
NEST SITE SELECTION AND BREEDING SUCCESS IN … · NEST SITE SELECTION AND BREEDING SUCCESS IN...
57
1 Running head: Murrelet Nest Site Selection and Success. NEST SITE SELECTION AND BREEDING SUCCESS IN MARBLED MURRELETS IN RELATION TO DISTANCES TO EDGES AND FOREST PATCH SIZE, AT DESOLATION AND CLAYOQUOT SOUNDS, BRITISH COLUMBIA, CANADA. Yuri Zharikov * , David B. Lank, Falk Huettmann † , Russell W. Bradley ‡ , Nadine Parker, Peggy P.-W. Yen ‡ , Laura McFarlane Tranquilla and Fred Cooke § Centre for Wildlife Ecology, Department of Biological Sciences, Simon Fraser University, 8888 University Drive, Burnaby B.C. V5A 1S6, Canada Yuri Zharikov: Department of Zoology and Entomology, University of Queensland, Brisbane, QLD 4072, Australia, Fax: +61-7-3365-1655; Email: [email protected] * Department of Zoology and Entomology, University of Queensland, Brisbane, QLD 4072, Australia. † Department of Geography, 2500 University Drive NW, University of Calgary, Calgary, AB T2N 1N4, Canada. ‡ PRBO Conservation Science, 4990 Shoreline Highway, Stinson Beach, California 94970, USA. § Larkin’s Cottage, 6 Lynn Road, Castle Rising, Norfolk. PE31 6AB, UK.
Transcript of NEST SITE SELECTION AND BREEDING SUCCESS IN … · NEST SITE SELECTION AND BREEDING SUCCESS IN...

1
Running head: Murrelet Nest Site Selection and Success.
NEST SITE SELECTION AND BREEDING SUCCESS IN MARBLED MURRELETS IN
RELATION TO DISTANCES TO EDGES AND FOREST PATCH SIZE, AT
DESOLATION AND CLAYOQUOT SOUNDS, BRITISH COLUMBIA, CANADA.
Yuri Zharikov*, David B. Lank, Falk Huettmann†, Russell W. Bradley‡, Nadine Parker, Peggy
P.-W. Yen‡, Laura McFarlane Tranquilla and Fred Cooke§
Centre for Wildlife Ecology, Department of Biological Sciences, Simon Fraser University, 8888
University Drive, Burnaby B.C. V5A 1S6, Canada
Yuri Zharikov: Department of Zoology and Entomology, University of Queensland, Brisbane,
QLD 4072, Australia, Fax: +61-7-3365-1655; Email: [email protected]
* Department of Zoology and Entomology, University of Queensland, Brisbane, QLD 4072,
Australia.
† Department of Geography, 2500 University Drive NW, University of Calgary, Calgary, AB
T2N 1N4, Canada.
‡ PRBO Conservation Science, 4990 Shoreline Highway, Stinson Beach, California 94970, USA.
§ Larkin’s Cottage, 6 Lynn Road, Castle Rising, Norfolk. PE31 6AB, UK.

2
ABSTRACT
We studied habitat selection and breeding success in Marbled Murrelets (Brachyramphus
marmoratus) in two areas in SW British Columbia, Canada. Murrelets selected old-growth
forest fragments either proportionately to their size frequency distribution in a landscape or
nested in disproportionately smaller fragments. Overall, however, that it was not the size of a
patch per se that was important to the birds but the proximity of a nest site to an abrupt transition
from old-growth to an unvegetated area. Streams seemed to be particularly favored probably
because they may be associated with good nesting substrate conditions and provide access route
to a nest. Interestingly, the murrelets also selected areas with numerous recent clear-cuts for
nesting, disproportionately to their availability in a landscape. Breeding success patterns
generally fit the site selection patters, i.e. the probability of breeding success appeared greater at
sites located closer to natural or artificial high contrast edges. Our results suggest that despite
the usual perception of the Marbled Murrelet as forest-fragmentation sensitive species, these
birds may both select for and do well in areas with moderate or even high degree of forest cover
fragmentation.

3
Quantitative studies of habitat selection are fundamental in designing strategies for the
conservation and management of endangered and habitat-alteration sensitive species (McGarigal
and McComb 2002, Manly et al. 2002). Conservation biologists and wildlife managers strive to
establish: (1) how the spatial distribution of landscape components correlates with the pattern of
occurrence of individuals and (2) how such fitness parameters as survival or breeding success of
individuals co-vary with landscape composition or proximity of particular landscape components
(Donázar et al. 1993, Fielding and Haworth 1995, Boal and Mannan 1998, Suárez et al. 2000,
Morris et al. 2001, Meyer and Miller 2002). Adequate land management decisions are critical
when landscape components essential for existence and successful reproduction of individuals of
a species of conservation concern are altered by human activities (e.g. clear-cutting,
afforestation, draining). In this case any land management decision, whether it is for commercial
or conservation purposes, has to be weighed against its potential impact on the population of the
species in question and the consequences of such an alteration need to be predicted. Likewise,
designation of protected areas for a particular organism has to be based on information about its
landscape-level habitat associations as reliable as possible.
The Marbled Murrelet (Brachyramphus marmoratus), is a unique Alcid that nests in
coniferous old-growth forests of the NW Pacific coast of North America (Nelson 1997). Due to
extensive clear-cutting of the forested terrain along the Pacific coast, considerable loss of nesting
habitat has occurred (Booth 1991, Ripple 1994) and continues to occur when stands of trees
suitable for nesting (Hamer and Nelson 1995, Manley 1999) are removed. The major apparent
impact of the direct loss of nesting habitat, has been declines in the species population size in
several parts of its range (Nelson 1997). These declines resulted in the Marbled Murrelet’s

4
receiving a conservation status in the USA (U.S. Fish and Wildlife Service 1997) and Canada.
Recovery plans have been conceived and are being actively pursued in both countries with the
purpose of reversing or at least controlling the perceived population downward trend by ensuring
the maintenance of sufficient amount of old-growth forest suitable for the species reproduction
(D. Bertram per. comm.).
So far, much of the research into breeding ecology of the Marbled Murrelet has been
driven by the need to understand the nature, scale, and severity of the impacts of habitat loss and
isolation on the populations of the species (Ralph et al. 1995, Raphael et al. 2002a). To do that,
first, landscape-level associations (as opposed to e.g. individual nesting tree or stand level,
Hamer and Nelson 1995, Manley et al. 1999) of the species nesting sites need to be reliably
established (Raphael et al 2002a). Specific, landscape-scale information on Marbled Murrelet
nesting habitat preferences is in dire need especially where protected sites are to be designated in
areas otherwise slotted for logging (e.g. for British Columbia, Mather and Chatwin 2001). This
is because designation of sites suitable for Marbled Murrelet nesting still remains largely based
on earlier perceptions (e.g. the need for a large (>200 ha) contiguous area of forest) not
necessarily supported by the most recent data (Huettmann et al. submitted).
In this study we examined landscape-level habitat associations and breeding success for
two sets of Marbled Murrelet nests located at two sites in SW British Columbia (Canada), which
differ substantially in their historical levels of tree-logging activity and forest fragmentation. We
attempted to determine whether the choice of a location for a nest and the probability of the nest
being successful co-vary with the Euclidean distance to several discrete landscape features

5
(Conner et al. 2003). These features were (1) the physical edge of the nest patch per se, (2) a
hard-edge clearcut, (3) a fuzzy-edge clearcut, (4) a logging road, (5) a stream, (6) a lake, (7) a
subalpine area, (8) a glacier, (9) a cliff or bare rock area and (10) the ocean shore. Our choice of
the features was based on the notion that they are related to (1) accessibility of a site to an
airborne murrelet, which, despite being a strong flier (G. Kaiser. pers. comm.), has poor flight
maneuverability (Pennycuick 1987, Spear and Ainley 1997); (2) travel costs between the
foraging (sea) and nesting (land) habitats; and (3) susceptibility to biotic or abiotic “edge effects”
(sensu Murcia 1995). Four other variables potentially affecting site choice and nesting success
known from previous studies – nesting patch area (Raphael et al. 2002a), terrain elevation and
slope (Bradley 2002, Huettmann et al. submitted), and nesting date (Bradley 2002) were also
included in our analyses.
Previous research based on radar counts of Marbled Murrelets flying inland (Burger
2002a, Raphael et al. 2002b) and audiovisual detections of the birds flying overland (e.g. Meyer
et al. 2002) during a breeding season showed that these birds are more abundant (more
frequently encountered) in coastal areas with large expanses of unfragmented old forest. Here,
we (1) considerably augment and refine the knowledge of the Marbled Murrelet habitat selection
by identifying specific, discrete landscape features, with which nest locations are associated
using data for two distinct areas and (2) determine how nest site attributes co-vary with breeding
success. There are at least four important improvements in this study over the previous Marbled
Murrelet work. (1) We analyze largest available sets of confirmed nest sites removing the
element of uncertainty and other potential biases associated with previously used radar count or
“occupied detection” approaches (e.g. Jodice and Collopy 2000, Rodway and Regehr 2000,

6
Anderson 2001, Meyer et al. 2002). (2) We apply our approach to two climatically similar areas
with distinct differences in the degree of forest cover fragmentation. (3) We apply a realistic
edge/fragmentation scenario hitherto unused in Marbled Murrelet work. (4) We provide an
assessment of fitness consequences associated with a particular habitat choice (Jones 2001a).
METHODS
STUDY AREA
The study sites were located in the northern part of the Strait of Georgia around Desolation
Sound (50o05’ N, 124o40’ W) and on the west coast of Vancouver Island at Clayoquot Sound
(49o12’ N, 126o06’ W), British Columbia, Canada (Fig. 1a). This choice was influenced by the
presence of large populations of Marbled Murrelets (Kaiser et al. 1991, Burger 2002a), and by
ongoing and/or recent forestry activities at both sites. The areas are mountainous with many
steep cliffs, high ridges, U-shaped glacial valleys and several large fjords and inlets. Overall, the
terrain at Clayoquot Sound (CS) is less rugged than at Desolation Sound (DS). Subalpine
avalanche chutes and stream courses creating gaps in arboreal cover are numerous on the
mountain slopes. Elevation ranges from sea level to ca. 2,500 m at DS and 2,200 m at CS. The
original ground cover of the lower slopes was/is old-growth forest: western red cedar (Tsuga
plicata) and western hemlock (T. heterophylla) with a transition to yellow cedar (Chamocyparis
nootkatensis) and mountain hemlock (T. mertensiana) at higher elevations (Green and Klinka
1994). Further up is an area of intermittent forest and scrub cover, and at the highest elevations
is alpine tundra and glaciers (Alaback and Podar 1998).

7
Desolation Sound has a long history of logging with early exploitation of old-growth
trees beginning at the turn of the last century. Systematic clear-cutting started in the 1930’s, and
the area is still being logged. In the absence of historical forest inventories (Nilsson et al. 1999),
it is difficult to know exactly how much of the original forest has disappeared from the study
area, but the loss has been estimated at ca. 80% (Huettmann et al. submitted). In contrast,
Clayoquot Sound is a relatively pristine area with a few major clear-cuts and logging roads.
Commercial-level logging activity started there only after 1954, and as late as 1993 at least 75%
of the original forest cover remained (Kelson et al. 1995).
GENERAL BIOLOGY AND ECOLOGY OF THE MARBLED MURRELET
The Marbled Murrelet is unique among Nearctic Alcids in that, while foraging in coastal waters,
it nests mainly on mossy platforms (Antittrichia curtipendula, Dicranum fuscesens and
Isothecium myosuroides) of thick branches of old coniferous trees (Singer et al. 1991, Naslund et
al. 1995, I. Manley pers. comm.). Whereas many aspects of biology of the Marbled Murrelet are
well described (e.g. Ralph et al. 1995, Nelson 1997), its nesting ecology and the landscape-level
habitat associations of nest sites are not (e.g. Raphael et al. 2002a).
RADIO-TELEMETRY
To obtain a sample of radio-tagged individuals, murrelets were captured prior to the breeding
season (early May – early June) on the water at night at DS (1998-2001) and CS (2000-2002),
using a spot-lighting technique (Whitworth et al. 1997). Radio transmitters from Advanced
Telemetry Systems (ATS; 2.2g ATS Model 394 in 1998 and 3.2g ATS Model 386 in 1999-2002)
were attached to individuals with a subcutaneous anchor and glue. The batteries remained

8
functional for approximately 60 days in 1998, and approximately 90 days in 1999-2002.
Following capture and tagging, birds were monitored daily on both water and inland locations
using a helicopter (L. Lougheed and R. Bradley pers. com.).
NEST MAPPING
We located nests using radio-telemetry during the incubation period (Bradley et al. 2002).
During daily flights over the marine area, all radio frequencies were scanned until detected. For
those individuals where incubation was suspected (Bradley 2002), flights were then extended
inland to search for the nest. Inland, once a signal was detected, its location was searched for,
and then confirmed by circling over the source of the signal. The location was photographed
from the air, marked on a map and recorded using a 12-channel Global Position System (Garmin
GPS 48). If the site was accessible to ground crews, locations were confirmed by tree climbing
and mapped. Nest locations were processed using GIS (ArcView 3.2 ESRI 2001). Locations
from the three sources, i.e. aerial photos, manually marked maps and GIS-plotted maps, were
evaluated and adjustments were made when necessary in order to obtain the real location as
found in the field. This method is believed to be accurate within ±30 m (Huettmann and
Sandmann, unpublished). In total 121 nests were located at DS and 36 at CS.
BREEDING SUCCESS
Nests were classified as either active (successful) or failed based on the following criterion: a
nest was considered as active if radio-marked birds were visiting the nest sites regularly for at
least 20 of the 30 – 40 days of chick rearing period. This corresponds to the 'mid-fledging' stage

9
(Bradley et al. 2002, Cam et al., in press). We restricted breeding success analysis to a subset of
107 DS nests (70 active and 37 failed) whose fate was determined with the highest degree of
certainty. The ‘mid-fledging’ stage was used as a proxy of breeding success because these data
could be determined for all active nests whereas fledging success could only be determined for
nests sites physically accessible to ground crews. Breeding success was not analyzed at CS due
to the small sample size.
GIS DATA COMPILATION AND MANIPULATIONS
Following Huettmann et al. (submitted) all GIS data manipulations were carried out in ArcView
3.2 and Spatial Analyst 1.1 employing necessary tools and extensions freely available from the
ESRI website. Forest cover, rock, stream and road themes for each study area were compiled
from 1:20,000 GIS maps obtained from three sources representing the major land-holders and
land managers in the area: the British Columbia Ministry of Forests (MOF), Weyerhaeuser Ltd.
and Interfor Ltd. Lake, subalpine zone-avalanche chute, glacier and ocean themes were
compiled from 1:250,000 LUP (Present Land Use) maps obtained from the MOF.
The original forest cover information had been derived from aerial photographs, which
were classified by aerial photo interpreters following approaches described by the B.C. Ministry
of Forests (1999). Some gaps in the available forest cover data existed at both DS and CS. At
DS, 24 nests fell into higher-elevation areas, for which no forest cover attribute data were
available. To alleviate the problem, sites within 1 km radius of each of these nests were
classified from available aerial photographs (Weyerhaeuser Ltd.) by a trained forest cover
interpreter following the standard MOF guidelines. This additional data set was merged with the

10
originally available forest coverage. At CS no nests fell into unclassified areas. Complete
coverage for all the other landscape features of interest was available for both study sites.
From the forest cover maps, we defined as “old-growth forest”, forested polygons with
either the primary or, where possible (CS – Vegetation Resource Inventory, RIC 2001)
secondary species being 141 years of age and older for DS (age classes 8 and 9) and 251 years
and older for CS (age class 9) with tree height of at least 15 m (height class 2 and above) for CS
and all height classes for DS. These selection criteria are conservative as they include (1) high
elevation tree stands, which are not tall but are old and may still be suitable for nesting and (2)
polygons containing trees or clumps of trees suitable for nesting embedded in a matrix of
unsuitable trees. These criteria are based on the preliminary data (L. Waterhouse, pers. comm.)
on Marbled Murrelet nest tree stand characteristics in both areas as classified from aerial photos
and our knowledge of vegetation patterns in the study areas.
We defined a forest patch as contiguous forest within the appropriate age category as
delimited by streams and roads. To create these patches, forest cover themes were processed in
the following way. (1) All polygons within the age and height range specified above were
converted to a 15x15 m grain raster map. This step dissolved the botanical (e.g. crown closure,
dominant tree species, height) boundaries between the original polygon “patches”. Such
boundaries are routinely created as a part of forest cover inventory but, here, are assumed not to
represent distinct real-life patch boundaries (edges) in a landscape. (2) Road and stream polyline
themes were also converted to 15x15 m grain raster maps. The grain size approximated the
width of a buffer (12 m) typically applied to mapped line features by the MOF (J. Sundé, pers.

11
comm.). (3) The forest cover and road and stream themes were overlaid and areas of overlap
were removed from the data set fragmenting the forest cover along the road and stream lines.
Finally (4) the raster forest cover data were converted back into the vector (polygon) format to
allow for the nest-to-edge distances and individual patch areas to be measured. (5) After this last
conversion we filtered out all fragments <0.045 ha (2 grid cells) in area to simplify the coverage.
These fragments made up <0.1% of the total forest area. We also visually examined all
unusually large (≥1000 ha) forest fragments. In cases when spatially separated parts of such a
fragment were joined by an isthmus ≤50 m wide, they were manually divided. Although
arbitrary, this step was taken, to minimize the chance of drawing the same fragment more than
once during site selection analyses (below). Only about 10 polygons (out of ca. 8000) had to be
manipulated in this fashion in each study area. These steps created a mosaic of old-growth forest
fragments as defined by both, the age/height attributes typical of the Marbled Murrelet nesting
habitat in the region, and streams and roads. These linear features fragment forest cover in real
terms but are not accounted for in standard forest cover maps. Such a design has not been
previously applied to forest patch definition in the studies of Marbled Murrelet habitat
associations (e.g. Meyer et al. 2002, Raphael et al. 2002b) or, to our knowledge, in other bird
forest habitat selection studies (e.g. Bayne and Hobson 2002).
To have the most up-to-date information on the locations of clear-cuts, they were
classified from two recent 30x30 m resolution LandSat 7 ETM images (DS 13 August 2000, CS
19 September 1999) obtained from the Geogratis Website (Geograstis 2002). Remote image
processing software eCognition® (Definiens Imaging GmbH 2002) was used for classification
and decision support (Steeby and Hall-Beyer, unpublished). eCognition has been developed to

12
overcome the limitations and weaknesses (e.g. high degree of pixel interspersion) of traditional
maximum likelihood pixel-based image processing methods for feature extraction from high
resolution images (Hofmann 2001, Waser and Schwartz 2001). Its strength lies in the fact that
important semantic information necessary to interpret an image, which is not represented in
single pixels but in meaningful image objects (e.g. buildings, roads, clear-cuts) and their mutual
relations, becomes incorporated into the process of image classification. Using eCognition
software, we were able to distinguish hard-edge clear-cuts, i.e. those completely devoid of trees
and fuzzy-edge clear-cuts, i.e. those containing small (≈900-1800 m2) tree patches. These
patches represented remnant old and secondary growth trees, which could not be reliably
separated from the surrounding clear-cut matrix to due their size. Presence of these trees resulted
in a different brightness signature of fuzzy-edge clear-cuts as opposed to hard-edge clear-cuts
allowing the distinction.
To determine whether nest site location differs from random we placed 1000 random
points within the old-growth stratum of a minimum convex polygon (MCP) encompassing the
distribution of nest sites with a 2.3 km (DS) or 3.1 km (CS) external buffer. The outside buffer
values represent the mean annual nearest-nest distances. MCP area (DS = 3.33*109 m2, CS =
1.52*109 m2) is assumed to represent potentially available breeding habitat for the birds captured
on the water (Fig. 1b, c). With coarse-scale habitat associations of nesting Marbled Murrelets
already known (reviewed in Ralph et al. 1995, Nelson 1997, Raphael et al. 2002a), the choice of
the extent of “available” area in this case aimed at (1) determining characteristics of locations
preferentially used within an established population (e.g. Erickson et al. 1998) and (2) removing
the degree of arbitrariness inevitable when available area is defined without taking organisms’

13
distribution into account (Gates and Evans 1998, Jones 2001a). We decided to use an unequal
sample size (1000 available random locations versus the appropriate number of nests) since this
allows to derive a more representative sample for the highly variable available area and does not
appear to bias the results (Nielson et al. submitted, see also Cramer 1999).
VARIABLES
A landscape feature was defined as per GIS database and represented a unique polygon for areal
features (forest patches, clear-cuts, lakes, ocean, etc) or unique polyline segment for line features
(roads, streams). Using the Nearest Feature 3.4 extension to Arcview 3.2 we measured
Euclidean distance to 1 m from each nest and random point location to the nearest edge of the
following features: (1) the forest patch, in which it is located; (2) three closest hard-edge clear-
cuts, (3) three closest fuzzy-edge clear-cuts, (4) logging road, (5) three closest streams, (6) lake,
(7) subalpine area-avalanche chute, (8) glacier, (9) bare rock area and (10) ocean shore. Point-
to-edge distances for three closest features for variables (2), (3) and (5) were measured to
account for a possible density effect on nest site selection and breeding success because these
features were visibly numerous within the landscapes. A mean of the three nearest distances was
treated as a separate variable along with the nearest feature distance per se. Area (ha) of the
underlying old-growth patch was recorded. Elevation above sea level (to 1 m) and slope (to
0.1°) indices for nests and random points were derived from a 25x25 m gridded Digital Elevation
Map (DEM) by recalculating each cell to the mean of its 9-cell neighborhood. The date of
initiation of incubation (1st April was set to 1) was determined from a persistent on-off radio-
tracking pattern reflecting switches by incubating parents (see Bradley 2002 for details). Thus,

14
the total initial number of candidate variables was 16 for site selection and 17 for breeding
success analyses. Three nests (out of 36) at CS and 35 nests (out of 121) at DS were missing
patch area (PA) and patch edge distance (PED) information because they fell into areas that did
not meet our “old-growth” definition criteria. The most likely reason for this is that in the
process of forest cover map compilation, areas with ≤10% of old-growth within the forest stand
are considered trivial by aerial photo interpreters and are ignored, when these small patches may
well be suitable for the birds (e.g. Naslund et al. 1995, I. Manley pers. comm.). We dealt with
the missing values as follows: the three missing values in the CS data set were replaced with the
overall mean; the DS data were analyzed (1) using all selected candidate variables and the
corresponding 86 nest data points and (2) excluding PA and PED and using only those variables
for which all 121 nest data points were available.
ANALYSES, VARIABLE SELECTION AND MODEL FITTING
Our general analytical approach to site selection was to compare the spatial distributions of
known nesting locations against a random set of available locations (Manly et al. 2002). This
approach does not rely on the assumption that the locations chosen at random are unused (Jones
2001a): the inference about selectivity is based on possible deviations from patterns expected
under the hypothesis that use is random with respect to the variables analysed.
Because old-growth forest patch size has major economic and aesthetic implications
(FEMAT 1993) the importance of the patch area variable on habitat selectivity in the Marbled
Murrelet is particularly relevant to the specification of management plans. Presently in Canada
individual old-growth fragment area (>200 ha) is the sole most important criterion in designating

15
protected sites for this species (IWMS 1999). To assess the importance of this variable for
Marbled Murrelets, PA was first exhaustively analyzed on its own.
We viewed the problem as follows. There is a large sample of k forest patches each of an
area αi, i=1,…, k. There are also N nests in the study area and we wish to test whether or not the
probability of a nest falling into a patch is proportional to its area. The number of known nests is
much smaller that the number of forest fragments (86 versus 10154 and 33 versus 6868 for DS
and CS respectively). If pi is the probability of finding a nest in a given patch, the number of
nests per patch is a Poisson distributed variable with the mean ii Npe = . The null hypothesis is
ii cap = where c is a constant estimated as N/A ( ∑= k
iiaA ).
Since we were specifically interested in knowing how the probability of finding a nest in
a patch varies with a monotonic increase in its area, the standard X2 statistic was not appropriate
as it does not take the order of cells into account. Therefore, a Cramér-von-Mises W2 statistic
was used (Choulakian et al. 1994). The statistic compares the cumulative theoretical distribution
with its estimate and is calculated as follows.
∑=
−=k
jjjtZNW
1
22 1
In this case let ∑ == j
iij oS
1 and ∑ =
= j
iij eT
1. Then Sj/N corresponds to the empirical distribution
in the continuous case. Suppose Zj = Sj – Tj, j = 1,…, k and let ti = (pi + pi+1 )/2.

16
Distribution under null hypothesis was developed using the basic Monte Carlo procedure
(Manly 1990) using the probabilities pi calculated as above and the observed values oi following
two steps. (1) We generated N random numbers from the uniform distribution between 0 and 1
and used these to allocate N nests in the k patches. (2) We calculated the W2 for the allocation.
These steps were repeated 1000 times and the P-values calculated. Additionally, Kolmogorov-
Smirnov Dmax-statistic was employed to determine the case with maximum difference |Zj|
between observed and predicted distributions.
For illustrative purposes we ranked individual old-growth fragments in the increasing
order of area and plotted cumulative old-growth area versus the cumulative number of nests
encompassed. It is expected that if no selection for a particular class size occurs, the resulting
plot will represent a straight line ( ii cap = see above). Otherwise deviations above or below the
null (neutral selection) pattern representing disproportionate use or avoidance respectively are
expected. Finally, the distribution of nest PA values binned in several classes was compared to
the one generated via placing 1000 random points into the landscape using goodness-of-fit tests.
This last step was also applied to PED values.
We used generalized linear models (GLZ module in Statistica©) with a logit link-
function to identify the set of variables that best separated nest sites from random points or active
from failed nests. Although data normality is not a formal requirement of logistic models, all
candidate distance and area variables were loge transformed to decrease sample variance
heterogeneity. Variable selection and model fitting were carried out following four steps. (1)
Univariate logistic regression models were used to test the null hypothesis of no difference

17
between the nest sites and random location (active and failed nest sites) for each continuous
landscape variable. All the variables with P≤0.11 were selected for multivariate analyses. (2) A
correlation matrix was constructed so that if two highly intercorrelated variables (r≥0.70, e.g.
Fielding and Haworth 1995) were found, one of them could be excluded. (3) The most
parsimonious set of variables (best approximating model, sensu Burnham and Anderson 2002)
describing nest location or probability of success was selected based on the Akaike’s Information
Criterion (AIC) or AICc if the ratio of the number of predictor variables to the sample size was
<1:40 (Burnham and Anderson 2002). “Best Subset” routine in the Statistica©’s GLZ module
was used for these analyses. A lower AIC score indicates a better approximating model, whereas
scores differing by less then 2 are considered similar in their ability to describe the data,
regardless of the magnitude of the AIC. Apart from the original variables, quadratic terms for
elevation and slope were included. This is because preference for intermediate values of these
environmental parameters may be displayed by the birds (Huettmann et al. submitted) resulting
in a ∩-shaped distribution. No similar a priori information was available for the other variables.
Therefore, no polynomial terms were included in the models. We also did not include first (or
higher) degree interaction terms since we could not define a priori any effect of interaction
between any of the two variables of site selection or breeding success. A full factorial design on
the other hand would have resulted in a large number of predictor variables increasing the danger
of overfitting a model and obtaining spurious results. (4) Multiple R2 and Kappa indices
representing improvement over random selection and the proportion of specific agreements in a
standard confusion matrix:
observed true/false versus predicted true/false (see Fielding and Bell 1997 for details)

18
as well as individual parameter coefficients were calculated for the best approximating model.
RESULTS
SITE SELECTION RELATIVE TO PATCH AREA AND PATCH EDGE DISTANCE
At Desolation Sound (86 nests), the mean sizes of nest patches and patches into which random
points fell were identical (Table 1). However, tests comparing the distributions showed
significant differences: the Cramér-von-Mises W2 = 0.59 (P = 0.021) and Dmax = 13.65 (P =
0.021) at fragment with the area of 9.8 ha. Therefore, we can reject the hypothesis that the nests
are distributed proportional to area. Specifically there was an apparent preference for fragments
ca. 10 and 200 ha in area (Fig. 2). Splitting the distribution of patch areas into 10 size classes
(Fig. 3) showed an identical pattern of two peaks in apparent preference: the smallest class (≤10
ha) and the intermediate size class (150-210 ha) (goodness-of-fit test, X29=29.3, P<0.001).
Overall, the mean patch size at Clayoquot Sound (33 nests) was 2-3 times larger than at
Desolation Sound (Table 2). Nest patches were not statistically different from patches associated
with random points (Table 2). Tests comparing the distributions also produced non-significant
results: the Cramér-von-Mises W2 = 0.08 (P = 0.69) and Dmax = 4.46 (P = 0.52). Therefore, we
cannot reject the hypothesis that the nests are distributed proportional to area. The cumulative
plot line displayed a linear trend except for an upward deviation at the higher end (Fig. 2).
However, splitting the distribution into a discrete number of classes (Fig. 3), still produced a
non-significant trend (X28=9.00, P=0.34).

19
At DS, nests on average were closer to the edge of an old-growth patch than random
points (Table 1). Although the distribution of nest to forest edge distances did not differ from
random, (Fig. 4, goodness-of-fit test, X26=8.59, P=0.20), nests tended to be found more
frequently within 30 m of edge (49%) of an old-growth forest patch than would be expected at
random (39%) (X21=3.35, P=0.067).
At CS the distribution of nest to forest edge distances did not differ from random (Table
2) whether seven (Fig. 4, X26=5.98, P=0.42) or two (≤30 m versus >30 m, X2
1=0.22, P=0.64)
distance classes were considered.
SITE SELECTION: MULTIVARIATE ANALYSES
Desolation Sound. Of 16 original candidate variables, five (distances to the nearest fuzzy-edge
clear-cut and the mean of three nearest FE clear-cuts, lake, ocean shore and subalpine area) were
eliminated as they did not differ from random in univariate analyses (see Table 1). Among the
remaining variables, distance to the nearest hard-edge clear-cut was strongly correlated with the
mean distance to the three nearest clear-cuts (variables 7 and 8, r=0.73, P<0.001). The same was
true for the variables 14 and 15 (r=0.75, P<0.001). Pair-wise correlation coefficients for all the
other variables were <0.42. Variables 7 and 15 were thus excluded due to their comparatively
weaker discriminatory performance in univariate logistic regressions. The rest of the variables
were entered into two multivariate regressions: (1) included all 11 remaining variables plus the
squared terms for Elevation and Slope Indices (n=85 nests, Table 3); (2) included 9 variables
(PA and PED excluded) plus square terms for Elevation and Slope Indices (n=121 nests, Table
3). In both models nests were located at sites with higher densities of hard-edge clear-cuts and

20
closer to streams, farther from glaciers, at lower elevations and on a steeper slope than would be
expected at random. Distance to the nearest rock-face or a logging road were model-specific
variables. Models with the full nest dataset resulted in a better discriminatory power (Қ) than the
models that included PA and PED as input variables and therefore a reduced number of data
points (missing cases). Since PA and PED did not consistently contribute to the multivariate
model anyway, it seems prudent to use the best approximating model based on the full data set
for discussion and interpretation purposes (Model 2, Table 4).
Clayoquot Sound. Of 16 original candidate variables, seven were initially selected as
input variables for multivariate logistic regression modeling. Two pairs of variables (10 and 12
and 14 and 15, see Table 2) were strongly correlated (r=0.71 and -0.70 respectively, P<0.001).
Variables 12 and 15 were excluded due to poorer fit to a univariate logistic model. As with the
DS data, quadratic terms for the Elevation and Slope Indices were included into the model
selection process.
A set of models including distances closer to streams and further from rock-faces provide
the best fit to the data (Table 5).
BREEDING SUCCESS
Of the eight candidate variables (Table 6) none were strongly inter-correlated (r<0.52). All were
entered into a multivariate logistic regression for the DS data set, and four accounted for the
largest difference between the active and failed nests (Model 1, Table 7). Active nests were
initiated earlier in the season and be located closer to the edge of an area of subalpine vegetation,
in a location with higher hard-edge clear-cut density, and at higher elevation, than failed nests.

21
DISCUSSION
Distance-based analyses of habitat selectivity are frequently used to analyze organism-habitat
associations. This is because they allow (1) for potentially important landscape features to
influence results of the analysis without making explicit assumptions about the scale of the
interaction (e.g. Morris et al. 2001) and (2) to detect the effect of features that lack area (i.e.
edges) (Conner et al. 2003). The scale at which Marbled Murrelets interact with their terrestrial
environment is unknown (Meyer and Miller 2002). Also the question of “edge effects” in
artificially fragmented habitats is gaining an ever increasing prominence in habitat selection
studies (e.g. Murcia 1995, Kremsater and Bunnell 1999). Therefore, we believe ours was an
appropriate approach to investigate habitat associations in the species.
Prior to this study the understanding of large-scale habitat associations in the Marbled
Murrelet, a protected species notoriously difficult to study due to its secretive nesting habits
(Jones 1993, Nelson 1997, Jones 2001b), was often based on count studies. Counts were
conducted in tandem with, usually, coarse-scale sampling of habitat data from the available GIS
databases (Jodice and Collopy 2000, Meyer et al. 2002, Miller et al. 2002, Raphael et al. 2002b).
Undoubtedly, these approaches provided much needed baseline information for the initial steps
in conservation and management of potential Marbled Murrelet nesting habitat. However, they
may have produced a somewhat biased view of the species habitat preferences due to non-
random sampling (Jodice and Collopy 2000, Rodway and Regehr 2000; see Anderson 2001 for a
critique of count studies). Also, they could not focus on the specific landscape-level associations

22
of these birds for a simple reason: a large and unbiased sample of confirmed nests from a single
location was not available. This study and a parallel study by Huettmann et al. (submitted) are
the first significant endeavors aimed at (1) determining the landscape components important in
nest site selection process and, importantly, (2) assessing the correlation between these
components of the breeding success of the murrelets using a large and unbiased data set. Such
information has important conservation implications and is indispensable in Marbled Murrelet
habitat management decision-making over its range of distribution.
PATCH SIZE
Univariate analyses suggested that at Desolation Sound Marbled Murrelets selected both smaller
and somewhat larger patches than would be expected at random. In addition, almost a third of
the nests were found in areas not classified as old forest. It seems reasonable to assume that the
“old-forest patch sizes” for these nests, if defined, would be small (≤10 ha, cf. Fig. 2). Thus our
conclusion that there is disproportionate usage of small patches within classified areas may
substantially under-represent the usage of small patches. On the other hand, there also is a
second, but smaller peak in patch size usage in the 200-250 ha range. This bimodality may be
real and the second peak may reflect usage of large but highly contorted areas with much edge
(Meyer and Miller 2002).
Overall, the results with regard to patch size selection are in contrast to the currently held
view of the species’ strong preference for large tracts of unfragmented old-growth landscapes
(e.g. Mather and Chatwin 2001, Meyer and Miller 2002, Meyer et al. 2002). We found
preference for smaller patches of old-growth forest (DS) and no clear patch size selection (CS,

23
see below). We believe the discrepancy is not so much due population differences or local
landscape patterns, as it is due to scale of investigation and confusion of preference (product of
positive selection) with the abundance or frequency of use (van Horn 1983). Gates et al. (1994)
for example showed a positive large-scale association between the European Skylark (Alauda
arvensis) abundance and the amount of area sown with grain-crops (Alauda larks inhabit open
environments). However, an intensive small-scale study showed that current farm management
practices preclude settlement and may have even been responsible for population declines
(Donald and Vickery 2000). Similarly, Marbled Murrelets are expected to be more abundant in
areas with much old-growth habitat, due to their very specific nesting requirements. However,
our results suggest that within these habitats, they actually select sites with moderate or even
high degree of forest cover fragmentation. The nature of fragmentation (natural or
anthropogenic, also see Manley et al. 1999), at least in the short-term (cf. Miller et al. 2002) does
not seem to matter. Strikingly, most of the studies (see Site Accessibility subsection) of Marbled
Murrelet – old-growth forest associations directly or indirectly acknowledge co-occurrence the
murrelets and high contrast edges without explicitly suggesting that forest fragmentation (i.e. the
“breaking apart” of a contiguous area) per se, as opposed to straightforward habitat loss (Fahrig
1999), may have no negative consequences for the species (but see Raphael et al. 2002a).
MICROCLIMATE, BIOTIC AND ABIOTIC EDGE EFFECTS
Neither the patch size nor the distance to the edge of a patch, were retained in the multivariate
models. We interpret the models as suggesting that proximity to a landscape feature producing
an abrupt transition from forest to a relatively open area was favored. Such a transition (1) is

24
likely to have a strong influence on the local microclimate patterns (e.g. Brosofske et al. 1997,
Chen et al. 1995, 1999, Butler 2001) and (2) may facilitate access to a nest site for this heavily
wing-loaded species (Manley et al. 1999, Waterhouse et al. 2002, Nelson et al. 2003).
Marbled Murrelets showed a strong tendency to nest near streams suggesting that this
landscape feature represents an important cue in nest site selection process by the birds. Riparian
zones are ecotones supporting a wide range of plant and animal life (Nilsson 1992). Specifically,
in the Pacific Northwest region, montaine riparian habitats support a considerably greater
diversity and abundance of epiphytic mosses (e.g. Jonsson 1997, Peck and Muir 2001, Rodway
and Regehr 2002), that typically form Marbled Murrelet nesting platforms (Singer et al. 1991,
Naslund et al. 1995, I. Manley pers. comm.), than upland habitats. This is because bryophytes
are only metabolically active when wet and are generally adapted to survive in mesic, low-light
conditions (e.g. Cleavitt 2002). Availability of thick mossy platforms on mid-canopy branches is
of critical importance to Marbled Murrelets because they do not build nests as such but lay their
single egg into a depression in the epiphyte substrate (Nelson 1997). This dependence on nesting
substrate availability is thought to be the main reason why the extent of epiphyte cover on
branches as well as the density of trees with mossy platforms per stand, are strongly correlated
with the small-scale nest site choice by the birds (Manley 1999, Rodway and Regehr 2002).
Thus, the observed tendency by the birds to nest closer to the streams than expected from
random may be explained by more favorable nesting substrate conditions there. On the basis of
a correlation between local epiphyte vegetation patterns and Marbled Murrelet habitat use, a
similar conclusion was reached by Rodway and Regehr (2002). In our study areas (DS and CS)
streams occur on steep mountain slopes. Therefore our site selection findings do not support

25
previously made suggestions that murrelets preferentially breed in old-growth forest at lower
elevation and in flat river valleys (e.g. Nelson 1997, Bahn 1998).
The relevance of the distance to glaciers seems less clear. Being large, permanent macro-
landscape features, glaciers are likely to have a strong affect of local climatic patterns (e.g.
Mizuno 1998, cf. Burger et al. 2000 for effects of the ocean). If sites located closer to a glacier
have an overall lower mean annual temperature, then their plant growing period will be shorter.
This will result in lower moss abundance and thus poorer nesting conditions (cf. Rodway and
Regehr 2002). Therefore such sites may be avoided by the murrelets.
Marbled Murrelets also nested closer to hard edge clear-cuts and at sites with higher
clear-cut density, than would be expected at random. This may be a counter-intuitive and even
controversial observation. However, the explanation for this pattern may simply reflect, and be
an artifact of, the limited availability of undisturbed old-growth forest (ca. 20% of original land
cover) at Desolation Sound (Huettmann et al. submitted). Our results could also explain why a
high inter-annual consistency in the nest distribution (and nearest-neighbor nest distance) was
observed at DS (F. Huettmann, unpublished).
Unlike with the streams, Marbled Murrelets did not select nest sites in the immediate
vicinity (i.e. tens of meters) of clear-cut edges (Table 1). Also, the distance to a clear-cut edge,
although highly variable, was on average well beyond the range at which open clear-cut
microclimatic conditions can penetrate into the old-growth forest interior (usually < 200 m) and
affect its biota (Chen et al. 1995, 1999). Rather, the birds nested in areas with numerous recent
(<15 – 20 years) clear-cuts. Our interpretation here is that the prime Marbled Murrelet habitat

26
and areas that were slotted for logging had very similar characteristics and/or were spatially
juxtaposed. Also, as will be shown below, there is no immediate fitness cost to the birds that
continued to nest in areas being logged or recently logged in terms of e.g. disturbance or
increased predation rates (Marzluff and Restani 1999, Raphael et al. 2002a). In Oregon, murrelet
old-growth habitat usage was lower in areas fragmented 20 years previously than in recently
similarly fragmented areas (Meyer et al. 2002). The argument put forth was that, whatever the
reason, these birds display a delayed response to habitat fragmentation. One likely mechanism
of such a time-lag may involve increased predation rates in fragmented areas (e.g. Manley et al.
1999). However, the relationship between nest depredation rates and degree of habitat
fragmentation is highly case-specific (Marzluff and Restani 1999, Lahti 2001, Raphael et al.
2002a) and no consistent relevant information is available for our study area. If mortality
exceeds recruitment, then a population will decline and eventually become extinct. However, in
our study area there is no evidence of population decrease in the Marbled Murrelet population
(Cam et al. in press). Thus, it is probably healthy.
The lag may also be due to the retarded growth and/or dispersal of old-growth forest
epiphytes near clear-cut edges (e.g. Esseen 1994, Renhorn et al. 1997, Sillet et al. 2000, Thomas
et al. 2001) due to adverse microclimatic conditions (Chen et al. 1999). Under this scenario old-
growth patches surrounded by regenerating forest (old clear-cuts) may have already reached the
lowest point in epiphyte growing conditions, whereas the stands adjacent to recently logged lots
still maintain their pre-disturbance condition and thus are not avoided.
SITE ACCESSIBILITY

27
Streams, clear-cuts and roads leading to them may also provide the birds with convenient
flyways to access potential nesting sites. Accessibility to nest sites may also account for the
relatively high usage of small stands not mapped as old forest. The physical characteristics of
these old trees embedded in the secondary growth matrix did not differ from those, found within
old forest, nor those reported for murrelets elsewhere (I. Manley. pers. comm.).
A detailed study of small-scale habitat associations in Marbled Murrelets just south of
our study area (Manley 1999) revealed a strong correlation between nest locations and proximity
of canopy gaps. Importantly, the nature of the gaps (e.g. riparian, industrial) did not seem to
matter to the birds. Similarly, Meyer and Miller (2002) reported a higher edge contrast index in
areas more frequently used by Marbled Murrelets suggesting that the birds’ abundance was
greater in landscapes with numerous abrupt transitions between forest and non-forest land cover
types. Furthermore, Bahn (1998), Burger (2002b), Waterhouse et al. (2002) and Nelson et al.
(2003) reported high canopy complexity as an important Marbled Murrelet habitat requirement.
Results of these studies and the pattern of habitat selection found by us strongly suggest that
murrelets specifically target sites with a high degree of vegetation and terrain vertical complexity
(also Huettmann et al. submitted). It is noteworthy that “typical” coastal Alcids select for similar
features in their nesting environments. Nettleship and Birkhead 1985 for example showed that
Atlantic Puffins (Fratercula arctica) selected steeper slopes and proximity to cliff-edges for their
nesting sites (burrows). Gaston (1992) reported a similar selection pattern in the Ancient
Murrelet (Synthliboramphus antiquus). This view meshes well with the poor flight
maneuverability of the Alcidae (Pennycuick 1987, Nettleship and Birkhead 1985). While being
unique in its nesting habitat requirements, the Marbled Murrelet is a typical Alcid when it comes

28
to fight agility. The birds use their nesting platforms much the same way as the coast-nesting
Alcids use cliff-ledges. They fly up from beneath and land by stalling. The reverse process
happens during a take-off: a bird drops from a branch and then levels off as it accelerates (e.g.
Jones 2001b). Importance of a clear flyway to a nest thus, seems quite apparent. Accessibility
of a convenient flyway may be even more critical for fledging young who, if they become
grounded during their maiden flight, have virtually no chances of taking off from the ground
(Carter and Sealy 1987). For a slowly reproducing, low fecundity bird, as all Alcids are, nest site
choice thus may be of critical adaptive significance (e.g. Harris et al. 1997). Furthermore,
landscape features creating gaps and abrupt edges in vegetation may be used by the commuting
murrelets as landmarks. Indeed, it has been observed (N. Parker, pers. comm.) that Marbled
Murrelets fly low above the surface/ground during periods of thick fog – a frequent occurrence
along the coast of BC. Therefore, a “flyway” feature does not necessarily need to lead directly to
a nest site (a local canopy gap may suffice there, Manley 1999), but it very well may be used for
an initial entry into a general nest locale (Burger 2002b).
Treeless rocky areas may serve a similar function as other gaps in arboreal vegetation, i.e.
to allow access to suitable nest sites. Effects of elevation and slope on nest site selection have
been discussed in detail by Huettmann et al. (submitted) and Bradley (2002) in terms of predator
abundance and site accessibility. Here we show that these topographic features still stand as
important predictors of habitat selectivity despite the inclusion of additional habitat variables.

29
SITE SELECTION AT CLAYOQUOT SOUND – IMPORTANCE OF STREAMS
Analysis of habitat selection at CS was based on a considerably smaller number of nests than at
DS. At CS, a much more pristine area, Marbled Murrelets used old-growth patches relative to
their availability in the study area. Shorter distance to the edge of a patch might have been
selected for as at DS, but the result was not significant due to a small sample size. Similar to DS,
murrelet nests found at CS, were located closer to a stream than would be expected at random.
In conjunction with the DS data and other information provided above, we suggest that the
observed association with stream-side old-growth forest may be a globally important habitat
selection criterion in the species regardless of the degree of habitat fragmentation and/or local
geomorphic patterns.
The selection for rocky areas at CS showed the opposite trend to that observed at
Desolation Sound. Overall, at DS rock made up 6.5% of the landscape cover and it was
uniformly distributed. By contrast at CS only 0.5% of the landscape cover was represented by
rock and its distribution was restricted to the NE part of the MCP. With the mean distance to the
nearest rocky site being 4 – 5 km (cf. 0.5 – 0.6 km at DS) we believe the pattern to be an artifact
of rarity and restricted location of the rocky sites irrelevant to selection by Marbled Murrelets.
CORRELATES OF BREEDING SUCCESS
If a temporal and/or spatial gradient in quality of available breeding sites exists, the breeding
success of individuals should vary across the system relative to the sites’ habitat characteristics
whether the latter could be explicitly identified or not.

30
As many other Alcid (Nettleship and Birkhead 1985, Hatchwell 1991, Hipfner et al.
1999) and non-Alcid species (Newton and Marquiss 1984, Brinkhof and Cave 1997, Hinsley et
al. 1999), Marbled Murrelets displayed a strongly negative seasonal trend in the probability of
breeding success. This decrease could occur either due to deteriorating environmental (e.g.
foraging) conditions for the parents or the quality of the parents themselves (Verhulst et al.
1995). Hipfner (1997) and Hipfner and Gaston (2002) attempted to disentangle these two factors
in an Alcid, the Thick-billed Murre (Uria lomvia) and found support for the parent quality
hypothesis, but not for the seasonal deterioration hypothesis. They suggested that the late
breeding of less capable birds is the likely proximate cause of many of the seasonal declines in
success reported in the Alcidae. In other Alcid species, however, the seasonal deterioration
hypothesis is more likely (e.g. Vermeer and Cullen 1979). At present there is no sufficient data
for the Marbled Murrelet populations in the study areas or elsewhere to suggest one way or the
other. Also, if seasonal deterioration of the environment plays a role, we do not know whether
negative changes occur in the marine environment with the food base (Vermeer and Cullen 1979,
Kitaysky and Golubova 2000) or in the terrestrial environment due to, for example, increased
predation risk (Hartman et al. 1997). Answering these questions will represent useful and
rewarding directions for future research. It is noteworthy, however, that earlier nesting birds
travelled farther from the ocean, suggesting that the cost of travel in this case is trivial. They
also selected steeper slopes which may provide for a better visibility of the surrounding area for
the parents and improve their chance of detecting predator threat (e.g. Northern Goshawk
(Accipiter gentilis) Ethier 1999 as cited by Burger 2002b). Higher breeding success was
recorded for pairs nesting on steeper slopes as has been found in the ground-nesting Alcids

31
(Nettleship and Birkhead 1985). This finding suggests that the location of a nest site in a more
broken terrain makes it less vulnerable to predators.
Our data show that nests located in areas with higher clear-cut density had a higher
chance of being active. It has been argued that the main reason of nest failure in the Marbled
Murrelet is predation by birds and mammals (Nelson 1997, Manley 1999). Fragmentation of
old-growth habitat will increase the risk from predation only if the newly created (edge) habitat
either supports a greater population of potential predators or allows for a better detectability of
prey’s nests. It is not at all clear that in our case such trends will occur due to creation of new
clear-cuts. Marbled Murrelets have a highly cryptic coloration (Nelson 1997) and they
commonly nest in the environments (stream-side forest stands) that support a higher than average
diversity and abundance of predators (e.g. for CS see Rodway and Regehr 2002). Therefore,
creation of additional edges probably does not make their nests more detectable to their potential
predators. Creation of edges will only increase local predator populations if it results in a
concurrent increase in staple food supply from anthropogenic (Raphael et al. 2002a) or natural
(Vickery et al. 1992) sources. Both almost never happen due to clear-cutting as opposed to e.g.
human settlement (Andrén 1995, Marzluff and Restani 1999, Song and Hannon 1999). As our
nest locations were relatively far from permanent human habitation (cf. Manley 1999), it is
unlikely that creation of clear-cuts resulted in a greater supply of food of anthropogenic origin.
At the same time it could have easily resulted in a decrease of natural food base for such typical
boreal species and known adult and nest predators of Marbled Murrelets (Nelson 1997) as the
Northern Goshawk (Ethier 1999), Common Raven (Corvus corax) and Gray Jay (Perisores
canadiensis) (McGarigal and McComb 1999, Raphael et al. 2002a). The amount of nesting

32
habitat suitable for potential predators could also decrease due to clear-cutting. This, along with
the habitat structure argument made above, may explain the higher breeding success in actively
logged areas.
If the proximity to a stream was important in nest site selection, why were murrelets
nesting near streams not more successful? If riparian microclimatic conditions improve
bryophyte growing conditions, then the reason may be due to the great abundance of possible
nesting platforms. Manley (1999) showed that micro-site selection by the Marbled Murrelets
relative to moss-cover thickness is highly dichotomous in a sense that platforms (trees) with
moss cover below 2 cm were rarely used. On the other hand those above this value were used
proportionately to their availability. This implies that while the nest structure condition is clearly
important and selected for in this species, it was probably relatively uniform, and abundant
relative to demand, across our study population and thus did not contribute to breeding success.
Avalanche chutes can be described as vertical non-forest strips of meadow, rocky ground
and shrubs with high-contrast edges functioning in a manner similar to streams (Suffling 1993,
Butler 2001). Although the parameter estimate for the distance to this feature was not
significant, this variable was consistently selected as contributing important information to the
best approximating model. Murrelet nests were more successful closer to avalanche chutes but
the birds did not seem to select for this feature. What can explain this contradiction?
Our coverage of avalanche chute features was obtained from an LUP mapping product
(MOF). This database combines avalanche chutes proper as defined by Suffling (1993) and
Butler (2001) (i.e. vertical features relative to a mountain slope) with the higher elevation

33
subalpine areas into one land cover class presumably due to their similar vegetation
characteristics. Higher elevations per se were selected against. If the murrelets actually selected
for avalanche chutes proper (e.g. Bradley 2002), then their avoidance of higher elevation
subalpine areas will cancel these trends out when selection relative to a coverage combining both
features is measured. This seems to be exactly what was observed (Table 1). If in fact the
murrelets do select for avalanche chutes and breed there more successfully, this may be
explained by their providing access routes to potential nesting sites as in the case of stream and
on the other by highly disturbed or even absent vegetation cover which may be avoided by
potential nest predators.
CONCLUSIONS
Our results show that in SW Canada, nesting Marbled Murrelets do not select disproportionately
large fragments of old-growth forest in a landscape, but either show no selection at all or prefer
smaller stands of forest relatively to what is available. However, as suggested by the
multivariate analyses, it is not the size of a forest fragment per se that is important to a nesting
murrelet but the proximity of a nesting site (tree) to an abrupt natural or artificial edge. Edges
created by streams appear to be particularly important. Also, the murrelet nesting success
appeared greater in environments with numerous high-contrast edges. Garman et al. (1999)
stated that in parts of the Pacific Northwest region of North America, present practices of old-
growth habitat management for wildlife may largely limit late-successional forests to riparian
corridors with little interior habitat. While this practice may not be optimal for some other
“umbrella” species typically associated with increased amounts of interior old-growth forest, e.g.

34
the Spotted Owl (Strix occidentalis) (Franklin and Gutiérrez 2002), it may, if no alternative is
possible, still provide for suitable habitat for the Marbled Murrelet. This management approach,
however, will only work in the long run if forest interior conditions essential for the maintenance
of epiphytic cover can be maintained within the remaining riparian buffers (Hylander et al.
2002). While the exact buffer width necessary to maintain such conditions will vary depending
on local topography and microclimate (Chen et al. 1999), it will probably need to be at least 100
m on either side of a stream course. The present forestry guide-lines on the other hand allow for
only a 90 m or less buffer-strip (Garman et al. 1999 and references therein). Whether such a
proposition can be accommodated in real terms needs to be determined.
ACKNOWLEDGEMENTS
The project was funded and/or supported logistically by the Forest Renewal BC (FRBC), Natural
Sciences and Engineering Research Council of Canada (NSERC), Canadian Wildlife Service
(CWS), Simon Fraser University, The British Columbia (B.C.) Ministry of Forests,
Weyerhaeuser Ltd., TimberWest Forest Ltd., International Forest Products Ltd., Western Forest
Products Ltd., Terminal Forest Products, and NCASI (National Council of the Paper Industry for
Air and Stream Improvement, Inc.). Interpretation of the findings in this paper does not
necessarily reflect the opinions of the above organizations. We acknowledge the hard and
diligent work of field crews, and the collaboration with the Centre of Wildlife Ecology, Simon
Fraser University, and CWS staff. E&B Helicopters provided the helicopter support. We would
like to thank the B.C. Ministry of Forests, Weyerhaeuser Ltd., Interfor Ltd., B.C. Ministry of

35
Environment, Lands and Parks, L. Waterhouse and J. Sundé for providing GIS data sets. M.
Stephens kindly developed and tested Monte Carlo models for patch size analyses. Mryka Hall-
Beyer and S. Steeby suggested and produced clear-cut classification.
LITERATURE CITED
ALABACK, P. AND J. PODAR. 1998. Vegetation from ridgetop to seashore. Pages 69-87 in The
Rainforest of Home. (P. K. Schoonmkaer, B. v. Hagen, and E. C. Wolf, Eds.). Island Press,
Washington.
ANDERSON, D. R. 2001. The need to get the basics right in wildlife field studies. Wildlife Society
Bulletin 29:1294-1297.
ANDRÉN, H. 1995. Effects of landscape composition on predation rates at habitat edges. Pages
225-255 in Mosaic Landscapes and Ecological Process. (L. Hansson, L. Fahrig, and G. Merriam,
Eds.). Chapman and Hall, London.
BAHN, V. 1998. Habitat requirements and habitat suitability index for the Marbled Murrelet
(Brachyramphus marmoratus) as a management target species in the Ursus Valley, British
Columbia. Diploma Thesis. Philips-University. Marburg.
BAYNE, E. M. AND K. A. HOBSON. 2002. Apparent survival of male ovenbirds in fragmented and
forested boreal landscapes. Ecology 83:1307-1316.

36
BOAL, C. W. AND R. W. MANNAN. 1998. Nest-site selection by Cooper's hawks in an urban
environment. Journal of Wildlife Management 62:864-871.
BOOTH, D. E. 1991. Estimating Prelogging Old-Growth in the Pacific-Northwest. Journal of
Forestry 89:25-29.
BRADLEY, R. W. 2002. Breeding ecology of radio-marked marbled murrelets (Brachyramphus
marmoratus) in Desolation Sound, British Columbia. M.S. thesis, Simon Fraser University,
Burnaby.
BRADLEY, R. W., L. A. MCFARLANE TRANQUILLA, B. A. VANDERKIST, AND F. COOKE. 2002. Sex
differences in nest visitation by chick-rearing Marbled Murrelets. Condor 104:178-183.
BRINKHOF, M. W. G. AND A. J. CAVE. 1997. Food supply and seasonal variation in breeding
success: An experiment in the European coot. Proceedings of the Royal Society, London B
264:291-296.
BROSOFSKE, K. D., J. CHEN, R. J. NAIMAN, AND J. F. FRANKLIN. 1997. Harvesting effects on
microclimatic gradients from small streams to uplands in western Washington. Ecological
Applications 7:1188-1200.
BURGER, A. E. 2002a. Radar Inventory and Watershed-level Habitat Association of Marbled
Murrelets in Clayoquot Sound, 1996-1998. Pages 121-137 in Multi-Scale Studies of Populations,
Distribution and Habitat Associations of Marbled Murrelets in Clayoquot Sound, British
Columbia (A. E. Burger and T. A. Chatwin, Eds.). Ministry of Water, Land and Air Protection,
Victoria, B.C.

37
BURGER, A. E. 2002b. Conservation assessment of Marbled Murrelets in British Columbia: a
review of the biology, populations, habitat associations, and conservation. Technical Report
Series No. 387. Canadian Wildlife Service, Pacific and Yukon Region, British Columbia.
BURGER, A. E., V. BAHN, AND A. R. M. TILLMANNS. 2000. Comparison of coastal fringe and
interior forests as reserves for Marbled Murrelets on Vancouver Island. Condor 102:915-920.
BURNHAM, K. P. AND D. R. ANDERSON. 2002. Model Selection and Multimodel Inference: A
Practical Information-Theoretic Approach. Springer-Verlag, New York.
BUTLER, D. R. 2001. Geomorphic process-disturbance corridors: a variation on a principle of
landscape ecology. Progress in Physical Geography 25:237-248.
CAM, E., L. LOUGHEED, R. W. BRADLEY, AND F. COOKE. In press. Demographic assessment of a
Marbled Murrelet population from capture-mark-recapture and radio telemetry data.
Conservation Biology 00:000-000.
CARTER, H. R. AND S. G. SEALY. 1987. Fish-Holding Behavior of Marbled Murrelets. Wilson
Bulletin 99:289-291.
CHEN, J., J. F. FRANKLIN, AND T. A. SPIES. 1995. Growing-season microclimatic gradients from
clearcut edges into old-growth Douglas-fir forests. Ecological Applications 5:74-86.
CHEN, J., S. C. SAUNDERS, T. R. CROW, R. J. NAIMAN, K. D. BROSOFKE, G. D. MROZ, B. L.
BROOKSHIRE, AND J. F. FRANKLIN. 1999. Microclimate in forest ecosystems and landscape
ecology. BioScience 49:288-297.

38
CHOULAKIAN, V., R. A. LOCKHART, AND M. A. STEPHENS. 1994. Cramér-von-Mises statistics for
discrete distributions. Canadian Journal of Statistics 22:125-137.
CLEAVITT, N. L. 2002. Stress tolerance of rare and common moss species in relation to their
occupied environments and asexual dispersal potential. Journal of Ecology 90:785-795.
CONNER, L. M., M. D. SMITH, AND L. W. BURGER. 2003. A comparison of distance-based and
classification-based analyses of habitat use. Ecology 84:526-531.
CRAMER, J. S. 1999. Predictive performance of the binary logit model in unbalanced samples.
Statistician 48:85-94.
DONALD, P. F. AND J. A. VICKERY. 2000. The importance of cereal fields for breeding and
wintering skylarks Alauda arvensis in the UK. Pages 140-150 in Ecology and Conservation of
Farmland Birds (N. J. Aebischer, P. V. Grice, A. D. Evans, and J. A. Vickery, Eds.). BOU,
Tring.
DONAZAR, J. A., F. HIRALDO, AND J. BUSTAMANTE. 1993. Factors influencing nest site selection,
breeding density and breeding success in the bearded vulture (Gypaetus barbatus). Journal of
Applied Ecology 30:504-514.
ERICKSON, W. P., T. L. MCDONALD, AND R. SKINNER. 1998. Habitat selection using GIS: a case
study. Journal of Agricultural, Biological and Environmental Statistics 3:296-310.
ESSEEN, P.-A. 1994. Tree mortality patterns after experimental fragmentation of an old-growth
conifer forest. Biological Conservation 68:19-28.

39
ETHIER, T. J. 1999. Breeding ecology and habitat of Northern Goshawks (Accipiter gentilis
laingi) on Vancouver Island: a hierarchical approach. M.S. thesis, University of Victoria,
Victoria.
FAHRIG, L. 1999. Forest loss and fragmentation: which has the greater effect on persistence of
forest dwelling animals? Pages 87-97 in Forest Fragmentation: Implications for Wildlife
Management (J. A. Rochelle, L. A. Lehmann, and J. Wisniewski, Eds.). Brill Academic
Publishers, Leiden.
FEMAT. 1993. Forest ecosystem management: An ecological, economic and social assessment.
Report of the forest ecosystem management assessment team. July 1993, Portland, OR, USA.
FIELDING, A. H. AND P. F. HAWORTH. 1995. Testing the generality of bird-habitat models.
Conservation Biology 9:1466-1481.
FIELDING, A. H. AND J. F. BELL. 1997. A review of methods for the assessment of prediction
errors in conservation presence absence models. Environmental Conservation 24:38-49.
FRANKLIN, A. B. AND R. J. GUTIÉRREZ. 2002. Spotted Owls, forest fragmentation and forest
heterogeneity. Studies of Avian Biology 25:203-220.
GARMAN, S. L., F. J. SWANSON, AND T. A. SPIES. 1999. Past, present, and future landscape
patterns in the Douglas-fir region of the Pacific Northwest. Pages 61-86 in Forest Fragmentation:
Implications for Wildlife Management (J. A. Rochelle, L. A. Lehmann, and J. Wisniewski, Eds.).
Brill Academic Publishers, Leiden.

40
GASTON, A. J. 1992. The Ancient Murrelet: A Natural History in the Queens Charlotte Islands. T.
and A.D. Poyser Ltd., London.
GATES, J. E. AND D. R. EVANS. 1998. Cowbirds breeding in the central Appalachians: Spatial and
temporal patterns and habitat selection. Ecological Applications 8:27-40.
GATES S., D. WINGFIELD-GIBBONS, P. C. LACK, AND R. J. FULLER. 1994. Declining farmland bird
species: modelling geographical patterns of abundance in Britain. Pages 153-178 in Large-scale
Ecology and Conservation Biology (P. J. Edwards, R. M. May, and N. R. Webb, Eds.).
Blackwell Scientific Publications, Oxford.
GREEN, R. A. AND K. KLINKA. 1994. A field guide to site identification and interpretation for the
Vancouver Forest Region. Land Management Handbook No. 28. Research Branch, Ministry of
Forests, Victoria, B.C.
HAMER, T. E. AND S. K. NELSON. 1995. Characteristics of Marbled Murrelet nest trees and
nesting stands. Pages 69-82 in Ecology and conservation of the Marbled Murrelet (C. J. Ralph,
G. L. Hunt, Jr., M. G. Raphael, and J. F. Piatt, Eds.). Pacific Southwest Research Station, Forest
Service, U.S. Department of Agriculture, Albany, California.
HARRIS, M. P. S. WANLESS, T. R. BARTON, AND D. A. ELSTON. 1997. Nest site characteristics,
duration of use and breeding success in the Guillemot Uria aalge. Ibis 139:468-476.
HARTMAN, L. H., A. J. GASTON, AND D. S. EASTMAN. 1997. Raccoon predation on ancient
murrelets on East Limestone Island, British Columbia. Journal of Wildlife Management 61:377-
388.

41
HATCHWELL, B. J. 1991. An experimental study of the effects of timing of breeding on the
reproductive success of Common Guillemots (Uria aalge). Journal of Animal Ecology 60:721-
736.
HINSLEY, S. A., P. ROTHERY, AND P. E. BELLAMY. 1999. Influence of woodland area on breeding
success in Great Tits Parus major and Blue Tits Parus caeruleus. Journal of Avian Biology
30:271-281.
HIPFNER, J. M. A. J. GASTON, D. L. MARTIN, AND I. L. JONES. 1999. Seasonal declines in
replacement egg-layings in a long-lived Arctic seabird: Costs of late breeding or variation in
female quality? Journal of Animal Ecology 68:988-998.
HIPFNER, J. M. AND A. J. GASTON. 2002. Growth of Thick-billed Murre (Uria lomvia) chicks in
relation to parental experience and hatching date. Auk 119:827-832.
HOFMANN, P. 2001. Detecting urban features from IKONOS data using an object-oriented
approach. Pages 28-33 in Proceedings of the First Annual Conference of the Remote Sensing and
Photogrammetry Society (Remote Sensing and Photogrammetry Society, Eds.).
HYLANDER, K. B., G. JONSSON, AND C. NILSSON. 2002. Evaluating buffer strips along boreal
streams using bryophytes as indicators. Ecological Applications 12:797-806.
IDENTIFIED WILDLIFE MANAGEMENT STRATEGY. 1999. Ministry of Water, Land and Air
Protection. Victoria, B.C. http://wlapwww.gov.bc.ca/wld/identified/index.htm

42
JODICE, P. G. R. AND M. W. COLLOPY. 2000. Activity patterns of Marbled Murrelets in Douglas-
fir Old-Growth Forests of the Oregon coast range. Condor 102:275-285.
JONES, J. 2001a. Habitat selection studies in Avian Ecology: A critical review. Auk 118:557-562.
JONES, P. H. 1993. Canada's first active Marbled Murrelet nest, Caren Range, Sechelt Peninsula,
BC. Discovery 22:147-150.
JONES, P. H. 2001b. The Marbled Murrelets of the Caren Range and Middlepoint Bight. Western
Canada Wilderness Committee, Vancouver, B.C.
JONSSON, B. G. 1997. Riparian bryophyte vegetation in the Cascade mountain range, Northwest
USA: Patterns at different spatial scales. Canadian Journal of Botany 75:744-761.
KAISER, G. W., T. E. MAHON, AND M. D. FAWCETT. 1991. Studies of Marbled Murrelets in
marine habitats, during 1990. Technical Report Series No. 131, Canadian Wildlife Service,
Pacific and Yukon Region, Delta, B.C.
KELSON, J. D., I. A. MANLEY, AND H. R. CARTER. 1995. Decline of the Marbled Murrelet in
Clayoquot Sound, British Columbia: 1982-1993. Northwestern Naturalist 76:90-98.
KITAYSKY, A. S. AND E. G. GOLUBOVA. 2000. Climate change causes contrasting trends in
reproductive performance of planktivorous and piscivorous alcids. Journal of Animal Ecology
69:248-262.
KREMSATER, L. AND F. L. BUNNELL. 1999. Edge effects: theory, evidence and implications to
management of western North American forests. Pages 117-153 in Forest Fragmentation:

43
Implications for Wildlife Management (J. A. Rochelle, L. A. Lehmann, and J. Wisniewski, Eds.).
Brill Academic Publishers, Leiden.
LAHTI, D. C. 2001. The "edge effect on nest predation" hypothesis after twenty years. Biological
Conservation 99:365-374.
MCGARIGAL, K. AND W. C. MCCOMB. 1999. Forest fragmentation effects on breeding bird
communities in the Oregon Coast range. Pages 223-246 in Forest Fragmentation: Implications
for Wildlife Management (J. A. Rochelle, L. A. Lehmann, and J. Wisniewski, Eds.). Brill
Academic Publishers, Leiden.
MANLEY, I. A. 1999. Behaviour and habitat selection of Marbled Murrelets nesting on the
Sunshine Coast. M.S. thesis, Simon Fraser University, Burnaby.
MANLEY, I. A., F. L. WATERHOUSE, AND A. S. HARESTAD. 1999. Nesting habitat of Marbled
Murrelets on the Sunshine Coast. Forest Service, British Columbia. Forest Research Extension
Note, EN-002 Wildlife. Nanaimo.
MANLY, B. F. J. 1990. Randomization and Monte Carlo Methods in Biology. Chapman and Hall,
London.
MANLY, B. F. L., L. L. MCDONALD, D. L. THOMAS, T. L. MCDONALD, AND W. P. ERICKSON.
2002. Resource Selection by Animals: Statistical Design and Analysis of Field Studies. Kluwer
Academic Publishers, Dordrecht.

44
MARZLUFF, J. M. AND M. RESTANI. 1999. The effects of forest fragmentation on avian nest
predation. Pages 155-169 in Forest Fragmentation: Implications for Wildlife Management (J. A.
Rochelle, L. A. Lehmann and J. Wisniewski, Eds.). Brill Academic Publishers, Leiden.
MATHER, M. H. AND T. A. CHATWIN. 2001. Marbled Murrelet nesting habitat: an evaluation of
Identified Wildlife Management Strategy (IWMS) and Landscape Unit Planning Guide (LUPG)
policy for protecting required old growth nesting habitat in selected Vancouver Island landscape
units. Unpublished report to Ministry of Environment, Lands and Parks, Nanaimo.
MEYER, C. B. AND S. L. MILLER. 2002. Use of fragmented landscapes by marbled murrelets for
nesting in Southern Oregon. Conservation Biology 16:755-766.
MEYER, C. B., S. L. MILLER, AND C. J. RALPH. 2002. Multi-scale landscape and seascape patterns
associated with marbled murrelet nesting areas on the U.S. west coast. Landscape Ecology
17:95-115.
MILLER, S. L., C. B. MEYER, AND C. J. RALPH. 2002. Land and seascape patterns associated with
marbled murrelet abundance offshore. Waterbirds 25:100-108.
MIZUNO, K. 1998. Succession processes of alpine vegetation in response to glacial fluctuations
of Tyndall Glacier, Mt. Kenya, Kenya. Arctic and Alpine Research 30:340-348.
MORRIS, A. J., M. J. WHITTINGHAM, R. B. BRADBURY, J. D. WILSON, A. KYRKOS, D. L.
BUCKINGHAM, AND A. D. EVANS. 2001. Foraging habitat selection by yellowhammers (Emberiza
citrinella) nesting in agriculturally contrasting regions of lowland England. Biological
Conservation 101:197-210.

45
MURCIA, C. 1995. Edge Effects in Fragmented Forests - Implications for Conservation. Trends in
Ecology and Evolution 10:58-62.
NASLUND, N. L., K. J. KULETZ, M. B. CODY, AND D. K. MARKS. 1995. Tree and habitat
characteristics and reproductive success at Marbled Murrelet tree nests in Alaska. Northwestern
Naturalist 76:12-25.
NELSON, S. K. 1997. Marbled Murrelet (Brachyramphus marmoratus). In The Birds of North
America, no. 276 (A. Poole and F. Gill, Eds.). Academy of Natural Sciences, Philadelphia, and
American Ornithologists' Union, Washington, D.C.
NELSON, S. K., T. E. HAMER, A. K. WILSON, AND D. J. MEEKINS. 2003. Characteristics of
Marbled Murrelet nest trees and nest sites in the Pacific Northwest. Proceedings of the 30th
Annual Pacific Seabird Group Meeting.
NETTLESHIP, D. N. AND T. R. BIRKHEAD. 1985. The Atlantic Alcidae. The Evolution, Distribution
and Biology of the Auks Inhabiting the Atlantic Ocean and Adjacent Water Areas. Academic
Press, San Diego.
NEWTON, I. AND M. MARQUISS. 1984. Seasonal trend in the breeding performance of
Sparrowhawks. Journal of Animal Ecology 53:809-829.
NILSSON, S., R. COLBERG, R. HAGLER, AND P. WOODBRIDGE. 1999. How sustainable are North
American Wood supplies? Interim Report IR-99-003/January. International Institute for Applied
Systems Analysis, Laxenburg, Austria.

46
PECK, J. E. AND P. S. MUIR. 2001. Estimating the biomass of harvestable epiphytic moss in
central western Oregon. Northwest Science 75:99-106.
PENNYCUICK, C. J. 1987. Flight of Auks (Alcidae) and Other Northern Seabirds Compared with
Southern Procellariiformes Ornithodolite Observations. Journal of Experimental Biology
128:335-347.
RALPH, C. J., G. L. HUNT, JR., M. G. RAPHAEL, AND J. F. PIATT. 1995. Ecology and conservation
of the Marbled Murrelet. Pacific Southwest Research Station, Forest Service, Albany, California,
U.S. Department of Agriculture, USA.
RAPHAEL, M., D. E. MACK, J. M. MARZLUFF, AND J. M. LUGINBUHL. 2002a. Effects of forest
fragmentation on populations of the Marbled Murrelet. Studies of Avian Biology 25:221-235.
RAPHAEL, M. G., D. E. MACK, AND B. A. COOPER. 2002b. Landscape-scale relationships between
abundance of marbled murrelets and distribution of nesting habitat. Condor 104:331-342.
RENHORN, K.-E., P.-A. ESSEEN, K. PALMQVIST, AND B. SUNDBERG. 1997. Growth and vitality of
epiphytic lichens .1. Responses to microclimate along a forest edge-interior gradient. Oecologia
109:1-9.
RIC (Resources Inventory Committee). 2001. Inventory methods for Marbled Murrelets in
marine and terrestrial habitats, Version 2.0. Standards for components of British Columbia’s
biodiversity, No. 10. Ministry of Environment, Lands and Parks, Resources Inventory
Committee, Victoria, B.C.

47
RIPPLE, W. J. 1994. Historic spatial patterns of old forests in western Oregon. Journal of Forestry
92:45-49.
RODWAY, M. S. AND H. M. REGEHR. 2000. Measuring Marbled Murrelets activity levels in valley
bottom habitat: bias due to station placement on stream channel flight corridors. Journal of Field
Ornithology 71:415-422.
RODWAY, M. S. AND H. M. REGEHR. 2002. Inland activity and forest structural characteristics as
indicators of Marbled Murrelet nesting habitat in Clayoquot Sound. Pages 59-90 in Multi-scale
studies of populations, distribution and habitat associations of Marbled Murrelets in Clayoquot
Sound, British Columbia (A. E. Burger and T. A. Chatwin, Eds.). B.C. Ministry of Water, Land
and Air Protection. Victoria, B.C.
SILLETT, S. C., B. MCCUNE, J. E. PECK, T. R. RAMBO, AND A. RUTCHY. 2000. Dispersal
limitations of epiphytic lichens result in species dependent on old-growth forests. Ecological
Applications 10:789-799.
SINGER, S. W., N. L. NASLUND, S. A. SINGER, AND C. J. RALPH. 1991. Discovery and observation
of two tree nests of the Marbled Murrelet. Condor 93:330-339.
SONG, S. J. AND S. J. HANNON. 1999. Predation in heterogeneous forests: a comparison at natural
and anthropogenic edges. Ecoscience 6:521-530.
SPEAR, L. B. AND D. G. AINLEY. 1997. Flight behaviour of seabirds in relation to wind direction
and wing morphology. Ibis 139:221-233.

48
SUÁREZ, S., J. BALBONTIN, AND M. FERRER. 2000. Nesting habitat selection by booted eagles
Hieraaetus pennatus and implications for management. Journal of Applied Ecology 37: 215-223.
SUFFLING, R. 1993. Induction of vertical zones in sub-alpine valley forests by avalanche-formed
fuel breaks. Landscape Ecology 8:127-138.
THOMAS, S. C., D. A. LIGUORI, AND C. B. HALPERN. 2001. Corticolous bryophytes in managed
Douglas-fir forests: habitat differentiation and responses to thinning and fertilization. Canadian
Journal of Botany 79:886-896.
VAN HORN, B. 1983. Density as a misleading indicator of habitat quality. Journal of Wildlife
Management 48:893-901.
VERHULST, S., J. H. VAN BALEN, AND J. M. TINGERGEN. 1995. Seasonal decline in reproductive
success of the Great Tit: variation in time or quality? Ecology 76:2392-2403.
VERMEER, K. AND L. CULLEN. 1979. Growth of Rhinoceros Auklets and Tufted Puffins, Triangle
Island, British Columbia. Ardea 67:22-27.
VICKERY, P. D., M. L. HUNTER AND J. V. WELLS. 1992. Evidence of incidental nest predation and
its effects on nests of threatened grassland birds. Oikos 63:281-288.
U.S. FISH AND WILDLIFE SERVICE. 1997. Recovery plan for the threatened Marbled Murrelet
(Brachyramphus marmoratus) in Washington, Oregon, and California. U.S. Fish and Wildlife
Service Region 1, Portland, OR.

49
WASER, L. AND M. SCHWARTZ. 2001. Using eCognition to detect storm losses in Swiss alpine
forest areas from high resolution satellite data. eCognition Application Note 2:11.
WATERHOUSE, F. L., R. W. BRADLEY, J. MARKILA, F. COOKE, AND L. LOUGHEED. 2002. Use of
airphotos to identify, describe, and manage forest structure of Marbled Murrelet nesting habitat
at a coastal British Columbia site. Forest Service, British Columbia. Forest Research Technical
Report, TR-016 Wildlife. Nanaimo.
WHITWORTH, D. R., J. Y. TAKEKAWA, H.R. CARTER, AND W.R. MCIVER. 1997. A night-technique
for at-sea capture of Xantus' Murrelets. Colonial Waterbirds 20:525-531.

50
Table 1. Characteristics (mean±SD) of Marbled Murrelet nest sites and random sites within the oldgrowth (>140 yr)
stratum in Desolation Sound. Statistics are for univariate logistic models. Nest sites, n=121 (n=86 for patch area
and patch edge distance); random sites, n=1000.
VARIABLE NEST SITE RANDOM SITE β±SE P
1. Elevation Indexa, m 700±345 880±384 -0.30±0.11 0.007
2. Slope Index, degreea 39.0±15.6 33.3±12.7 0.03±0.01 <0.001
3. Patch Areaa, ha 161.2±278.8 161.2±219.1 -0.15±0.07 0.040
4. Patch Edge Distancea 46.2±51.0 63.3±63.0 -0.30±0.09 0.002
5. Distance to FEb Clear-cut 486±473 590±648 -0.01±0.04 0.74
6. Mean of three nearest
FE Clear-cut Distances 780±520 912±707 -0.14±0.11 0.18
7. Distance to HEb Clear-cut 1451±1031 1580±1381 -0.13±0.06 0.027
8. Mean of three nearest
HE Clear-cut Distancesa 1897±1342 2357±1463 -0.50±0.13 <0.001
9. Distance to Roada 1406±1603 1739±1641 -0.32±0.09 <0.001
10. Distance to Glaciera 6491±5330 5602±4763 0.27±0.12 0.021
11. Distance to Lake 3522±1968 3494±2131 0.08±0.12 0.41
12. Distance to Ocean 8769±6451 8392±6774 0.07±0.08 0.38
13. Distance to Rocky Areaa 605±830 746±850 -0.18±0.05 <0.001
14. Distance to Streama 135±139 176±178 -0.46±0.01 <0.001
15. Mean of three nearest
Stream Distances 240±194 271±200 -0.48±0.14 0.001
16. Distance to Subalpine Area 1625±2691 1543±2584 0.003±0.04 0.93
aVariables entered into a multivariate logistic regression. bFE=Fuzzy-edge, HE=Hard-edge

51
Table 2. Characteristics (mean±SD) of Marbled Murrelet nest sites and random sites within the oldgrowth (>250 yr,
>15 m) stratum in Clayoquot Sound. Statistics are for univariate logistic models. Nest sites, n=36 (n=33 for patch
area and patch edge distance), random sites, n=1000.
VARIABLE NEST SITE RANDOM SITE β±SE P
1. Elevation Index, m 565±334 609±343 -0.08±0.17 0.63
2. Slope Index, degree 31.3±11.4 29.2±11.9 0.02±0.01 0.28
3. Patch Area, ha 489.8±465.7 405.7±426.7 0.18±0.12 0.13
4. Patch Edge Distancea 76.1±64.0 86.1±76.2 -0.22±0.14 0.11
5. Distance to FEb Clear-cut 885±885 934±993 -0.004±0.084 0.96
6. Mean of three nearest
FE Clear-cut Distances 1109±993 1119±1017 0.02±0.15 0.88
7. Distance to HEb Clear-cut 2202±1643 2496±1687 -0.13±0.12 0.31
8. Mean of three nearest
HE Clear-cut Distances 2457±1671 2789±1698 -0.18±0.19 0.34
9. Distance to Road 4027±3294 4658±3128 -0.15±0.14 0.28
10. Distance to Glaciera 13796±8658 10781±8057 0.50±0.22 0.016
11. Distance to Lake 4336±2420 4085±2457 0.14±0.21 0.49
12. Distance to Ocean 6143±8053 9380±8536 -0.17±0.09 0.086
13. Distance to Rocky Areaa 5051±2694 4101±2966 0.51±0.22 0.009
14. Distance to Streama 119±158 142±101 -0.49±0.18 0.008
15. Mean of three nearest
Stream Distances 219±188 237±120 -0.47±0.27 0.085
16. Distance to Subalpine Areaa 5450±6272 3582±5030 0.20±0.11 0.043
aVariables entered into a multivariate logistic regression. bFE=Fuzzy-edge, HE=Hard-edge

52
Table 3. AIC and log-likelihood ratio chi-square values, for the top five models ranked in the
order of AIC for Desolation Sound. Models judged as “best” based on the number of parameters
and the AIC value given in bold.
MODEL with 11 input variables np AIC LL Ratio X2
1. Elev+Elev2+Slope2+Road+MNHEa+Stream+Glacier 7 554.4 57.67
2. Elev+Elev2+Slope2+PEDa+Road+MNHE+Stream+Glacier 8 554.8 59.27
3. Elev+Slope+Slope2+Road+MNHE+Stream+Glacier 7 555.0 57.14
4. Elev+Elev2+Slope2+PED+Road+Stream+Glacier 7 555.2 56.94
5. Elev+Elev2+Slope2+Road+Stream 5 555.2 52.87
MODEL with 9 input variables
1. Elev+Slope+Elev2+Slope2+MNHE+Stream+Glacier+Rock 8 674.9 110.3
2. Elev+Elev2+Slope2+MNHE+Stream+Glacier+Rock 7 675.9 107.3
3. Elev+Slope+Elev2+Slope2+Road+MNHE+Str.+Glacier+Rock 9 676.1 111.1
4. Elev+Slope+Elev2+Slope2+MNHE+Stream+Glacier+Rock 8 677.0 110.3
5. Elev+Elev2+Slope2+Rd+MNHE+Stream+Glacier+Rock 8 677.9 108.1
aMNHE = Mean of three nearest Hard-edge Clear-cut Distances
bPED = Patch edge distance

53
Table 4. Parameters of best approximating models presented in Table 3.
TERM ESTIMATE SE X2 P
MODEL 1 (X27,1085=25.9, P<0.001, multiple R2=0.097, Kappa=0.00)
Intercept 7.65 2.36 10.56 0.0012
Elevation Index -1.07 0.23 21.09 <0.0001
(Elevation Index)2 -0.36 0.13 7.87 0.0050
(Slope Index)2 0.0006 0.0001 29.73 <0.0001
Mean of three hard-edge clear-cuts -0.51 0.14 12.63 0.0004
Stream -0.53 0.11 23.49 <0.0001
Glacier 0.39 0.15 6.91 0.0086
Rock -0.12 0.05 5.41 0.0200
MODEL 2 (X27,1121=107.3, P<0.001, multiple R2=0.140, Kappa=0.126)
Intercept 6.76 2.70 6.27 0.0123
Elevation Index -1.02 0.29 12.27 0.0005
(Elevation Index)2 -0.49 0.20 5.94 0.0148
(Slope Index)2 0.0005 0.0001 16.08 <0.0001
Road -0.21 0.13 2.81 0.0934
Mean of three hard-edge clear-cuts -0.29 0.17 2.86 0.0910
Stream -0.40 0.13 10.35 0.0013
Glacier 0.29 0.17 2.82 0.0932

54
Table 5. AIC and log-likelihood ratio chi-square values, for the top five models ranked in the
order of AIC and parameters of the best approximating model for Clayoquot Sound. Models
judged as “best” based on the number of parameters and the AIC value given in bold.
MODEL np AIC LL Ratio X2
1. Stream+Rock 2 302.9 15.8
2. Stream+Subalpine+Rock 3 303.7 16.90
3. Stream+Glacier+Rock 3 304.1 16.55
4. PED+Stream+Rock 3 304.1 16.54
5. Stream+Glacier 2 305.0 13.64
Best approximating model parameters
(X22,1036=15.8, P<0.001, multiple R2=0.050, Kappa=0.00)
TERM ESTIMATE SE X2 P
Intercept -5.74 1.96 8.57 0.0034
Stream -0.55 0.18 9.60 0.0019
Rock 0.60 0.23 6.80 0.0091

55
Table 6. Characteristics (mean±SD) of successful and failed Marbled Murrelet nests in Desolation Sound. Statistics
refer to univariate logistic regressions. Sample size: successful nests, n=70, failed nests n=37.
VARIABLE SUCCESSFUL FAILED β±SE P
Julian Date (1=1st April)a 53.4±13.1 64.0±12.2 -0.07±0.02 <0.001
Elevation Indexa, m 742±319 611±340 0.86±0.38 0.021
Slope Indexa, degree 41.4±15.0 34.7±15.1 0.03±0.01 0.031
Patch Areaa, ha 153.3±279.8 220.0±318.9 -0.27±0.14 0.054
Patch Edge Distance 41.9±46.2 56.8±64.4 -0.22±0.18 0.21
Distance to FEb Clear-cut 501±495 455±481 0.08±0.09 0.37
Mean of three nearest
FE Clear-cut Distances 809±543 736±530 0.17±0.23 0.47
Distance to HEb Clear-cut 1022±935 1379±1142 -0.14±0.15 0.33
Mean of three nearest
HE Clear-cut Distancesa 1718±1218 2305±1579 -0.46±0.29 0.11
Distance to Road 1468±1719 1182±1258 0.13±0.19 0.50
Distance to Glacier 5863±4277 7945±7023 -0.21±0.26 0.42
Distance to Lake 3495±1867 3616±2237 0.01±0.30 0.98
Distance to Oceana 9457±6518 6937±5915 0.39±0.19 0.037
Distance to Rocky Areaa 501±675 946±1098 -0.15±0.10 0.11
Distance to Stream 127±104 128±165 0.19±0.20 0.39
Distance to Subalpine Areaa 1207±1955 2773±3809 -0.17±0.10 0.067
aVariables entered into a multivariate logistic regression. bFE=Fuzzy-edge, HE=Hard-edge
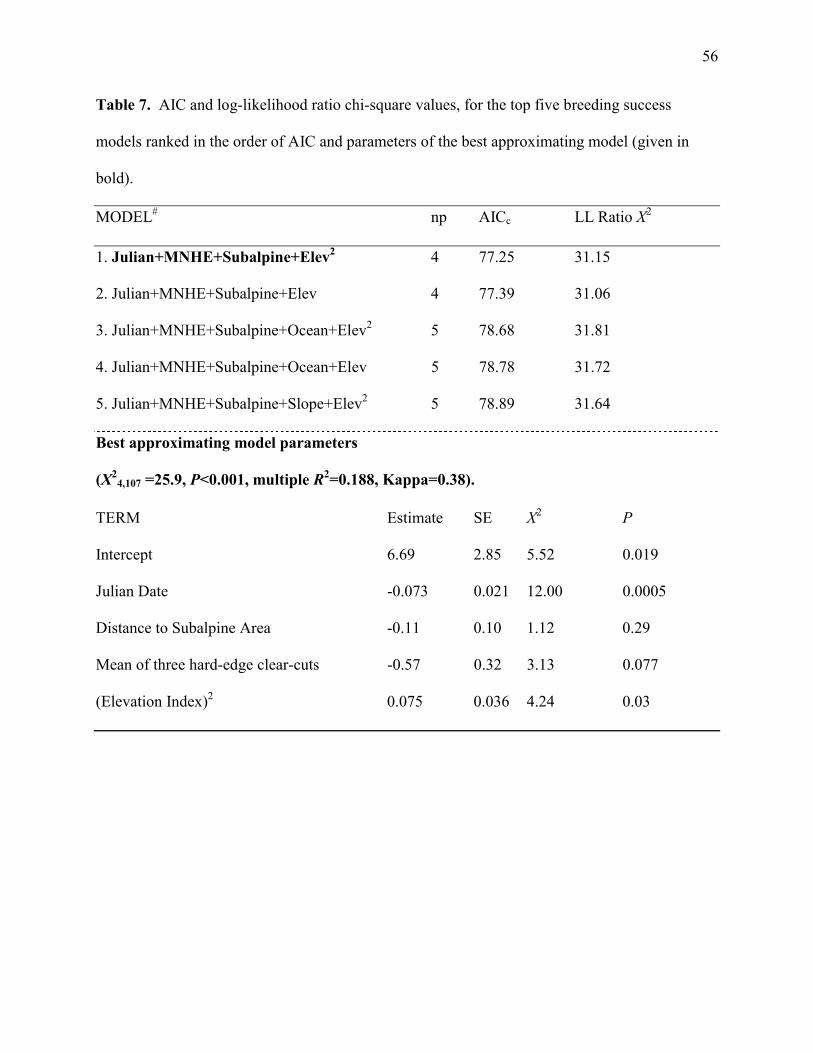
56
Table 7. AIC and log-likelihood ratio chi-square values, for the top five breeding success
models ranked in the order of AIC and parameters of the best approximating model (given in
bold).
MODEL# np AICc LL Ratio X2
1. Julian+MNHE+Subalpine+Elev2 4 77.25 31.15
2. Julian+MNHE+Subalpine+Elev 4 77.39 31.06
3. Julian+MNHE+Subalpine+Ocean+Elev2 5 78.68 31.81
4. Julian+MNHE+Subalpine+Ocean+Elev 5 78.78 31.72
5. Julian+MNHE+Subalpine+Slope+Elev2 5 78.89 31.64
Best approximating model parameters
(X24,107 =25.9, P<0.001, multiple R2=0.188, Kappa=0.38).
TERM Estimate SE X2 P
Intercept 6.69 2.85 5.52 0.019
Julian Date -0.073 0.021 12.00 0.0005
Distance to Subalpine Area -0.11 0.10 1.12 0.29
Mean of three hard-edge clear-cuts -0.57 0.32 3.13 0.077
(Elevation Index)2 0.075 0.036 4.24 0.03

57
Fig. 1. Overall location of study areas in relation to the western Canada coastline (a) and
distribution of old-growth forest cover (grey shade) and nest locations (black dots) within the
buffered minimum convex polygons (black line) for Desolation Sound (b) and Clayoquot Sound
(c).
Fig. 2. Cumulative plots of area under old-growth forest cover and the number of
Marbled Murrelet nests encompassed. The linear trend represents a neutral selection for
fragment size (pi = cαi) where c estimated as N/A the slope represents (see Methods). Boxes
represent the approximate individual fragment sizes which show the greatest deviation from the
neutral trend.
Fig. 3. Percent frequency distribution of nest (black) and random (grey) forest fragments
for each study area.
Fig. 4. Percent frequency distribution of distance to edge values for Marbled Murrelet
nests (black) and random points (grey) for each study area.


















