
NEST PREDATION, CLUTCH SIZE, AND PHYSIOLOGICAL COSTS OF EGG PRODUCTION...
99
NEST PREDATION, CLUTCH SIZE, AND PHYSIOLOGICAL COSTS OF EGG PRODUCTION IN THE SONG SPARROW (MELOSPIZA MELODIA). by Marc Travers B.Sc. Bishop's University THESIS SUBMITTED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF SCIENCE In the Department of Biological Sciences © Marc Travers 2009 SIMON FRASER UNIVERSITY Spring 2009 All rights reserved. This work may not be reproduced in whole or in part, by photocopy or other means, without pennission of the author.
Transcript of NEST PREDATION, CLUTCH SIZE, AND PHYSIOLOGICAL COSTS OF EGG PRODUCTION...

NEST PREDATION, CLUTCH SIZE, AND PHYSIOLOGICAL� COSTS OF EGG PRODUCTION IN THE SONG SPARROW�
(MELOSPIZA MELODIA).
by
Marc Travers� B.Sc. Bishop's University�
THESIS SUBMITTED IN PARTIAL FULFILLMENT OF� THE REQUIREMENTS FOR THE DEGREE OF�
MASTER OF SCIENCE�
In the� Department�
of� Biological Sciences�
© Marc Travers 2009
SIMON FRASER UNIVERSITY
Spring 2009
All rights reserved. This work may not be� reproduced in whole or in part, by photocopy�
or other means, without pennission of the author.�

APPROVAL
Name: Marc Travers
Degree: Master of Science
Title of Thesis:
Nest predation, clutch size, and physiological costs of egg production in the song sparrow (Melospiza melodia)
Examining Committee:
Chair: Dr. J. Guttman, Assistant Professor
Dr. T. Williams, Professor, Co-Senior Supervisor� Department of Biological Sciences, S.F.U.�
Dr. L. Zanette, Associate Professor, Co-Senior Supervisor Department of Biology, University of Western Ontario
Dr. D. Green, Assistant Professor� Department of Biological Sciences, S.F.U.�
Dr. B. Roitberg, Professor� Department of Biological Sciences, S.F. U.� Public Examiner�
22 January 20_0.,-9 _ Date Approved
11

SIMON� FRASER UNIVERSITY LIBRARY
Declaration of Partial Copyright Licence
The author, whose copyright is declared on the title page of this work, has granted to Simon Fraser University the right to lend this thesis, project or extended essay to users of the Simon Fraser University Library, and to make partial or single copies only for such users or in response to a request from the library of any other university, or other educational institution, on its own behalf or for one of its users.
The author has further granted permission to Simon Fraser University to keep or make a digital copy for use in its circulating collection (currently available to the public at the "Institutional Repository" link of the SFU Library website <www.lib.sfu.ca> at: <http://ir.lib.sfu.ca/handle/1892/112>) and, without changing the content, to translate the thesis/project or extended essays, if technically possible, to any medium or format for the purpose of preservation of the digital work.
The author has further agreed that permission for multiple copying of this work for scholarly purposes may be granted by either the author or the Dean of Graduate Studies.
It is understood that copying or publication of this work for financial gain shall not be allowed without the authors written permission.
Permission for pUblic performance, or limited permission for private scholarly use, of any multimedia materials forming part of this work, may have been granted by the author. This information may be found on the separately catalogued multimedia material and in the signed Partial Copyright Licence.
While licensing SFU to permit the above uses, the author retains copyright in the thesis, project or extended essays, including the right to change the work for subsequent purposes, including editing and pUblishing the work in whole or in part, and licensing other parties, as the author may desire.
The original Partial Copyright Licence attesting to these terms, and signed by this author, may be found in the original bound copy of this work, retained in the Simon Fraser University Archive.
Simon Fraser University Library Burnaby, BC, Canada
Revised: Fall 2007

SIM()N FRASER UNIVEnSlTY THINK~NG OF THE WORLD
STATEMENT OF ETHICS APPROVAL
The author, whose name appears on the title page of this work, has obtained, for the research described in this work, either:
(a) Hu man research ethics approval from the Simon Fraser University Office of Research Ethics.
or
(b)� Advance approval of the animal care protocol from the University Animal Care Committee of Simon Fraser University;
or has conducted the research
(c)� as a co-investigator, in a research project approved in advance,
or
(d)� as a member of a course approved in advance for minimal risk human research, by the Office of Research Ethics.
A copy of the approval letter has been filed at the Theses Office of the University Library at the time of submission of this thesis or project.
The original application for approval and letter of approval are filed with the relevant offices. Inquiries may be directed to those authorities.
Bennett Library Simon Fraser University
Burnaby, BC, Canada
l.Ast: "'''.SIOI''I. Summer 2007

ABSTRACT
We examined the effects of nest predation on both clutch Size and the
physiological cost of egg production using a clutch removal experiment in free-living
song sparrows (Melospiza melodia), inducing "high nest predation" (HNP) females to
produce many replacement clutches compared to "low nest predation" (LNP) females. In
a preliminary analysis we investigated the utility of multiple measures to assess
"physiological condition", including inter-correlations between physiological traits, sex
differences, and the relationship between physiological traits and reproductive
perfonnance (laying date). In our main study, experimental nest predation resulted in
HNP females laying 11 % fewer eggs per replacement clutch. As a result of frequent re-
nesting, HNP birds produced 57% more clutches (3.0 vs 4.7) and laid 41 % more eggs in
total. Physiological condition of HNP females' declined during the experiment associated
with the increase in egg production and we suggest these results are consistent with
physiological costs of egg production.
Keywords: clutch size, nest predation, cost of reproduction, condition, hematology, oxidative stress, song sparrows (Melospiza melodia).
Subject Terms: Predation, clutch size, physiology, song sparrows (Melospiza melodia)
III

ACKNOWLEDGEMENTS
1 would like to thank my supervisors, Dr. Tony Williams, Dr. Liana Zanette, and
Dr. Mike Clinchy for their support and guidance throughout this project. I have learned a
great deal from all of your advice. 1 would also like to thank Dr. David Green for sitting
on my supervisory committee, and providing helpful comments and statistics advice
during this thesis. Sophie Bourgeon, Oliver Love, and Emily Wagner deserve a big thank
you for putting in many hours helping me with lab work. I would also like to thank the
entire Williams lab for helpful comments, advice and coffee breaks. Robert DeCaire,
Katie Pagnucco, and Renaud Rincent also deserve a big thank you for putting in long
days and conducting meticulous work in the field. I would like to thank Beryl Clinchy for
providing wonderful meals and a warm place to stay at the start of the field season. I
would like to thank my family for having provided me with unconditional support in any
interest or endeavour in my life. I could not have made it to SFU without your help. It is
impossible to express how much help Marlena has provided me over the last few years.
From providing stats advice to mental support, she was always there for me and I am
deeply indebted to her.
jV

TABLE OF CONTENTS
Approval ii�
Abstract iii�
Acknowledgements iv�
Table of Contents v�
List of Figures vii�
List of Tables viii�
Chapter 1 General Introduction 1�
I. I Introduction: 2� 1.2 Nest Predation and Clutch Size 3� 1.3 Cost of Egg Production 4� 1.4 Study species- The Song Sparrow (Melospiza melodia) 8� 1.5 Summary of Thesis Chapters 9� 1.6 References II�
Chapter 2 Multivariate analysis of physiological condition in relation to� reproductive quaHty and sex 16�
2.1 Introduction 17� 2.2 Materials & Methods 19�
2.2.1 Study Species and Field Methods 19� 2.2.2 Measurement of Physiological Traits 21� 2.2.3 Statistical Analysis 24�
2.3 Results 25� 2.3.1 Relationship Between Physiological Traits 25� 2.3.2 Differences Between Sexes 27� 2.3.3 Physiology and Proximity to Egg-Laying 27� 2.3.4 Physiology and Initiation Date 28�
2.4 Discussion 28� 2.4.1 Overall Relationship Between Physiological Traits 28� 2.4.2 Individual Relationships Between Physiological Traits 30� 2.4.3 Differences Between Sexes 31� 2.4.4 Physiology and Proximity to Egg-Laying " 32� 2.4.5 Physiology and Initiation Date 33�
2.5 References 34�
v

44 Chapter 3 Experimental evidence that nest predation affects clutch size and the cost of reproduction in a free living song bird
3.1� Introduction 45 3.2� Methods 49
3.2.1� Study Species 49 3.2.2� Food Supplementation 50 3.2.3� Experimental Manipulation of Nest Predation and Clutch Number. 51 3.2.4� Measurement of Physiological Traits 53 3.2.5� Hatching Success and Nestling Growth 56 3.2.6� Statistical Analyses 56
3.3 3.3.1� Clutch Size Effects 59 3.3.2� Clutch Mass & Egg Mass Effects 60 3.3.3� Cumulative Effects of Nest Predation on Total Egg Production 61 3.3.4� Physiological Cost of Egg Production 61 3.3.5� Hatching Success and Nestling Growth 62
3.4� Discussion 62
3.5� References 72
3.4.1
Results 59
Total Cumulative Egg Production & the Physiological Cost of Egg Production 66
Chapter 4 General sythesis and future directions 83�
4.1 Synthesis 83� 4.2 Future Directions 85� 4.3 References 87�
VI

LIST OF FIGURES
Figure 2.1 Male and female mean ± SE of all distinct correlations presented in Table 2.1 40
Figure 3.1 Treatment effects on the observed mean ± SE of a) clutch size b) clutch mass C) and total seasonal egg number laid by HNP (black) and LNP (grey) females in replacement clutches 78
Figure 3.2 Relationship between treatment effects and date on a) clutch size and b) clutch mass predicted by the mixed model (REML) for HNP (solid line) and LNP (dashed line) females. Dotted lines indicate the SE around the predicted line for HNP females and dashed/dotted lines indicate SE for LNP females. Numbers 1-4.7 represent clutch number and horizontal bars indicate the SE for the average date clutches were produce. For HNP females clutch number 4 indicates the date of the 4th
clutch for females that produced a total of 5 clutches. Clutch 4.7 is the final clutch for all HNP females (i.e. either 4 or 5) 79
Figure 3.3 Proportion of3, 4, and 5 egg clutches produced by HNP (black) and LNP (grey) females across the breeding season 80
Figure 3.4 Standardized effects sizes for treatment effects on HNP females relative to LNP females. Effect size; small := 0.2, medium := 0.5, larger := 0.8 (Cohen 1992). TAC = Total antioxidant capacity, TOS := Total oxidative status 82
VlI

LIST OF TABLES�
Table 2.1 Spearman rank correlations of condition indices. Female correlations are presented below diagonal 1s with male correlation above. * Significant correlation after controlling for multiple comparisons. t Moderate to large correlations (i.e. r> 0.4 Cohen 1988) 41
Table 2.2 Mean ± SE of female and male physiological traits. Paired t-test were conducted to determine statistical differences between social pairs. * Indicates significant values after controlling for multiple comparisons. Fat score was tested using a Wilcox Sign-rank test. TAC == total antioxidant capacity, TAC-res == residuals ofTAC after controlling for uric acid. Cart == corticosterone, NEFA == non-esterified free fatty acid 42
Table 2.3 Summary of regression analysis for female and male physiological traits correlated with the days until initiation of laying. NEFA == nonesterified free fatty acids. TAC-residuals == antioxidant capacity of plasma controlling for uric acid .. "'''''''''''' 43
Table 3.1 The effect of nest predation on physiological condition. Means ± SE for physiological traits measured in females at pre-breeding and post manipulation (incubation). See chapter 2 for units. Descriptive statistics from the OFA along with standardized effect sizes for each index. Effect size; small == 0.2, medium == 0.5, larger == 0.8 (Cohen 1992) 8.l
V!l[

CHAPTER 1 GENERAL INTRODUCTION

1.1 Introduction:
Predators can affect prey demography through consumption of individuals or their
offspring and through non-consumptive costs resulting from the response of prey to
predators (Creel and Christianson 2008). Non-consumptive costs originate from
interactions between predators and their prey (i.e. failed attack, consumption of
conspecifics/young) that do not result in death to the individual (Peckarsky et a1. 1993),
but that alter prey behaviour (e.g. altered reproductive strategies, increased parental
effort) or physiology (Boonstra et a1. 1998, Carter et al. 2008, Morat et a1. 2008).
Although these responses may be the best strategy to cope with predation, they ultimately
lead to reductions in reproductive success (Nelson et a1. 2004, Creel et al. 2007, Pangle et
a1. 2007, Carter et al. 2008), physiological condition (Boonstra et a1. 1998, Clinchy et al.
2004) and survival (Creel and Christianson 2008). Therefore, in addition to the
consumptive cost of predation, a full understanding of predator-prey interactions requires
an examination of non-consumpti ve predator effects (Peckarsky et a!. 1993, Lima 1998).
In a wide range of avian species, nest predation (i.e. a consumptive cost) accounts
for the loss of approximately half of all nesting attempts and more than 80% of all nest
failures (Ricklefs 1969, Martin 1992, Martin 1995, Newton 1998). Although much is
known about the frequency and universality of nest predation, little experimental work
has examined the non-consumptive cost that may occur as a result of nest predation.
Therefore, the main objective of this thesis was to examine two potential non
consumptive costs that may result from the predator-prey interactions of nest predation;
reductions in clutch size and increased costs of egg production. What follows is a brief
2

overview of the evidence and proposed mechanistic explanations for both, the
relationship between nest predation and clutch size, and the cost of egg production.
1.2 Nest Predation and Clutch Size
In a 1947 article, Alexander Skutch first proposed that nest predators may limit
avian clutch size. Since that time, numerous descriptive studies have reported negative
correlations between clutch size and nest predation rates across species (Martin 1995,
Martin et al. 2006), habitats (Ferretti and Martin 2005, Zanette et al. 2006b, Olsen et al.
2008), and years (Julliard and Pemns 1997, Zanette et al. 2006b). Descriptive studies like
these provide valuable insight into factors associated with clutch size variation, but do not
allow us to infer a causal relationship. To date, two experimental studies have reported a
relationship between nest predation and clutch size. Eggers et al. (2006) found that
Siberian jays (Perisoreus in/austus) reduced their clutch size in response to recordings of
nest predator calls presented on their territories. Doligez and Clobert (2003)
experimentally elevated nest predation rates and displayed predator models to collared
flycatchers (Ficedula albicollis) and reported a reduced clutch size for the population in
the following year.
Slagsvold (1982) summarized the many proposed mechanisms by which nest
predation may reduce clutch size, classifying them into mechanisms that assume clutch
size affects the probability of nest predation occurring and those that assume nest
predation is independent of clutch size. With regards to the former, smaller clutches, and
the subsequent smaller broods, may incur less nest predation because of both a shorter
nest duration (i.e. fewer exposure days) and reduced nest conspicuousness. For example,
3

smaller clutches take fewer days to lay and smaller broods may fledge early if parents can
provide each nestling with more food (Perrins 1977). Smaller broods may also be less
conspicuous because better fed nestlings will beg less or because parents can make fewer
feeding trips to and from the nest (Eggers et at. 2006).
If nest predation is independent of clutch size, females may also reduce clutch
size to limit any costs (i.e. date effects or physiological) they may incur if nest predation
does take place (bet hedging strategy, Slagsvold 1984). For example, smaller clutches
require less time to produce allowing females to re-nest earlier in the season when
nestlings have a better chance of survival to adulthood (Hochacka 1990). Smaller
clutches should also reflect lower energetic or physiological investment, thus increasing a
female's ability to re-nest or her probability of survival should nest predation occur
(Slagsvold 1984).
1.3 Cost of Egg Production
Nest predation frequently results in re-nesting (Newton 1998, Grzybowski and
Pease 2005), which necessarily leads to an increase in egg production. As egg production
can incur costs, it follows that re-nesting in response to nest predation may increase the
costs that result from egg production. Experiments that have increased egg production
can provide examples for costs that may result from nest predation induced re-nesting.
Most egg production experiments are conducted on "indeterminate layers", birds
that continue to lay when eggs are removed from their nest. In these experiments, females
forced to lay additional eggs were found to have reduced physiological condition
(Kalmbach et al. 2004), reduced brood rearing capabilities (Monaghan et at. 1998), and
4

reduced local survival (Nager et al. 2001, Visser and LesseHs 2001), as well as delays in
the onset of subsequent breeding seasons (Kalmbach et al. 2004). Furthennore, increased
laying resulted in females laying poorer quality eggs (Nager and Houston 2000) that
hatch less often (Kalmbach et al. 2004), and their chicks had slower growth and increased
early mortality (Monaghan et al. 1995). Although, the continuous laying of indeterminate
layers may differ from the way predation increases egg production (i.e. through
mcreasmg complete laying bouts or number of clutches), the type of costs (those
mentioned above) and the underlying mechanisms are likely similar. Below are
descriptions of the two dominant perspectives thought to explain the mechanism(s)
underlying the cost of egg production; resource and non-resource based costs (Williams
2005).
The costs of egg laying are traditionally assumed to result from resource based
trade-offs, wherein females allocate a limited amount of resources between egg
production and their own physiological requirements (Williams 1966). During laying,
females must obtain the energy and nutrients required for self-maintenance (e.g. energy
balance, muscle repair, immune function) as well as the energy and nutrients required by
the organs involved in egg production (e.g. ovary, liver) and the egg (e.g. fat, protein,
calcium, carotenoids). If a female is un-able to obtain sufficient resources, one or all of
these functions must suffer (Partridge 2005).
In many avian systems, resource availability appears to limit egg production.
Numerous studies have found that providing supplemental food to egg laying females
will increase their clutch size (Carlson 1989, Nager et al. 1997, Clifford and Anderson
200], Zanette et al. 2006b). Resource availability has also been found to limit other
5�

aspects of egg production, such as clutch mass and clutch number. Nager (2006) reported
that females with lower endogenous protein stores prior to laying produced significantly
lighter clutches. When a subset of these females were given supplemental protein they
produced heavier clutches than control females with similar initial protein stores (Nager
2006). Blount et al. (2004) found that females supplemented with carotenoids produce
similar clutch sizes compared with control birds, but were one third more likely to
produce an additional clutch. These studies clearly show that resource availability
frequently limits egg production (i.e. reduction in current reproductive success).
However, resource limitation does not necessarily result in long tenn trade-offs as a result
of reallocation of nutrients between egg production and maternal health, as females may
simply match their investment in eggs with food availability.
More recently, it has been proposed that reproduction itself, or the regulatory
(physiological) processes controlling reproduction, may generate costs (Partridge et a!.
2005, Harshman and Zera 2007) which might be independent of resource allocation per
se (i.e. non-resource based costs, Williams (2005». Specifically, egg production costs
would be expected to occur even when a female has access to sufficient resources
(Wagner et a!. 2008b). Kalmbach et al (2004) forced two groups of great skuas
(Slercorarius skua) to lay additional eggs, one with and one without food
supplementation, and compared their timing of laying one year later with un-manipulated
controls (fewer eggs and no food supplementation). Increased egg production was found
to delay the onset of breeding similarly in both fed and un-fed birds compared to un-fed
controls (Kalmbach et a!. 2004), suggesting that additional resources during laying did
not eliminate the cost of egg production. Next, I will discuss the two most commonly
6

cited explanation for non-resource based costs: oxidative stress and negative honnone
pleiotropy (Williams 2005, Harshman and Zera 2007).
Oxidative stress is the imbalance between reactive oxygen specles (ROS) and
antioxidant defenses (Finkel and Holbrook 2000). ROS can damage bio-molecules such
as DNA, lipids and proteins (Beckman and Ames 1998), and it is the cumulative effect of
oxidative damage that is believed to result in reproductive senescence and reduced
longevity, i.e. the free radical theory of aging (Hannan 1956, Beckman and Ames 1998,
Finkel and Holbrook 2000). Oxidative stress is considered a compelling candidate for
mediating the cost of egg production in part because both the free radical theory of aging
and the cost of egg production (i.e. life history theory) similarly predict senescence and
reduced survival (Harshman and Zera 2007). In addition, egg production likely increases
a number of metabolic processes that result in the production of oxidants, including
general energy production, organ function (e.g. liver), and lipid peroxidation (Beckman
and Ames 1998). Furthermore, several studies have found that increased egg production
is associated with reduced resistance to oxidative stress. For example, fruit fly's
stimulated to lay more eggs showed increased susceptibility to experimentally elevated
oxidative stress (subjects died more rapidly), suggesting that diminished resistance to
oxidative stress is a direct physiological cost of egg production (Salmon et al. 2001,
Wang et al. 2001). In addition, Bertrand et a1. (2006) report that whole blood from zebra
finches that laid more eggs had lower resistance to oxidants (i.e. faster lysis of red blood
cells). Furthermore, Alonzo-Alverez et al (2006) found that zebra finches that laid more
clutches had lower resistance to oxidants and that resistance to oxidants was a significant
predictor of mortality up to three months after sampling.
7

Hormones are also considered likely mediators of the cost of reproduction
because of their plieotropic (multiple) effects on physiology (Ketterson and Nolan 1992,
Ketterson and Nolan 1999, Williams 2005). Honnone pleiotropy may result in costs
specific to egg production because hormones are essential regulators of egg production
(i.e. present during egg production, likely at elevated levels), but elevated plasma
hormone levels are also known to have negative effects on some aspects of physiological
condition. In birds, the erythropoietic (red blood cell production) suppressive effect of the
hormone estradiol has been proposed as one mechanistic explanation for the cost of egg
production (Kalmbach et al. 2004, Williams et a1. 2004, Williams 2005, Wagner et al.
2008a, b). Marked increases in estradiol observed during egg production (Williams et al.
2004) are known to regulate the production of yolk precursors (Christians and Williams
1999, Walzem et a1. 1999), oviduct development (Yu et a1. 1971) and reproductive
behaviour (Balthazart 1983). Yet high levels of estradiol can suppress red blood cell
production (Clermont and Schraer 1979, Blobel and Orkin 1996), which may explain the
anaemia reported in a wide range of egg laying birds (Williams 2005, Wagner et al.
2008b). Because the level of circulating red blood cells is related to flight perfonnance
and aerobic capacity (Viscor et a1. 1985, Hammond et al. 2000), egg laying induced
anaemia may ultimately lead to reduced reproductive success or survival it~ for example,
females with lower hematocrit are less able to rear broods or escape predation.
1.4 Study species- The Song Sparrow (Melospiza melodia)
Unless otherwise stated, the following was obtained from the Arcese et a1. (2002)
review in the Birds of North America. Song sparrows are one of the most widespread
8

songbirds in North America, found in habitats ranging from arctic-alpine, rain forest, to
desert scrub. Peak breeding densities are found in riparian areas, on islands, and
bordering tidal marshes. The diet of Song sparrows consists of seeds, fruits, and
invertebrates. Stomach content analysis suggests that plant material accounts for 86% of
the diet in winter, 54% in spring, 60% in summer, and 92% in fall. Song sparrows are
socially monogamous (high territory fidelity), although extra pair patemity rates of 15%
have been documented. Clutch sizes range from 2-5 eggs, egg mass ranges from 2.43
3.28g, and females can produce 3-4 successful clutches and up to 8 unsuccessful clutches
- totalling 28 eggs laid in a season (Zanette et a1. 2006b, Travers unpublished data).
Females solely build nests (open cup), incubate eggs and brood nestlings. Males and
females share the feeding of nestlings and fledglings. Nests are not re-used after
successful or unsuccessful breeding attempts. The rate of nest predation varies
considerably by habitat. At the study site used for this thesis, 48-69% of nests were
consumed between 2000-2002 (Zanette et a1. 2006a). The maximum recorded life span of
a song sparrow is 8-9 years. On Mandarte Island, British Columbia, 56% of the variation
in life time reproductive success (number of recruits to the island) was partitioned to
offspring survival to independence, 10% to life span, and 5% to the number of eggs
produced (all variables were positively correlated with life time reproductive success,
Smith 1988).
1.5 Summary of Thesis Chapters
In chapter 2, we investigated the utility of using multiple physiological traits to
assess "physiological condition". We examine (1) whether a suite of physiological traits
9

are likely to provide redundant infonnation on physiological condition (inter-conelations
between traits), (2) sources of variation in physiological traits (sex differences), and (3)
the relationship between physiological traits and reproductive perfonnance (laying date).
In chapter 3, we experimentally test if nest predation affects clutch size and the
physiological condition of females. Using a clutch removal manipulation, we induced
"high nest predation" (HNP) females to produce many replacement clutches compared to
"low nest predation" (LNP) females. We predicted that high nest predation would
decrease clutch size, however, we also predicted that by inducing re-nesting predation
would increase total egg production, resulting in elevated physiological costs of egg
production in HNP females. We test for nest predation affects on average clutch size and
on the seasonal pattern of clutch size, total seasonal egg production, and female
physiological state, and we evaluate if physiological effects are consistent with the
various predictions for the cost of egg production. Because females with insufficient
resources may trade-off their offspring quality to maintain their own condition or because
costs incuned by mothers may be passed on to offspring, we test for predation affects on
hatching success of eggs and on nestling size. Throughout the experiment we provided
all birds with unlimited access to high quality feed For this reason we expected that
treatment effects would not be attributed to resource limitation.
In chapter 4, 1 briefly review implications of the main results from the preceding
chapters, and discuss possibilities for future research.
10

1.6 References
Alonso-Alvarez, c., S. Bertrand, G. Devevey, J. Prost, B. Faivre, O. Chastel, and G. Sorci. 2006. An experimental manipulation of life-history trajectories and resistance to oxidative stress. Evolution 60: 1913-1924.
Arcese, P., M. K. Sogge, A. B. MalT, and M. A. Patten. 2002. Song SpalTow (Melospiza melodia).in A. G. Poole, F, editor. The Birds of North America. The Birds of NOlih America, Inc., Philadelphia.
Balthazart, J. 1983. Hormonal cOlTelates of behaviour. Pages 221-365 in D. S. Farner, J. R. King, and K. C. Parkes, editors. Avian Biology. Academic Press, New York,.
Beckman, K. B. and B. N. Ames. 1998. The free radical theory of aging matures. Physiological Reviews 78:547-581.
Bertrand, S., C. Alonso-Alvarez, G. Devevey, B. Faivre, 1. Prost, and G. Sorci. 2006. Carotenoids modulate the trade-off between egg production and resistance to oxidative stress in zebra finches. Oecologia 147:576-584.
Blobel, G. A. and S. H. Orkin. 1996. Estrogen-induced apoptosis by inhibition of the erythroid transcription factor GATA-I. Molecular and Cellular Biology 16: 16871694.
Blount, J., D. C. Houston, P. F. Surai, and A. P. Moller. 2004. Egg-laying capacity is limited by carotenoid pigment availability in wild gulls Lams fuscus. Proceedings of The Royal Society of London Series B-Biological Sciences 271:S79-S81.
Boonstra, R., D. Hik, G. R. Singleton, and A. Tinnikov. 1998. The impact of predatorinduced stress on the snowshoe hare cycle. Ecological Monographs 68:371-394.
Carlson, A. 1989. Courtship feeding and clutch size in red-backed shrikes (laniuscollurio). American Naturalist 133:454-457.
Carter, M. J., C. Vega-Retter, and R. Ramos-Jiliberto. 2008. Non-lethal effects of invertebrate predators on Daphnia: morphological and life-history consequences of water mite kairomone. Freshwater Biology 53: 1857-1867.
Christians, J. K. and T. D. Williams. 1999. Effects of exogenous 17 beta-estradiol on tee reproductive physiology and reproductive perfonnance of European starlings (Sturnus vulgaris). Journal of Experimental Biology 202:2679-2685.
Clermont, C. P. and H. Schraer. 1979. Effect of estrogen on rate offe-59 uptake by hematopoietic-tissue in japanese quail. American Journal of Physiology 236:E342-E346.
Clifford, L. D. and D. J. Anderson. 2001. Food limitation explains most clutch size variation in the Nazca booby. Journal of Animal Ecology 70:539-545.
11

Clinchy, M., L. Zanette, R. Boonstra. J. C. Wingfield, and J. N. M. Smith. 2004. Balancing food and predator pressure induces chronic stress in songbirds. Proceedings of the Royal Society Biological Sciences Series B 271:2473-2479.
Creel, S. and D. Christianson. 2008. Relationships between direct predation and risk effects. Trends In Ecology & Evolution 23:194-201.
Creel, S.) D. Christianson, S. Liley, and J. A. Winnie, Jr. 2007. Predation risk affects reproductive physiology and demography of elk. Science 315:960.
Doligez, B. and J. Clobert. 2003. Clutch size reduction as a response to increased nest predation rate in the collared flycatcher. Ecology 84:2582-2588.
Eggers, S., M. Griesser, M. Nystrand, and J. Ekman. 2006. Predation risk induces changes in nest-site selection and clutch size in the Siberian jay. Proceedings of the Royal Society Biological Sciences Series B 273:701-706.
Ferretti, V. L. P. E. and T. E. Martin. 2005. Life-history variation of a neotropical thrush challenges food limitation theory. Proceedings of The Royal Society B-Biological Sciences 272:769-773.
Finkel, T. and N. J. Holbrook. 2000. Oxidants, oxidative stress and the biology of ageing. Nature 408:239-247.
Grzybowski, J. and C. M. Pease. 2005. Renesting detennines seasonal fecundity in songbirds: What do we know? What should we assume? Auk 122:280-292.
Hammond, K. A., M. A. Chappell, R. A. Cardullo, R. S. Lin, and T. S. Johnsen. 2000. The mechanistic basis of aerobic perfonnance variation in red junglefowl. Journal of Experimental Biology 203:2053-2064.
Harman, D. 1956. Aging - A theory based on free-radical and radiation-chemistry. Journals of Gerontology 11:298-300.
Harshman, L. and A. J. Zera. 2007. The cost of reproduction: the devil in the details. Trends In Ecology & Evolution 22:80-86.
Hochacka, W. 1990. Seasonal decline in reproductive performance of song sparrows. Ecology 71: 1279-1288.
Julliard, R. M. R. H. C. J. and C. M. Perrins. 1997. Phenotypic adjustment of clutch size due to nest predation in the Great Tit. Ecology 78:394-404.
Kalmbach, E., R. Griffiths, J. E. Crane, and R. W. Furness. 2004. Effects of experimentally increased egg production on female body condition and laying dates in the great skua Stercorarius skua. Journal of Avian Biology 35:501-514.
Ketterson, E. and V. Nolan. 1999. Adaptation, exaptation, and constraint: A honnonal perspective. American Naturalist 154:S4-S25.
Ketterson, E. D. and V. Nolan, Jr. 1992. Hormones and life histories: An integrative approach. American Naturalist 140:S33-S62.
12

Lima, S. L. 1998. Nonlethal effects in the ecology of predator-prey interactions - What are the ecological effects of anti-predator decision-making? Bioscience 48:25-34.
Martin, T. 1995. Avian life-history evolution in relation to nest sites, nest predation, and food. Ecological Monographs 65:101-127.
Martin, T., R. D. Bassar, S. K. Bassar, J. 1. Fontaine, P. Lloyd, H. A. Mathewson, A. M. Niklison, and A. Chalfoun. 2006. Life-history and ecological correlates of geographic variation in egg and clutch mass among passerine species. Evolution 60:390-398.
Martin, T. E. 1992. Interaction of nest predation and food limitation in reproductive strategies. Current Ornithology 9: 163-197.
Monaghan, P., M. Bolton, and D. C. Houston. 1995. Egg-production constraints and the evolution of avian clutch size. Proceedings of The Royal Society of London Series B-Biological Sciences 259: 189-191.
Monaghan, P., R. G. Nager, and D. C. Houston. 1998. The price of eggs: increased investment in egg production reduces the offspring rearing capacity of parents. Proceedings ofThe Royal Society of London Series B-Biological Sciences 265: 173 I -173 5.
Morat, F., S. Betoulle, M. Robert, A. F. Thailly, S. Biagianti-Risbourg, and R. LecomteFiniger. 2008. Parental care in response to natural variation in nest predation pressure in six sunfish (Centrarchidae: Teleostei) species. Ecology of Freshwater Fish 17:617-627.
Nager, R. 2006. The challenges of making eggs. Ardea 94:323-346.
Nager, R., P. Monaghan, and D. C. Houston. 2001. The cost of egg production: increased egg production reduces future fitness in gulls. Journal of Avian Biology 32: 159166.
Nager, R. G., C. Ruegger, and A. J. VanNoordwijk. 1997. Nutrient or energy limitation on egg fOnTIation: A feeding experiment in great tits. Journal of Animal Ecology 66:495-507
Nager, R. M. P. and D. C. Houston. 2000. Within-clutch trade-offs between the number and quality of eggs: Experimental manipulations in gulls. Ecology 81: 1339-1350.
Nelson, E. H., C. E. Matthews, and J. A. Rosenheim. 2004. Predators reduce prey population growth by inducing changes in prey behavior. Ecology 85: 1853-1858.
Newton, 1. 1998. Population limitation in birds. Academic Press, London, UK
Olsen, B., J. Felch, R. Greenberg, and J. Walters. 2008. Causes of reduced clutch size in a tidal marsh endemic. Oecologia 158:421-435.
t3

Pangle, K. L., S. D. Peacor, and O. E. Johannsson. 2007. Large nonlethal effects of an invasive invertebrate predator on zooplankton population growth rate. Ecology 88:402-412.
Partridge, L., D. Gems, and D. J. Withers. 2005. Sex and death: What is the connection? Cell 120:461-472.
Peckarsky, B. L., C. A. Cowan, M. A. Penton, and C. Anderson. 1993. Sublethal consequences of stream-dwelling predatory stoneflies on mayfly growth and fecundi ty. Ecology 74: 1836-1846.
Perrins, C. M. 1977. The role of predation in the evolution of clutch size. Pages 181-191 in P. CM, editor. Evolutionary Ecology. Macmillian Press, London.
Ricklefs, R. E. 1969. An analysis of nesting mortality in birds. Smith. Contribution to Zoolology 9: 1-48.
Salmon, A. B., D. B. Marx, and L. G. Harshman. 2001. A cost of reproduction in Drosophila melanogaster: Stress susceptibility. Evolution 55: 1600-1608.
Skutch, A. F. 1949. Do tropical birds raise as many young as they can nourish? Ibis 91 :430-455.
Slagsvold, T. 1984. Clutch size variation of birds in relation to nest predation - on the cost of reproduction. Journal of Animal Ecology 53:945-953.
Smith,.I. N. M. 1988. Determinants oflifetime reproductive success in the Song Sparrow. Pages 154-172 in T. H. Clutton-Brock, editor. Reproductive Success. University of Chicago Press, Chicago.
Viscor, G., M. S. Marques, and 1. Palomeque. 1985. Cardiovascular and organ weight adaptations as related to flight activity in birds. Comparative Biochemistry and Physiology a-Physiology 82:597-599.
Visser, M. E. and C. M. Lessells. 2001. The costs of egg production and incubation in great tits (Porus major). Proceedings of The Royal Society of London Series Biological Sciences 268:1271-1277.
Wagner, E., J. S. Prevolsek, K. E. Wynne-Edwards, and T. D. Williams. 2008a. Hematological changes associated with egg production: estrogen dependence and repeatability. Journal of Experimental Biology 211 :400-408.
Wagner, E. C., C. A. Stables, and T. D. Williams. 2008b. Hematological changes associated with egg production: direct evidence for changes in erythropoiesis but a lack of resource dependence? Journal of Experimental Biology 211:2960-2968.
Walzem, R. L., R. J. Hansen, D. L. Williams, and R. L. Hamilton. 1999. Estrogen induction of VLDLy assembly in egg-laying hens. Pages 467S-472S. Amer Inst Nutrition.
14

Wang, Y., A. B. Salmon, and L. G. Harshman. 2001. A cost ofreproduction: oxidative stress susceptibility is associated with increased egg production in Drosophila melanogaster. Experimental Gerontology 36: 1349-1359.
Williams, G. 1966. Natural selection costs of reproduction and a refinement of lacks principle. American Naturalist 100:687-&.
Williams, T. 2005. Mechanisms underlying the costs of egg production. Bioscience 55:39-48.
Williams, T., W. O. Challenger, 1. K. Christians, M. Evanson, O. Love, and F. Vezina. 2004. What causes the decrease in hematocrit during egg production? Functional Ecology 18:330-336.
Yu, J. Y. L., L. D. Campbell, and R. R. Marquardt. 1971. SEX HORMONE Control mechanisms part 1 effect of estrogen and progesterone on major cellular components in chicken gallus-domesticus oviducts. Canadian Journal of Biochemistry 49:348-356.
Zanette, L., M. Clinchy, and J. N. M. Smith. 2006a. Combined food and predator effects on songbird nest survival and annual reproductive success: results from a bifactorial experiment. Oecologia 147:632-640.
Zanette, L., M. Clinchy, and 1. N. M. Smith. 2006b. Food and predators affect egg production in song sparrows. Ecology 87:2459-2467.
15

CHAPTER 2 Multivariate analysis of physiological condition in relation to reproductive quality and sex
Marc Travers l ,2, Liana Zanette2 Michael Clinch/, Tony D. Williams I
'Simon Fraser University, Department of Biological Sciences, Burnaby, B.C., Canada 2University of Westem Ontario, Department of Biological Sciences, London, Ontario, Canada 3University of Victoria, Department of Biological Sciences, Victoria, B.c. Canada
16

2.1 Introduction
Body condition is of particular interest in studies of free-living animals because it
is thought to indicate aspects of individual quality that are related to performance in many
quality-dependent traits such as lay date (Bety et a!. 2003, Ninni et a1. 2004), clutch size
(Andersson and Gustafsson 1995), egg size (Smith and Moore 2003) offspring size
(Atkinson and Ramsay 1995), and survival (Romero and Wekelski 200 I, Blums et al.
2005). Body condition is frequently measured with morphometric based indices like the
body condition index (BCl), which is a measure of mass controlling for structural body
size. The BCl is widely used because it is non-invasive, easy to obtain in field studies,
and may reflect aspects of physiological state widely considered important to an animal's
condition such as nutrient status and fat storage (Brown 1996, Glazier 2000, Smith and
Moore 2003). However, some authors have questioned the validity of using the BCl
because the basic statistical and allometric principles of the method are unlikely to be met
in most field studies (Green 2001, Hayes and Shonkwiler 2001). While some empirical
studies have found that Bel is significantly correlated with actual body fat (Ardia 2005,
Schulte-Hostedde et a1. 2005), others have concluded that BCl may not be a broadly
applicable index of condition because it accurately reflects fat content in some species
but not others (Spengler et a!. 1995).
More recently, physiological indices have been increasingly used to measure
condition and assess individual quality. Physiological indices are promising condition
indicators because these assays can be used on a wide range of species to measure diverse
aspects of physiological state. For example, researchers have used physiological indices
to measure energetic state (Jenni-Eiermann and lenni 1997, Williams et a1. 1999).
17

immune function (Bourgeon et a1. 2007), hormone profiles (Love et al. 2005), blood
constituents (Totzke et al. 1999) and oxidative stress (Alonso-Alvarez et a1. 2004).
Importantly, physiological indices have been related to fitness related traits like arrival
date following migration (Piersma 1996), timing of breeding and clutch size (Andersson
and Gustafsson 1995) and survival (Nadolski et aJ. 2006).
Most studies that have used physiological indices have attempted to measure
condition with a single physiological trait or index. However, the complexity of
organismal physiology would suggest that measuring a single aspect (e.g. oxygen
carrying capacity or fat content) of physiological state may not provide a complete
understanding of condition. Yet to date, there has been little work examining how much
individual measures tell us about other aspects of overall physiological state. If multiple
physiological indices are highly correlated with one another and are indicative of many
aspects of physiological state, then individual indices may be expected to provide
considerable infonnation about overall condition. However, if physiological traits are not
highly related to one another measurement of multiple physiological traits might be
required for a more accurate assessment of true "condition" (Seeman et a1. 2001).
Here we take a comprehensive approach to measuring physiological condition in
male and female song sparrows (Melospiza melodia) during the pre-breeding and brood
rearing period. Specifically, we measured multiple physiological traits as putative indices
of condition (15 indices in total) relating to energetic state, hematology, oxidative stress,
and immune function. We first test for correlations between the individual physiological
traits we measured to examine the overall strength of relationships between common
indices of condition. We then compare physiological traits in female and male social
18

pminers to test whether sex is an important source of variation in the indices measured
here. Male and female physiology may differ as a result of differing roles (demands)
during some stages of breeding. Alternatively, sexes may show similar physiological
scores due to assortative mating or territory quality. Basic knowledge of sources of
variation should aid future studies in differentiating between environmental noise and
experimental effects. Final1y, we use a multivariate approach to test if an integrated
assessment of condition explains variation in the date females initiate breeding, and we
conduct these correlations in both females and their male social partners. Male condition
may be related to lay date if females paired to high quality males lay earlier or if territory
quality is also an important factor influencing lay date. Lay date is a good trait to
examine if the physiological traits measured here could be used as an indicator of
individual quality because 1) there is considerable evidence linking lay date with
reproductive success in birds (Daan et al. 1990, Verhulst and Tinbergen 1991, Sanz 1999,
Brinkhof et a1. 2002), indicating that lay date is a measure of reproductive quality and
because 2) lay date is considered a condition dependent trait (Bearhop et a1. 1999,
Kalmbach et a1. 2004).
2.2 Materials & Methods
2.2.1 Study Species and Field Methods
We studied song sparrows (Mefospiza mefodia) on the Southern Gulf Islands of
British Columbia, Canada. Song sparrows in this area are resident year round and multi
brooded. Breeding typically begins mid April and ends in early July. Pre-breeding song
19

sparrows were captured between March 6 and March 28 (Julian capture date mean ± SD
= 76.8 ± 7.4) using baited potter traps. We pre-baited potter traps with 2.5ml of white
millet per day, for an average of 5 days prior to capture. Once captured, all birds were
colour-banded for individual identification and a blood sample (up to 300j..d) was
collected from the brachial vein using heparanized capillary tubes. We measured blood
glucose concentration (see below) and prepared blood smears immediately after releasing
the bird. Blood smears were air dried and stored in slide boxes containing desiccation
packs. We refrigerated blood in cooler packs for a maximum of 10 hours before
separating the plasma from the red blood cells. Plasma was stored on dry ice until
pennanent laboratory storage at -20°e. For all females sampled during the pre-breeding
period we attempted to locate their first nest. If a nest was found after incubation had
begun, we back calculated the lay date of the first egg by aging the eggs with the aid of
an egg candler. Laying date or initiation of breeding was then defined as the date when a
female laid her first egg in her first nest of the season (Julian lay date mean ± SD = I 07.8
± 8.8). We also captured and collected blood from females and males during the breeding
season. Breeding birds were sampled on the 61h day of the nestling phase using mist nets
placed across the flight path from the nest to their foraging areas. Blood used for
corticosterone analysis was collected in under 3 min from the time the bird hit the net.
Other than capture methods, blood collection procedures and storage were identical to
pre-breeding methods.
20�

2.2.2 Measurement of Physiological Traits
We examined fifteen physiological traits measuring components of hematological
state and oxygen carrying capacity (hematocrit, hemoglobin, and polychromatic, i.e.
proportion of reticulocytes or immature red blood cells), oxidative stress (total oxidative
status (TOS), total antioxidant capacity (TAC)), uric acid levels, plasma carotenoids, and
TAC-residuals controlling for uric acid, (i.e. antioxidant capacity not attributed to uric
acid), immunological status (plasma immunoglobulin levels), honnonal (total
corticosterone), and energetic state (plasma glucose, NEFA, triglyceride, glycerol). In
addition, we measured more widely used indices of body condition: body mass (g), body
condition index and fat score. Body condition index scores are mass divided by tarsus
length. We quantified visible subcutaneous fat stores under the wing, and in the furcular
and abdominal region using a 7 point scale (DeSante et a1. 2008). Physiological traits
were measured as follows:
We used colorimetric assays to measure plasma levels of total antioxidant
capacity (TAC), uric acid, total oxidant status (TOS), carotenoids, immunoglobulin (lg),
triglycerides, free glycerol, NEFA, and whole blood levels of hemoglobin. We
detennined assay variation using a hen plasma standard. Intra-assay variation was 4.6%,
5.4%, 2.7%, 2.6% 1.7%, 3%, 5.7% and 2.2% for triglyceride, uric acid, NEFA, 19,
carotenoids, TAC, TOS, and hemoglobin respectively. Inter-assay variation was 2.4%,
4.4%, 1.1 %, 19.4%, 5.9%, 2.3%, 9.2% for triglyceride, uric acid, NEFA, Ig, carotenoids,
TAC, and TOS respectively (we did not obtain an inter-assay CV for hemoglobin).
21

Hematological status: Hematocrit was measured using standard techniques (Campbell
1995) after centrifugation for 10mln using a Zipocrit (Laboratory Essentials, USA)
portable centrifuge. Hemoglobin concentration in whole blood was determined using the
cyanomethemoglobin method (Drabkin and Austin 1932) modified for a colorimetric
assay, using 5~1 of whole blood diluted in 1.25ml Drabkin's reagent (05941, Sigma
Aldrich, Canada) with absorbance measured at 540nm. Polychromasia was counted on
smears stained with Wright-Giemsa (Sigma-Aldrich Canada, Ltd) and was calculated as
the proportion of red blood cells (RBC) that are immature (Campbell 1995). A single
observer, who was blind to experimental 10, counted (>500 RBC per count) all smears in
triplicate and had an intra-count variation of <4%. Polychromasia scores and intra-count
variation were calculated from the two most similar scores of 3 repeated counts of the
same smear.
Oxidative stress: Total antioxidant capacity (TAC) was determined using a modified
Randox-TEAC assay described by Ere! (2004). TAC results are reported in mmol Trolox
equivalent.L-1 (238813. Sigma-Aldrich). Plasma uric acid concentrations were measured
in duplicate using the QuantiChrom™ uric acid kit (DIUA-250; BioAssay Systems,
USA). Total oxidative status (TaS) was determined as described by Erel (2005). TOS
results are reported in H202 equivalent.L- I. We prepared the carotenoid assay (see
Alonso-Alvarez et a1. 2004) by vortexing 20~1 of plasma diluted in 180~1 of absolute
ethanol, centrifuging the plasma/ethanol at 1500g for 10min, and recovering the
supernatant. Plasma carotenoid concentrations were determined in triplicate using 50~1 of
supernatant per well, with absorbance measured at 450nm and concentrations calculated
22�

usmg a standard curve of lutein (minimum 70% xanthophyll from alfalfa; Sigma
Aldrich).
Immune status: Plasma immunoglobulins (Ig) were determined using the ELISA method
with commercial anti-chicken antibodies as reported by Martinez et al. (2003). We
adapted the method for use in song sparrows by determining the sparrow appropriate
plasma dilution (1/8000). Ig results are presented in units of absorbance.
Hormonal. The concentration of total corticosterone (see Breuner and Orchinik 2002,
Love et al. 2004, 2005) in non-extracted plasma was determined using a corticosterone
Enzyme-linked-immunoabsorbent Assay (EIA - Assay Designs Inc., Michigan USA,
catalog # 901-097) with a 4-parameter logistic fit.
Energetic status: The plasma concentration of free glycerol and total triglycerides were
determined in duplicate samples with absorbance measured at 540nm after 10min of
incubation at 37°C (see Seaman et al. 2006), using Sigma-Aldrich reagents (Triglyceride
reagent & Free glycerol reagent, Sigma-Aldrich Canada). Triglyceride concentrations
were calculated by subtracting free glycerol from total triglyceride. Non-esterified free
fatty acids concentrations were determined in triplicate using a NEFA kit (NEFA-HR(2);
Wako Diagnostics, USA). Blood glucose scores were measured at the time of capture
using Acensia® glucose meter (Bayer Inc).
23�

2.2.3 Statistical Analysis
We used Speannan rank cOlTelations to examine the relationships between all
physiological traits to limit the influence of outliers on our statistical tests (Myers and
Well 2003). The r-values from the speannan rank correlations are presented in Table 2.1
as a correlation matrix. We first test whether all correlations (r-values) within the matrix
(separately for males and females) were significantly larger than would be expected by
chance when conducting this many correlations (i .e. were the matrix wide correlations
significantly different from zero given our sample size and number of correlations). We
used the program Multicorr to conduct the global test of the correlation matrix (Steiger
1979, 1980,2005). We then tested whether individual correlations within the matrix were
significant using standard t-tests, setting alpha at 0.05 and adjusting P-values for the
number of statistical comparisons conducted using the false discovery method (FOR)
(Benjamini and Hochberg 1995). For these correlations, we did not control for any
potential environmental effects like date, time of day or temperature because we wanted
to see how strongly traits are actually related to each other. Fore example, we know that
time of day affects some traits and not others. This indicates that traits are not necessarily
or always functionally linked, and the extent to which traits are, or are not, linked is
exactly what we are interested in testing. We used paired t-tests to examine sex
differences between social pairs, allowing us to test sex differences controlling for
territory quality. We again adjusted P-values using FOR. We used forward stepwise
regressions to examine if physiological traits of females are 1) related to the proximity
(days) from sampling to the beginning of egg-laying so that we could examine whether
physiology changes as females approach egg-laying and; 2) related to initiation date (date
on which the first egg of the season is laid) to assess whether early breeders are
24

physiologically distinct from those that begin their breeding season later. For males, we
are testing whether male physiology is related to 1) the proximity of sampling to their
partner's initiation of breeding and 2) the date their pattner started breeding. For the
stepwise regression analysis we set the F to enter at I, which allowed non-signiflcant
contributors to enter the model in some cases. Prior to the regression analyses we found
that lay date and the number of days from blood sampling to laying were highly
correlated requiring us to statistically partition out their shared affects on physiology in
the two models described above. To do this we entered lay date as an independent
variable in the model with days to laying as the dependent variable (model 1) and vice
versa for the model with lay date as the dependent variable (model 2). For the
physiological variables entered into the regression analyses we first examined lf capture
date, time, and temperature were significantly related to physiological traits. For traits
correlated with one or more of these confounding variables, we Obtained the residuals of
the regression, which were then used in the regression analyses in which proximity to
egg-laying and lay date were dependent variables. It should be noted that polychromasia
was not measured in males and corticosterone was not measured in pre-breeding samples.
Prior to any analyses all variables were Box-cox transformed (Krebs 1999) and tested for i
nonnality using the Shapiro-Wilks test.
2.3 Results
2.3.1 Relationship Between Physiological Traits
Overall, the inter-correlations between physiological traits (Table 2.1) were
significantly different from zero when simultaneously considering all possible
25

correlations (females: X2 120 = 256.2, P< 0.0001; Males: X2
105 = 213.1, P< 0.0001).
However, the magnitude of the correlations between all the physiological traits were
generally weak to moderate (Table 2.1), considering Cohens (1988) guidelines for the
strength of correlation effects (i.e. small= 0.1, medium= 0.3, large= 0.5). The average
correlation after removing the direction of etIect (- or +), was 0.22 (SO = 0.16, range =0
91) for females, and 0.25 (SD = 0.17, range = 0-89) for males.
After controlling for the number of comparisons conducted, significant positive
correlations were found between total antioxidant capacity (TAC) and uric acid (Females:
t20= 9.66, P< 0.001, spearman r = 0.91; Males: tl7= 4.60, P< 0.001, spearman r = 0.74 )
and total oxidative status (TOS) and triglycerides (Females: t20= 3.97, P< 0.001,
spearman r = 0.66; Males: tl7= 3.58, P< 0.001, spearman r = 0.66) in both females and
males (Table 2.1). In Table 2.1, we also highlight moderate to large (r > OA, Cohen 1988)
correlations in female and male physiological traits.
2.3.1.1� Mass, Body Condition Index, and Fat Score
These more commonly used indices of condition also generally had a weak to
moderate average correlation with other traits (Cohen 1988) and, mass and condition
generally had similar relationships with other condition indices (Table 2.1). Female mass
and BCI had mainly negative relationships with other indices, with the notable exception
that mass and BCI tended (after correction for multiple comparisons) to be positively
related with NEFA (Table 2.1). In males, mass was significantly negatively correlated
with TAC (tI7= 3.92, P< 0.001, spearman r = -0.69), with BCI following a similar trend.
In males only, fat scores tended non-significantly to be positively related to uric acid and
glucose, while female fat scores were weakly correlated with all other variables.
26

2.3.2� Differences Between Sexes
Females were significantly lighter than their social mate at pre-breeding capture
(Table 2.2). Females tended to have higher levels of antioxidants other than uric acid (i.e.
TAC-residuals), and higher levels of oxidants (TOS), although both were not significant
after controlling for multiple comparisons. During the nestling phase, females were again
significantly lighter than their social mate, but also had a significantly lower body
condition index (Table 2.2). Females also tended to have lower hemoglobin during the
nestling phase (P= 0.051).
2.3.3� Physiology and Proximity to Egg-Laying
Several physiological traits in non-breeding females and males were significantly
related to the number of days until females laid their first clutch, i.e. days from sampling
to initiation of breeding. In females, hematocrit, triglycerides, and immunoglobulin (Ig),
loaded into the stepwise procedure as significantly positively correlated with the days
until laying (i.e. females with lower levels of these variables were closer to laying, Table
2.3). TAC-residual, which is a measure of antioxidant capacity not attributed to uric acid,
tended to be positively correlated with days until laying (Table 2.3). In contrast, glucose,
fat score, and uric acid were significantly negatively correlated with days to egg-laying.
In mates, the traits fat score and TAC-residuals were significantly positively correlated,
while NEFA (non-esterified free fatty acids) were significantly negatively correlated with
27�

days until their partner begins egg-laying (Table 2.3). Triglycerides and hemoglobin were
non-significant contributors in the stepwise procedure.
2.3.4 Physiology and Initiation Date
Several physiological traits were related to initiation date after controlling for the
proximity of blood sampling to egg-laying date. Females that staried breeding earlier in
the season had lower immunoglobulin scores (~= 0.38, t16= 2.38, P= 0.03) and tended to
have higher levels of free glycerol (~= -0.35, t16= -2.04, P= 0.058) than females that
initiated later, while hematocrit and NEFA were non-significant contributors in the
stepwise procedure (hematocrit: p= -0.08, t16= -0.59, P= 0.56; NEFA: p= 0.18, t16= 1.33,
P= 0.19). Males paired to females that initiated breeding earlier in the season had higher
TAC-residual (p= -0.24, tl3= -2.19, P= 0.047), and fat (P= -0.32, t13= -2.35, P= 0.03) and
tended to have higher hemoglobin (P= -0.25, tl3= -2.12, P= 0.054) and lower free glycerol
(P= 0.21, tl3= 1.72, P= 0.11).
2.4 Discussion
2.4.1 Overall Relationship Between Physiological Traits
We found significant overall relationships (global assessment) between all the
physiological traits measured, as would be expected if each trait measures some aspect of
overall condition. That is, we would predict that animals in good condition would have
better scores on many or all traits with the opposite occurring in poor condition
28�

individuals. However, if these traits do measure aspects of overall condition or quality it
is somewhat surprising that the relationships between traits were generally weak to
moderate. Particularly interesting is body condition index (BCI), which is frequently used
as an indicator of general nutritional state, but as are results show had only a small to
moderate correlation with our other variables including those that measure energetic and
nutritional status. Furthermore, our results indicate that several traits were related to each
other in a manner inconsistent with individuals being in either good or poor condition
(i.e. traits were inversely related with respect to interpretation of condition). For example,
mass and BCI were negatively related to many traits in both sexes, indicating that lighter
birds were in better condition. 'Unexpected inverse correlations' could arise if birds in
better condition can better afford to lose mass while gaining in some other aspect of
reproduction, such as greater territorial defence or increased foraging effort for specific
nutrients. Alternatively, birds may facultatively reduce mass (and BCI) to increase flight
performance when they are highly active (Norberg 1981, Gaston and Jones 1989).
Regardless of the exact reason for 'unexpected inverse correlations', such results
suggest that jf physiological traits are measured i.n isolation researchers may come to
incorrect conclusions about an animal's condition. Furthermore, the small to moderate
cOlTelations found between all the traits suggest that no single trait is likely to tell us
much about the variation in an animal's overall physiological state; implying that
individual physiological traits may also not be strongly related to quality dependent traits
like survival or fecundity, etc. In support ofthis idea, Seeman et al. (2001), in a study on
human aging, found that individually none of the 10 physiological traits they measured
predicted the timing of death. However, the cumulative measure of condition produced
29�

after combining the 10 traits did in fact significantly predict mortality. Seeman et a1.
(2001) suggested that to better understand condition we need to measure the combined
'physiological burden' incurred by individuals using measures that incorporate multiple
aspect of physiological state (see also chapter 3).
2.4.2 Individual Relationships Between Physiological Traits
Our results indicate that there were several moderate to strong correlations
between individual traits, however only a few correlations related to oxidative stress
remained significant after controlling for multiple comparisons. Here TAC (total plasma
antioxidant capacity) and uric acid were significantly positively correlated in both males
and females, which is consistent with uric acid acting as a strong antioxidant (Erel 2004).
In a comparison across 200 avian species, Cohen (2007) found an average correlation of
0.79 between uric acid and TAC, which is similar to the correlations (0.74 males, 0.91
females) found in this study. We also found strong positive correlations between TOS
(total plasma oxidative status) and triglycerides in both males and females. Positive
correlations between TOS and triglycerides may be explained by findings that
triglycerides are correlated with the release of ROS (reactive oxygen species) from white
blood cells in humans (Araujo et a1. 1995, Katsuki et al. 2004, Mazor et a1. 2008). In
males, mass and TAC were negatively correlated. Similar inverse relationships were
evident between mass and both uric acid and TAC residuals (antioxidant capacity not
attributed to uric acid), indicating that the higher TAC of lighter males was due to higher
levels of both uric acid and TAC residuals.
30

2.4.3 Differences Between Sexes
Most physiological traits were similar between male and female social mates at
both pre-breeding and brood rearing, with the exception that brood rearing females had
lower body condition index scores. The results of previous sex comparisons of
physiological traits vary considerably across species. Several studies have found that
measures of energetics, hematology and immune function differed between the sexes at
brood rearing (Kern et al. 2005, Owen et al. 2005, Kilgas et al. 2006) and pre-breeding
(Horak et al. 1998) while others have found no sex differences at various times
throughout the annual cycle (Acquarone et a1. 2002, Hauptmanova et al. 2002, Masello
and Quillfeldt 2004, Sanchez-Guzman et al. 2004). Owen et al. (2005) attributed sex
differences in physiological traits during brood rearing to the greater work load of males
who not only provision nestlings but defend territories as well. In a similar vein,
behavioural similarities between the sexes at brood rearing may explain why song
sparrows in the present study had comparable scores for most physiological traits. Here,
males and females were captured on the 6lh day of brood rearing; a time when the sexes
appear behaviourally the most similar during the breeding season. By this stage both
sexes appear to be equally focused on nestling provisioning as females greatly reduce
brooding during day light hours at this time and males appear less territorial as indicated
by less singing and a lack of response to song play backs (per. observation). At pre
breeding, similarities in the physiology of male and female partners also may indicate
similar behaviour and energetic demands, or potentially different behaviour with similar
net energetic demands or effects on physiology. Although not tested here, it may also be
possible that small sex differences when added across the suite of physiological traits
31

may in fact indicate a cumulative physiological difference between the sexes at both
stages examined here (Seeman et a1. 2001).
2.4.4 Physiology and Proximity to Egg-Laying
Several physiological traits in both males and females appear to be related to the
proximity of egg-laying. Males had lower fat scores, less circulating antioxidants and
higher levels of free fatty acids (NEFA). These data suggest that males were more active
as the date their partner initiated egg-laying approached, but that this activity was not
directed towards increasing fat reserves or obtaining antioxidant rich foods. The lower fat
and higher free fatty acid scores of males suggest that fat is being converted to fatty acids
to fuel flight muscles (Butler and Bishop 2000) and high activity levels are potentially
metabolising antioxidants or reducing the foraging opportunities to replace them. Higher
activity levels of male song sparrows may be a result of increased territorial behaviour
(prior to their social partners entering oestrous), which has been shown to result in
reduced male energetic state (Komdeur 2001). Females appeared to differ from males in
several ways, though these differences were not due to egg laying per say, as birds in this
study were sampled an average of 31 days (range 56-12) before laying their first egg and
would not have started to produce eggs or have started to nest build. In contrast to males,
females sampled closer to their date of egg-laying had higher fat scores, yet their lower
triglyceride levels suggest they were not putting on fat at the time of sampling. Closer to
egg-laying females also had higher uric acid, indicating potentially higher dietary
consumption of protein (or catabolism), as well as higher glucose scores. Glucose is
known to power leg muscles in birds (Butler and Bishop 2000), and in contrast to males,
32�

high glucose but not high fatty acids, may indicate that females are more actively
foraging (song sparrows predominantly forage on the ground through hopping and
scratching) for protein (high uric acid) or other nutrients like calcium that are known to
be important in egg-laying (Graveland et al. 1994, Selman and Houston 1996). The lower
levels of Ig of females closer to egg-laying may result from increased locomotor activity
(Pedersen and Hoffman-Goetz 2000, Merino et al. 2006) associated with increased
foraging (see below for further discussion of Ig). The reduced hematocrit observed in
female song sparrows as lay date approached may also result from an increase in
locomotor activity (Birkhead et al. 1998), however inCrease oxygen demands (generally
associated with increased activity) are also reported to increase hematocrit (Viscor et a1.
1985).
2.4.5 Physiology and Initiation Date
Out of the suite of 15 physiology traits only Ig was significantly related to
initiation date. Here, females that laid earlier in the season had lower 19 scores. This may
indicate that early laying birds had lower parasite loads as blue tits Cyanistes caeruleus
infected with the blood parasite Haemoproteus were reported to have elevated Ig scores
(Ots and Horak 1998) and antibiotic-treated blue tits were found to have fewer parasites
and lower 19 relative to controls (Tomas et al. 2007). However, interpreting Ig scores can
be difficult as low scores have been reported for animals considered in both good (see
above) and poor health (Apanius and Nisbet 2006, Merino et al. 2006). In our case, no
traits other than 19 were strongly related to initiation date, giving us no reference point to
further interpret why early layers had lower Ig. It is surprising that other physiological
33�

traits were not correlated with initiation date, particularly energetic measures, as lay date
is thought to be condition dependent (Kalmbach et al. 2004). Potentially other factors like
age/experience, which are known to affect initiation date (Vergara et al. 2007), may not
be directly related to physiological state and therefore obscure potential relationships
between physiology and lay date. However, males paired to females that initiated
breeding earlier did appear to be in better condition based on higher levels of antioxidants
and fat and tended to have higher hemoglobin. This may indicate that male quality
influences female lay date (Bearhop et al. 1999) or potentially that pairs that started
breeding earlier had higher quality territories, and the physiological signature of territory
quality was, for some reason, only evident in males at the time of sampling.
2.5 References
Acquarone, C., M. Cucco, and G. Malacarne. 2002. Annual variation of immune condition in the Hooded Crow (Corvus carone cornix). Journal Fur Ornithologie 143:351-355.
Alonso-Alvarez, c., S. Bertrand, G. Devevey, J. Prost, B. Faivre, and G. Sorci. 2004. Increased susceptibility to oxidative stress as a proximate cost of reproduction. Ecology Letters 7:363-368.
Andersson, M. and L. Gustafsson. 1995. Glycosylated hemoglobin - a new measure of condition in birds. Proceedings of The Royal Society of London Series BBiological Sciences 260:299-303.
Apanius, V. and I. C. T. Nisbet. 2006. Serum immunoglobulin G levels are positively related to reproductive performance in a long-lived seabird, the common tern (Sterna hirundo). Oecologia 147:12-23.
Araujo, F. B., D. S. Barbosa, C. Y. Hsin, R. C. Maranhao, and D. S. P. Abdalla. 1995. Evaluation of oxidative stress in patients with hyperlipidemia. Atherosclerosis 117:61-71.
34

Ardia, D. 2005. Super size me: an experimental test of the factors affecting lipid content and the ability of residual body mass to predict lipid stores in nestling European Starlings. Functional Ecology 19:414-420.
Atkinson, S. and M. A. Ramsay. 1995. The effects of prolonged fasting of the bodycomposition and reproductive success of female polar bears (Ursus-maritimus). Functional Ecology 9:559-567.
Bearhop, S., R. Griffiths, K. Orrm, and R. W. Furness. 1999. Mean corpuscular volume (MCV) as a measure of condition in birds. Ecology Letters 2:352-356.
Benjamini, Y. and Y. Hochberg. 1995. Controlling the false discovery rate - a practical and powerful approach to multiple testing. Journal of the Royal Statistical Society Series B-Methodological 57:289-300.
Bety, J., G. Gauthier, and J. F. Giroux. 2003. Body condition, migration, and timing of reproduction in snow geese: A test of the condition-dependent model of optimal clutch size. American Naturalist 162: 110-121.
Birkhead, T. R., F. Fletcher, and E. 1. Pellatt. 1998. Sexual selection in the zebra finch Taeniopygia gut/ata: condition, sex traits and immune capacity. Behavioral Ecology and Sociobiology 44: 179-191.
Blums, P., J. D. Nichols, J. E. Hines, M. S. Lindberg, and A. Mednis. 2005. Individual quality, survival variation and patterns of phenotypic selection on body condition and timing of nesting in birds. Oecologia 143:365-376.
Bourgeon, S., T. RacIot, Y. Le Maho, D. Ricquier, and F. Criscuolo. 2007. Innate immunity, assessed by plasma NO measurements, is not suppressed during the incubation fast in eiders. Developmental & Comparative Immunology 31:720728.
Breuner, C. W. and M. Orchinik. 2002. Plasma binding proteins as mediators of corticosteroid action in vertebrates. Pages 99-112. Soc Endocrinology.
Brinkhot~ M. W. G., A. J. Cave, S. Daan, and A. C. Perdeck. 2002. Timing of current reproduction directly affects future reproductive output in European coots. Evolution 56:400-411.
Brown, M. E. 1996. Assessing body condition in birds. Pages 67-135 in V. Nolan and E. D. Ketterson, editors. Current Ornithology. Plenum Press, New York.
Butler, P. J. and C. M. Bishop. 2000. Flight. Pages 391-435 in C. G. Whittow, editor. Sturkie's avian physiology. Academic, San Diego.
Campbell, T. W., editor. 1995. Avian Hematology and Cytology. 2nd edition. Iowa State Press Ames.
Cohen, A., K. Klasing, and R. Ricklefs. 2007. Measuring circulating antioxidants in wild birds. Comparative Biochemistry and Physiology Biochemistry & Molecular Biology 147:110-121.
35

Daan, S., C. Dijkstra, and J. M. Tinbergen. 1990. Family-planning in the kestrel ifalcotinnunculus) - the ultimate control of covariation of laying date and clutch size. Behaviour 114:83-116.
DeSante, D. F., K. M. Burton, P. Velez, D. Froehlich, and D. R. Kaschube. 2008. MAPS Manual: 2008 Protocol. Contribution No. 127 of the Institute for Bird Populations.
Drabkin, D. L. and .T. H. Austin. 1932. Spectrophotometric studies I. Spectrophotometric constants for common hemoglobin derivatives in human, dog, and rabbit blood. Journal of Biological Chemistry 98:719-733.
Erel, O. 2004. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clinical Biochemistry 37:277-285.
Erel, O. 2005. A new automated colorimetric method for measuring total oxidant status. Clinical Biochemistry 38: 11 03-1111.
Gaston, A. J. and I. L. Jones. 1989. The relative importance of stress and programmed anorexia in determining mass-loss by incubating ancient murrelets. Auk 106:653658.
Glazier, D. 2000. Is fatter fitter? Body storage and reproduction in ten populations of the freshwater amphipod Gammarus minus. Oecologia 122:335-345.
Graveland, .T., R. Vanderwal, .T. H. Vanbalen, and A. J. Vannoordwijk. 1994. Poor reproduction in forest passerines from decline of snail abundance on acidified soils. Nature 368:446-448.
Green, A. 2001. Mass/length residuals: Measures of body condition or generators of spurious results? Ecology 82:1473-1483.
Hauptmanova, K., I. Literak, and E. Bartova. 2002. Haematology and 1eucocytozoonosis of great tits (Pants major L.) during winter. Acta Veterinaria Bmo 71: 199-+.
Hayes, J. P. and S. J. Shonkwiler. 2001. Morphological indicators of body condition: useful or wishful thinking? in J. R. Speakman, edi tor. Body composition analysis of animals: a handbook of non-destructive methods. Cambridge University Press, Cambridge.
Horak, P., S. Jenni-Eiermann, I. Ots, and L. Tegelmann. 1998. Health and reproduction: The sex-specific clinical profile of great tits (Pants major) in relation to breeding. Canadian Journal of Zoology 76:2235-2244.
Jenni-Eiennann, S. and L. Jenni. 1997. Diurnal variation of metabolic responses to shorttenn fasting in passerine birds during the postbreeding, molting and migratory period. Condor99:1J3-122.
36�

Kalmbach, E., R. Griffiths, J. E. Crane, and R. W. Furness. 2004. Effects of experimentally increased egg production on female body condition and laying dates in the great skua Stercorarius skua. Journal of Avian Biology 35:501-514.
Katsuki, A., Y. Sumida, H. Urakawa, E. C. Gabazza, S. Murashima, K. Nakatani, Y. Yano, and Y. Adachi. 2004. Increased oxidative stress is associated with serum levels of triglyceride, insulin resistance, and hyperinsulinemia in Japanese metabolically obese, nonnal-weight men. Diabetes Care 27:631-632.
Kern, M., W. Bacon, D. Long, and R. J. Cowie. 2005. Blood metabolite and corticosterone levels in breeding adult Pied Flycatchers. Condor 107:665-677.
Kilgas, P., R. Mand, M. Magi, and V. Tilgar. 2006. Hematological parameters in broodrearing great tits in relation to habitat, multiple breeding and sex. Comparative Biochemistry and Physiology Part A Molecular & Integrative Physiology 144:224-231.
Komdeur, J. 2001. Mate guarding in the Seychelles warbler is energetically costly and adjusted to paternity risk. Proceedings of The Royal Society of London Series BBiological Sciences 268:2103-2111.
Krebs, C. J. 1999. Ecological methodology. 2n'd edition. Benjamin & Cummings, Menlo Park.
Love, O. P., C. W. Breuner, F. Vezina, and T. D. Williams. 2004. Mediation of a corticosterone-induced reproductive conflict. Honnones and Behavior 46:59-65.
Love, O. P., E. H. Chin, K. E. Wynne-Edwards, and T. D. Williams. 2005. Stress honnones: A link between maternal condition and sex-biased reproductive investment. American Naturalist 166:751-766.
Martinez, J., G. Tomas, S. Merino, E. Arriero, and J. Moreno. 2003. Detection of serum immunoglobulins in wild birds by direct ELISA: a methodological study to validate the technique in different species using antichicken antibodies. Functional Ecology 17:700-706.
Masello,1. F. and P. Quillfeldt. 2004. Are haematological parameters related to body condition, ornamentation and breeding success in wild burrowing parrots Cyanoliseus patagonus? Journal of Avian Biology 35:445-454.
Mazor, R., R. Shurtz-Swirski, R. Farah, B. Kristal, G. Shapiro, F. Dorlechter, M. CohenMazor, E. Meilin, S. Tamara, and S. Sela. 2008. Primed polymorphonuclear leukocytes constitute a possible link between inflammation and oxidative stress in hyperlipidemic patients. Atherosclerosis 197: 93 7-943.
Merino, S., J. Moreno, G. Tomas, J. Martinez, J. Morales, J. Martinez-De la Puente, and J. L. Osorno. 2006. Effects of parental effort on blood stress protein HSP60 and immunoglobulins in female blue tits: a brood size manipulation experiment. Journal of Animal Ecology 75:1147-1153.
37

Myers, J. L. and A. D. Well. 2003. Research design and statistical analysis. 2nd edition. Lawrence Erlbaum Associates, London.
Nadolski, J., J. Skwarska, A. Kalinski, M. Banbura, R. Sniegu1a, and J. Banbura. 2006. Blood parameters as consistent predictors of nestling performance in great tits (Pants major) in the wild. Comparative Biochemistry And Physiology AMolecular & Integrative Physiology 143:50-54.
Ninni, P., F. de Lope, N. Saino, C. Haussy, and A. P. Moller. 2004. Antioxidants and condition-dependence of arrival date in a migratory passerine. Oikos 105:55-64.
Norberg, R. A. 1981. Temporary weight decrease in breeding birds may result in more fledged young. American Naturalist 118:838-850.
Ots, 1. and P. Horak. 1998. Health impact of blood parasites in breeding great tits. Oeco10gia 116:441-448.
Owen, J. c., M. K. Sogge, and M. D. Kern. 2005. Habitat and sex differences in physiological condition of breeding Southwestern Willow Flycatchers (Empidonax traillii extimus). Auk 122:1261-1270.
Pedersen, B. K. and L. Hoffman-Goetz. 2000. Exercise and the immune system: regulation, integration, and adaptation. Physiological Reviews 80: I055-1 081.
Piersma, T. E. J. M. 1996. Build-up of red blood cells in refuelling bar-tailed godwits in relation to individual migratory quality. Condor 98:363-370.
Romero, M. L. and M. Wekelski. 2001. Corticosterone levels predict survival probabilities of Galapagos marine iguanas during E1 Nino events. Proceedings of the National Academy of Sciences of the United States of America 98:7366-7370.
Sanchez-Guzman, J. M., A. Villegas, C. Corbacho, R. Moran, A. Marzal, and R. Real. 2004. Response of the hematocrit to body condition changes in Northern Bald Ibis Geronticus eremita. Comparative Biochemistry And Physiology A-Molecular & Integrative Physiology 139:41-47.
Sanz, J. J. 1999. Seasonal variation in reproductive success and post-nuptial moult of blue tits in southern Europe: an experimental study. Oecologia 121:377-382.
Schulte-Hostedde, A., B. Zinner, 1. S. Millar, and G. 1. Hickling. 2005. Restitution of mass-size residuals: Validating body condition indices. Ecology 86: 155-163.
Seaman, D., C. G. Guglielmo, R. W. Elner, and T. D. Williams. 2006. Landscape-scale physiology: Site differences in refueling rates indicated by plasma metabolite analysis in free-living migratory sandpipers. Auk 123:563-574.
Seeman, T. E., B. S. McEwen, J. W. Rowe, and B. H. Singer. 2001. Allostatic load as a marker of cumulative biological risk: MacArthur studies of successful aging. Proceedings of the National Academy of Sciences of the United States of America 98:4770-4775.
38

Selman, R. and D. C. Houston. 1996. The effect of prebreeding diet on reproductive output in zebra finches. Proceedings of the Royal Society of London Series B Biological Sciences 263: 1585-1588.
Smith, R. and F. R. Moore. 2003. Arrival fat and reproductive perfonnance in a longdistance passerine migrant. Oecologia 134:325-331.
Spengler, 1., P. L. Leberg, and W. C. Barrow. 1995. Comparison of condition indexes in migratory passerines at a stopover site in coastal louisiana. Condor 97:438-444.
Steiger, J. H. 1979. MULTICORR: A computer program for fast, accurate, small-sample testing of correlational pattern hypotheses. Educational and Psychological Measurement 39:677-680.
Steiger, J. H. 1980. Tests for comparing elements of a correlation matrix. Psychological Bulletin 87:245-251.
Steiger, J. H. 2005. Comparing correlations: Pattern hypothesis tests between and/or within independent samples. Pages 377-414 in A. Maydeu-Olivares and J. J. McArdle, editors. Contemporary psychometrics: a festschrift for Roderick P. McDonald Lawrence Erlbaum Associates, Mahwah.
Tomas, G., S. Merino, J. Moreno, J. Morales, and J. Martinez-de la Puente. 2007. Impact of blood parasites on immunoglobulin level and parental effort: a medication field experiment on a wild passerine. Functional Ecology 21:125-133.
Totzke, U., M. Fenske, O. Huppop, H. Raabe, and N. Schach. 1999. The influence of fasting on blood and plasma composition of herring gulls (Larus argentatus). Physiological And Biochemical Zoology 72:426-437.
Vergara, P., J. I. Aguirre, and M. Fernandez-Cruz. 2007. Arrival date, age and breeding success in white stork Ciconia ciconia. Journal of Avian Biology 38:573-579.
Verhulst, S. and J. M. Tinbergen. 1991. Experimental-evidence for a causal relationship between timing and success of reproduction in the great tit Parus-m-major. Journal of Animal Ecology 60:269-282.
Viscor, G., M. S. Marques, and J. Palomeque. 1985. Cardiovascular and organ weight adaptations as related to flight activity in birds. Comparative Biochemistry and Physiology 82:597-599.
Williams,1. D., C. G. Guglielmo, O. Egeler, and C. J. Martyniuk. 1999. Plasma lipid metabolites provide infonnation on mass change over several days in captive Western Sandpipers. Auk 116:994-1000
39

Figure 2.1 Male and female mean ± SE of all distinct correlations presented in Table 2.1.
1.0
~
"'-""- 0.8 c Q)
u :E Q)
0.6 0 u c 0
-..:::; 0.4 CO Q) "" - ~
0 0
0.2
0.0 ,
Females Males
40�
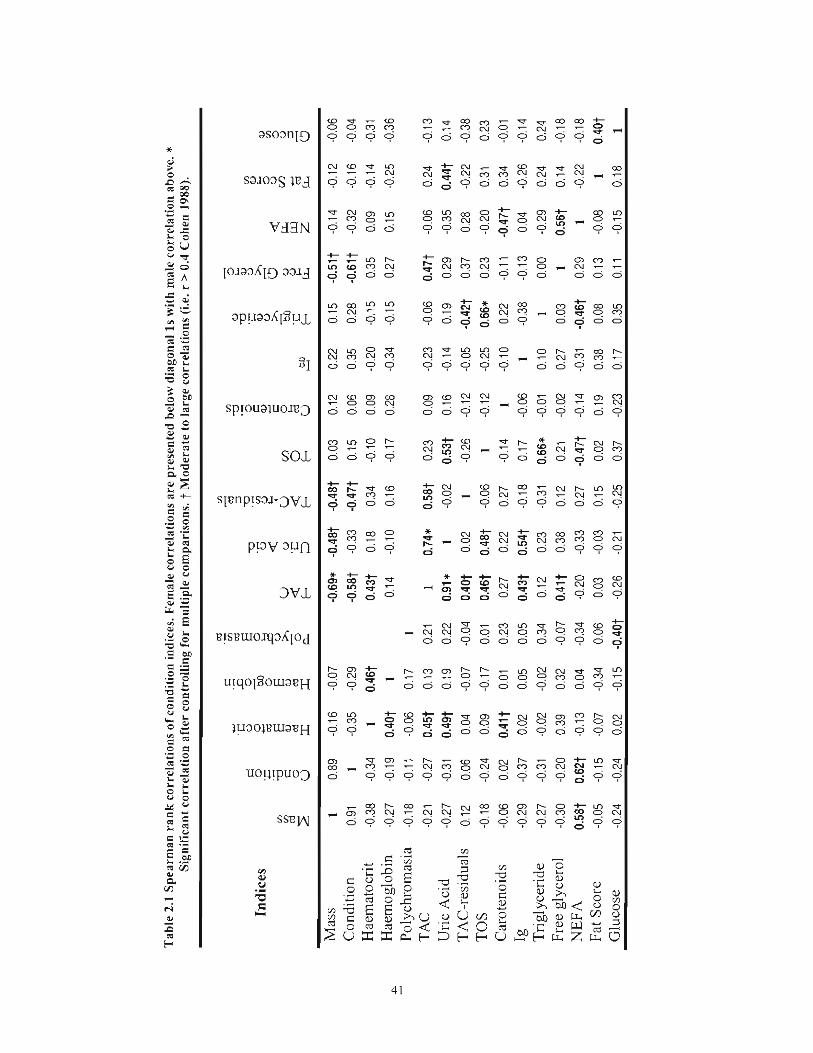
Tab
le 2
.1 S
pea
rman
ran
k c
orre
lati
ons
of c
ondi
tion
ind
ices
. F
emal
e co
rrel
atio
ns a
re p
rese
nted
bel
ow d
iag
on
alls
wit
h m
ale
corr
elat
ion
abov
e. *
S
igni
fica
nt c
orre
lati
on a
fter
con
trol
ling
for
mul
tipl
e co
mpa
riso
ns. t
Mo
der
ate
to l
arge
cor
rela
tion
s (i
.e. r>
0.4
Coh
en 1
988)
.
- cd
VJ
Indi
ces
VJ
VJ
cd
~
I=:
0 ....., 'U
I=:
0 u
...., 'r::
()
0 .....
cd 6 Il
) cd
.:r::
t::
B
0 ~
0 6 Il)
cd
.:r::
'[;j
cd
6 0 E
()
>--.
0 0...
U « b
-0
'0 « (
)
'i: :::J
'@
;:l
-0
'[;j
Il
) .....
I U « b
if)
0 b
C/"J
'U
0 c: <l) ...., c: 0 @
U
CD
.....
..
Il)
'U
'i:
Il)
()
>-.
b1 'r::
b
-0
.....
Il)
()
>--.
<3 Il)
Il) .....
f.L<
« ~ U-l
Z
<.rJ
Il
) .... 0 (
)
if)
......
co;:l
f.L<
Il)
VJ 0 ()
;:l - 0
Mas
s I
0.89
-0
.16
·007
-0
.69*
-0
.48t
-0
.48t
0.
03
0.12
0.
22
0.15
-0
.51t
-0
.14
-0.1
2 -0
.06
Con
diti
on
0.91
I
-0.3
5 -0
.29
-0.5
8t
-0.3
3 -O
,4lt
0.15
0.
06
0.35
0.
28
-0.6
1t
-0.3
2 -0
.16
-0.0
4 H
aem
atoc
rit
-0.3
8 -0
.34
I 0,
46t
0.43
t 0.
18
0.34
-0
.10
0.09
-0
.20
-0.1
5 0.
35
0.09
-0
.14
-0.3
1
.j>
. -H
aem
oglo
bin
-0.2
7 -0
.19
0.40
t 1
0.14
-0
.10
0.16
-0
.17
0.28
-0
.34
-0.1
5 0.
27
0.15
-0
.25
-0.3
6 P
olyc
hrom
asia
-0
.18
-0.1
1 -0
.06
017
I T
AC
-0
.21
-0.2
7 0.
45t
0.13
0.
21
1 0.
74*
0.58
t 0.
23
0.09
-0
.23
-0.0
6 0.
47t
-0.0
6 0.
24
-0.1
3 U
ric
Aci
d -0
.27
-0.3
1 0.
49t
0.19
0.
22
0.91
* I
-0.0
2 0,
53t
0.16
-0
.14
0.19
0.
29
-0.3
5 0.
44t
0.14
T
AC
-res
idua
l s
0.12
0.
06
0.04
-0
.07
-0.0
4 0.
40t
0.02
I
-0.2
6 -0
.12
-0.0
5 -0
,42t
0.
37
0.28
-0
.22
-0.3
8 T
OS
-0
.18
-0.2
4 0.
09
-0.1
7 0.Q
1 0.
46t
0.48
t -0
.06
1 -0
.12
-0.2
5 0,
66*
0.23
-0
20
031
0.23
C
arot
enoi
ds
-0.0
6 0.
02
0.41
t 0.
01
0.23
0.
27
0.22
0.
27
-0.1
4 1
-0.1
0 0.
22
-0.1
1 -0
,47t
0.
34
-0.0
1 [g
-0
.29
-037
0.
02
0.05
0.
05
0.43
t 0.
54t
-0.1
8 0.
17
-0.0
6 1
-0.3
8 -0
.13
0.04
-0
.26
-014
T
rig]
yce
ride
-0
.27
-0.3
1 -0
.02
-0.0
2 0.
34
0.12
0.
23
-0.3
1 0.
66*
-0.0
1 0.
10
1 00
0 -0
.29
0.24
0.
24
Fre
e gl
ycer
ol
-0.3
0 -0
20
0.39
03
2 -0
07
0.41
t 0.
38
0.12
0.
21
-0.0
2 0.
27
0.03
I
0.56
t 0.
14
-0.1
8 N
EF
A
0.58
t 0.
62t
-0.1
3 0.
04
-0.3
4 -0
.20
-0.3
3 0.
27
-0.4
7t
-0.1
4 -0
.31
·0.4
6t
0.29
1
-022
-0
.18
Fat
Sco
re
-0.0
5 -0
.15
-0.0
7 -0
.34
006
0.03
-0
03
0.15
0.
02
0.19
0.
38
0.08
0.
13
-0.0
8 1
0.40
t G
luco
se
-0.2
4 -0
.24
0.02
-0
.15
-0.4
0t
-0.2
6 -0
.21
-0.2
5 0.
37
-0.2
3 0.
17
0.35
0.
11
-0.1
5 0.
18
1
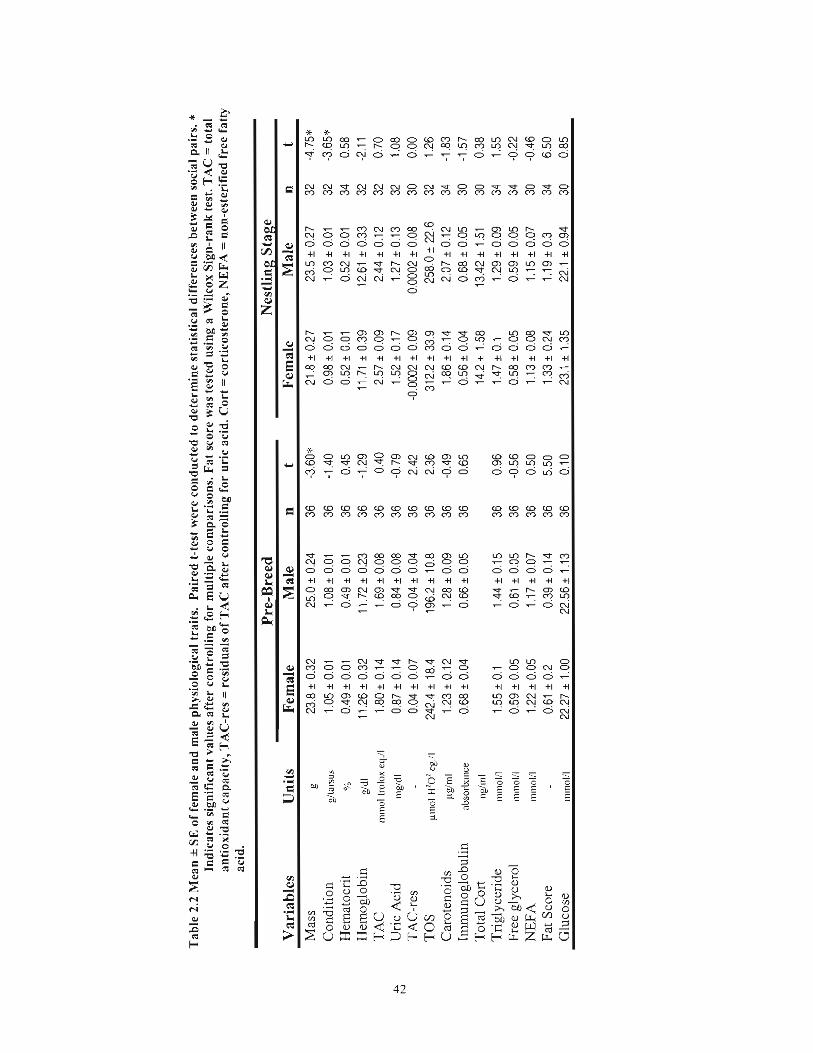
Tab
le 2
.2 M
ean
± S
E o
f fe
mal
e an
d m
ale
phys
iolo
gica
l tr
aits
. P
aire
d t-
test
wer
e co
nduc
ted
to d
eter
min
e st
atis
tica
l di
ffer
ence
s be
twee
n so
cial
pai
rs. *
Indi
cate
s si
gnif
ican
t va
lues
aft
er c
ontr
olli
ng f
or m
ulti
ple
com
pari
sons
. F
at s
core
was
tes
ted
usin
g a
Wil
cox
Sig
n-r
ank
test
. TA
C =
= to
tal
anti
oxid
ant
capa
city
, TA
C-r
es =
= re
sidu
als
of T
AC
aft
er c
ontr
olli
ng f
or u
ric
acid
. C
ort
== c
orti
cost
eron
e, N
EF
A =
= no
n-es
teri
fied
fre
e fa
tty
acid
.
Pre
-Bre
ed
Nes
tlin
g S
tage
V
aria
bles
U
nits
F
emal
e M
ale
n t
Fem
ale
Mal
e n
t M
ass
g 23
8 ±
0.3
2 25
0 ±
0.2
4 36
-3
.60*
21
.8 ±
0.2
7 23
.5 ±
0.2
7 32
-4
.75*
C
ondi
tion
g!
lars
us
1.05
± 0
.01
1.08
± 0
.01
36
-1.4
0 0.
98 ±
0.0
1 1.
03 ±
0.0
1 32
-3
.65*
H
emat
ocri
t %
0.
49 ±
0.0
1 0.
49 ±
0.0
1 36
0.
45
0.52
± 0
.01
0.52
± 0
.01
34
0.58
H
emog
lobi
n gl
dl
11.2
6 ±
0.3
2 11
.72
± 0
.23
36
-1.2
9 11
.71
± 0
.39
12.6
1 ±
0.3
3 32
-2
.11
TA
C
Illil
lollr
olox
eq.
!1
1.80
±0.1
4 1.
69 ±
008
36
0.
40
2.57
± 0
.09
2.44
±0.1
2 32
0.
70
N+:-
Uri
c A
cid
Illg/
dl
0.87
± 0
.14
0.84
± 0
.08
36
-0.7
9 1.
52 ±
0.1
7 1.
27±0
.13
32
1.08
T
AC
-res
0.
04 ±
0.0
7 -0
.04
± 0
.04
36
2.42
-0
.000
2 ±
0.0
9 0.
0002
± 0
08
30
0.00
T
OS
~'
'l10
1
H'O
' eg.
!1
242.
4 ±
18.
4 19
6.2
± 1
0.8
36
2.36
31
2.2
± 3
3.9
258.
0 ±
22.
6 32
1.
26
Car
oten
oids
'lg
lml
1.23
±0.1
2 1.
28 ±
0.0
9 36
-0
.49
1.86
± 0.
14
2.07
± 0
.12
34
-1.8
3 Im
mun
oglo
buli
n ab
sorb
ance
0.
68 ±
0.0
4 0.
66 ±
0.0
5 36
0.
65
0.56
± 0
.04
0.68
± 0
.05
30
-1.5
7 T
otal
Cor
t ng
!1ll1
14
.2 ±
1.5
8 13
.42
± 1
.51
30
0.38
T
rigl
ycer
ide
III Il
lOIII
1.
55 ±
0.1
1.
44 ±
015
36
09
6 1.
47±0
.1
1.29
± 0
.09
34
1.55
F
ree
glyc
erol
Ill
lllol
Il 0.
59 ±
0.0
5 0.
61 ±
0.0
5 36
-0
.56
0.58
± 0
.05
0.59
± 0
.05
34
-0.2
2 N
EF
A
III Il
lO V
I 1.
22 ±
0.0
5 1.
17±0
.07
36
0.50
1.
13±0
.08
1.15
± 0
.07
30
-0.4
6 F
at S
core
0.
61 ±
0.2
0.
39±0
.14
36
5.50
1.
33 ±
0.2
4 1.
19 ±
0.3
34
6.
50
Glu
cose
II
I IllO
III
22.2
7 ±
1.0
0 22
.56±
1.1
3 36
0.
10
23.1
± 1
.35
22.1
± 0
.94
30
085

Table 2.3 Summary of regression analysis for female and male physiological traits correlated with the days until initia tion of laying. NEFA =non-esterified free fa tty acids. TAC-resid uals = antioxidant capacity of plasma controlling for uric acid.
Variables Beta SE of Beta t P
Female Measures Hematocrit 0.40 0.09 4.27 0.001 Tliglycerides 0.34 0.09 3.79 0.002 Glucose -0.35 0.09 -3.67 0.003 Fat Score -0.25 0.09 -2.81 0.02 Uric Acid -0.49 0.13 -3.72 0.003 Immunoglobulin 0.43 0.14 3.04 0,009 TAC-residual 0,18 0.08 2,11 0.055
Male Measures Fat Score 0.29 0.10 3.03 0.01 NEFA -0.28 0.11 -2.59 0.02 TAC-residuals 0.22 0,10 2.24 0.045 Triglycerides -0.13 0.10 -1,23 0.24 Hemoglobin 0.12 0,10 1,13 0.28
43

CHAPTER 3 EXPERIMENTAL EVIDENCE THAT NEST PREDATION AFFECTS CLUTCH SIZE AND THE COST OF REPRODUCTION IN A FREE LIVING SONG BIRD
Marc Travers l,2, Michael Clinchy3, Tony D. Williams l
, Liana Zanette2
lSimon Fraser University, Department of Biological Science, Burnaby, B.C., Canada 2University of Western Ontario, Department of Biological Science, London, Ontario, Canada 3University of Victoria, Department of Biological Science, Victoria, B.c. Canada
44

3.1 Introduction
For decades ecologists have been interested in whether clutch size in birds is
affected more by food availability or nest predation (Lack 1947, Skutch 1949, Pemns
1977, Martin 1987, Newton 1998). Whereas numerous food supplementation experiments
have demonstrated that food availability has proximate effects on clutch size (reviewed in
Nager 2006), only two experimental studies to date have shown that nest predation can
have proximate effects on clutch size. Eggers et al. (2006) showed that Siberian jays
(Perisoreus in/austus) reduced their clutch size in response to recordings of nest predator
calls presented on their territories. Doligez and Colbert (2003) experimentally elevated
nest predation rates and displayed predator models to collared flycatchers (Ficedula
albicollis) and reported a reduced clutch size for the population in the subsequent year.
In both studies clutch size effects were reported to be in response to predator cues
indicating the presence of predators rather than an individual's direct experience of nest
predation (i.e. offspring loss). To the best of our knowledge, no experimental study to
date has demonstrated that an individual's experience of nest predation affects the size of
subsequent clutches. Because direct nest predation is a common experience across many
avian species (Ricklefs 1969, Martin and Li 1992, Martin 1995), such data are critical for
furthering our understanding of predation effects on clutch size.
Martin (1995) conducted a comparative study examining the relationship between
direct nest predation and clutch size in 123 species of Passeriformes and Piciformes, and
reported an inverse correlation between direct nest predation and clutch size. Marti n
(1995) also found that nest predation was more strongly correlated with clutch number
(i.e. the number of clutches laid in a season) than with clutch size and that clutch number
45�

and clutch size were inversely correlated. Zanette et a!. (2006b) reported results from a
bi-factorial food supplementation and natural predator reduction experiment on song
sparrows (Melospiza melodia) that suggested direct nest predation may have proximate
effects on clutch size similar to those described by Martin (1995) at the inter-specific
level. Zanette et a1. (2006b) found that nest predation increased clutch number and that
clutch number and average clutch size were inversely correlated, which suggested that
birds under high nest predation were forced to re-nest more frequently and consequently
laid smaller clutches in each nest (Zanette et a1. 2006b). These studies suggest that direct
nest predation may in fact reduce clutch size, but that clutch size effects may be the result
of predation effects on clutch number.
After a predation event, many avian species can re-nest (Grzybowski and Pease
2005). While re-nesting increases the chances that parents will successfully rear at least
one offspring in a breeding season, re-nesting necessarily leads to an increase in clutch
number and therefore egg production. Martin (1995) proposed that increases in egg
production as a consequence of predation may increase the energetic demands on
female's, which may lead to energy trade-offs resulting in the inverse relationship
between clutch number and clutch size; i.e. females not able to obtain sufficient resources
to meet the demands of increases in clutch number lay smaller clutches. Clearly increases
in egg production require females to obtain some amount of additional resources, and
supplemental feed high in energy and nutrients has been shown to increase clutch size in
song sparrows (Zanette et a1. 2006b) and other species (Nager 2006). However, Zanette et
al. (2006b) reported a similar inverse relationship between clutch number and clutch size
in both food supplemented and non food supplemented song sparrows; suggesting that, at
46�

least at the intra-specific level, the inverse relationship between clutch number and clutch
size is not the result of a resource-based mechanism. Therefore, experimentally
examining the effects of nest predation on clutch number and clutch size when food is
super abundant, is necessary to determine if direct nest predation does in fact affect
clutch size, and whether, as suggested by Zanette et a1. (2006b), the effects can occur
when food is abundant.
The demands of increased egg production have been shown experimentally to
negatively affect a female's physiological condition (Kalmbach et a1. 2004) as well as the
quality of the offspring she can produce; indicated by reduced hatching success
(Kalmbach et a1. 2004) and lower mass of nestlings that do hatch (Monaghan et a1. 1995).
The costs of egg laying are traditionally assumed to result from resource based trade-offs
where females allocate a limited amount of resources between egg production and their
own physiological systems (Williams 1966); thus, resource allocation trade-offs should
be evident only when resources are scarce. More recently, it has been proposed that
reproduction itself, or the regulatory (physiological) processes controlling reproduction,
may generate physiological costs to the female (Partridge et al. 2005, Harshman and Zera
2007) which might be independent of allocation of resources per se (i.e. non-resource
based costs, Williams (2005)). As one example, the hormone estradiol (E2) is necessary
to produce eggs but increased levels during egg production can cause a transient
inhibition of red blood cell production resulting in reproduction-induced anaemIa
(Williams et a1. 2004, Wagner et a1. 2008a). Despite some progress, the physiological
mechanisms underlying the costs of egg production remain poorly understood (Williams
2005, Harshman and Zera 2007). Therefore, incorporating measurements of physiological
47

condition into experimental studies which manipulate egg production and resource (food)
availability might identify specific mechanisms underlying costs of egg production (e.g.
resource vs non-resource) and if this is done in the context of nest predation may further
our understanding of the ways in which predation can impact prey. Furthennore, if the
inverse relationship between clutch number and clutch size was the result of an energetic
trade-off (sensu Martin 1995) we would expect that nest predation induced increases in
clutch number/egg production would result in either females ending up in a poor
energetic state, or that their offspring were be of poorer quality, or both. Yet, if costs of
egg production exist even when females are not limited by food availability, the
possibility would remain that the inverse relationship between clutch number and clutch
size is the result of egg production costs (Martin 1995), but of a non-resource based type.
We conducted a clutch removal manipulation to examine the effects of
experimental nest predation on clutch number and clutch size in a multi-brooded species,
the song sparrow (Melospiza melodia). Following Zanette et aI's (2006b) study we were
specificall y interested in testing the effects of nest predation independent of food
availability and so we provided all birds with un-limited access to high quality food
throughout the entire breeding season. In addition to testing the affects of nest predation
on clutch size, we examined the cumulative affects of nest predation on total seasonal egg
production to investigate whether predation resulted in increased physiological 'costs of
egg production' for mothers as well as reduced hatching success and growth of offspring.
We assessed physiological costs of egg production using a range of physiological traits
which should reflect the specific mechanisms introduced above (resource-based
mechanisms affecting energetic balance, and non-resource based mechanisms affecting
48�

oxidative stress and hematology), along with general indices of condition. We predicted
that in food-supplemented females with access to ad libitum high quality food; 1)
experimentally-increased nest predation will increase clutch number and decrease clutch
size, 2) that high-predation females with a higher frequency of re-nesting (i.e. elevated
total seasonal egg production) will be in poorer physiological condition reflecting
increased costs of egg production, and 3) because of food supplementation, we expect
costs will be consistent with non-resource rather than resource based hypotheses for the
costs of egg production. Finally, Zanette et al. (2006a,b) showed that food availability
reduces the occurrence of nest predation. If nest predation affects clutch size even when
food is not limiting, then because food affects nest predation, food and predators would
have combined effects on clutch size. Whereas speaking of either food or predator effects
may help us disentangle specific mechanisms, in many real-world situations this either/or
likely represents a false dichotomy (Zanette et al. 2006b).
3.2 Methods
3.2.1 Study Species
We worked on a colour banded population of song sparrows (Melospiza melodia)
on the Southern Gulf Islands, British Columbia, Canada. Song sparrows in this area are
resident year round and multi-brooded. Song sparrows in the region can successfully rear
3-4 broods, and when nest predation is high females can produce up to 8 clutches and 28
eggs in a season (Zanette et aJ. 2006b). Clutch size can range from 2-5 eggs but IS
typically 3-4 eggs and egg-laying typically begins in April and ends in early July.
49�

3.2.2 Food Supplementation
As described in the introduction, we were interested in investigating the effects of
nest predation on clutch size and physiological costs of egg production independent of
food availability, i.e. at high-resource levels. To remove the potential effects of limiting
resources on egg production and female condition we provided ad libitum supplemental
food (from March 31 to the end of the breeding season) to all females via feeders placed
on each territory (following Zanette et a!. 2003, 2006a,b). Previous work has shown that
song sparrows supplemented with food high in carbohydrates, fat, protein, and calcium
had greater foraging efficiency (Duncan-Rastogi et a1. 2006), nested earlier and for
longer, produced larger clutches, more clutches, and increased total seasonal egg
production (Zanette et al. 2006b), were in better physiological condition (Clinchy et a!.
2004), and had higher annul reproductive success (Zanette et a1. 2003, Zanette et a!.
2006a). Here, we further improved on the supplemental food provided by adding whole
egg powder and carotenoids. Whole egg powder was used to provide the sulphur rich
amino acids (Houston et aL 1995) in the appropriate ratios (Ramsay and Houston 1998)
thought to limit clutch size and egg mass (Selman and Houston 1996). In general, whole
egg powder would also presumably provide all nutrients or ratios of nutrients impOltant
for egg production. Carotenoids have been shown to limit re-nesting in gulls (Blount et
a!. 2004) and may also have beneficial effects on physiological condition (McGraw et al.
2006) and nestling development (De Neve et a1. 2008). The feed used in this study
consisted per kilogram of: 600.90g white millet, 315.47g of high-fat high-protein pellets
(Aquamax Grower 400), 2I.03g of crushed oyster shell, 60g of dried whole egg powder,
and 2.6g of Oro Glo pigrnenter, Kemin industries (equivalent to O.04g xanthophyll
carotenoids/kg of feed). We estimated that 60g of dried whole egg is equivalent to 240g
50

of wet egg based on the 75% water content of eggs reported by Carey et al. (1980). To
continn that specitic components of the supplemental feed were consumed and absorbed
we conducted a feeding validation using plasma carotenoid levels from fed and un-fed
birds as a marker of dietary intake.
3.2.3 Experimental Manipulation of Nest Predation and Clutch Number
To experimentally manipUlate nest predation we needed to control the level of
natural nest predation. Consequently, we conducted our nest predation manipulation on
islands with comparatively low levels of natural nest predation (Zanette et al. 2006a)
where, as part of another experiment, natural nest predation was further reduced by the
removal of a principal nest predator (the brown-headed cowbird).
We experimentally manipulated nest predation to generate a High Nest Predation
(HNP; n = 14 females) and a Low Nest Predation (LNP; n = 11 females) group. To
generate high nest predation we removed all eggs from a female's nest on the 6th day of
incubation which caused females to re-nest. We repeated this process until females in the
HNP group had produced 4 or 5 clutches. We generated low nest predation by removing
all eggs from a female's nest on the 6th day of incubation and replacing them with clay
eggs fashioned to resemble those of song sparrows. Females in the LNP group incubated
the day eggs and because the eggs did not hatch, the duration of the incubation period
was extended (normal duration of incubation = 13 days; mean extension ± S. D.= 3.65 ±
1.93 days). Females in the LNP group eventually abandoned these nests, giving them the
opportunity to re-nest. We repeated the clay egg manipulation until LNP females had
produced 3 clutches over the breeding season. Incubating a clutch that does not hatch is
51

an experience females song sparrows sometimes undergo because the clutch is infertile,
but more frequently in this region this is due to a female cowbird having visited the nest
and punctured the eggs unbeknownst to the mother sparrow. Mother sparrows in this
population have consequently been recorded incubating up to an additional II days (i.e.
nearly double the normal incubation peliod; CJinchy & Zanette unpublished data). Here
our aim was to address factors that result from increased egg production, as opposed to
brood rearing. For this reason, we followed the clay egg procedure for the LNP group to
ensure that these females, like their HNP counterparts, only experienced egg-stage
related phenomenon, and did not raise young during the course of the experiment. The
term low nest predation treatment refers to how LNP nests lasted longer (rather than LNP
females actually experiencing predation), thus simulating longer nest durations that
would occur in low predation areas relative to high predation areas. That is, when the
predation rate is high nests are likely to be consumed early in the cycle. Conversely,
when nest predation is lower nest are likely to be consumed later in the cycle, thus lasting
longer.
To experimentally control for initiation date (date of the first egg laid in the
season) we assigned females to treatments by pairing birds with similar initiation dates
and then randomly assigning each female to one of the treatments. For HNP females'
penultimate clutch (3 or 4) only, we timed the removal of the clutch to ensure that HNP
females produced their final experimental clutch (clutch # 4 or 5) at the same time as
LNP females produced their third clutch.
52

3.2.4 Measurement of Physiological Traits
We captured pre-breeding song sparrows between March 6-282007 using baited
potter traps in order to obtain a baseline measure of physiological state. Once captured,
we colour banded all birds for individual identification and collected up to 300111 of blood
from the brachial vein using heparanized capillary tubes. We measured blood glucose
concentration (see below) and prepared blood smears immediately after releasing the
bird. Blood smears were air dried and stored in slide boxes containing desiccation packs.
We refiigerated blood in cooler packs for a maximum of 10 hours before separating the
plasma from the red blood cells. Plasma was stored on dry ice until pennanent laboratory
storage at -20°C. Females were captured and blood sampled a second time on the 8-1 Olh
day of incubation of their final experimental clutch using mist nets. Blood used for
corticosterone analysis was collected in under 3 min from the time the bird hit the net.
Other than capture methods, blood collection procedures and storage were identical to
pre-breeding methods. Treatments did not differ in the date of blood sampling at the
beginning (pre-breeding tlll= 0.33, P= 0.75) or end of the experiment (final clutch: tl8=
0.03, p= 0.97)
We used colorimetric assays to measure plasma levels of triglycerides, NEFA,
immunoglobulin (Ig), carotenoids, total antioxidants capacity (TAC), total oxidant status
(TOS) and whole blood levels of hemoglobin. We detennined assay variation using a hen
plasma standard. Intra-assay variation range was 4.6%, 2.7%, 2.6% 1.7%, 3%, 5.7% and
2.2% for triglyceride, NEFA, Ig, carotenoids, TAC, TOS, and hemoglobin respectively.
Inter-assay variation was 2.4%, 1.1 %, 19.4%, 5.9%, 2.3%, 9.2% for triglyceride, NEFA,
19, carotenoids, TAC, and TOS respectively (we did not obtain an inter-assay CY for
hemoglobin).
53

3.2.4.1� Energetic measures
Plasma concentrations of total triglycerides were determined in duplicate samples
with absorbance measured at 540nrn after 10min of incubation at 37°C (see Seaman et al.
2006), using Sigma-Aldrich reagents (Triglyceride reagent & free glycerol reagent,
Sigma-Aldlich Canada). Triglyceride concentrations were calculated by subtracting free
glycerol from total triglyceride. Non-esterified free fatty acids concentrations were
determined in triplicate using a NEFA kit (NEFA-HR(2); Wako Diagnostics, USA).
Blood glucose scores were measured at the time of capture using Acensia® glucose meter
(Bayer Inc). We quantified visible subcutaneous fat stores under the wing, and in the
furcular and abdominal region using a 7 point scale (DeSante et al. 2008)
3.2.4.2� Non-energetic measures
This included measures of hematological (hematocrit, hemoglobin,
polychromasia), immunological (lg), corticosterone and oxidative stress (TAC, TOS,
carotenoids) status. Hematocrit was measured using standard techniques (Campbell and
Ellis 2007) after centrifugation for lOmin using a Zipocrit (Laboratory Essentials, USA)
portable centrifuge. Hemoglobin concentration in whole blood was determined using the
cyanomethemoglobin method (Drabkin and Austin 1932) modified for a colorimetric
assay, using 5Jll of whole blood diluted in 1.25ml Drabkin's reagent (D5941, Sigma
Aldrich, Canada) with absorbance measured at 540nm. Polychromasia was counted on
smears stained with Wright-Giemsa (Sigma-Aldrich Canada, Ltd) and was calculated as
54

the proportion of red blood cells (RBC) that are immature (Campbell and Ellis 2007). A
single observer, who was blind to experimental ID, counted (>500 RBC per count) all
smears in triplicate and had an intra-count variation of <4%. Polychromasia scores and
intra-count variation were calculated from the two most similar scores of 3 repeated
counts of the same smear. Plasma immunoglobulins (lg) were detennined using the
ELISA method with commercial anti-chicken antibodies as reported by Martinez et al.
(2003). We adapted the method for use in song sparrows by determining the sparrow
appropriate plasma dilution (1/8000). 19 results are presented in units of absorbance. The
concentration of total corticosterone (see Breuner and Orchinik 2002, Love et al. 2004,
2005) in non-extracted plasma was detennined using a corticosterone Enzyrne-linked
immunoabsorbent Assay (EIA - Assay Designs Inc., Michigan USA, catalog # 901-097)
with a 4-parameter logistic fit. Total antioxidant capacity (TAC) was detennined using a
modified Randox-TEAC assay described by Ere1 (2004). TAC results are reported in
mmol Trolox equivalent.L- 1 (238813, Sigma-Aldrich). Total oxidative status (TOS) was
determined as described by Erel (2005). TOS results are reported in Ilmo1 H202
equivalent.C'. We prepared the carotenoid assay (see Alonso-Alvarez et al. 2004) by
vortexing 20111 of plasma diluted in 180111 of absolute ethanol, centrifuged the
plasma/ethanol at 1500g for 10min, and recovered the supernatant. Plasma carotenoid
concentrations were detennined in triplicate using SOIlI of supernatant per well, with
absorbance measured at 450nm and compared with a standard curve of lutein (minimum
70% xanthophyll from alfalfa; Sigma-Aldrich).
55

3.2.5 Hatching Success and Nestling Growth
On the final experimental clutch (clutch 3 for LNP and clutch 4 or 5 for HNP) we
removed one egg from each nest for egg composition analysis (results not discussed in
this paper) and the remaining eggs were left un-manipulated in the nest. For those nests
that survived natural predation, we examined treatment affects on post-manipulation
hatching success and nestling growth.
3.2.6 Statistical Analyses
We obtained complete information on clutch size for 85 nests and egg/clutch mass
for 68 nests and used these in our analyses. We conducted 3 sets of analyses. In the first
set, we evaluated treatment effects on clutch size, clutch mass, and egg mass using mixed
models. We first tested for a main effect oftreatment on each egg variable in replacement
clutches (i.e. those laid after the first clutch of the season) by conducting a one-way
ANOVA with female identification as a random effect. We then examined how our 3
egg variables changed over the season in the two groups by conducting a one-way
ANCOVA with female entered as a random effect and treatment and date as fixed effects.
In the latter analysis we include all clutches including the first clutch of the season (i.e.
clutch sizes produced before the beginning of the experiment) and enter both date and the
quadratic of date (date2) into the model to test for trt*date and trt*date2 interactions. Both
date tenns are important because song sparrows often follow a curvilinear clutch size
pattern across the season. Clutch size often increases from first to latter clutches because
sparrows at our site do not produce first clutches of 5 eggs, but do produce 5 egg clutches
later. Though, clutch size often decreases after mid season, potentially as a result of a
56�

negative relationship between lay date and recruitment reported in song sparrows <1 Okm
away on Mandarte island (Hochacka 1990). We also examine treatment affects on the
distribution of clutch sizes laid using a chi squared test.
In the second set of analyses we evaluate treatment effects on total cumulative
egg production (clutch number and total egg number) and we compare HNP and LNP
females' experimental season length, nest duration, and re-nesting period (i.e. days from
clutch removal or abandonment to the laying of the first egg in replacement clutch) to
detennine a) how these are driving the clutch number and egg number difference, and 2)
potentially influencing any treatment effects on clutch size and other egg production
variables. We use a non-parametric Mann-Whitney U test to evaluate treatment effects
on the number of clutches produced by HNP and LNP females and we use a t-test to
compare treatment effects on total egg production (total = clutch number * dutch size).
We restrict the latter analysis to females (n= 9 HNP, n= 8 LNP in which we obtained
complete clutch size information on every clutch produced throughout the season. (i.e.
actual total egg production), rather than multiplying average clutch number by average
clutch size for each individual; although these two methods result in virtually identical
outcomes. We use a t-test to evaluate treatment differences in season length, nest
duration, and average inter-nest interval. Nest duration and inter-nest interval were
calculated as an average for each female across all the nests they produced.
In the third set of analyses, we evaluate the cumulative effects of nest predation
on the physiological profile of females in our two groups using a discriminant function
analysis (DFA). Model significance was determined using a pennutation test iterated
10,000 times in R statistical software (see Mundry and Sommer 2007 for a description of
57

OFA with pennutation test (pDFA)). Our test statistic for the pennutation test was the
eigen-value from the DFA, rather than the classification accuracy used in Mundry &
Sommer (2007). Much like the F statistic, the eigen-value is the between-group variation
divided by the within group variation (in canonical scores) (McGarigal et al. 2000), and
therefore does not have an upper bound (maximum value) like classification accuracy
(i.e. 100%), thus removing the problem of over-fitting when using many discriminating
variables. We report descriptive statistics from the discriminant function analysis (Beta
values) along with standardized effect sizes (Cohen 1992) for each variable in the model
(Table 3.1). We first conducted a DFA comparing the pre-breeding (base line) physiology
of HNP and LNP females testing for initial treatment biases in condition. To test for
predation affects on physiology we use a repeated measures design in the fonn of
difference scores (second capture score - first capture score) in the pDFA. We include
only second capture scores of corticosterone because corticosterone was not assayed in
pre-breeding samples. We used a chi squared test to examine treatment effects on
hatching success of the final experimental clutches (i.e. clutch 3 LNP & clutch 4 or 5
HNP). Finally, we used a t-test to examine treatment affects on nestling mass and
structural size. We used residuals for mass after controlling for the time of day sampled.
Nestling structural size was a combined measure of wing cord and tarsus length produced
using a principle components analysis.
All mixed models were fitted using the restricted maximum likelihood procedure
in Genstat. In the mixed models we test for un-equal variances and deviations from
nonnality using residual and nonnaI probability plots. For all t-tests, we conduct similar
tests using Levene's test of un-equal variance and the Shapiro-Wilk's nonnality test.
58

3.3� Results
Our stratified random design resulted in females in both treatments having similar
initial lay dates (Julian date: HNP females = 94.1 ± 9.1, LNP females = 93.3 ± 8.3; t23 =
0.23, P = 0.83). Also, HNP and LNP females laid their final experimental clutch on a
similar date (HNP females = 149 ± 8.8, LNP females =147 ± 7.8; t22 = -0.57, P = 0.57).
We found no significant differences in clutch size (Mann-Whitney U= 31.5, N ,= 9, N2=
9, p= 0.42), clutch mass (t8= 0.63, P= 0.55), or egg mass (t8= 0.25, P= 0.80) for the first
clutches of females subsequently assigned to either the high predation or low predation
treatments. We confirmed that the experimental diet was consumed and specific
components absorbed based on analysis of plasma carotenoid levels: fed and un-fed birds
were captured on day 6 of brood rearing, after the clutch number manipulation, at which
time fed birds had significantly higher plasma levels of carotenoids than un-fed birds (t41
= -2.93, P < 0.01).
3.3.1� Clutch Size Effects
Experimental nest predation significantly increase clutch number and significantly
decreased clutch size: HNP females laid 57% more clutches in total and in replacement
clutches laid 11 % fewer eggs per clutch or nearly half an egg less, compared to LNP
females (Fig 3.1a). Mean clutch number was 4.71 ± 0.47 for HNP females and 3.00 ±
0.00 for LNP females (Mann-Whitney U= 0.00, P< 0.001). Mean clutch size for
replacement clutches was 3.75 ± 0.07 for HNP females and 4.21 ± 0.14 for LNP females
59

(treatment effect, F L 64 = 10.70, P = 0.002). When considering clutch size across the
entire season, the effect of date on clutch size significantly varied between the two
treatments (treatment*Date, Fl,78 = 8.01, P < 0.01; treatment*date-) = FUBI = 7.21, P <
0.01; Fig 3.2a). The nest predation treatment had significant effects on the distribution of
clutch sizes laid by HNP and LNP females (X22= 14.23, P < 0.001), with HNP females
laying proportionally more small and medium sized clutches, but fewer large clutches.
HNP females laid 8.5% and 11.2% more 3 and 4 egg clutches respectively, but 19.7%
fewer 5 egg clutches (Fig 3.3).
3.3.2� Clutch Mass & Egg Mass Effects
The effect of experimentally-increased nest predation on total clutch mass
followed a similar trend as that seen for clutch size: HNP females laid replacement
clutches which were on average 6.5% lighter (Fig 3.1 b) and the effect of date on clutch
mass tended to vary between the treatments mirroring the clutch size results (Fig 3.2b).
However, the effect of experimental nest predation on the average clutch mass of
replacement clutches was marginally non-significant (mean ± SE (g), HNP == 11.24 ±
0.28, LNP = 12.04 ± 0.42; treahnent effect, F I, 55. = 2.46, P = 0.12), as was the interaction
between date and treatment on the seasonal pattern of clutch mass (treatment*date = F 1,61
== 2.77, P =0.10; treatment*date2, F 1.61 = 3.01, P = 0.09). Egg mass was not affected by
the treatment (F 1.56 = 0.24, P == 0.63) or by treatment by date interactions (treatment*date
)
= FI.62 = 0.05, P = 0.81; treatment*date-, FI.62 = 0.01, P = 0.91).
60�

3.3.3� Cumulative Effects of Nest Predation on Total Egg Production
As a result of producing 57% more clutches HNP females laid 41.5% more eggs
(Fig 3.1b) over the course of the experiment (HNP = 17.33 ± 0.5, LNP = 12.25 ± 0.37;
t15= 8.02, P < 0.001), again revealing that HNP females laid fewer eggs per replacement
clutch since the increase in total egg production (41 %) was less than expected from the
increase in clutch number (57 %). The clutch number difference (and the clutch number
affect on egg number) occurred within the same number of days, i.e. the length of the
experimental breeding season was similar when comparing HNP and LNP females
(season length, t22 = -0.31, P == 0.75). Constrained within the same season length, the
clutch number difference must be due to treatment effects on either the duration of each
nesting attempt (i.e. days from laying the first egg to clutch removal or abandonment)
and/or the duration of the re-nesting period between nesting attempts. HNP females nests
lasted 64% fewer days than LNP females' nests (HNP = 6.52 ± 0.45, LNP = 17.95 ±
0.79; t23= -13.26, P< 0.001), but HNP and LNP females did not differ in the length of the
re-nesting period (t23= -0.74, P= 0.47). This indicates that the clutch number difference
was solely due to predation effects on nest duration.
3.3.4� Physiological Cost of Egg Production
Prior to the start of breeding HNP and LNP females were in similar physiological
state when compared using our integrated measure of condition based on a suite of
physiological traits (DFA Eigen-value = 2.32, Wilks' Lambda = 0.30, pDFA P = 0.51).
However, at the tennination of the experimental nest predation manipulation HNP and
LNP females had significantly different physiological profiles (DFA Eigen-value = 32.7,
61

Wilks' Lambda = 0.03, pDFA P = 0.0012; Table 3.1). Looking at effect sizes> 0.2
(Table 3.1 & Fig 3.4), following Cohen's (1992) guidelines for the magnitude of effect
sizes (i.e. small= 0.2, medium== 0.5, large== 0.8), females in the high nest predation
treatment had higher polychromasia (immature red blood cells), a larger imbalance
between total plasma antioxidants (TAC) and oxidative status (TOS), lower plasma
carotenoids and plasma imunoglobulin levels, higher plasma corticosterone levels, and
lower body fat.
3.3.5 Hatching Success and Nestling Growth
Hatching success of eggs within the final experimental clutches did not differ
between HNP and LNP females (X2, == 0.13, P == 0.72). There was a trend for HNP
nestlings to be lighter 6 days after hatching, however the difference was non-significant
(ts = -1. 74, P = 0.12), and nestlings of HNP and LNP females were found to be of similar
structural size (ts = -0.44, P = 0.73).
3.4 Discussion
Consistent with our first prediction, our results clearly show that experimentalIy
increased nest predation increased clutch number but decreased clutch size of
replacement clutches. In tenns of clutch size, females that experienced nest predation a)
laid smaller replacement clutches, and b) had a significantly altered pattern of seasonal
variation in clutch size. As a result, females that experienced high nest predation laid
proportionally more 3- and 4-egg clutches but fewer 5-egg clutches. Given the effects of
62�

nest predation on clutch size it was surprising that there were no significant predation
affects on clutch mass, although the affects of nest predation on clutch mass did closely
mirror those of predation on clutch size. 11 appears that indlvidual variation in egg size
and a slight non-significant increase in egg size by HNP females dampened nest
predation affects on clutch mass. Nest predation dld not significantly affect the mass of
eggs laid in replacement clutches or the seasonal pattern of variation in egg mass. In line
with our second prediction, our results show that high predation females (HNP) with
increased total egg production were in worse physiological condition, consistent with nest
predation inducing an increased cost of egg production. We suggest that treatment effects
on physiological condition were largely consistent with the hypothesis of non-resource
based mechanisms underlying costs of egg production, i.e. this involved changes in traits
related to hematology and oXldative stress.
Our study has demonstrated experimentally that the direct effect of nest predation
per se affects the clutch size of replacement clutches laid in subsequent nests, a finding
that contrasts with that of previous studies. For example, Doligez & Colbert (2003)
manipulated nest predation and also the visual cues of common nest predators (i.e. they
presented stuffed models at the nest) but found that clutch size did not differ between
birds that experienced nest predation and those that had not. Rather, Doligez & Colbert
(2003) found that clutch size was reduced in the experimental areas in the year following
experimental manipulation because females laid smaller clutches when paired to males
that had resided in the experimental area during the manipulation and laid larger clutches
when paired to newly immigrated males. Doligez & Colbert (2003) concluded that clutch
size adjustments were a result of infOlmation on predation risk, but that this information
63

was not based on direct nest predation. Eggers et al. (2006) examined the effects of
auditory cues to predator presence and found that Siberian jays reduced clutch size in
response to increased vocalizations of common nest predators. Fountain and Martin
(2006), using a predator removal experiment, show that in habitats with both more
predator cues and higher levels of nest predation, birds produced smaller eggs, but not
smaller clutches. In our study we manipulated nest predation only, and not the visual or
auditory cues of predators. In this case both treatments were equally exposed to the nest
predator (the researcher) and would presumably have similar background cues from
natural predators at our study site. The reduction in clutch size that we report thus
demonstrates that nest predation itself can affect clutch size through effects on clutch loss
and is, to the best of our knowledge, the first experimental evidence that direct nest
predation affects clutch size.
As we discussed in the introduction, Martin (1995) proposed that the inverse
correlation between clutch number and clutch size observed across species may have
been the result of an energetic trade-off. In the present study nest predation increased
clutch number and decreased clutch size when energy and nutrients were super-abundant,
suggesting that food availability can not eliminate the effects of direct nest predation on
clutch size in song sparrows. Yet, it is possible that food limitation may remain the
underlying mechanism for the clutch size reduction that we report. For example, the
amount of food consumed by prey may be affected by predator' intimidation', resulting
in predators affecting clutch size indirectly through effects on food availability. Martin
(1992) described one such means where nest predation affects "perching time" (i.e.,
vigilance), which affects "foraging time," which through reduced food consumption
64�

affects clutch size. However, our work and previous work on song sparrows suggests that
predator intimidation did not limit song sparrows access to food. In a bi-factorial study of
food and predators, Zanette et al. (2006b) found that food availability is an important
factor detennining the length of time required for re-nesting, with fed birds re-nesting in
significantly fewer days than un-fed birds. Therefore, if predator intimidation reduced
song sparrows access to food we would expect that predators would then also affect the
re-nesting duration. Zanette et al. (2006b) reported that predators did not affect the re
nesting duration for both fed and un-fed birds. In the present study we also found the nest
predation did not affect the re-nesting duration suggesting that predator intimidation did
not affect song sparrows access to food. Overall, we suggest that our results demonstrate
that nest predation in song sparrows affects clutch size by a mechanism that is
independent of food availability.
In showing that nest predation can affect clutch size when food is abundant, our
results also confinn that food and predators can have combined effects on clutch size via
food acting on the rate of nest predation. Zantte et al. (2006a) found that food availability
reduced the occurrence of nest predation in song sparrows and Duncan-Rastogi et aJ.
(2006) showed this was likely because fed birds spent more time on the nest and made
fewer foraging trips of shorter duration then un-fed birds. Combined these results
suggested that if nest predation does affect clutch size then because food availability
reduced the occurrence of nest predation, food may reduce the opportunity for predation
to affect clutch size (i.e. a food and predator interaction on clutch size) (Zanette et al.
2006b). Therefore, our results demonstrating that predators can affect clutch size also
confinns that food and predators can have combined affects on clutch size (Zanette et al.
6S�

2006b). Taken together, we suggest that work on food and predators in song sparrows
sheds light on the decades long dichotomy over whether food or predators affect clutch
size (Lack 1947, Pemns 1977, Martin 1987, Newton 1998).
3.4.1� Total Cumulative Egg Production & the Physiological Cost of Egg Production
Our results suggest that there is a physiological cost of increased egg production
as a result of direct nest predation stimulating more frequent re-nesting and thus, total egg
production. Here experimentally-increased nest predation increased total egg production
with HNP females laying an average of 1.7 more clutches, which resulted in HNP
females producing an additional 5 eggs over the season. We believe that the effects of
nest predation on egg production that we found are not specific to our study. In birds,
direct nest predation 1s the primary source of nest failure (Ricklefs 1969, Martin 1995),
and has frequently been reported to stimulate re-nesting and thus increase clutch number
in many species (Grzybowski and Pease 2005, Zanette et a1. 2006b). For example, Catlin
& Rosenberg (2008) found that nest predation in burrowing owls (Athene mniculari)
resulted in both increased clutch number and total egg number. Other studies directly
manipulating egg production by egg removal, have shown that increased egg production
effort can have negative affects on female survival (Nager et a!. 200 I, Visser and Lessells
2001), a female's ability to rear chicks (Monaghan et a1. 1998), and can delay subsequent
breeding (Kalmbach et a1. 2004), negatively impact a female's physiological condition,
reduced hatching success of eggs (Kalmbach et a1. 2004), and reduced nestling size
(Monaghan et a!. 1995).
66

In our study, HNP females were in worse overall physiological condition at the
end of the season, consistent with costs incurred from greater total seasonal egg
production. We found that treatment effects on condition indices were largely consistent
with predictions for the non-resource based costs of egg production discussed in the
introduction (Williams 2005). However, traditionally the cost of egg production is
explained by a resource based trade-off where females are forced to allocate a limiting
amount of food resources between there own energetic needs or those of their offspring.
For example, Kalmbach et al. (2004) found that increased egg production in great skuas
(Stercorarius skua) resulted in both a decrease in female mass and reduced hatching
success of later laid eggs. In the present study, we supplemented all birds a high quality
diet, and therefore did not expect to find resource based trade-offs. Even if we assumed
resources were still limiting, but by an internal limitation (e.g. absorption rate) rather than
external quantity (Speakman and Krol 2005), we would expect treatment effects on
energetic traits and or a trade-off with offspring quality. However, we generally did not
detect evidence for resource based trade-off. Here, there were significant increases in our
two measures of egg production, yet there were generally weak and inconsistent
treatment affects on female energetic balance (with the exception of fat); in fact there was
a slight positive affect on HNP mass. Furthermore, female energetic condition does not
appear to have been maintained at the expense of offspring quality. HNP females actually
laid slightly heavier eggs (3.00g vs 2.89g, non-significant difference) in their final
clutches, and there were no treatment effects on hatching success, or nestling growth. We
suggest that the general lack of effects, or trade-off, predicted by the resource based
hypothesis, indicates that females were not limited by food availability; which is
67

consistent with the super-abundance of food and is similar to our clutch size results. That
said, there was a large negative treatment effect on fat scores, suggesting that despite an
abundance of food HNP females were not able to absorb or assimilate energy at a high
enough rate to maintain fat stores during increased egg production (Speakman and Krol
2005). Alternatively, temporal differences in access to food may have influenced fat
dynamics between treatments. HNP females spent less time incubating nests and more
time re-nesting as a result of predation, reducing the number of days spent in continuous
short term incubation fasts. Energy during short term fasts is fuelled by fat stores (Jenni
Eiermann and Jenni 1997), however, because converting food into fat and then
metabolizing fat into usable energy is inefficient (Cope 2003), animals with continuous
access to food may facultatively reduce fat storage (i.e. similar to the programmed
anorexia hypothesis Norberg 1981, Gaston and Jones 1989).
In contrast to effects on "energetic" physiological traits, overall there were
consistent negative effects on non-energetic indices. In terms of the different non
resource based mechanisms for costs of egg production described in the introduction,
HNP females had a greater imbalance between antioxidants and oxidative status (reactive
oxygen species) consistent with oxidative stress as an underlying mechanism for the cost
of egg production (Salmon et al. 2001, Wang et al. 2001, Alonso-Alvarez et al. 2006,
Bize et al. 2008). HNP females also had higher polychromasia (immature red blood cells)
as predicted by the idea of reproductive anaemia as a result of estrogen-dependent
transient (i.e. during egg laying) suppression of red blood cell (RBC) production
(Williams et al. 2004, Wagner et al. 2008a). This suggests that a greater number of bouts
of egg laying (i.e. clutch number) with high nest predation might involve more frequent
68�

exposure to reproductive hormones which would enhance the negative pleiotropic
(multiple) effects of gonadal estrogens. The slightly higher levels of hematocrit (which
appears inconsistent with estrogen suppression) of HNP females may be due to the
increased proportion of the larger (volume) immature red blood cells (Campbell and Ellis
2007) found in HNP females blood. We obtained blood samples approximately 9 days
after estrogens would have decreased (i.e. at clutch completion (Williams et al. 2004),
which is sufficient time for regenerative anaemia to recover hematocrit levels (Domm
and Taber 1946, Clark et al. 1988) through increasing cell numbers but also simply
because of the larger contribution of each immature cell to hematocrit volume (Fernandez
and Grindem 2006). Our general indices of condition, immunogloblins and
corticosterone, are also consistent with HNP females being in worse condition, and
potentially, at least in the case of imunoglobulins, as a result of increased egg production.
Merino et al. (2006) found that increased breeding effort reduced immunoglobulin levels
and Ochs et a1. (2008) found that females that produced more eggs (i.e. larger clutches)
had declines in general measures of immune function. Finally, numerous studies have
shown that plasma corticosterone levels are negatively correlated with condition
(Kitaysky et al. 1999, Romero and Wekelski 2001, Lobato et a1. 2008).
A further explanation for our results is that many of these physiological effects we
observed might also be associated with the elevated corticosterone levels in HNP
females, potentially because increased nest predation induced higher 'stress'. The chronic
stress hypothesis (conflict between obtaining food and avoiding being eating) proposed
by Boonstra et al. (1998) for mammals and followed up by Clinchy et a1. (2004) in birds,
also predicts a number of broad scale physiological changes observed here (excluding
69�

oxidative stress). To examIne this hypothesis we looked at the direction of the
relationship between corticosterone (stress honnone) and all other indices of condition at
post breeding (but not reported in this paper). We found that the direction of the
relationship between indices and corticosterone was not consistent with treatment effects
on indices (7 out 12 times), suggesting corticosterone did not cause many of the treatment
effects. Although, corticosterone can have wide spread influence on many variables we
examined, we suggest that it does not provide a better explanation then predictions made
by the cost of egg production but may have also contributed to our results.
Our results provide initial evidence consistent with the possibility that the effect
of nest predation on increased clutch number and decreased clutch size is the result of a
trade-off influenced by physiological costs of egg production. Gasparini et al. (2006)
found that experimentally inducing re-nesting (i.e. increasing clutch number) in
kittiwakes resulted in reduced clutch mass in both fed and un-fed birds replacement
clutches (i .e. simi lar to our findings food did not eliminate the inverse relationship),
which they suggest may be the result of a physiological constraint influencing egg
production decisions. If in the present study clutch size reductions are the result of
physiological cost of egg production, the influence of such costs would have to occur
early in the season because HNP females produced smaller clutches as early as the
second clutch, i.e. the first replacement clutch. However, in the present study we are
unable to detennine if increased physiological costs of egg production are due to a greater
total egg production (i.e. more bouts and more eggs), which would not fully explain the
early clutch size reduction or, due to the fact that high-predation females had an increased
frequency of egg laying with less "recovery" time between laying events; which would
70�

be consistent with an early reduction of clutch size. With regards to shorter recovery
explanation, incubation, although energetically demanding, can allow for physiological
recovery from egg production (Hario et al. 1991, Alonso-Alvarez et al. 2002, Williams et
al. 2004, Navarro et al. 2007, Wagner et aJ. 2008b). In our study, increased nest predation
shortened the incubation period reducing the potential for incubation recovery. As well,
shorter incubation periods were not compensated for by longer re-nesting duration. Such
behaviour may be due to the fact that annual reproductive success in song sparrows and
other species is strongly influence by clutch number (Nagy and Holmes 2004, Zanette et
al. 2006a), therefore birds may be 'driven' to re-nest without fully recovering
physiological state, with physiological state then influencing clutch size decisions.
In conclusion, our results further highlight how predators can negatively affect
prey beyond simply killing them or their offspring (Lima 1998, Creel and Christianson
2008). Here, song sparrows that experienced nest predation laid smaller subsequent
clutches and ending up in worse physiological condition, consistent with predation effects
increasing the cost of egg production. Furthermore, our results suggest that direct nest
predation does affect clutch size and can affect physiological condition independent of
food availability, but importantly, we do not imply that this means that food does not also
affect the variables we measured. Our results confinn a previous hypothesis that food and
predators have combined effects on clutch size (Zanette et al. 200Gb), and highlights the
importance of simultaneously considering the affects of food and predators.
71

3.5 References
Alonso-Alvarez, C, S. Bertrand, G. Devevey, J. Prost, B. Faivre, O. Chastel, and G.� Sorci. 2006. An experimental manipulation of life-history trajectories and� resistance to oxidative stress. Evolution 60: 1913-1924.�
Alonso-Alvarez, C, S. Bertrand, G. Devevey, J. Prost, B. Faivre, and G. Sorci. 2004. Increased susceptibility to oxidative stress as a proximate cost ofreproduction. Ecology Letters 7:363-368.
Alonso-Alvarez, C, A. Velando, M. Ferrer, and J. A. R. Veira. 2002. Changes in plasma biochemistry and body mass during incubation in the Yellow-legged Gull. Waterbirds 25:253-258.
Bize, P., G. Devevey, P. Monaghan, B. Doligez, and P. Christe. 2008. Fecundity and survival in relation to resistance to oxidative stress in a free-living bird. Ecology 89:2584-2593.
Blount, J., D. C. Houston, P. F. Surai, and A. P. Moller. 2004. Egg-laying capacity is limited by carotenoid pigment availability in wild gulls Larusfuscus. Proceedings of The Royal Society of London Series B-Biological Sciences 271: S79-S81.
Boonstra, R., D. Hik, G. R. Singleton, and A. Tinnikov. 1998. The impact of predatorinduced stress on the snowshoe hare cycle. Ecological Monographs 68:371-394.
Breuner, C W. and M. Orchinik. 2002. Plasma binding proteins as mediators of corticosteroid action in vertebrates. Journal of Endocrinology 175:99-112.
Campbell, T. W. and C Ellis, editors. 2007. Avian and Exotic Animal Hematology and Cytology. 3rd edition. Iowa State Press, Ames.
Carey, C. R. H. and P. Parisi. 1980. Calories, water, lipid and yolk in avian eggs. Condor 82:335-343.
Catlin, D. H. and D. K. Rosenberg. 2008. Breeding dispersal and nesting behavior of burrowing owls following experimental nest predation. American Midland Naturalist 159:1-7.
Clark, M. W., R. P. Gildersleeve, J. P. Thaxton, C R. Parkhurst, and D. 1. McRee. ] 988. Hematological effects of ethyl methanesulfonate, paraquat and phenylhydrazine in japanese quail. Comparative Biochemistry and Physiology C-Pharmacology Toxicology & Endocrinology 89: 15-30.
Clinchy, M., L. Zanette, R. Boonstra, J. C Wingfield, and 1. N. M. Smith. 2004. Balancing food and predator pressure induces chronic stress in songbirds. Proceedings of the Royal Society Biological Sciences Series B 271:2473-2479.
Cohen, J. 1992. A power primer. Psychological Bulletin 112:155-159.
72

Cope, D. R. 2003. Variation in daily and seasonal foraging routines of non-breeding barnacle geese (Branta leucopsis): working harder does not overcome environmental constraints. Journal of Zoology 260:65-71.
Creel, S. and D. Christianson. 2008. Relationships between direct predation and risk effects. Trends In Ecology & Evolution 23: 194-201.
De Neve, L., J. A. Fargallo, P. Vergara, J. A. Lemus, J.-G. M., and L. 1. 2008. Effects of maternal carotenoid availability in relation to sex, parasite infection and health status of nestling kestrels (Falco tinnunculus). Journal of Experimental Biology 211: 1414-1425.
DeSante, D. F., K. M. Burton, P. Velez, D. Froehlich, and D. R. Kaschube. 2008. MAPS Manual: 2008 Protocol. Contribution No. 127 of the Institute for Bird Populations
Doligez, B. and 1. CIobert. 2003. Clutch size reduction as a response to increased nest predation rate in the collared flycatcher. Ecology 84:2582-2588.
Domm, L. V. and E. Taber. 1946. Endocrine factors controlling erythrocyte concentration in the blood of the domestic fowl. Physiological Zoology 19:258-281.
Drabkin, D. L. and J. H. Austin. 1932. Spectrophotometric studies 1. Spectrophotometric constants for common hemoglobin derivatives in human, dog, and rabbit blood. Journal of Biological Chemistry 98:719-733.
Duncan-Rastogi, A., L. Zanette, and M. Clinchy. 2006. Food availability affects diurnal nest predation and adult antipredator behaviour in song sparrows, Melospiza melodia. Animal Behaviour 72:933-940.
Eggers, S., M. Griesser, M. Nystrand, and J. Ekman. 2006. Predation risk induces changes in nest-site selection and clutch size in the Siberian jay. Proceedings of the Royal Society Biological Sciences Series B 273:701-706.
Erel, O. 2004. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clinical Biochemistry 37:277-285.
Erel, O. 2005. A new automated colorimetric method for measuring total oxidant status. Clinical Biochemistry 38: 11 03-1 Ill.
Fernandez, F. R. and C. B. Grindem, editors. 2006. Reticulocyte response. 5th edition. Blackwell, Ames.
Fontaine, J. and T. E. Martin. 2006. Parent birds assess nest predation risk and adjust their reproductive strategies. Ecology Letters 9:428-434.
Gasparini, J., A. Roulin, V. A. Gill, S. A. Hatch, and T. Boulinier. 2006. Kittiwakes strategically reduce investment in replacement clutches. Proceedings of The Royal Society Biological Sciences 273: 1551-1554.
73

Gaston, A. J. and 1. L. Jones. 1989. The relative importance of stress and programmed anorexia in determining mass-loss by incubating ancient murrelets. Auk 106:653658.
Grzybowski, J. and C. M. Pease. 2005. Renesting detennines seasonal fecundity in songbirds: What do we know? What should we assume? Auk 122:280-292.
Hario, M., M. Kilpi, and K. Selin. 1991. Parental investment by the sexes in the herring gull - the use of energy reserves during early breeding. Ornis Scandinavica 22:308-312.
Harshman, L. and A. J. Zera. 2007. The cost of reproduction: the devil in the details. Trends In Ecology & Evolution 22:80-86.
Hochacka, W. 1990. Seasonal decline in reproductive perfonnance of song sparrows. Ecology 71: 1279-1288.
Houston, D., D. Donnan, P. Jones, 1. Hamilton, and D. Osborne. 1995. Changes in the muscle condition of female zebra finches poephila-guttata during egg-laying and the role of protein storage in bird skeletal-muscle. Ibis 137:322-328.
Jenni-Eiennann, S. and L. Jenni. 1997. Diurnal variation ofmetabolic responses to shortterm fasting in passerine birds during the postbreeding, molting and migratory period. Condor 99:113-122.
Kalmbach, E., R. Griffiths, J. E. Crane, and R. W. Furness. 2004. Effects of experimentally increased egg production on female body condition and laying dates in the great skua Stercorarius skua. Journal of Avian Biology 35:501-514.
Kitaysky, A., J. C. Wingfield, and 1. F. Piatt. 1999. Dynamics of food availability, body condition and physiological stress response in breeding Black-legged Kittiwakes. Functional Ecology 13:577-584.
Lack, D. 1947. The significance of clutch-size. Ibis 89:302-352.
Lima, S. L. 1998. Nonlethal effects in the ecology of predator-prey interactions - What are the ecological effects of anti-predator decision-making? Bioscience 48:25-34.
Lobato, E., S. Merino, J. Moreno, J. Morales, G. Tomas, J. M. la Puente, J. L. Osorno, A. Kuchar, and E. Moestl. 2008. Corticosterone metabolites in blue tit and pied flycatcher droppings: Effects of brood size, ectoparasites and temperature. Honnones and Behavior 53:295-305.
Love, O. P., C. W. Breuner, F. Vezina, and T. D. Williams. 2004. Mediation of a corticosterone-induced reproductive conflict. Hormones and Behavior 46:59-65.
Love, O. P., E. H. Chin, K. E. Wynne-Edwards, and T. D. Williams. 2005. Stress hormones: A link between maternal condition and sex-biased reproductive investment. American Naturalist 166:751-766.
74

Martin, T. 1987. Food as a limit on breeding birds a life-history perspective. Annual Review of Ecology and Systematics:453-488.
Martin, T. 1995. Avian life-history evolution in relation to nest sites, nest predation, and food. Ecological Monographs 65:101-127.
Martin, T. E. 1992. Interaction of nest predation and food limitation in reproductive strategies. Current Ornithology 9: 163 -197.
Martin, T. E. and P. J. Li. 1992. Life-history traits of open-nesting vs cavity-nesting birds. Ecology 73:579-592.
Martinez, J., G. Tomas, S. Merino, E. Arriero, and J. Moreno. 2003. Detection of serum immunoglobulins in wild birds by direct ELISA: a methodological study to validate the technique in different species using antichicken antibodies. Functional Ecology 17:700-706.
McGarigal, K., S. Cushman, and S. Stafferd, editors. 2000. Multivariate statistics for wildlife and ecological research. Springer-Verlag, New York, USA.
McGraw, K., O. L. Crino, W. Medina-Jerez, and P. M. Nolan. 2006. Effect of dietary carotenoid supplementation on food intake and immune function in a songbird with no carotenoid coloration. Ethology 112: 1209-1216.
Merino, S., J. Moreno, G. Tomas, J. Martinez, J. Morales, J. Martinez-De la Puente, and J. L. Osorno. 2006. Effects of parental effort on blood stress protein HSP60 and immunoglobulins in female blue tits: a brood size manipulation experiment. Journal of Animal Ecology 75: 1147-1153.
Monaghan, P., M. Bolton, and D. C. Houston. 1995 Egg-production constraints and the evolution of avian clutch size. Proceedings of The Royal Society of London Series B-Biological Sciences 259: 189-191.
Monaghan, P., R. G. Nager, and D. C. Houston. 1998. The price of eggs: increased investment in egg production reduces the offspring rearing capacity of parents. Proceedings of The Royal Society of London Series B-Biological Sciences 265: 1731-1735.
Mundry, R. and C. Sommer. 2007. Discriminant function analysis with nonindependent data: consequences and an alternative. Animal Behaviour 74:965-976.
Nager, R. 2006. The challenges of making eggs. Ardea 94:323-346.
Nager, R., P. Monaghan, and D. C. Houston. 2001. The cost of egg production: increased egg production reduces future fitness in gulls. Journal of Avian Biology 32: 159166.
Nagy, L. and R. Holmes. 2004. Factors influencing fecundity in migratory songbirds: is nest predation the most important? Journal of Avian Biology 35:487-491.
75

Navarro, J., J. Gonzalez-Solis, and G. Viscor. 2007. Nutritional and feeding ecology in Cory's shearwater Calonectris diomedea during breeding. Marine Ecology Progress Series 351:261-271.
Newton,1. 1998. Population limitation in birds. Academic Press, London, UK
Norberg, R. A. 1981. Temporary weight decrease in breeding birds may result in more fledged young. American Naturalist 118:838-850.
Ochs, C. L. and R. D. Dawson. 2008. Patterns of variation in leucocyte counts of female tree swallows, Tachycineta bicolor: Repeatability over time and relationships with condition and costs of reproduction. Comparative Biochemistry and Physiology A-Molecular & Integrative Physiology 150:326-331.
Partridge, L., D. Gems, and D. J. Withers. 2005. Sex and death: What is the connection? Cell 120:461-472.
Perrins, C. M. 1977. The role of predation in the evolution of clutch size. University Park Press, Baltimore, USA.
Ramsay, S. and D. C. Houston. 1998. The effect of dietary amino acid composition on egg production in blue tits. Proceedings of the Royal Society of London Series bBiological Sciences 265:1401-1405.
Ricklefs, R. E. 1969. An analysis of nesting mortality in birds. Smith. Contr. Zool 9: 1-48.
Romero, M. L. and M. Wekelski. 2001. Corticosterone levels predict survival probabilities of Galapagos marine iguanas during E1 Nino events. Proceedings of the National Academy of Sciences of the United States of America 98:7366-7370.
Salmon, A. B., D. B. Marx, and L. G. Harshman. 2001. A cost ofreproduction in Drosophila melanogaster: Stress susceptibility. Evol ution 55: 1600-1608.
Seaman, D., C. G. Guglielmo, R. W. Elner, and T. D. Williams. 2006. Landscape-scale physiology: Site differences in refueling rates indicated by plasma metabolite analysis in free-hving migratory sandpipers. Auk 123:563-574.
Selman, R. and D. C. Houston. 1996. The effect of prebreeding diet on reproductive output in zebra finches. Proceedings of the Royal Society of London Series B Biological Sciences 263: 1585-1588.
Skutch, A. F. 1949. Do tropical birds raise as many young as they can nourish? Ibis 91 :430-455.
Speakman, J. R. and E. Kro!. 2005. Limits to sustained energy intake IX: a review of hypotheses. Journal of Comparative Physiology B-Biochemical Systemic and Environmental Physiology 175:375-394.
Visser, M. and C. M. Lessells. 2001. The costs of egg production and incubation in great tits (Panls major). Proceedings of The Royal Society of London Series BBiological Sciences 268: 1271-1277.
76

Wagner, E., J. S. Prevolsek, K. E. Wynne-Edwards, and T. D. Williams. 2008a. Hematological changes associated with egg production: estrogen dependence and repeatability. Journal of Experimental Biology 211 :400-408.
Wagner, E. c., C. A. Stables, and T. D. Williams. 2008b. Hematological changes associated with egg production: direct evidence for changes in erythropoiesis but a lack of resource dependence? Journal of Experimental Biology 211 :2960-2968.
Wang, Y, A. B. Salmon, and L. G. Harshman. 2001. A cost of reproduction: oxidative stress susceptibility is associated wi th increased egg production in Drosophila melanogaster. Experimental Gerontology 36: 1349-1359.
Williams, G. 1966. Natural selection costs of reproduction and a refinement oflacks principle. American Naturalist 100:687-&.
Williams, T. D. 2005. Mechanisms underlying the costs of egg production. Bioscience 55:39-48.
Williams, T. D., W. O. Challenger, J. K. Christians, M. Evanson, O. Love, and F. Vezina. 2004. What causes the decrease in hematocrit during egg production? Functional Ecology 18:330-336.
Zanette, L., M. Clinchy, and J. N. M. Smith. 2006a. Combined food and predator effects on songbird nest survival and annual reproductive success: results from a bifactorial experiment. Oecologia 147:632-640.
Zanette, L., M. Clinchy, and J. N. M. Smith. 2006b. Food and predators affect egg production in song sparrows. Ecology 87:2459-2467.
Zanette, L., J. N. M. Smith, H. v. Oort, and M. Clinchy. 2003. Synergistic effects offood and predators on annual reproductive success in song sparrows. Proceedings of The Royal Society of London Series B-Biological Sciences 270:799-803.
77
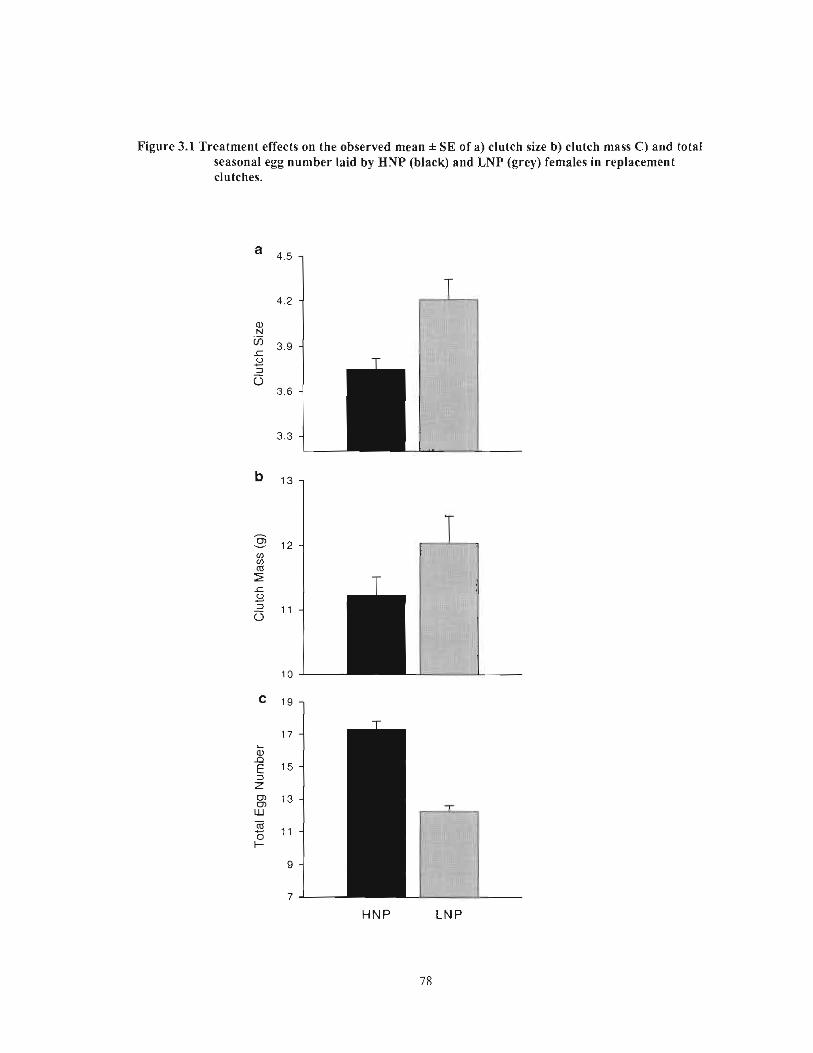
Figure 3.1 Treatment effects on the observed mean ± SE of a) clutch size b) clutch mass C) and total seasonal egg number laid by HNP (black) and LNP (grey) females in replacement clutches.
a
aJ N (]j ..c .8 ::::l
C3
b
:§ (J) (J) ro 2 £: £ ::::l (3
C
~
aJ .0 E ::::l Z OJ OJ
UJ
ro 0 I
4.5
4.2
3.9
3.6
3.3
13
12
11
10
19
17
15
13
11
9
7
HNP LNP
78
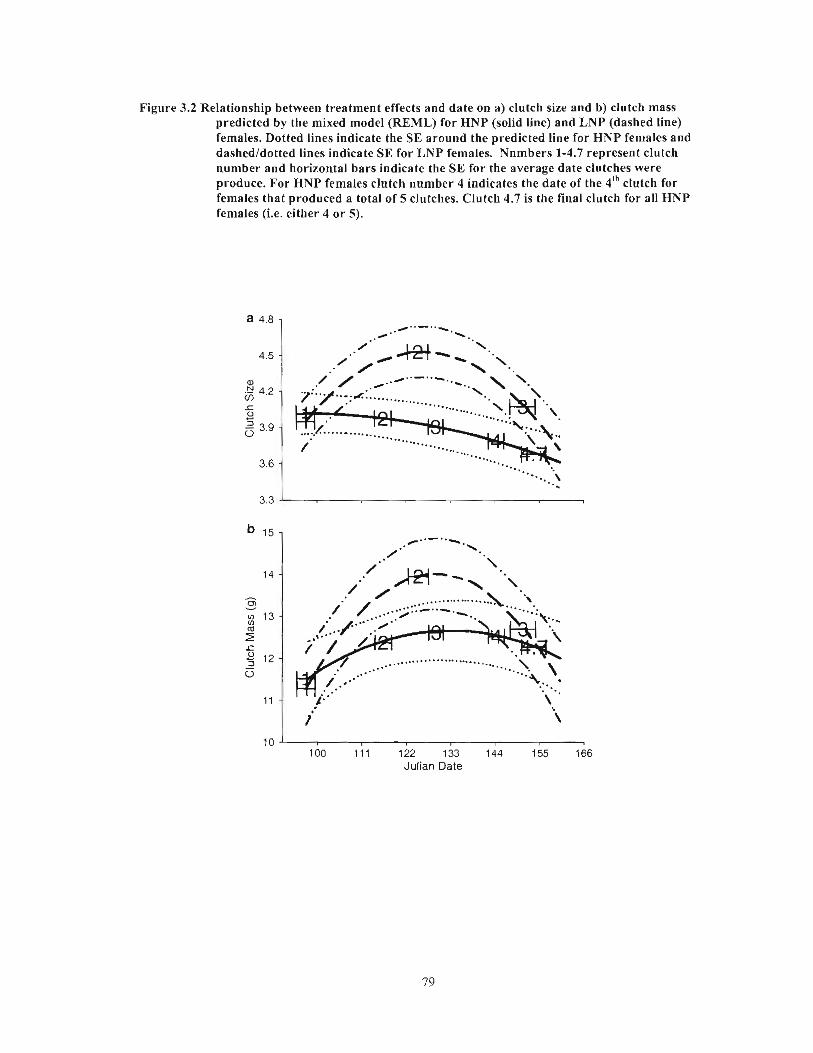
Figure 3.2 Relationship between treatment effects and date on a) clutch size and b) clutch mass predicted by the mixed model (REML) for HNP (solid line) and LNP (dashed line) females. Dotted lines indicate the SE around the predicted line for HNP females and dashed/dotted lines indicate SE for LNP females. Nnmbers 1-4.7 represent clutch number and horizontal bars indicate the SE for the average date clutches were produce. For HNP females clutch number 4 indicates the date of the 41h clutch for females that produced a total of 5 clutches. Clutch 4.7 is the final clutch for all HNP females (i.e. either 4 or 5).
a 4.8 ." ~..". .. _...............
./., l...!:l.J. ",
./ .' .- ....,..-z:::j - - "" .' /' , '.
Q)
4.5
/ / .. -"-"-.. "" M4.2 7::/""";:::-:':." ,.. ~'\. ~ .... . ~:~ "'.(J
~ 3.9 ...../................. :,..,..�,/ .
.... .... 3.6 '" " ........ \� 3.3 ..L-_~----.----r--~--~---,------,
b 15 ......- ......... ./
§ (J) (J) etl
~ £
14
13
/ '\.
/ )-2"l--, '\. /' ' .
.' / '- .. './ -- .. --......... . ; .. / " '. \"~ I .' " ./',/' /.., \
(J
"5 0
12 "7 .. -
. /........
'\. \ . . ....,':-"
. f' \
::J .I ' \
100 111 122 133 144 155 166 Julian Date
79�
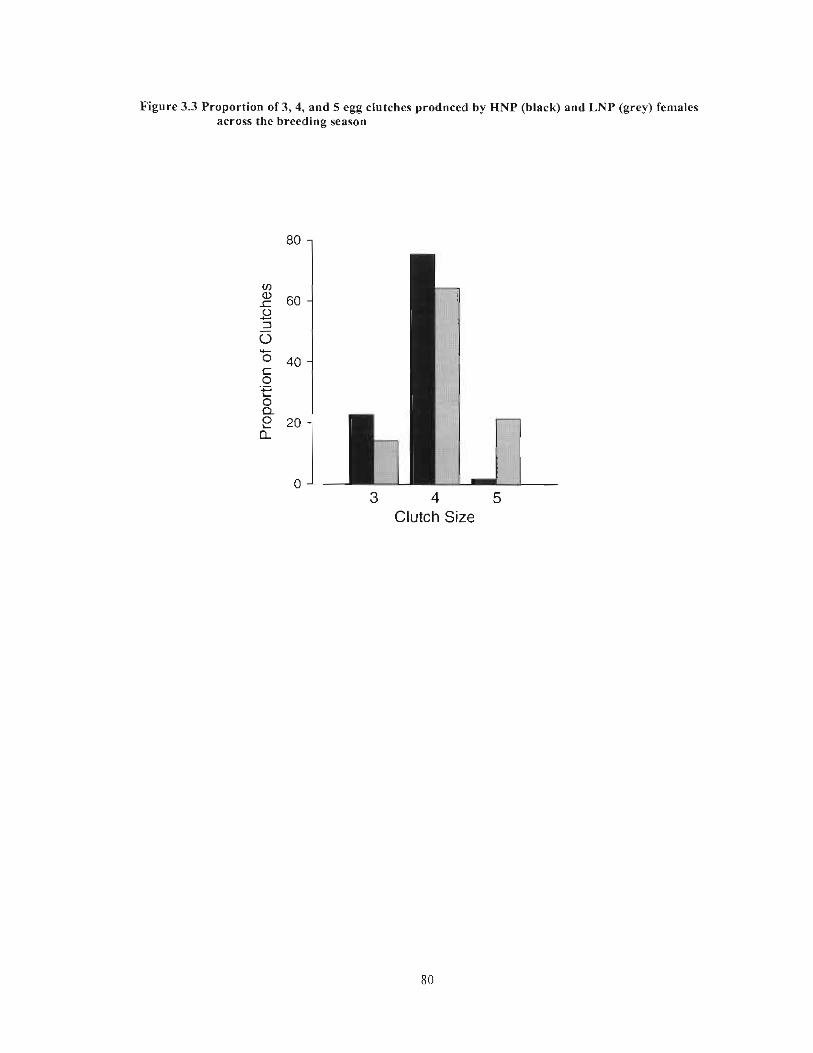
Figure 3.3 Proportion of 3,4, and 5 egg clutches prodnced by HNP (black) and LNP (grey) females across the breeding season
80
(f) Q) £. 60 0.......� ::J
-00 40 c 0
"+=;..... 0 Q..� 0� ..... 20 n..
0 3 4 5
Clutch Size
80�

Tab
le 3
.1 T
he e
ffec
t o
f ne
st p
reda
tion
on
phys
iolo
gica
l co
ndit
ion.
Mea
ns ±
SE
for
phy
siol
ogic
al t
rait
s m
easu
red
in f
emal
es a
t pr
e-br
eedi
ng a
nd p
ost
man
ipul
atio
n (i
ncub
atio
n).
See
cha
pter
2 f
or u
nits
. D
escr
ipti
ve s
tati
stic
s fr
om t
he D
FA
alo
ng w
ith
stan
dard
ized
eff
ect
size
s fo
r ea
ch i
ndex
. E
ffec
t si
ze;
smal
l =
0.2
, m
ediu
m =
0.5
, la
rger
= 0
.8 (
Coh
en 1
992)
.
Pre
-Bre
edin
g In
cuba
tion
~in
HN
P
Can
onic
al
Uni
vari
ate
Sca
le
Ind
ices
R
elat
ive
Coe
ffic
ient
E
ffec
t Si
ze
HN
P
LN
P
HN
P
LN
P
to L
NP
(B
eta)
(d
)
Hem
atol
ogy
Hem
atoc
rit
0.49
3 ±
0.0
10
0.50
0 ±
0.0
09
0.52
2 ±
0.0
1 0.
527
± 0
.006
i
0.50
0.
09
Hem
oglo
bin
10.7
2 ±
0.3
8 11
.41
± 0
.37
11.4
0 ±
0.3
3 12
.24
± 0
.30
1 1.
69
-0.1
2
Pol
ychr
omas
ia
3.10
±0.2
6 3.
21 ±
0.3
3 4.
11 ±
0.5
4 2.
94 ±
0.2
0 i
4.53
0.
63
00
O
xida
tive
Str
ess
Ant
ioxi
dant
Sta
tus
1.94
± 0
.17
1.50
±0.1
0 2.
29±0
.13
2.10
± 0.
12
1 7.
25
-0.4
3
Oxi
dati
ve S
tatu
s 21
9±
16
239
±26
29
4±
27
273
± 3
9 i
1.13
0.
34
Car
oten
oids
1.
40 ±
0.1
5 1.
34 ±
0.1
8 1
95
±0
19
2.
25±0
.19
1 01
8 -0
.52
Imm
une
Inun
unog
lobu
lin
0.75
± 0
.05
0.62
± 0
.04
0.75
± 0
.08
0.69
± 0
.04
1 2.
36
-0.3
7
Hon
nona
l C
orti
cost
eron
e 8.
17 ±
1.2
9 6.
24 ±
1.3
5 i
4.24
0.
46
Ene
rget
ic S
tatu
s�
Tri
glyc
erid
es
1.42
± 0
.11
1.59
± 0
.15
1.31
±0.1
6 1.
36±0
.16
i 0.
61
0.18
NE
FA
1.26
± 0
.06
1.20
± 0
.10
1.03
± 0
.11
0.93
± 0
.07
i 22
4 00
9
Glu
cose
21
.65±
1.19
23
.99
± 1
.73
20.7
9 ±
1.5
4 21
.60
± 0
.90
i 4.
04
0.19
Fat
1.09
± 0
.28
0.22
± 0
.15
3.05
± 0
.35
3.44
± 0
31
1 3.
58
-0.8
9
Mas
s 23
.87
± 0
33
23.8
8 ±
0.4
4 24
.21
± 0
.42
23.9
6 ±
0.3
7 i
1.61
0.
18
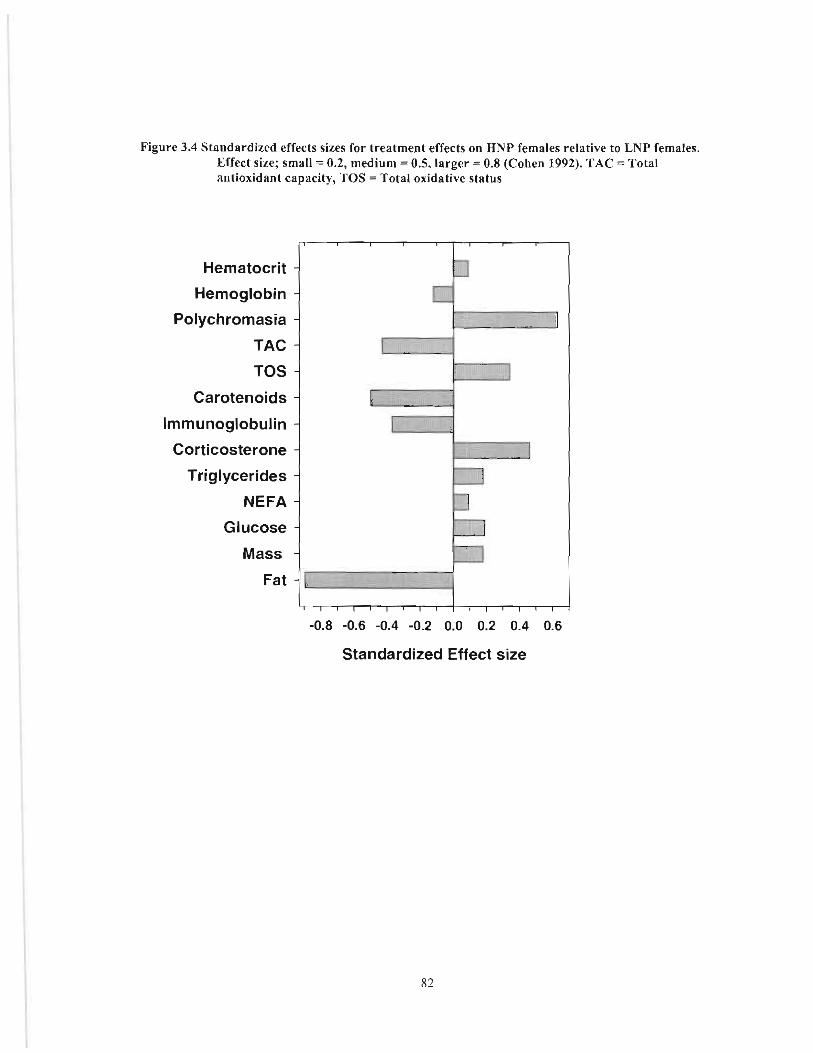
Figure 3.4 Standardized effects sizes for treatment effects on HNP females relative to LNP females. Effect size; small = 0.2, medium == 0.5. larger == 0.8 (Cohen 1992). TAC == Total antioxidant capacity, TOS == Total oxidative status
Hematocrit
Hemoglobin
Polychromasia
TAC
TOS
Carotenoids
Immunoglobulin
Corticosterone
Triglycerides
NEFA
Glucose
Mass�
Fat�
-0.8 ~0.6 -0.4 -0.2 0.0 0.2 0.4 0.6
Standardized Effect size
82�

CHAPTER 4 GENERAL SYTHESIS AND FUTURE DIRECTIONS
4.1 Synthesis
What follows is a discussion of our results from chapter 2 and 3 usmg a new
conceptual frame work (allostasis) to integrate the findings of both chapters. I have
chosen to discuss allostasis because I believe that ecophysiology studies could benefit
from working within the allostasis framework. In addition, although not discussed below,
I have found that the analytical tools used in studies of allostasis may also be beneficial to
ecologists that use multiple variables to measure physiological state.
A goal of many studies of free living animals is to identify physiological traits
that indicate the health and fitness potential of an individual (Bortolotti et al. 2002, Hill
and Fanner 2005, Ochs and Dawson 2008). Numerous studies report relationships
between hormonal, immunological or energetic measures and traits related to
reproductive success and survival (Andersson and Gustafsson 1995, Atkinson and
Ramsay 1995, Romero and Wekelski 2001, Bety et al. 2003, Blums et al. 2005).
However, no universal trait has yet immerged that ecologist, or doctors for that matter,
use to determine the health or fitness potential of an individual. Our results (chapter 2)
suggest that this may be due to the fact that no single trait explains much of the variation
in overall physiological state. From this, we concluded that measurements of condition
should integrate multiple traits to obtain a better understanding of global physiological
state. The results of chapter 3 provide further support for this earlier finding. Rather than
83

large effects on a few physiological traits, our analysis of condition revealed that it was
the cumulative effect of predation spread across a number of traits that resulted in
treatment differences.
The results from a number of human epidemiology studies appears to support our
conclusion that a more integrated measure will aid in more accurately assessing condition
(McEwen 1998, Seeman et al. 2001, Glei et al. 2007, Nelson et al. 2007), but these
studies also provide a framework to better understand physiological results. Many
medical studies are examining physiology and health within the conceptual framework of
allostasis (Stewart 2006). Allostasis is the maintenance of stability (in traits key to
immediate survival-pH, glucose, temperature, oxygen levels) through change in other
physiological systems or through behavioural changes (e.g. the laying of smaller clutches
found in chapter 3) (McEwen and Seeman 1999). When the body is challenged (e.g. as
mundane as standing up in the morning or as serious as fleeing a predator), stability in the
four key traits is maintained by honnone regulated changes in physiology (e.g. mobilise
fat stores when glucose is low)(McEwen and Wingfield 2003). When larger challenges
are frequent and long lasting, the bodies physiological response inevitably leads to wear
and tear on its own system (i .e. allostatic load) (Seeman et al. 200 I, McEwen 2008).
Allostasis and allostatic load appear to provide a good explanation for why we
found significant but weak correlation among traits (chapter 2) and why we might expect
that predation effects were only moderate but spread over many traits (chapter 3).
Allostasis is regulated by honnones which act (sometimes in detrimental fashion) on
many tissues throughout the body and can have suppressive and stimulatory effects on
other honnones; e.g. as one mediator increases there are compensatory changes else
84�

where (McEwen 2008). This highlights the interconnected nature of organismal
physiology and would likely result in correlations between traits like we found in chapter
2. However, for much of the same reasons, the allostatic response can be very complex;
with multiple pathways to the same end point (McEwen 2008). In this case, the
relationship between traits would be weakened. Furthennore, multiple physiological
pathways to solve one challenge (e.g. recovery from egg laying) may result in spreading
physiological 'costs' over multiple traits. For these reasons, researchers who use the
concept of allostasis propose that obtaining a more comprehensive measure of
physiological condition will provide a better health index (Seeman et al. 2001).
One mechanism that generates allostatic load is repeated events that result JO
elevations of the al10static response (Logan and Barksdale 2008). It seems that nest
predation induced laying of addition clutches could be such a repeated event. A second
type of allostatic load involves the failure to habituate to the same stressor (i.e. the failure
ofthe body to dampen the honnonal response) (McEwen and Seeman 1999). Again, with
respect to predation, females that are laying additional clutches can not 'habituate' to the
reproductive honnones necessary for egg production. In this sense, exposure to estradiol
and subsequent proposed suppression of erythropoieses during laying would represent the
second type of alJostatic load.
4.2 Future Directions
Future studies should test if reductions in condition like those reported here affect
reproduction and survival outside of the experimental breeding season, as these data
85

would be critical to quantify the long tenn demographic costs of predation and increased
egg production. Directly manipulating condition in a manner predicted by the cost of egg
production would seem like the most expedient way to examine the long tenn effects of
increased egg laying. Administration of caffeine (Olcina et al. 2008) or nicotine (Neogy
et al. 2008) has previously been used to increase oxidative stress. And injections of
phenylhydrazine hydrochloride can be used to induce anaemia (Clark et al. 1988, Wagner
2008). These manipulations could be conducted at the end of the breeding season to
examine the effects of egg production on over winter survival and subsequent breeding
perfonnance.
In the present study we were not able to determine whether clutch size reductions
we reported are the result of egg production costs influencing clutch size decisions (i.e.
bet hedging strategy, Slagsvold 1984) or whether females reduced clutch size as a
strategy to limit the likelihood of future predation (i.e. nest survival strategy, Skutch
1949). As we suggested in chapter 3, are data are consistent with costs of egg production
influencing clutch size. However, our results are not inconsistent with females having
reduced clutch size to limit predation occurring on subsequent nest (i.e. reduced nest
duration or reduced conspicuousness hypothesis, Slagsvold 1982). Indirect evidence
reported in song sparrows suggests that reduction in clutch size may in fact reduce the
likelihood of predation occurring. Zanette et al. (2006) found that fed birds were less
likely to have their nests predated, which may result from fed females having longer
bouts on and shorter bouts off the nest during incubation (Duncan-Rastogi et al. 2006).
These results suggest that activity at the nest may in fact be an important factor
influencing predation. A valuable first step in differentiating between these two
86�

hypothesis would be to first test the relationship between clutch size and condition. This
could be accomplished using the phenotypic manipulations of oxidative stress and
hematocrit mentioned above. Regardless, of the specific nest predation hypothesis, this
work may provide valuable insight into proximate mechanisms affecting clutch size,
particularly as previous studies have found correlations between clutch size and both
oxidative stress (Bize et a1. 2008) and hematocrit (Dufva 1996).
4.3 References
Andersson, M. and L. Gustafsson. 1995. Glycosylated hemoglobin - a new measure of condition in birds. Proceedings of The Royal Society of London Series BBiological Sciences 260:299-303.
Atkinson, S. and M. A. Ramsay. 1995. The effects of prolonged fasting of the bodycomposition and reproductive success of female polar bears (Ursus-maritimus). Functional Ecology 9:559-567.
Bety, 1., G. Gauthier, and 1. F. Giroux. 2003. Body condition, migration, and timing of reproduction in snow geese: A test of the condition-dependent model of optimal clutch size. American Naturalist 162:110-121.
Bize, P., G. Devevey, P. Monaghan, B. Doligez, and P. Christe. 2008. Fecundity and survival in relation to resistance to oxidative stress in a free-living bird. Ecology 89:2584-2593.
Slums, P., J. D. Nichols, J. E. Hines, M. S. Lindberg, and A. Mednis. 2005. Individual quality, survival variation and patterns of phenotypic selection on body condition and timing of nesting in birds. Oecologia 143:365-376.
Bortolotti, G. R., R. D. Dawson, and G. L. Murza. 2002. Stress during feather development predicts fitness potential. Journal of Animal Ecology 71 :333-342.
Clark, M. W., R. P. Gildersleeve, J. P. Thaxton, C. R. Parkhurst, and D. 1. McRee. 1988, Hematological effects of ethyl methanesulfonate, paraquat and phenylhydrazine in japanese quail. Comparative Biochemistry and Physiology C-Phannacology Toxicology & Endocrinology 89: 15-30.
87

Dufva, R. 1996. Blood parasites, health, reproductive success, and egg volume in female Great Tits Panls major. Journal of Avian Biology 27:83-87.
Duncan-Rastogi, A, L. Zanette, and M. Clinchy. 2006. Food availability affects diurnal nest predation and adult antipredator behaviour in song sparrows, MeLospiza melodia. Animal Behaviour 72:933-940.
Glei, D. A., N. Goldman, Y. L. Chuang, and M. Weinstein. 2007. Do chronic stressors lead to physiological dysregulation? Testing the theory of allostatic load. Psychosomatic Medicine 69:769-776.
Hill, G. E. and K. L. Fanner. 2005. Carotenoid-based plumage coloration predicts resistance to a novel parasite in the house finch. Naturwissenschaften 92:30-34.
Logan, J. G. and D. J. Barksdale. 2008. Allostasis and allostatic load: expanding the discourse on stress and cardiovascular disease. Journal of Clinical Nursing 17:201-208.
McEwen, B. S. 1998. Protective and damaging effects of stress mediators. New England Joumal of Medicine 338: 171-179.
McEwen, B. S. 2008. Central effects of stress honnones in health and disease: Understanding the protective and damaging effects of stress and stress mediators. European Journal of Phannacology 583: 174-185.
McEwen, B. S. and T. Seeman. 1999. Protective and damaging effects of mediators of stress - Elaborating and testing the concepts of allostasis and allostatic load. Pages 30-47. New York Acad Sciences.
McEwen, B. S. and 1. C. Wingfield. 2003. The concept of allostasis in biology and biomedicine. Honnones and behavior 43:2-15.
Nelson, K. M., G. Reiber, T. Kohler, and E. J. Boyko. 2007. Peripheral arterial disease in a multiethnic national sample: The role of conventional risk factors and allostatic load. Ethnicity & Disease 17:669-675.
Neogy, S., S. Das, S. K. Mahapatra, N. Mandai, and S. Roy. 2008. Amelioratory effect of Andrographis paniculata Nees on liver, kidney, heari, lung and spleen during nicotine induced oxidative stress. Environmental Toxicology and Phannacology 25:321-328.
Ochs, C. L. and R. D. Dawson. 2008. Patterns of variation in leucocyte counts of female tree swallows, Taehycineta hieoLor: Repeatability over time and relationships with condition and costs of reproduction. Comparative Biochemistry and Physiology A-Molecular & Integrative Physiology 150:326-331.
88�

Olcina, G. J., R. Timon, D. Munoz, J. J. Maynar, M. J. Caballero, and M. Maynar. 2008. Caffeine ingestion effects on oxidative stress in a steady-state test at 75% V-02 (max). Science & Sports 23:87-90.
Romero, M. L. and M. Wekelski. 2001. Corticosterone levels predict survival probabilities of Galapagos marine iguanas during El Nino events. Proceedings of the National Academy of Sciences of the United States of America 98:7366-7370.
Seeman, T. E., B. S. McEwen, J. W. Rowe, and B. H. Singer. 2001. Allostatic load as a marker of cumulative biological risk: MacArthur studies of successful aging. Proceedings of the National Academy of Sciences of the United States of America 98:4770-4775.
Skutch, A. F. 1949. Do tropical birds raise as many young as they can nourish? Ibis 91:430-455.
Slagsvold, T. 1982. Clutch size vanatlOn In passenne birds - the nest predation hypothesis. Oecologia 54:159-169.
Slagsvold, T. 1984. Clutch size variation of birds in relation to nest predation - on the cost of reproduction. Journal of Animal Ecology 53:945-953.
Stewart, J. A. 2006. The Detrimental Effects of Allostasis:Allostatic Load as a Measure of Cumulative Stress. Journal of physiological Anthropology 25: 133-145.
Wagner, E. 2008. Anemia: A pysiological mechanism underlying the cost of egg production. Simon Fraser University, Burnaby.
Zanette, L., M. Clinchy, and J. N. M. Smith. 2006. Combined food and predator effects on songbird nest survival and annual reproductive success: results from a bifactorial experiment. Oecologia 147:632-640.
89�


















