
Neurogenic airway microvascular leakage induced by toluene inhalation in rats
6
Pulmonary, Gastrointestinal and Urogenital Pharmacology Neurogenic airway microvascular leakage induced by toluene inhalation in rats Tatsuo Sakamoto a, ⁎, Michihiro Kamijima b , Mio Miyake c a Department of Hygiene, Yamaguchi University Graduate School of Medicine, Kogushi 1-1-1, Ube 755-8505, Japan b Department of Occupational and Environmental Health, Nagoya City University Graduate School of Medicine, 1 Kawasumi, Mizuho-cho, Mizuho-ku, Nagoya 467-8601, Japan c Department of Pediatrics, Nagoya University Graduate School of Medicine, 65 Tsurumai-cho, Showa-ku, Nagoya 466-8550, Japan abstract article info Article history: Received 19 October 2010 Received in revised form 11 April 2012 Accepted 12 April 2012 Available online 24 April 2012 Keywords: Airway microvascular leakage CP-99,994 Mast cell degranulation Neutral endopeptidase Tachykinin NK 1 receptor Toluene is a representative airborne occupational and domestic pollutant that causes eye and respiratory tract irritation. We investigated whether a single inhalation of toluene elicits microvascular leakage in the rat airway. We also evaluated the effects of CP-99,994, a tachykinin NK 1 receptor antagonist, and ketotifen, a histamine H1 receptor antagonist with mast cell-stabilizing properties, on the airway response. The content of Evans blue dye that extravasated into the tissues was measured as an index of plasma leakage. Toluene (18–450 ppm, 10 min) concentration-dependently induced dye leakage into the trachea and main bronchi of anesthetized and mechanically ventilated rats. Toluene at concentrations of ≥50 and ≥30 ppm caused sig- nificant responses in the trachea and main bronchi, respectively, which both peaked after exposure to 135 ppm toluene for 10 min. This response was abolished by CP-99,994 (5 mg/kg i.v.), but not by ketotifen (1 mg/kg i.v.). Nebulized phosphoramidon (1 mM, 1 min), a neutral endopeptidase 24.11 inhibitor, signifi- cantly enhanced the response induced by toluene (135 ppm, 10 min) compared with nebulized 0.9% saline (1 min). These results show that toluene can rapidly increase airway plasma leakage that is predominantly mediated by tachykinins endogenously released from airway sensory nerves. However, mast cell activation might not be important in this airway response. © 2012 Elsevier B.V. All rights reserved. 1. Introduction Toluene is widely used in industry to produce benzene, trinitrotol- uene, nylon, plastics and polyurethanes, and it is found in gasoline, paint, ink, lacquer, paint thinner, adhesives, fingernail polish, cleaning agents and rubber (American Conference of Governmental Industrial Hygienists, ACGIH, 2010). Thus, occupational and domestic settings contain many possible emitters of toluene. Cigarette smoke and vehi- cle exhaust also contain some toluene. Concerns about health hazards from this pollutant have consequently increased (Saijo et al., 2004; Samet et al., 1988). A threshold time weighted average (TLV-TWA) of 20 ppm is recommended as the limit to protect employees in the workplace from subclinical changes in blue–yellow color vision and the potential for spontaneous abortion in female workers (ACGIH, 2010). Toluene causes eye and respiratory tract irritation. Toluene inhala- tion (100 ppm, 6 h) irritates the eyes and upper respiratory tracts of healthy volunteers, but does not cause cough or bronchoconstriction (Andersen et al., 1983). However, toluene (≥3300 ppm) decreases respiratory rates by at least 50% in mice (Schaper, 1993), indicating irritation of the lower respiratory tract. Fujimaki et al. (2007) showed that long-term (12 weeks) exposure to 50 ppm toluene decreases substance P levels in bronchoalveolar lavage fluid in mice and concur- rently increases neurotrophin-3. Tin-Tin-Win-Shwe et al. (2007) showed that seven inhalations of 9 ppm toluene for 30 min each en- hance plasma levels of nerve growth factor after provocation with ov- albumin in ovalbumin-sensitized mice. These findings suggest that toluene modulates the mechanisms associated with neurogenic air- way inflammation. However, whether toluene stimulates sensory nerves in the airway and subsequently leads to acute airway inflam- mation such as plasma leakage via tachykinin release has not been assessed. The present study investigates whether a single inhalation of tolu- ene elicits microvascular leakage and bronchoconstriction in rat air- way. To further investigate the role of tachykinin release induced by toluene, we evaluated effects of a tachykinin NK 1 receptor antagonist, CP-99,994 [(+)-(2s,3s)-3-(2-methoxybenzylamino)-2-phenylpiperi- dine] (McLean et al., 1993; Piedimonte et al., 1993), on airway micro- vascular leakage induced by toluene. Previous studies of isolated rat peritoneal mast cells (Devillier et al., 1989; Mazurek et al., 1981) have suggested that tachykinins such as substance P induce mast cell degranulation without binding to a specific tachykinin receptor. Rat lung mast cells release histamine in response to high doses of substance P in vitro (Ali et al., 1986). On the other hand, Pauwels et al. (1995) showed that tachykinin NK 1 receptor-mediated mast cell activation (namely, release of 5- hydroxytryptamine) is partly responsible for airway plasma leakage European Journal of Pharmacology 685 (2012) 180–185 ⁎ Corresponding author. Tel.: + 81 836 22 2229; fax: + 81 836 22 2345. E-mail address: [email protected] (T. Sakamoto). 0014-2999/$ – see front matter © 2012 Elsevier B.V. All rights reserved. doi:10.1016/j.ejphar.2012.04.035 Contents lists available at SciVerse ScienceDirect European Journal of Pharmacology journal homepage: www.elsevier.com/locate/ejphar
-
Upload
tatsuo-sakamoto -
Category
Documents
-
view
216 -
download
2
Transcript of Neurogenic airway microvascular leakage induced by toluene inhalation in rats

European Journal of Pharmacology 685 (2012) 180–185
Contents lists available at SciVerse ScienceDirect
European Journal of Pharmacology
j ourna l homepage: www.e lsev ie r .com/ locate /e jphar
Pulmonary, Gastrointestinal and Urogenital Pharmacology
Neurogenic airway microvascular leakage induced by toluene inhalation in rats
Tatsuo Sakamoto a,⁎, Michihiro Kamijima b, Mio Miyake c
a Department of Hygiene, Yamaguchi University Graduate School of Medicine, Kogushi 1-1-1, Ube 755-8505, Japanb Department of Occupational and Environmental Health, Nagoya City University Graduate School of Medicine, 1 Kawasumi, Mizuho-cho, Mizuho-ku, Nagoya 467-8601, Japanc Department of Pediatrics, Nagoya University Graduate School of Medicine, 65 Tsurumai-cho, Showa-ku, Nagoya 466-8550, Japan
⁎ Corresponding author. Tel.: +81 836 22 2229; fax:E-mail address: [email protected] (T. Sa
0014-2999/$ – see front matter © 2012 Elsevier B.V. Alldoi:10.1016/j.ejphar.2012.04.035
a b s t r a c t
a r t i c l e i n f oArticle history:Received 19 October 2010Received in revised form 11 April 2012Accepted 12 April 2012Available online 24 April 2012
Keywords:Airway microvascular leakageCP-99,994Mast cell degranulationNeutral endopeptidaseTachykinin NK1 receptor
Toluene is a representative airborne occupational and domestic pollutant that causes eye and respiratorytract irritation. We investigated whether a single inhalation of toluene elicits microvascular leakage in therat airway. We also evaluated the effects of CP-99,994, a tachykinin NK1 receptor antagonist, and ketotifen,a histamine H1 receptor antagonist with mast cell-stabilizing properties, on the airway response. The contentof Evans blue dye that extravasated into the tissues was measured as an index of plasma leakage. Toluene(18–450 ppm, 10 min) concentration-dependently induced dye leakage into the trachea and main bronchiof anesthetized and mechanically ventilated rats. Toluene at concentrations of ≥50 and≥30 ppm caused sig-nificant responses in the trachea and main bronchi, respectively, which both peaked after exposure to135 ppm toluene for 10 min. This response was abolished by CP-99,994 (5 mg/kg i.v.), but not by ketotifen(1 mg/kg i.v.). Nebulized phosphoramidon (1 mM, 1 min), a neutral endopeptidase 24.11 inhibitor, signifi-cantly enhanced the response induced by toluene (135 ppm, 10 min) compared with nebulized 0.9% saline(1 min). These results show that toluene can rapidly increase airway plasma leakage that is predominantlymediated by tachykinins endogenously released from airway sensory nerves. However, mast cell activationmight not be important in this airway response.
© 2012 Elsevier B.V. All rights reserved.
1. Introduction
Toluene is widely used in industry to produce benzene, trinitrotol-uene, nylon, plastics and polyurethanes, and it is found in gasoline,paint, ink, lacquer, paint thinner, adhesives, fingernail polish, cleaningagents and rubber (American Conference of Governmental IndustrialHygienists, ACGIH, 2010). Thus, occupational and domestic settingscontain many possible emitters of toluene. Cigarette smoke and vehi-cle exhaust also contain some toluene. Concerns about health hazardsfrom this pollutant have consequently increased (Saijo et al., 2004;Samet et al., 1988). A threshold time weighted average (TLV-TWA)of 20 ppm is recommended as the limit to protect employees in theworkplace from subclinical changes in blue–yellow color vision andthe potential for spontaneous abortion in female workers (ACGIH,2010).
Toluene causes eye and respiratory tract irritation. Toluene inhala-tion (100 ppm, 6 h) irritates the eyes and upper respiratory tracts ofhealthy volunteers, but does not cause cough or bronchoconstriction(Andersen et al., 1983). However, toluene (≥3300 ppm) decreasesrespiratory rates by at least 50% in mice (Schaper, 1993), indicatingirritation of the lower respiratory tract. Fujimaki et al. (2007) showed
+81 836 22 2345.kamoto).
rights reserved.
that long-term (12 weeks) exposure to 50 ppm toluene decreasessubstance P levels in bronchoalveolar lavage fluid in mice and concur-rently increases neurotrophin-3. Tin-Tin-Win-Shwe et al. (2007)showed that seven inhalations of 9 ppm toluene for 30 min each en-hance plasma levels of nerve growth factor after provocation with ov-albumin in ovalbumin-sensitized mice. These findings suggest thattoluene modulates the mechanisms associated with neurogenic air-way inflammation. However, whether toluene stimulates sensorynerves in the airway and subsequently leads to acute airway inflam-mation such as plasma leakage via tachykinin release has not beenassessed.
The present study investigates whether a single inhalation of tolu-ene elicits microvascular leakage and bronchoconstriction in rat air-way. To further investigate the role of tachykinin release induced bytoluene, we evaluated effects of a tachykinin NK1 receptor antagonist,CP-99,994 [(+)-(2s,3s)-3-(2-methoxybenzylamino)-2-phenylpiperi-dine] (McLean et al., 1993; Piedimonte et al., 1993), on airway micro-vascular leakage induced by toluene.
Previous studies of isolated rat peritoneal mast cells (Devillier etal., 1989; Mazurek et al., 1981) have suggested that tachykininssuch as substance P induce mast cell degranulation without bindingto a specific tachykinin receptor. Rat lung mast cells release histaminein response to high doses of substance P in vitro (Ali et al., 1986). Onthe other hand, Pauwels et al. (1995) showed that tachykinin NK1
receptor-mediated mast cell activation (namely, release of 5-hydroxytryptamine) is partly responsible for airway plasma leakage

181T. Sakamoto et al. / European Journal of Pharmacology 685 (2012) 180–185
induced by substance P and capsaicin in F344-, but not in BDE-rats.Therefore, if toluene stimulates sensory nerves in the airway, releasedtachykinins can potentially cause mast cell degranulation. In addition,toluene might directly activate mast cells. We therefore investigatedthe role of mast cells in plasma leakage induced by toluene in the air-ways of rats using ketotifen (4-(1-methyl-4-piperidylidene)4H-benzo[4,5] cycloheptal[1,2-b]thiophen-10(9H)-one hydrogen fuma-rate), a histamine H1 receptor antagonist with mast cell-stabilizingproperties (Martin and Römer, 1978).
2. Materials and methods
2.1. Animals
Experiments proceeded with the approval of the Animal EthicsCommittee of the Institute for Laboratory Animal Research, NagoyaUniversity Graduate School of Medicine. This committee operates inaccordance with the Guidelines for the Care and Use of LaboratoryAnimals of Nagoya University (2007; Nagoya, Japan). Five days beforeexperimentation, specific pathogen-free male Wistar rats (8 weeks ofage, 260–280 g; Japan SLC Inc., Hamamatsu, Japan) were housed in atemperature-controlled room and given free access to food andwater.
2.2. Reagents
CP-99,994 and ketotifen were donated by Pfizer (Groton, CT) andNovartis Pharma (Tokyo, Japan), respectively. Other materials and theirrespective sources included: ethanol, Evans blue dye, 37% (w/v) formalinand toluene from Sigma Aldrich Japan (Tokyo, Japan); thiamylal sodiumfrom Kyorin Pharmaceutical (Tokyo, Japan); 0.9% saline from OtsukaChemical (Tokyo, Japan); and phosphoramidon, a neutral endopeptidase24.11 inhibitor, from Peptide Institute (Osaka, Japan). CP-99,994, ketoti-fen, Evans blue dye and phosphoramidon were dissolved in 0.9% saline.
2.3. Measurement of pulmonary insufflation pressure
Rats were given thiamylal (100 mg/kg i.p.) and placed on a homeo-thermic blanket (KN-474, Natume, Tokyo, Japan) to maintain rectal tem-perature at about 37 °C. A tracheal cannula (length, 8 mm; innerdiameter, 1.3 mm) was inserted into the lumen of the cervical tracheathrough a tracheostomy and snugly sutured. A polyethylene catheterwas inserted into the left carotid artery tomonitormean systemic arterialblood pressure with a pressure transducer (TP-400T; Nihon Koden,Tokyo, Japan). The right external jugular vein was cannulated for intrave-nous administration. The animals were connected to a ventilator (Model131, New England Medical Instruments, Medway, MA) set at a tidal vol-ume of 8 mL/kg and a frequency of 90 strokes/min. Pulmonary insuffla-tion pressure (PIP) was measured using a pressure transducer (TP-400T) connected to a side arm of the expiratory limb of the ventilatingtubing. Signals of PIP and mean systemic arterial blood pressure fromthe transducers were amplified (AP-601G, Nihon Koden) and recordedwith a two-channel recorder (RTA-1100, Nihon Koden).
2.4. Measurement of airway microvascular leakage
Evans blue dye (0.5 mL/kg; 40 mg/ml) was injected via the rightexternal jugular vein before provocation and then tissue sampleswere collected as follows. The thoracic cavity was opened after prov-ocation and then a cannula was inserted into the aorta through theleft ventricle and the heart was perfused with 40 mL of 0.9% salineat a pressure of 120 mm Hg. Blood and perfusates were expelledthrough an incision in the right atrium. The pulmonary circulationwas also perfused with 20 mL of 0.9% saline at the same pressure.The lungs were removed and then the connective tissues and paren-chyma were gently scraped off. The lower trachea (length, 6 mm) and
main bronchi were dissected, blotted dry and weighed. Evans bluedye that leaked into the tissues was measured as described (Ito etal., 1996) and is expressed as ng/mg of wet tissue.
2.5. Toluene and formaldehyde gas supply systems
Constant concentrations of gaseous toluene from liquid toluenewere generated using a dynamic standard gas generator (PermeaterModel PD-1B, Gastec, Ayase, Japan). Neat toluene (3 mL) was placedin D-10, D-20 and D-30 diffusion tubes (Gastec) to yield 18, 30 and50 ppm toluene, respectively. A D-30 (1/2 tube length) diffusiontube was used to produce 135 and 450 ppm toluene. Each tube wasthen fixed in a dilution–permeation chamber under a constant tem-perature (50 °C). A steady flow of air, passed through the chamber,was adjusted to 1 L/min for 18, 30, 50, and 135 ppm of toluene, andto 0.3 L/min for 450 ppm. Each experimental condition using the tol-uene supply system was determined by six measurements of the lossof liquid toluene (by weight) remaining in the diffusion tube and thetotal flow volume for 24 h. The difference among repeated measure-ments of the weight loss for 24 h was within±3.5% of the theoretical-ly targeted value. Formaldehyde gas (45 ppm) was generated usingthe Permeater Model PD-1B (Gastec) as described (Ito et al., 1996).Room air passed through the chamber (1 L/min, 50 °C) served asclean air. Each gas was delivered to the ventilatory circuit throughthe air inlet of the ventilator. The temperature of the gases at theport of the tracheal cannula was almost 20 °C, which was similar tothat of room air.
2.6. Protocol
2.6.1. Effects of different concentrations of gaseous tolueneRats were assigned to five groups (n=5 per group) to investigate the
concentration-dependent effects of the inhalation of 18, 30, 50, 135 or450 ppm of toluene on airway microvascular leakage and bronchocon-striction (Fig. 1A). Another two groups of rats (n=5 per group) inhaledformaldehyde (45 ppm) or clean air (Fig. 1A).We alsomonitored PIP andmean systemic arterial blood pressure throughout the experiments.Baseline PIP andmean systemic arterial blood pressurewere determinedimmediately before exposure to each gas tested. Our previous study (Itoet al., 1996) showed that formaldehyde (5–45 ppm) concentration-dependently induces airway microvascular leakage in rats, a responsethat reaches a peak at 45 ppm. In addition, the response induced by45 ppm formaldehyde was almost equivalent to that induced by an ex-cess (100 mg/kg) dose of intramuscular capsaicin (Kimura et al., 2010),indicating that 45 ppm formaldehyde caused maximal levels of airwaymicrovascular leakage induced by tachykinins released from sensorynerves.
2.6.2. Airway microvascular leakage during a 30-min period of tolueneinhalation
Airway microvascular leakage was evaluated in six groups of rats(n=5per group) during three consecutive 10-min periods of toluene in-halation (50 or 135 ppm; Fig. 1B). Rats (n=5) were sham-stimulatedwith clean air for 10 min.
2.6.3. Effects of CP-99,994 and ketotifen on toluene-induced airway mi-crovascular leakage
Rats were assigned to five groups (n=5 per group) to assess theeffects of CP-99,994, ketotifen or vehicle (0.9% saline) (Fig. 1C). Weapplied CP-99,994 (0.8, 2 and 5 mg/kg i.v.) according to a previousstudy showing that CP-99,994 at doses of >3 mg/kg i.v. abolish theairway microvascular leakage induced by inhaled formaldehyde(15 ppm, 10 min) (Ito et al., 1996). The dose of ketotifen (1 mg/kgi.v.) was determined on the basis of a preliminary study (Ito et al.,1996). In brief, the dose of ketotifen obviously inhibited the responsein the trachea (98.8±8.5 ng/mg, n=5, to 64.9±6.8 ng/mg, n=4;

Fig. 1. Experimental protocols for evaluating airway microvascular leakage induced by inhaled toluene in rats. (A) Concentration-dependent effects of inhaled toluene on airwaymicrovascular leakage in rats. (B) Time-course changes of airway microvascular leakage during 30-min of toluene inhalation in rats. (C) Effects of tachykinin NK1 receptor antag-onist CP-99,994 and of histamine H1 receptor antagonist ketotifen with mast cell-stabilizing properties, on microvascular leakage induced by inhaled toluene in rats.
182 T. Sakamoto et al. / European Journal of Pharmacology 685 (2012) 180–185
Pb0.05, unpaired Student's t test) and main bronchi (92.2±11.9 ng/mg,n=5, to 38.3±3.1 ng/mg, n=4; Pb0.05, Welch's test) induced by aero-solized ovalbumin in passively ovalbumin-sensitized rats.
2.6.4. Effects of phosphoramidon on toluene-induced airway microvascu-lar leakage
We investigated the effects of inhibiting neutral endopeptidase ac-tivity on the airway microvascular leakage induced by inhaled toluene.Four groups of rats (n=5 per group) were anesthetized, connected tothe ventilator and then administered with phosphoramidon (1 mM)or 0.9% saline for 1 min generated from an ultrasonic nebulizer(Model 5000; Devilbiss, Somerset, PA). The respective aerosolswere de-livered through a separate system that bypassed the inspiratory limb ofthe ventilation tubing. Evans blue dyewas administered 1 min later, fol-lowed by a 10-min inhalation of 135 ppm toluene or clean air. Tissuesamples were then collected to measure dye leakage. This dose of neb-ulized phosphoramidon maximally enhances the bronchoconstrictioninduced by 5 Hz of vagal nerve stimulation in guinea pigs (Lötvall etal., 1991). A lower dose of phosphoramidon (0.1 mM for 1 min) also in-creases pulmonary resistance induced by inhaled capsaicin or substanceP in guinea pigs (Dusser et al., 1989).
2.7. Statistical analysis
All values are expressed as means±S.E.M. We used an unpaired Stu-dent's t test (two-tailed) to evaluate the significance of the difference inplasma leakage in each airway level between two independent groupswith equal variance when assessed with the F test. We otherwise usedWelch's test (two-tailed). One-way analysis of variance (ANOVA)was ap-plied to assess the concentration-dependent effect of toluene on airwayplasma leakage and the treatment effect of CP-99,994 on the airway re-sponse. When appropriate, Dunnett's test was conducted for multiplecomparisons with one control. The significance of time and treatment ef-fects on PIP andmean systemic arterial blood pressurewas assessed usinga two-way ANOVA with Bonferroni correction for multiple comparisons.The effect of phosphoramidon on airway microvascular leakage induced
by inhalation of toluene and control air was analyzed with this statisticalmethod. A level of Pb0.05 was considered statistically significant.
3. Results
3.1. Effects of different concentrations of inhaled toluene
Baseline PIP and mean systemic arterial blood pressure did notsignificantly differ in rats that inhaled toluene, formaldehyde orclean air (Table 1). Neither PIP nor mean systemic arterial blood pres-sure was altered by inhaling toluene (18–450 ppm) or formaldehyde(45 ppm) for 10 min or during a subsequent 10-min observation(Table 1). Toluene concentration-dependently caused significant dyeleakage into the trachea and main bronchi (Fig. 2). The threshold con-centrations of toluene for this response were 50 and 30 ppm in thetrachea and main bronchi, respectively, and 135 ppm of toluene elic-ited the maximal response in both sites. However, the peak responsewas significantly smaller than that elicited by 45 ppm of formalde-hyde (Pb0.01, unpaired Student's t test). Dye did not leak into the pa-renchyma macroscopically after 450 ppm of toluene was inhaled for10 min.
3.2. Airway microvascular leakage during toluene inhalation for 30 min
The amount of dye that leaked into the trachea and main bronchiwas almost maximal during the first 10 min of toluene inhalation (50and 135 ppm, Fig. 3). The response in the trachea persisted somewhatduring a second 10-min inhalation of 135 ppm of toluene.
3.3. Effects of CP-99,994 and ketotifen on toluene-induced airwaymicrovascular leakage
Baseline PIP and mean systemic arterial blood pressure, deter-mined immediately before exposure to toluene, did not significantlydiffer among the five treated groups given CP-99,994 (0.8, 2, or5 mg/kg i.v.), ketotifen (1 mg/kg i.v.) or 0.9% saline (0.5 ml/kg i.v.)(data not shown). The increased dye leakage into the trachea and
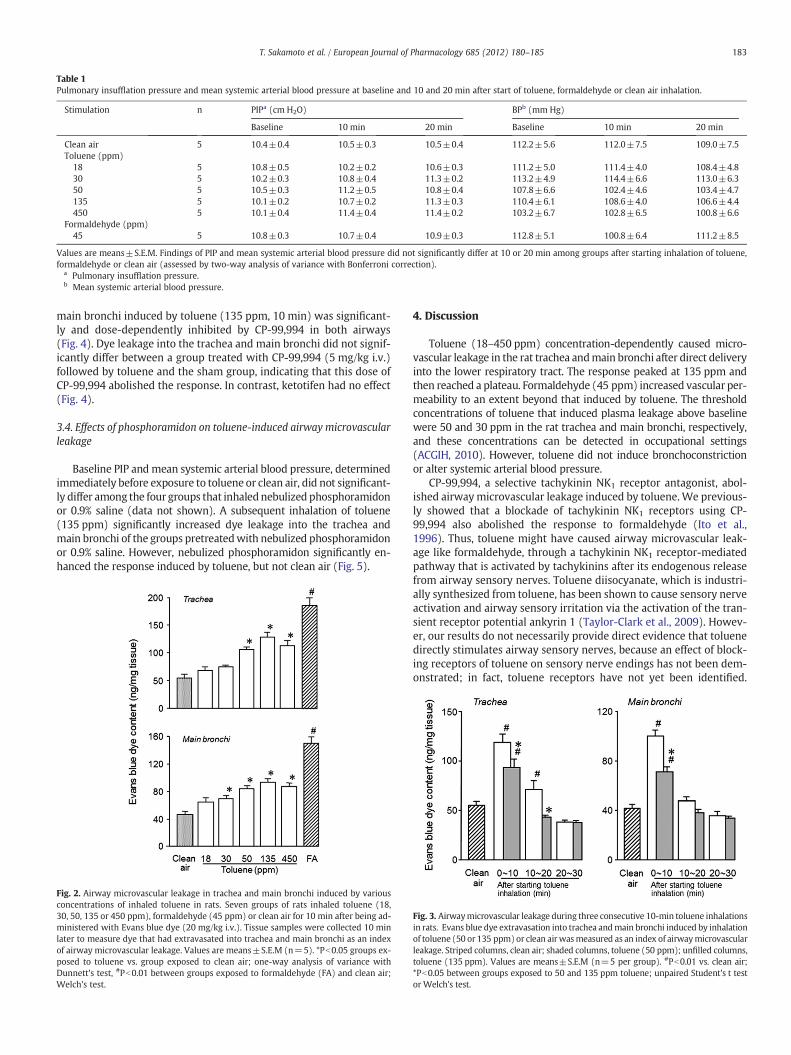
Table 1Pulmonary insufflation pressure and mean systemic arterial blood pressure at baseline and 10 and 20 min after start of toluene, formaldehyde or clean air inhalation.
Stimulation n PIPa (cm H2O) BPb (mm Hg)
Baseline 10 min 20 min Baseline 10 min 20 min
Clean air 5 10.4±0.4 10.5±0.3 10.5±0.4 112.2±5.6 112.0±7.5 109.0±7.5Toluene (ppm)
18 5 10.8±0.5 10.2±0.2 10.6±0.3 111.2±5.0 111.4±4.0 108.4±4.830 5 10.2±0.3 10.8±0.4 11.3±0.2 113.2±4.9 114.4±6.6 113.0±6.350 5 10.5±0.3 11.2±0.5 10.8±0.4 107.8±6.6 102.4±4.6 103.4±4.7135 5 10.1±0.2 10.7±0.2 11.3±0.3 110.4±6.1 108.6±4.0 106.6±4.4450 5 10.1±0.4 11.4±0.4 11.4±0.2 103.2±6.7 102.8±6.5 100.8±6.6
Formaldehyde (ppm)45 5 10.8±0.3 10.7±0.4 10.9±0.3 112.8±5.1 100.8±6.4 111.2±8.5
Values are means±S.E.M. Findings of PIP and mean systemic arterial blood pressure did not significantly differ at 10 or 20 min among groups after starting inhalation of toluene,formaldehyde or clean air (assessed by two-way analysis of variance with Bonferroni correction).
a Pulmonary insufflation pressure.b Mean systemic arterial blood pressure.
183T. Sakamoto et al. / European Journal of Pharmacology 685 (2012) 180–185
main bronchi induced by toluene (135 ppm, 10 min) was significant-ly and dose-dependently inhibited by CP-99,994 in both airways(Fig. 4). Dye leakage into the trachea and main bronchi did not signif-icantly differ between a group treated with CP-99,994 (5 mg/kg i.v.)followed by toluene and the sham group, indicating that this dose ofCP-99,994 abolished the response. In contrast, ketotifen had no effect(Fig. 4).
3.4. Effects of phosphoramidon on toluene-induced airway microvascularleakage
Baseline PIP and mean systemic arterial blood pressure, determinedimmediately before exposure to toluene or clean air, did not significant-ly differ among the four groups that inhalednebulized phosphoramidonor 0.9% saline (data not shown). A subsequent inhalation of toluene(135 ppm) significantly increased dye leakage into the trachea andmain bronchi of the groups pretreatedwith nebulized phosphoramidonor 0.9% saline. However, nebulized phosphoramidon significantly en-hanced the response induced by toluene, but not clean air (Fig. 5).
Fig. 2. Airway microvascular leakage in trachea and main bronchi induced by variousconcentrations of inhaled toluene in rats. Seven groups of rats inhaled toluene (18,30, 50, 135 or 450 ppm), formaldehyde (45 ppm) or clean air for 10 min after being ad-ministered with Evans blue dye (20 mg/kg i.v.). Tissue samples were collected 10 minlater to measure dye that had extravasated into trachea and main bronchi as an indexof airway microvascular leakage. Values are means±S.E.M (n=5). *Pb0.05 groups ex-posed to toluene vs. group exposed to clean air; one-way analysis of variance withDunnett's test, #Pb0.01 between groups exposed to formaldehyde (FA) and clean air;Welch's test.
4. Discussion
Toluene (18–450 ppm) concentration-dependently caused micro-vascular leakage in the rat trachea andmain bronchi after direct deliveryinto the lower respiratory tract. The response peaked at 135 ppm andthen reached a plateau. Formaldehyde (45 ppm) increased vascular per-meability to an extent beyond that induced by toluene. The thresholdconcentrations of toluene that induced plasma leakage above baselinewere 50 and 30 ppm in the rat trachea and main bronchi, respectively,and these concentrations can be detected in occupational settings(ACGIH, 2010). However, toluene did not induce bronchoconstrictionor alter systemic arterial blood pressure.
CP-99,994, a selective tachykinin NK1 receptor antagonist, abol-ished airwaymicrovascular leakage induced by toluene. We previous-ly showed that a blockade of tachykinin NK1 receptors using CP-99,994 also abolished the response to formaldehyde (Ito et al.,1996). Thus, toluene might have caused airway microvascular leak-age like formaldehyde, through a tachykinin NK1 receptor-mediatedpathway that is activated by tachykinins after its endogenous releasefrom airway sensory nerves. Toluene diisocyanate, which is industri-ally synthesized from toluene, has been shown to cause sensory nerveactivation and airway sensory irritation via the activation of the tran-sient receptor potential ankyrin 1 (Taylor-Clark et al., 2009). Howev-er, our results do not necessarily provide direct evidence that toluenedirectly stimulates airway sensory nerves, because an effect of block-ing receptors of toluene on sensory nerve endings has not been dem-onstrated; in fact, toluene receptors have not yet been identified.
Fig. 3. Airwaymicrovascular leakage during three consecutive 10-min toluene inhalationsin rats. Evans blue dye extravasation into trachea andmain bronchi induced by inhalationof toluene (50 or 135 ppm) or clean air wasmeasured as an index of airwaymicrovascularleakage. Striped columns, clean air; shaded columns, toluene (50 ppm); unfilled columns,toluene (135 ppm). Values are means±S.E.M (n=5 per group). #Pb0.01 vs. clean air;*Pb0.05 between groups exposed to 50 and 135 ppm toluene; unpaired Student's t testor Welch's test.

Fig. 4. Effects of CP-99,994 (tachykinin NK1 receptor antagonist) and ketotifen (hista-mine H1 receptor antagonist with mast cell-stabilizing properties) on microvascularleakage in trachea and main bronchi induced by inhaled toluene in rats. Rats were ad-ministered with CP-99,994 (0.8, 2 or 5 mg/kg i.v.), ketotifen (1 mg/kg i.v.) or vehicle(0.9% saline, 0.5 ml/kg i.v.) and then inhaled toluene (135 ppm) for 10 min. Tissuesamples were then collected to measure amounts of dye leakage into trachea andmain bronchi as indices of airway microvascular leakage. Sham group received 0.9% sa-line (0.5 ml/kg i.v.) and then inhaled clean air for 10 min. Values are means±S.E.M(n=5 per group). *Pb0.05 CP-99,994 followed by toluene vs. 0.9% saline (Con) fol-lowed by toluene; one-way analysis of variance with Dunnett's test. #Pb0.01 vs.sham group; unpaired Student's t test or Welch's test.
184 T. Sakamoto et al. / European Journal of Pharmacology 685 (2012) 180–185
Bradykinin that is produced in the airway via the conversion ofkininogens in plasma exudate (Proud and Kaplan, 1988) mightcause tachykinin release from sensory nerves via the activation ofbradykinin B2 receptors (Groneberg et al., 2004), leading to airwayplasma leakage. However, such a role of bradykinin has not been de-termined in microvascular leakage induced by formaldehyde in ratairways (Ito et al., 1996).
Formaldehyde at 45 ppm causes maximal airway microvascularleakage mediated by tachykinins released from sensory nerves(Kimura et al., 2010). We found here that the maximal responsewas much greater than that elicited by toluene (135 ppm), indicatingthat even toluene at the maximal tested concentration did notcompletely deplete tachykinins from sensory nerve endings in theairway. The reason for the different maximal responses induced by
Fig. 5. Effects of phosphoramidon (neutral endopeptidase 24.11 inhibitor) on subse-quent microvascular leakage in rat airways induced by toluene inhalation. Rats inhalednebulized phosphoramidon (1 mM) or 0.9% saline for 1 min. Evans blue dye was theninjected i.v. and then rats inhaled toluene (135 ppm) or clean air for 10 min. Tissuesamples were obtained to measure the amounts of dye that leaked into trachea andmain bronchi as indices of airway microvascular leakage. Values are mean±S.E.M(n=5 per group). #Pb0.01; inhaled toluene vs. clean air, *Pb0.01; nebulized phos-phoramidon vs. 0.9% saline; two-way ANOVA with Bonferroni correction.
these chemicals is unclear. Formaldehyde might be more easily dif-fused and delivered to sensory nerve endings in the airway than tol-uene, since formaldehyde is far more soluble than toluene.
Tachyphylactic responses in airway microvascular leakage inducedby airway sensory nerve stimulation may involve decreases in tachyki-nins within sensory nerve endings, impaired tachykinin release fromsensory nerves that leads to decreased amounts of tachykinins releasedfrom sensory nerves and desensitization of tachykinin NK1 receptors onendothelial cells in post-capillary venules (Bowden et al., 1994;Groneberg et al., 2004). We showed that toluene rapidly induced neu-rogenic airway microvascular leakage, as the response almost ceasedwithin 10 min despite constant inhalation. The tachyphylactic responseinduced by toluene is unlikely due to complete depletion of tachykininsin sensory nerve endings as discussed above. The response induced byformaldehyde at submaximal concentrations also ceased in the tracheaandmain bronchiwithin 15 min after the start of inhalation, despite theconstant exposure to formaldehyde (Kimura et al., 2010). This tachy-phylactic response is thought to be due to a decrease in the amount oftachykinins released from sensory nerves at exposed sites induced byinitially administered formaldehyde (Kimura et al., 2010). Thus, initiallyadministered toluene similarly appears to decrease the amount oftachykinins released from sensory nerves or cause desensitization oftachykinin NK1 receptors on endothelial cells in post-capillary venulesat exposed sites. However, further studies should determine the mech-anisms underlying the tachyphylactic response elicited during tolueneinhalation.
Ketotifen did not affect airwaymicrovascular leakage induced by tol-uene, whereas the dose of ketotifen applied herein obviously inhibitedallergen-induced airway microvascular leakage in rat airway (seeSection 2.6.3). Ketotifen has both the antagonism of histamine H1 recep-tors and mast cell-stabilizing properties (Martin and Römer, 1978).Therefore, mediators derived from mast cells including histamine areprobably not involved in this response, although the effects of higherdoses of ketotifen on this response were not evaluated, since weak ago-nistic activity, namely hypotension, occurs at i.v. doses of >1 mg/kg.That is to say, our data provided no evidence that toluene-induced air-way plasma leakage involves the degranulation of mast cells via aneurogenic-mediated mechanism, tachykinin release via a mast cell-mediated pathway or direct stimulation of toluene to mast cells. The in-volvement of mast cell degranulation in neurogenic plasma leakage inthe respiratory tract remains controversial (Ito et al., 1996; Kowalski etal., 1997; Lilly et al., 1995). Pauwels et al. (1995) showed strain-relateddifferences in a rat model. However, our data are consistent with thefinding that it has no effect on the response induced by formaldehyde(Ito et al., 1996). In addition, we previously showed that topical toluene,as well as formalin and capsaicin, causes plasma leakage in rat skin viathe stimulation of tachykinin NK1 receptors but not through a mastcell-mediated pathway (Futamura et al., 2009).
Exogenous tachykinins produce bronchoconstriction (Joos et al.,1994) and hypotension in the rat (Lembeck et al., 1992; Maggi et al.,1987). However, the present study showed that tachykinin release in-duced by toluene in the airway did not result in these effects, whichwas consistent with our previous findings on the effects of formalde-hyde on the airway (Ito et al., 1996). The reason for this discrepancy isnot clear. The site of tachykinin release might be distant from airwayand arterial smooth muscles, because toluene probably induces the re-lease of tachykinins from sensory nerve endingswithin and beneath theepithelium, where most sensory nerves containing tachykinin in theairway are located (Baluk et al., 1992). Released tachykinins might dif-fuse and be actively degraded by several peptidases, including neutralendopeptidase and angiotensin-converting enzyme (Dusser et al.,1989; Lötvall et al., 1991), before reaching their sites of action.We dem-onstrated neutral endopeptidase activity in the rat airway using theneutral endopeptidase inhibitor phosphoramidon, which enhanced air-way microvascular leakage induced by toluene. However, phosphora-midon did not alter PIP or mean systemic arterial blood pressure

185T. Sakamoto et al. / European Journal of Pharmacology 685 (2012) 180–185
during and 10 min after toluene inhalation. Li et al. (2005) showed thatendogenously-released nitric oxide inhibits capsaicin-induced bronch-oconstriction but not plasma extravasation in rat airways. Such a senso-ry nerve-mediated inhibitory response that mediates relaxation in theairway could be an alternative explanation for the failure of toluene toevokebronchoconstriction. However, further studies are required tode-termine the underlying mechanisms that account for such findings.
In conclusion, toluene rapidly induces airway microvascular leak-age in rats through a tachykinin NK1 receptor-mediated pathwaystimulated by tachykinins released from sensory nerve endings. How-ever, this response is probably not large enough to cause narrowing ofthe airway lumen (Chung et al., 1990). Our experimental data suggestthat toluene inhalation causes and exaggerates neurogenic inflamma-tion in human airways, particularly when they are already inflamedand when neutral endopeptidase activity is impaired (Di Maria etal., 1998).
Acknowledgements
This studywas funded in part by theNational Institute ofOccupationalSafety and Health, Japan [KIBAN34284].
References
Ali, H., Leung, K.B.P., Pearce, F.L., Hayes, N.A., Foreman, J.C., 1986. Comparison of thehistamine-releasing action of substance-P on mast cells and basophils from differ-ent species and tissues. Int. Arch. Allergy Appl. Immunol. 79, 413–418.
American Conference of Governmental Industrial Hygienists (ACGIH), 2010. 2010 TLVsand BEIs. ACGIH Publication, Cincinnati, Ohio.
Andersen, I., Lundqvist, G.R., Mølhave, L., Pedersen, O.F., Proctor, D.F., Vaeth, M., Wyon,D.P., 1983. Human response to controlled levels of toluene in six-hour exposures.Scand. J. Work Environ. Health 9, 405–418.
Baluk, P., Nadel, J.A., McDonald, D.M., 1992. Substance P-immunoreactive sensoryaxons in the rat respiratory tract: a quantitative study of their distribution androle in neurogenic inflammation. J. Comp. Neurol. 319, 586–598.
Bowden, J.J., Garland, A.M., Baluk, P., Lefevre, P., Grady, E.F., Vigna, S.R., Bunnett, N.W.,McDonald, D.M., 1994. Direct observation of substance P-induced internalizationof neurokinin-1 (NK1) receptors at sites of inflammation. Proc. Natl. Acad. Sci. 91,8964–8968.
Chung, K.F., Rogers, D.F., Barnes, P.J., Evans, T.W., 1990. The role of increased airwaymi-crovascular permeability and plasma exudation in asthma. Eur. Respir. J. 3,329–337.
Devillier, P., Drapeau, G., Renoux, M., Regoli, D., 1989. Role of the N-terminal arginine inthe histamine-releasing activity of substance-P, bradykinin and related peptides.Eur. J. Pharmacol. 168, 53–60.
Di Maria, G.U., Bellofiore, S., Geppetti, P., 1998. Regulation of airway neurogenic inflam-mation by neutral endopeptidase. Eur. Respir. J. 12, 1454–1462.
Dusser, D.J., Jacoby, D.B., Djokic, T.D., Rubinstein, I., Borson, D.B., Nadel, J.A., 1989. Virusinduces airway hyperresponsiveness to tachykinins: role of neutral endopeptidase.J. Appl. Physiol. 67, 1504–1511.
Fujimaki, H., Yamamoto, S., Tin-Tin-Win-Shwe, Hojo, R., Sato, F., Kunugita, N.,Arashidani, K., 2007. Effect of long-term exposure to low-level toluene on airwayinflammatory response in mice. Toxicol. Lett. 30, 132–139.
Futamura, M., Goto, S., Kimura, R., Kimoto, I., Miyake, M., Ito, K., Sakamoto, T., 2009. Dif-ferential effects of topically applied formalin and aromatic compounds onneurogenic-mediated microvascular leakage in rat skin. Toxicology 255, 100–106.
Groneberg, D.A., Quarcoo, D., Frossard, N., Fischer, A., 2004. Neurogenic mechanisms inbronchial inflammatory diseases. Allergy 59, 1139–1152.
Ito, K., Sakamoto, T., Hayashi, Y., Morishita, M., Shibata, E., Sakai, K., Takeuchi, Y., Torii,S., 1996. Role of tachykinin and bradykinin receptors and mast cells in gaseousformaldehyde-induced airway microvascular leakage in rats. Eur. J. Pharmacol.307, 291–298.
Joos, G.F., Kips, J.C., Pauwels, R.A., 1994. In vivo characterization of the tachykinin re-ceptors involved in the direct and indirect bronchoconstrictor effect of tachykininsin two inbred rat strains. Am. J. Respir. Crit. Care Med. 149, 1160–1166.
Kimura, R., Kimoto, I., Takeda, M., Miyake, M., Sakamoto, T., 2010. Alteration in airwaymicrovascular leakage induced by sensorineural stimulation in rats exposed to in-haled formaldehyde. Toxicol. Lett. 199, 254–260.
Kowalski, M.L., Didier, A., Lundgren, J.D., Igarashi, Y., Kaliner, M.A., 1997. Role of senso-ry innervation and mast cells in neurogenic plasma protein exudation into the air-way lumen. Respirology 2, 267–274.
Lembeck, F., Donnerer, J., Tsuchiya, M., Nagahisa, A., 1992. The non-peptide tachykininantagonist, CP-96,345, is a potent inhibitor of neurogenic inflammation. Br. J. Phar-macol. 105, 527–530.
Li, P.-C., Shaw, C.F., Kuo, T.F., Chien, C.T., 2005. Inducible nitric oxide synthase evokednitric oxide counteracts capsaicin-induced airway smooth muscle contraction,but exacerbates plasma extravasation. Neurosci. Lett. 378, 117–122.
Lilly, C.M., Hall, A.E., Rodger, I.W., Kobzik, L., Haley, K.J., Drazen, J.M., 1995. Substance P-induced histamine release in tracheally perfused guinea pig lungs. J. Appl. Physiol.78, 1234–1241.
Lötvall, J.O., Tokuyama, K., Löfdahl, C.G., Ullman, A., Barnes, P.J., Chung, K.F., 1991. Pep-tidase modulation of noncholinergic vagal bronchoconstriction and airway micro-vascular leakage. J. Appl. Physiol. 70, 2730–2735.
Maggi, C.A., Giuliani, S., Santicioli, P., Regoli, D., Meli, A., 1987. Peripheral effects of neu-rokinins: functional evidence for the existence of multiple receptors. J. Auton.Pharmacol. 7, 11–32.
Martin, U., Römer, D., 1978. Pharmacological properties of a new, orally active antiana-phylactic compound: ketotifen, a benzocycloheptathiophene. Arzneimittel-forschung 28, 770–782.
Mazurek, N., Pecht, I., Teichberg, V.I., Blumberg, S., 1981. The role of the N-terminal tet-rapeptide in the histamine releasing action of substance-P. Neuropharmacology20, 1025–1027.
McLean, S., Ganong, A., Seymour, P.A., Snider, R.M., Desai, M.C., Rosen, T., Bryce, D.K.,Longo, K.P., Reynolds, L.S., Robinson, G., Schmidt, A.W., Siok, C., Heym, J., 1993.Pharmacology of CP-99,994; a nonpeptide antagonist of the tachykininneurokinin-1 receptor. J. Pharmacol. Exp. Ther. 267, 472–479.
Pauwels, R.A., Germonpré, P.R., Kips, J.C., Joos, G.F., 1995. Genetic control of indirect air-way responsiveness in the rat. Clin. Exp. Allergy 25, 55–60.
Piedimonte, G., Bertrand, C., Geppetti, P., Snider, R.M., Desai, M.C., Nadel, J.A., 1993. Anew NK1 receptor antagonist (CP-99,994) prevents the increase in tracheal vascu-lar permeability produced by hypertonic saline. J. Pharmacol. Exp. Ther. 266,270–273.
Proud, D., Kaplan, A.P., 1988. Kinin formation: mechanisms and role in inflammatorydisorders. Annu. Rev. Immunol. 6, 49–83.
Saijo, Y., Kishi, R., Sata, F., Katakura, Y., Urashima, Y., Hatakeyama, A., Kobayashi, S., Jin,K., Kurahashi, N., Kondo, T., Gong, Y.Y., Umemura, T., 2004. Symptoms in relation tochemicals and dampness in newly built dwellings. Int. Arch. Occup. Environ.Health 77, 461–470.
Samet, J.M., Marbury, M.C., Spengler, J.D., 1988. Health effects and sources of indoor air-pollution. Part II. Am. Rev. Respir. Dis. 137, 221–242.
Schaper, M., 1993. Development of a database for sensory irritants and its use in estab-lishing occupational exposure limits. Am. Ind. Hyg. Assoc. J. 54, 488–544.
Taylor-Clark, T.E., Kiros, F., Carr, M.J., McAlexander, M.A., 2009. Transient receptor po-tential ankyrin 1 mediates toluene diisocyanate-evoked respiratory irritation.Am. J. Respir. Cell Mol. Biol. 40, 756–762.
Tin-Tin-Win-Shwe, Yamamoto, S., Nakajima, D., Furuyama, A., Fukushima, A., Ahmed,S., Goto, S., Fujimaki, H., 2007. Modulation of neurological related allergic reactionin mice exposed to low-level toluene. Toxicol. Appl. Pharmacol. 222, 17–24.


















