
Molecular phylogeny of the Bacidiaceae (Lecanorales, lichenized ...
15
783 Stefan EKMAN Department of Botany, University of Bergen, Alle U gaten 41, N-5007 Bergen, Norway. E-mail : stefan.ekman!bot.uib.no Received 28 September 2000 ; accepted 25 January 2001. The phylogeny of the family Bacidiaceae (Lecanorales, Ascomycota) was investigated using 65 nuclear ITS1-5–8S-ITS2 ribosomal DNA sequences, 63 of which were newly determined. After exclusion of ambiguous alignment, the data set contained 285 variable characters, 212 of which were parsimony-informative. Phylogenetic estimations were performed with maximum parsimony (unweighted and weighted) and maximum likelihood optimality criteria. Four different phylogenetic hypotheses were tested using a parametric bootstrap approach to simulate the expected null distribution of the difference between the globally optimal tree and the best (constrained) tree agreeing with the null hypotheses under unweighted and weighted parsimony, and maximum likelihood : (1) the genus Bacidia is monophyletic ; (2) the genus Bacidina is monophyletic ; (3) the genus Toninia is monophyletic ; and (4) the family Ramalinaceae is monophyletic and distinct from a monophyletic Bacidiaceae. The monophyly of Bacidia, Toninia, and the Ramalinaceae was rejected under all circumstances. Hence, Bacidiaceae is likely to be a younger synonym of Ramalinaceae. The monophyly of Bacidina was not rejected under any optimality criterion. Furthermore, the data set suggests that the Bacidia beckhausii and B. sabuletorum groups are unrelated to Bacidia s. str., that Megalaria is monophyletic, and that Lecania auct. is polyphyletic. INTRODUCTION The Bacidiaceae belongs in the Lecanorales, the largest of the orders among the ascomycetes. Its presently most widely used circumscription was established by Eriksson & Hawks- worth (1987, 1993, 1998), who included 28 genera. In their sense, the family included crustose and some squamulose, foliose, and subfruticose taxa with a chlorococcoid photobiont, biatorine (or sometimes lecanorine) apothecia, a more or less well-developed annular proper exciple of hyphae dissimilar to the paraphyses, a well-developed hypothecium, a hymenium of sparsely branched paraphyses and asci with an amyloid apex of the ‘ Bacidia-type ’ or ‘ Biatora-type ’ (Hafellner (1984), non- or transversely septate (rarely muriform) ascospores, uni- or plurilocular pycnidia with conidiophores of types II, III, and VI (Vobis 1980), and ellipsoid, bacilliform or filiform conidia. Historically, these lichens were included in the gigantic Lecideaceae (Zahlbruckner (1905, 1921–40), which included most crustose lichens with biatorine or lecideine apothecia and a non-Trentepohlia photobiont. They were treated as such until the comprehensive reclassification of the Lecanoraceae and Lecideaceae by Hafellner (1984). In that work, the taxa belonging to the family now known as Bacidiaceae (as circumscribed by Eriksson & Hawksworth, loc. cit.) were dispersed over the Bacidiaceae, Biatoraceae, Catinariaceae, Lecaniaceae, Phyllopsoraceae, Tephromelataceae, Schadoniaceae, and Squamarinaceae. Later, Hafellner (1988 : 45) reduced the Biatoraceae and Lecaniaceae into synonymy with the Bacidiaceae. Even though Eriksson & Hawksworth (1987, 1993, 1998) amalgamated several of Hafellner’s families, the delimitation of the family has been questioned. It was suggested by Rambold (1989 : 21, 73), Rambold & Triebel (1992 : 52, 61), and Hertel & Rambold (1995) that the Lecanoraceae and Bacidiaceae as used by Eriksson & Hawksworth were insufficiently delimited and should be treated as one family, the name of which would then be Lecanoraceae. This view was tentatively supported by Ekman (1996 a). Based on SSU rDNA data, however, Ekman & Wedin (2000), demonstrated that the Bacidiaceae and Lecanoraceae are likely to represent distinct families, although the genera Tephromela and Scolicio- sporum probably do not belong in either of the families. A factor that may have contributed to the confusion around the distinction between the Lecanoraceae and Bacidiaceae is the different interpretation of single genera. Bacidina was first treated in the Lecanoraceae (e.g. Eriksson & Hawksworth 1987, 1993), and subsequently in the Bacidiaceae (Ekman 1996 a, Eriksson & Hawksworth 1998). The genera Megalaria and Tylothallia have been treated in the Lecanoraceae (Eriksson & Hawksworth 1998), but Ekman & Tønsberg (1996) presented data speaking in favour of including Megalaria in the Bacidiaceae, and Ekman (1997) considered Tylothallia to be closely related to Cliostomum, a member of the Bacidiaceae (Eriksson & Hawksworth 1987, 1993, 1998, Ekman & Wedin 2000). Although not a matter of much debate, the delimitation of the Bacidiaceae relative to the Catillariaceae as used by Eriksson & Hawksworth is also troubled by misclassified Mycol. Res. 105 (7) : 783–797 (July 2001). Printed in the United Kingdom. Molecular phylogeny of the Bacidiaceae (Lecanorales, lichenized Ascomycota)
Transcript of Molecular phylogeny of the Bacidiaceae (Lecanorales, lichenized ...

783
Stefan EKMAN
Department of Botany, University of Bergen, AlleUgaten 41, N-5007 Bergen, Norway.
E-mail : stefan.ekman!bot.uib.no
Received 28 September 2000 ; accepted 25 January 2001.
The phylogeny of the family Bacidiaceae (Lecanorales, Ascomycota) was investigated using 65 nuclear ITS1-5±8S-ITS2 ribosomal DNA
sequences, 63 of which were newly determined. After exclusion of ambiguous alignment, the data set contained 285 variable
characters, 212 of which were parsimony-informative. Phylogenetic estimations were performed with maximum parsimony
(unweighted and weighted) and maximum likelihood optimality criteria. Four different phylogenetic hypotheses were tested using a
parametric bootstrap approach to simulate the expected null distribution of the difference between the globally optimal tree and the
best (constrained) tree agreeing with the null hypotheses under unweighted and weighted parsimony, and maximum likelihood : (1)
the genus Bacidia is monophyletic ; (2) the genus Bacidina is monophyletic ; (3) the genus Toninia is monophyletic ; and (4) the family
Ramalinaceae is monophyletic and distinct from a monophyletic Bacidiaceae. The monophyly of Bacidia, Toninia, and the Ramalinaceae
was rejected under all circumstances. Hence, Bacidiaceae is likely to be a younger synonym of Ramalinaceae. The monophyly of
Bacidina was not rejected under any optimality criterion. Furthermore, the data set suggests that the Bacidia beckhausii and B.
sabuletorum groups are unrelated to Bacidia s. str., that Megalaria is monophyletic, and that Lecania auct. is polyphyletic.
INTRODUCTION
The Bacidiaceae belongs in the Lecanorales, the largest of the
orders among the ascomycetes. Its presently most widely
used circumscription was established by Eriksson & Hawks-
worth (1987, 1993, 1998), who included 28 genera. In their
sense, the family included crustose and some squamulose,
foliose, and subfruticose taxa with a chlorococcoid photobiont,
biatorine (or sometimes lecanorine) apothecia, a more or less
well-developed annular proper exciple of hyphae dissimilar to
the paraphyses, a well-developed hypothecium, a hymenium
of sparsely branched paraphyses and asci with an amyloid
apex of the ‘Bacidia-type ’ or ‘Biatora-type ’ (Hafellner (1984),
non- or transversely septate (rarely muriform) ascospores, uni-
or plurilocular pycnidia with conidiophores of types II, III,
and VI (Vobis 1980), and ellipsoid, bacilliform or filiform
conidia. Historically, these lichens were included in the
gigantic Lecideaceae (Zahlbruckner (1905, 1921–40), which
included most crustose lichens with biatorine or lecideine
apothecia and a non-Trentepohlia photobiont. They were
treated as such until the comprehensive reclassification of the
Lecanoraceae and Lecideaceae by Hafellner (1984). In that work,
the taxa belonging to the family now known as Bacidiaceae (as
circumscribed by Eriksson & Hawksworth, loc. cit.) were
dispersed over the Bacidiaceae, Biatoraceae, Catinariaceae,
Lecaniaceae, Phyllopsoraceae, Tephromelataceae, Schadoniaceae,
and Squamarinaceae. Later, Hafellner (1988 : 45) reduced the
Biatoraceae and Lecaniaceae into synonymy with the Bacidiaceae.
Even though Eriksson & Hawksworth (1987, 1993, 1998)
amalgamated several of Hafellner’s families, the delimitation
of the family has been questioned. It was suggested by
Rambold (1989 : 21, 73), Rambold & Triebel (1992 : 52, 61),
and Hertel & Rambold (1995) that the Lecanoraceae and
Bacidiaceae as used by Eriksson & Hawksworth were
insufficiently delimited and should be treated as one family,
the name of which would then be Lecanoraceae. This view was
tentatively supported by Ekman (1996a). Based on SSU
rDNA data, however, Ekman & Wedin (2000), demonstrated
that the Bacidiaceae and Lecanoraceae are likely to represent
distinct families, although the genera Tephromela and Scolicio-
sporum probably do not belong in either of the families.
A factor that may have contributed to the confusion around
the distinction between the Lecanoraceae and Bacidiaceae is the
different interpretation of single genera. Bacidina was first
treated in the Lecanoraceae (e.g. Eriksson & Hawksworth 1987,
1993), and subsequently in the Bacidiaceae (Ekman 1996a,
Eriksson & Hawksworth 1998). The genera Megalaria and
Tylothallia have been treated in the Lecanoraceae (Eriksson &
Hawksworth 1998), but Ekman & Tønsberg (1996) presented
data speaking in favour of including Megalaria in the
Bacidiaceae, and Ekman (1997) considered Tylothallia to be
closely related to Cliostomum, a member of the Bacidiaceae
(Eriksson & Hawksworth 1987, 1993, 1998, Ekman & Wedin
2000). Although not a matter of much debate, the delimitation
of the Bacidiaceae relative to the Catillariaceae as used by
Eriksson & Hawksworth is also troubled by misclassified
Mycol. Res. 105 (7) : 783–797 (July 2001). Printed in the United Kingdom.
Molecular phylogeny of the Bacidiaceae (Lecanorales, lichenizedAscomycota)

Molecular phylogeny of the Bacidiaceae 784
genera. Classified in the Catillariaceae by Eriksson &
Hawksworth (1993, 1998), Arthrosporum and Toninia were
considered closely related to, possibly even indistinctly
delimited from, Bacidia, the largest of the genera in the
Bacidiaceae, by Ekman (1996a) and Timdal (1991). For a more
detailed discussion on the relationships between genera of the
Bacidiaceae, see Timdal (1991), Printzen (1995), and Ekman
(1996a).
A further family with unclear relationships to the Bacidiaceae
is the Ramalinaceae, containing the well-known fruticose
lichens Ramalina and Niebla. The Ramalinaceae possess an
amyloid ascus apex that is virtually identical to the one in the
Bacidiaceae, transversely septate spores are common in both
families, and a rare group of secondary metabolites, the
orcinol meta-depsides, are present in both families. With the
inclusion of the newly described crustose genus Ramalinora
(Lumbsch, Rambold & Elix 1995) in the Ramalinaceae, it has
become difficult to draw a clear-cut line between the families.
Although different in important characters, e.g. apothecium
ontogeny, the similarities between Ramalinora, with its only
known species R. glaucolivida (Lumbsch et al. 1995), and
Cliostomum, particularly C. tenerum (Ekman 1997), are striking.
The relationships between the Bacidiaceae and the Ramalinaceae
are definitively in need of scrutiny.
The relationships between the genera within the Bacidiaceae,
whatever circumscription is used, and between species and
species groups within its genera is largely unknown. There is
so far not a single published work dealing with the phylogeny
of the Bacidiaceae or any of its constituent genera (apart from
the outline by Ekman & Wedin 2000). As already mentioned,
Timdal (1991 : 23) pointed to problems with delimiting
Toninia from Bacidia. The genus Bacidina in the sense of Ekman
(1996a) contains a high degree of variation, and its monophyly
can be questioned. Lecania in its traditional sense appears
heterogeneous (Ekman 1996a). Hafellner (1984) questioned
the distinctness of the subfruticose genus Thamnolecania
relative to the crustose Lecania. Ekman (1996a : 45–46)
excluded the ‘Bacidia beckhausii group ’, the ‘Bacidia lutescens
group ’, and the ‘Bacidia sabuletorum group ’ from Bacidia s. str.
but was unable to refer them to any other described genus.
Several of the genera accepted in the Bacidiaceae by Eriksson
& Hawksworth (1998) include only a few known species (e.g.
Adelolecia, Boreoplaca, Herteliana, Rolfidium, Schadonia, Speer-
schneidera, Squamacidia, and Waynea), and their relationships
with the variable, species-rich genera of the family (Bacidia,
Biatora, Lecania, and Toninia) are poorly understood.
In addition to the taxonomic considerations, nomenclature
has been in dispute. The name Bacidiaceae was validly
described by Watson (1929). Both Biatoraceae (Stizenberger
1862 : 163) and Phyllopsoraceae (Zahlbruckner 1905) are older
synonyms, but the validity of the former needs to be
investigated further (Art. 33±7 ; Greuter et al. 2000). However,
the use of Bacidiaceae is maintained here, as it is a relatively
well-established name for the group in question (Eriksson &
Hawksworth 1987, 1993, 1998).
The problems involved in understanding familial and
generic boundaries in this group of lichens stem mainly from
the hazards of interpreting and homologizing morphological
characters. The aim of this investigation is to better understand
the Biatoraceae and its phylogeny using molecular data, in this
case DNA sequences from the ITS1-5±8S-ITS2 ribosomal
DNA. Particular attention is paid to the testing of four null
hypotheses : (1) Bacidia in the sense of Ekman (1996a) forms
a monophyletic group ; (2) Bacidina in the sense of Ekman
(1996a) forms a monophyletic group ; (3) Toninia in the sense
of Timdal (1991) forms a monophyletic group ; and (4) the
Ramalinaceae in the sense of many authors (e.g. Eriksson &
Hawksworth 1998) form a monophyletic group, which is
distinct from a monophyletic Bacidiaceae.
MATERIALS AND METHODS
Specimens
New sequences were obtained from 66 species listed in Table
1. The vast majority, 63 species, belong to the Bacidiaceae as
circumscribed by Eriksson & Hawksworth (1998) with the
corrections suggested by Ekman (1996a, 1997) and Ekman &
Wedin (2000) (Table 1). The remaining three species,
Scoliciosporum umbrinum, Tephromela atra, and T. aglaea, were
included as potential outgroup taxa.
DNA extraction, PCR amplification, sequencing, and
editing
DNA was extracted using the DNeasy Plant Mini Kit4(Qiagen). Complete PCR amplification of the nuclear ITS1-
5±8S-ITS2 ribosomal DNA region was performed using the
primers ITS1F (Gardes & Bruns 1993) and ITS4 (White et al.
1990). The PCR cocktail, the total volume of which was 50 µl,
contained, in addition to extracted DNA, 2±5 mM MgCl#,
200 µM of each of the four dNTPs, 0±7 µM of each primer,
1±5 U of a DNA polymerase (either AmpliTaq or AmpliTaq
Gold, PE Biosystems), together with a Mg#+ free buffer in the
concentration recommended by the manufacturer. The fol-
lowing PCR cycling parameters were used : a two-minute
(AmpliTaq) or a nine-minute hold (AmpliTaq Gold) at 94 °Cfollowed by six cycles including denaturation at 94 °(AmpliTaq) or 95 ° (AmpliTaq Gold) for 60 s, annealing at
62 ° (decreasing 1 ° each cycle) for 60 s, and extension at 72 °for 105 s, then 34 cycles with denaturation at 94 ° (AmpliTaq)
or 95 ° (AmpliTaq Gold) for 30 s, annealing at 56 ° for 30 s,
and extension at 72 ° for 105 s plus an addition of three
seconds each cycle, and finally a 10-min hold at 72 °, after
which the reaction was cooled to a constant 4 °. PCR products
were electrophorized in a 1% agarose gel and visualized using
ethidium bromide. They were subsequently cleaned using the
QiaQuick Spin kit (Qiagen) or, in case of an impure product,
the QiaQuick Gel Extraction kit (Qiagen). The cleaned PCR
product was sequenced with the ITS1F and ITS4 primers,
sometimes also the ITS2 and ITS3 primers (White et al. 1990)
when a long PCR product was obtained due to the presence
of group I introns at the very end of the SSU (Gargas,
DePriest & Taylor 1995). The Big Dye Terminator kit (PE
Biosystems) was used according to the manufacturer’s
instructions except that half-size instead of full-size reactions
were used. The final extension product was cleaned using a
NaAc precipitation protocol according to the manufacturer’s

S. Ekman 785
Table 1. Species from which new ITS1-5±8S-ITS2 nuclear ribosomal DNA sequences were obtained.
Species Generic affiliation CollectionGenBank}EMBLaccession no.
Arthrosporum populorum Haugan 4430 (O) AF282106Bacidia absistens Ekman 3223 (BG) AF282085B. arceutina Ekman 3110 (BG) AF282083B. auerswaldii P. Johansson 20 (UPS) AF282122B. bagliettoana Ekman 3137 (BG) AF282123B. beckhausii Not Bacidia s. str. Holien 6744 (TRH) AF282071B. biatorina Knutsson 94-148 (hb. Knutsson) AF282079B. caligans Bacidina P. Johansson 21 (UPS) AF282096B. circumspecta Ekman L1330 (LD) AF282124B. diffracta Wetmore 26401 (MIN) AF282090B. fraxinea T. Johansson 1620 (BG) AF282088B. hemipolia Not Bacidia s. str. Tønsberg 25091 (BG) AF282072B. hostheleoides 1996, Seaward (hb. Seaward 108121) AF282081B. incompta Ekman 3144 (BG) AF282092B. laurocerasi subsp. laurocerasi Wetmore 74318 (MIN) AF282078B. lutescens ? Ekman L1161 (LD) AF282082B. medialis Ekman L1193 (LD) AF282102B. polychroa Knutsson 91-215 (hb. Knutsson) AF282089B. rosella Ekman 3117 (BG) AF282086B. rubella auct. Ekman 3021 (BG) AF282087B. sabuletorum Not Bacidia s. str. Ekman 3091 (BG) AF282069B. schweinitzii Wetmore 72619 (MIN) AF282080B. scopulicola Ekman 3106 (BG) AF282084B. subincompta auct. Ekman 3413 (BG) AF282125B. suffusa Wetmore 74771 (MIN) AF282091B. vermifera T. Johansson 1619 (BG) AF282109Bacidina arnoldiana Ekman 3157 (BG) AF282093B. chloroticula Tønsberg 18642 (BG) AF282098B. delicata 1996, Fritz (BG) AF282097B. egenula Ekman 3003 (BG) AF282095B. inundata Ekman 3187 (BG) AF282094B. phacodes Ekman 3414 (BG) AF282100Biatora sphaeroides Mycobilimbia Ekman 3454 (BG) AF282068B. vernalis Tønsberg 23757 (BG) AF282070Catillaria globulosa Not Catillaria s. str. Ekman 3142 (BG) AF282073Cliostomum griffithii Ekman 3022 (BG) AF282076Lecania cyrtella Ekman 3017 (BG) AF282067L. naegelii Ekman 3401 (BG) AF282101Megalaria grossa Ekman 3466 (BG) AF282074M. laureri Ekman 3119 (BG) AF282075Thamnolecania brialmontii Convey 121 (AAS) AF282066Toninia alutacea Haugan & Timdal 4824 (O) AF282116T. aromatica Haugan & Timdal 4819 (O) AF282126T. candida Bratli & Timdal 8733 (O) AF282117T. cinereovirens Haugan & Timdal 7953 (O) AF282104T. coelestina Haugan 5985 (O) AF282127T. lutosa Timdal SON28}08 (O) AF282114T. nordlandica Haugan & Timdal 8129 (O) AF282113T. opuntioides Haugan & Timdal 8057 (O) AF282119T. pennina Haugan & Timdal 8122 (O) AF282111T. philippea Haugan & Timdal H3750 (O) AF282112T. plumbina Haugan 4352 (O) AF282107T. rosulata Timdal 8640 (O) AF282121T. sculpturata Haugan & Timdal 7829 (O) AF282110T. sedifolia Knutsson 97-407 (BG) AF282120T. squalida Haugan 4970 (O) AF282103T. talparum Timdal SON120}01 (O) AF282108T. taurica Haugan & Timdal 8060 (O) AF282118T. toniniana TuX rk 20721 (O) AF282115T. tristis subsp. tristis Haugan & Timdal 8109 (O) AF282105T. verrucarioides Bratli & Timdal 8709 (O) AF282128Tylothallia biformigera Ekman 3096 (BG) AF282077Waynea californica Tønsberg 21048 (BG) AF282099
Generic affiliations follow Ekman (1996), Printzen (1995), or Timdal (1991) if other than the one indicated by the generic name of the taxon (‘ ? ’ indicates
uncertain affinity different from the generic name). Abbreviations of public herbaria in which source collections are deposited follow Holmgren, Holmgren &
Barnett (1990). Private herbaria are denoted ‘hb ’.

Molecular phylogeny of the Bacidiaceae 786
recommendations. Extension products were subjected to
automatic sequencing on an ABI 377 with the XL upgrade (PE
Biosystems). Sequence fragments were assembled and edited
using Sequencher 3±0 and 3±1.1 (Gene Codes). Partial SSU and
LSU rDNA sequences, sometimes including an intron, at the
beginning and the end of the resulting sequence were
removed before alignment. The start of ITS1 and end of ITS2
were defined using the nuclear rDNA part of the Saccharomyces
cerevisiae chromosome XII sequence Z73326 obtained from
GenBank.
Sequence alignment
The 66 new sequences were aligned together with four
additional sequences : Leifidium tenerum (GenBank accession
number AF117998), Ramalina fastigiata (U84583), Ramalina
siliquosa (U84587), and Sphaerophorus globosus (AF282129).
First, a preliminary alignment including all taxa was performed
using SAM 3±0 (Sequence Alignment and Modeling Software
System) (Krogh et al. 1994, Hughey & Krogh 1996, Hughey,
Karplus & Krogh 1999, Durbin et al. 1998), which is available
on-line at Institut Pasteur (Paris ; http :}}bioweb.pasteur.fr}-
seqanal}motif}sam-uk.html). From the resulting preliminary
alignment it was evident that Scoliciosporum umbrinum,
Tephromela aglaea, and T. atra were dubiously alignable with
the other taxa, and they were hence excluded from the further
analysis. The SAM analysis was reiterated with the remaining
67 taxa. Finally, the alignment was manually optimized.
However, a series of gap-rich regions in the ITS1 and ITS2,
clearly remained ambiguously aligned. These sites were
consequently excluded and the remaining ones were used in
the subsequent phylogenetic analyses and analysis of
phylogenetic signal. The final alignment was submitted
to TreeBASE (Harvard ; http :}}www.herbaria.harvard.edu}treebase) where it is filed under matrix accession number
M849.
Phylogenetic signal
For an a priori estimation of the amount of phylogenetic
signal, the Relative Apparent Synapomorphy Analysis (RASA)
technique was applied (Lyons-Weiler, Hoelzer & Tausch
1996), as implemented in the software RASA 2±3 (Lyons-
Weiler 1999). Unlike other measures of phylogenetic signal,
RASA is tree-independent and is not based on the assumptions
of a tree-building optimality criterion. A rooted test was
performed, and the inferred signal content was compared to
an analytical null model. Here and in the subsequent
phylogenetic analyses, Sphaerophorus globosus and Leifidium
tenerum (Sphaerophoraceae) were used as outgroup taxa
following a suggestion by Ekman & Wedin (2000) that the
Sphaerophoraceae constitute a possible sister-group to the
Bacidiaceae.
Phylogenetic analyses
Phylogenetic analyses were performed on an Apple Power-
Macintosh G3}266 using maximum (unweighted) parsimony
(MP), maximum weighted parsimony (MWP), and maximum
likelihood (ML) optimality criteria as implemented in the
computer programme PAUP* 4±0b2a, except for the analyses
of simulated data, which were analysed using PAUP* 4±0b3aand 4±0b4a (Swofford 1999).
Unweighted parsimony
Gaps were treated as missing data. A heuristic search with
1000 random-addition sequence replicates was performed
using tree bisection-reconnection (TBR) branch-swapping, and
with the MulTrees option on and the steepest descent and
collapse zero-length branches options off. Multiple character
states were interpreted as uncertainties. Branch lengths were
assigned using ACCTRAN character state optimization.
Branch support was estimated using a jackknife analysis
with 10000 replicates with the same search parameters as
above, except that 100 random-addition replicates were tested
in each replicate and that MulTrees was off. The nominal
exclusion of characters was 36±79% (Farris et al. 1996) and
JAC resampling was emulated using PAUP*.
Weighted parsimony
Settings were identical to the unweighted analysis, except that
character state transformations were weighted unequally
according to a symmetric step matrix. Weights were obtained
by charting the absolute average number of changes on 100
equiprobable random trees using MacClade 3±08 (Maddison
& Maddison 1992, Lutzoni 1997). Reciprocal substitutions
were summed, and the relative frequency of each substitution
type was calculated. Frequencies were converted to costs of
changes using the negative natural logarithm of the frequency
and rounded to nearest single-decimal number. The resulting
step matrix was used in PAUP* by applying user-defined
character types. Violations against the triangle inequality were
not present. Branch support was estimated using a jackknife
analysis with 300 replicates. All other search parameters were
identical to the unweighted parsimony jackknife.
Maximum likelihood
In order to investigate what likelihood model best fitted the
data, a likelihood ratio test (Huelsenbeck & Crandall 1997 ;
Huelsenbeck & Rannala 1997) was performed as implemented
in the computer programme MODELTEST 2±1 (Posada &
Crandall 1998). The critical value of rejection was set to 0±008in order to maintain an overall type I error rate of 0±05. The
results favoured a model with unequal nucleotide frequencies
and six different time-reversible substitution types. This
corresponds to the GTR model (e.g. Rodrı!guez et al. 1990,
Yang 1994a). Furthermore, the test favoured the assumption
of invariability (I) in a fraction of sites, pinv
(Gu et al. 1995), and
substitution rate heterogeneity among nucleotide sites
according to a gamma model (Yang 1993).
A χ# test of homogeneity of base frequencies across taxa
was performed, as implemented in PAUP*, since the GTR
likelihood model employed here assumes stationary base
frequencies.
Initial values of the six relative substitution rates (the R
matrix), the proportion of invariable sites (pinv
), and the

S. Ekman 787
gamma curve shape parameter (α) were estimated from one of
the most parsimonious trees using maximum likelihood, and
set constant during the search. The likelihood of this tree was
calculated with four, six, eight, and ten discrete gamma
categories (Yang 1994b), and it was found that the likelihood
was maximized with six categories. Hence, a discrete gamma
model with six categories was applied (dΓ6), and the average
rates of the categories were represented by their means. A tree
search in three parts was conducted : (1) A heuristic search
with 50 random-addition sequence replicates was performed
using nearest-neighbour interchanges (NNI) branch-swapping ;
(2) the most likely trees found in the first search were input
into a second round of branch-swapping with subtree pruning-
regrafting (SPR) ; and (3) the most likely tree(s) found in the
second search were input into a third round of branch-
swapping with tree bisection-reconnection (TBR). After the
search was completed, likelihood parameters were reestimated
according to the most likely tree found, and the heuristic
search was reiterated until tree topology and likelihood
parameters had converged and continued iteration was
unnecessary. Convergence occurred after the third search.
During all searches, the MulTrees option and collapse
effectively zero-length branches options were on, and the
steepest descent option was off.
Each branch in the resulting tree was subjected to a
likelihood ratio test (as implemented in PAUP*) of the null
hypothesis that the branch has zero length. This test was
performed with full reoptimization after forcing a branch
length to zero.
Branch support was estimated using a jackknife analysis
with 100 replicates. Starting trees were obtained using
neighbour-joining (with maximum likelihood distances), which
were subjected to nearest-neighbour interchanges (NNI)
branch-swapping. MulTrees was on, and nchuck and chuck-
score was set to 10 and 1, respectively.
Hypothesis testing
First, the optimal trees agreeing with each of the constraints
inherent in the four null hypotheses were searched for under
unweighted and weighted parsimony, and maximum like-
lihood. The following constraints were used :
Null hypothesis 1 : Bacidia is monophyletic, i.e. B. absistens,
B. arceutina, B. auerswaldii, B. bagliettoana, B. biatorina, B.
circumspecta, B. diffracta, B. fraxinea, B. hostheleoides, B. incompta,
B. laurocerasi, B. medialis, B. polychroa, B. rosella, B. rubella, B.
schweinitzii, B. scopulicola, B. subincompta, B. suffusa, and B.
vermifera form a monophyletic group.
Null hypothesis 2 : Bacidina is monophyletic, i.e. B.
arnoldiana, B. caligans, B. chloroticula, B. delicata, B. egenula, B.
inundata, and B. phacodes form a monophyletic group.
Null hypothesis 3 : Toninia is monophyletic, i.e. T. alutacea,
T. aromatica, T. candida, T. cinereovirens, T. coelestina, T. lutosa,
T. nordlandica, T. opuntioides, T. pennina, T. philippea, T.
plumbina, T. rosulata, T. sculpturata, T. squalida, T. talparum, T.
taurica, T. toniniana, T. tristis, T. sedifolia, and T. verrucarioides
form a monophyletic group.
Null hypothesis 4 : Ramalinaceae is distinct from Bacidiaceae,
i.e. Ramalina fastigiata and R. siliquosa form a monophyletic
group, and all other ingroup taxa form another monophyletic
group.
Search parameters under MP, MWP, and ML were identi-
cal to the searches for the unconstrained trees, except
that six discrete gamma categories (dΓ6) were assumed
rather than tested for in the ML analysis. Different trees
may have different optimal likelihood functions and hence
likelihood parameters in the ML analysis were optimized
for the particular tree topology, rather than, e.g., kept
identical to the parameters of the globally most likely
tree.
When a search yielded more than a single optimal tree
(unweighted and weighted parsimony), the one with the best
likelihood was chosen to represent the null hypothesis. The
likelihood of all the most parsimonious, unweighted and
weighted, trees was calculated using a GTRIdΓ6 model
with the R matrix, pinv
, and α either estimated from the tree
topology (weighted parsimony) or, due to computational
effort, kept constant at the parameters of the globally most
likely tree. Polytomies occurring in the most likely constrained
trees (since effectively zero-length branches were collapsed in
the constrained ML searches) were randomly dichotomized
using MacClade 3±08 (Maddison & Maddison 1992). Branch
lengths of all trees (MP, MWP, ML) selected to represent the
null hypotheses were optimized under a GTRIdΓ6
likelihood model with the R matrix, pinv
, and α estimated.
Branch lengths and likelihood parameters were saved for later
use in the parametric bootstrap. None of the trees representing
the same null hypothesis were identical in topology, which
means that each hypothesis test involved a unique pair of
trees being compared.
The four phylogenetic null hypotheses were tested using a
parametric bootstrap approach (Efron 1985, Felsenstein 1988,
Huelsenbeck, Hillis & Jones 1996, Swofford et al. 1996 : 506,
Hillis, Mable & Moritz 1996 : 523–526, Huelsenbeck, Hillis &
Nielsen 1996). Under a parametric bootstrap, a null distribution
of the null hypothesis is generated by recording the difference
in length}likelihood between the optimal topology and the
best topology agreeing with a null hypothesis for each of
many data sets that were simulated along the best null
hypothesis tree from the original data set. The difference in
length or likelihood should be zero in the ideal case, but due
to stochastic variation or systematic error of the reconstruction
method this is not necessarily so. The difference in
length}likelihood obtained from the original data set (δ) can
then be compared to this null distribution, and the probability
of the null hypothesis being true can be calculated. The
popular Kishino–Hasegawa (Kishino & Hasegawa 1989) and
Templeton tests (Templeton 1983, Felsenstein 1985) were not
employed, since the null distributions that these tests rely on
apply only when the trees being compared can be considered
drawn at random from the population of all possible trees, i.e.
when the trees are fully specified a priori (Goldman, Anderson
& Rodrigo 2000). Consequently, these tests are inapplicable
when one or both of the trees being compared originate from
the analysis of the data set at hand. In fact, most published
applications of these tests are invalid.
The parametric bootstrap was performed in the following
way : 249 data sets per null hypothesis (1–4) per optimality
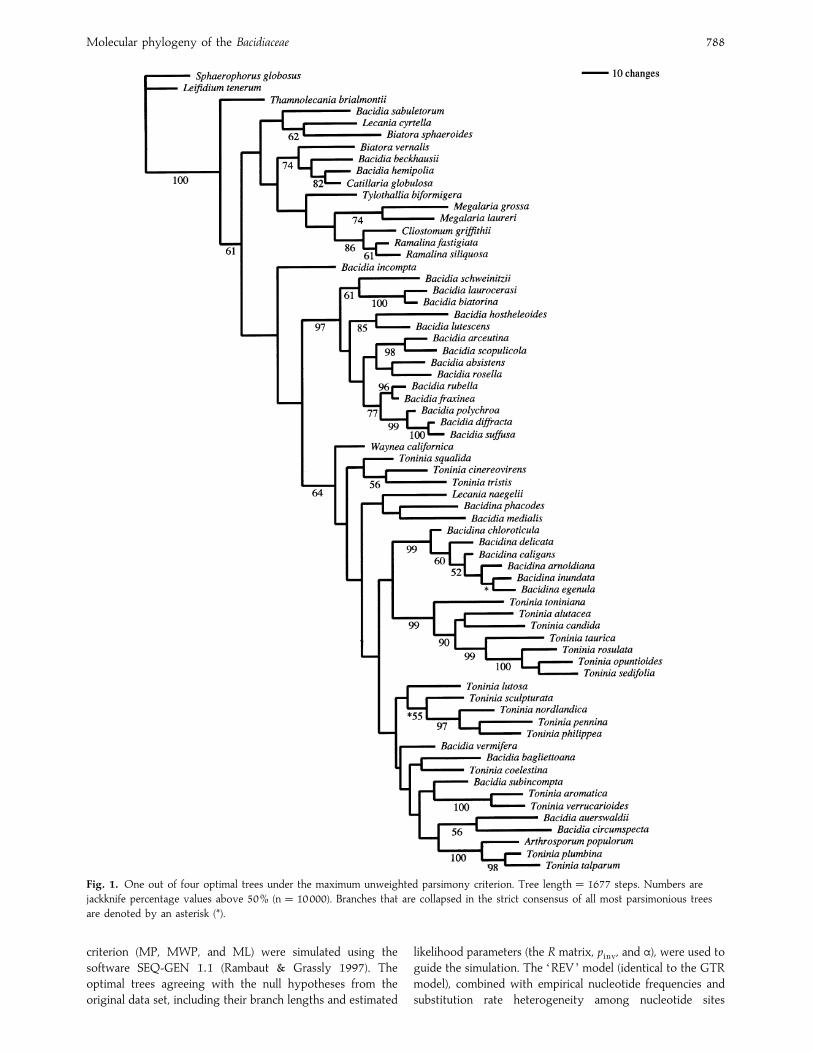
Molecular phylogeny of the Bacidiaceae 788
Fig. 1. One out of four optimal trees under the maximum unweighted parsimony criterion. Tree length¯ 1677 steps. Numbers are
jackknife percentage values above 50% (n¯ 10000). Branches that are collapsed in the strict consensus of all most parsimonious trees
are denoted by an asterisk (*).
criterion (MP, MWP, and ML) were simulated using the
software SEQ-GEN 1±1 (Rambaut & Grassly 1997). The
optimal trees agreeing with the null hypotheses from the
original data set, including their branch lengths and estimated
likelihood parameters (the R matrix, pinv
, and α), were used to
guide the simulation. The ‘REV ’ model (identical to the GTR
model), combined with empirical nucleotide frequencies and
substitution rate heterogeneity among nucleotide sites

S. Ekman 789
Fig. 2. One out of three optimal trees under the maximum weighted parsimony criterion. Tree length¯ 2644±1 steps. A symmetric
step matrix was used, which included weights obtained by charting the absolute average number of changes on 100 equiprobable
random trees (see text). Numbers are jackknife percentage values above 50% (n¯ 300). Branches that are collapsed in the strict
consensus of all most parsimonious trees are denoted by an asterisk (*).

Molecular phylogeny of the Bacidiaceae 790
Fig. 3. The single optimal tree under the maximum likelihood criterion. –Ln likelihood¯ 8166.1434. A GTRIdΓ6 model was used,
since it was shown to best explain the observed data according to a likelihood ratio test. Numbers are jackknife percentage values
above 50% (n¯ 100). An asterisk (*) denotes a branch that is not significantly longer than zero under a likelihood ratio test with full
reoptimization after forcing a branch length to zero. The nomenclaturally correct generic names of some clades discussed in the text and
supported by this tree (sometimes also the unweighted and weighted parsimony trees) have been marked. Light grey (as opposed to
dark grey) shading refers to optional inclusion. Informal names of species groups refer to the discussion of Ekman (1996a : 45–46).

S. Ekman 791
according to a continuous gamma model, was used. The
length of the simulated sequences corresponded to the
number of potentially variable sites in the original matrix, i.e.
505¬(1®pinv
) (invariable sites were not added to the
matrices, since only parsimony was used for the analysis ; see
below). A total of (four null hypotheses¬three optimality
criteria¬249¯) 2988 new matrices were simulated.
Each of the (4 null hypotheses¬249¯) 996 data sets
simulated along the optimal constrained MP trees was
analysed in an unconstrained analysis (100 random-addition
sequence replicates, TBR branch-swapping, MulTrees option
on, steepest descent, collapse zero-length branches options
off, nchuck¯ 500, and chuckscore¯ 1), and in a constrained
analysis (same search parameters, the constraint being the
same as the one used to obtain the tree used for simulation).
To facilitate analysis, it was executed in PAUP* as a
continuous series of 996 DATA blocks, each followed by a
PAUP block containing the desired settings and search
commands. For each of the 249 data sets representing a null
hypothesis, the difference between the optimal unconstrained
and the optimal constrained tree was recorded and plotted in
a histogram.
Each of the 996 data sets simulated along the optimal
constrained MWP trees was analysed in a similar way, except
that character state changes were weighted using the same
step matrix as in the initial search for the globally most
optimal MWP tree.
Ideally, the 996 data sets simulated along the optimal
constrained ML trees should be analyzed under maximum
likelihood, since the difference in likelihood between the
optimal unconstrained and constrained trees obtained from
the original data matrix was calculated under this optimality
criterion. However, such an analysis is unrealistic and would
take years to complete even with very moderately aggressive
search parameters. Therefore, differences in parsimony scores
were used in place of differences in likelihood scores for
testing the null hypotheses obtained under the ML optimality
criterion. Although a switch in the optimality criterion can be
motivated, a slight increase in the stochastic variation of the
test can be anticipated, since the difference in parsimony score
will sometimes underestimate and sometimes overestimate
the difference in likelihood scores.
The probability of a null hypothesis being correct is
r}(m1), where r is the rank order of δ among all score
differences and m is the number of bootstrap replicates
(Goldman 1993). Raw probability values were Dunn–S) ida! kcorrected for multiple comparisons (Sokal & Rohlf 1995) to
maintain a total type I error rate of 0±05.The complete analysis as described here, including some
mistakes in need of correction, required approximately four
and a half months of dedicated CPU time.
RESULTS
The final alignment of the 67 taxa was 752 positions in length.
Altogether 247 positions representing 25 blocks of contiguous
alignment sites were excluded due to alignment problems.
The number of included alignment positions was 505. In this
Table 2. A comparison of optimal trees obtained under maximum
parsimony (MPa-d), maximum weighted parsimony (WMPa-c), and
maximum likelihood (MLa).
Tree
Tree length (un-
weighted parsimony)
Tree length
(weighted parsimony) -Ln likelihood
MPa 1677 2654±2 8189±2804MPb 1677 2655±4 8189±4138MPc 1677 2656±6 8189±5331MPd 1677 2657±8 8189±6683MWPa 1690 2644±1 8174±7531MWPb 1690 2644±1 8175±4004MWPc 1690 2644±1 8175±4004MLa 1701 2673±5 8166±1434
The likelihood of each tree was calculated using a GTRIdΓ6 model
with empirical base frequencies, and the R matrix, proportion of invariable
sites pinv
, and the gamma curve shape parameter α estimated.
alignment, 285 characters were variable, and 212 were
parsimony-informative.
The test for phylogenetic signal was highly significant
(βnull
¯ 17±7, βobs
¯ 28±0, ..¯ 2012, rooted tRASA
¯ 38±5,P' 0±001), and it was concluded that the matrix contained a
significant amount of hierarchical signal potentially useful for
phylogenetic analysis.
The heuristic search under the maximum unweighted
parsimony optimality criterion yielded four equally par-
simonious trees of 1677 steps, all with consistency index (CI)
¯ 0±26, retention index (RI)¯ 0±50, and rescaled consistency
index (RC)¯ 0±13 (indices calculated with uninformative
characters excluded). One of the most parsimonious trees,
with jackknife branch support, is shown in Fig. 1. The heuristic
search under the maximum weighted parsimony optimality
criterion yielded three equally parsimonius trees of 2644±1steps, all with consistency index (CI)¯ 0±26, retention index
(RI)¯ 0±52, and rescaled consistency index (RC)¯ 0±14. One
of these trees, with jackknife branch support, is shown in Fig.
2. The heuristic search under the maximum likelihood
optimality criterion yielded a single most likely tree with ln
likelihood¯®8166±1434. Likelihood parameters were rAC
¯2±66, r
AG¯ 3±93, r
AT¯ 1±93, r
CG¯ 0±76, r
CT¯ 7±29 (as-
suming rGT
¯ 1), pinv
¯ 0±19, and α¯ 0±58. This tree, with
jackknife support values, is shown in Fig. 3. The chi-square test
of homogeneity of base frequencies across taxa revealed no
significant differences (χ#¯ 68±3, ..¯ 138, P¯ 1±00). The
optimal trees obtained under the three optimality criteria are
being compared in Table 2. Figs 4–7 demonstrate the outcome
of the parametric bootstrap analyses of the four null
hypotheses under maximum unweighted parsimony, maxi-
mum weighted parsimony, and maximum likelihood. Dunn–
S) ida! k corrected probabilities of the null hypothesis being
correct are provided in Table 3. The parametric bootstrap
rejected the monophyly of Bacidia, Toninia, and the Rama-
linaceae no matter what optimality criterion had been used to
generate the tree representing the null hypotheses. The
monophyly of Bacidina, on the other hand, was not rejected
under any optimality criterion.
DISCUSSION
Common to the optimal trees obtained under maximum
unweighted and weighted parsimony as well as maximum
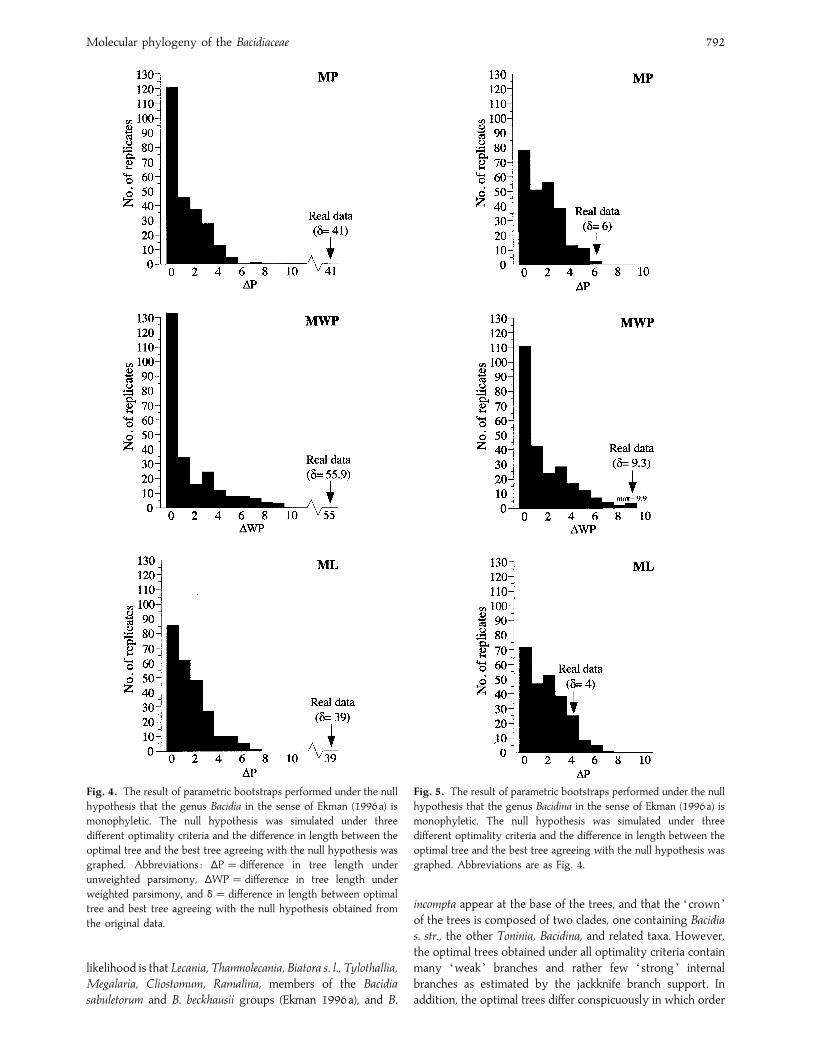
Molecular phylogeny of the Bacidiaceae 792
Fig. 4. The result of parametric bootstraps performed under the null
hypothesis that the genus Bacidia in the sense of Ekman (1996a) is
monophyletic. The null hypothesis was simulated under three
different optimality criteria and the difference in length between the
optimal tree and the best tree agreeing with the null hypothesis was
graphed. Abbreviations : ∆P¯ difference in tree length under
unweighted parsimony, ∆WP¯ difference in tree length under
weighted parsimony, and δ¯ difference in length between optimal
tree and best tree agreeing with the null hypothesis obtained from
the original data.
likelihood is that Lecania, Thamnolecania, Biatora s. l., Tylothallia,
Megalaria, Cliostomum, Ramalina, members of the Bacidia
sabuletorum and B. beckhausii groups (Ekman 1996a), and B.
Fig. 5. The result of parametric bootstraps performed under the null
hypothesis that the genus Bacidina in the sense of Ekman (1996a) is
monophyletic. The null hypothesis was simulated under three
different optimality criteria and the difference in length between the
optimal tree and the best tree agreeing with the null hypothesis was
graphed. Abbreviations are as Fig. 4.
incompta appear at the base of the trees, and that the ‘crown ’
of the trees is composed of two clades, one containing Bacidia
s. str., the other Toninia, Bacidina, and related taxa. However,
the optimal trees obtained under all optimality criteria contain
many ‘weak ’ branches and rather few ‘strong ’ internal
branches as estimated by the jackknife branch support. In
addition, the optimal trees differ conspicuously in which order

S. Ekman 793
Fig. 6. The result of parametric bootstraps performed under the null
hypothesis that the genus Toninia in the sense of Timdal (1991) is
monophyletic. The null hypothesis was simulated under three
different optimality criteria and the difference in length between the
optimal tree and the best tree agreeing with the null hypothesis was
graphed. Abbreviations are as Fig. 4.
some clades appear in the tree. The position of a number of
species is notoriously unstable, e.g. Bacidia bagliettoana, B.
incompta, Thamnolecania brialmontii, and Waynea californica.
Presumably, the rather different trees obtained under the
different optimality criteria is a result of substantial homoplasy
in the matrix (as indicated by, e.g. the low CI, RI, and RC), by
Fig. 7. The result of parametric bootstraps performed under the null
hypothesis that the family Ramalinaceae auct. is monophyletic and
distinct from a monophyletic Bacidiaceae. The null hypothesis was
simulated under three different optimality criteria and the difference
in length between the optimal tree and the best tree agreeing with
the null hypothesis was graphed. Abbreviations are as Fig. 4.
substantially different nucleotide substitution rates (as indi-
cated by the R matrix of the most likely tree ; CT substitutions
are approximately ten timesmore likely thanCG substitutions),
and by substantial rate heterogeneity among nucleotide sites
(as indicated by the rather low value of α, 0±58, of the most
likely tree). The two latter factors are known to create more

Molecular phylogeny of the Bacidiaceae 794
Table 3. Probabilities of null hypotheses 1–4 being correct under unweighted and weighted parsimony, and maximum likelihood. ∆P and ∆WP refer to
differences in unweighted or weighted parsimony scores between constrained and optimal trees obtained from simulated data matrices. Raw probabilities
(calculated according to the formula of Goldman 1993) were Dunn-S) ida! k corrected for multiple comparisons (Sokal & Rohlf 1995) to maintain a total
type I error rate of 0±05. The actual null hypothesis at test was that the best tree agreeing with the null hypothesis (the constrained tree) does not differ
significantly from the globally best tree (given an optimality criterion). Hypotheses were rejected if p% 0±05. When the rank order of δ is an interval, p
was calculated from the worst rank (see text for the calculation of p). The lowest probability possible to obtain in this experiment is p¯ 0±047.
Optimality
criterion
Difference of tree
length}likelihood (δ)
Highest ∆P or ∆WP
recorded in simulation Rank order of δ p
(1) Bacidia in the sense of Ekman (1996a) is monophyletic
MP 41 7 1 0±047*MWP 55±9 9±9 1 0±047*ML 39}97±8947 7 1 0±047*
(2) Bacidina in the sense of Ekman (1996a) is monophyletic
MP 6 6 1–3 0±135MWP 9±3 9±9 2 0±091ML 4}3±5009 7 15–40 0±877
(3) Toninia in the sense of Timdal (1991) is monophyletic
MP 25 7 1 0±047*MWP 37±3 10±6 1 0±047*ML 23}63±7593 8 1 0±047*
(4) Ramalinaceae auct. is monophyletic
MP 15 9 1 0±047*MWP 22±9 10±2 1 0±047*ML 14}22±1206 8 1 0±047*
problems to maximum parsimony analysis than to maximum
likelihood analysis, at least when a correct likelihood model is
used (e.g. Yang 1996a,b, 1998). Although still susceptible to
errors due to rate heterogeneity among nucleotide sites,
weighted parsimony is less adversely affected than unweighted
parsimony by unbalanced nucleotide substitution rates, and it
was considered to ‘often provide a close approximation of the
likelihood solutions ’ by Swofford et al. (1996). A parameter-
rich maximum likelihood model like the one used here is
assumed to be more or less immune to these problems. On the
other hand, the likelihood model used may be a poor predictor
of the actual evolutionary events, and even if a correct model
is used, cases are known where other models may reconstruct
the true phylogeny with greater accuracy (Yang 1997). In
addition, likelihood parameter estimates are just estimates ;
they contain errors that may add up to produce an incorrect
phylogeny. If the maximum likelihood tree obtained in this
study is anywhere near the true phylogeny, with several short
internal branches (18 of which are not significantly longer than
zero), the hope of ever obtaining an accurate and well-
supported estimate of the phylogeny of the Bacidiaceae may
have been thwarted by nature itself.
Despite problems with phylogeny estimation and low
branch support, the data set contains sufficient information to
discriminate between alternative phylogenetic scenarios. The
hypotheses that Bacidia in the sense of Ekman (1996a) and
Toninia in the sense of Timdal (1991) are monophyletic were
emphatically rejected by the parametric bootstraps irrespective
of optimality criterion (hypothesis 1 and 3 of Table 3 ; Figs 4,
6), despite the fact that these monographers excluded
numerous species that were previously referred to the two
genera.
A suitable candidate for a redefined Bacidia s. str. obtained
in this study is the clade containing B. hostheleoides and B.
lutescens at the bottom (Figs 1–3). This clade has a 97–100%
jackknife support depending on optimality criterion. The
overall problem with Bacidia in the sense of Ekman (1996a)
seems to be that too many species have been included. In
particular, species with blue-green pigmentation in the
epithecium and}or with fusiform or bacilliform spores (as
opposed to acicular, i.e. with one blunt end and one tapering
end) appear not to be closely related to Bacidia s. str. as
suggested here, but rather to species groups referred to
Toninia as circumscribed by Timdal (1991). Unlike most other
species in need of exclusion from Bacidia s. str., B. medialis has
usually pale apothecia with little pigment. It was considered to
belong to Bacidia s. str. and being closely related to B.
hostheleoides by Ekman (1996a), although a difference in
structure of the proper exciple between the two species was
noted. In the present study, however, it becomes apparent
that B. medialis has little to do with B. hostheleoides, the former
being more closely allied with Bacidina phacodes and Lecania
naegelii. In addition, the affinities of Bacidia incompta to Bacidia
s. str. remain doubtful. As suggested by Ekman (1996a), the B.
beckhausii group appears to be distantly related to Bacidia s. str.
and possibly represents an as yet undescribed genus (although
poorly supported as revealed by jackknife values ; Figs 1–3).
However, Catillaria globulosa seems to belong in this group as
well. This species bears an overall striking similarity with the
other species of the group, except that the spores are 1-septate
instead of 3-septate. Catillaria globulosa was referred to Bacidia
by Wirth (1987), and this treatment has been followed by a
number of authors. The data also lends support to the
exclusion of B. sabuletorum and its relatives from Bacidia s. str.
(Ekman 1996a). Instead, the B. sabuletorum group seems to be
more closely related to Thamnolecania and Lecania (assuming
that L. cyrtella is a member of Lecania s. str.). Unlike the
prediction by Ekman (1996a), however, the B. lutescens group

S. Ekman 795
appears not to bear any close relationship with the B.
beckhausii group, but instead seems to be a part of Bacidia s. str.
Toninia as understood by Timdal (1991) appears to be a
polyphyletic assemblage of species. Apparently, some species
groups are more closely related to taxa traditionally referred
to Arthrosporum, Bacidia, Bacidina, or Waynea than they are to
each other. Unfortunately, the data does not contain resolving
power enough to detail any alternative taxonomic framework
for this group of taxa. The data suggests, however, that some
of the generic names treated as synonyms of Toninia (Timdal
1991 : 30) may have to be resurrected in the future, e.g.
Thalloidima (Fig. 3). A few well-supported groups (here taken
to mean a jackknife support of ca 75% or more under all
optimality criteria) can be discerned (Figs 1–3) : (1) Arthro-
sporum populorum, Toninia plumbina, and T. talparum ; (2)
Toninia aromatica and T. verrucarioides ; (3) Toninia nordlandica,
T. pennina, and T. philippea ; and (4) Toninia toniniana, T.
alutacea, T. candida, T. taurica, T. opuntioides, T. sedifolia, and T.
rosulata. The first group would constitute a slightly expanded
version of the genus Arthrosporum, A. populorum being the
type species, whereas the last group would constitute the
genus Thalloidima, the type species of which is T. candida. The
circumscription of Toninia s. str. is very uncertain. The optimal
trees obtained in this study suggest that Toninia would
include T. cinereovirens (the type species), T. squalida, T. tristis,
and possibly also one or a few species previously referred to
Bacidia, viz. B. bagliettoana, B. auerswaldii, and B. circumspecta.
Great care should be taken in the interpretation, however. If
the position of Bacidina}Woessia}Lecania naegelii as nested
within clades dominated by species of Toninia turns out to be
artefactual, Toninia can be conveniently redefined and become
monophyletic by including in it Arthrosporum, the majority of
misclassified Bacidia species with blue-green pigmentation in
the epithecium and}or with fusiform or bacilliform spores, and
possibly also Waynea.
The case for or against a monophyletic genus Bacidina is
more uncertain. The parametric bootstrap test does not reject
the null hypothesis of a monophyletic Bacidina under any
optimality criterion (hypothesis 2 of Table 3 ; Fig. 5). This
should not be taken to mean that Bacidina is necessarily
monophyletic, just that the data set at hand does not contain
the necessary resolving power to tell whether the genus is
monophyletic or not. A ‘core group ’ of species, Bacidina
chloroticula, B. delicata, B. arnoldiana, B. inundata, B. egenula, and
Bacidia caligans constitute a well-supported group (with
98–100% jackknife support depending on optimality criterion ;
Figs 1–3). This group of species is sometimes referred to
Woessia, the type species being W. fusarioides, a synonym of
Bacidina arnoldiana or W. arnoldiana (see e.g. Diederich &
Se! rusiaux 2000). B. phacodes, the type species of Bacidina, may
or may not be associated with this group. It is interesting to
note that in all of the optimal trees, B. phacodes is more or less
closely associated with Lecania naegelii, since these species are
the only known representatives of the Bacidiaceae that possess
straight macroconidia with multiple transverse septa. Bacidina
may or may not be synonymous with Woessia, and one or
both of Bacidia medialis and Lecania naegelii may have to be
included in Bacidina. It has been proposed to conserve Bacidina
against Woessia if these names are congeneric (Ekman 1996b).
The null hypothesis that the Ramalinaceae is monophyletic
and distinct from a monophyletic Bacidiaceae was rejected by
the parametric bootstraps under all optimality criteria
(hypothesis 4 of Table 3 ; Fig. 7). The jackknife support for
Cliostomum being the sistergroup of Ramalina is very high in
this study, 86–97% depending on optimality criterion (Figs
1–3). However, this should not be immediately interpreted to
mean that Cliostomum is the closest living crustose relative of
the Ramalinaceae, since Ramalinora glaucolivida (Lumbsch et al.
1995) was not part of the study. Accomodating the fruticose
Ramalinaceae and the basically crustose Bacidiaceae in the same
family may seem surprising, but disregarding growth habit,
other morphological characters are similar (see ‘ Introduction ’).
Other families in which the presence of several growth forms
have been confirmed by molecular evidence include the
Roccellaceae (Myllys et al. 1999) and Physciaceae (Lohtander
2000). If the status of the Bacidiaceae and Ramalinaceae as
synonyms is confirmed by further testing, this has implications
for nomenclature, since Ramalinaceae (described by Agardh
1821 : 93) is much older than Bacidiaceae as well as
Phyllopsoraceae and Biatoraceae. Interestingly, the best tree
under weighted parsimony would theoretically allow the
delimitation of a Ramalinaceae in a new, wider sense, and
the Bacidiaceae in a narrower sense without impairing the
monophyly of any of the families (as long as Bacidia incompta
is excluded from both families). The Ramalinaceae in this sense
would consequently have to include Tylothallia, Megalaria,
Cliostomum, Thamnolecania, Biatora, the Bacidia sabuletorum
and B. beckhausii groups, and at least parts of Lecania in
addition to Ramalina and possibly other fruticose genera now
referred to the Ramalinaceae that were not part of this
investigation. However, in such a classification it would be
exceedingly difficult to tell the families apart on morphology.
Synonymizing Ramalinaceae and Bacidiaceae then seems like a
more attractive alternative.
A few additional observations emanate from this data set,
but without having been properly treated in a framework of
phylogenetic hypothesis testing. First of all, the data does not
lend support to the notion that Thamnolecania may be
congeneric with Lecania (Hafellner 1984). Biatora vernalis, the
type species of Biatora, forms a monophyletic group together
with the Bacidia beckhausii group (including Catillaria globulosa)
with high jackknife support (74–88% depending on optimality
criterion ; Figs 1–3). On the other hand, B. sphaeroides, which
is sometimes treated as a member of Biatora (e.g. Coppins
1992), is not immediately associated with B. vernalis. This is
congruent with the idea laid out by Printzen (1995) that B.
sphaeroides, along with a few others (i.e. B. carneoalbida, B.
epixanthoides, and B. tetramera), belong in a separate genus,
Mycobilimbia. Later, Printzen & Lumbsch (2000) informally re-
included Mycobilimbia in Biatora. However, there is no real
incongruence between the tree topologies presented in that
work and the ones presented here, since the studies differ
significantly in taxon sampling. Megalaria grossa, the type
species of the genus, and M. laureri form a monophyletic
group with high jackknife support (74–94% depending on
optimality criterion ; Figs 1–3), which lends some support to
the generic circumscription of Megalaria proposed by Ekman
& Tønsberg (1996). Finally, the genus Lecania as treated, for

Molecular phylogeny of the Bacidiaceae 796
example, by Ekman (1996a) and Diederich & Se! rusiaux (2000),
appears to be polyphyletic. Unfortunately, material of the
type species of the genus, L. fuscella, was not available for
sequencing. All of these observations, however, need to be
properly tested with better taxon representation than was
possible here.
The most common use for the ITS rDNA region in
phylogenetics is at the interspecific level (Bridge & Hawks-
worth 1998). However, a study by Hershkovitz & Lewis
(1996) has demonstrated that ITS variation is non-random at
very deep phylogenetic levels, and that it contains conserved
motifs that can resolve even interkingdom relationships. The
study presented here demonstrates that the ITS region can aid
in resolving phylogeny at the intergeneric level within a
family of ascomycetes. There is a price attached to this goal,
however : a fairly large amount of ambiguous alignment had
to be excluded. In addition, phylogenetic reconstruction is
complicated by difficult conditions (unbalanced nucleotide
substitution rates, rate heterogeneity among nucleotide sites)
and rather poor resolution (many short internal branches, low
branch support). Furthermore, taxon sampling is still rather
uneven. The representation of taxa in this study that have
been referred to Bacidia, Bacidina, and Toninia is fairly good.
Other groups are still poorly sampled, particularly members of
potentially ‘basal ’ taxa, (e.g. Lecania, Biatora, Phyllopsora, and
the Ramalinaceae auct.). Before any definite conclusions on the
phylogeny of the Bacidiaceae can be drawn, and the
accompanying nomenclatural novelties can be proposed, more
DNA sequence data from more taxa is needed.
ACKNOWLEDGEMENTS
I am indebted to Christian Printzen for providing me with valuable comments
on the manuscript, to O$ rjan Fritz, Ha/ kon Holien, Per Johansson, Thomas
Johansson, Tommy Knutsson, Mark Seaward, and the curators of AAS, BG,
MIN, and O for providing me with freshly collected herbarium material for
DNA sequencing, and to Mats Wedin for letting me use his Sphaerophoraceae
sequences before they were published in GenBank. I gratefully acknowledge
the staff at the Sequencing Facility at the University of Bergen. I thank the
Research Council of Norway for financial support, and Dag E. Helland for
providing me with space and facilities in his laboratory.
REFERENCES
Agardh, C. A. (1821) Aphorismi botanici. [Dissertation No. 7.] Berling, Lund.
Bridge, P. D. & Hawksworth, D. L. (1998) What molecular biology has to tell
us at the species level in lichenized fungi. Lichenologist 30 : 307–320.
Coppins, B. J. (1992) Biatora Fr. (1817). In The Lichen Flora of Great Britain and
Ireland (O. W. Purvis, B. J. Coppins, D. L. Hawksworth, P. W. James &
D. M. Moore, eds) : 118–121. Natural History Museum Publications,
London.
Diederich, P. & Se! rusiaux, E. (2000) The Lichens and Lichenicolous Fungi of
Belgium and Luxembourg. An annotated checklist. Muse! e National d’Histoire
Naturelle, Luxembourg.
Durbin, R., Eddy, S., Krogh, A. & Mitchison, G. (1998) Biological Sequence
Analysis.Probabilisticmodels of proteins and nucleic acids. CambridgeUniversity
Press, Cambridge, UK.
Efron, B. (1985) Bootstrap confidence intervals for a class of parametric
problems. Biometrika 72 : 45–58.
Ekman, S. (1996a) The corticolous and lignicolous species of Bacidia and
Bacidina in North America. Opera Botanica 127 : 1–148.
Ekman, S. (1996b) Proposal to conserve the name Bacidina against Lichingoldia
and Woessia (lichenized Ascomycotina). Taxon 45 : 687–688.
Ekman, S. (1997) The genus Cliostomum revisited. Symbolae Botanicae
Upsalienses 32 (1) : 17–28.
Ekman, S. & Tønsberg, T. (1996) A new species of Megalaria from the North
American West Coast, and notes on the generic circumscription. The
Bryologist 99 : 34–40.
Ekman, S. & Wedin, M. (2000) The phylogeny of the families Lecanoraceae and
Bacidiaceae (lichenized Ascomycota) inferred from nuclear SSU rDNA
sequences. Plant Biology 2 : 350–360.
Eriksson, O. E. & Hawksworth, D. L. (1987) Outline of the ascomycetes –
1987. Systema Ascomycetum 6 : 259–337.
Eriksson, O. E. & Hawksworth, D. L. (1993) Outline of the ascomycetes –
1993. Systema Ascomycetum 12 : 51–257.
Eriksson, O. E. & Hawksworth, D. L. (1998) Outline of the ascomycetes –
1998. Systema Ascomycetum 16 : 83–296.
Farris, J. S., Albert, V. A., Ka$ llersjo$ , M., Lipscomb, D. & Kluge, A. G. (1996)
Parsimony jackknifing outperforms neighbour-joining. Cladistics 12 :
99–124.
Felsenstein, J. (1985) Confidence limits on phylogenies with a molecular clock.
Systematic Zoology 34 : 152–161.
Felsenstein, J. (1988) Phylogenies from molecular sequences : inference and
reliability. Annual Review of Genetics 22 : 521–565.
Gardes, M. & Bruns, T. D. (1993) ITS primers with enhanced specificity for
basidiomycetes – application to the identification of mycorrhizae and rusts.
Molecular Ecology 2 : 113–118.
Gargas, A., DePriest, P. T. & Taylor, J. W. (1995) Positions of multiple
insertions in SSU rDNA of lichen-forming fungi. Molecular Biology and
Evolution 12 : 208–218.
Goldman, N. (1993) Statistical tests of models of DNA substitution. Journal of
Molecular Evolution 36 : 182–198.
Goldman, N., Anderson, J. P. & Rodrigo, A. G. (2000) Likelihood-based tests
of topologies in phylogenetics. Systematic Biology 49 : 652–670.
Greuter, W., McNeill, J., Barrie, F. R., Burdet, H. M., Demoulin, V., Filgueiras,
T. S., Nicolson, D. H., Silva, P. C., Skog, J. E., Trehane, P., Turland, N. J. &
Hawksworth, D. L. (2000) International Code of Botanical Nomenclature (Saint
Louis Code). [Regnum Vegetabile Vol. 138.] Koeltz Scientific Books,
Ko$ nigstein.
Gu, X., Fu, Y.-X. & Li, W.-H. (1995) Maximum likelihood estimation of the
heterogeneity of substitution rate among nucleotide sites. Molecular Biology
and Evolution 12 : 546–557.
Hafellner, J. (1984) Studien in Richtung einer natu$ rlicheren Gliederung der
Sammelfamilien Lecanoraceae und Lecideaceae. Nova Hedwigia, Beiheft 79 :
241–371.
Hafellner, J. (1988) Principles of classification and main taxonomic groups. In
Handbook of lichenology (M. Galun, ed.) 3: 41–52. CRC Press, Boca Raton,
FL.
Hershkovitz, M. A. & Lewis, L. A. (1996) Deep-level diagnostic value of the
rDNA-ITS region. Molecular Biology and Evolution 13 : 1276–1295.
Hertel, H. & Rambold, G. (1985) Lecidea sect. Armeniacae : lecideoide Arten der
Flechtengattung Lecanora und Tephromela (Lecanorales). Botanische JahrbuX cher107 : 469–501.
Hertel, H. & Rambold, G. (1995) On the genus Adelolecia (lichenized
Ascomycotina, Lecanorales). Bibliotheca Lichenologica 57 : 211–230.
Hillis, D. M., Mable, B. K. & Moritz, C. (1996) Applications of molecular
systematics : the state of the field and a look to the future. In Molecular
Systematics (D. M. Hillis, C. Moritz & B. K. Mable, eds) : 515–543. 2nd edn.
Sinauer Associates, Sunderland, MA.
Holmgren, P. K., Holmgren, N. H. & Barnett, L. C. (1990) Index Herbariorum.
Part I : The herbaria of the world. New York Botanical Garden, New York,
NY
Huelsenbeck, J. P. & Crandall, K. A. (1997) Phylogeny estimation and
hypothesis testing using maximum likelihood. Annual Review of Ecology and
Systematics 28 : 437–466.
Huelsenbeck, J. P., Hillis, D. M. & Jones, R. (1996) Parametric bootstrapping
in molecular phylogenetics : applications and performance. In Molecular
Zoology : advances, strategies and protocols (J. D. Ferraris & S. R. Palumbi, eds) :
19–45. Wiley-Liss, New York, NY.
Huelsenbeck, J. P., Hillis, D. M. & Nielsen, R. (1996) A likelihood-ratio test of
monophyly. Systematic Biology 45 : 546–558.
Huelsenbeck, J. P. & Rannala, B. (1997) Phylogenetic methods come of age :
testing hypotheses in an evolutionary context. Science 276 : 227–232.

S. Ekman 797
Hughey, R., Karplus, K. & Krogh, A. (1999) SAM: Sequence alignment and
modeling software system. Baskin Center for Computer Engineering and
Science, University of California, Santa Cruz, CA. Internet : http :}}www.cse.ucsc.edu}research}compbio}papers}samjdoc}samjdoc.html.
Hughey, R. & Krogh, A. (1996) Hidden Markov models for sequence analysis :
extension and analysis of the basic method. Computer Applications in the
Biosciences 12 : 95–107.
Kishino, H. & Hasegawa, M. (1989) Evaluation of the maximum likelihood
estimate of the evolutionary tree topologies from DNA sequence data, and
the branching order in Hominoidea. Journal of Molecular Evolution 29 :
170–179.
Krogh, A., Brown, M., Mian, I. S., Sjo$ lander, K. & Haussler, D. (1994) Hidden
Markovmodels in computational biology : applications to protein modeling.
Journal of Molecular Biology 235 : 1501–1531.
Lohtander, K. (2000) Phylogeny and species delimitation in Roccellaceae and
Physciaceae (Euascomycetes). Doctoral dissertation, Botaniska Institutionen,
Stockholms Universitet, Stockholm.
Lumbsch, H. T., Rambold, G. & Elix, J. A. (1995) Ramalinora (Ramalinaceae) – a
new lichen genus from Australia. Australian Systematic Botany 8 : 521–530.
Lutzoni, F. M. (1997) Phylogeny of lichen- and non-lichen-forming omphalin-
oid mushrooms and the utility of testing combinability among multiple
data sets. Systematic Biology 46 : 373–406.
Lyons-Weiler, J. (1999) RASA 2±3 for Macintosh and manual. Pennsylvania
State University, PA. Internet : http.}}test1.bio.psu.edu}LW}rasatext.html.
Lyons-Weiler, J., Hoelzer, G. A. & Tausch, R J. (1996) Relative apparent
synapomorphy analysis (RASA) I : The statistical measurement of
phylogenetic signal. Molecular Biology and Evolution 13 : 749–757.
Maddison, W. P. & Maddison, D. R. (1992) MacClade Version 3. Sinauer
Associates, Sunderland, MA.
Myllys, L., Lohtander, K., Ka$ llersjo$ , M. & Tehler, A. (1999) Applicability of
ITS data in Roccellaceae (Arthoniales, Euascomycetes) phylogeny. The
Lichenologist 31 : 461–476.
Posada, D. & Crandall, K. A. (1998) MODELTEST: testing the model of DNA
substitution. Bioinformatics 9 : 817–818.
Printzen, C. (1995) Die Flechtengattung Biatora in Europa. Bibliotheca
Lichenologica 60 : 1–275.
Printzen, C. & Lumbsch, H. T. (2000) Molecular evidence for the diversification
of extant lichens in the Late Cretaceous and Tertiary. Molecular Phylogenetics
and Evolution 17 : 379–387.
Rambaut, A. & Grassly, N. C. (1997) Seq-Gen : an application for the Monte
Carlo simulation of DNA sequence evolution along phylogenetic trees.
Computer Applications in the Biosciences 13 : 235–238.
Rambold, G. (1989) A monograph of the saxicolous lecideoid lichens of
Australia (excl. Tasmania). Bibliotheca Lichenologica 34 : 1–345.
Rambold, G. & Triebel, D. (1992) The inter-lecanoralean associations.
Bibliotheca Lichenologica 48 : 1–201.
Rodrı!guez, F., Oliver, J. F., Marin, A. & Medina, J. R. (1990) The general
stochastic model of nucleotide substitution. Journal of Theoretical Biology.
142 : 485–501.
Sokal, R. R. & Rohlf, F. J. (1995) Biometry. 3rd edn. W. H. Freeman, New York.
Stizenberger, E. (1862) Beitrag zur Flechtensystematik. Bericht uX ber die
ThaX tigkeit der St Gallischen naturwissenschaftlichen Gesellschaft 1861–1862 :
124–182.
Swofford, D. L. (1999) PAUP*. Phylogenetic Analysis Using Parsimony (*and
Other Methods). Version 4. Sinauer Associates, Sunderland, MA.
Swofford, D. L., Olsen, G. J., Waddell, P. J. & Hillis, D. M. (1996) Phylogenetic
inference. In Molecular Systematics (D. M. Hillis, C. Moritz & B. K. Mable,
eds) : 407–514. 2nd ed. Sinauer Associates, Sunderland, MA.
Templeton, A. R. (1983) Phylogenetic inference from restriction endonuclease
cleavage site maps with particular reference to the evolution of humans and
the apes. Evolution 37 : 221–244.
Timdal, E. (1991) A monograph of the genus Toninia (Lecideaceae, Ascomycetes).
Opera Botanica 110 : 1–137.
Vobis, G. (1980) Bau und Entwicklung der Flechten-Pycnidien und ihrer
Conidien. Bibliotheca Lichenologica 14 : 1–141.
Watson, W. (1929) The classification of lichens. New Phytologist 28 : 1–36.
White, T. J., Bruns, T. D., Lee, S. & Taylor, J. (1990) Amplification and direct
sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR
Protocols : a guide to methods and applications (M. A. Innis, D. H. Gelfand, J. J.
Sninsky & T. J. White, eds) : 315–322. Academic Press, San Diego, CA.
Wirth, V. (1987) Die Flechten Baden-WuX rttembergs. Eugen Ulmer, Stuttgart,
Germany.
Yang, Z. (1993) Maximum-likelihood estimation of phylogeny from DNA
sequences when substitution rates differ over sites. Molecular Biology and
Evolution 10 : 1396–1401.
Yang, Z. (1994a) Estimating the pattern of nucleotide substitution. Journal of
Molecular Evolution 39 : 105–111.
Yang, Z. (1994b) Maximum likelihood phylogenetic estimation from DNA
sequences with variable rates over sites : approximate methods. Journal of
Molecular Evolution 39 : 306–314.
Yang, Z. (1996a) Phylogenetic analysis using parsimony and likelihood
methods. Journal of Molecular Evolution 42 : 294–307.
Yang, Z. (1996b) Among-site rate variation and its impact on phylogenetic
analyses. Trends in Ecology and Evolution 11 : 367–372.
Yang, Z. (1997) How often do wrong models produce better phylogenies?
Molecular Biology and Evolution 14 : 105–108.
Yang, Z. (1998) On the best evolutionary rate for phylogenetic analysis.
Systematic Biology 47 : 125–133.
Zahlbruckner, A. (1905) Ascolichenes. Lieferung 3. In Die natuX rlichen Pflanzen-
familien nebst ihren Gattungen und wichtigeren Arten insbesondere den
Nutzpflanzen (A. Engler and K. Prantl, eds), 1 (1*): 97–144. Engelmann,
Leipzig.
Zahlbruckner, A. (1921–40) Catalogus Lichenum Universalis. 10 vols. Gebru$ derBorntraeger, Leipzig.
Corresponding Editor : D. L. Hawksworth


















