Multidrug resistance proteins in rheumatoid arthritis, role in disease‐modifying antirheumatic...
12
REVIEW Multidrug resistance proteins in rheumatoid arthritis, role in disease-modifying antirheumatic drug efficacy and inflammatory processes: an overview G Jansen 1 , RJ Scheper 2 , BAC Dijkmans 1 Departments of 1 Rheumatology and 2 Pathology, VU University Medical Center, Amsterdam, The Netherlands Drug resistance is generally accepted as an important cause of treatment failure for patients with neoplastic or infectious diseases. Molecular mechanisms underlying drug resistance include the action of drug efflux pumps belonging to the super-family of ATP binding cassette (ABC) proteins, which mediate the cellular extrusion of a large variety of therapeutic drugs, a phenotype that is referred to as multidrug resistance (MDR). Unlike neoplastic and infectious diseases, chronic inflammatory diseases have received little attention. The potential role of ABC transporters in determining the efficacy of anti-rheumatic drugs, notably disease modifying anti- rheumatic drugs (DMARDs), in patients with rheumatoid arthritis is unclear. Based on knowledge from the field of oncology and immunology, this review concentrates on the pharmacological role of MDR proteins in the (clinical) efficacy of several DMARDs, as well as the physiological role of MDR proteins in transporting signalling molecules important in inflammatory processes. Key words: rheumatoid arthritis, DMARDs, multidrug resistance, ABC transporters, P-glycoprotein, multidrug resistance associated protein, methotrexate, sulphasalazine, chloroquine, review Rheumatoid arthritis (RA) is a common autoim- mune disease that is characterized by a chronic inflammation of the synovial joints, and infiltration by blood-derived cells (T cells, macrophages), leading to progressive destruction of cartilage and bone (1). Current medical therapy of patients with RA consists of treatment with: . Non-steroidal anti-inflammatory drugs (NSAIDs), which reduce pain and inflammation but do not alter the course of the disease . Disease modifying anti-rheumatic drugs (DMARDs), which can influence or modify the course of RA. Over the past decade ‘biologicals’ have also been introduced in the treatment of RA (2). These biologicals are specifically aimed at antibody-guided blocking of pro-inflammatory cytokines [e.g. tumour necrosis factor a (TNFa)] that play a major role in the pathophysiology of RA (3). Novel anti-TNFa agents, such as infliximab (4) or etanercept (5), displayed marked anti-rheumatic activity, but their long-term superiority over DMARDs — with known efficacy and low costs — still needs to be established, especially in the treatment of early RA. For this reason, DMARDs or combinations of DMARDs remain a common treatment option for patients with RA (6 – 9). However, it is well recognized that with successive treatment with DMARDs, many RA patients demonstrate loss of drug efficacy over time. Whether this is due to the onset of acquired drug resistance has not been extensively investigated. Meta-analyses of DMARD efficacy for treatment of RA patients showed that (hydroxy)chloroquine and sulphasalazine were among the DMARDs with the relatively shortest duration of efficacy (1 – 2 years), while methotrexate (MTX) had the longest- lasting efficacy (6 – 7 years) (10 – 13). Recently, a study by Morgan (14) showed that a small population of RA patients may be primary unresponsive to DMARDs, possibly via a multidrug resistance (MDR)-related mechanism. This review will further elaborate on the possible role of MDR proteins in the efficacy of DMARDs in RA treatment. However, it should be kept in mind that, as with anti-cancer drugs, DMARD resistance may be a multifactorial event, not only involving enhanced drug efflux via specific MDR proteins, but also other mechanisms, outlined in Figure 1A. In short, other mechanism of drug resistance can include: . Defective drug delivery (pharmacokinetic resis- tance) . Impaired drug uptake . Impaired drug activation . Enhanced drug detoxification/sequestration Gerrit Jansen, Department of Rheumatology, Rm 4A42, VU University Medical Center, De Boelelaan 1117, 1081 HV Amsterdam, The Netherlands. E-mail: [email protected] Received 11 September 2003 Accepted 11 September 2003 Scand J Rheumatol 2003;32:325–336 325 www.scandjrheumatol.dk # 2003 Taylor & Francis on license from Scandinavian Rheumatology Research Foundation DOI: 10.1080/03009740310004333 Scand J Rheumatol Downloaded from informahealthcare.com by QUT Queensland University of Tech on 11/01/14 For personal use only.
Transcript of Multidrug resistance proteins in rheumatoid arthritis, role in disease‐modifying antirheumatic...

REVIEW
Multidrug resistance proteins in rheumatoid arthritis, role indisease-modifying antirheumatic drug efficacy and inflammatoryprocesses: an overview
G Jansen1, RJ Scheper2, BAC Dijkmans1
Departments of 1Rheumatology and 2Pathology, VU University Medical Center, Amsterdam, The Netherlands
Drug resistance is generally accepted as an important cause of treatment failure for patients with neoplastic or
infectious diseases. Molecular mechanisms underlying drug resistance include the action of drug efflux pumps
belonging to the super-family of ATP binding cassette (ABC) proteins, which mediate the cellular extrusion of
a large variety of therapeutic drugs, a phenotype that is referred to as multidrug resistance (MDR). Unlike
neoplastic and infectious diseases, chronic inflammatory diseases have received little attention. The potential
role of ABC transporters in determining the efficacy of anti-rheumatic drugs, notably disease modifying anti-
rheumatic drugs (DMARDs), in patients with rheumatoid arthritis is unclear. Based on knowledge from the
field of oncology and immunology, this review concentrates on the pharmacological role of MDR proteins inthe (clinical) efficacy of several DMARDs, as well as the physiological role of MDR proteins in transporting
signalling molecules important in inflammatory processes.
Key words: rheumatoid arthritis, DMARDs, multidrug resistance, ABC transporters, P-glycoprotein, multidrug resistanceassociated protein, methotrexate, sulphasalazine, chloroquine, review
Rheumatoid arthritis (RA) is a common autoim-
mune disease that is characterized by a chronic
inflammation of the synovial joints, and infiltration
by blood-derived cells (T cells, macrophages),
leading to progressive destruction of cartilage and
bone (1). Current medical therapy of patients with
RA consists of treatment with:
. Non-steroidal anti-inflammatory drugs (NSAIDs),
which reduce pain and inflammation but do not
alter the course of the disease
. Disease modifying anti-rheumatic drugs (DMARDs),
which can influence or modify the course of RA.
Over the past decade ‘biologicals’ have also been
introduced in the treatment of RA (2). These
biologicals are specifically aimed at antibody-guided
blocking of pro-inflammatory cytokines [e.g. tumour
necrosis factor a (TNFa)] that play a major role in
the pathophysiology of RA (3). Novel anti-TNFaagents, such as infliximab (4) or etanercept (5),
displayed marked anti-rheumatic activity, but their
long-term superiority over DMARDs — with known
efficacy and low costs — still needs to be established,
especially in the treatment of early RA. For this
reason, DMARDs or combinations of DMARDs
remain a common treatment option for patients with
RA (6 – 9). However, it is well recognized that with
successive treatment with DMARDs, many RA
patients demonstrate loss of drug efficacy over time.
Whether this is due to the onset of acquired drug
resistance has not been extensively investigated.Meta-analyses of DMARD efficacy for treatment
of RA patients showed that (hydroxy)chloroquine
and sulphasalazine were among the DMARDs with
the relatively shortest duration of efficacy (1 – 2
years), while methotrexate (MTX) had the longest-
lasting efficacy (6 – 7 years) (10 – 13).
Recently, a study by Morgan (14) showed that a
small population of RA patients may be primary
unresponsive to DMARDs, possibly via a multidrug
resistance (MDR)-related mechanism.
This review will further elaborate on the possible
role of MDR proteins in the efficacy of DMARDs in
RA treatment. However, it should be kept in mind
that, as with anti-cancer drugs, DMARD resistance
may be a multifactorial event, not only involving
enhanced drug efflux via specific MDR proteins, but
also other mechanisms, outlined in Figure 1A. In
short, other mechanism of drug resistance can
include:
. Defective drug delivery (pharmacokinetic resis-
tance)
. Impaired drug uptake
. Impaired drug activation
. Enhanced drug detoxification/sequestration
Gerrit Jansen, Department of Rheumatology, Rm 4A42, VU
University Medical Center, De Boelelaan 1117, 1081 HV
Amsterdam, The Netherlands.
E-mail: [email protected]
Received 11 September 2003
Accepted 11 September 2003
Scand J Rheumatol 2003;32:325–336 325
www.scandjrheumatol.dk
# 2003 Taylor & Francis on license from Scandinavian Rheumatology Research Foundation
DOI: 10.1080/03009740310004333
Scan
d J
Rhe
umat
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y Q
UT
Que
ensl
and
Uni
vers
ity o
f T
ech
on 1
1/01
/14
For
pers
onal
use
onl
y.

. Altered (downstream) effects after drug – target
interaction (15).
(Multi) drug resistance
Drug resistance can be an inherent/intrinsic property
of cells, indicating that cells are resistant to specific
classes of drugs without ever been exposed to these
drugs previously. Alternatively, drug resistance
can be acquired, indicating that cells were primary
sensitive, but develop resistance over successive drugtreatments. Drug resistance can be restricted to one
specific class of drugs, e.g. antifolates like MTX,
or can extend to multiple structurally unrelated
drugs, a phenotype designated as MDR (16 – 21).
Molecular mechanisms of cellular drug resistance
can take place at various levels, involving,
for example, impaired cellular uptake, enhanced
metabolism, increased levels of target enzymes,defective apoptotic pathways and/or enhanced cel-
lular extrusion of drugs (15, 22).
This latter mechanism can be mediated by the
up-regulation of specific members of the family of
ATP binding cassette (ABC) transporters (17, 21,
23 – 26). Drug extrusion via these transporters is an
active process requiring ATP as an energy source
(Figure 1B). Data from the human genome projectrevealed that the ABC transporter family includes 48
proteins (25, 27, 28), divided into seven distinct
subfamilies (ABC A–G), some of which are
extensively characterized, while the functional prop-
erties of others are still unknown.
Several review papers have appeared over the past
few years covering specific aspects of ABC trans-
porters in relation to resistance to anti-cancer drugs
(17 – 19, 21, 23 – 26, 29 – 34). ABC transporters that
have an established or putative role in drug
resistance include (Table 1):
. ABCB1 [P-glycoprotein (Pgp), primary MDR
(MDR1)] (16, 35 – 37)
. ABCC1-7, ABCC10/11, also denoted as MDR-
associated protein 1 – 9 (MRP1 – 9) (38 – 49)
. ABCG2, also known as breast-cancer resistance
protein (BCRP) (20, 34, 50 – 53).
Pgp and MRPs are ‘full transporters’ with 12 – 17
transmembrane domains (TMDs) assembling a pore-
forming protein (25). In contrast, ABCG2 is referred
to as a ‘half transporter’ (six TMDs) requiring
homo- or hetero-dimerization to form a functionally
active protein (21, 54).
Although there is overlap in substrate specificity,
Pgp is known to prefer neutral hydrophobic sub-
strates, while MRPs and ABCG2 transport anionic
compounds, mostly as conjugates of glutathione,
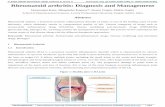
Figure 1. (A) Molecular mechanisms of cellular drug resistance. (1) Efficient drug delivery to target cells/tissues is a critical step in the
efficacy of drugs. Drug resistance due to diminished drug delivery is referred to as pharmacokinetic resistance. (2) Cell membrane
translocation of drugs can proceed via various routes, passive diffusion (a), facilitated diffusion (b) or via a carrier-mediated process
(c) (which is the case for MTX). Defective cellular uptake is a common resistance mechanism. (3) Prior to drug activation, cellular
extrusion may take place via specific efflux transporters, e.g. ABC transporters, thereby reducing the intracellular concentration of drug
available for therapeutic activity. (4) Drug activation, e.g. phosphorylation may be required for better intracellular retention and better
inhibition of the target. This process can be counteracted by enhanced drug sequestration (5), e.g. in subcellular compartments (e.g.
lysosomes), or detoxification (6), e.g. dephoshorylation, both of which can lead to drug resistance. Drug targeting (7) should contribute
to drug efficacy by damaging the target and inhibiting its function. (8) Several modes of escape from drug interactions exist, thereby
conferring resistance: they include up-regulation of target levels (e.g. increased DHFR in case of MTX resistance), modified/mutated
targets, enhanced repair of damage, impaired capacity for cells to go into apoptosis, or salvage of the damage by exogenous com-
pounds (e.g. purines and pyrimidines as a bypass for MTX effects (modified from Reference 22). (B) Energy-dependent cellular drug
extrusion via MDR transporters. Following entry of drugs (1 – 2), drugs can be subjected to extrusion (3 – 5) via an energy (ATP)-
dependent process involving an ABC transporter (4). Drugs can be extruded in co-transport with glutathione, or after conversion to
glutathione, glucuronide or sulphate conjugates.
326 G Jansen et al
www.scandjrheumatol.dk
Scan
d J
Rhe
umat
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y Q
UT
Que
ensl
and
Uni
vers
ity o
f T
ech
on 1
1/01
/14
For
pers
onal
use
onl
y.

glucuronides or sulphates (25, 55 – 60). Tissue
distribution studies showed that MDR proteins are
expressed at locations and by tissues that are heavily
exposed to toxic agents, microbials and exogenous
compounds (31, 53, 61, 62). Consistently, one of the
primary functions of MDR proteins is the extrusion
of toxic substances.MDR proteins Pgp and MRP1 have been detected
in immunologically relevant cells [T cells, monocytes,
dendritic cells (DCs) and macrophages] (61, 63 – 73),
where they are thought to play not only a
pharmacological, but also a physiological role in
inflammatory processes (see later sections). Interest-
ingly, ABCG2 has been identified in a side popula-
tion of CD34z haematopoietic progenitor cells (74 –76), although its pharmacological/physiological rele-
vance is still unclear.
MDR proteins and DMARDs/anti-inflammatory drugs
Substrate specificity studies suggest that several
DMARDs, NSAIDs and other anti-inflammatory
drugs (e.g. glucocorticoids) can be transported by,
or inhibit, specific MDR proteins (summarized inTable 2). The putative role of MDR proteins in loss
of efficacy/resistance for these anti-rheumatic drugs
is discussed in greater detail below.
Methotrexate
The folate antagonist MTX is the anchor drug in the
treatment of RA patients. The precise mechanism of
the anti-rheumatic action of MTX has still not been
fully elucidated, but involves the inhibition of keyenzymes in folate metabolism and purine biosynthesis
de novo, both of these are required for DNA
synthesis (77). This inhibition provokes folate
depletion and enhanced adenosine release, leading
to apoptosis (78) and/or interaction with adenosine
receptors, resulting in an anti-inflammatory effect
(79). Cell entry of MTX requires a specific
transporter, the reduced folate carrier (RFC), which
normally internalizes natural folates (80 – 82). Once
taken up intracellularly, MTX is complexed to
polyglutamate forms, which prevents its leakage
from the cells (83, 84). In cancer patients, the most
common mechanisms of acquired resistance to MTX
include impaired RFC transport, elevated levels of
dihydrofolate reductase (DHFR), and/or impaired
polyglutamylation (85 – 87).Originally, the MDR transporter Pgp was pro-
posed to play a role in MTX efflux in a T-cell line
that displayed resistance to supra-pharmacological
concentrations of MTX (88). However, the dominant
mechanism of MTX resistance in this cell line was
defective RFC- mediated uptake of MTX, rather
than Pgp-mediated efflux. Consistent with the notion
that Pgp preferentially transports neutral hydrophic
compounds, rather than a divalent anion like MTX,
there is currently broad consensus that Pgp is not a
major player in MTX efflux (16, 25, 55, 89). Instead
of Pgp, there is compelling evidence that the MDR
transporters MRP1-4 and ABCG2 have the capacity
to extrude non-polyglutamated MTX (90 – 99). Of
note is that ABCG2 appears to be capable of
transporting di- and tri-glutamate forms of MTX
(58).Transport kinetic data from these studies showed
that MTX efflux via MRP1-4 and BCRP is a low
affinity/high capacity process, in contrast to MTX
Table 1. Overview of selected ABC transporters/MDR proteins with known involvement of drug resistance (to anti-cancer drugs).
ABCcode
Commonname
Chromosomelocalization
Size(AA)
Normal tissuedistribution
Immumologicalcells
Preferredsubstrates Refs
ABCB1 P-gp,MDR1
7q21 1279 Many tissues T cells (CD4, CD8)DCs, monocytes
Neutral hydrophobiccompounds
(18, 25, 68, 71,125, 65, 69, 73)
ABCC1 MRP1 16p13.1 1531 Ubiquitous T cells (CD56, CD8,CD4), CD34z, DC, Mw
Anionic conjugates(glutathione, glucoronide)
(25, 38, 61, 70,204, 73, 205)
ABCC2 MRP2 10q24 1545 Liver, intestine,kidney
Anionic conjugates,glutathione
(19, 39)
ABCC3 MRP3 17q21.3 1527 Liver Anionic conjugates (25, 91)ABCC4 MRP4 13q32 1325 Many tissues
(kidney)Nucleoside analogues (39, 41, 93, 113)
ABCC5 MRP5 3q27 1437 Many tissues Nucleoside analogues (39, 43, 113)ABCC6 MRP6 16p13.1 1503 Liver, kidney Glutathione conjugates,
small peptides(45, 206)
ABCC10 MRP7 6p21 1513 Low in alltissues
? (46, 207)
ABCC11 MRP8 16q12.1 1382 Low in alltissues
Fluoropyrimidines (47, 48)
ABCG2 BCRP 4q22 655 Liver, breast,placenta
CD34z (side population) Amphipatic drugs (21, 51, 53, 74)
AA~Number of amino acids, Mw~macrophages. For timely updates on properties of ABC transporters, see http://www.nutrigene.4t.com/humanabc.htm
MDR and rheumatoid arthritis 327
www.scandjrheumatol.dk
Scan
d J
Rhe
umat
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y Q
UT
Que
ensl
and
Uni
vers
ity o
f T
ech
on 1
1/01
/14
For
pers
onal
use
onl
y.

influx via RFC, which is a high affinity/low capacity
process (80, 82). Apparently, a dynamic balance
exists between the influx and efflux routes in con-
trolling intracellular levels of MTX. In this respect,
MTX exposure time, which allows conversion to
non-effluxable polyglutamated forms, can also be
a determining factor in the efficacy of MTX. This is
illustrated in Table 3, where the efficacy (anti-
proliferative effect) of MTX was evaluated against
Pgp-, MRP1- and BCRP-over-expressing cells com-
pared with controls after either a 72 h continuous
exposure to MTX (permissive for polyglutamyla-
tion), or a short-term (4 h) exposure to MTX
(incomplete polyglutamylation) followed by a 68 h
drug-free period. Following a 4 h exposure to MTX,
only cells over-expressing MRP1 and BCRP dis-
played a high level of resistance to MTX, indicating
that MRP1 and BCRP, but not Pgp, were able to
extrude unpolyglutamated MTX, resulting in lower
intracellular levels and diminished efficacy. Follow-
ing a 72 h exposure to MTX, low levels of resistance
were still noted for the MRP1 and BCRP1, but not
for the Pgp over-expressing cells.
Finally, it is of importance to note that MRP1-3
can also mediate the efflux of natural folates (100).In this respect, diminished expression of MRP1 may
lead to increased intracellular folate pools, which can
antagonize MTX activity (101).
Sulphasalazine
Sulphasalazine (SSZ) is a potent DMARD used as
a single agent and in combination regimens with
MTX, hydroxychloroquine and prednisone (8, 9).
SSZ is an organic anion that, based on structure –
activity relationships, could be a potential substratefor members of the ABCC family of MDR
transporters. In fact, a role for MRP1 in SSZ
transport was indicated from studies by Lightfoot
(102): however, studies from our laboratory (unpub-
lished) did not demonstrate resistance to SSZ in
MRP1-overexpressing cells. This may be caused by
Table 2. Overview of selected ABC transporters/MDR proteins with known physiologic and pharmacologic substrates/inhibitors(DMARDs and other anti-inflammatory drugs).
ABCcode
Commonname
Pharmacologicsubstrate (DMARD/anti-inflammatory drug) Physiologic substrate Inhibitor(s) Refs
ABCB1 P-gp,MDR1
Chloroquine, CSA Steroids, lipids, PAF Verapamil, PSC833 (18, 25, 123, 126, 127)
Prednisone,dexamethasone
LY335979, CsA (106, 120, 190)
ABCC1 MRP1 MTX, Chloroquine LTC4, LTB4, LTE4 MK571, probenecid, (25, 90, 108, 128, 165)Folates NSAIDs, flavonoids, CHQ (158, 183, 208)
ABCC2 MRP2 MTX Bile salts, LTC4, steroids Sulphinpyrazone (25, 90)ABCC3 MRP3 MTX Bile salts, LTC4, steroids Probenecid (91, 92, 94, 95)ABCC4 MRP4 MTX, Azathioprine
(6-MP)LTE4, cAMP, cGMP Probenecid, NSAIDs (93, 113, 114, 135, 136)
ABCC5 MRP5 Azathioprine (6-MP) cGMP, cAMP Nitroprusside (43, 113, 114, 134)ABCC6 MRP6 LTC4 (45)ABCC10 MRP7 LTC4, steroids CSA (46)ABCC11 MRP8 cAMP/cGMP (48)ABCG2 BCRP MTX, SSZ (?) Steroids, chlorophyll
metabolitesGF120918, Ko143 (58, 59, 99, 167, 168)
Fumitremorgin C (20, 26, 185, 186, 188, 209)
E217bG~17b-oestradiol glucuronide.
Table 3. Role of MDR proteins in MTX resistance.
Exposure time MTX
Resistance Factor(#)
ABCB1(##) (Pgp) ABCC1(###) (MRP1) ABCG2(####) (BCRP)
72 h 0.8 1.9 1.74 h (z68 h drug free) 1.1 w80 28
(#)Resistance factor is defined as the ratio of IC50 concentration (MTX concentration resulting in 50% inhibition of cell proliferation) of cellswith over-expression of the indicated ABC transporter versus control cells without over-expression of the ABC transporter.(##)ABCB1/Pgp-overexpressing cells were vinblastin-resistant CEM cells (106).(###)ABCC1/MRP1-overexpressing cells were MRP1 transfected 2008 cells (90).(####)ABCG2/BCRP-overexpressing cells were mitoxantrone-resistant MCF-7/MR cells (50).
328 G Jansen et al
www.scandjrheumatol.dk
Scan
d J
Rhe
umat
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y Q
UT
Que
ensl
and
Uni
vers
ity o
f T
ech
on 1
1/01
/14
For
pers
onal
use
onl
y.

the fact that SSZ is also an inhibitior of glutathione-
S-transferase (103), the enzyme responsible for
glutathione conjugation prior to MRP1 transport.Recent studies by our laboratory have shown that
acquired resistance to SSZ was accompanied by
induction of ABCG2 (see below).
(Hydroxy)chloroquine
The antimalarial DMARD (hydroxy)chloroquine
has relatively little activity as a single agent, but is
used in combinations with other DMARDs (6, 104,105). Chloroquine has been identified as a substrate,
as well as an inhibitor of Pgp and MRP1 (106 – 108).
Its inhibitory activity suggests its use as a chemo-
sensitizer, as it elevates the intracellular concentra-
tion and efficacy of therapeutic drugs that are
substrates for these MDR proteins (see below).
Leflunomide
Leflunomide is a DMARD with immunosuppressiveactivity due to inhibition of dihydroorotate dehy-
drogenase (DHODH), a key enzyme in pyrimidine
biosynthesis de novo (109, 110). To date, it is
unknown whether leflunomide, or its active meta-
bolite A771726, is a substrate for MDR proteins, or
whether over-expression of specific MDR proteins
confers resistance to leflunomide. Acquired resis-
tance to leflunomide does, however, seem to beassociated with increased activity and/or structural
alterations of the target enzyme DHODH (111).
Azathioprine
Azathioprine is a prodrug of 6-mercaptopurine (6-
MP) and has an established place in RA treatment
(112). 6-MP, along with 6-thioguanine and other
nucleoside analogues, proved to be a potentialsubstrate for MRP4/ABCC4 and MRP5/ABCC5
(41, 43, 113 – 115). Over-expression of MRP4 or
MRP5 is associated with resistance to this class of
compounds. As for MTX, where polyglutamylation
abolished efflux, metabolism of 6-MP to its diphos-
phate and triphosphate metabolites diminished
substrate affinity for MRP4 and MRP5.
Auranofin/aurothiomalate
Although gold preparations belong to the first
generation of DMARDs, they are still prescribed
in some cases of RA. Glennas and colleagues (116,
117) are the only investigators who have studied
mechanisms of acquired resistance to aurothioma-
late. They observed the onset of resistance, but
an underlying mechanism was not revealed. No
evidence for an increase in methallothionein wasobserved (118). Evidence has been presented that
MRP1 and MRP2 can transport glutathione con-
jugates of heavy metal ions (arsenite, cis-platin) (19)
but whether this also includes gold preparationsremains to be established.
Cyclosporin A
In RA treatment, cyclosporin A (CSA) displayedactivity in combination with MTX (119). CSA is well
recognized for its inhibitory interaction with P-gp
(35, 120) (see also below). Data on mechanisms of
acquired resistance to CSA are limited, but surpris-
ingly do not seem to involve up-regulation of Pgp
expression (121, 122).
Glucocorticoids
Glucocorticoids (GC) display DMARD activity.
Given their hydrophobic nature, GCs like pred-
nisone and dexamethasone were identified as sub-strates for Pgp (18, 123). Over-expression of Pgp
reduces the cellular uptake of dexamethasone, which
can be normalized by blocking of Pgp (124).
Altogether, there is ample evidence that several
DMARDs can be substrates for MDR proteins. To
what extent specific MDR proteins actually con-
tribute to clinical DMARD resistance requires
further investigations.
Physiological role of MDR proteins
MDR proteins have received most attention for their
capacity to extrude therapeutic drugs from cells.Less well understood is the putative physiological
role of MDR proteins in transporting signalling
molecules, peptides or small proteins that are
involved in immunological/inflammatory processes
(Table 2). Pgp is expressed on CD4z T cells and
CD14z DCs, where it may mediate interferon-c(IFN-c) and interleukin-12 (IL-12) release (125),
and DC migration to draining lymph nodes (68).Whether these processes involve the Pgp-mediated
transport of lipid molecules, such as platelet-
activating factor (PAF), is the subject of ongoing
studies (126, 127).
An established substrate for MRP1 and other
members of the MRP family is leucotriene C4
(LTC4) (25, 128). Leucotrienes are biologically
active metabolites derived from arachidonic acidthat can activate inflammatory processes by interac-
tion with specific cysteinyl leucotriene receptors,
which are coupled to G proteins and downstream
signalling pathways (129, 130). LTC4 is a glu-
tathione conjugate of LTA4, which explains its
substrate affinity for MRP1 as this protein prefer-
entially transports glutathion conjugates. As for Pgp,
Robbiani (72) has demonstrated an important rolefor MRP1 and LTC4 in antigen-presenting DC
MDR and rheumatoid arthritis 329
www.scandjrheumatol.dk
Scan
d J
Rhe
umat
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y Q
UT
Que
ensl
and
Uni
vers
ity o
f T
ech
on 1
1/01
/14
For
pers
onal
use
onl
y.

migration to lymph nodes. MRP1-mediated trans-
port of LTC4 initiated binding to cysteinyl leuco-
triene receptors, which promoted CCL19-dependent
trafficking of DCs to the lymph nodes. MRP12/2
knock-out mice consistently demonstrated a mark-
edly impaired DC mobilization capacity, which
could be restored by exogenous LTC4. Of note,
studies by Wijnholds (131) had shown that MRP12/2
mice had a decreased inflammatory response (ear
oedema) after arachidonic acid, LTC4 or phorbol
ester treatment.
Steroids are another class of biologically active
compounds that are potential physiological sub-
strates for Pgp, MRPs and BCRP (25, 57, 59, 132,
133). A well-known high-affinity substrate for
these MDR transporters is 17b-oestradiol 17b-D-glucuronide (E217bG). Recently, cyclic adenosine
monophosphate (cAMP) and cyclic guanosine
monophosphate (cGMP) were found to be high-
affinity substrates for MRP4 and MRP5 (93, 114,
134, 135). Since cyclic nucleotides play essential
roles as activators of protein kinases and signalling
pathways, one may speculate that MRP4 and MRP5
may (indirectly) contribute to control these processes
by extrusion of cAMP and cGMP.
Finally, Reid (136) reported that MRP4 is a major
efflux transporter for prostaglandins (PG) such as
PGE1, PGE2 and PGA1. Interestingly, several non-
steroidal antiinflammatory drugs (NSAIDs), notably
indomethacin and indoprofen, proved to be potent
inhibitors of PG efflux via MRP4. This suggests
that, beyond inhibition of PG synthesis, part of the
anti-inflammatory effects of these NSAIDs may be
mediated via inhibition of MRP4.In an early report, Salmon and Dalton (137)
hypothesized that MDR proteins, such as P-gp,
could function as efflux transporters for pro-
inflammatory cytokines, such as TNFa. The anti-
inflammatory effect of a DMARD like CSA could
then be compatible with it being an inhibitor of Pgp.
In line with this hypothesis were studies by Drach
(138) and Raghu (139), demonstrating that inhibi-
tion of Pgp blocked the release of Il-2, IL4 and
IFNc. In contrast, however, Eisenbraum and Miller
(140) showed that T cells from Pgp2/2 mice had
unaltered release of IL2, IL4 and IFNc compared
with T cells from wild-type mice. Thus, a direct role
for Pgp in transport of cytokines with a molecular
weight of 10 – 25 kD is still controversial.
An alternative explanation for these observations
may be that Pgp effluxes compounds that control the
release of cytokines via other routes. A few reports
also suggested a role for MRP1 in the transmem-
brane transport of distinct cytokines, e.g. basic
fibroblast growth factor and IL-4 (141, 142).
Vellenga (143) reported a marked increase in the
release of IL-6 from activated monocytes following
inhibition of MRP1 by MK571. Again, as for Pgp,
conclusive evidence for MRP1-mediated efflux of
cytokines will require additional studies, including
competition studies of putative protein substrateswith established MRP1 substrates, such as LTC4.
Rather than for proteins, there is substantial
evidence that Pgp and MRP1 can efflux small
(cyclic) peptides (up to 10 mer) (144, 145).
ANLL (N-acetyl-leu-leu-norleucinal) is a tripep-
tide with Pgp and MRP1 substrate affinity (144,
145). It is an inhibitor of 26S-proteasome activity,
which plays a key role in the activation of nuclearfactor kB (NFkB), the nuclear transcription factor
that mediates the transcription of TNFa (146, 147).
Hence, cellular Pgp and MRP1 expression should be
considered as factors determining the efficacy of
targeting NFkB signaling by peptide-based 26S-
proteasome inhibitors.
In summary, beyond transport of therapeutic
drugs, some MDR proteins have the capacity totransport small signalling molecules/peptides. A
direct role in transporting higher molecular weight
cytokines is still under debate, although (in)direct
effects on cytokine release seem evident.
Regulation of MDR protein expression
Drug-induced up-regulation of MDR protein expres-
sion is well documented, however, mechanisms in
control of the transcriptional regulation of MDR
genes are still relatively poorly understood. There is
accumulating evidence that endogenous or exogen-
ous stress signals (e.g. oxidative stress, toxic drugs,
inflammation, cytokines and growth factors) play acentral role in the transcriptional regulation of Pgp
and MRP1 (148): for most other MDR proteins
this needs to be established. In this respect, several
downstream signal transduction pathways/proteins
(MAPK, JNK) and transcription factors (NFkB,and notably cAMP-responsive transcription factors
AP1 and SP1) have been implicated in the expression
of Pgp and MRP1 (149 – 151).NFkB binding sites were identified in the promo-
ter region of the Pgp gene (152, 153), and actually
appeared to control the regulation of Pgp gene
expression (154). The status/activity of p53, a key
regulatory gene in tumourogenesis, as well as chronic
inflammation (155), also appeared to be of impor-
tance in the regulation of MRP1 expression: wild
type p53 suppresses (156), while mutated p53enhances (157) MRP1 gene transcription. Finally,
a recent study by Assaraf (158) showed that cellular
folate depletion resulted in down-regulation of
MRP1 expression, tentatively as a physiological
response to prevent loss of intracellular folates.
A number of studies showed that cytokines can
have differential effects on MDR protein expression.
In fact, TNFa (159, 160) and IL-1b and IL-6 (148)down-regulated expression of Pgp, whereas IFN-c
330 G Jansen et al
www.scandjrheumatol.dk
Scan
d J
Rhe
umat
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y Q
UT
Que
ensl
and
Uni
vers
ity o
f T
ech
on 1
1/01
/14
For
pers
onal
use
onl
y.

can up-regulate Pgp expression (161). TNFa was
found to up-regulate expression of MRP1, but not
of LRP (162). Effects of cytokines/chemokines onexpression levels of MDR proteins other than Pgp
and MRP1 are currently unknown.
Cell differentiation and maturation are processes
that are associated with changes in MDR protein
expression. For example, an increase in Pgp and
MRP1 expression was noted during monocyte –DC
differentiation, and during DC maturation (71, 73).
Of note are also observations that MDR expressionmay alter as a function of ageing (163). In particular,
Pgp expression in CD4z and CD8z T cells increased
during ageing (164). Hence, clinical studies with RA
patients for the evaluation of MDR protein expres-
sion should include age-matched controls. Taken
together, these studies demonstrate that up/down-
regulated expression of MDR proteins can be either
drug-induced or a response to stress signals, cyto-kines and other inflammatory effects.
MDR protein induction by DMARDs
Induction of MDR proteins during development of
resistance to anti-cancer drugs is well established.With the exception of MTX, it is unknown whether
acquired resistance to DMARDs can also be asso-
ciated with the induction of specific MDR proteins.
In recent studies from our laboratory (165 – 168),
resistance was provoked in vitro in human T cells
(CEM cell line) to the DMARDs choloroquine
(CHQ) and sulphasalazine (SSZ). These two
DMARDs were selected as, in clinical practice, theypresent a relatively more rapid onset of diminished
efficacy than other DMARDs (10, 12, 13). Following
exposure of CEM to stepwise increasing concentra-
tions of CHQ or SSZ for 6 months, appreciable
levels of resistance to CHQ (3.5-fold) and SSZ (6.5-
fold) were noted. Analysis of MDR protein expres-
sion revealed a marked induction of MRP1 in
CHQ- resistant cells and BRCP in SSZ-resistantcells. CHQ-resistant cells retained sensitivity to most
other DMARDs (MTX, CSA, leflunomide and
SSZ), but were highly cross-resistant (w1000-fold)
to the glucocorticoid dexamethasone. In contrast,
SSZ-resistant T cells displayed markedly increased
(13-fold) sensitivity to dexamethasone compared to
parental cells. This may actually reflect the improved
efficacy of glucocorticoids (prednisone) in combina-tion with MTXzSSZ, compared to MTXzSSZ
alone for the treatment of RA patients (9). SSZ-
resistant cells showed unaltered sensitivity to CHQ
and CsA.
Interestingly, SSZ-resistant cells displayed a low
level of cross-resistance to MTX (v2-fold), which
markedly increased (w300-fold) when MTX sensi-
tivity was tested in the presence of the selectiveconcentrations of SSZ. This loss of MTX efficacy
appeared to be due an unexpected and hitherto
unknown inhibition by SSZ of the reduced folate
carrier, the cell membrane transporter for MTX,which abolished cell entry of MTX to confer resis-
tance (169). Finally, an altered release of cytokines
was observed in SSZ-resistant cells, as indicated by
a 3-fold higher secretion of TNFa compared with
control CEM cells.
Collectively, these in vitro results demonstrate that
the onset of DMARD resistance can be accompa-
nied by induction of specific MDR proteins, altereddrug sensitivity and increased TNFa release.
MDR proteins in RA patients under DMARD therapy
A large number of cancer therapy studies have
included an evaluation of MDR protein expression,notably Pgp, MRP1-5 and BCRP, as part of an
assessment of the efficacy of chemotherapeutic drugs
(36, 170 – 173). Other than in a pilot setting, similar
studies have not been carried out for RA patients to
assess the (long term) efficacy of various DMARDs.
In one of four studies published on the putative
role of DMARD resistance and MDR expression,
Maillefert (174) observed a significant increase in thepercentage of Pgp-positive peripheral blood lympho-
cytes in a group of RA patients (n~68, mean disease
duration 10.7 years) treated with prednisone (n~40,
10.7% Pgpz) compared with RA patients treated
with various DMARDs (n~25) and NSAIDs
(n~34), or controls (n~44) (3.3% Pgpz). Pgp
expression did not correlate with gender, disease
duration, past resistances to DMARDs, RF, or C-reactive protein (CRP). Consistent with prednisone
being a potential substrate for Pgp, these results may
suggest a role for prednisone in Pgp induction.
Yudoh (175) studied percentages of Pgp positive
cells in Th1 and Th2 subpopulations of CD4z T
cells of 14 RA patients and eight RA patients non-
responsive to DMARD therapy (sulphasalazine/
bucillamine). The percentage of Pgpz cells wassignificantly higher in Th1 cells of non-responders
versus responders, both at baseline (9.6% versus
3.3%) and two months after therapy (15.4% versus
3.3%). No differences in Pgp expression (2.2 – 2.6%)
were observed in Th2 cells of responders and non-
responders.
Llorente (176) analysed the percentage of periph-
eral blood lymphocytes for Pgp activity (based ondaunorubicin efflux) in 24 controls, eight therapy-
responsive RA patients and eight therapy non-
responsive RA patients. Therapy non-responsive
patients revealed a markedly higher percentage of
Pgp-active cells (34.4%) compared with therapy
responsive patients (3.9%) or controls (v1.5%).
Finally, in a prospective study, Maillefert (177)
analysed Pgp expression (by indirect immunofluor-escence) on CD4z and CD8z T cells, B cells and
MDR and rheumatoid arthritis 331
www.scandjrheumatol.dk
Scan
d J
Rhe
umat
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y Q
UT
Que
ensl
and
Uni
vers
ity o
f T
ech
on 1
1/01
/14
For
pers
onal
use
onl
y.

monocytes in a group of RA patients (n~24, mean
age 56 years, disease duration 5.9 years). Pgp
expression was analysed at baseline and 6 monthsafter therapy with one of the following DMARDs:
MTX, sulphasalazine, leflunomide or gold salts. At
this stage, 17 out of 19 evaluable patients displayed
a clinical response. Most significant differences at
baseline and after 6 months therapy were a decrease
in the percentage of Pgpz monocytes in responders:
16% versus 0%, respectively, suggesting that Pgp
expression on monocytes may be a marker ofresponse to DMARDs.
To date, only one study, by Jorgensen (178), has
reported on the expression of Pgp in synovial tissue
of RA patients. Immunohistochemical staining of
Pgp was observed in synovial membranes of five out
of 16 RA patients treated with various DMARDs.
Surprisingly, however, Pgp mRNA was identified
in only one out of these five patients, from whichthe investigators suggested they have identified an
atypical MDR phenotype, distinct from classical
Pgp, that was previously described in a T-cell line
selected for extremely high concentrations of MTX
(179).
In summary, studies on MDR expression on
clinical specimens of RA patients are currently
limited to the analysis of Pgp expression onperipheral blood lymphocytes. Data on expression
of other MDR proteins and in synovial cells are not
readily available.
MDR reversal agents/inhibitors
In cancer chemotherapy, blocking or modulating of
MDR activity has received considerable attention
as a possible strategy to overcome diminished drug
efficacy due to MDR-mediated drug efflux. Inhibi-
tors/modulators for various MDR proteins are now
available (Table 2), which confer in vitro reversal
of resistance to chemotherapeutic drugs transported
by the MDR proteins. Verapamil and CSA are firstgeneration inhibitors of Pgp activity (37, 180, 181).
PSC833 (120), a structural analogue of CSA devoid
of immunosuppressive effects, is a second generation
Pgp inhibitor currently undergoing clinical trials.
Inhibitors of MRP1 include probenecid, which
also inhibits MRP2-4 (25, 90, 93), and MK571, a
leucotriene D4 analogue (182). Interestingly, several
NSAIDs also appeared to be potent inhibitorsof MRP1 (e.g. sulindac and indomethacin) (183)
and MRP4 (e.g. indoprofen, indomethacin) (136).
Recently identified inhibitors of BCRP include
GF120918 (184, 185) and a fumitremorgine analogue
Ko143 (186). Based on in vitro potency to reverse
drug resistance, several MDR inhibitors are cur-
rently being evaluated in a (pre)clinical setting (29,
187, 188) for their ability to modulate clinical drugresistance.
Evaluating the outcome of the various MDR
reversal trials, van Zuylen (189) concluded that in
the cancer chemotherapeutic setting the clinicalbenefit of MDR modulators is rather modest. This
may be due to several reasons:
. Plasma levels of MDR modulators are insufficient
to achieve optimal reversal
. Pharmacokinetic drug interact with some MDR
modulators, requiring dose reductions
. A multifactorial basis for drug resistance rendersreversal of the resistant phenotype incomplete.
Obviously, these issues should be resolved in order
to reveal a greater potential of the MDR inhibitors/
modulators, but recent studies with third generation
MDR reversal agents look promising in this respect
(190). Another novel clinical application of MDR
inhibitors may be in improving oral drug treatment
by temporary inhibition of drug transporters in thegastrointestinal tract (191).
From a rheumatology perspective, lessons can to
be learned for the exploitation of MDR reversal
agents as possible modulators of DMARD resis-
tance. First, in order to utilize the appropriate MDR
modulator, the MDR protein(s) contributing to
efflux of various DMARDs should be identified.
Second, different endpoints may reflect the efficacyof MDR modulators in cancer treatment (restora-
tion of antiproliferative effects) and RA treat-
ment (restoration of anti-inflammatory effects of
DMARDs). These differential effects should not
necessarily be achieved at identical concentrations
of MDR modulators. Ideally, if modulation of
DMARD resistance is effective at lower concentra-
tions of MDR modulators than for anticancer drugs,this would improve the therapeutic window of the
modulators. Third, it is apparent that MDR proteins
have physiological functions in transporting inflam-
matory mediators, e.g. LTC4 via MRP1. Hence,
blocking of MDR proteins may turn out to be a
double-edged sword, as it may reduce the anti-
inflammatory response capacity — as in MRP1 2/2
knock out mice (131).In conclusion, although MDR modulators are
available, their full (in vitro) potential still needs
to be translated into a markedly enhanced clinical
benefit of cancer chemotherapeutic drugs. Explora-
tion of MDR modulators to improve the efficacy of
DMARDs may warrant further attention.
Perspectives
Drug resistance, inherent or acquired, is generally
accepted as a common cause of treatment failure in
the field of infectious diseases and cancer, but has
received relatively little attention in clinical rheuma-tology in relation to DMARD resistance. Expanding
332 G Jansen et al
www.scandjrheumatol.dk
Scan
d J
Rhe
umat
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y Q
UT
Que
ensl
and
Uni
vers
ity o
f T
ech
on 1
1/01
/14
For
pers
onal
use
onl
y.

knowledge on drug resistance has revealed that there
is considerable overlap in molecular mechanisms of
drugs, regardless of whether these drugs have anti-
infectious, anti-cancer or anti-rheumatic effects.
Although drug resistance can be a multifactorial
event, one common mode of cellular drug resistance
is enhanced efflux by several members of the ABC
transporter family, giving rise to the MDR pheno-
type. There is a growing body of evidence that
various ABC transporters are expressed in cell types
that play a role in inflammatory processes, and
which are targets for DMARD therapy. In this
respect it will be important that future studies
categorize in clinical specimens from RA patients
(peripheral blood cells, synovial cells and tissue) the
expression of those ABC transporters that have an
established role in drug resistance. These studies are
now achievable, as the technology required to
analyse mRNA levels of all 48 ABC transporters
is available (192), just as flow cytometric assays are
available to analyse the functional activity of Pgp,
MRP1 and BCRP (193). However, for an assess-
ment of the various MDR transporters in clinical
specimens containing many different cell types,
immunohistochemistry remains the mainstay of the
analytical platform (40, 53).
Other important issues requiring further attention
are:
. The assessing of whether up-regulation of MDR
proteins in RA patients is: a DMARD-induced
phenomenon, with possible variability dependent
on the DMARD used, and/or a disease-related
phenomenon. In other words, is MDR expression/
functional activity a reflection of disease activity
related to the physiological function of some MDR
proteins in transporting inflammatory mediators.
. Patient variability in response to DMARDs may
be envisioned given the identification of a large
variety of Pgp, MRP1 and BCRP polymorphisms
(194 – 200). The functional implications of all these
polymorphisms need to be established. Further-
more, several distinct mutations in the Pgp, MRP1
and BCRP gene have been reported that may
affect transport of MTX (58, 99, 201, 202).
. Studies on drug resistance will have broader
implications in inflammatory diseases rather than
in RA alone, as expression of Pgp was also identified
in blood cells of other autoimmune diseases, e.g.
systemic lupus erythematosus (SLE) (203).
. Along with clinical studies, the use of in vitro
models with acquired resistance to DMARDs
resistance can provide further insight into the
molecular mechanisms of resistance to various
DMARDs, the involvement of MDR proteins
and/or other mechanisms of resistance, and
the design of strategies to overcome DMARD
resistance.
Answering these questions will undoubtedly con-
tribute to better treatment options with DMARDs
for patients with RA or other chronic inflammatorydiseases.
Acknowledgements
Drs Ruud Oerlemans, Willem Lems, Joost van der Heijden and
George Scheffer are acknowledged for critical reading of the
manuscript and helpful suggestions.
This study was supported by a grant (NRF 03I-40) from the
Dutch League against Rheumatism (Nationaal Reumafonds).
Abbreviations
MDR1; Multi Drug Resistance, Pgp; P-glycoprotein, MRP;
Multidrug Resistance associated Protein, MTX; methotrexate,
CsA; cyclosporin A, BCRP; Breast Cancer Resistance Proteins,
ABC; ATP Binding Cassette, GC; Glucocorticoid, cAMP; Cyclic
Adenosine Mono-Phosphate, NFkB; Nuclear Factor kB, DHFR;
Dihydrofolate reductase, FPGS; Folylpolyglutamate synthetase,
RFC; Reduced Folate Carrier, RA; Rheumatoid Arthritis.
References
Selected key references are shown below. The online version of
this review includes a complete reference list. Online access: www.
scandjrheumatol.dk
..1. Feldmann M, Brennan FM, Maini RN. Rheumatoid arthritis.
Cell 1996;85:307 – 10.
2. Smolen JS, Steiner G. Therapeutic strategies for rheumatoid
arthritis. Nat Rev Drug Discov 2003;2:473 – 88.
6. Fathy N, Furst DE. Combination therapy for autoimmune
diseases: the rheumatoid arthritis model. Springer Semin
Immunopathol 2001;23:5 – 26.
8. O’Dell JR, Leff R, Paulsen G, Haire C, Mallek J, Eckhoff PJ,
et al. Treatment of rheumatoid arthritis with methotrexate
and hydroxychloroquine, methotrexate and sulfasalazine, or a
combination of the three medications: results of a two-year,
randomized, double-blind, placebo-controlled trial. Arthritis
Rheum 2002;46:1164 – 70.
9. Landewe RB, Boers M, Verhoeven AC, Westhovens R, van de
Laar MA, Markusse HM, et al. COBRA combination therapy
in patients with early rheumatoid arthritis: long-term
structural benefits of a brief intervention. Arthritis Rheum
2002;46:347 – 56.
10. Wolfe F. The epidemiology of drug treatment failure in
rheumatoid arthritis. Baillieres Clin Rheumatol 1995;9:619 –
32.
12. Maetzel A, Wong A, Strand V, Tugwell P, Wells G,
Bombardier C. Meta-analysis of treatment termination
rates among rheumatoid arthritis patients receiving disease-
modifying anti-rheumatic drugs. Rheumatology (Oxford)
2000;39:975 – 81.
14. Morgan C, Lunt M, Brightwell H, Bradburn P, Fallow W,
Lay M, et al. Contribution of patient related differences to
multidrug resistance in rheumatoid arthritis. Ann Rheum Dis
2003;62:15 – 9.
16. Endicott JA, Ling V. The biochemistry of P-glycoprotein-
mediated multidrug resistance. Annu Rev Biochem 1989;
58:137 – 71.
20. Gottesman MM, Fojo T, Bates SE. Multidrug resistance in
cancer: role of ATP-dependent transporters. Nat Rev Cancer
2002;2:48 – 58.
21. Litman T, Druley TE, Stein WD, Bates SE. From MDR to
MXR: new understanding of multidrug resistance systems,
MDR and rheumatoid arthritis 333
www.scandjrheumatol.dk
Scan
d J
Rhe
umat
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y Q
UT
Que
ensl
and
Uni
vers
ity o
f T
ech
on 1
1/01
/14
For
pers
onal
use
onl
y.

their properties and clinical significance. Cell Mol Life Sci
2001;58:931 – 59.
22. Hayes JD, Wolf CR. Molecular mechanisms of drug
resistance. Biochem J 1990;272:281 – 95.
25. Borst P, Oude Elferink RP. Mammalian ABC transporters in
health and disease. Annu Rev Biochem 2002;71:537 – 92.
27. Dean M, Rzhetsky A, Allikmets R. The human ATP-binding
cassette (ABC) transporter superfamily. Genome Res
2001;11:1156 – 66.
31. Leslie EM, Deeley RG, Cole SP. Toxicological relevance of
the multidrug resistance protein 1, MRP1 (ABCC1) and
related transporters. Toxicology 2001;167:3 – 23.
33. Lehnert M. Clinical multidrug resistance in cancer: a multi-
factorial problem. Eur J Cancer 1996;32A:912 – 20.
37. Germann UA. P-glycoprotein — a mediator of multidrug
resistance in tumour cells. Eur J Cancer 1996;32A:927 – 44.
38. Cole SP, Bhardwaj G, Gerlach JH, Mackie JE, Grant CE,
Almquist KC, et al. Over-expression of a transporter gene in a
multidrug-resistant human lung cancer cell line. Science
1992;258:1650 – 4.
40. Scheffer GL, Kool M, Heijn M, de Haas M, Pijnenborg AC,
Wijnholds J, et al. Specific detection of multidrug resistance
proteins MRP1, MRP2, MRP3, MRP5, and MDR3 P-
glycoprotein with a panel of monoclonal antibodies. Cancer
Res 2000;60:5269 – 77.
41. Schuetz JD, Connelly MC, Sun D, Paibir SG, Flynn PM,
Srinivas RV, et al. MRP4: A previously unidentified factor in
resistance to nucleoside-based antiviral drugs. Nat Med
1999;5:1048 – 51.
43. Wijnholds J, Mol CA, van Deemter L, de Haas M, Scheffer
GL, Baas F, et al. Multidrug-resistance protein 5 is a
multispecific organic anion transporter able to transport
nucleotide analogs. Proc Natl Acad Sci USA 2000;97:7476 –
81.
44. Kool M, van der Linden M, de Haas M, Baas F, Borst P.
Expression of human MRP6, a homologue of the multidrug
resistance protein gene MRP1, in tissues and cancer cells.
Cancer Res 1999;59:175 – 82.
46. Chen ZS, Hopper-Borge E, Belinsky MG, Shchaveleva I,
Kotova E, Kruh GD. Characterization of the transport
properties of human multidrug resistance protein 7 (MRP7,
ABCC10). Mol Pharmacol 2003;63:351 – 8.
47. Bera TK, Lee S, Salvatore G, Lee B, Pastan I. MRP8, a new
member of ABC transporter superfamily, identified by EST
database mining and gene prediction program, is highly
expressed in breast cancer. Mol Med 2001;7:509 – 16.
50. Taylor CW, Dalton WS, Parrish PR, Gleason MC, Bellamy
WT, Thompson FH, et al. Different mechanisms of decreased
drug accumulation in doxorubicin and mitoxantrone resistant
variants of the MCF7 human breast cancer cell line. Br J
Cancer 1991;63:923 – 9.
51. Doyle LA, Yang W, Abruzzo LV, Krogmann T, Gao Y, Rishi
AK, Ross DD. A multidrug resistance transporter from
human MCF-7 breast cancer cells. Proc Natl Acad Sci USA
1998;95:15665 – 70.
53. Maliepaard M, Scheffer GL, Faneyte IF, van Gastelen MA,
Pijnenborg AC, Schinkel AH, et al. Subcellular localization
and distribution of the breast cancer resistance protein
transporter in normal human tissues. Cancer Res
2001;61:3458 – 64.
55. Borgnia MJ, Eytan GD, Assaraf YG. Competition of
hydrophobic peptides, cytotoxic drugs, and chemosensitizers
on a common P-glycoprotein pharmacophore as revealed by
its ATPase activity. J Biol Chem 1996;271:3163 – 71.
58. Chen ZS, Robey RW, Belinsky MG, Shchaveleva I, Ren XQ,
Sugimoto Y, et al. Transport of methotrexate, methotrexate
polyglutamates, and 17beta-estradiol 17-(beta-D-glucuronide)
by ABCG2: effects of acquired mutations at R482 on
methotrexate transport. Cancer Res 2003;63:4048 – 54.
60. Jedlitschky G, Leier I, Buchholz U, Barnouin K, Kurz G,
Keppler D. Transport of glutathione, glucuronate, and sulfate
conjugates by the MRP gene-encoded conjugate export pump.
Cancer Res 1996;56:988 – 94.
61. Flens MJ, Zaman GJ, van der Valk P, Izquierdo MA,
Schroeijers AB, Scheffer GL, et al. Tissue distribution of the
multidrug resistance protein. Am J Pathol 1996;148:1237 – 47.
66. Legrand O, Perrot JY, Tang R, Simonin G, Gurbuxani S,
Zittoun R, Marie JP. Expression of the multidrug resistance-
associated protein (MRP) mRNA and protein in normal
peripheral blood and bone marrow haemopoietic cells. Br J
Haematol 1996;94:23 – 33.
68. Randolph GJ, Beaulieu S, Pope M, Sugawara I, Hoffman L,
Steinman RM, Muller WA. A physiologic function for
p-glycoprotein (MDR-1) during the migration of dendritic
cells from skin via afferent lymphatic vessels. Proc Natl Acad
Sci USA 1998;95:6924 – 9.
70. Laupeze B, Amiot L, Payen L, Drenou B, Grosset JM, Lehne
G, et al. Multidrug resistance protein (MRP) activity in
normal mature leukocytes and CD34-positive hematopoietic
cells from peripheral blood. Life Sci 2001;68:1323 – 31.
71. Laupeze B, Amiot L, Bertho N, Grosset JM, Lehne G,
Fauchet R, Fardel O. Differential expression of the efflux
pumps P-glycoprotein and multidrug resistance-associated
protein in human monocyte-derived dendritic cells. Hum
Immunol 2001;62:1073 – 80.
72. Robbiani DF, Finch RA, Jager D, Muller WA, Sartorelli AC,
Randolph GJ. The leukotriene C(4) transporter MRP1
regulates CCL19 (MIP-3beta, ELC)- dependent mobilization
of dendritic cells to lymph nodes. Cell 2000;103:757 – 68.
73. Schroeijers AB, Reurs AW, Scheffer GL, Stam AG, de
Jong MC, Rustemeyer T, et al. Up-regulation of drug
resistance-related vaults during dendritic cell development.
J Immunol 2002;168:1572 – 8.
74. Zhou S, Schuetz JD, Bunting KD, Colapietro AM, Sampath
J, Morris JJ, et al. The ABC transporter Bcrp1/ABCG2 is
expressed in a wide variety of stem cells and is a molecular
determinant of the side-population phenotype. Nat Med
2001;7:1028 – 34.
77. Cutolo M, Sulli A, Pizzorni C, Seriolo B, Straub RH. Anti-
inflammatory mechanisms of methotrexate in rheumatoid
arthritis. Ann Rheum Dis 2001;60:729 – 35.
78. Genestier L, Paillot R, Quemeneur L, Izeradjene K, Revillard
JP. Mechanisms of action of methotrexate. Immunopharma-
cology 2000;47:247 – 57.
79. Cronstein BN. Molecular therapeutics. Methotrexate and its
mechanism of action. Arthritis Rheum 1996;39:1951 – 60.
80. Jansen G. Receptor- and carrier-mediated transport systems
for folates and antifolates. In: Jackman AL, editor. Anti-
cancer drug development guide: antifolate drugs in cancer
therapy. Totowa, NJ: Humana Press Inc; 1999. p. 293 – 321.
84. Rots MG, Pieters R, Peters GJ, Noordhuis P, Van Zantwijk
CH, Kaspers GJ, et al. Role of folylpolyglutamate synthetase
and folylpolyglutamate hydrolase in methotrexate accumula-
tion and polyglutamylation in childhood leukemia. Blood
1999;93:1677 – 83.
85. Gorlick R, Goker E, Trippett T, Waltham M, Banerjee D,
Bertino JR. Intrinsic and acquired resistance to methotrexate
in acute leukemia. N Engl J Med 1996;335:1041 – 8.
87. Ranganathan P, Eisen S, Yokoyama WM, McLeod HL. Will
pharmacogenetics allow better prediction of methotrexate
toxicity and efficacy in patients with rheumatoid arthritis?
Ann Rheum Dis 2003;62:4 – 9.
90. Hooijberg JH, Broxterman HJ, Kool M, Assaraf YG, Peters
GJ, Noordhuis P, et al. Antifolate resistance mediated by the
multidrug resistance proteins MRP1 and MRP2. Cancer Res
1999;59:2532 – 5.
91. Kool M, van der Linden M, de Haas M, Scheffer GL, de Vree
JM, Smith AJ, et al. MRP3, an organic anion transporter able
334 G Jansen et al
www.scandjrheumatol.dk
Scan
d J
Rhe
umat
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y Q
UT
Que
ensl
and
Uni
vers
ity o
f T
ech
on 1
1/01
/14
For
pers
onal
use
onl
y.

to transport anti-cancer drugs. Proc Natl Acad Sci USA
1999;96:6914 – 9.
97. Chen ZS, Lee K, Walther S, Raftogianis RB, Kuwano M,
Zeng H, Kruh GD. Analysis of methotrexate and folate
transport by multidrug resistance protein 4 (ABCC4): MRP4
is a component of the methotrexate efflux system. Cancer Res
2002;62:3144 – 50.
99. Volk EL, Farley KM, Wu Y, Li F, Robey RW, Schneider E.
Over-expression of wild-type breast cancer resistance protein
mediates methotrexate resistance. Cancer Res 2002;62:5035– 40.
100. Hooijberg JH, Peters GJ, Assaraf YG, Kathmann I, Priest
DG, Bunni MA, et al. The role of multidrug resistance
proteins MRP1, MRP2 and MRP3 in cellular folate
homeostasis. Biochem Pharmacol 2003;65:765 – 71.
101. Stark M, Rothem L, Jansen G, Scheffer GL, Goldman ID,
Assaraf YG. Antifolate resistance associated with loss of
MRP1 expression and function in Chinese hamster ovary
cells with markedly impaired export of folate and cholate.
Mol Pharmacol 2003;64:220 – 7.
102. Liang E, Proudfoot J, Yazdanian M. Mechanisms of
transport and structure – permeability relationship of sulfa-
salazine and its analogs in Caco-2 cell monolayers. Pharm
Res 2000;17:1168 – 74.
105. Goekoop YP, Allaart CF, Breedveld FC, Dijkmans BA.
Combination therapy in rheumatoid arthritis. Curr Opin
Rheumatol 2001;13:177 – 83.
107. Vezmar M, Georges E. Direct binding of chloroquine to the
multidrug resistance protein (MRP): possible role for MRP
in chloroquine drug transport and resistance in tumor cells.
Biochem Pharmacol 1998;56:733 – 42.
110. Breedveld FC, Dayer JM. Leflunomide: mode of action in
the treatment of rheumatoid arthritis. Ann Rheum Dis
2000;59:841 – 9.
112. Seidman EG, Furst DE. Pharmacogenetics for the indivi-
dualization of treatment of rheumatic disorders using
azathioprine. J Rheumatol 2002;29:2484 – 7.
113. Wielinga PR, Reid G, Challa EE, van der Heijden I, van
Deemter L, de Haas M, et al. Thiopurine metabolism and
identification of the thiopurine metabolites transported by
MRP4 and MRP5 overexpressed in human embryonic
kidney cells. Mol Pharmacol 2002;62:1321 – 31.
117. Glennas A, Rugstad HE. Cultured human cells can acquire
resistance to the antiproliferative effect of sodium aurothio-
malate. Ann Rheum Dis 1986;45:389 – 95.
119. Tugwell P, Pincus T, Yocum D, Stein M, Gluck O, Kraag G,
et al. Combination therapy with cyclosporine and metho-
trexate in severe rheumatoid arthritis [The Methotrexate –
Cyclosporine Combination Study Group]. N Engl J Med
1995;333:137 – 41.
121. Wright KA, Twentyman PR. Derivation and characterisa-
tion of a mouse tumour cell line with acquired resistance to
cyclosporin A. Eur J Cancer 1993;29A:389 – 94.
124. Maillefert JF, Duchamp O, Solary E, Genne P, Tavernier C.
Effects of cyclosporin at various concentrations on dex-
amethasone intracellular uptake in multidrug resistant cells.
Ann Rheum Dis 2000;59:146 – 8.
125. Frank M, Denton M, Alexander S, Khoury S, Sayegh M,
Briscoe D. Specific MDR1 P-glycoprotein blockade inhibits
human alloimmune T cell activation in vitro. J Immunol
2001;166:2451 – 9.
127. Randolph GJ. Dendritic cell migration to lymph nodes:
cytokines, chemokines, and lipid mediators. Semin Immunol
2001;13:267 – 74.
131. Wijnholds J, Evers R, van Leusden MR, Mol CA, Zaman
GJ, Mayer U, et al. Increased sensitivity to anticancer drugs
and decreased inflammatory response in mice lacking
the multidrug resistance-associated protein. Nat Med
1997;3:1275 – 9.
134. Jedlitschky G, Burchell B, Keppler D. The multidrug
resistance protein 5 functions as an ATP-dependent export
pump for cyclic nucleotides. J Biol Chem 2000;275:30069 –
74.
136. Reid G, Wielinga P, Zelcer N, van der Heijden I, Kuil A,
de Haas M, et al. The human multidrug resistance protein
MRP4 functions as a prostaglandin efflux transporter and is
inhibited by nonsteroidal antiinflammatory drugs. Proc Natl
Acad Sci USA 2003;100:9244 – 9.
137. Salmon SE, Dalton WS. Relevance of multidrug resistance to
rheumatoid arthritis: development of a new therapeutic
hypothesis. J Rheumatol Suppl 1996;44:97 – 101.
138. Drach J, Gsur A, Hamilton G, Zhao S, Angerler J, Fiegl M,
et al. Involvement of P-glycoprotein in the transmembrane
transport of interleukin-2 (IL-2), IL-4, and interferon-gamma
in normal human T lymphocytes. Blood 1996;88:1747 – 54.
140. Eisenbraun MD, Miller RA. mdr1a-encoded P-glycoprotein
is not required for peripheral T cell proliferation, cytokine
release, or cytotoxic effector function in mice. J Immunol
1999;163:2621 – 7.
143. Vellenga E, Tuyt L, Wierenga BJ, Muller M, Dokter W.
Interleukin-6 production by activated human monocytic cells
is enhanced by MK-571, a specific inhibitor of the multi-drug
resistance protein-1. Br J Pharmacol 1999;127:441 – 8.
144. de Jong MC, Slootstra JW, Scheffer GL, Schroeijers AB,
Puijk WC, Dinkelberg R, et al. Peptide transport by the
multidrug resistance protein MRP1. Cancer Res 2001;
61:2552 – 7.
146. Lee DH, Goldberg AL. Proteasome inhibitors: valuable new
tools for cell biologists. Trends Cell Biol 1998;8:397 – 403.
148. Sukhai M, Piquette-Miller M. Regulation of the multidrug
resistance genes by stress signals. J Pharm Pharm Sci
2000;3:268 – 80.
151. Rohlff C, Glazer RI. Regulation of multidrug resistance
through the cAMP and EGF signalling pathways. Cell Signal
1995;7:431 – 43.
154. Bentires-Alj M, Barbu V, Fillet M, Chariot A, Relic B,
Jacobs N, et al. NF-kappaB transcription factor induces
drug resistance through MDR1 expression in cancer cells.
Oncogene 2003;22:90 – 7.
155. Tak PP, Smeets TJ, Boyle DL, Kraan MC, Shi Y, Zhuang S,
et al. p53 overexpression in synovial tissue from patients with
early and longstanding rheumatoid arthritis compared with
patients with reactive arthritis and osteoarthritis. Arthritis
Rheum 1999;42:948 – 53.
158. Assaraf YG, Rothem L, Hooijberg JH, Stark M, Ifergan I,
Kathmann GA, et al. Loss of multidrug resistance protein 1
(MRP1) expression and folate efflux activity results in a
highly concentrative folate transport in human leukemia
cells. J Biol Chem 2003;278:6680 – 6.
160. Stein U, Walther W, Shoemaker RH. Modulation of mdr1
expression by cytokines in human colon carcinoma cells: an
approach for reversal of multidrug resistance. Br J Cancer
1996;74:1384 – 91.
162. Stein U, Walther W, Laurencot CM, Scheffer GL, Scheper
RJ, Shoemaker RH. Tumor necrosis factor-alpha and
expression of the multidrug resistance- associated genes
LRP and MRP. J Natl Cancer Inst 1997;89:807 – 13.
163. Efferth T. Adenosine triphosphate-binding cassette trans-
porter genes in ageing and age-related diseases. Ageing Res
Rev 2003;2:11 – 24.
167. Van der Heijden J, de Jong MC, Dijkmans BAC, Lems WF,
Oerlemans R, Kathmann GA, et al. Development of
sulphasalazine resistance in human T cells induces expression
of the multidrug resistance transporter ABCG2 (BCRP) and
augmented production of TNFalpha. Ann Rheum Dis. In
press 2003.
168. Van der Heijden J, de Jong MC, Dijkmans BAC, Lems WF,
Oerlemans R, Kathmann GA, et al. Acquired resistance of
human T cells to sulphasalazine: stability of the resistant
MDR and rheumatoid arthritis 335
www.scandjrheumatol.dk
Scan
d J
Rhe
umat
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y Q
UT
Que
ensl
and
Uni
vers
ity o
f T
ech
on 1
1/01
/14
For
pers
onal
use
onl
y.

phenotype and sensitivity to non-related DMARDs. Ann
Rheum Dis. In press 2003.
169. Jansen G, Oerlemans R, Van der Heijden J, Lems WF,
Scheper RJ, Assaraf YG, Dijkmans BAC. Sulphasalazine is a
potent inhibitor of the reduced folate carrier, the dominant
cell membrane transporter for natural folates and metho-
trexate [Abstract]. Arthritis Rheum. In press 2003.
174. Maillefert JF, Maynadie M, Tebib JG, Aho S, Walker P,
Chatard C, et al. Expression of the multidrug resistance
glycoprotein 170 in the peripheral blood lymphocytes of
rheumatoid arthritis patients. The percentage of lymphocytes
expressing glycoprotein 170 is increased in patients treated
with prednisolone. Br J Rheumatol 1996;35:430 – 5.
175. Yudoh K, Matsuno H, Nakazawa F, Yonezawa T, Kimura
T. Increased expression of multidrug resistance of P-
glycoprotein on Th1 cells correlates with drug resistance in
rheumatoid arthritis. Arthritis Rheum 1999;42:2014 – 5.
176. Llorente L, Richaud-Patin Y, Diaz-Borjon A, Alvarado
DLB, Jakez-Ocampo J, De La FH, et al. Multidrug
resistance-1 (MDR-1) in rheumatic autoimmune disorders.
Part I: Increased P-glycoprotein activity in lymphocytes from
rheumatoid arthritis patients might influence disease out-
come. Joint Bone Spine 2000;67:30 – 9.
177. Maillefert JF, Puechal X, Falgarone G, Daumen-Legre V,
Liote F, Cherasse A, et al. Expression of the multidrug
resistance p-glycoprotein on peripheral blood monocytes as a
predictor of response to second-line therapy in rheumatoid
arthritis. Arthritis Rheum 2002;46:S134.
178. Jorgensen C, Sun R, Rossi JF, Costes J, Richard D, Bologna
C, Sany J. Expression of a multidrug resistance gene in
human rheumatoid synovium. Rheumatol Int 1995;15:
83 – 6.
181. Fisher GA, Lum BL, Sikic BI. The reversal of multidrug
resistance. Cancer Treat Res 1995;78:45 – 70.
183. Duffy CP, Elliott CJ, O’Connor RA, Heenan MM, Coyle S,
Cleary IM, et al. Enhancement of chemotherapeutic drug
toxicity to human tumour cells in vitro by a subset of non-
steroidal anti-inflammatory drugs (NSAIDs). Eur J Cancer
1998;34:1250 – 9.
186. Allen JD, van Loevezijn A, Lakhai JM, van der Valk M, van
Tellingen O, Reid G, et al. Potent and specific inhibition
of the breast cancer resistance protein multidrug transporter
in vitro and in mouse intestine by a novel analogue of
fumitremorgin C. Molecular Cancer Therapeutics 2002;
1:417 – 25.
187. Tan B, Piwnica-Worms D, Ratner L. Multidrug resistance
transporters and modulation. Curr Opin Oncol 2000;
12:450 – 8.
189. van Zuylen L, Nooter K, Sparreboom A, Verweij J.
Development of multidrug-resistance convertors: sense or
nonsense? Invest New Drugs 2000;18:205 – 20.
190. Thomas H, Coley HM. Overcoming multidrug resistance
in cancer: an update on the clinical strategy of inhibiting
p-glycoprotein. Cancer Control 2003;10:159 – 65.
192. Langmann T, Mauerer R, Zahn A, Moehle C, Probst M,
Stremmel W, Schmitz G. Real-time reverse transcription-
PCR expression profiling of the complete human ATP-
binding cassette transporter superfamily in various tissues.
Clin Chem 2003;49:230 – 8.
193. van der Pol MA, Broxterman HJ, Pater JM, Feller N,
van der MM, Weijers GW, et al. Function of the ABC
transporters, P-glycoprotein, multidrug resistance protein
and breast cancer resistance protein, in minimal residual
disease in acute myeloid leukemia. Haematologica
2003;88:134 – 47.
194. Kerb R, Hoffmeyer S, Brinkmann U. ABC drug transpor-
ters: hereditary polymorphisms and pharmacological impact
in MDR1, MRP1 and MRP2. Pharmacogenomics
2001;2:51 – 64.
199. Zamber CP, Lamba JK, Yasuda K, Farnum J, Thummel K,
Schuetz JD, Schuetz EG. Natural allelic variants of breast
cancer resistance protein (BCRP) and their relationship to
BCRP expression in human intestine. Pharmacogenetics
2003;13:19 – 28.
201. Ito K, Oleschuk CJ, Westlake C, Vasa MZ, Deeley RG, Cole
SP. Mutation of Trp1254 in the multispecific organic anion
transporter, multidrug resistance protein 2 (MRP2)
(ABCC2), alters substrate specificity and results in loss
of methotrexate transport activity. J Biol Chem 2001;
276:38108 – 14.
203. Diaz-Borjon A, Richaud-Patin Y, Alvarado DLB, Jakez-
Ocampo J, Ruiz-Arguelles A, Llorente L. Multidrug
resistance-1 (MDR-1) in rheumatic autoimmune disorders.
Part II: Increased P-glycoprotein activity in lymphocytes
from systemic lupus erythematosus patients might affect
steroid requirements for disease control. Joint Bone Spine
2000;67:40 – 8.
336 G Jansen et al
www.scandjrheumatol.dk
Scan
d J
Rhe
umat
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y Q
UT
Que
ensl
and
Uni
vers
ity o
f T
ech
on 1
1/01
/14
For
pers
onal
use
onl
y.