
Morphological Evidence Phylogeny of Oriental Voles (Rodentia
14
BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. Phylogeny of Oriental Voles (Rodentia: Muridae: Arvicolinae): Molecular and Morphological Evidence Author(s): Shaoying Liu, Yang Liu, Peng Guo, Zhiyu Sun, Robert W. Murphy, Zhenxin Fan, Jianrong Fu and Yaping Zhang Source: Zoological Science, 29(9):610-622. 2012. Published By: Zoological Society of Japan DOI: http://dx.doi.org/10.2108/zsj.29.610 URL: http://www.bioone.org/doi/full/10.2108/zsj.29.610 BioOne (www.bioone.org ) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use . Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.
Transcript of Morphological Evidence Phylogeny of Oriental Voles (Rodentia

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions,research libraries, and research funders in the common goal of maximizing access to critical research.
Phylogeny of Oriental Voles (Rodentia: Muridae: Arvicolinae): Molecular andMorphological EvidenceAuthor(s): Shaoying Liu, Yang Liu, Peng Guo, Zhiyu Sun, Robert W. Murphy, Zhenxin Fan, JianrongFu and Yaping ZhangSource: Zoological Science, 29(9):610-622. 2012.Published By: Zoological Society of JapanDOI: http://dx.doi.org/10.2108/zsj.29.610URL: http://www.bioone.org/doi/full/10.2108/zsj.29.610
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological,and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and bookspublished by nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercialinquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.

2012 Zoological Society of JapanZOOLOGICAL SCIENCE 29: 610–622 (2012)
Phylogeny of Oriental Voles (Rodentia: Muridae: Arvicolinae):
Molecular and Morphological Evidence
Shaoying Liu1*, Yang Liu1, Peng Guo2, Zhiyu Sun1, Robert W. Murphy3,4,
Zhenxin Fan5, Jianrong Fu1, and Yaping Zhang3
1Sichuan Academy of Forestry, Chengdu, Sichuan 610066, China2College of Life Sciences and Food Engineering, Yibin University, Yibin, Sichuan 644007, China
3Kunming Institute of Zoology, the Chinese Academy of Science, Kunming, Yunnan 650223, China4Centre for Biodiversity and Conservation Biology, Royal Ontario Museum,
100 Queen’s Park, Toronto, Ontario, M5S 2C6 Canada5College of Life Sciences, Sichuan University,
Chengdu, Sichuan 610051, China
The systematics of Oriental voles remains controversial despite numerous previous studies. In this
study, we explore the systematics of all species of Oriental voles, except Eothenomys wardi, using
a combination of DNA sequences and morphological data. Our molecular phylogeny, based on two
mitochondrial genes (COI and cyt b), resolves the Oriental voles as a monophyletic group with
strong support. Four distinct lineages are resolved: Eothenomys, Anteliomys, Caryomys, and the
new subgenus Ermites. Based on morphology, we consider Caryomys and Eothenomys to be valid
genera. Eothenomys, Anteliomys, and Ermites are subgenera of Eothenomys. The molecular phy-
logeny resolves subgenera Anteliomys and Ermites as sister taxa. Subgenus Eothenomys is sister
to the clade Anteliomys + Ermites. Caryomys is the sister group to genus Eothenomys. Further, the
subspecies E. custos hintoni and E. chinensis tarquinius do not cluster with E. custos custos and
E. chinensis chinensis, respectively, and the former two taxa are elevated to species level and
assigned to the new subgenus Ermites.
Key words: Anteliomys, Caryomys, Eothenomys, Ermites, DNA barcoding
INTRODUCTION
Eothenomys (sensu lato) is a genus of Oriental voles
that mainly occurs in the Hengduan Mountains of southwest-
ern China, northeastern Burma, and Assam, India. Initially
erected as a subgenus of Microtus (Miller, 1896), Hinton
(1923) elevated the taxon to the level of genus based on
morphology. This arrangement is widely accepted.
The intrageneric taxonomy and systematics of Oriental
voles is controversial. Hinton (1923), followed by Gromov
and Polyakov (1977), did not recognize any subgenera of
Eothenomys. Hinton (1923) proposed recognition of Caryomys
and Anteliomys and considered them to be closely related
genera. Later, Hinton (1926), followed by Ellerman (1941)
and Ellerman and Morrison-Scott (1951), recognized
Eothenomys and Anteliomys as valid genera, and he con-
sidered Caryomys to be immature individuals of the subspe-
cies Evotomys rufocanus shanseius. Ma and Jiang (1996)
reinstated Caryomys as a valid genus. This rearrangement
has been accepted by many systematists (Luo et al., 2000;
Wang, 2003; Musser and Carleton, 2005). The recognition
of subgenera also varies. Some systematists recognize
Anteliomys as a subgenus of Eothenomys (Ellerman and
Morrison-Scott, 1951; Ma and Jiang, 1996; Luo et al., 2000;
Ye et al., 2002; Wang, 2003; Musser and Carleton, 2005).
In contrast, Allen (1940) suggested that both Caryomys and
Anteliomys should be subgenera of Eothenomys, an
arrangement that is accepted by most systematists (Corbet,
1978; Honacki et al., 1982; Nowak and Paradiso, 1983;
Corbet and Hill, 1992; Musser and Carleton, 1993; Nowak,
1999).
Taxonomic uncertainty is common in Eothenomys (sensu
lato). To date, 25 nominal species have been included at
least once in this genus, yet today only between five and 12
species are recognized; others have been either regarded to
be synonyms or subspecies of other species, or transferred
to other genera (Table 1). The main controversies focus on
whether these are valid species and subspecies. For exam-
ple, Thomas (1921) described E. cachinus as a full species
and Luo et al. (2000) and Wang (2003) recognized it. How-
ever, most mammalogists consider the taxon to be a
subspecies or synonym of E. melanogaster (Hinton, 1923;
Ellerman, 1941; Ellerman and Morrison-Scott, 1951; Corbet,
1978; Musser and Carleton, 1993), or a synonym of E.
miletus (Corbet and Hill, 1992). Recently, Musser and
Carleton (2005) elevated it back to a species (Table 1). Sim-
ilarly, controversy exists as to whether E. miletus (Thomas,
* Corresponding author. Tel. : +86-28-83226632;
Fax : +86-28-83226547;
E-mail: [email protected]
Supplemental material for this article is available online.
doi:10.2108/zsj.29.610
Phylogeny of Oriental Voles 611
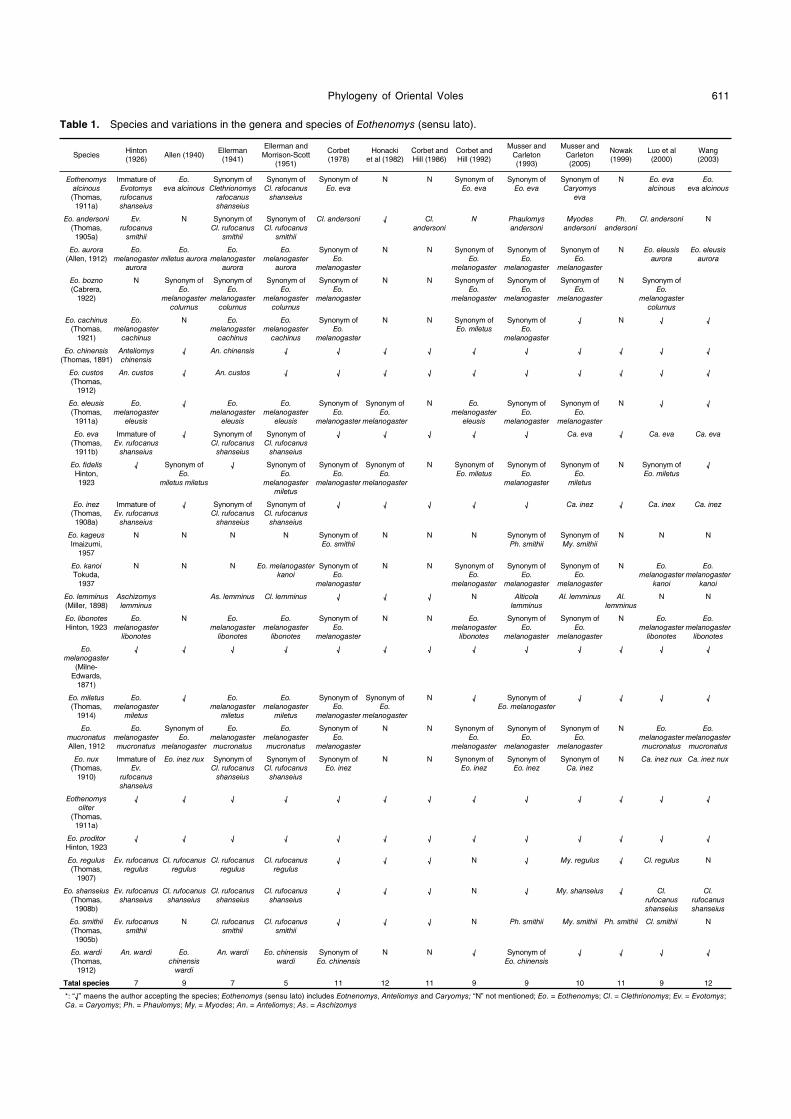
Table 1. Species and variations in the genera and species of Eothenomys (sensu lato).
SpeciesHinton(1926)
Allen (1940)Ellerman(1941)
Ellerman andMorrison-Scott
(1951)
Corbet(1978)
Honackiet al (1982)
Corbet andHill (1986)
Corbet andHill (1992)
Musser andCarleton(1993)
Musser andCarleton(2005)
Nowak(1999)
Luo et al(2000)
Wang(2003)
Eothenomysalcinous(Thomas,1911a)
Immature ofEvotomysrufocanusshanseius
Eo.eva alcinous
Synonym ofClethrionomys
rafocanusshanseius
Synonym ofCl. rafocanus
shanseius
Synonym ofEo. eva
N N Synonym ofEo. eva
Synonym ofEo. eva
Synonym ofCaryomys
eva
N Eo. evaalcinous
Eo.eva alcinous
Eo. andersoni(Thomas,1905a)
Ev.rufocanus
smithii
N Synonym ofCl. rufocanus
smithii
Synonym ofCl. rufocanus
smithii
Cl. andersoni √ Cl.andersoni
N Phaulomysandersoni
Myodesandersoni
Ph.andersoni
Cl. andersoni N
Eo. aurora(Allen, 1912)
Eo.melanogaster
aurora
Eo.miletus aurora
Eo.melanogaster
aurora
Eo.melanogaster
aurora
Synonym ofEo.
melanogaster
N N Synonym ofEo.
melanogaster
Synonym ofEo.
melanogaster
Synonym ofEo.
melanogaster
N Eo. eleusisaurora
Eo. eleusisaurora
Eo. bozno(Cabrera,
1922)
N Synonym ofEo.
melanogastercolurnus
Synonym ofEo.
melanogastercolurnus
Synonym ofEo.
melanogastercolurnus
Synonym ofEo.
melanogaster
N N Synonym ofEo.
melanogaster
Synonym ofEo.
melanogaster
Synonym ofEo.
melanogaster
N Synonym ofEo.
melanogastercolurnus
Eo. cachinus(Thomas,
1921)
Eo.melanogaster
cachinus
N Eo.melanogaster
cachinus
Eo.melanogaster
cachinus
Synonym ofEo.
melanogaster
N N Synonym ofEo. miletus
Synonym ofEo.
melanogaster
√ N √ √
Eo. chinensis(Thomas, 1891)
Anteliomyschinensis
√ An. chinensis √ √ √ √ √ √ √ √ √ √
Eo. custos(Thomas,
1912)
An. custos √ An. custos √ √ √ √ √ √ √ √ √ √
Eo. eleusis(Thomas,1911a)
Eo.melanogaster
eleusis
√ Eo.melanogaster
eleusis
Eo.melanogaster
eleusis
Synonym ofEo.
melanogaster
Synonym ofEo.
melanogaster
N Eo.melanogaster
eleusis
Synonym ofEo.
melanogaster
Synonym ofEo.
melanogaster
N √ √
Eo. eva(Thomas,1911b)
Immature ofEv. rufocanus
shanseius
√ Synonym ofCl. rufocanus
shanseius
Synonym ofCl. rufocanus
shanseius
√ √ √ √ √ Ca. eva √ Ca. eva Ca. eva
Eo. fidelisHinton,1923
√ Synonym ofEo.
miletus miletus
√ Synonym ofEo.
melanogastermiletus
Synonym ofEo.
melanogaster
Synonym ofEo.
melanogaster
N Synonym ofEo. miletus
Synonym ofEo.
melanogaster
Synonym ofEo.
miletus
N Synonym ofEo. miletus
√
Eo. inez(Thomas,1908a)
Immature ofEv. rufocanus
shanseius
√ Synonym ofCl. rufocanus
shanseius
Synonym ofCl. rufocanus
shanseius
√ √ √ √ √ Ca. inez √ Ca. inex Ca. inez
Eo. kageusImaizumi,
1957
N N N N Synonym ofEo. smithii
N N N Synonym ofPh. smithii
Synonym ofMy. smithii
N N N
Eo. kanoiTokuda,
1937
N N N Eo. melanogasterkanoi
Synonym ofEo.
melanogaster
N N Synonym ofEo.
melanogaster
Synonym ofEo.
melanogaster
Synonym ofEo.
melanogaster
N Eo.melanogaster
kanoi
Eo.melanogaster
kanoi
Eo. lemminus(Miller, 1898)
Aschizomyslemminus
As. lemminus Cl. lemminus √ √ √ N Alticolalemminus
Al. lemminus Al.lemminus
N N
Eo. libonotesHinton, 1923
Eo.melanogaster
libonotes
N Eo.melanogaster
libonotes
Eo.melanogaster
libonotes
Synonym ofEo.
melanogaster
N N Eo.melanogaster
libonotes
Synonym ofEo.
melanogaster
Synonym ofEo.
melanogaster
N Eo.melanogaster
libonotes
Eo.melanogaster
libonotes
Eo.melanogaster
(Milne-Edwards,
1871)
√ √ √ √ √ √ √ √ √ √ √ √ √
Eo. miletus(Thomas,
1914)
Eo.melanogaster
miletus
√ Eo.melanogaster
miletus
Eo.melanogaster
miletus
Synonym ofEo.
melanogaster
Synonym ofEo.
melanogaster
N √ Synonym ofEo. melanogaster
√ √ √ √
Eo.mucronatusAllen, 1912
Eo.melanogastermucronatus
Synonym ofEo.
melanogaster
Eo.melanogastermucronatus
Eo.melanogastermucronatus
Synonym ofEo.
melanogaster
N N Synonym ofEo.
melanogaster
Synonym ofEo.
melanogaster
Synonym ofEo.
melanogaster
N Eo.melanogastermucronatus
Eo.melanogastermucronatus
Eo. nux(Thomas,
1910)
Immature ofEv.
rufocanusshanseius
Eo. inez nux Synonym ofCl. rufocanus
shanseius
Synonym ofCl. rufocanus
shanseius
Synonym ofEo. inez
N N Synonym ofEo. inez
Synonym ofEo. inez
Synonym ofCa. inez
N Ca. inez nux Ca. inez nux
Eothenomysoliter
(Thomas,1911a)
√ √ √ √ √ √ √ √ √ √ √ √ √
Eo. proditorHinton, 1923
√ √ √ √ √ √ √ √ √ √ √ √ √
Eo. regulus(Thomas,
1907)
Ev. rufocanusregulus
Cl. rufocanusregulus
Cl. rufocanusregulus
Cl. rufocanusregulus
√ √ √ N √ My. regulus √ Cl. regulus N
Eo. shanseius(Thomas,1908b)
Ev. rufocanusshanseius
Cl. rufocanusshanseius
Cl. rufocanusshanseius
Cl. rufocanusshanseius
√ √ √ N √ My. shanseius √ Cl.rufocanusshanseius
Cl.rufocanusshanseius
Eo. smithii(Thomas,1905b)
Ev. rufocanussmithii
N Cl. rufocanussmithii
Cl. rufocanussmithii
√ √ √ N Ph. smithii My. smithii Ph. smithii Cl. smithii N
Eo. wardi(Thomas,
1912)
An. wardi Eo.chinensis
wardi
An. wardi Eo. chinensiswardi
Synonym ofEo. chinensis
N N √ Synonym ofEo. chinensis
√ √ √ √
Tatal species 7 9 7 5 11 12 11 9 9 10 11 9 12
*: “√” maens the author accepting the species; Eothenomys (sensu lato) includes Eotnenomys, Anteliomys and Caryomys; “N” not mentioned; Eo. = Eothenomys; Cl. = Clethrionomys; Ev. = Evotomys; Ca. = Caryomys; Ph. = Phaulomys; My. = Myodes; An. = Anteliomys; As. = Aschizomys

S. Liu et al.612
1914) is a subspecies or synonym of E. melanogaster
(Hinton, 1926; Ellerman and Morrison-Scott, 1951; Corbet,
1978; Honacki, 1982; Musser and Carleton, 1993), or a full
species (Allen, 1940; Corbet and Hill, 1992; Nowak, 1999;
Luo et al., 2000; Wang, 2003; Musser and Carleton, 2005)
(Table 1). Of course, these issues skirt the question of
whether or not any subspecies are valid taxa (Frost and
Hillis, 1990; Burbrink et al., 2000).
Most previous studies on the systematics of Eothenomys
(sensu lato) have focused on morphological comparisons,
particularly external morphology. Molecular analyses of the
group are rare. Yang et al. (1998) summarized all available
karyological data and discussed the putative evolutionary
relationships among the main lineages of the Clethrionomyini.
All taxa are diploid, generally with 54–56 chromosomes,
having a fundamental arm-number between 54 and 60
(excluding E. proditor). However, analyses of cytological
data provide discordant results, and no analyses unambigu-
ously elucidate the phylogenetic relationships within this
group. Luo et al. (2004) first explored the phylogeny of
Eothenomys (sensu lato) using nucleotide sequence data
from the mitochondrial DNA (mtDNA) cytochrome b gene
(cyt b). Their results suggest that Oriental voles (eight species
included) form a monophyletic group with two distinct mater-
nal lineages, the subgenera Eothenomys and Anteliomys.
Because Caryomys was not included in their study, the
validity of the genus, as well as the relationships among
Anteliomys, Caryomys, and Eothenomys, remain unre-
solved. Some recent molecular phylogenies include speci-
mens of Eothenomys (sensu lato), but they have not
focused on the genus itself (Buzan et al., 2008; Robovský
et al., 2008).
Herein we use nucleotide sequences from two mtDNA
genes—cytochrome oxidase subunit I (COI) and cytochrome
b (cyt b)—to explore the
evolutionary history and
systematics of Eothenomys
(sensu lato). We include a
morphological comparison
that includes all putative
species other than E.
wardi. Our aims are to: (1)
test whether Caryomys and
Anteliomys are monophyl-
etic genera; (2) hypothe-
size the phylogenetic rela-
tionships of species within
Caryomys, Anteliomys, and
Eothenomys; (3) test
whether cryptic taxa occur
in this group; and (4) test
the efficacy of DNA barcod-
ing for the group.
MATERIALS AND
METHODS
Samples and data collection
A total of 458 specimens
representing 11 species
(including 15 subspecies) and
two unidentified forms of
Eothenomys (sensu lato) were collected. All specimens have been
deposited in Sichuan Academy of Forestry. Morphological data
were obtained from 426 specimens (Supplementary Text S1
online). We sequenced 110 specimens for cyt b and the standard
DNA barcoding gene COI. The sampling localities were indicated in
Fig. 1 and detailed in Supplementary Table S1 online; eight repre-
sentatives of the allied genera (Myodes, Alticola, and Microtus) in
Cricetidae were included for de novo sequencing to evaluate the
monophyly of the genera.
DNA extraction, amplification, and sequencing
Total DNA was extracted from 95% ethanol-preserved liver or
muscle tissue using the standard proteinase K and phenol/chloroform
method (Sambrook and Russell, 2002). The entire cyt b and COI
genes were amplified using primers described by Burbrink et al.
(2000) and Delisle and Strobeck (2002), respectively. For cyt b, we
followed the cycling sequence protocol of Burbrink et al. (2000) but
with an annealing temperature of 56°C. Prior to sequencing, PCR
products were purified using a commercial kit (Watson BioMedical).
Double-stranded PCR products were directly sequenced from both
directions using an ABI 3730 Genetic Analyzer (Applied Biosystems)
following the manufacturer’s protocols.
Alignment of cyt b and COI was straightforward because indels
did not occur. The inadvertent amplification of pseudogenes (Zhang
and Hewitt, 1996) was checked by translating the sequences into
amino acids using MEGA 5 (Tamura et al., 2011) and aligning them
with published sequences. Kimura’s (1980) two-parameter genetic
distances (K2P) were calculated for the cyt b and COI data using
MEGA 5.
Matrilineal genealogy reconstruction
We used multiple methods to infer phylogenetic (matrilineal)
relationships, including maximum parsimony (MP), maximum likeli-
hood (ML), and Bayesian inference (BI). MP trees were obtained
using PAUP* 4.0b10 (Swofford, 2003) with a heuristic search using
1,000 random sequence addition replicates and TBR (tree bisection-
reconnection) branch swapping. Bootstrap support values (BS) for
lineages were calculated from 1,000 pseudoreplicates (Felsenstein,
Fig. 1. The collection localities of samples of voles sequenced in this work.

Phylogeny of Oriental Voles 613
1985). ML analyses were implemented in RAxML (Stamatakis et al.,
2008). For BI, the nucleotide sequences were partitioned by codon
position. Six partitions were given, one each for codon positions 1,
2, and 3 independently for both cyt b and COI. The best-fit models
of evolution for each partition were chosen using MrModeltest 2.3
(Huelsenbeck and Crandall, 1997; Posada and Crandall, 1998,
2001). BI was implemented using MrBayes 3.1 (Huelsenbeck and
Ronquist, 2001; Ronquist and Huelsenbeck, 2003). Three runs
were performed with four Markov chains (three heated chains and
one cold chain) starting from a random tree. Each of these runs was
conducted with a total of 5 million generations, and sampled every
100 generations. Stationarity was confirmed by inspecting plots of ln
(L) against generations using Tracer 1.3 (Rambaut and Drummond,
2003), and the first 1,000 generations were discarded as burn-in.
The frequencies of nodal resolution in the consensus tree were
termed posterior probabilities (PP).
As only 68 samples had sequence data from both genes, we
reconstructed matrilineal relationships using three different data-
sets: COI, cyt b, and a combined dataset (COI + cyt b). When mul-
tiple samples were sequenced from the same species, only unique
haplotypes were used to construct the trees.
Morphological analysis
A total of 426 individuals representing 11 species and two
unidentified forms of Eothenomys (sensu lato), including 15 subspe-
cies, were examined morphologically from the following taxa:
Eothenomys cachinus, E. chinensis chinensis, E. ch. tarquinius, E.
custos custos, E. cu. hintoni, E. cu. rubellus, E. eleusis eleusis, E.
el. aurora, E. eva eva, E. ev. alcinous, E. fidelis, E. inez inez, E. i.
nux, E. melanogaster chengduenus, E. me. melanogaster, E. miletus
miletus, E. olitor hypolitor, and E. proditor (Supplementary Text S1
online). Morphological structures of the teeth rows were scored.
Head plus body length, tail length, hind foot length, and greatest
length of skull were measured. External measurements were taken
in the field on freshly captured specimens and recorded to the near-
est 0.5 mm. Cranial measurements were taken with a vernier cali-
per to the nearest 0.02 mm. Morphological measurements followed
the protocol described by Liu et al. (2007). Measurements were
abbreviated as follows: HBL, head and body length, measuring from
snout to anus; TL, tail length; HFL, hind foot length excluding claws;
and SGL, skull greatest length.
Statistical analyses of the morphological data were performed
using SPSS v.12.0 for Windows (SPSS Inc., 1999). Descriptive sta-
tistics (mean, standard deviation, and observed range) were com-
puted for each species. Principal component analysis (PCA) was
used to obtain a general view of variation. We projected the most
informative factors of individuals to detect differentiation of morpho-
logical measurements between genera, subgenera, and species.
When the groups could not be clearly distinguished, t-tests were
used to determine if they differed statistically significantly while
assuming a priori a P < 0.05 to be significant.
Fieldwork was conducted following the animal care and use
guidelines of the American Society of Mammalogists (Gannon et al.,
2007).
RESULTS
Molecular data analysis
After alignment, a fragment of 1,539 bp was obtained for
COI from a total of 88 samples of Oriental voles. The frag-
ment was larger than the standard used for DNA barcoding
(Hebert et al., 2003). Seventy-five haplotypes were
screened de novo and 59 complete sequences were used
to hypothesize matrilineal relationships. Partial sequences
only were used in the combined datasets. Eight haplotypes
from the allied genera were also sequenced de novo. These
included Alticola stracheyi (one haplotype), Microtus fortis
(one), Myodes rufocanus (three), My. rutilis (two), and
Neodon irene (one). The following five additional sequences
were retrieved from GenBank as a part of outgroup based
on the study of Steppane et al. (2004): Hylaeamys yunganus
(Cricetidae: Sigmodontinae), Mesocricetus auratus
(Cricetidae: Cricetinae), Neacomys guianae (Cricetidae:
Sigmodontinae), Rattus exulans (Muridae: Murinae), and
Tatera indica (Muridae: Gerbillinae). For cyt b, up to a total
of 1,140 bp were obtained from each of the 99 samples suc-
cessfully sequenced de novo, and 81 unique haplotypes
were resolved, of which 69 complete sequences were used
to reconstruct matrilineal relationships. The remaining partial
fragments were used in the combined datasets only. Five
unique haplotypes of Myodes were also sequenced de
novo. From GenBank, we retrieved 16 fragments from
Oriental voles, 20 from the allied genera, and five from the
outgroup taxa. For the combined data, 68 Oriental voles had
both cyt b and COI gene sequences (de novo). Sequences
from either cyt b or COI only were available from 28 speci-
mens (de novo). Of six specimens of the allied genera
(Myodes and Alticola), five specimens of Myodes had
sequences from both cyt b and COI (de novo); one speci-
men of Alticola stracheyi had sequence data from COI only
(de novo). Sequences of cyt b only were retrieved from
GenBank for 20 samples of the allied genera. Two complete
outgroup sequences from the Muridae were retrieved from
GenBank. In our de novo sequences, no deletions, inser-
tions, or stop codons were found in either gene, indicating
that paralogous nuclear insertions had not been amplified.
Novel sequences have been deposited in GenBank
(GenBank accession numbers as follows: COI, HM165276–
HM165358; and cyt b, HM165359–HM165444) (Supple-
mentary Table S1 online).
Sequence divergence
Average K2P genetic distances for the standard barcod-
ing gene, COI, were generally sufficient to diagnose species
(Table S2) and lineages (not given in table). Within
Eothenomys, average interspecific divergences ranged from
0.011 to 0.086 and inter-individual differences averaged
0.056. Levels of divergence were greater than those values
within a new subgenus (erected below), where they ranged
from 0.020 to 0.052 and inter-individual differences aver-
aged 0.027. Clade Anteliomys had the highest average level
of interspecific divergence (0.106–0.134) and inter-individual
differences averaged 0.087. Within Eothenomys, the bar-
codes did not always unambiguously identify morphological
taxa. In contrast, sequence divergence for COI between
unambiguously identified the two species of Caryomys
(average divergence 0.066) and species within Myodes
(average divergence 0.134). Levels of divergence for cyt b
were generally equivalent to those of COI (Table S2).
Phylogenetic/matrilineal analyses
The average base frequencies for COI were as follows:
T = 26.6%, C = 29.4%, A = 34.6%, and G = 9.4%; for cyt b
T = 23.5%, C = 33.6%, A = 36.6%, and G = 6.3%. A total of
502 nucleotide sites (33%) were variable in COI, of which 454
(29%) were potentially parsimony-informative when including
the outgroup taxa. For cyt b, 445 sites (39%) were variable
and 393 (34%) were potentially parsimony-informative when

S. Liu et al.614
including the outgroup taxa.
MP analysis of the COI sequences produced two most
parsimonious trees of 1,595 steps (CI = 0.433, RI = 0.859,
RC = 0.372). MrModeltest was employed to identify the
following models of sequence evolution in the BI and ML
analyses of the partitioned data under the AIC criterion:
GTR + I + Г for the first codon position, HKY for the second,
and GTR + Г for the third.
For cyt b, MP resolved 45133 most parsimonious trees
of 2,783 steps with CI = 0.309, RI = 0.767, RC = 0.237. For
Fig. 2. Fifty percent majority rule consensus tree from a Bayesian analysis of cyt b nucleotide sequence data. Numbers represent nodal sup-
port of key nodes inferred from Bayesian posterior probability, ML bootstrap, and MP bootstrap analysis, respectively. Asterisks indicate nodes not supported by the MP analysis (< 50). Morphological correlates are as follows: triangles of the first lower molar of Caryomys interlaced; trian-gles of the first lower molar of the Eothenomys fuse transversely; the first upper molar of subgenus Eothenomys has four inner angles; the first upper molar of subgenera Anteliomys and Ermites has three inner angles; structure of the third upper molar of subgenus Anteliomys non-variable; and structure of the third upper molar of subgenus Ermites multivariable.

Phylogeny of Oriental Voles 615
the BI and ML analyses, the models SYM + I + Г, HKY + I,
and GTR + I + Г were selected for three codon positions,
respectively, under the AIC criterion.
Despite the different datasets and methods of tree con-
struction, all generated trees were largely congruent. Minor
disagreements were associated with poorly supported
nodes (Figs. 2–4).
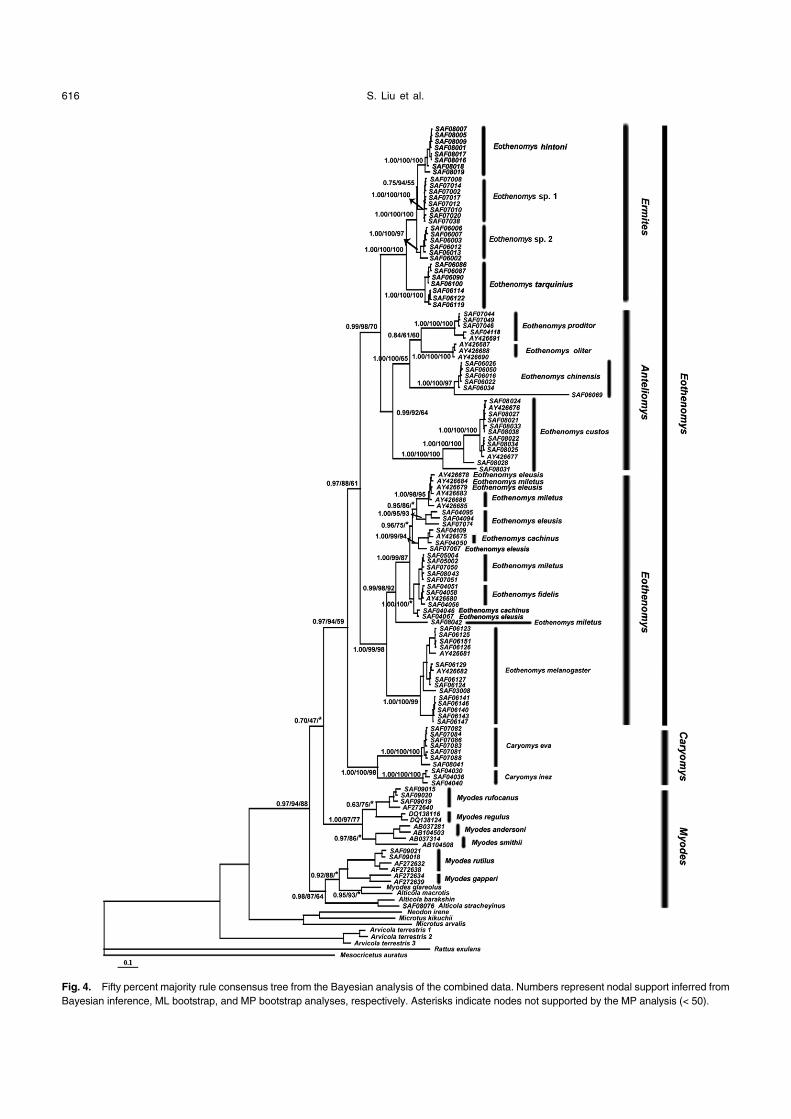
Analyses of the sequence data always resolved a mono-
phyletic Eothenomys (sensu lato) and frequently with strong
support. In the BI analyses of the cyt b and combined data-
sets, PP = 100% and PP = 97%, respectively. The ML anal-
yses for COI and the combined datasets obtained BS = 77
and BS = 94, respectively. In the MP tree for COI, BS = 90.
Four maternal lineages of Oriental voles were recovered.
Three of those were accompanied with high support values
in all analyses, corresponding to Eothenomys, Caryomys,
and a new taxon. The fourth lineage, Anteliomys, was
strongly supported in the BI analyses. However, for this lin-
eage, the ML and MP analyses for COI had very low support
values (BS < 50%), the ML analysis for cyt b obtained mod-
erate support (BS = 74%), and the MP analysis for the com-
bined dataset had low support (BS = 64%) (Figs. 2–4).
The sister-group relationship between the new taxon
and Anteliomys was strongly supported with the exception of
the MP tree for the combined dataset (BS = 70; Fig. 4). The
sister-group relationship of Eothenomys with the new taxon
and Anteliomys was highly supported in both the BI and ML
trees for COI data (PP = 100%; BS = 93%) and the com-
bined dataset (PP = 99%; BS = 98%), but the results from
cyt b showed low support values in all three tree construc-
tion methods (Fig. 2).
The major lineages contained the same species
irrespective of method of analysis and dataset. In most
cases, the sublineages, including samples from the same
Fig. 3. Fifty percent majority rule consensus tree from a Bayesian analysis of COI nucleotide sequence data. Numbers are nodal support val-
ues for key nodes inferred from Bayesian posterior probability, ML bootstrap, and MP bootstrap analyses, respectively. Asterisks indicate
nodes not supported by the MP analysis (< 50).
S. Liu et al.616
Fig. 4. Fifty percent majority rule consensus tree from the Bayesian analysis of the combined data. Numbers represent nodal support inferred from
Bayesian inference, ML bootstrap, and MP bootstrap analyses, respectively. Asterisks indicate nodes not supported by the MP analysis (< 50).

Phylogeny of Oriental Voles 617
species, received strong support values. Most of the mor-
phologically identified subspecies correctly clustered in their
species. However, E. eleusis eleusis, E. el. aurora and E. el.
yinyjiangensis did not cluster together. The new taxon con-
tained two putative subspecies (E. chinensis tarquinius and
E. custos hintoni) and two undescribed forms. The former
two subspecies were distantly related to their congeners E.
ch. chinensis and E. cu. custos, respectively. Anteliomys
contained E. proditor, E. custos, E. chinensis, and E. olitor.
Eothenomys contained five species: Eothenomys cachinus,
E. eleusis, E. fidelis, E. melanogaster, and E.
miletus. Notably, three species (E. cachinus,
E. eleusis, and E. miletus) nested together,
but these were not resolved as monophyl-
etic (monogenealogical sensu Murphy and
Méndez de la Cruz, 2010) species. Mono-
phyletic Caryomys was comprised of E. inez
and E. eva.
Morphological groups
The Oriental voles were divided into
several morphological groups. (1) They
formed two groups based on the condition of
the first lower molar: Caryomys (Fig. 5A)
was distinguished from Eothenomys,
Anteliomys, and the new taxon by having
triangles of the first lower molar arranged
alternately; the triangles of the first lower
molar were confluent transversely in
Eothenomys, Anteliomys, and the new taxon
(Fig. 5B). (2) The first upper molar of
Eothenomys had four inner angles (Fig. 5C),
whereas that of Anteliomys and the new
taxon had three inner angles (Fig. 5D). (3)
The third upper molar of the new taxon
(including two previously described subspe-
cies, E. chinensis tarquinius and E. custos
hintoni, as well as E. sp. 1 and E. sp. 2) was
complex and variable (Fig. 5E1–3, F1–3, G1–
4), whereas that of Anteliomys was either
complex or simple, but without variation (Fig. 5H–K). (4)
Three species-groups were defined based on the proportion
of TL to HBL. In the first group, the tail was more than half
of the HBL. This group included Eothenomys chinensis
chinensis, E. ch. tarquinius, E. custos hintoni, and E. eva. The
second group consisted of Eothenomys custos, E. eleusis, E.
fidelis, E. inez, E. melanogaster, E. miletus, E. olitor, and E.
proditor. Here, TL was less than half of HBL. The last group
included the two unidentified forms. The TL was equal to half
of HBL (Table 2). (5) Three taxa (E. chinensis chinensis, E.
Fig. 5. Teeth from the right side of Oriental voles. (A) first lower molar of Caryomys (C.
eva); (B) first lower molar of genus Eothenomys (E. melanogaster); (C) first upper molar of
subgenus Eothenomys (E. melanogaster); (D) first upper molar of subgenera Anteliomys
and Ermites (E. custos); (E1–3) third upper molar of E. tarquinius; (F1–3) third upper
molar of E. hintoni; (G1–4) third upper molar of E. sp. 1; (H) third upper molar of E.
chinensis; (I) third upper molar of E. custos; (J) third upper molar of E. proditor; (K) third
upper molar of E. olitor.
Table 2. Morphological comparison of Oriental voles. Length values are shown as mean with range in parentheses (mm).
genera subgenera species HBL TL HTL TL/HBL SGL n 1st lower molar1st upper
molar3rd upper molar
CaryomysC. eva 87.73 (75–98) 51.61 (43–61) 16.36 (15–19) 0.59 22.98 (21.00–25.70) 33 Triangles of the
first lower rangealternately
With 3 innerand 3 outer
angles
With 3 innerand 3
outer anglesC. inez 92.14 (80–99) 34.86 (30–38) 15.46 (13–18) 0.38 23.79 (22.10–25.40) 14
Eothenomys
Eothenomys
E. melanogaster 97.92 (80–110) 37.10 (28.5–52)16.16 (14–19) 0.38 24.51 (21.76–26.08) 55
Triangles of thefirst lower molarwere confluenttransversely
With 4 innerangles
with 3–4 inner and3–4 outer angles.
Structures ofintraspecies
steady
E. fidelis 110.22 (98–121) 42.39 (38–49) 17.94 (15–19) 0.38 27.47 (26.56–28.30) 18
E. miletus 103.33 (91–115) 42.17 (36–51) 17.06 (15–19) 0.41 26.06 (24.42–28.42) 25
E. cachinus 97.67 (78–112) 39.08 (34–46) 17.79 (15–20) 0.40 25.17 (24.42–26.18) 13
E. eleusis 99.42 (82–112) 42.16 (36–50) 17.20 (13–21) 0.42 24.75 (22.30–28.28) 19
Anteliomys
E. chinensis 121.66 (102–134)64.12 (57–71) 22.41 (21–25) 0.53 29.92 (27.52–31.14) 82
With 3 innerangles
with 3–5 inner and3–5 outer angles,
structure ofintraspeciesmolar steady
E. custos 99.04 (79–110) 37.42 (32–50) 17.06 (15–20) 0.38 24.43 (22.76–25.96) 26
E. proditor 96.57 (90–110) 36.57 (35–40) 18.00 (15–20) 0.38 25.77 (24.62–27.46) 7
E. olitor 86.75 (86–88) 27.50 (26–29) 14.25 (14–15) 0.32 20.09 (21.78–22.40) 4
Ermites
E. tarquinius 114.31 (104–125)66.63 (62–75) 22.40 (18–24) 0.58 27.37 (26.10–28.72) 48
With 3inner angles
With 4–6 innerand 4–6 outer
angles,structure ofintraspeciescomplex andmultivariate
E. hintoni 98.79 (92–105) 56.11 (52–60) 18.00 (16–19) 0.57 24.39 (23.52–25.62) 19
E. sp. 1 96.27 (86–110) 46.21 (42–53) 17.02 (16–20) 0.48 22.91 (21.74–24.40) 42
E.sp. 2 97.19 (87–110) 47.24 (39–55) 17.38 (16–19) 0.49 23.44 (22.56–24.40) 22
HBL: head and body length; TL: tail length; HFL: hind foot length; SGL: skull greatest length; n: numbers of specimens.
S. Liu et al.618
ch. tarquinius, and E. fidelis) had
the largest HBL (> 110 mm),
nine taxa (E. cachinus, E. custos
hintoni, E. custos [E. cu. custos
and E. cu. rubellus], E. eleusis,
E. melanogaster, E. miletus, E.
proditor, and the two unidenti-
fied forms) had medium HBLs
(averages range from 95–105
mm), and the remaining taxa had
HBL < 95 mm (E. eva, E. olitor,
and E. inez). A detailed compar-
ison of the morphology was given
in Table 2. Based on morpholog-
ical characteristics, Eothenomys
chinensis chinensis, E. ch.
tarquinius, E. custos custos, E.
cu. hintoni, E. eva, E. inez, E.
olitor, and the two unidentified
forms were identified unambigu-
ously. All other subspecies were
correctly identified to their spe-
cies. However, Eothenomys
eleusis, E. fidelis, and E. miletus
could be distinguished by SGL
measurements only.
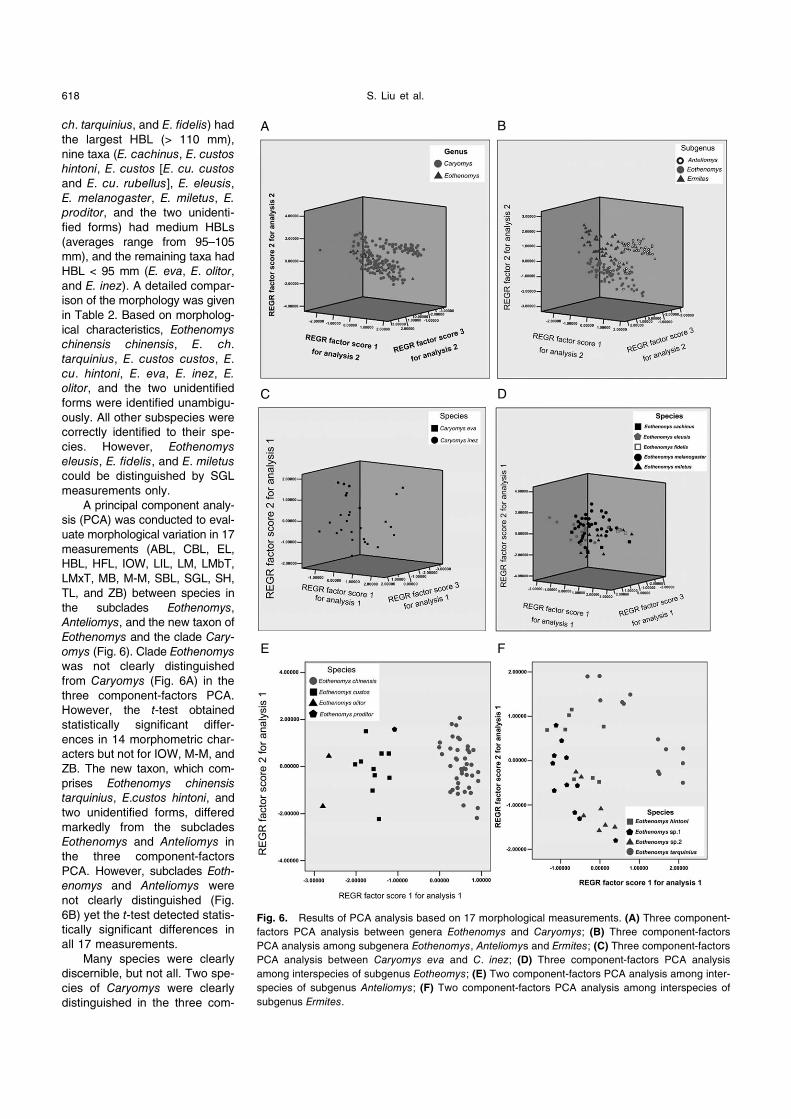
A principal component analy-
sis (PCA) was conducted to eval-
uate morphological variation in 17
measurements (ABL, CBL, EL,
HBL, HFL, IOW, LIL, LM, LMbT,
LMxT, MB, M-M, SBL, SGL, SH,
TL, and ZB) between species in
the subclades Eothenomys,
Anteliomys, and the new taxon of
Eothenomys and the clade Cary-
omys (Fig. 6). Clade Eothenomys
was not clearly distinguished
from Caryomys (Fig. 6A) in the
three component-factors PCA.
However, the t-test obtained
statistically significant differ-
ences in 14 morphometric char-
acters but not for IOW, M-M, and
ZB. The new taxon, which com-
prises Eothenomys chinensis
tarquinius, E.custos hintoni, and
two unidentified forms, differed
markedly from the subclades
Eothenomys and Anteliomys in
the three component-factors
PCA. However, subclades Eoth-
enomys and Anteliomys were
not clearly distinguished (Fig.
6B) yet the t-test detected statis-
tically significant differences in
all 17 measurements.
Many species were clearly
discernible, but not all. Two spe-
cies of Caryomys were clearly
distinguished in the three com-
A B
C D
E F
Fig. 6. Results of PCA analysis based on 17 morphological measurements. (A) Three component-
factors PCA analysis between genera Eothenomys and Caryomys; (B) Three component-factors
PCA analysis among subgenera Eothenomys, Anteliomys and Ermites; (C) Three component-factors
PCA analysis between Caryomys eva and C. inez; (D) Three component-factors PCA analysis
among interspecies of subgenus Eotheomys; (E) Two component-factors PCA analysis among inter-
species of subgenus Anteliomys; (F) Two component-factors PCA analysis among interspecies of
subgenus Ermites.

Phylogeny of Oriental Voles 619
ponent-factors PCA (Fig. 6C). In contrast, within subclade
Eothenomys, five species were not clearly distinguished in
the three component-factors PCA (Fig. 6D). The two com-
ponent-factors PCA clearly distinguished the following four
species in subclade Anteliomys: Eothenomys chinensis, E.
custos, E. olitor, and E. proditor (Fig. 6E). Within the new
taxon, Eothenomys chinensis tarquinius was clearly distin-
guished from the others. Eothenomys custos hintoni and the
other two unidentified forms were slightly differentiated by
two component-factors in the PCA (Fig. 6F). The two
unidentified forms differed statistically significantly by five
measurements: EL, LM, M-M, SH, and ZB.
The molecular and morphological results yielded two pri-
mary conclusions. First, Cayomys and Eothenomys are valid
genera based on the condition of the lower molar and the
molecular phylogeny. The latter genus included three sub-
genera: Eothenomys, Anteliomys, and a new subgenus.
Second, although matrilineal genealogies did not reject the
null hypothesis of panmixia, the historical relationships sug-
gest the existence of species. Our trees suggested that
Eothenomys chinensis tarquinius and E. custos hintoni were
valid taxa based on historical relationships and the extent of
lineage divergence. Our morphological results clearly diag-
nosed these taxa, and suggest the absence of gene flow.
Taken together, the genealogy and morphological analyses
served to reject the null hypothesis of conspecificity of E.
chinensis tarquinius and E. ch. chinensis as well as E.
custos hintoni and E. cu. custos. Therefore, we regarded E.
tarquinius and E. hintoni as distinct species. Further, the two
unidentified forms appeared to be undescribed species. The
lineage that included E. hintoni, E. tarquinius, and two
unidentified species formed a morphologically distinct mono-
phyletic group. These taxa could have been assigned to the
subgenus Anteliomys yet recognition of a new subgenus for-
mally united these taxa and the corresponding taxonomy
more finely reflected phyletic history. Thus, we found it
desirable to erect a new subgenus, whose diagnostic traits
and content are as follows:
Systematic account
Family: Cricetidae
Subfamily: Arvicolinae
Genus: Eothenomys
Subgenus: Ermites Liu, Liu, Guo, Sun, Murphy, Fan, Fu
and Zhang subgen. nov.
Type species: Eothenomys hintoni (as published in the
trinomen Eothenomys custos hintoni Osgood, 1932; herein
elevated to the rank of species)
Etymology.—The subgeneric name, masculine in gen-
der, is a patronym honoring Professor and Academician
Ermi Zhao, an internationally recognized herpetologist and
our teacher.
Diagnosis.—The palate is typical of Eothenomys. The
teeth are slender. Average skull length is less than 27.5 mm,
average head and body length is less than 115 mm. The tri-
angle of the first lower molar is confluent transversely. The
second upper molar lacks a second inner triangle. The third
upper molar is very complex, multivariate, and has more
than four inner and four outer angles. The tail is longer than
or equal to half of the body and head length.
This subgenus includes four species: Eothenomys
hintoni (= Eothenomys custos hintoni), E. tarquinius (=
Eothenomys chinensis tarquinius), and two undescribed
species (Figs. 2–4; E. sp. 1 and E. sp. 2).
Systematics of Oriental voles
Combined, the morphology of the first lower molar and
the matrilineal genealogy resolved the taxonomy of Oriental
voles as follows:
Caryomys Thomas, 1911
Caryomys inez (Thomas, 1908) [type species]
Caryomys eva (Thomas, 1911)
Eothenomys Miller, 1896
E. (Anteliomys)
E. (A.) chinensis (Thomas, 1891) [type species]
E. (A.) olitor (Thomas, 1911)
E. (A.) custos (Thomas, 1912)
E. (A.) proditor Hinton, 1923
E. (Ermites) subgen. nov.
E. (Er.) tarquinius (Thomas, 1912)
E. (Er.) hintoni Osgood, 1932 [type species]
E. (Er.) sp. 1
E. (Er.) sp. 2
E. (Eothenomys) Miller, 1896
E. (Eo.) melanogaster (Milne-Edwards, 1871) [type
species]
E. (Eo.) eleusis (Thomas, 1911) [species inquirenda]
E. (Eo.) miletus (Thomas, 1914) [species inquirenda]
E. (Eo.) cachinus (Thomas, 1921) [species inquirenda]
E. (Eo.) fidelis Hinton, 1923 [species inquirenda]
Because of a lack of specimens, the status of Eothenomys
wardi was not investigated.
DISCUSSION
The different datasets and methods of tree construction
yield a highly resolved, robust matrilineal genealogy and we
take this to be reflective of the phylogeny. Most nodes are
highly supported. In most analyses, the Oriental voles clus-
ter together with strong support. Morphological structure cor-
responds with the matrilineal genealogy.
Historically, Oriental and Japanese red-backed voles
are placed in the genus Eothenomys (sensu lato) (Corbet,
1978; Corbet and Hill, 1986). Using a molecular phylogeny,
Luo et al. (2004) suggested that Japanese red-backed voles
should be removed from Eothenomys and placed in the
genus Phaulomys, as initially defined by Thomas (1905b).
Subsequently, Musser and Carleton (2005) assigned
Japanese red-backed voles to Myodes. A detailed discus-
sion of the classification of Japanese red-backed voles is
beyond the scope of our work, but our results strongly sup-
port the exclusion of Japanese red-backed voles from the
Oriental voles. Additionally, our molecular phylogeny sug-
gests that the genus Myodes, as defined presently, is not a
monophyletic group. Here too, further work is needed.
The systematics of Oriental voles is fraught with differing
opinions. Validity of the genera and subgenera remains con-
tentious. Luo et al. (2004) considered Eothenomys and
Anteliomys as subgenera of genus Eothenomys. Although
their work did not include Caryomys, they recognized the
genus and suggested it might be the sister-group of
Eothenomys. Our analysis, with more extensive samples
and data from two genes, resolves the historical associa-

S. Liu et al.620
tions of the Oriental voles and confirms their prediction. The
group contains four distinct lineages and the nodes generally
receive high support values. These four lineages correspond
to the genera Caryomys and Eothenomys (sensu stricto), the
latter with the subgenera Eothenomys, Anteliomys, and
Ermites. Morphologically, all species of Caryomys possess
inter-bedded molar triangles on the first and second lower
molars; in Eothenomys, triangles on the first lower molar are
confluent transversely. The upper molars of subgenus
Eothenomys have three outer and four inner salient angles
and the first upper molars of subgenus Anteliomys have
three outer and three inner salient angles. In both subgen-
era, TL is less than half of HBL, except for E. chinensis. In
this exception, TL is greater than 60 mm, more than half of
the HBL, and the skull length averages more than 29 mm.
Finally, subgenus Ermites is diagnosed by the following
characteristics: first upper molars have three outer and three
inner salient angles, but the third upper molar is variable but
has more than four inner and four outer angles; TL is equal
to or longer than half of HBL; average skull length is less
than 27.5 mm; and HBL averages less than 115 mm (Table
2).
The interspecific relationships of subgenus Ermites are
well resolved. Traditionally, E. hintoni and E. tarquinius are
assigned to subgenus Anteliomys, but as subspecies of
either Eothenomys custos or E. chinensis (Ellerman and
Morrison-Scott, 1951; Corbet, 1978; Honacki et al., 1982;
Corbet and Hill, 1986; Musser and Carleton, 1993, 2005;
Nowak, 1999; Luo et al., 2000; Wang, 2003). However, our
genealogy does not cluster E. hintoni and E. tarquinius
together with E. custos and E. chinensis, respectively, and
they appear in different lineages (Figs. 2–4). Consequently,
we recognize E. hintoni and E. tarquinius as valid species
and assign them to subgenus Ermites.
Subgenus Anteliomys has a strongly supported sister
relationship with Ermites, in most cases. Anteliomys con-
tains four species. Individuals of the same species cluster
together with strong support and the interspecific relation-
ships are also well resolved. Yang et al. (1998) concluded
that karyologically the Yulong vole (E. proditor) substantially
differs from allied voles and they suggested further study
was required to determine the status of E. olitor and E.
proditor. Our molecular analyses obtain very high support
values for the group. Traditionally, E. wardi is placed in sub-
genus Anteliomys. Some systematists recognize this taxon
as a subspecies of E. chinensis (Allen, 1940; Ellerman and
Morrison-Scott, 1951; Corbet, 1978; Musser and Carleton,
1993), but others regard it as a valid species (Hinton, 1926;
Ellerman, 1941; Corbet and Hill, 1992; Kaneko, 1996;
Nowak, 1999; Luo et al., 2000; Wang, 2003; Musser and
Carleton, 2005). Unfortunately, the absence of samples of E.
wardi precludes us from making any taxonomic inference.
Subgenus Eothenomys is the most complex group of
Oriental voles. Often, this subgenus is considered to contain
five species: E. melanogaster, E. cachinus, E. eleusis, E.
fidelis, and E. miletus. Frequently the latter four taxa are
considered to be subspecies of E. melanogaster (Ellerman
and Morrison-Scott, 1951; Corbet, 1978; Honacki et al.,
1982; Corbet and Hill, 1986; Musser and Carleton, 1993).
Nowak (1999) accorded E. melanogaster and E. miletus
species status. Musser and Carleton (2005) recognized four
species, and Wang (2003) accepted five species. Our molec-
ular analyses unite four species (E. cachinus, E. eleusis, E.
fidelis, and E. miletus) exclusive of E. melanogaster.
Whereas all individuals of E. melanogaster and E. fidelis
cluster together as discrete lineages, and while noting that
E. fidelis has a very small sample size, individuals of E.
cachinus, E. eleusis, and E. miletus are intermixed in the
matrilineal genealogy (Figs. 2–4). Incomplete lineage sort-
ing, historical hybridization, and potential conspecificity all
provide possible explanations for the pattern. In order to test
these possibilities, further study involving more intensive
sampling and nuclear genes is required. Irrespective of the
above, our study does not support the proposals of Musser
and Carleton (2005), which recognize eleusis as a subspe-
cies of E. melanogaster.
Genus Caryomys is comprised of two species: C. inez
and C. eva. Recognition of these two species is well sup-
ported.
DNA barcoding reflects the morphological results. Spe-
cies in the subgenera Ermites and Anteliomys, and genus
Caryomys, are unambiguously identified. Within subgenus
Eothenomys, DNA barcoding also fails to segregate E.
cachinus and E. eleusis; the inability to sequence multiple
specimens of E. miletus precludes a comparison of the COI
and cyt b data, in which the latter gene fails to diagnose this
taxon. Regardless, DNA barcoding appears to be an effi-
cient approach to identifying unambiguous species as well
as being an efficient way to discern where additional
research is required.
In China, most species of Oriental voles are endemic to
Yunnan and Sichuan. They occur in the southern part of the
Hengduan Mountains. This region was severely affected by
several orogenic events associated with the Qinghai-Tibetan
Plateau (Luo et al., 2004). The most intense and frequent
orogenic events, known as the Qingzang Movement (Zheng
et al., 2000; An et al., 2001), are dated at between 3.6 and
1.7 million years ago (Ma). Subsequent events, termed the
Kun-Huang Movement (1.2–0.6 Ma; Jing et al., 2007), are
responsible for further shaping the geomorphology of this
area. These geological events likely isolated populations of
voles and in doing so drove speciation. Geological pro-
cesses greatly influence the divergence of species endemic
to this region (Luo et al., 2004; Yang et al., 2006; Zhang and
Jiang, 2006; Jin et al., 2008; Fan et al., 2009, 2011; Zhang et
al., 2010). Among Oriental voles, the subgenera Eothenomys
and Ermites occur in this region only. The Gonggashan
Mountains—the southern Hengduan Mountains in western
Sichuan: highest peak 7556 m—has the greatest concentra-
tion of Oriental voles with six species: Eothenomys
melanogaster, E. cachinus, E. eleusis, E. miletus, E. hintoni,
and E. tarquinius.
The oldest fossil of E. melanogaster is dated at 2.03 Ma
(Zheng, 1993), E. chinensis is dated at 1.8 Ma (Zheng,
1993), and Caryomys eva and C. inez are dated at 0.5 Ma
(Middle or Late Pleistocene) (Li and Xue, 1996). Caryomys
roots at the base of the genealogy yet the fossils are far
more recent than those of Eothenomys. Zheng (1993) pro-
poses that E. melanogaster lay at the center of diversifica-
tion of Oriental voles because extant specimens appear to
be identical to those from the Pleistocene. Our results do not
support his proposal because Oriental voles form the sister-

Phylogeny of Oriental Voles 621
group of Caryomys. However, the absence of fossil series
precludes a determination of the initial date of origin.
In order to identify the Oriental voles, a key to the gen-
era and subgenera is given as follows:
A. Triangles of the first lower molar ranged alternately .
........................................................................ Caryomys
B. Triangles of the first lower molar are confluent trans-
versely ..........................................................Eothenomys
a. The first upper molar with four inner triangles .....
.............................................. Subgenus Eothenomys
b. The first upper molar with three inner triangles
(a) Tail shorter or longer than half of the head and
body length (if tail is longer than half of the head and body
length, the body is very large, average head and body
length 120 mm, average skull length larger than 29 mm),
structure of the third upper molar non-variable ....................
............................................................ Subgenus Anteliomys
(b) Tail equal to or longer than half of the head and
body length (if tail is longer than half of the head and body
length, the body is medium, average head and body length
less than 115 mm, average skull length smaller than 27.5 mm),
structure of the third upper molar complex and variable......
..................................................................Subgenus Ermites
ACKNOWLEDGMENTS
This research was funded by the National Natural Science
Foundation of China (NSFC 30970330) and National Basic
Research Program of China (973 Project: 2007 CB109106) and to
R.W.M. by a Visiting Professorship for Senior International Scientists
from the Chinese Academy of Sciences and a NSERC Discovery
Grant (3148). R. Johnston (University of California, Los Angeles)
provided editorial assistance and R. Liao and J. Zhao assisted in
fieldwork. We thank S. Huang (Huangshan University) for help in
analyzing molecular data. Special thanks to J. H. Bai for drawing
the skulls and X. L. Jiang (Kunming Institute of Zoology) for provid-
ing some specimens.
REFERENCES
Allen GM (1912) Some Chinese vertebrates. Mammalia. Mem Mus
Comp Zool Harvard College 59: 1–14
Allen GM (1924) Microtinae collection by the American Museum
Asian Expeditions. Amer Mus Novi 133: 1–13
Allen GM (1940) The Mammals of China and Mongolia. Natural
History of Central Asia. Vol. XI. The American Museum of Natu-
ral History, New York
An Z, John EK, Warren LP, Stephen CP (2001) Evolution of Asian
monsoons and phased uplift of the Himalaya-Tibetan plateau
since Late Miocene times. Nature 411: 62–66
Burbrink FT, Lawson R, Slowinski JB (2000) Mitochondrial DNA
phylogeography of the polytypic North American rat snake
(Elaphe obsoleta): a critique of the subspecies concept. Evolu-
tion 54: 2107–2118
Buzan EV, Krystufek B, Hänfling B, Hutchinson WF (2008) Mito-
chondrial phylogeny of Arvicolinae using comprehensive taxo-
nomic sampling yields new insights. Biol J Linn Soc 94: 825–
835
Cabrera A (1922) Sobre algunos mammiferos de la China oriental.
Bol R Soc Esp Hist Nat 22: 162–170
Corbet GB (1978) The Mammals of the Palaearctic Region: A Taxo-
nomic Review. British Museum (Natural History), London and
Cornell University Press, Ithaca, NY
Corbet GB, Hill JE (1986) A world list of mammalian species. Sec-
ond edition. British Museum (Natural History), London
Corbet GB, Hill JE (1992) Mammals of Indomalayan Region. A Sys-
tematic Review. Oxford University Press, Oxford
Delisle I, Strobeck C (2002) Conserved primers for rapid sequenc-
ing of the complete mitochondrial genome from carnivores,
applied to three species of bears. Mol Biol Evol 19: 358–361
Ellerman JR (1941) The Families and Genera of Living Rodents.
British Museum (Natural History), London
Ellerman JR, Morrison-Scott TCS (1951) Checklist of Palaearctic
and Indian Mammals 1758 to 1946. British Museum (Natural
History), London
Fan ZX, Liu SY, Liu Y, Zeng B, Zhang XY, Cuo C, et al. (2009)
Molecular phylogeny and taxonomic reconsideration of the sub-
family Zapodiane (Rodentia: Dipodidae), with an emphasis on
Chinese species. Mol Phylogenet Evol 51: 447–453
Fan ZX, Liu SY, Liu Y, Zhang XY, Yue BS (2011) How Quaternary
geologic and climatic events in the southeastern margin of the
Tibetan Plateau influence the genetic structure of small mam-
mals: inferences from phylogeography of two rodents, Neodon
irene and Apodemus latronum. Genetica 139: 339–351
Felsenstein J (1985) Confidence limits on phylogenies: an approach
using the bootstrap. Evolution 39: 783–791
Frost DR, Hillis DM (1990) Species concepts in practice: herpetolog-
ical applications. Herpetologica 46: 115–128
Gannon WL, Sikes RS, the Animal Care and Use Committee of the
American Society of Mammalogists (2007) Guidelines of the
American Society of Mammalogists for the use of wild mam-
mals in research. J Mammal 88: 809–823
Gromov IM, Polyakov IYa (1977) Fauna of the USSR Mammals,
Vol. III, No. 8, Voles (Microtinae). English Edition: 1992, Douglas
Siegel-Causey and Robert S. Hoffmann. Smithsonian Institution
Libraries and the National Science Foundation, Washington DC
Hebert PDN, Cywinska A, Ball SL, deWaard JR (2003) Biological
identifications through DNA barcodes. Proc Royal Soc London
B 270: 313–721
Hinton MAC (1923) On the voles collected by Mr. G. Forrest in
Yunnan, with remarks upon the genera Eothenomys and
Neodon and upon their allies. Ann Mag Nat Hist, Ser. 9, 6: 146–
163
Hinton MAC (1926) Monograph of the Voles and Lemmings
(Microtinae) Living and Extinct. British Museum (Natural History),
London
Honacki JH, Kinman KE, Koeppl JW (1982) Mammal Species of the
World. Published as a Joint Venture of Allen Press, Inc. and the
Association of Systematics Collection, Lawrence, Kansas
Huelsenbeck JP, Crandall KA (1997) Phylogeny estimation and
hypothesis testing using maximum likelihood. Ann Rev Ecol
Syst 28: 437–466
Huelsenbeck JP, Ronquist FR (2001) MrBayes: Bayesian inference
of phylogenetic trees. Bioinformatics 17: 754–755
Imaizumi Y (1957) Taxonomic studies on the red-backed vole from
Japan. Pt. 1, major divisions of the vole and description of
Eothenomys with a new species. Bull Nat Sci Mus 3: 195–216
Jin YT, Brown RP, Liu NF (2008) Cladogenesis and phylogeography
of the lizard Phrynocephalus vlangalii (Agamidae) on the
Tibetan plateau. Mol Ecol 17: 1971–1982
Jing MD, Yu HT, Wu SH, Wang W, Zheng XG (2007) Phylogenetic
relationships in genus Niviventer (Rodentia: Muridae) in China
inferred from complete mitochondrial cytochrome b gene. Mol
Phylogenet Evol 44: 521–529
Kaneko Y (1996) Morphological variation, and latitudinal and altitudi-
nal distribution of Eothenomys chinensis, E. wardi, E. custos, E.
proditor, and E. olitor (Rodentia, Arvicolidae) in China. Mam
Study 21: 89–114
Kimura M (1980) A simple method for estimating evolutionary rates
of base substitutions through comparative studies of nucleotide
sequences. J Mol Evol 16: 111–120
Li CL, Xue XX (1996) Biogeography and the age of the fossil
rodents fauna from Xishuidong Nantian, Shaanxi. Vertebrata

S. Liu et al.622
Pal Asiatica 34: 156–162
Liu SY, Sun ZY, Zeng ZY, Zhao EM (2007) A new species
(Proedomys: Aricolinae: Murida) from Sichuan province, China.
J Mammal 88: 1170–1178
Luo J, Yang DM, Hitoshi S, Wang YX, Chen WJ, Campbell KL, et al.
(2004) Molecular phylogeny and biogeography of Oriental voles:
genus Eothenomys (Muridae, Mammalia). Mol Phylogenet Evol
33: 349–362
Luo ZX, Chen W, Guo B (2000) Fauna Sinica. Mammalia, Vol. 6,
Rodentia, Part III: Cricetidae. Scientific Publisher, Beijing
Ma Y, Jiang JQ (1996) The reinstatement of the status of genus
Caryomys (Rodentia: Microtinae). Acta Zootax Sinica 21: 493–
497
Miller GS (1896) Genera and subgenera of voles and lemmings. N
Amer Fauna 12: 1–86
Miller GS (1898) Description of a new genus and species of micro-
tine rodent from Siberia. Proc Acad Nat Sci Philadelphia 50:
368–371
Milne-Edwards A (1871) Note sur une nouvelle espece de semnopi-
theque. Nouv Arch Mus Hist Paris Bull 7: 93
Murphy RW, Méndez de la Cruz FR (2010) The herpetofauna of
Baja California and its associated islands: a conservation
assessment and priorities. In “Conservation of Mesoamerican
Amphibians and Reptiles” Ed by LD Wilson, JH Townsend and
JD Johnson, Eagle Mountain Publishing, LC, Eagle Mountain,
Utah, pp 238–273
Musser GG, Carleton MD (1993) Family Muridae. In “Mammal Spe-
cies of the World: a Taxonomic and Geographic Reference” Ed
by DE Wilson and DM Reeder, 2nd ed, Smithsonian Institution
Press, Washington, DC, pp 501–535
Musser GG, Carleton MD (2005) Family Cricetidae. In “Mammal
Species of the World: a Taxonomic and Geographic Reference”
Ed by DE Wilson and DM Reeder, 3rd ed, The Johns Hopkins
Press, Baltimore, MD, pp 955–1039
Nowak RM (1999) Walker’s Mammals of the World. 6th ed, The
Johns Hopkins University Press, Baltimore and London
Nowak RM, Paradiso JL (1983) Walker’s Mammals of the World. 4th
ed, The Johns Hopkins University Press, Baltimore and London
Osgood WH (1932) Mammals of the Kelley-Roosevelts and Dela-
cour Asiatic Expeditions. Field Mus Nat Hist Zool 18: 193–339
Posada D, Crandall KA (1998) ModelTest: testing the model of DNA
substitution. Bioinformatics 14: 817–818
Posada D, Crandall KA (2001) Selecting the best-fit model of nucle-
otide substitution. Syst Biol 50: 580–601
Rambaut A, Drummond AJ (2003) Tracer v1.3. Available from http://
evolve.zoo.ox.ac.uk
Robovský J, Ra Ánková V, Zrzavý J (2008) Phylogeny of Arvicolinae
(Mammalia, Cricetidae): utility of morphological and molecular
datasets in a recently radiating clade. Zool Scripta 37: 571–590
Ronquist F, Huelsenbeck JP (2003) MRBAYES 3: Bayesian phylo-
genetic inference under mixed models. Bioinformatics 19:
1572–1574
Sambrook J, Russell DW (2002) Molecular Cloning: A Laboratory
Manual. 3rd ed, Cold Spring Harbor Laboratory Press
Stamatakis A, Hoover P, Rougemont J (2008) A rapid bootstrap
algorithm for the RAxML web-servers. Syst Biol 75: 758–771
Steppan SJ, Adkins RM, Anderson J (2004) Phylogeny and diver-
gence-date estimates of rapid radiations in muroid rodents
based on multiple nuclear genes. Syst Biol 53: 533–553
Swofford D (2003) PAUP*: Phylogenetic Analysis Using Parsimony
and Other Methods. version 4.0, Sinauer, Sunderland,
Massachusetts
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S
(2011) MEGA5: Molecular Evolutionary Genetics Analysis
using maximum likelihood, evolutionary distance, and maxi-
mum parsimony methods. Mol Phylogenet Evol 28: 2731–2739
Thomas O (1891) Description of a new vole from China. Ann Mag
Nat Hist Ser 6, 8: 117–118
Thomas O (1905a) The Duke of Bedford’s zoological exploration in
eastern Asia.—1. List of mammals obtained by Mr. M. P.
Anderson in Japan. Proc Zool Soc Lond 1906 (2): 331–363
Thomas O (1905b) On some new Japanese mammals presented to
the British Museum by Mr. R. Gordon Smith. Ann Mag Nat Hist
Ser 7, 15: 487–495
Thomas O (1907) The Duke of Bedford’s zoological expedition in
eastern Asia. List of small mammals from Korea and Quelpart.
Proc Zool Soc Lond (1906): 863–865
Thomas O (1908a) Mammals collected in the provinces of Shan-si
and Shen-si, Northern China. Abstr Proc Zool Soc Lond 63: 44–
45
Thomas O (1908b) List of mammals from the provinces Chih-li and
Shan-si northern China. Proc Zool Soc Lond 1908: 635–646
Thomas O (1910) Exhibition of mammals from China. Abstr Proc
Zool Soc Lond 83: 25–26
Thomas O (1911a) Mammals in west of China. Abstr Proc Zool Soc
Lond 100: 50
Thomas O (1911b) Mammals collected in the province of Kan-su
and Sze-chwan, western China, by Mr. Malcolm Anderson, for
the Duke of Bedford’s exploration of Eastern Asia. Abstr Proc
Zool Soc Lond 90: 3–5
Thomas O (1912) On insectivores and rodents collected by Mr. F.
Kingdom Ward in Northwest Yunnan. Ann Mag Nat Hist Ser 8,
9: 513–519
Thomas O (1914) Second list of small mammals from western Yunnan
collected by Mr. F. Kingdom Ward. Ann Mag Nat Hist, Ser. 8,
14: 472–475
Thomas O (1921) On small mammals from the Kachin Province,
northern Burma. J Bombay Nat Hist Soc 27: 499–505
Tokuda M (1937) The alpine Muridae from Formosa. Bot Zool Tokyo
5: 1115–1122
Wang YX (2003) A Complete Checklist of Mammal Species and
Subspecies in China: A Taxonomic and Geographical Refer-
ence. China Forestry Publishing House, Beijing
Yang DM, Liu RQ, Zhang YP, Chen ZP, Wang YX (1998) Chromo-
some study of Yulong vole (Eothenomys proditor). Cytologia
65: 435–440
Yang SJ, Yin ZH, Ma XM, Lei FM (2006) Phylogeography of ground
tit (Pseudopodoces humilis) based on mtDNA: Evidence of
past fragmentation on the Tibetan Plateau. Mol Phylogenet
Evol 41: 257–265
Ye XD, Ma Y, Zhang JS, Wang CL, Wang ZK (2002) A summary of
Eothenomi (Rodentia: Cricetidae: Microtinae). Acta Zootax
Sinica 27: 173–182
Zhang DR, Chen MY, Murphy RW, Che J, Pang JF, JS Hu, et al.
(2010) Genealogy and palaeodrainage basins in Yunnan Prov-
ince: phylogeography of the Yunnan spiny frog, Nanorana
yunnanensis (Dicroglossidae). Mol Ecol 19: 3406–3420
Zhang DX, Hewitt GM (1996) Nuclear integrations: challenges for
mitochondrial DNA markers. Trends Ecol Evol 11: 247–251
Zhang FF, Jiang ZG (2006) Mitochondrial phylogeography and
genetic diversity of Tibetan gazelle (Procapra picticaudata):
implications for conservation. Mol Phylogenet Evol 41: 313–321
Zheng HB, Powell CM, An ZS, Dong GR (2000) Pliocene uplift of the
northern Tibet Plateau. Geology 8: 715–718
Zheng SH (1993) Quaternary Rodents of Sichuan and Guizhou
Area, China. Science Press, Beijing
(Received April 4, 2011 / Accepted February 28, 2012)


















