human prefrontal cortex remains elusive.
For the low NMDA mode (E) the excitatory syn-aptic transmission was dominated by AMPA receptorcurrents. As previous work suggested, a substantialAMPA component will destabilize the network by theinduced synchronous activity of the neurons (Wang1999; Compte et al. 2000; Gutkin et al. 2001). TheNMDA-to-AMPA transmission ratio was graduallyreduced while preserving the total synaptic input currentuntil the persistent activity state was lost. For the LowNMDA mode we used a ratio of 30% NMDA receptorcurrent. The conductances of the NMDA and AMPAreceptors were calculated using their relative contribu-tion to the time integral of a unitary excitatory postsyn-aptic current (EPSC) at −65 mV (Compte et al. 2000).
2.2.3 Network simulation protocol
The simulation protocol was chosen to replicate thebehavioral distractor trial protocol. However, only the
cue with the following distractor have been simulated inthe network instances. The models received only back-ground activity the �rst 500 ms of the simulation. Bothcues and distractors were presented as synaptic inputswhich lasted for 150 ms and had identical stimulationamplitudes. The cue consisted of a 0.8µA/cm2 currentinjected in the distal dendritic compartment which wasadministrated to a population of 20 pyramidal neuronshaving the preferred angle between 160◦ and 180◦. Thedistractor current had identical characteristics as the cue,and was elicited 750 ms after the cue onset at ten differ-ent locations on the cell ring, ranging from 22.5◦ to 180◦and including the �ve locations from the behavioral trial.
3 Results
We studied the relationship between spatial distance todistractors and mnemonic performance in a behavioralWM task. The experimental protocol was used in a com-puter simulation of sustained activity in the prefrontalcortex. Five distinct neural mechanisms were tested indifferent modes of a recurrent network model.
3.1 Visuo-spatial WM task
The behavioral results for the 31 participants are pre-sented in Fig. 3. The average accuracies (±SEM) asa function of the distractor distance were 7.4± 3.8◦,7.2± 2.7◦, 6.8± 2.9◦, 6.6± 2.4◦, 6.5± 3.1◦, for the dis-tractor distances of 22.5◦, 45◦, 67.5◦, 90◦, 180◦, respec-tively, and 6.1± 2.6◦ for the undistracted control.A t-test for the difference between average accuraciesof distractor distances and control was signi�cant onlyfor the closest distractor (P < 0.05 for 22.5◦ and P > 0.05for longer distractor distances). If the individual aver-ages were taken into consideration, a one-way ANO-VA testing the dependency on the distractor distancewas found signi�cant (P < 0.05). The post-test for lin-ear trend between the means for every distance washighly signi�cant ( P < 0.01, slope−0.55). Furthermore,accuracy slopes calculated for all participants were sig-ni�cantly different form the null hypothesis ( t-test, P <
0.05). The behavioral results suggest that a cue which issimilar to a distractor will perturb the remembered tar-get location with the highest effect being achieved withthe smallest distractor–target distance.
3.2 Neural network model
3.2.1 Spatiotemporal firing patterns
Figure 4 shows the raster plots for the network modelusing the reference parameters. All network instances

412 Biol Cybern (2007) 96:407–419
Fig. 3 Behavioral task results. The average accuracy drop-off forall participants as a function of the distractor distance: higher dis-tance leads to better accuracy (lower values). Visual distractorsappeared at 22.5◦, 45◦, 67.5◦, 90◦ and 180◦. The control baselinerepresents average accuracy for undistracted targets. The pointsare �tted with an exponential decay curve
used the same stimulation protocol as the behavioral dis-tractor trial. The initial state of the network (0–500 ms)was dominated by background activity. A transient cuestimulus of 150 ms triggers the formation of a spatiallylocalized persistent activity state (bump attractor) whichwas sustained during the whole simulation time (Fig.4a).The raster plot in Fig. 4b exempli�es a simulation withdistractor induced at 56.25◦ distance. We used the accu-racy of the bump state location to quantify the distractordistance effect. The accuracy was calculated as the aver-age difference(θA ) between the initial cue location rightafter the cue onset time and the bump state location aftera delay period of 4 s (Fig.4c).
3.2.2 Population firing profile
Electrophysiological studies performed on theprefrontal cortex of monkeys have reported neuronal�ring rates of around 20 Hz for the neurons involvedin the delay activity of delayed response tasks (Fuster1973;Funahashi et al. 1989). However, more recent stud-ies have observed �ring rates up to more than 60 Hzwhen the cue is preferred (Constantinidis et al. 2001;Compte et al. 2003). By allowing the slow NMDA cur-rents to dominate the synaptic transmission, cortical net-work models report �ring rates in the range of 20–40 Hz(Compte et al. 2000; Durstewitz et al. 2000).
Figure 5a shows the bump population �ring pro�leaveraged over the last 500 ms of the simulation time.The reference mode (A) demonstrated a narrow tun-ing curve with low �ring rates for the bump attrac-tor population (average 22.9 Hz). The high E–E mode(B) approached to the highest degree biophysical data,showing a wide tuning curve, close to physiologicalshapes found in monkeys (Funahashi et al. 1989) and
Fig. 4 Spatially localized persistent activity state of the networkmodels. The dots correspond to action potentials from pyramidalcells. The x axis represents time, whiley axis the preferred cue(0◦–360◦) of the neurons. The cue is presented during 150 ms, to asmall fraction of the neurons with a preferred cue at 180◦. A per-sistent activity state is induced by the cue which lasts during theentire delay period. The lower trace represents the average �ringrate (100 ms bins) for the bump attractor population. a Networksimulation of the reference mode. b Reference mode with a dis-tractor induced at 56.25◦ from the memorized cue. c Example ofaccuracy drop-off of the memorized cue calculation for the refer-ence mode with distractor presented at 56◦ (not shown). The angleθA is the average difference between the initial bump location andpost-delay location
bump �ring rates still within physiological range (aver-age 37.5 Hz). The reduced adjacent E–E connectionstrength of the wide pro�le mode (C) resulted in lowbump �ring rates (average 20.5 Hz). The bistable unitsof the bistable mode (D) showed as expected elevated�ring rates (average 32.1 Hz). The low NMDA mode(E) had a narrow tuning curve with very high �ringrates (average 45 Hz). If a distractor is induced in thenetwork the �ring rates of the initial bump measuredimmediately after the distractor onset will be lowered,with the highest effect observed for the closer distrac-tors (Fig. 5b). Resistance to distractor in�uence provesto be positively correlated with the �ring rates acrossthe �ve network models tested.
3.2.3 Dependence on distractor distance
Figure 6a shows the accuracy of the memorized cue forall �ve studied neural mechanisms as a function of the

Biol Cybern (2007) 96:407–419 413
Fig. 5 Firing rate plots. a Population �ring pro�le. Average �r-ing rates of the undistracted memory bump for the last 500 ms ofthe simulation time. All modes except the low NMDA mode gen-erated �ring rates within physiological values. Reference mode:average 22.9, peak 36 Hz. High E-E mode: average 37.5, averagepeak 56 Hz. Wide pro�le mode: average 20.5, peak 38 Hz. Bistablemode: average 32.1 peak, 52 Hz. Low NMDA mode: average 45.0,peak 78 Hz. (B) Firing rates of the memory bump in a time windowof 500 ms directly after distractor onset as function of distractordistance. The points are average of eight noisy simulations and�tted with an exponential decay curve
distractor distance. The modeled mechanism demon-strated to have a clear distractor distance effect, withthe highest effect observed at the two closest distractordistances (22.5◦ and 33.75◦). This distance dependencyresembles the connectivity curve of the E population(Fig. 2b) which presented a Gaussian drop-off in gEE
with the E–E distance. The smooth drop-off in accu-racy with distractor distance of the high E–E (B) andlow NMDA (E) modes resembled to the highest degreethe behavioral results. Differentiating these mechanismsfrom the mechanisms with low excitatory connectivity,the reference (A), wide pro�le (C) and bistable (D)modes which showed a similar close distractor effectseems straightforward. However, the similar behaviorof the unrelated mechanisms of high E–E (B) and lowNMDA (E) modes as well as the dissimilar performanceof the closely related reference (A) and high E–E (B)modes indicate that the neural mechanism per se isinsuf�cient to account for the distractor distance effect.We therefore tested whether the difference in the accu-racies of the memorized cue is mainly attributed to the�ring rate of the bump attractor. For this, we adjusted the
inhibitory feedback (gIE ) to the excitatory cells in orderto alter the �ring rates of the �ve mechanisms and bringthem in the smallest range possible. Figure6b showsaccuracy as function of distractor distance for modes A,B, C and D with bump �ring rates adjusted to 24.8, 26.5,26.3 and 23.4 Hz, respectively. The �ring rates of the lowNMDA (E) mode could not be reduced to values lowerthan 44.8 Hz, thus this mechanism was ineligible. For theclosest distractor distance (22.5◦), the four mechanismsfall within an accuracy range of 7.8◦ when operating atoriginal �ring rates (Fig. 6a). This range decreased to2.9◦ with the adjusted �ring rates. For the longer dis-tractor distances the range was reduced from 3.9 to 1.9◦.
The low NMDA mode (E) was surprisingly robust todistractors. We explain this as a consequence of soar-ing �ring rates of the preferred cue neuronal population(average 45 Hz). The drift of the bump is stabilized dueto the high level of feedback inhibition that promptlyrejects any new incoming stimuli. Given the fast decayof AMPA currents, AMPA synapses remain unsaturatedwhile NMDA receptors saturate during the persistentactivity state of the network. For the same input current,AMPA needs to surpass NMDA receptors peak conduc-tance, consequently a dominant AMPA component tothe total synaptic drive leads to elevated �ring rates.
In a subsequent analysis we mapped the whole feasi-ble range of �ring rates for the �ve neural mechanismsas a function of accuracy. All network modes showeda clear correlation between �ring rate and accuracy(Fig. 6c). Although showing the same accuracy—�ringrate correlation, the low NMDA (E) mode can be iso-lated from the rest of the studied mechanisms due toelevated bump �ring rates.
These results suggest that the �ring rate of the bumpattractor is the key factor in�uencing the accuracy of thememorized cue. However, the �ring rate may not be theonly factor responsible for the memory drift, but the par-ticular �ring pattern given by a neural mechanism mayaccount for some variation as all points in Fig. 6c didnot fall on a universal line. The low NMDA (E) mode inparticular, which generated highly synchronous activity,showed higher accuracy drop-offs than expected for itsbump �ring rates.
3.2.4 Mode-dependent parameter range
We mapped the span of the excitatory connectionstrength mediated by the NMDA receptor channelswithin the limits imposed in order to preserve the dyna-mic stability of the network. We gradually increased thepeak conductance of NMDA transmission from 1.67to 1.82 mS/cm2 (Fig. 7a). All other parameters wereunchanged. The lower limit marks the least synaptic

414 Biol Cybern (2007) 96:407–419
Fig. 6 Accuracy drop-off analysis for the �ve studied mecha-nisms.a Accuracy drop-off as a function of the distractor distance.Using the standard simulation protocol all �ve neural mechanismswere tested with distractors induced at ten distances, spanningfrom 22.5◦ to 180◦ from the initial location of the memorized cues.The series are averages of eight noisy simulations with the con-trol baseline (no distractor) subtracted. The points are �tted withan exponential decay curve. The distractor distance effect wasfound to be of similar magnitude in the behavioral experimentcompared to the High E-E and Low NMDA modes. b The �ringrates of the �ve network modes were adjusted to similar valuesproducing a much narrower range of accuracy drop-offs betweendifferent modes. c Accuracy drop-offs as a function of �ring rateof the memory bump. The entire �ring rate range of all modes wasmapped by means of inhibitory feedback to excitatory cells (gIE )
variation. The standard simulation protocol with a distractor at33.75◦ was used
current required to uphold the cue-induced bumpattractor during the entire simulation time when a dis-tractor is applied at 33.75◦. The value chosen as refer-ence (1.67 mS/cm2) was the smallest value to satisfy this
condition. Beyond the higher limit random persistentactivity destabilizes the network. We used gEE = 1.82mS/cm2for the high excitatory connectivity simulations.The highest accuracy (lowest value) was observed in themiddle of the interval (1.73< gEE < 1.78). Widening theconnectivity footprint also seemed to improve the accu-racy (Fig. 7b). σ was increased from the reference value0.050 to 0.062, value that rendered �ring rates of thebump which exceeded physiological values. Increasingthe gCAN value increased the depolarizing currents thatentered the dendritic compartments resulting in almostmonotonic accuracy improvement (Fig. 7c)
The fast excitation mediated by the AMPA recep-tors followed by a slower inhibition mediated by theGABA receptors promotes oscillations which destabi-lize the network ( Wang 1999). Although the relativecontribution of NMDA and AMPA receptors is still amatter of debate, physiological studies have found per-sistent activity to be more sensitive to NMDA recep-tor block than to AMPA receptor block ( Dudkin et al.1997; Shima and Tanji 1998; Aura and Riekkinen 1999).Additionally, the human prefrontal cortex is reportedto have the highest NMDA receptor density of all cor-tical areas (Scherzer et al. 1998). While preserving thetotal synaptic drive we gradually decreased the NMDA-to-AMPA ratio until the persistent activity state wasdestabilized (Fig. 7d). If the NMDA receptor contri-bution is dominant the neuronal �ring is asynchronous( > 75% NMDA). A decreased NMDA receptor compo-nent synchronizes the �ring pattern and the sustainedactivity is lost (<25% NMDA). Interestingly, increas-ing the AMPA component improves the accuracy of thememorized cue, with the highest accuracy attained in aninterval between 40 and 90% NMDA contribution tothe total synaptic drive.
We further investigated how the accuracy criterion iscorrelated to the distractibility of the network model.Distractibility re�ects the capacity to uphold the persis-tent activity state in the presence of distractingstimuli. Accuracy was altered by adjusting the excit-atory connection strengths within physiological range(gEE = 1.57− 1.72 mS/cm2). We mapped the small-est intensity of the distractor current that successfullyled to the extinction of the bump attractor (Fig. 7e).We found an inverse correlation between accuracy anddistractibility of a memorized cue.
4 Discussion
Our results strengthen the current hypothesis whichattributes the temporary storage of the information tostimulus selective persistent neural activity—the
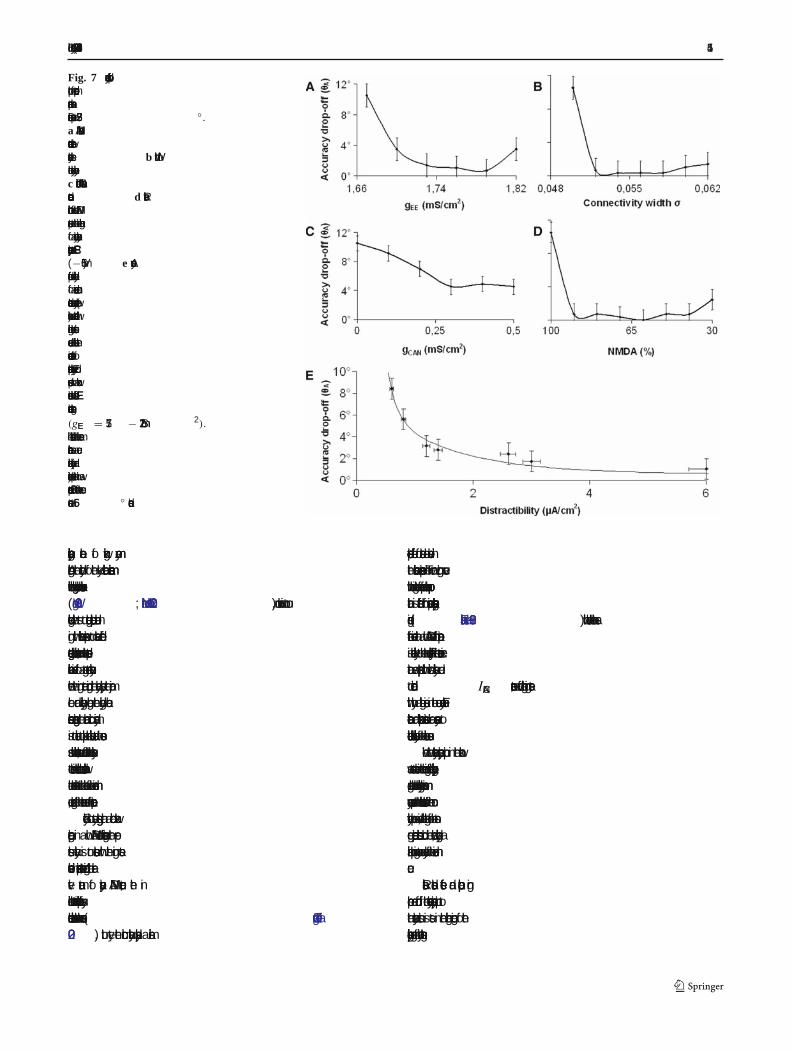
Biol Cybern (2007) 96:407–419 415
Fig. 7 Accuracy drop-offplot for mode-dependentparameter variation.Distractor present at 33.75◦.a NMDA channelconductance betweenexcitatory cells. b Width ofthe excitatory connectivity.c Dendritic CAN channelconductance.d Relativecontribution of the NMDAreceptors to the time integralof a unitary excitatorypostsynaptic current EPSC(−65 mV). e Accuracydrop-off versus distractibilityof a memorized cue.Increased cue accuracy (lowervalues) was correlated withhigher distractor currentsneeded to abolish thecue-induced state ofpersistent activity. Every dotcorresponds to a networkinstance with different E-Econnections strengths(gEE = 1.57− 1.72 mS/cm2).The distractor had the samecharacteristics as a cuestimulus (intensity andduration) except that it waspresented 500 ms after the cueonset at 56◦ distance
neurophysiological correlate of working memory.Although the identity of the key cellular mechanismwhich underlies working memory has been controversial(Wang 2001; Miller and Cohen 2001), our work is to ourknowledge the �rst to integrate computational model-ing with behavioral experiments to evaluate differentsuggested neural mechanisms. Here, we have presentedbehavioral evidence for a strong recurrent connectivitywhere the �ring rate during the delay plays the majorrole and interestingly enough, the underlying neuralmechanisms generating the neuronal circuit dynamicsis not crucial to explain the behavioral data. In consen-sus with behavioral results, we found that the sensitivityto distractors in recurrent cortical models decreases withthe distractor distance to the location of the memorizedcue, regardless of the neuronal mode of operation.
Secondly our study suggests that a cortical networkoperating in a low NMDA mode sustaining the per-sistent activity is not consistent with the �ring ratesobserved in experiments. Moreover, reducing the rela-tive amount of synaptic NMDA current, the insilico network demonstrated increased potential for syn-chronous oscillation between the neurons (Tegnér et al.2002), but yet the circuit actually displays a similar
distance-dependent effect of distractors as observed inthe behavioral experiment. The narrow tuning curvewith elevated �ring rates of preferred cue neuronal pop-ulation is different from previous electrophysiological�ndings ( Funahashi et al. 1989) which could be taken asfurther evidence that a low NMDA mode of operationis less likely to be the case. Finally, if the recurrent exci-tation was supplemented with intrinsic bistability dueto dendritic ICAN currents, we found higher �ring rateswith only a slight increase in the cue accuracy. Furthertheoretical and experimental studies are necessary toelucidate cellular bistability of the cortical neurons.
When the excitatory synaptic input in the networkwas set to values in the lower regime of the physiologicalrange, the model showed a signi�cantly lower mnemonicaccuracy compared to the behavioral results. If the con-nectivity pro�le was widened, the strength of the stron-ger adjacent connections could be lowered, yielding asimilar bump �ring rate and accuracy of the memorizedcue.
Realistic distractor effects and population �ringpro�le are found if the reverberatory synaptic input tothe excitatory units is set in the higher region of thephysiological range of connectivity strengths.

416 Biol Cybern (2007) 96:407–419
4.1 Accuracy of memory location
For the simulation study, we considered the differencebetween the accuracy drop-offs of the undisrupted cuesand cues followed by distractors. This difference wascompared with the corresponding behavioral values andthe similarity gave the quantitative basis when evaluat-ing the �ve tested neural mechanisms. We deliberatelydid not use the absolute accuracy drop-off values forevaluation purposes as the undisrupted “base line” accu-racy drop-off depends not only on the memory decay butalso on factors like the maximum value allowed for theaccuracy drop-off (20◦ here, see Sect.2) and the frac-tion of the visual �eld the presentation screen covers.As the network model did not account for these sourcesof uncertainty a comparison of the behavioral and simu-lated absolute accuracy drop-offs cannot be performed.
Following experimental results like ( Miller et al. 1996)several previous studies (Koulakov et al. 2002; Brodyet al. 2003) have stressed the necessity of robustness todistractors of cortical networks representing WM. Theelevated non-selective inhibition rate of a recurrent net-work in persistent activity state offers resistance to inter-vening stimuli. This is true as long as distractor intensityis low or the distance between the bump state and dis-tractor stimulus is suf�ciently large. The persistent activ-ity state of the network will eventually subside for highdistractor amplitudes. The E to E connectivity structureof the model is the key factor understanding the distrac-tor distance effect observed in recurrent models.
The observed instability of the memorized target loca-tion after a variable delay period has been previouslystudied in humans (Ploner et al. 1998) and monkeys(White et al. 1994). The loss in accuracy was attributedto both systematic errors that were similar in size forany studied delay intervals and to variable errors thatincreased monotonically as delay intervals were length-ened. Theoretical studies have accredited the randomdrift of memory to tuned inhibitory feedback on excit-atory cells (Ben-Yishai et al. 1997). Our computationalstudy suggests that the magnitude of the memory drift iscorrelated to the distractibility of the cue. High memorydrifts imply high distractibility. The �nding is howeveryet to be observed in behavioral experiments.
4.2 Relation to multiple-item representations
The present computational study represents one targetcue along with the following distractor stimulus. Thisapparent difference between the computational modeland the experimental design could be interpreted asweakness of the study. However, because this class ofWM models can maintain multiple items ( Macoveanu
et al. 2006), simulating an isolated cue and distractor ishighly representative even for a multiple-item model ofthe delayed-response task. The accuracy criterion usedre�ects intrinsic functional characteristics of the neuro-nal units and their recurrent connections, being indepen-dent of the number of represented cues. Our �nding thatthe �ring rate is the main determinant of the effect of adistractor further supports the notion that the particularneuronal mechanisms underlying a multi-item mode ofoperation is of less relevance here.
4.3 Experimental design considerations for distractingthe human working memory
An important property of the prefrontal cortex is theextent to which it can retain information in WM in spiteof distraction ( Cornette et al. 2001; Sakai et al. 2002).There are numerous studies showing how working mem-ory capacity interacts with interference. Subjects withlower working memory capacity are more prone to makeerroneous saccades to irrelevant stimulus locations(Roberts et al. 1994; Kane et al. 2001). WM showed to becrucial for directing attention appropriately in selectiveattention tasks through the active maintenance of stim-ulus priorities. Consequently, a high working memoryload increases the interfering effect of a distractor (deFockert et al. 2001; Lavie et al. 2004). It thus seems asthe relationship between working memory capacity andworking memory load determines how much interferingeffect a distractor has. Our behavioral task incorporatesprevious psychological and experimental knowledge ofworking memory capacity and distractibility. Interest-ingly, initially it proved dif�cult to distract the humansubjects. We had therefore to consider the experimentaldesign carefully. The delayed-response task was con-structed to maximize the task demand thus facilitatingdistraction. This was achieved by using a number ofcues that tapped the general visuo-spatial WM storagecapacity (Todd and Marois 2004; Vogel and Machizawa2004) and distractors which resembled the visual stim-uli ( Treisman and Gelade 1980). Furthermore, we con-sidered the distracted cue from the middle of the cuesequence because of the lower retention probabilityof the target as predicted by the serial position effect(Thomas 1968).
4.4 Outlook and experimental predictions
The present study makes several statements and pre-dictions. For the ensuing discussion it should be notedthat a small accuracy drop-off means that a manip-ulation has a small effect on the accuracy; the mne-monic accuracy is therefore high under this condition.

Biol Cybern (2007) 96:407–419 417
First, mnemonic accuracy is directly proportional to thespatial distance between cue and distractor, as validatedby both behavioral (Fig. 3) and computational exper-iments (Fig. 6a, b). A nearby distractor perturbs thecue more than a distant distractor and the behavioralresponse is consequently less accurate. Secondly, thecomputational analysis demonstrates that accuracy isalso inversely proportional to distractibility (Fig. 7e).Third, it can therefore be inferred that the distancebetween the cue and distractor is inversely proportionalto distractibility. Hence, a behaviorally weak distractorat small distance is predicted to have a similar effect onthe mnemonic accuracy as behaviorally stronger or morerelevant distractor at a larger distance from the encodedcue. To the best of our knowledge this has not yet beenexamined experimentally. Fourth, the study revealedthat the �ring rate and not the neural mechanism per se isdirectly proportional to the mnemonic accuracy (Fig. 6).Fifth, recent experimental studies (Klingberg et al. 2005)have demonstrated that the capacity of working mem-ory, the ability to maintain several items across timecan be improved through computerized working mem-ory training. A neuronal correlate to this remarkableresult has recently been identi�ed. Imaging experiments(Olesen et al. 2004) detected that working memorytraining induces a selected increase of the brain activ-ity. However, the neuronal mechanisms underlying theeffects of the training are yet unclear. The present study,which demonstrates the positive correlation betweenthe mnemonic accuracy and �ring rate, suggests thatthe observed increase in brain activity could correlatewith an increased �ring rate, and therefore increasedmnemonic accuracy. A recent experimental study con-ducted by Olesen et al. (2005) con�rms a tight rela-tion between WM capacity, delay related elevated brainactivity and mnemonic accuracy. Those results in con-junction with the results of the present study predict that,after the memory training, stronger distracting stimuliare required to disrupt the mnemonic activity during adelay-period task. This is a testable suggestion. Finally,if the working memory training affects neural mecha-nisms in such a manner that the delay-related �ring rateincreases, then the present study accounts for the bene-�cial effects on mnemonic accuracy and distractibility.
References
Abeles M (1991) Corticonics: neural circuits of the cerebral cortex.Cambridge University Press, Cambridge
Amari S (1977) Dynamics of pattern formation in lateral-inhibi-tion type neural �elds. Biol Cybern 27:77–87
Amit DJ (1995) The Hebbian paradigm reintegrated: localreverberations as internal representations. Behav Brain Sci18:617–626
Amit DJ, Brunel N (1997) Model of global spontaneous activityand local structured activity during delay periods in the cerebralcortex. Cereb Cortex 7:237–252
Aura J, Riekkinen P Jr (1999) Blockade of NMDA receptorslocated at the dorsomedial prefrontal cortex impairs spatialworking memory in rats. Neuroreport 10:243–248
Baddeley A (1986) Working memory. Oxford University Press,New York
Ben-Yishai R, Bar-Or RL, Sompolinsky H (1995) Theory oforientation tuning in visual cortex. Proc Natl Acad Sci USA92:3844–3848
Ben-Yishai R, Hansel D, Sompolinsky H (1997) Traveling wavesand the processing of weakly tuned inputs in a cortical networkmodule. J Comput Neurosci 4:57–77
Brody CD, Romo R, Kepecs A (2003) Basic mechanisms forgraded persistent activity: discrete attractors, continuous attrac-tors, and dynamic representations. Curr Opin Neurobiol13:204–211
Brunel N (1996) Hebbian learning of context in recurrent neuralnetworks. Neural Comput 8:1677–1710
Compte A, Brunel N, Goldman-Rakic PS, Wang XJ (2000) Syn-aptic mechanisms and network dynamics underlying spatialworking memory in a cortical network model. Cereb Cortex10:910–923
Compte A, Constantinidis C, Tegnér J, Raghavachari S, ChafeeMV, Goldman-Rakic PS, Wang XJ (2003) Temporally irregularmnemonic persistent activity in prefrontal neurons of monkeysduring a delayed response task. J Neurophysiol 90:3441–3454
Constantinidis C, Franowicz MN, Goldman-Rakic PS (2001) Cod-ing speci�city in cortical microcircuits: a multiple-electrodeanalysis of primate prefrontal cortex. J Neurosci 21:3646–3655
Cornette L, Dupont P, Salmon E, Orban GA (2001) The neu-ral substrate of orientation working memory. J Cogn Neurosci13:813–828
de Fockert JW, Rees G, Frith CD, Lavie N (2001) The role of work-ing memory in visual selective attention. Science 291:1803–1806
Diesmann M, Gewaltig MO, Aertsen A (1999) Stable propaga-tion of synchronous spiking in cortical neural networks. Nature402:529–533
Dudkin KN, Kruchinin VK, Chueva IV (1997) Effect of NMDA onthe activity of cortical glutaminergic structures in delayed visualdifferentiation in monkeys. Neurosci Behav Physiol 27:153–158
Durstewitz D, Seamans JK, Sejnowski TJ (2000) Dopamine-mediated stabilization of delay-period activity in a networkmodel of prefrontal cortex. J Neurophysiol 83:1733–1750
Egorov AV, Hamam BN, Fransen E, Hasselmo ME, Alonso AA(2002) Graded persistent activity in entorhinal cortex neurons.Nature 420:173–178
Fransen E, Lansner A (1998) A model of cortical associativememory based on a horizontal network of connected columns.Network 9:235–264
Funahashi S, Bruce CJ, Goldman-Rakic PS (1989) Mnemoniccoding of visual space in the monkey’s dorsolateral prefrontalcortex. J Neurophysiol 61:331–349
Fuster JM (1973) Unit activity in prefrontal cortex during delayed-response performance: neuronal correlates of transientmemory. J Neurophysiol 36:61–78
Fuster JM (1995) Memory in the cerebral cortex. MIT press,Cambrige
Fuster JM (1997) The prefrontal cortex: anatomy, physiology, andneuropsychology of the frontal lobe. Lippincott-Raven, NewYork

418 Biol Cybern (2007) 96:407–419
Gewaltig MO, Diesmann M, Aertsen A (2001) Propagation ofcortical syn�re activity: survival probability in single trials andstability in the mean. Neural Netw 14:657–673
Goldman MS, Levine JH, Major G, Tank DW, Seung HS(2003) Robust persistent neural activity in a model integratorwith multiple hysteretic dendrites per neuron. Cereb Cortex13:1185–1195
Goldman-Rakic PS (1987) Circuitry of primate prefrontal cor-tex and regulation of behavior by representational memory. In:Handbook of physiology. American Physiology Society, Beth-esda, pp 373–417
Goldman-Rakic PS (1995) Cellular basis of working memory.Neuron 14:477–485.
Gutkin BS, Laing CR, Colby CL, Chow CC, Ermentrout GB(2001) Turning on and off with excitation: the role of spike-timing asynchrony and synchrony in sustained neural activity.J Comput Neurosci 11:121–134
Haj-Dahmane S, Andrade R (1998) Ionic mechanism of the slowafterdepolarization induced by muscarinic receptor activationin rat prefrontal cortex. J Neurophysiol 80:1197–1210
Hebb DO (1949) The organization of behaviour. Wiley, New YorkHestrin S, Sah P, Nicoll RA (1990) Mechanisms generating the
time course of dual component excitatory synaptic currentsrecorded in hippocampal slices. Neuron 5:247–253
Hop�eld JJ (1982) Neural networks and physical systems withemergent collective computational abilities. Proc Natl AcadSci USA 79:2554–2558
Jahr CE, Stevens CF (1990) Voltage dependence of NMDA-activated macroscopic conductances predicted by single-chan-nel kinetics. J Neurosci 10:3178–3182
Kane MJ, Bleckley MK, Conway ARA, Engle RW (2001) Acontrolled-attention view of working-memory capacity. J ExpPsychol Gen 130:169–183
Klingberg T, Fernell E, Olesen PJ, Johnson M, Gustafsson P,Dahlstrom K, Gillberg CG, Forssberg H, Westerberg H (2005)Computerized training of working memory in children withADHD—a randomized, controlled trial. J Am Acad ChildAdolesc Psychiatry 44:177–186
Koulakov AA, Raghavachari S, Kepecs A, Lisman JE (2002)Model for a robust neural integrator. Nat Neurosci 5:775–782
Kritzer MF, Goldman-Rakic PS (1995) Intrinsic circuit organi-zation of the major layers and sublayers of the dorsolateralprefrontal cortex in the rhesus monkey. J Comp Neurol359:131–143
Lavie N, Hirst A, de Fockert JW, Viding E (2004) Load theoryof selective attention and cognitive control. J Exp Psychol Gen133:339–354
Lee RH, Heckman CJ (1998) Bistability in spinal motoneuronsin vivo: systematic variations in persistent inward currents. JNeurophysiol 80:583–593
Levitt JB, Lewis DA, Yoshioka T, Lund JS (1993) Topographyof pyramidal neuron intrinsic connections in macaque monkeyprefrontal cortex (areas 9 and 46). J Comp Neurol 338:360–376
Lisman JE, Fellous JM, Wang XJ (1998) A role for NMDA-receptor channels in working memory. Nat Neurosci 1:273–275
Loewenstein Y, Sompolinsky H (2003) Temporal integration bycalcium dynamics in a model neuron. Nat Neurosci 6:961–967
Macoveanu J, Klingberg T, Tegner J (2006) A biophysical modelof multiple-item working memory: a computational and neu-roimaging study. Neuroscience 141:1611–1618
Markram H, Lubke J, Frotscher M, Roth A, Sakmann B (1997)Physiology and anatomy of synaptic connections between thicktufted pyramidal neurones in the developing rat neocortex. JPhysiol 500(Pt 2):409–440
McCormick DA, Connors BW, Lighthall JW, Prince DA (1985)Comparative electrophysiology of pyramidal and sparsely
spiny stellate neurons of the neocortex. J Neurophysiol 54:782–806
Miller EK, Cohen JD (2001) An integrative theory of prefrontalcortex function. Annu Rev Neurosci 24:167–202
Miller EK, Erickson CA, Desimone R (1996) Neural mechanismsof visual working memory in prefrontal cortex of the macaque.J Neurosci 16:5154–5167
Mountcastle VB (1997) The columnar organization of the neocor-tex. Brain 120(Pt 4):701–722
Norman DA (1970) Models of human memory. Academic, NewYork
Olesen PJ, Westerberg H, Klingberg T (2004) Increased prefrontaland parietal activity after training of working memory. Nat Neu-rosci 7:75–79
Olesen PJ, Macoveanu J, Klingberg T (2005) Childhood develop-ment of brain activity related to working memory and distractorprocessing (unpublished)
Ploner CJ, Gaymard B, Rivaud S, Agid Y, Pierrot-Deseilligny C(1998) Temporal limits of spatial working memory in humans.Eur J Neurosci 10:794–797
Roberts RJ, Hager LD, Heron C (1994) Prefrontal cognitive pro-cesses working memory and inhibition in the antisaccade task.J Exp Psychol Gen 123:374–393
Sakai K, Rowe JB, Passingham RE (2002) Active maintenancein prefrontal area 46 creates distractor-resistant memory. NatNeurosci 5:479–484
Salin PA, Prince DA (1996) Electrophysiological mapping of GA-BAA receptor-mediated inhibition in adult rat somatosensorycortex. J Neurophysiol 75:1589–1600
Sandberg A, Tegnér J, Lansner A (2003) A working memorymodel based on fast Hebbian learning. Network 14:789–802
Scherzer CR, Landwehrmeyer GB, Kerner JA, Counihan TJ,Kosinski CM, Standaert DG, Daggett LP, Velicelebi G, Pen-ney JB, Young AB (1998) Expression of N-methyl-D-aspartatereceptor subunit mRNAs in the human brain: hippocampus andcortex. J Comp Neurol 390:75–90
Schiller J, Schiller Y (2001) NMDA receptor-mediated den-dritic spikes and coincident signal ampli�cation. Curr OpinNeurobiol 11:343–348
Seung HS (1996) How the brain keeps the eyes still. Proc NatlAcad Sci USA 93:13339–13344
Seung HS, Lee DD, Reis BY, Tank DW (2000) Stability of thememory of eye position in a recurrent network of conductance-based model neurons. Neuron 26:259–271
Shima K, Tanji J (1998) Involvement of NMDA and non-NMDAreceptors in the neuronal responses of the primary motor cortexto input from the supplementary motor area and somatosen-sory cortex: studies of task-performing monkeys. Jpn J Physiol48:275–290
Singer W (1993) Synchronization of cortical activity and its puta-tive role in information processing and learning. Annu RevPhysiol 55:349–374
Spruston N, Jonas P, Sakmann B (1995) Dendritic glutamate recep-tor channels in rat hippocampal CA3 and CA1 pyramidal neu-rons. J Physiol 482(Pt 2):325–352
Tegnér J, Compte A, Wang XJ (2002) The dynamical stability ofreverberatory neural circuits. Biol Cybern 87:471–481
Thomas HB (1968) An information-theoretic model for the serialposition effect. Psychol Rev 75:409–420
Todd JJ, Marois R (2004) Capacity limit of visual short-term mem-ory in human posterior parietal cortex. Nature 428:751–754
Treisman AM, Gelade G (1980) A feature-integration theory ofattention. Cognit Psychol 12:97–136
Vogel EK, Machizawa MG (2004) Neural activity predicts indi-vidual differences in visual working memory capacity. Nature428:748–751

Biol Cybern (2007) 96:407–419 419
Wang XJ (1999) Synaptic basis of cortical persistent activity: theimportance of NMDA receptors to working memory. J Neuro-sci 19:9587–9603
Wang XJ (2001) Synaptic reverberation underlying mnemonicpersistent activity. Trends Neurosci 24:455–463
Wang XJ, Tegnér J, Constantinidis C, Goldman-Rakic PS (2004)Division of labor among distinct subtypes of inhibitory neuronsin a cortical microcircuit of working memory. Proc Natl AcadSci USA 101:1368–1373
White JM, Sparks DL, Stanford TR (1994) Saccades to remem-bered target locations: an analysis of systematic and variableerrors. Vis Res 34:79–92
Xiang Z, Huguenard JR, Prince DA (2002) Synaptic inhibition ofpyramidal cells evoked by different interneuronal subtypes inlayer v of rat visual cortex. J Neurophysiol 88:740–750


















