Negative Regulation of Expression of the Nitrate As ... · J. Bacteriol. doi:10.1128/JB.01668-09 JB...
40
1 Negative Regulation of Expression of the Nitrate Assimilation nirA Operon in the Heterocyst-Forming Cyanobacterium Anabaena sp. Strain PCC 7120 José Enrique Frías and Enrique Flores* Instituto de Bioquímica Vegetal y Fotosíntesis, Consejo Superior de Investigaciones Científicas y Universidad de Sevilla, Centro de Investigaciones Científicas Isla de la Cartuja, E-41092, Seville, Spain. *Corresponding author. Instituto de Bioquímica Vegetal y Fotosíntesis, Centro de Investigaciones Científicas Isla de la Cartuja, Avenida Américo Vespucio 49, E-41092, Seville, Spain. Tel.: +34 954489523; fax: +34 954460065. E-mail address: [email protected] (E. Flores) Running title: Regulation of the nirA operon Keywords: Anabaena, cyanobacteria, nitrate assimilation, regulation Copyright © 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. J. Bacteriol. doi:10.1128/JB.01668-09 JB Accepts, published online ahead of print on 26 March 2010 on February 2, 2019 by guest http://jb.asm.org/ Downloaded from
Transcript of Negative Regulation of Expression of the Nitrate As ... · J. Bacteriol. doi:10.1128/JB.01668-09 JB...

1
Negative Regulation of Expression of the Nitrate Assimilation
nirA Operon in the Heterocyst-Forming Cyanobacterium
Anabaena sp. Strain PCC 7120
José Enrique Frías and Enrique Flores*
Instituto de Bioquímica Vegetal y Fotosíntesis, Consejo Superior de
Investigaciones Científicas y Universidad de Sevilla, Centro de
Investigaciones Científicas Isla de la Cartuja, E-41092, Seville, Spain.
*Corresponding author. Instituto de Bioquímica Vegetal y Fotosíntesis, Centro de
Investigaciones Científicas Isla de la Cartuja, Avenida Américo Vespucio 49, E-41092,
Seville, Spain. Tel.: +34 954489523; fax: +34 954460065.
E-mail address: [email protected] (E. Flores)
Running title: Regulation of the nirA operon
Keywords: Anabaena, cyanobacteria, nitrate assimilation, regulation
Copyright © 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.J. Bacteriol. doi:10.1128/JB.01668-09 JB Accepts, published online ahead of print on 26 March 2010
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

2
Abstract
In the filamentous, heterocyst-forming cyanobacterium Anabaena sp. strain
PCC 7120, expression of the nitrate assimilation nirA operon takes place in the
absence of ammonium and the presence of nitrate or nitrite. Several positive-
action proteins that are required for expression of the nitrate assimilation nirA
operon have been identified. Whereas NtcA and NtcB exert their action by direct
binding to the nirA operon promoter, CnaT acts by an as-yet-unknown
mechanism. In the genome of this cyanobacterium, ORF all0605 (the nirB gene) is
found between the nirA (encoding nitrite reductase) and ntcB genes. A nirB mutant
was able to grow at the expenses of nitrate as a nitrogen source and showed
abnormally high levels of nirA operon mRNA both in the presence and in the
absence of nitrate. This mutant showed increased nitrate reductase activity but
decreased nitrite reductase activity, an imbalance that resulted in excretion of
nitrite, which accumulated in the extracellular medium when the nirB mutant was
grown in the presence of nitrate. A deletion-in-frame nirA mutant also showed a
phenotype of increased expression of the nirA operon in the absence of ammonium
independent of the presence of nitrate in the medium. Both NirB and NirA are
therefore needed to keep low levels of expression of the nirA operon in the absence
of an inducer. Because NirB is also needed to attain high levels of nitrite reductase
activity, NirA appears to be a negative element in the nitrate-regulation of the
expression of the nirA operon in Anabaena sp. strain PCC 7120.
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

3
Assimilatory nitrate reduction is carried out by many plants, algae, fungi and
bacteria. It involves the uptake of nitrate into the cell and its two-step reduction via
nitrite to ammonium, which is incorporated into carbon skeletons. In bacteria, uptake is
carried out by ABC-type or MFS transporters, and reduction involves the direct transfer
of electrons to nitrate and nitrite via nitrate reductase and nitrite reductase, respectively,
from iron-sulfur or flavin-containing donor proteins (28). Expression of the nitrate
assimilation system is frequently subjected to a dual regulation: repression by
ammonium and induction by nitrate (or nitrite). Whereas repression is exerted by the
general nitrogen control system of the bacterium, a variety of different mechanisms
appear to exist that mediate induction (28).
Cyanobacteria are photoautotrophs that carry out oxygenic photosynthesis.
Nitrate and ammonium are excellent sources of nitrogen for cyanobacteria in general,
and many strains are able to use urea or to fix atmospheric nitrogen (13, 15). In
cyanobacteria, reduction of nitrate to ammonium is catalyzed by two ferredoxin-
dependent enzymes, nitrate reductase and nitrite reductase. Genes encoding nitrite
reductase (nirA), an ABC-type nitrate/nitrite uptake transporter (nrtABCD), and nitrate
reductase (narB) are clustered together constituting the nirA operon (nirA-nrtABCD-
narB) in the genomes of Synechococcus elongatus strain PCC 7942 (hereafter referred
to as S. elongatus) and Anabaena sp. strain PCC 7120(13). Several genes involved in
the biosynthesis of the nitrate reductase molybdenum cofactor (molybdopterin guanine
dinucleotide) and two additional genes, narM and nirB, that affect nitrate reductase and
nitrite reductase activity levels, respectively, have also been identified in S. elongatus
(13). The nirB gene has been shown to be required for attaining maximum levels of
nitrite reductase, and its inactivation provokes an imbalance between nitrate and nitrite
reduction resulting in release of nitrite to the external medium (36).
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

4
Nitrate reductase and nitrite reductase activities are lower in ammonium-grown
than in nitrate-grown cyanobacterial cells (13, 15). Expression of these enzyme
activities takes place at appreciable levels in the absence of nitrate or nitrite in some
cyanobacteria such as S. elongatus, but not in the heterocyst-forming, N2-fixing
cyanobacteria such as Anabaena sp. strain PCC 7120. Thus, in the non-N2-fixing
cyanobacteria the nitrate assimilation system is mainly subjected to ammonium-
promoted repression, whereas in the N2-fixing cyanobacteria, in addition to repression
by ammonium, induction by nitrate or nitrite is also required for attaining high levels of
expression, giving rise to a “nitrate effect” in this type of cyanobacteria (13, 15).
Expression of the nirA operon upon ammonium withdrawal is promoted by the
NtcA protein, a CAP-family transcription factor that is widespread among
cyanobacteria (25). NtcA activity is enhanced by 2-oxoglutarate, a putative C to N
balance signal in the cyanobacterial cell (17, 30) that can act on NtcA both directly (3,
33, 37-39) and indirectly via the signal-transduction protein PII (13, 33). In addition to
NtcA, a route-specific, LysR-type trancriptional regulator, NtcB, is involved in the
regulation of nirA operon expression (1, 2, 18, 27). In contrast to NtcA that is strictly
necessary for expression of the nirA operon in all investigated cyanobacterial strains,
NtcB is involved in regulation with different stringency levels depending on the
cyanobacterial strain. In the case of Anabaena sp. strain PCC 7120, the NtcB protein is
strictly required for expression of the nirA operon and for growth at the expense of
nitrate, and expression of ntcB itself takes place from an NtcA-dependent promoter (18).
A third positive regulatory element of nirA operon expression in Anabaena sp. strain
PCC 7120 is the CnaT protein (20), which shows overall sequence similarity to glycosyl
transferases. An Anabaena cnaT insertional mutant is unable to use nitrate as a nitrogen
source due to a defect in activation of transcription of the nirA operon. However, CnaT
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

5
does not appear to be a DNA-binding protein and, consequently, the effect of CnaT on
nirA operon expression could be indirect.
Four open reading frames (ORFs all0603, all0604, all0605 and all0606) are
located in the genome of Anabaena sp. strain PCC 7120 between the nirA and ntcB
genes, with the same orientation as ntcB (Fig. 1). all0603 would encode a 101-amino
acid transcriptional regulator of the XRE-family. all0604 would encode a 119-amino
acid polypeptide that shows no homology with any protein of known function. all0605
(previously designated orf398 (19) encodes a protein with sequence similarity to several
proteins previously characterized in S. elongatus: the phycocyanobilin lyase alpha
subunit CpcE (26 % identity in a 163 amino acid overlap), the NblB polypeptide
involved in phycobilisome degradation (22 % identity in a N-terminal 180 amino acid
overlap plus 23 % identity in a C-terminal 148 amino acid overlap), and the NirB
protein (23 % identity in a 331 amino acid overlap). all0606 (previously designated
orf136 (19) would encode a protein similar to the cytochrome b6f-complex iron-sulfur
protein PetC. Besides all0606, three other Anabaena sp. strain PCC 7120 ORFs,
all2453, all4511, and all1512, show overall homology to the cytochrome b6f-complex
iron-sulfur protein PetC (26).
In this study, we show that, in addition to the positive elements described above,
the expression of the Anabaena nirA operon is subjected to the action of two negative
elements, the products of ORF all0605 (the Anabaena nirB gene) and nirA, which
repress the expression of the nirA operon when nitrate is absent from the culture
medium giving rise to the aforementioned nitrate effect in Anabaena sp. strain PCC
7120.
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

6
MATERIALS AND METHODS
Strains and growth conditions. Anabaena sp. (also known as Nostoc sp.) strain
PCC 7120 was routinely grown photoautotrophically at 30°C under white light (about
25 µE s-1
m-2
), with shaking for liquid cultures. Media used for growth were BG11
(NaNO3 as the nitrogen source)(34), BG110 (BG11 without nitrate) or BG110NH4+
(BG110 supplemented with 2 mM NH4Cl and 4 mM N-tris(hydroxymethyl)methyl-2-
aminoethane sulfonic acid (TES)-NaOH buffer, pH 7.5). For growth on plates, medium
solidified with separately autoclaved 1% agar (Difco) was used. When appropriate,
antibiotics were added to plates at the following final concentrations: streptomycin
(Sm), 5 µg/ml; spectinomycin (Sp), 5 µg/ml; and neomycin (Nm), 30 µg/ml. In liquid
cultures, antibiotic concentrations used were as follows: Sm, 2 µg/ml; Sp, 2 µg/ml; and
Nm, 5 µg/ml. Strains CSE17 (19), CSE23, CSE271 and CSE272 were routinely grown
in BG110NH4+ medium supplemented with Sm and Sp. Strains EF116B and CSE27B
were routinely grown in BG110NH4+ medium supplemented with Nm. CSE27 and
CSE172 were routinely grown in BG110NH4+ medium.
For derepression experiments, cells grown in BG110NH4+ (BG110 supplemented
with 4 mM NH4Cl and 8 mM TES-NaOH buffer, pH 7.5) medium bubbled with a
mixture of air and CO2 (1% of CO2, v/v) at 30°C in the light (75 to 100 µE s-1
m-2
) were
harvested by filtration and washed with BG110 medium, resuspended in the media
indicated in each experiment and incubated under the same conditions. All media used
for derepression experiments were supplemented with 12 mM NaHCO3.
Escherichia coli DH5α, HB101, XL1-Blue and ED8654 were grown in Luria-
Bertani medium as described previously (4).
Generation of mutant strains. The method of sacB-mediated positive selection
for double recombinants in Anabaena sp. (7) was used to generate mutant strains
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

7
EF116B (nrtB::C.K1), CSE27 (∆nirA), CSE27B (∆nirA nrtB::C.K1) and CSE23
(all0604::C.S3), and to complement mutant strain CSE17 (all0605::C.S3). Plasmids
pCSE111B (for CSE23), pCSE129 (for CSE172), pCSE142B (for CSE27), pCSE149
(for CSE271 and CSE272) and pCSE152B (for EF116B and CSE27B) were transferred
to the cyanobacterial parental strain by conjugation (11). See Table 1 for a description
of strains and plasmids. Plasmid pRL623 was used as helper plasmid in conjugations,
except for the generation of strain CSE23 for which plasmids pRL528 and pRL591-
W45 were used as helper plasmids. In all cases plasmid pRL443 was used as
conjugative plasmid. For generation of strains CSE27 and CSE172, some sucrose-
sensitive exconjugants (SmrSp
r for CSE27; Sm
rSp
rNm
r for CSE172) were grown in
BG110NH4+ liquid medium without antibiotics. These cultures were sonicated in a
cleaning bath and plated on BG110NH4+ solid medium containing 5% sucrose. Double
recombinants were identified by their sucrose-resistant antibiotic-sensitive phenotype.
In CSE27, a 666-bp internal fragment of nirA, corresponding to nucleotides 598-1263
of the 1611-nucleotide-long coding region, was deleted from the genome, and as a
consequence of the in-frame deletion of nucleotides, the modified nirA should encode a
protein of 304 amino acid residues. In all cases the genomic structure of the resultant
Anabaena mutant strain was checked by Southern analysis or PCR analysis to confirm
the absence of parental chromosomes in the resultant Anabaena strains.
DNA isolation and Southern blot analysis. Isolation of DNA from Anabaena
sp. was performed as previously described (7). For Southern blots, restriction
endonuclease-digested DNA was subjected to electrophoresis in agarose gels and
transferred to Hybond™-N+ membranes following the instructions of the manufacturer.
Labeling of probes with 32
P and hybridization was performed as described previously
(9, 21).
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

8
RNA isolation and analysis. RNA from Anabaena sp. was prepared as
described previously (29). The resulting RNA preparations were treated with RNase-
free DNase I to eliminate contaminating DNA. For Northern blot analysis, RNA
(approximately 20-25 µg) was subjected to electrophoresis in denaturing formaldehyde
gels, transferred to Hybond™-N+ membranes, and subjected to hybridization at 65°C as
described previously (9). DNA probes (see Table 2) used in Northern experiments
were: nirA probe, PCR-generated DNA fragment using primers nir-7120-15 and nir-
7120-16 (probe a), nir-7120-23 and nir-7120-25 (probe b), or nir-7120-30 and nir-7120-
31 (probe c); narB probe, PCR-generated DNA fragment using primers N-narB-7120
and C-narB-7120; ntcA probe, NcoI/SalI DNA fragment from pCSAM61 (29); ntcB
probe, PCR-generated DNA fragment using primers Nc-ntcB and ntcB-3 (18); cnaT
probe, HincII/BstXI DNA fragment from pCSE118 (20) bearing most of the Anabaena
cnaT gene; all0603 probe, PCR-generated DNA fragment using primers all0603-1 and
all0603-2; all0604 probe, PCR-generated DNA fragment using primers all0604-1 and
all0604-2; and all0605 (nirB) probe, PCR-generated DNA fragment using primers
orf398-2 and nir-7120-10. For PCR-generated probes, Anabaena chromosomal DNA,
plasmid pCSE21 (19) or plasmid pCSE95 (18) was used as a template. Primer extension
experiments were performed as described elsewhere (4), using 20-25 µg of RNA and
primer nir-1 (19). Results were visualized and quantified with a Cyclone storage
phosphor system and OptiQuant image analysis software (Packard).
Nitrate uptake. Nitrate uptake assays were performed as previously described
(14). Ammonium-grown cells (4 to 5 µg of chlorophyll a/ml) were derepressed by
incubation for 4 h in BG11 medium. Cells were then harvested by filtration, washed
with 10 mM Tricine-NaOH buffer (pH 8.1), resuspended in the same buffer to 10 µg of
chlorophyll a/ml, and incubated under culture conditions. Uptake assays were started by
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

9
addition of NaNO3 (100 to 110 µM, final concentration). Nitrate disappearance was
determined by estimating the concentration of nitrate in the medium in aliquots of the
cell suspensions after removal of the cells by filtration through Millipore HA 0.45-mm-
pore-size filters. Nitrate concentration was determinated by HPLC using a Partisil 10
SAX WCS Analytical Column (4.6 mm x 250 mm; 10 µm) from Whatman International
Ltd (England).
Enzyme activities. Nitrate reductase (23) and nitrite reductase (24) were
measured with dithionite-reduced methyl viologen as the reductant in cells made
permeable with mixed alkyltrimethylammonium bromide. The amount of cells added to
an enzymatic assay for nitrate reductase and nitrite reductase contained 5 and 25 µg of
chlorophyll a, respectively. Activity units correspond to µmol of nitrite produced
(nitrate reductase) or removed (nitrite reductase) per minute.
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

10
RESULTS AND DISCUSSION
Verification of strain CSE17. Construction of mutant strain CSE17
(all0605::C.S3 or nirB::C.S3) has been previously described (19). When ammonium-
grown cells of this mutant were incubated in the absence of ammonium in cultures
bubbled with CO2-enriched air, they showed abnormally low levels of nitrite reductase
activity in nitrate-containing medium and abnormally high levels of nitrate reductase
activity in medium containing no combined nitrogen (see below). This data confirm
previously reported data (19).
To verify that the phenotype shown by strain CSE17 results from inactivation of
all0605 (nirB) and not from any additional mutation elsewhere in the genome, the
mutated version of nirB in CSE17 was replaced by a wild-type version of the gene (see
Materials and Methods for details). In strain CSE172, the genomic structure in the nirB
region was identical to that of the wild-type strain PCC 7120 (not shown). Nitrate and
nitrite reductase levels in strain CSE172 were similar to those of the wild type rather
than to those of its parental strain CSE17 showing that the phenotype of strain CSE17
results from the inactivation of nirB.
Two small ORFs, all0604 and all0603, whose coding sequences overlap by 3
nucleotides, are located downstream of all0605 (Fig. 1). The gene-cassette inserted into
all0605 in strain CSE17, C.S3 (12), contains transcriptional terminators that are
effective in Anabaena sp. strain PCC 7120 (see, e. g., 19). To verify that the phenotype
of strain CSE17 results from inactivation of all0605 and not from a polar effect on a
downstream gene, the expression of all0604-all0603 was investigated and a mutant of
all0604 was constructed with the same cassette, C.S3 (see Fig. 1 and Materials and
Methods for details). Mutant strain CSE23 (all0604::C.S3) was able to grow in media
containing ammonium, nitrate or no combined nitrogen and exhibited nitrate reductase
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

11
and nitrite reductase activity levels that were similar (within 10 %) to those of the wild-
type strain. Additionally, the expression of nirA and narB in mutant strain CSE23,
investigated by Northern analysis, showed no appreciable difference to that observed in
the wild-type strain PCC 7120 (not shown).
To investigate the expression of all0604 and all0603, Northern analysis was
performed with probes of each of these ORFs and RNA isolated from strains PCC 7120,
CSE17 and CSE172. The all0604 probe hybridized to bands of about 0.33 kb (likely
corresponding to an all0604 transcript) and 0.87 kb (likely corresponding to a transcript
covering both all0604 and all0603) (Fig. 2). Only the latter band was obtained using an
all0603 probe and no expression of all0603 was observed in mutant strain CSE23
(all0604::C.S3) (not shown). Thus, a bicistronic transcript is produced, but a
monocistronic transcript is also observed that may result from premature transcription
termination downstream of all0604. The 0.87-kb bicistronic transcript accumulated at
an increased level after 24 h incubation in the absence of combined nitrogen. No
significant differences in the expression profiles of all0603 and all0604 were observed
between strains PCC 7120, CSE17, and CSE172 (Fig. 2). The results of analysis of the
nirB mutant (all0605::C.S3) and of expression of all0604-all0603 both indicate that the
phenotype of CSE17 cannot be ascribed to a polar effect on the expression of
downstream genes.
Effect of inactivation of nirB on the expression of nitrate assimilation
enzyme activities and genes. The development of the nitrate and nitrite reductase
activities was analyzed in cultures of wild-type strain PCC 7120 and mutant strain
CSE17 during a 24-h induction experiment in nitrate-containing media. In the wild type
(Fig. 3A), both enzymatic activities reached a plateau after 6 h of incubation in nitrate-
containing media, with activities of about 89 mU (mg of protein)-1
for nitrate reductase
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

12
and about 23 mU (mg of protein)-1
for nitrite reductase. In the mutant (Fig. 3B), the
nitrite reductase activity also reached a plateau at 6 h, showing an activity of about 11
mU (mg of protein)-1
, whereas the nitrate reductase activity increased during the course
of the experiment to reach an activity of about 131 mU (mg of protein)-1
at 24 h. An
accumulation of nitrite in the culture medium was observed specifically for strain
CSE17 indicating that its nitrite reductase is unable to reduce all the nitrite produced by
nitrate reductase, the excess nitrite being excreted.
The development of nitrate reductase in strains PCC 7120, CSE17 and CSE172
was also studied in media lacking combined nitrogen (Fig. 4). Expression was observed,
and after reaching a peak shortly after the beginning of the incubation, the activity
decayed in the wild type but much less in strain CSE17. Strain CSE172 behaved like the
wild type. The level of nitrate reductase activity after 24 h in the absence of nitrate was
about 15-fold higher in mutant CSE17 than in the wild type. These results show that
inactivation of nirB has profound effects on the enzyme activities in the nitrate
assimilation system resulting in low levels of nitrite reductase and, especially when the
cells are incubated in the absence of nitrate, abnormally high levels of nitrate reductase.
To test whether the abnormal levels of nitrate and nitrite reductase activities
shown by strain CSE17 resulted from an altered expression of the nirA operon,
Northern analyses were carried out with probes of the nirA and narB genes.
Hybridizations were performed with total RNA isolated from cells of strains PCC 7120,
CSE17, and CSE172 incubated under different nitrogen regimes. As previously reported
for the nirA operon (19), which produces a long transcript of close to 10 kb that is
unstable, only a smear of degraded RNA products could be detected with both the nirA
and narB probes (Fig. 5). In strains PCC 7120 and CSE172, a high level of expression
of narB took place only in media without ammonium in the presence of nitrate. In the
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

13
nirB mutant (strain CSE17), however, a high level of expression was observed in the
absence of ammonium both in the presence and absence of nitrate (Fig. 5A). A similar
expression profile was obtained with the nirA probe (Fig. 5C). Levels of the 5’ region of
the nirA operon transcript in strain CSE17 were also analyzed by primer extension. The
obtained data (Fig. 6) corroborated those obtained by Northern experiments, suggesting
that the increased nirA operon transcript levels observed in strain CSE17 in the absence
of combined nitrogen corresponded to a higher utilization of the nirA operon promoter
in this strain.
To test whether the expression of the positive regulatory elements NtcA, NtcB
and CnaT was affected in mutant strain CSE17, Northern analysis was performed. The
expression patterns observed for the ntcA, ntcB and cnaT genes under the different
nitrogen regimes tested were similar in strains PCC 7120, CSE17 and CSE172 (Fig. 5)
and similar to those previously reported (18, 20, 29). These results indicate that NirB is
not involved in the regulation of transcription of these genes and, additionally, that the
altered levels of nirA operon expression observed in strain CSE17 do not result from an
altered expression of the positive regulatory elements NtcA, NtcB and CnaT. Therefore,
NirB appears to be a factor inhibiting the expression of the nirA operon, especially in
the absence of nitrate, in Anabaena sp. strain PCC 7120.
The accumulation of nirA operon transcripts in strain CSE17, observed with
probes of nirA and narB, explains the higher nitrate reductase activity observed in this
mutant in both the presence and absence of nitrate; however, it does not explain the low
levels of nitrite reductase activity observed in the presence of nitrate. A similar
phenotype of expression of low levels of nitrite reductase activity has been observed for
a nirB mutant of S. elongatus, although no effect of the corresponding mutation on the
nirA operon transcript level has been reported (35). It has been proposed that NirB is
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

14
required as a chaperone or scaffold for expression of maximum nitrite reductase activity
(35). Similarity of NirB to CpcE and NblB is based on the presence in these proteins of
a number of HEAT-like repeats, which consist of tandemly repeated amino acid
modules that appear to function in protein-protein interactions and may have a
scaffolding role (22). Anabaena NirB appears to have six such HEAT repeats. A role
based on the presence of these structural motifs would explain why NirB from S.
elongatus and Anabaena sp. strain PCC 7120, two homologous proteins that appear to
have a similar function in nitrate reduction and are therefore orthologues, show such a
low identity degree (23 %). If NirB works through protein-protein interactions, it is
conceivable that NirB interacts with the nitrite reductase allowing it to reach a
maximally active conformation for the development of its enzymatic activity. In turn,
this raises the possibility of a regulation of the nirA operon by nitrite reductase itself
rather than by NirB.
Study of a nirA mutant. Different mutant strains in the Anabaena nirA operon
generated by lux-tagged Tn5 transposon mutagenesis have been described (6). One of
those mutants, strain TLN10 (nirA::Tn5), shows a phenotype similar to that of the nirB
mutant. When cells are transferred from ammonium-containing medium to medium
lacking combined nitrogen, a luciferase activity develops in strain TLN10 that is very
high as compared to that showed by the mutant strains TLN12 (nrtC::Tn5) and TLN21
(Tn5 inserted in the nrtD-narB intergenic region) (6). Given that in both cases, the nirB
mutant and strain TLN10, the nitrite reductase activity is affected (we assume that the
Tn5 insertion inactivates nitrite reductase), we decided to mutate the nirA gene in order
to check whether the nitrite reductase is involved in the regulation of expression of the
nirA operon.
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

15
A nirA mutant, strain CSE27, was constructed by deleting in frame a 666-bp
internal fragment of nirA, corresponding to nucleotides 598-1263 of the 1611-
nucleotide-long coding region (see Fig. 1 and Materials and Methods for details). When
nitrate was used as a nitrogen source, strain CSE27 was unable to grow and nitrite
accumulated in the culture medium (not shown). The expression of nirA in mutant strain
CSE27 was investigated by Northern analysis using RNA isolated from strains PCC
7120, CSE17 and CSE27 (Fig. 7). As is the case for strain CSE17, a high expression of
the nirA operon in the absence of ammonium in both the presence and absence of nitrate
was observed in strain CSE27, and the observed levels of expression were even higher
than in strain CSE17.
To verify that the phenotype shown by strain CSE27 resulted from inactivation
of nirA, a wild-type version of the nirA gene bearing the complete nirA operon promoter
was introduced into strain CSE27. Plasmid pCSE149 (see Table 1), a derivative of
pCSEL24 (31), can recombine with the nirA region in the chromosome and with the
nucA region in the Anabaena alpha megaplasmid. In the exconjugant strains CSE271
and CSE272, pCSE149 was integrated into the alpha megaplasmid (Fig. 8A) and into
the chromosome (Fig. 8B), respectively, in such a way that in both cases one copy of
the nirA operon was maintained as in CSE27 and an intact nirA gene together with its
promoter was additionally present. The genomic structure of these strains was
confirmed by PCR analysis using different primer pairs (nui-7120-4 and nir-7120-28;
nir-7120-27 and nrtA-7120-2; nir-7120-6 and orf398-4; ntcs3 (19) and nrtA-7120-2;
ntcs3 and nrtA-7120-3; see Table 2). Both strains were able to grow using nitrate as a
nitrogen source. The expression of narB was investigated by Northern analysis using
total RNA isolated from strains CSE271 and CSE272 and, as controls, PCC 7120 and
CSE27 (Fig. 8C). In contrast to strain CSE27, strains CSE271 and CSE272 presented an
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

16
expression of narB similar to that shown by strain PCC 7120. These results indicate that
the introduction of a wild-type copy of nirA into strain CSE27 restored a wild-type
phenotype of nirA operon expression at the mRNA level. Using the DNA fragment
deleted in CSE27 as a nirA probe, we were also able to corroborate an ammonium- and
nitrate-regulated expression of the wild-type copy of nirA introduced in strains CSE271
and CSE272 (Fig. 8D). Interestingly, however, the nirA copy in the alpha megaplasmid
(strain CSE271) was expressed at higher levels than that present in the chromosome
(strain CSE272).
As described in the Introduction, a high level of expression of the nirA operon in
Anabaena sp. strain PCC 7120 requires both the absence of ammonium and the
presence of nitrate or nitrite, which act as inducers, in the culture medium. The results
described above suggest that nitrite reductase has, in addition to its role in nitrite
reduction to ammonium during nitrate assimilation, a negative role in the expression of
the nirA operon when this cyanobacterium is incubated in the absence of both
ammonium and nitrate in the culture medium. A simple interpretation of our results
would be that in the nirB and nirA mutants, which have low levels or lack nitrite
reductase, respectively, nitrite accumulates in the cells up to levels that strongly induce
the expression of the nirA operon. However, whereas this interpretation would be
evident for those cultures supplemented with nitrate, we observed similarly increased
nirA operon mRNA levels in the nirB and nirA mutants incubated in the absence as well
as in the presence of added nitrate. The possibility remains, nevertheless, that traces of
nitrate or nitrite present in the BG110 medium are concentrated within the cells to act as
inducers.
Mutants of the NrtABCD transporter. An Anabaena ABC-type nitrate
transporter, NrtABCD, has been previously identified (19). To check whether nitrate
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

17
concentrated within the cells from traces present in the medium could be acting as an
inducer, the nrtB gene, which encodes the transmembrane component of the nitrate
transporter, was mutated in strain CSE27 and in its parental strain EF116 (see Table 1
for details). For this, the C.K1 cassette, which confers resistance to neomycin and does
not bear transcriptional terminators, was introduced into the ScaI restriction site of nrtB
in the same orientation as this gene (see Fig. 1 and Materials and Methods for details).
Two mutants, strains EF116B (nrtB::C.K1) and CSE27B (∆nirA, nrtB::C.K1), were
obtained that were defective in the active transport of nitrate (Fig. 9A). The expression
of the nirA gene was studied by Northern analysis, and both mutants presented an
expression profile similar to that shown by their respective parental strains (Fig. 9B).
The fact that similar expression levels were observed in the presence and absence of
added nitrate in spite of the absence of an active transporter makes it unlikely the
possibility of a role of traces of nitrate in regulation.
The Anabaena NrtABCD transporter also mediates the uptake of nitrite into the
cells (ref. 32 and our unpublished results), but nitrite can enter into the cyanobacterial
cells also by means of diffusion of nitrous acid (16). Diffusion is not however a
mechanism that could concentrate nitrite within the cells, and direct determination of
nitrite in cell extracts from filaments incubated without added nitrate under the
conditions described in this work rendered negligible levels of nitrite (not shown). Our
results argue against accumulation of an inducer (nitrate or nitrite) as responsible for the
increased levels of the expression of the nirA operon observed in nirB and nirA mutants.
They suggest instead that NirA (the nitrite reductase) and NirB (a possible nitrite
reductase chaperone) somehow inhibit the expression of the nirA operon when the wild-
type Anabaena filaments are incubated in the absence of both ammonium and nitrate.
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

18
Concluding remarks. As mentioned above, both Anabaena mutants, nirA and
nirB, have lost the nitrate effect on expression of the nirA operon (see Fig. 7). We do
not know whether the observed phenotype of the nirB mutant just results from the lack
of NirB protein or is related to the possible role of NirB as scaffolding protein in the
maturation of the nitrite reductase. If the latter were the case, the nitrite reductase would
be the key element in the negative regulation of the nirA operon in the absence of
ammonium. How nitrite reductase could exert this negative role remains to be
elucidated. A plausible hypothesis is that the nitrite reductase acts as a modulator of a
transcription factor. Because NtcA is a general N-control transcription factor and NtcB
is a nitrate assimilation pathway-specific transcriptional activator, NtcB could be
favored as a candidate target for nitrite reductase. Indeed, a role of NtcB mediating a
nitrite effect in the expression of the nirA operon in S. elongatus has been suggested (1).
The nitrite reductase could switch from being an inhibitor of the NtcB transcriptional
activator in the absence of nitrate or nitrite to acting as an enzyme in the presence of
these nitrogen sources. It has recently been found that some enzymes, termed ‘trigger
enzymes’, can control gene expression in response to the availability of their substrates
through different mechanisms: by binding to either DNA or RNA or by modulating of
the activity of transcription factors through either covalent modification or protein-
protein interactions (8). Our results suggest that nitrite reductase from Anabaena sp.
strain PCC 7120 is a ‘trigger enzyme’ that, in addition to its role in nitrite reduction to
ammonium, exerts a regulatory role on the nitrate assimilation system.
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

19
ACKNOWLEDGMENTS
We thank A. M. Muro-Pastor and A. Valladares for their help during this work
and A. Herrero for discussion. Use of DNA sequences from the DOE-Joint Genome
Institute (USA) and Kazusa DNA Research Institute (Japan) databases is acknowledged.
This work was supported by grant numbers BFU2005-07672 and BFU2008-03811 from
the Ministerio de Ciencia y Tecnología (Spain).
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

20
REFERENCES
1. Aichi, M., and T. Omata. 1997. Involvement of NtcB, a LysR family
transcription factor, in nitrite activation of the nitrate assimilation operon in the
cyanobacterium Synechococcus sp. strain PCC 7942. J. Bacteriol. 179:4671-
4675.
2. Aichi, M., N. Takatani, and T. Omata. 2001. Role of NtcB in activation of
nitrate assimilation genes in the cyanobacterium Synechocystis sp. strain PCC
6803. J. Bacteriol. 183:5840-5847.
3. Aldehni, M. F., J. Sauer, C. Spielhaupter, R. Schmid, and K. Forchhammer.
2003. Signal transduction protein P(II) is required for NtcA-regulated gene
expression during nitrogen deprivation in the cyanobacterium Synechococcus
elongatus strain PCC 7942. J. Bacteriol. 185:2582-2591.
4. Ausubel, F. M., R. Brent, R. E. Kingston, D. Moore, J. G. Seidman, J. A.
Smith, and K. Struhl. 2009. Current protocols in Molecular Biology. Greene
Publishing and Wiley-Interscience, New York.
5. Black, T. A., Y. Cai, and C. P. Wolk. 1993. Spatial expression and
autoregulation of hetR, a gene involved in the control of heterocyst development
in Anabaena. Mol. Microbiol. 9:77-84.
6. Cai, Y., and C. P. Wolk. 1997. Nitrogen deprivation of Anabaena sp. strain
PCC 7120 elicits rapid activation of a gene cluster that is essential for uptake
and utilization of nitrate. J. Bacteriol. 179:258-266.
7. Cai, Y. P., and C. P. Wolk. 1990. Use of a conditionally lethal gene in
Anabaena sp. strain PCC 7120 to select for double recombinants and to entrap
insertion sequences. J. Bacteriol. 172:3138-3145.
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

21
8. Commichau, F. M., and J. Stülke. 2008. Trigger enzymes: bifunctional
proteins active in metabolism and in controlling gene expression. Mol.
Microbiol. 67:692-702.
9. Church, G. M., and W. Gilbert. 1984. Genomic sequencing. Proc. Natl. Acad.
Sci. U S A 81:1991-1995.
10. Elhai, J., A. Vepritskiy, A. M. Muro-Pastor, E. Flores, and C. P. Wolk.
1997. Reduction of conjugal transfer efficiency by three restriction activities of
Anabaena sp. strain PCC 7120. J. Bacteriol. 179:1998-2005.
11. Elhai, J., and C. P. Wolk. 1988. Conjugal transfer of DNA to cyanobacteria.
Methods Enzymol. 167:747-754.
12. Elhai, J., and C. P. Wolk. 1988. A versatile class of positive-selection vectors
based on the nonviability of palindrome-containing plasmids that allows cloning
into long polylinkers. Gene 68:119-138.
13. Flores, E., J. E. Frías, L. M. Rubio, and A. Herrero. 2005. Photosynthetic
nitrate assimilation in cyanobacteria. Photosynth. Res. 83:117-133.
14. Flores, E., M. G. Guerrero, and M. Losada. 1983. Photosynthetic nature of
nitrate uptake and reduction in the cyanobacterium Anacystis nidulans. Biochim.
Biophys. Acta. 722:408-416.
15. Flores, E., and A. Herrero. 1994. Assimilatory nitrogen metabolism and its
regulation, p. 487-517. In D. A. Bryant (ed.), The Molecular Biology of
Cyanobacteria. Kluwer Academic Publishers, Dordrecht.
16. Flores, E., A. Herrero, and M. G. Guerrero. 1987. Nitrite uptake and its
regulation in the cyanobacterium Anacystis nidulans. Biochim. Biophys. Acta
896:103-108.
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

22
17. Forchhammer, K. 1999. The PII protein in Synechococcus PCC 7942 senses
and signals 2-oxoglutarate under ATP-replete conditions, p. 549-553. In P. GA,
L. W, and G. Schmetterer (ed.), The Phototrophic Prokaryotes. Kluwer
Academic/Plenum Publishers, New York.
18. Frías, J. E., E. Flores, and A. Herrero. 2000. Activation of the Anabaena nir
operon promoter requires both NtcA (CAP family) and NtcB (LysR family)
transcription factors. Mol. Microbiol. 38:613-625.
19. Frías, J. E., E. Flores, and A. Herrero. 1997. Nitrate assimilation gene cluster
from the heterocyst-forming cyanobacterium Anabaena sp. strain PCC 7120. J.
Bacteriol. 179:477-486.
20. Frías, J. E., A. Herrero, and E. Flores. 2003. Open reading frame all0601
from Anabaena sp. strain PCC 7120 represents a novel gene, cnaT, required for
expression of the nitrate assimilation nir operon. J. Bacteriol. 185:5037-5044.
21. Frías, J. E., A. Mérida, A. Herrero, J. Martín-Nieto, and E. Flores. 1993.
General distribution of the nitrogen control gene ntcA in cyanobacteria. J.
Bacteriol. 175:5710-5713.
22. Groves, M. R., and D. Barford. 1999. Topological characteristics of helical
repeat proteins. Curr. Opin. Struct. Biol. 9:383-389.
23. Herrero, A., E. Flores, and M. G. Guerrero. 1985. Regulation of nitrate
reductase cellular levels in the cyanobacteria Anabaena variabilis and
Synechocystis sp. FEMS Microbiol. Lett. 26:21-25.
24. Herrero, A., and M. G. Guerrero. 1986. Regulation of nitrite reductase in the
cyanobacterium Anacystis nidulans. J. Gen. Microbiol. 132:2463-2468.
25. Herrero, A., A. M. Muro-Pastor, and E. Flores. 2001. Nitrogen control in
cyanobacteria. J. Bacteriol. 183:411-425.
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

23
26. Kaneko, T., Y. Nakamura, C. P. Wolk, T. Kuritz, S. Sasamoto, A.
Watanabe, M. Iriguchi, A. Ishikawa, K. Kawashima, T. Kimura, Y.
Kishida, M. Kohara, M. Matsumoto, A. Matsuno, A. Muraki, N. Nakazaki,
S. Shimpo, M. Sugimoto, M. Takazawa, M. Yamada, M. Yasuda, and S.
Tabata. 2001. Complete genomic sequence of the filamentous nitrogen-fixing
cyanobacterium Anabaena sp. strain PCC 7120. DNA Res. 8:205-213.
27. Maeda, S., M. Okamura, M. Kobayashi, and T. Omata. 1998. Nitrite-specific
active transport system of the cyanobacterium Synechococcus sp. strain PCC
7942. J. Bacteriol. 180:6761-6763.
28. Moreno-Vivián, C., and E. Flores. 2007. Nitrate Assimilation in Bacteria, p.
263-282. In H. Bothe, S. J. Ferguson, and W. E. Newton (ed.), Biology of the
Nitrogen Cycle. Elsevier B.V.
29. Muro-Pastor, A. M., A. Valladares, E. Flores, and A. Herrero. 2002. Mutual
dependence of the expression of the cell differentiation regulatory protein HetR
and the global nitrogen regulator NtcA during heterocyst development. Mol.
Microbiol. 44:1377-1385.
30. Muro-Pastor, M. I., J. C. Reyes, and F. J. Florencio. 2001. Cyanobacteria
perceive nitrogen status by sensing intracellular 2-oxoglutarate levels. J. Biol.
Chem. 276:38320-38328.
31. Olmedo-Verd, E., A. M. Muro-Pastor, E. Flores, and A. Herrero. 2006.
Localized induction of the ntcA regulatory gene in developing heterocysts of
Anabaena sp. strain PCC 7120. J. Bacteriol. 188:6694-6699.
32. Paz-Yepes, J., E. Flores, and A. Herrero. 2009. Expression and mutational
analysis of the glnB genomic region in the heterocyst-forming Cyanobacterium
Anabaena sp. strain PCC 7120. J. Bacteriol. 191:2353-2361.
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

24
33. Paz-Yepes, J., E. Flores, and A. Herrero. 2003. Transcriptional effects of the
signal transduction protein P(II) (glnB gene product) on NtcA-dependent genes
in Synechococcus sp. PCC 7942. FEBS Lett. 543:42-46.
34. Rippka, R., J. Deruelles, J. B. Waterbury, M. Herdman, and R. Y. Stanier.
1979. Generic assignments, strain histories and properties of pure cultures of
cyanobacteria. J. Gen. Microbiol. 111:1-61.
35. Suzuki, I., N. Horie, T. Sugiyama, and T. Omata. 1995. Identification and
characterization of two nitrogen-regulated genes of the cyanobacterium
Synechococcus sp. strain PCC7942 required for maximum efficiency of nitrogen
assimilation. J. Bacteriol. 177:290-296.
36. Suzuki, I., H. Kikuchi, S. Nakanishi, Y. Fujita, T. Sugiyama, and T. Omata.
1995. A novel nitrite reductase gene from the cyanobacterium Plectonema
boryanum. J. Bacteriol. 177:6137-6143.
37. Tanigawa, R., M. Shirokane, S. Maeda Si, T. Omata, K. Tanaka, and H.
Takahashi. 2002. Transcriptional activation of NtcA-dependent promoters of
Synechococcus sp. PCC 7942 by 2-oxoglutarate in vitro. Proc. Natl. Acad. Sci.
U S A 99:4251-4255.
38. Valladares, A., E. Flores, and A. Herrero. 2008. Transcription activation by
NtcA and 2-oxoglutarate of three genes involved in heterocyst differentiation in
the cyanobacterium Anabaena sp. strain PCC 7120. J. Bacteriol. 190:6126-6133.
39. Vázquez-Bermúdez, M. F., A. Herrero, and E. Flores. 2002. 2-Oxoglutarate
increases the binding affinity of the NtcA (nitrogen control) transcription factor
for the Synechococcus glnA promoter. FEBS Lett. 512:71-74.
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

25
40. Vioque, A. 1997. The RNase P RNA from cyanobacteria: short tandemly
repeated repetitive (STRR) sequences are present within the RNase P RNA gene
in heterocyst-forming cyanobacteria. Nucleic Acids Res. 25:3471-3477.
41. Wolk, C. P., Y. Cai, L. Cardemil, E. Flores, B. Hohn, M. Murry, G.
Schmetterer, B. Schrautemeier, and R. Wilson. 1988. Isolation and
complementation of mutants of Anabaena sp. strain PCC 7120 unable to grow
aerobically on dinitrogen. J. Bacteriol. 170:1239-1244.
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

26
Figure legends
FIG. 1. Genomic region of Anabaena sp. strain PCC 7120 bearing the nitrate
assimilation gene cluster. Genes and ORFs are indicated by thick arrows, which also
show the direction of transcription. Black arrows correspond to the ORFs investigated
in this work. The location of the restriction sites into which gene-cassettes (C.S3 for
strains CSE17 and CSE23; and C.K1 for strains EF116B and CSE27B) were inserted is
indicated. The region deleted from nirA in strain CSE27 is indicated as a ruled bar.
Abbreviations for some restriction endonuclease sites: B, BglI; C, ClaI; E5, EcoRV; H,
HindIII; P, PvuII; S, SpeI; Sc, ScaI; X, XbaI. Horizontal lines below the genes denote
probes used for Northern analyses.
FIG. 2. Northern analysis of the expression of all0604 in strains PCC 7120,
CSE17, and CSE172. Hybridization assays were carried out using RNA isolated from
cells grown with ammonium (NH4+) or grown with ammonium and incubated for 4 or
24 h in medium containing nitrate (NO3-) or no combined nitrogen (-N). Hybridization
to rnpB (40) served as a loading and transfer control (lower panel). The position and
size (indicated in kb) of the detected transcripts is shown on the left.
FIG. 3. Changes in levels of nitrate and nitrite reductase activities, and nitrite
accumulation in the culture medium after transfer of ammonium-grown cells to nitrate-
containing medium. (A) Strain PCC 7120 (WT); (B) strain CSE17. Nitrate reductase
activity (diamonds); nitrite reductase activity (squares); nitrite concentration in the
medium (triangles).
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

27
FIG. 4. Changes in levels of nitrate reductase activity after transfer of
ammonium-grown cells to a medium containing no combined nitrogen. Strain PCC
7120 (circles); strain CSE17 (squares); strain CSE172 (triangles).
FIG. 5. Northern analysis of the expression of the nirA operon and of some
regulatory genes in strains PCC 7120, CSE17, and CSE172. Hybridization assays were
carried out using RNA isolated from cells grown with ammonium (NH4+) or grown with
ammonium and incubated for 4 or 24 h in medium containing nitrate (NO3-) or no
combined nitrogen (-N). The hybridization probes used (see Materials and Methods for
details) corresponded to narB (A), cnaT (B), nirA (C; fragment “a” in Fig. 1), ntcA (D)
or ntcB (E). Hybridization to rnpB (40) served as a loading and transfer control for each
of the two filters used. The position of some size markers (A, C) or identified transcripts
(B, D, E) is shown, indicated in kb, on the left.
FIG. 6. Primer extension analysis of the expression of the nirA gene in strains
PCC 7120 and CSE17. Primer extension assays were carried out using oligonucleotide
nir-1 as a primer and RNA isolated from cells grown with ammonium (NH4+) or grown
with ammonium and incubated for 4 h in medium containing nitrate (NO3-) or no
combined nitrogen (-N). The arrowhead points to the extension product identifying the
Anabaena nirA operon tsp. The sequencing ladders presented were generated with the
same primer used in the primer extension reactions and plasmid pCSE26 as template.
FIG. 7. Northern analysis of the expression of nirA in strains PCC 7120, CSE17,
and CSE27. Hybridization assays were carried out using RNA isolated from cells grown
with ammonium (NH4+) or grown with ammonium and incubated for 4 in medium
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

28
containing nitrate (NO3-) or no combined nitrogen (-N). Fragment “b” in Fig. 1 was
used as nirA probe (see Materials and Methods for details). Hybridization to rnpB (40)
served as a loading and transfer control (lower panel). The position and size (indicated
in kb) of some size standards is shown on the left. The small changes in the pattern of
hybridization bands in strain CSE27 as compared to strains CSE17 and PCC 7120 result
from the deletion of 666 bp from nirA in CSE27.
FIG. 8. Genomic structure of nirA complementing constructs in strains CSE271
and CSE272 and Northern analysis of the expression of narB and nirA in strains PCC
7120, CSE27, CSE271 and CSE272. The complementing constructs included ORF
all0606, the nirA operon promoter region and the nirA gene incorporated into the nucA-
nuiA region of the alpha megaplasmid (A) or in the chromosomal nirA region (B).
Hybridization assays were carried out using RNA isolated from cells of the indicated
strains grown with ammonium (NH4+) or grown with ammonium and incubated for 4 in
medium containing nitrate (NO3-) or no combined nitrogen (-N). The hybridization
probes used (see Materials and Methods) corresponded to narB (C) and nirA (D;
fragment “c” in Fig. 1). Hybridization to rnpB (40) served as a loading and transfer
control. The position and size (indicated in kb) of some size standards is shown on the
left.
FIG. 9. Nitrate uptake and Northern analysis of the expression of nirA in strains
PCC 7120, EF116B, CSE27 and CSE27B. (A) Ammonium-grown cells of strains
PCC7120 (closed diamonds), EF116B (open diamonds), CSE27 (closed triangles) and
CSE27B (open triangles) were washed, resuspended in BG11 medium (17,6 mM
NaNO3), and incubated for 4 h as indicated in Materials and Methods for derepression
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

29
of the nitrate assimilation system. Cells were then harvested and used for nitrate uptake
assays (see Materials and Methods). At the indicated times, nitrate was measured by
HPLC using aliquots withdrawn from the assay mixture. (B) Hybridization assays were
carried out using RNA isolated from cells of the indicated strains grown with
ammonium (NH4+) or grown with ammonium and incubated for 4 in medium containing
nitrate (NO3-) or no combined nitrogen (-N). Fragment “b” in Fig. 1 was used as nirA
probe (see Materials and Methods). Hybridization to rnpB (40) served as a loading and
transfer control (lower panel). The position and size (indicated in kb) of some size
standards is shown on the left.
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

30
Table 1. Cyanobacterial strains and plasmids used in this study.
Strain or
plasmid
Relevant characteristics Reference
Strains
PCC 7120 Wild-type Anabaena strain (34)
EF116 Derivative of Anabaena sp. strain PCC 7120 unable to fix nitrogen under aerobic conditions (41)
EF116B Nmr derivative of strain EF116; nrtB::C.K1 This work
CSE17 SmrSp
r derivative of strain PCC 7120; all0605::C.S3 (19)
CSE172 SmsSp
s derivative of strain CSE17; all0605::C.S3 substituted by a wild-type version of
all0605
This work
CSE23 SmrSp
r derivative of strain PCC 7120; all0604::C.S3 This work
CSE27 Derivative of strain EF116; ∆nirA This work
CSE27B Nmr derivative of strain CSE27; ∆nirA nrtB::C.K1 This work
CSE271 SmrSp
r derivative of strain CSE27; pCSE149 plasmid integrated into nucA region of the
alpha megaplasmid
This work
CSE272 SmrSp
r derivative of strain CSE27; pCSE149 plasmid integrated into nirA region of the
Anabaena chromosome
This work
Plasmids
pCSE111B 2.79-kb SpeI fragment from pCSE73 (19), bearing AccI-ended gene-cassette C.S3 (12) inserted into the site ClaI of all0604, cloned in pRL278; used to generate mutant strain
CSE23
This work
pCSE129 1.94-kb XbaI/HindIII fragment from pCSE26 (19), that contains part of all0605 and the
whole all0606, cloned in pRL278; used to complement mutant strain CSE17
This work
pCSE142 1166-bp product of PCR with primers nir-7120-23, nir-7120-25, nir-7120-26 and nrtA-7120-
3, with pCSE26 as template; presents a deletion of a 666-bp internal segment of nirA,
corresponding to nucleotides 598-1263 of the 1611-nucleotide-long coding region
This work
pCSE142B 1.3-kb PvuII/SalI fragment from pCSE142 between the sites NruI/XhoI of pRL277; used to
generate mutant strain CSE27
This work
pCSE152B 2.9-kb ClaI/XbaI fragment from pCSE2 (19), bearing HincII-ended gene-cassette C.K1 (12)
inserted into the ScaI site of nrtB (in the same direction as nrtB), cloned in pRL277; used to
generate mutant strains EF116B, CSE17B and CSE27B
This work
pCSE149 5.3-kb EcoRV/PvuII fragment from pCSE78B (19) inserted between the sites EcoRV/NruI
of pCSEL24(31); used to complement mutant strain CSE27
This work
pRL277 SmrSp
r, sacB-carrying, mobilizable vector (5)
pRL278 Nmr, sacB-carrying, mobilizable vector (5)
pRL443 Kms derivative of conjugative plasmid RP4 (11)
pRL528 Mobilization-helper; encodes M.AvaI and M.Eco47II. (11)
pRL591-W45 Mobilization-helper; encodes M.EcoT22I (10)
pRL623 Mobilization-helper; encodes M.AvaI, M.Eco47II, and M.EcoT22I (10)
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

31
Table 2.
Oligodeoxynucleotide primers used in this work
Primer Sequence (5’-3’) nir-7120-6 GGT GTT GGT CGT GGG TAC
nir-7120-10 GCA AGC GAT CGC ACT GCC
nir-7120-15 GCA ACA GAC CGA GAT CAT CG
nir-7120-16 CCC CAT TCA TCA ATT AGC C
nir-7120-23 CTA CCC CCA AAG CCA GCC TC
nir-7120-25 GAG GGC GAA CGC ATG AAC TGA ATT ATC CC
nir-7120-26 ACG TTC ATG CGT TCG CCC TCA TCG AAA CC
nir-7120-27 CGG GAT AAT TCA GTT CAT GC
nir-7120-28 TTC CGG CAC GGG CGC ACA ATT TGG CAA CTT CG
nir-7120-30 GGA AAT CAA CGA TTT AGC CTT TGT TCC
nir-7120-31 GTT GCA AAA TTG TGC GCC CGT G
orf398-2 CAT AGT GCA GAT GAT TTG TC
orf398-4 TCA GTG CAG CGA TCG CAT GG
nrtA-7120-2 TCC AAT CTT GCC GCA TAC
nrtA-7120-3 TCT AGA GGA AGT ACA GCC ATG TAC C
N-narB-7120 GGA GCG AAG CGA CGT GAC
C-narB-7120 GGT CAG TTG GGT AAA CTC
all0603-1 CGA AGC CAT TTG ATG AAC
all0603-2 CAA TCG AAC TGA GAA ATC AC
all0604-1 AAT GAC CTG GGA GGT AGA G
all0604-2 TCA TAG GAA CCC CTC TG
nui-7120-4 ATG AGT GAG TCT GAA TAC CC
Introduced restriction enzyme cutting sites are shown in bold face.
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

B
Fig.1
(Frías and Flores)
E5
(CSE23) C.S3
HC PS XS
cnaT ntcB all0603 all0605 all0606 nirA nrtA nrtB nrtC nrtD narB
all0604 (nirB)
C.S3 (CSE17)
1 kbCSE27Sc
C.K1 (EF116B/CSE27B)
(a)(b)
(c)
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

24 h NO3-
NH4+
4 h NO3-
4 h -N
24 h -N
PC
C 7
12
0 C
SE
17
CS
E17
2
24 h NO3-
NH4+
4 h NO3-
4 h -N
24 h -N
24 h NO3-
NH4+
4 h NO3-
4 h -N
24 h -N
0.8
7
0.3
3
Fig
. 2
(Frías an
dF
lores)
(all0
604
)
(rnp
B)
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

0
20
40
60
80
100
120
140
0 5 10 15 20 25
time (h)
0
100
200
300
400
500
600
700
nit
rite
( µµ µµM
)
En
zym
ati
cact
ivit
y
(mU
/mg
pro
t)
20
40
60
80
100
120
140
100
200
300
400
500
600
700
nit
rite
( µµ µµM
)
A
B
En
zym
ati
cact
ivit
y
(mU
/mg
pro
t)
Fig. 3
(Frías and Flores)
WT
CSE17
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

0
10
20
30
40
50
60
0 5 10 15 20 25time (h)
En
zym
ati
cact
ivit
y
(mU
/mg
pro
t)
Fig. 4
(Frías and Flores)
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

Fig. 5
(Frías and Flores)
PCC 7120 CSE17 CSE172
24 h
NO
3-
NH
4+
4 h
NO
3-
4 h
-N
24 h
-N
24 h
NO
3-
NH
4+
4 h
NO
3-
4 h
-N
24 h
-N
24 h
NO
3-
NH
4+
4 h
NO
3-
4 h
-N
24 h
-N
A
2.82.3
1.5
0.5
(narB)
C
2.82.31.5
0.5
(nirA)
0.95
E(ntcB)
D
1.5
(ntcA)
B
1.1
(cnaT)
(rnpB)
(rnpB)
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

NH
4+
NO
3-
-N
PCC 7120 CSE17
NH
4+
NO
3-
-NA C G T
Fig. 6
(Frías and Flores)
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

Fig. 7
(Frías and Flores)
NO
3-
-N NH
4+
PCC 7120 CSE17 CSE27
2.8
2.3
1.5
NO
3-
-N NH
4+
NO
3-
-N NH
4+
(nirA)
(rnpB)
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

Fig. 8
(Frías and Flores)
1 kb
BE5
C.S3
nuiA nucA nirA all0606 nuiA nucA
X
(Alpha megaplasmid)
A6
7
B E5P X B
C.S3
E5XCSE27
all0605 all0606 nirA nucA nuiA all0606 nirA nrtA nrtB nrtC nrtD narB
B (Chromosome)
3
54
1
2
8
PCC 7120 CSE27 CSE271 CSE272
NO
3-
-N NH
4+
NO
3-
-N NH
4+
NO
3-
-N NH
4+
NO
3-
-N NH
4+
2.82.3
1.5
C(narB)
2.82.3
1.5
PCC 7120 CSE27 CSE271 CSE272
NO
3-
-N NH
4+
NO
3-
-N NH
4+
NO
3-
-N NH
4+
NO
3-
-N NH
4+
D(nirA)
(rnpB) (rnpB)
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
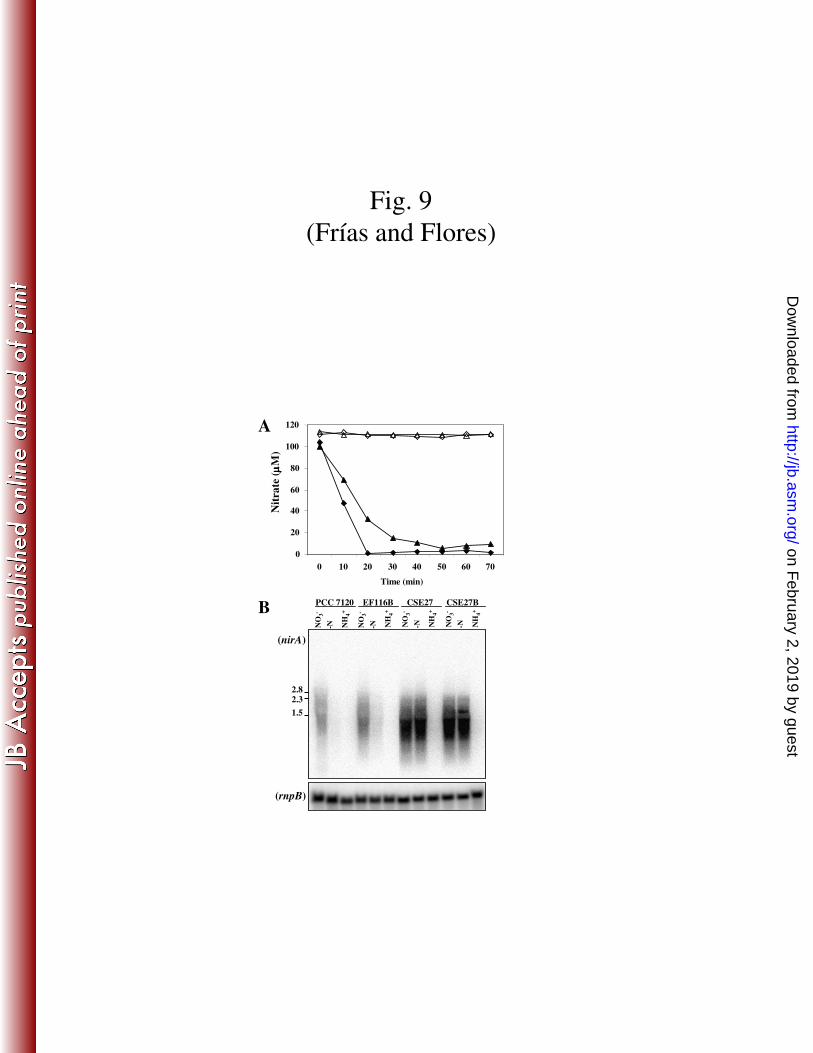
Fig. 9
(Frías and Flores)
0
20
40
60
80
100
120
0 10 20 30 40 50 60 70
Time (min)
Nit
ra
te (µµ µµ
M)
A
PCC 7120 EF116B CSE27 CSE27B
NO
3-
-N NH
4+
NO
3-
-N NH
4+
NO
3-
-N NH
4+
NO
3-
-N NH
4+
2.8
2.3
1.5
B
(nirA)
(rnpB)
on February 2, 2019 by guest
http://jb.asm.org/
Dow
nloaded from