
Morphology and Anatomy of the Flower of Meliosma (Sabiaceae)- Implications for Pollination
13
Morphology and anatomy of the flower of Meliosma (Sabiaceae): implications for pollination biology L. P. Ronse De Craene, 1 L. Wanntorp 2 1 Royal Botanic Garden Edinburgh, Edinburgh, UK 2 Swedish Museum of Natural History, Stockholm, Sweden Received 29 June 2007; Accepted 9 October 2007; Published online 7 December 2007 Ó Springer-Verlag 2007 Summary. The structure and anatomy of mature flowers of four species of Meliosma is investigated using scanning electron and light microscopy. The vasculature of the flower, including the structure of the gynoecium, is described in detail. The mechanism of stamen maturation and pollen release is illustrated and discussed. The existence of an explosive pollina- tion mechanism is questioned for at least part of the species. Flowers are proterandrous and fertile stamens are kept spatially separate from the style by a ring of large staminodes. Anthers are disporangiate by the loss of the adaxial pollen sacs. During maturation the filament bends progressively outwards and releases the pollen on the extension of the connective that acts as a secondary pollen presentation system. The nectary has five appendages topped with stomata secreting abundant nectar. The relationships of Sabi- aceae are discussed relative to other early diverging eudicots. The significance of Sabiaceae as an isolated clade is highlighted, although some features point to a link with Menispermaceae. Keywords: Meliosma; Sabia; Sabiaceae; early diverging eudicots; disporangiate anthers; floral anat- omy; staminodes; nectary; pollination Introduction The Sabiaceae is a small family of three genera Sabia Colebr., Ophiocaryon Endl. and Meliosma Bl. distributed in Eastern and South Eastern Asia and tropical Central and South America. Melios- ma with about 25–70 species has the widest distribution occurring both in Asia and America, while Sabia with about 19–50 species is restricted to Asia, and Ophiocaryon with about 7 species is only found in tropical South America (Chen 1943, van Beusekom 1971, Barneby 1972, Kubitzki 2007). Occasionally two separate fam- ilies Sabiaceae and Meliosmaceae have been considered (e.g. Dahlgren 1981, Takhtajan 1997), although a unified Sabiaceae is to be preferred (e.g. Warburg 1895, Cronquist 1981, Stevens 2007). Earlier classifications placed Sabiaceae either in the Sapindales (e.g. Bentham and Hooker 1862, Dahlgren 1981, Takhtajan 1997), or in the Ranunculales close to Menispermaceae (e.g. Warburg 1895, Cronquist 1981). However, all recent phylogenetic studies based on molecular Correspondence: Louis P. Ronse De Craene, Royal Botanic Garden Edinburgh, 20A Inverleith Row, Edinburgh EH3 5LR, UK e-mail: [email protected] Pl Syst Evol 271: 79–91 (2008) DOI 10.1007/s00606-007-0618-y Printed in The Netherlands Plant Systematics and Evolution
Transcript of Morphology and Anatomy of the Flower of Meliosma (Sabiaceae)- Implications for Pollination

Morphology and anatomy of the flower of Meliosma (Sabiaceae):implications for pollination biology
L. P. Ronse De Craene,1 L. Wanntorp2
1Royal Botanic Garden Edinburgh, Edinburgh, UK2Swedish Museum of Natural History, Stockholm, Sweden
Received 29 June 2007; Accepted 9 October 2007; Published online 7 December 2007
� Springer-Verlag 2007
Summary. The structure and anatomy of mature
flowers of four species of Meliosma is investigated
using scanning electron and light microscopy. The
vasculature of the flower, including the structure of
the gynoecium, is described in detail. The mechanism
of stamen maturation and pollen release is illustrated
and discussed. The existence of an explosive pollina-
tion mechanism is questioned for at least part of the
species. Flowers are proterandrous and fertile stamens
are kept spatially separate from the style by a ring of
large staminodes. Anthers are disporangiate by the
loss of the adaxial pollen sacs. During maturation the
filament bends progressively outwards and releases
the pollen on the extension of the connective that acts
as a secondary pollen presentation system. The
nectary has five appendages topped with stomata
secreting abundant nectar. The relationships of Sabi-
aceae are discussed relative to other early diverging
eudicots. The significance of Sabiaceae as an isolated
clade is highlighted, although some features point to a
link with Menispermaceae.
Keywords: Meliosma; Sabia; Sabiaceae; early
diverging eudicots; disporangiate anthers; floral anat-
omy; staminodes; nectary; pollination
Introduction
The Sabiaceae is a small family of three genera
Sabia Colebr., Ophiocaryon Endl. and MeliosmaBl. distributed in Eastern and South Eastern Asia
and tropical Central and South America. Melios-ma with about 25–70 species has the widest
distribution occurring both in Asia and America,
while Sabia with about 19–50 species is
restricted to Asia, and Ophiocaryon with about
7 species is only found in tropical South America
(Chen 1943, van Beusekom 1971, Barneby 1972,
Kubitzki 2007). Occasionally two separate fam-
ilies Sabiaceae and Meliosmaceae have been
considered (e.g. Dahlgren 1981, Takhtajan 1997),
although a unified Sabiaceae is to be preferred
(e.g. Warburg 1895, Cronquist 1981, Stevens
2007).
Earlier classifications placed Sabiaceae either
in the Sapindales (e.g. Bentham and Hooker
1862, Dahlgren 1981, Takhtajan 1997), or in the
Ranunculales close to Menispermaceae (e.g.
Warburg 1895, Cronquist 1981). However, all
recent phylogenetic studies based on molecular
Correspondence: Louis P. Ronse De Craene, Royal Botanic Garden Edinburgh, 20A Inverleith Row, Edinburgh EH3 5LR, UK
e-mail: [email protected]
Pl Syst Evol 271: 79–91 (2008)
DOI 10.1007/s00606-007-0618-y
Printed in The Netherlands
Plant Systematicsand Evolution

data have placed the family in the early diverging
eudicots, but uncertainties remain about its
position relative to Proteales and Buxales, or
even core eudicots (e.g. Hilu et al. 2003; Soltis
et al. 2003, 2005; Furness et al. 2007; Worberg
et al. 2007).
Wanntorp and Ronse De Craene (2007)
investigated the floral development of selected
species of Meliosma and found that the pentam-
erous flowers of Sabiaceae have a unique origin
with a spiral initiation throughout, adding support
to the hypothesis that pentamery has arisen
independently in the family.
The flowers of Meliosma are deceivingly
complex (Wanntorp and Ronse De Craene
2007). They consist of four to five small sepals,
five petals of two different sizes, three stamin-
odes, two fertile stamens, and a superior,
bicarpellate ovary. The staminodes are situated
opposite the large petals and are basally adnate
to them. The stamens are dorsally fused with a
small petal. Stamens and staminodes are
arranged in a closely coherent unit. Anthers
consist of a broad basal platform, in some
species extending into a crenulate rim, and bear
two globular pollen sacs in an apical-adaxial
position. The platform goes over into a narrow
flattened filament. Staminodes form pouches in
which the globular pollen sacs fit tightly when in
bud. Two of the staminodes are asymmetrically
built because only one pouch is developed, and
the odd one has two pouches. The four pollen
sacs fit with these four pouches. The staminodes
form a tightly connected rim encircling the style.
The gynoecium consists of two fused carpels
with two connivent, occasionally twisted styles.
The base of the ovary is surrounded by a
conspicuous nectary with five prominent
appendages alternating with the stamens and
staminodes. A floral diagram illustrating the
arrangement of floral parts is given in Wanntorp
and Ronse de Craene (2007).
Descriptions of the morphology of the flowers
of Meliosma often lack detail and are inaccurate
(e.g., Gagnepain 1950, see Discussion in Wann-
torp and Ronse De Craene 2007). Little is known
about the pollination of the tiny flowers and
almost nothing is known regarding the anatomy
of the flower. Warburg (1895) and van Beusekom
(1971) argue that the flowers of Meliosma have
an explosive pollination mechanism, whereas the
stamens are held under tension by the staminodial
appendages. In this study we analyze the floral
anatomy and the mature morphology of selected
species of Meliosma to understand the internal
structure of the flower and clarify the pollination
mechanism. Comparison is made with the flower
of the sister genus Sabia. Knowledge of the floral
morphology of families of the early diverging
eudicots becomes increasingly important to
understand the floral evolution of the angio-
sperms. This paper aims to contribute to that
goal.
Materials and methods
Flower buds of four species of Meliosma were used:
M. veitchiorum Hemsl. (accession number 19521006),
M. dilleniifolia (Wall.) Wall. aff. ssp. tenuis (Maxim.)
Beus. (accession number 19632056; M. dilleniifolia(Wall.) Wall. ssp. cuneifolia (Franch.) Beus. (syn. M.cuneifolia Franch., also used by Wanntorp and Ronse
De Craene 2007: accession number 19381038) in
cultivation at the Royal Botanic Garden Edinburgh
(RBGE), and M. pinnata (Roxb.) Walp. ssp. arnotti-ana (Walp.) Beus. var. oldhamii (Maxim.) Beus. (syn.
M. oldhamii Maxim.) from Kyoto Botanical Garden
(Japan). Floral buds were collected, fixed in FAA (5%
acetic acid, 5% formaldehyde, 90% ethanol 70%), and
subsequently stored in 70% alcohol. Additional
observations were made on flowering trees of M.veitchiorum and M. alba (Schlechtend.) Walp. (syn.
M. beaniana Rehd. & Wils.: accession number:
19081007) cultivated at RBGE. Meliosma veitchiorumand M. alba belong to subgenus Kingsboroughia,
while M. dilleniifolia and M. pinnata belong to
subgenus Meliosma. Buds and mature flowers were
dissected and prepared using a Wild MZ8 stereo-
microscope (Leica, Wetzlar, Germany), dehydrated in
an ethanol–acetone series, and critical point dried with
a K850 Critical Point dryer (Emitech Ltd, Ashford,
Kent, UK). The dried material was later coated with
platinum using an Emitech K575X sputter coater
(Emitech Ltd, Ashford, Kent, UK) and examined with
a Supra 55VP scanning electron microscope (LEO
Electron Microscopy Ltd, Cambridge, UK). Reference
material (in ethanol) (831 Led, 832 Led, 833 Led, 876
Led) is kept at RBGE.
80 L. P. Ronse De Craene, L. Wanntorp: Floral anatomy and pollination mechanism in Meliosma

For light microscopy flower buds were embedded
in Kulzer’s Technovit (2-hydroxyethyl methacrylate),
as described in Igersheim and Cichocki (1996) and
sectioned with a Leitz Minot 1212 rotary microtome
fitted with metal blade. The sections (about 8 l thick)
were stained with ruthenium red and toluidine blue
and enclosed in DMX.
Results
Floral anatomy. Floral anatomy was investigated
in flower buds of Meliosma veitchiorum, M.pinnata, and M. dilleniifolia ssp. cuneifolia. No
separate descriptions are given for individual
species as anatomy was generally found to be
similar. A closed vascular system is found at the
base of the flower. Traces for sepals and bracts arise
at different levels corresponding to their initiation
sequence. The five sepals are supplied by a single
vascular trace that divides into one median and two
laterals higher in the sepal (Fig. 1). Five common
stamen–petal traces are given off at a higher level
following the departure of sepal traces (Fig. 2).
Stamens/staminodes and the corresponding petals
are detached as common organs and the vascular
bundles branch off into the respective organs (Figs.
3, 4). The division of bundles to the stamens occurs
below that of the staminodes. At this level a nectary
is visible as small-celled tissue and two traces
diverge from the central stele (Fig. 4, arrows). The
gynoecium is delimited from the nectary and the
two traces differentiate as the two dorsal bundles of
the carpels (Figs. 4, 5). While the locules are visible
the central vascular tissue becomes reorganized in
two areas, each becoming the ventral traces for the
respective ovules of each carpel by halving (Figs. 5,
6, 12). The ventral traces are used up in supplying
the ovules. At the top of the ovary the dorsals fade
out and no traces run into the styles. Ovules are
initially inserted parallelly (only one of a pair
visible in Fig. 10) but in older buds they become
superposed and a septum separates the two locules
(Figs. 11–13, 15). The ovules fill the ovarian cavity
completely. The placenta is covered with darkly
staining secretory trichomes (Fig. 13) and secretion
extends along the whole stylar canal (Fig. 16). In M.pinnata and M. dilleniifolia the ovules have a single
integument (Figs. 11, 12, 15, 16). In M. veitchiorum
two integuments are formed as a rim around the
nucellus (Figs. 6, 14, 17). We occasionally found
that the outer integument is incompletely
developed, suggesting that the single integument
is the inner one in M. pinnata and M. dilleniifolia.
The styles extend beyond the ring of staminodes
(Figs. 8, 9, 18). Styles are weakly to strongly
appressed but not fused (Figs. 9, 18, 39–42). In M.pinnata and M. dilleniifolia the two styles are erect
and terminate in a common slit-like stigmatic area
(Figs. 9, 39, 40, 42). Styles are not closely
appressed to each other and are often intertwined
in M. veitchiorum (Fig. 27). No vascular tissue runs
in the styles but the two stylar canals are filled with
a secretion (Fig. 18). At the top of the styles the
stylar canals open adaxially into a broad slit. Anther
development has been described in Wanntorp and
Ronse De Craene (2007). Ovule development
corresponds with microsporogenesis (Fig. 25). In
young anthers the anther wall is composed of an
epidermis, an endothecium which is one cell thick,
a few intermediate layers, and an inner secretory
tapetum layer of the secretory type (Figs. 19, 20). In
older anthers the tapetum layer is partly resolved
and the endodermis shows tangential thickenings
(Figs. 21–23). The ovary wall, as well as the
anthers, contains cells with a broad prismatic
calcium oxalate crystal. In the anthers these are
mainly situated in the area of the vascular bundle
(Figs. 23, 24). By disintegration of the anther wall,
the crystals become intermingled with the pollen.
Pollen grains are tricolporate, about 15 l in size,
and with a reticulate exine.
Flowering process. Flowers are proter-
androus. There is a difference to the extent of
development of the style between different
species. In M. veitchiorum and M. pinnata the
styles grow between the tightly connivent
staminodes before the buds opens (Figs. 18,
27). In M. dilleniifolia ssp. cuneifolia the
development of the style lags behind and the
closely interconnected styles emerge when the
anthers are already open (Figs. 28, 30, 36). In
young flower buds the staminodes surround the
young styles in a coherent unit and the young
anthers fit in adaxial folds formed by the
staminodes (Figs. 9, 27, 36). At this stage the
filaments are abruptly bent inwards in the middle
L. P. Ronse De Craene, L. Wanntorp: Floral anatomy and pollination mechanism in Meliosma 81
82 L. P. Ronse De Craene, L. Wanntorp: Floral anatomy and pollination mechanism in Meliosma

and the anthers are hidden from view (Fig. 27).
One of the three staminodes is symmetrically
developed and encapsulates one pollen sac of
each anther; the two other staminodes have only a
single lobe that encloses the other pollen sac
(Figs. 6, 8, 26, 36). In this way the androecium
forms a tightly closed dome covering disc and
ovary. When the flowers open, the style protrudes
between the staminodes before the stamens are
clearly visible. By pressing on the filament with a
needle, the anthers pop out of the pouch formed
by the staminodes (Figs. 27–30). When the flower
expands, the filaments bend outwards and the
anthers become detached from the tight grip of
the staminodes. This is a progressive process in
M. veitchiorum and M. alba, as we could not see
the explosive mechanism described by van
Beusekom (1971) and Warburg (1895). In M.dilleniifolia we often found fewer stamens in
opened flowers (Figs. 28, 30), suggesting that
they drop off at an early stage, possibly by an
explosive mechanism. While the filaments curve
outwards the pollen sacs dehisce by a slit situated
on the inner surface (Figs. 27, 28, 30, 34, 35).
The single pollen sacs open inward out; the
anther wall curves outwards and exposes the
pollen grains that are collected in clumps on the
broad connective expansion (Figs. 29, 34, 35).
This broad dish-like area differs in shape between
different species, being shallow and extending as
flaps next to the pollen sacs in M. veitchiorum(Figs. 9, 27), and forming two ventral auricles in
M. dilleniifolia (Figs. 31–33) and M. pinnata(Fig. 34). The sticky pollen remains in clumps on
the protuberance for the insect visitor to collect
(Figs. 29, 34). When pollen is released the
anthers continue their outward curve (Fig. 30).
Soon after pollen release the stamens and
staminodes drop off with the petals. The
pollination mechanism might explain the
heteromorphy of the petals; the broad erect
petals enclose the bud and hold the staminodes
erect (Figs. 27, 28), while the smaller often bifid
petals do allow for the filament to curve outwards
(Figs. 32, 33). After the flower has dropped its
petals and stamens the ovary continues its
development sheathed by the nectary. The
ovary wall is covered with uniseriate multi-
cellular trichomes (Fig. 44).
The nectary in Meliosma is a broad disc
surrounding the superior ovary. There is no
vascular tissue but the secretory cells are small
with large nuclei (Figs. 4, 12, 18, 25, 38). At
maturity the nectary is either developed in the
shape of a tire or a buoy with weak crenellations
(Fig. 41), or it resembles a crown bearing five
prominent appendages topped with one to three
stomata (Figs. 39, 40, 43). Both types were
observed in M. dilleniifolia (Figs. 40, 41), while
in the other species only the crenulate disk was
found. Abundant secretion occurs through these
distal stomata (Figs. 45, 46). The appendages are
not equidistant, but four are grouped in pairs, while
the odd one is situated at the other end of the disc
(Figs. 40, 43). The arrangement of appendages
corresponds with the insertion of the stamens—the
pairs are close to the fertile stamen in the median
plane of the flower. As the staminodes tightly
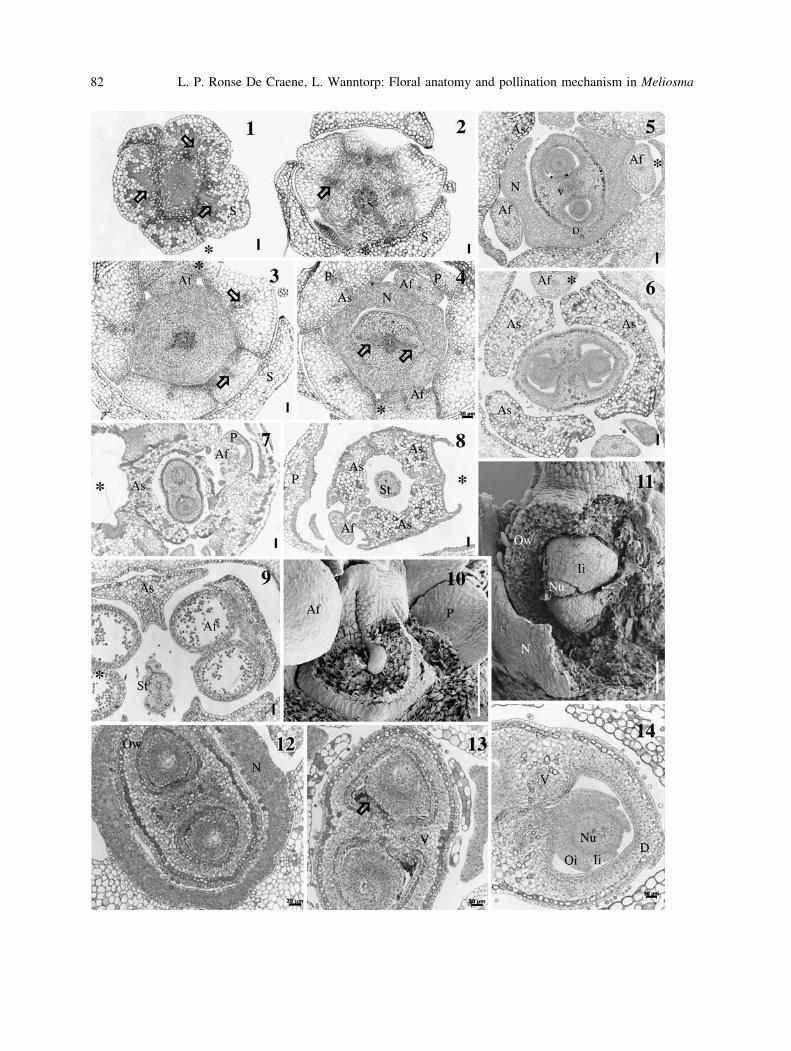
Figs. 1–14. Transverse and longitudinal sections of the flower of Meliosma. 1, 7, 8, 10–13 M. dilleniifolia ssp.
cuneifolia; 2–6, 9, 14 M. veitchiorum. 1 TS at base of flower with departure of traces to each sepal (arrows). 2, 3Successive TS with departure of common petal–stamen traces (arrows). 4 Division of stamen petal traces and
separation of respective organs. Note nectary and departure of two dorsals (arrows) from the central stele. 5 TS
through the upper part of the nectary. 6 TS through the middle of the ovary with ovule attachment. 7 TS through
the upper part of the ovary. 8 TS through the base of the style. Note the connected staminodes. 9 TS through the
stylar and anther regions. 10 Young ovary showing one of the developing ovules. 11 Section of older ovary
showing two ovules within a locule. 12 TS in the lower part of the ovary showing two ovules and the nectary. 13TS through the middle of the ovary with placental tissue. The arrow points to secretory trichomes. 14 TS of one
locule showing an ovule with two integuments and attachment to the placenta. 1–3, 5–9 Bar 50 lm, 4 12,
13 = 20 lm; 10, 11 = 100 lm. Af fertile stamen, As sterile stamen, D dorsal trace, Ii inner integument, Nnectary, Nu, nucellus, Oi outer integument, Ow ovary wall, P petal or petal trace, S sepal, St style, V ventral trace.
Asterisks point to the adaxial side of the flower
b
L. P. Ronse De Craene, L. Wanntorp: Floral anatomy and pollination mechanism in Meliosma 83
84 L. P. Ronse De Craene, L. Wanntorp: Floral anatomy and pollination mechanism in Meliosma

cover the nectary as a lid (Figs. 6–8,38), nectar is
produced and accumulates between the broad
disc lobes. Access to nectar is only possible
through slits between the staminodial lobes—this
induces the pollinating insects to wander close to
the anther tissue.
Discussion
Anther dehiscence. The broad dish-like append-
age of the anthers has been described as an
expanded connective by various authors (e.g.
Baillon 1874, Warburg 1895, Takhtajan 1997).
However, the fact that the anthers are
disporangiate has been overlooked by most
authors (except for brief mentions in the
literature (e.g. Warburg 1895, Baillon 1874;
Chen 1943, Kubitzki 2007). Sabiaceae were
left out in the global review of anthers of
Endress and Stumpf (1990). Sabia shares the
disporangiate anthers with Meliosma (Ronse De
Craene unpubl. obs.; van de Water 1980). One
tricky question is whether the adaxial or the
abaxial pair of pollen sacs is reduced.
Developmentally it is not possible to observe
any traces of the second set of pollen sacs
(Wanntorp and Ronse De Craene 2007). The
normal dehiscence pattern for tetrasporangiate
anthers in most angiosperms is laterally with the
lobes curving towards the exterior (Fig. 47A).
When dehiscing, the anther walls of Meliosmacurve inward out, exposing the pollen on the
broad connective. A plausible explanation for
this pattern is that the adaxial (ventral) pollen
sacs have been lost and that the broadened
connective has pushed the two remaining pollen
sacs towards each other (Fig. 47B). In the case of
tetrasporangiate anthers opening in a similar
fashion one would expect the abaxial pollen sacs
to curve out in the same way as in Meliosma.
Van de Water (1980) described a similar
dehiscence pattern for Sabia. Sabia has a
broad connective; the anthers open along the
connection with the connective at the adaxial
side and the wall turns inside out. When fully
opened the anthers give the impression of being
extrorse. Disporangiate anthers are fairly
common in basal angiosperms (e.g. Lauraceae,
Monimiaceae, Hernandiaceae) and their location
is difficult to assess. They have been correlated
with a specialized floral biology such as a
secondary pollen presentation on the gynoecium
or a precise deposition of pollen linked to the
dehiscence of the thecae by narrow slits (Endress
and Stumpf 1990). In the case of Meliosma the
anthers are set widely apart and separated by the
broad staminodes; pollen is not deposited on the
gynoecium, but on the extended connective
appendage. It appears that Meliosma shows a
unique pattern of secondary pollen presentation
by accumulating pollen on the extensive
connective-dish. The gradual movement of the
anthers exposes the dish to visiting insects.
However, more experimental work needs to be
carried out in other species to verify the
statement of van Beusekom whether an
explosive mechanism can be invoked. In that
case the broad connective can act as a container
catapulting the pollen on the insect. Zhilin (1981)
suggested that the explosive mechanism is
restricted to subgenus Meliosma to which M.dilleniifolia and M. pinnata belongs. Subgenus
Kingsboroughia containing M. veitchiorum and
M. alba is said to lack the explosive mechanism
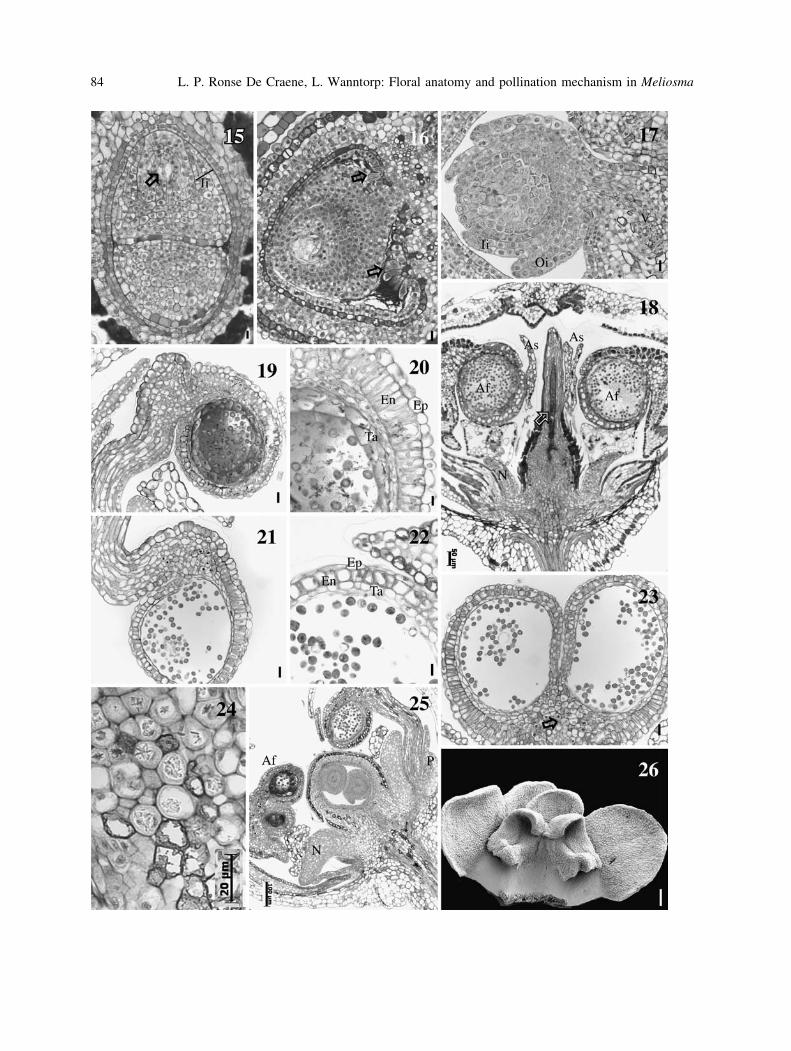
Figs. 15–26. Transverse and longitudinal sections of the flower of Meliosma. 15, 16, 18 M. pinnata; 17, 19–26M. veitchiorum. 15 LS showing two superposed ovules. In the upper ovule an embryo sac is visible (arrow). 16TS through locule. Note the single integument and secretory hairs on the placenta (arrow). 17 TS with detail of
ovule with two integuments. 18 LS through young flower. Note the style filled with secretion reaching into the
ovary (arrow). 19 LS of anther at meiosis. 20 LS of anther wall. 21 LS of mature anther. 22 TS of anther wall. 23TS through the middle of mature anther. Arrow points to the position of calcium oxalate crystals. 24 Detail of
cells in the connective area. 25 LS of part of the flower showing two superposed ovules. 26 Adaxial view of
staminode with two interconnected petals. 18, Bar = 50 lm, Figs. 19, 21, 23 = 10 lm, 15–17, 20, 22,
24 = 20 lm, 26 = 200 lm. Af fertile stamen, As sterile stamen, En endodermis, Ep epidermis, Ii inner
integument, N nectary, Nu, nucellus, Oi outer integument, P petal, Ta tapetum, V ventral trace
b
L. P. Ronse De Craene, L. Wanntorp: Floral anatomy and pollination mechanism in Meliosma 85
86 L. P. Ronse De Craene, L. Wanntorp: Floral anatomy and pollination mechanism in Meliosma

and the flowers remain open for a longer time.
This corresponds with our observations.
Floral anatomy. The ovary is syncarpous
but the styles are postgenitally united. The ovules
are initially positioned side by side in each locule
but they become superposed by the restricted
space for their further development. The
embryology of Sabiaceae has been briefly
described by Mauritzon (1936), Raju (1952),
and Sharanina (1996). They mention that the
ovules are unitegmic and have no micropyle. We
report the presence of two integuments in
Meliosma veitchiorum, contrary to descriptions
of unitegmy in textbooks obviously taken over
from earlier studies (e.g. Cronquist 1981,
Takhtajan 1997). However, the other species
studied were unitegmic. Endress and Igersheim
(1999) briefly described the gynoecium structure
of Sabia and reported the presence of mucilage in
style and ovary. We report the presence of intra-
ovarian secretory trichomes situated on the
placenta. These trichomes produce ovarian
mucilage which is also found in the pollen-
transmitting tract of the style and play an obvious
role in facilitating pollen tube growth (Rudall
et al. 1998). Secretory intra-ovarian trichomes are
rare in the eudicots, but are found more regularly
in the monocots and some basal angiosperms.
There is no discussion on crystals in anthers,
but Raju (1952) mentions crystals in the ovary
wall cells. We found that calcium oxalate crystals
were often present in open anthers mixed with
pollen grains. D’Arcy et al. (1996) studied the
presence of calcium oxalate in anthers of differ-
ent families, not including Sabiaceae. Contrary to
species where calcium oxalate occurs in packages
at the level of anther dehiscence, in Meliosma
calcium oxalate occurs scattered in idioblasts
around the connective area. D’Arcy et al. (1996)
discussed several hypotheses for the presence of
calcium oxalate packages, including a role in
anther dehiscence, as a metabolic sink, discour-
agement of herbivores or as a food source. It is
possible that the association of calcium oxalate
with pollen in Meliosma is accidental without the
significance of the packages associated with
specialized flowers found in the asterids.
The apical stomata on the nectary have been
overlooked by most authors, who describe disc
and glands, and nectaries as separate entities (e.g.
Chen 1943). The whole disc-like structure with
appendages needs to be described as a nectary, as
it contains densely staining tissue responsible for
nectar secretion. Chen (1943) and van de Water
(1980) observed a similar variation in the nectary
of Sabia as in Meliosma in the extent of
development of the appendages, as well as
presence of glandular tissue on top of the
appendages.
Systematic considerations. The flower mor-
phology of Meliosma is reminiscent of Sabia in
many ways and supports the recognition of a
single family (Chen 1943, van de Water 1980,
Kubitzki 2007, Wanntorp and Ronse De Craene
2007). Sabia and Meliosma share several floral
characters including the filaments adhering to the
base of the petals, disporangiate anthers with a
similar pollen morphology, the five-lobed
nectary, and similar ovary and ovules. The
connective in Sabia is thick, although not
developed to the extent found in Meliosma. A
comparative study including the polarization of
character evolution will only be possible by
including the third genus of Sabiaceae,
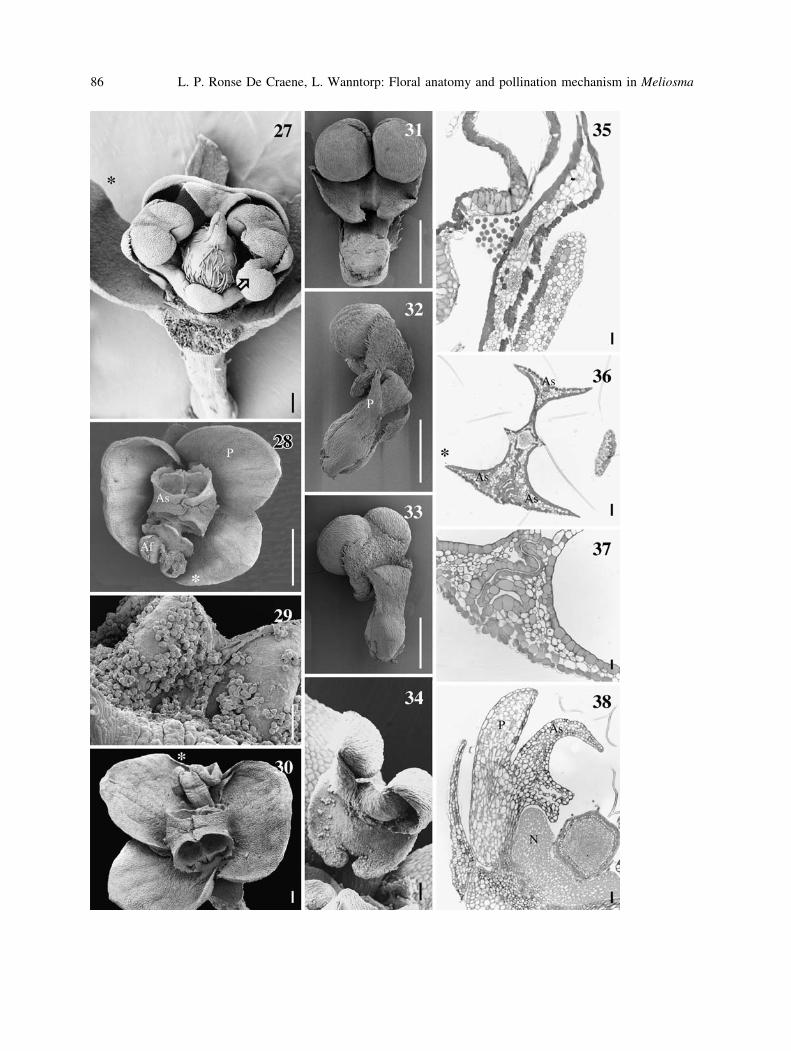
Figs. 27–38. Anthesis and anatomy of staminodes in Meliosma. 27, 29, 38 M. veitchiorum; 28, 30–33, 35–37:
M. dilleniifolia ssp. cuneifolia; 27, 34: M. pinnata. 27. Lateral view of flower at anthesis, two petals and two
staminodes removed. The right anther starts dehiscing (arrow). 28 Apical view of flower at the onset of
dehiscence. 29 Pollen grains on the exposed anther wall. 30 Flower with recurved filament. 31–33 adaxial,
lateral, and abaxial views of young anthers and their associated petals. 34 Dehisced anther with pollen grains on
the basal appendage. 35 TS of dehisced anther with pollen grains and the connective. 36 TS of staminodes
surrounding the style. 37 TS of two connected staminodes. 38 LS of nectary partly covered by staminodial
appendage. 35, 37, Bar = 20 lm, 36, 38 = 50 lm, 34 = 100 lm, 30–33 = 200 lm, 29–31 = 500 lm,
28 = 1 mm. Af fertile stamen, As sterile stamen, N nectary, P petal. Asterisks point to the adaxial side of the
flower
b
L. P. Ronse De Craene, L. Wanntorp: Floral anatomy and pollination mechanism in Meliosma 87
Ophiocaryon, for which material is currently not
available.
Affinities of Sabiaceae are still unclear.
Table 1 compares Sabiaceae with other poten-
tially related families in a number of characters.
A closer link with Menispermaceae (cf. Cron-
quist 1981) cannot be excluded based on pollen,
embryological and ovary characters. Some Meni-
spermaceae share dithecal anthers with Sabiaceae
(Endress and Stumpf 1990) but details of their
arrangement are not known. Interestingly some
Menispermaceae and Ranunculaceae have
unitegmic ovules with a rudimentary outer
integument (reported in Endress and Igersheim
1999). Sabiaceae share the presence of hemian-
atropous ovules with Menispermaceae and some
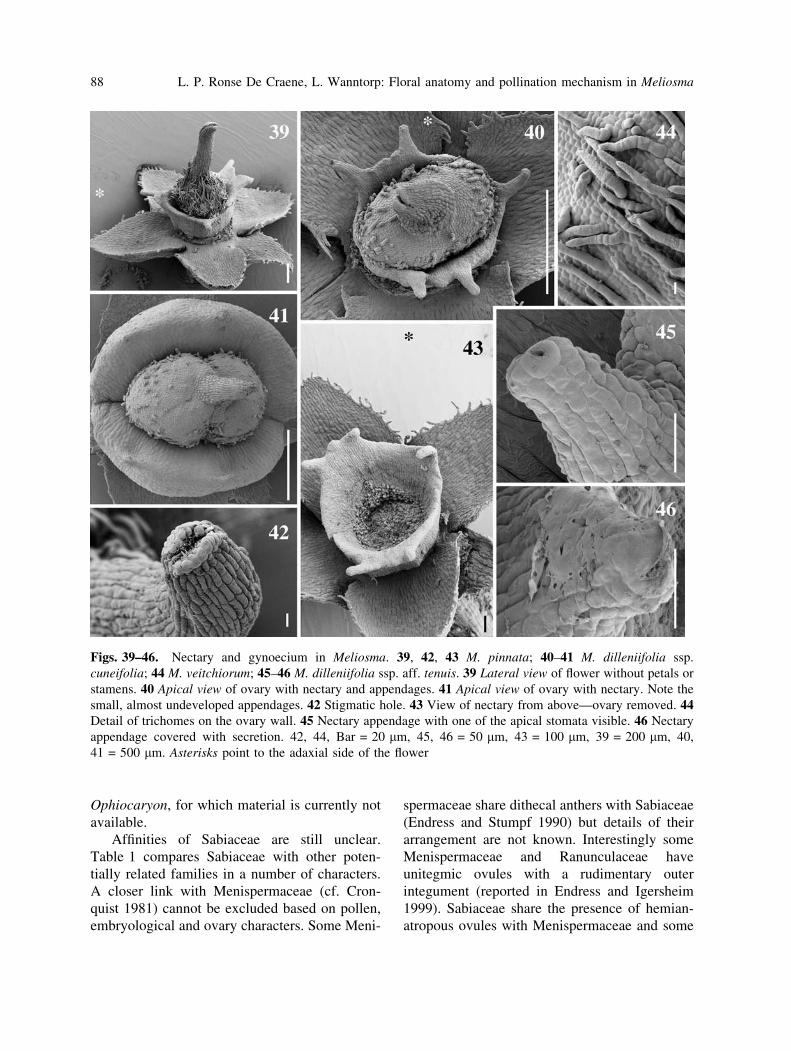
Figs. 39–46. Nectary and gynoecium in Meliosma. 39, 42, 43 M. pinnata; 40–41 M. dilleniifolia ssp.
cuneifolia; 44 M. veitchiorum; 45–46 M. dilleniifolia ssp. aff. tenuis. 39 Lateral view of flower without petals or
stamens. 40 Apical view of ovary with nectary and appendages. 41 Apical view of ovary with nectary. Note the
small, almost undeveloped appendages. 42 Stigmatic hole. 43 View of nectary from above—ovary removed. 44Detail of trichomes on the ovary wall. 45 Nectary appendage with one of the apical stomata visible. 46 Nectary
appendage covered with secretion. 42, 44, Bar = 20 lm, 45, 46 = 50 lm, 43 = 100 lm, 39 = 200 lm, 40,
41 = 500 lm. Asterisks point to the adaxial side of the flower
88 L. P. Ronse De Craene, L. Wanntorp: Floral anatomy and pollination mechanism in Meliosma
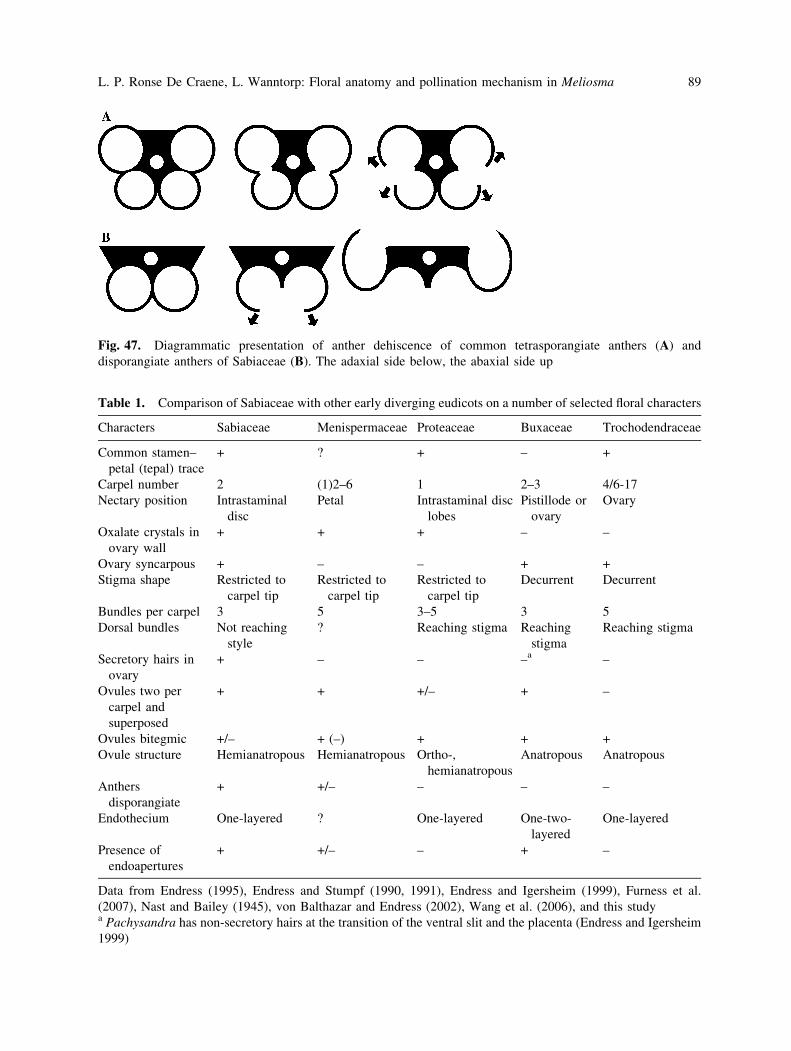
Fig. 47. Diagrammatic presentation of anther dehiscence of common tetrasporangiate anthers (A) and
disporangiate anthers of Sabiaceae (B). The adaxial side below, the abaxial side up
Table 1. Comparison of Sabiaceae with other early diverging eudicots on a number of selected floral characters
Characters Sabiaceae Menispermaceae Proteaceae Buxaceae Trochodendraceae
Common stamen–
petal (tepal) trace
+ ? + – +
Carpel number 2 (1)2–6 1 2–3 4/6-17
Nectary position Intrastaminal
disc
Petal Intrastaminal disc
lobes
Pistillode or
ovary
Ovary
Oxalate crystals in
ovary wall
+ + + – –
Ovary syncarpous + – – + +
Stigma shape Restricted to
carpel tip
Restricted to
carpel tip
Restricted to
carpel tip
Decurrent Decurrent
Bundles per carpel 3 5 3–5 3 5
Dorsal bundles Not reaching
style
? Reaching stigma Reaching
stigma
Reaching stigma
Secretory hairs in
ovary
+ – – –a –
Ovules two per
carpel and
superposed
+ + +/– + –
Ovules bitegmic +/– + (–) + + +
Ovule structure Hemianatropous Hemianatropous Ortho-,
hemianatropous
Anatropous Anatropous
Anthers
disporangiate
+ +/– – – –
Endothecium One-layered ? One-layered One-two-
layered
One-layered
Presence of
endoapertures
+ +/– – + –
Data from Endress (1995), Endress and Stumpf (1990, 1991), Endress and Igersheim (1999), Furness et al.
(2007), Nast and Bailey (1945), von Balthazar and Endress (2002), Wang et al. (2006), and this studya Pachysandra has non-secretory hairs at the transition of the ventral slit and the placenta (Endress and Igersheim
1999)
L. P. Ronse De Craene, L. Wanntorp: Floral anatomy and pollination mechanism in Meliosma 89

Proteaceae (Endress and Igersheim 1999). Pollen
grains are of a generalized type (tricolporate,
subprolate and reticulate, with endoapertures:
Erdtman 1952; Furness et al. 2007). Sabiaceae
share endoapertures only with two other fami-
lies of early-diverging eudicots, Buxaceae and
Menispermaceae. Recent molecular evidence
(Worberg et al. 2007) supports the monophyly
of Sabiaceae and places the family as the second
branch in the basal eudicot grade with inconclu-
sive support. This places the family in an isolated
position in relation to the other early diverging
eudicots as might be expected from the unusual
floral morphology which evolved clearly sepa-
rately with little evidence of a close association
with other families (Wanntorp and Ronse De
Craene 2007).
We thank Frieda Christie and Dr. Chris Jeffree for
assistance with the SEM. We are also grateful to Dr.
Toru Tokuoka for collecting flower buds of Meliosmapinnata. Travel for LW to RBGE was possible
through SYNTHESYS grant GB-TAF 1626. Helpful
comments by Dmitry Sokoloff and an anonymous
reviewer are gratefully acknowledged.
References
Baillon H (1874) Serie des Sabia. Histoire des Plantes
V: 345–348. Paris, Hachette
Barneby RC (1972) Meliosmaceae—Ophiocaryon. In:
Maguire B, et al (eds) The flora of the Guayana
Highlands—Part IX. Mem New York Bot Gard 23:
114–120
Bentham G, Hooker F JD (1862) Genera Plantarum 1.
London
Chen L (1943) A revision of the genus SabiaColebrooke. Sargentia 3: 1–75
Cronquist A (1981) An integrated system of classifi-
cation of the flowering plants. Columbia University
Press, New York
Dahlgren RMT (1981) A revised classification of the
angiosperms with comments on correlation between
chemical and other characters. In: Young DA,
Seigler S (eds) Phytochemistry and angiosperm
phylogeny. Praeger, New York, pp 149–204
D’Arcy WG, Keating RC, Buchmann SL (1996) The
calcium oxalate package or so-called resorption
tissue in some angiosperm anthers. In: D’Arcy WG,
Keating RC (eds) The anther. Form, function and
phylogeny. Cambridge University Press, Cam-
bridge, pp 159–191
Endress PK (1995) Floral structure and evolution in
Ranunculanae. Pl Syst Evol 9(Suppl): 47–61
Endress PK, Igersheim A (1999) Gynoecium diversity
and systematics of the basal eudicots. Bot J Linn
Soc 130: 305–393
Endress PK, Stumpf S (1990) Non-tetrasporangiate
stamens in the angiosperms: structure, systematic
distribution and evolutionary aspects. Bot Jahrb
Syst 112: 193–240
Endress PK, Stumpf S (1991) The diversity of stamen
structures in ‘lower’ Rosidae (Rosales, Fabales,
Proteales, Sapindales). Bot J Linn Soc 107: 217–
293
Erdtman G (1952) Pollen morphology and plant
taxonomy, Almqvist and Wiksell, Stockholm
Furness CA, Magallon S, Rudall PJ (2007) Evolution
of endoapertures in early-divergent eudicots, with
particular reference to pollen morphology in Sabi-
aceae. Pl Syst Evol 263: 77–92
Gagnepain F (1950) Meliosma (Sabiacee): sa fleur.
Bull Soc Bot Fr 97: 89–90
Hilu KW, Borsch T, Muller K, Soltis DE, Soltis PS,
Savolainen V, Chase MW, Powell MP, Alice LA,
Evans C, Sauquet H, Neinhuis R, Slotta TAB,
Rohwer JG, Campbell CS, Chatrou LW (2003)
Angiosperm phylogeny based on matK sequence
information. Amer J Bot 90: 1758–1776
Igersheim A, Cichocki O (1996) A simple method for
microtome sectioning of prehistoric charcoal spec-
imens embedded in 2-hydroxyethyl methacrylate
(HEMA). Rev Palaeobot Palyn 92: 389–393
Kubitzki K (2007) Sabiaceae. In: Kubitzki K (ed) The
families and genera of vascular plants, vol IX.
Springer, Berlin, pp 413–417
Mauritzon J (1936) Zur Embryologie und systemati-
schen Abgrenzung der Reihen Terebinthales und
Celastrales. Bot Not 1936: 161–212
Nast CG, Bailey IW (1945) Morphology and rela-
tionships of Trochodendron and Tetracentron II.
Inflorescence, flower, and fruit. J Arnold Arbor 46:
267–276
Raju MVS (1952) Embryology of Sabiaceae. Curr Sci
21: 107–108
Rudall PJ, Prychid CJ, Jones C (1998) Intra-ovarian
trichomes, mucilage secretion and hollow styles in
monocotyledons. In: Owens SJ, Rudall PJ (eds)
Reproductive biology. Royal Botanic Gardens,
Kew, pp 219–230
90 L. P. Ronse De Craene, L. Wanntorp: Floral anatomy and pollination mechanism in Meliosma

Sharanina EA (1996) Sabiaceae. In: Takhtajan A,
Danilova M (eds) Comparative anatomy of seeds,
vol. 5. Mir & Semja, St. Petersburg, pp 356–359
Soltis DE, Senters AE, Zanis MJ, Kim S, Thompson
JD, Soltis PS, Ronse De Craene LP, Endress PK,
Farris JS (2003) Gunnerales are sister to other core
eudicots: implications for the evolution of penta-
mery. Amer J Bot 90: 461–470
Soltis DE, Soltis PS, Endress PK, Chase MW (2005)
Phylogeny and evolution of angiosperms, Sinauer,
Sunderland
Stevens PF (2007) Angiosperm Phylogeny Website.
http://www.mobot.org/MOBOT/research/APweb/.
Version 8, June 2007
Takhtajan A (1997) Diversity and classification of
flowering plants, Columbia University Press, New
York
Van Beusekom CF (1971) Revision of Meliosma(Sabiaceae), section Lorenzanea excepted, living
and fossil, geography and pylogeny. Blumea 19:
355–529
Van de Water TPM (1980) A taxonomic revision of
the genus Sabia (Sabiaceae). Blumea 26: 1–64
von Balthazar M, Endress PK (2002) Reproductive
structures and systematics of Buxaceae. Bot J Linn
Soc 140: 193–228
Wang H, Meng A, Li J, Feng M, Chen Z, Wang W
(2006) Floral organogenesis of Cocculus orbicul-atus and Stephania dielsiana (Menispermaceae).
Int J Pl Sci 167: 951–960
Wanntorp L, Ronse De Craene LP (2007) Floral
development of Meliosma (Sabiaceae). Evidence
for multiple origins of pentamery in the eudicots.
Amer J Bot 94 (in press)
Warburg O (1895) Sabiaceae. In: Engler A, Prantl K
(eds) Die naturlichen Pflanzenfamilien III, 5: 367–
374. Engelmann, Leipzig
Worberg A, Quandt D, Barniske A-M, Lohne C, Hilu
KW, Borsch T (2007) Phylogeny of basal eudicots:
insights from non-coding and rapidly evolving
DNA. Org Div Evol 7: 55–77
Zhilin SG (1981) Sabiaceae. In: Takhtajan AL (ed)
Plant life, part 2 vol. 5. Prosveshchenie, Moscow,
pp 268–270
L. P. Ronse De Craene, L. Wanntorp: Floral anatomy and pollination mechanism in Meliosma 91







![Predistorter for Power Amplifier using Flower Pollination ... · flower pollination algorithm ([3] and [6]) which is developed based on the flower pollination process of flowering](https://static.fdocuments.in/doc/165x107/6036a860c9608f042126548f/predistorter-for-power-amplifier-using-flower-pollination-flower-pollination.jpg)










