
Molecular Mechanism of Anti-Prostate Cancer Activity of ...mail.encognitive.com/files/Molecular...
12
NUTRITION AND CANCER, 57(1), 100–110 Copyright C 2007, Lawrence Erlbaum Associates, Inc. Molecular Mechanism of Anti-Prostate Cancer Activity of Scutellaria Baicalensis Extract Fei Ye, Shiquan Jiang, Helen Volshonok, Josephine Wu, and David Y. Zhang Abstract: Scutellaria baicalensis is a widely used Chinese herbal medicine historically used in antiinflammatory and anticancer therapy. The goals of the study were to 1) deter- mine its in vitro and in vivo anti-prostate cancer activity, 2) investigate its molecular mechanism directed at cell proliferation control including cyclooxygenase-2(COX-2) prostaglandin E 2 (PGE 2 ) and cyclins/cdks pathways, and 3) compare it with those of PC-SPES (PC stands for prostate cancer and spes is Latin for hope), a former herbal mix- ture for prostate cancer treatment of which S. baicalensis is a major constituent. Two human prostate cancer cell lines (LNCaP, androgen dependent, and PC-3, androgen inde- pendent) were assessed for growth inhibition. S. baicalensis exerted dose- and time-dependent increased growth inhibi- tion in both cell lines. However, the PC-3 cells IC 50 (50% growth inhibition concentration) were slightly more sensi- tive than LNCaP cells (IC 50 = 0.15 mg/ml), although the former is androgen independent. S. baicalensis was more ef- fective in inhibition of cell growth compared with PC-SPES (IC 50 = 0.38 mg/ml for PC-3 cells). Significant reduction of PGE 2 synthesis in both cells after treatment with S. baicalen- sis resulted from direct inhibition of COX-2 activity rather than COX-2 protein suppression. S. baicalensis also inhib- ited prostate-specific antigen production in LNCaP cells. Finally, S. baicalensis suppressed expression of cyclin D1 in LNCaP cells, resulting in a G 1 phase arrest, while in- hibiting cdk1 expression and kinase activity in PC-3 cells, ultimately leading to a G 2 /M cell cycle arrest. Animal stud- ies showed a 50% reduction in tumor volume after a 7-wk treatment period. This study demonstrated that S. baicalensis may be a novel anticancer agent for the treatment of prostate cancer. Introduction Prostate cancer is the most frequently diagnosed malig- nancy in males and the second leading cause of cancer-related death in men after lung cancer. In 2006, the American Can- cer Society reported an expected total of 234,460 newly di- agnosed prostate cancers and 27,350 prostate cancer–related All authors are affiliated with the Department of Pathology, Mount Sinai School of Medicine, New York University, New York, NY 10029. deaths in the United States (1). Despite significant advances in prostate cancer treatment, the survival rate for advanced stages of this particular cancer has not significantly improved during the past decade (2). The high mortality rate observed in advanced prostate cancer patients is due to loss of an- drogen dependency for cancer cell growth that results in resistance to androgen ablation therapy (3). Furthermore, standard treatment options for localized prostate cancer (sur- gical, radiation, and hormonal therapy) are associated with morbidities that often impair patient quality of life, such as urinary incontinence and sexual dysfunction (4). Therefore, prostate cancer patients seek complementary and alternative medicine (CAM) modalities in hopes of halting or slowing disease progression to improve the general quality of life (5). Recent surveys demonstrated that approximately 40% of prostate cancer patients utilize various CAM modalities as a component of therapy (6). PC-SPES, a proprietary herbal mixture, recently received significant attention in the general public as well as scien- tific and medical communities. PC-SPES contains Scutel- laria baicalensis and seven other herbal extracts (7). PC- SPES has been shown to reduce serum prostate-specific anti- gen (PSA) levels and proven effective in patients with both androgen-sensitive and androgen-resistant types of prostate cancer (7–9). Importantly, all studies reported a >50% re- duction of PSA levels in more than half of patients with ad- vanced androgen-refractory prostate cancer. Unfortunately, PC-SPES was voluntarily removed from the consumer mar- ket by its manufacturers due to undeclared substance contam- ination (that is, warfarin and diethylstilbestrol; http://www. meb.unibonn.de/cancer.gov/CDR0000404384.html). In June 2002, four National Institutes of Health–sponsored PC- SPES–associated studies, one of which included a clinical trial, were placed on hold. Since then, three laboratory stud- ies have resumed their investigations (http://nccam.nih.gov/ health/alerts/spes/). S. baicalensis Georgi, also known as huang qin, or Chi- nese skullcap, has been used for thousands of years in tra- ditional Chinese medicine as an antiinflammatory, antibac- terial, and anticancer remedy to treat respiratory infections, gastrointestinal disorders, hepatitis, and cancer. Our recent
Transcript of Molecular Mechanism of Anti-Prostate Cancer Activity of ...mail.encognitive.com/files/Molecular...

NUTRITION AND CANCER, 57(1), 100–110Copyright C© 2007, Lawrence Erlbaum Associates, Inc.
Molecular Mechanism of Anti-Prostate Cancer Activity of ScutellariaBaicalensis Extract
Fei Ye, Shiquan Jiang, Helen Volshonok, Josephine Wu, and David Y. Zhang
Abstract: Scutellaria baicalensis is a widely used Chineseherbal medicine historically used in antiinflammatory andanticancer therapy. The goals of the study were to 1) deter-mine its in vitro and in vivo anti-prostate cancer activity,2) investigate its molecular mechanism directed at cellproliferation control including cyclooxygenase-2(COX-2)prostaglandin E2 (PGE2) and cyclins/cdks pathways, and3) compare it with those of PC-SPES (PC stands for prostatecancer and spes is Latin for hope), a former herbal mix-ture for prostate cancer treatment of which S. baicalensis isa major constituent. Two human prostate cancer cell lines(LNCaP, androgen dependent, and PC-3, androgen inde-pendent) were assessed for growth inhibition. S. baicalensisexerted dose- and time-dependent increased growth inhibi-tion in both cell lines. However, the PC-3 cells IC50 (50%growth inhibition concentration) were slightly more sensi-tive than LNCaP cells (IC50 = 0.15 mg/ml), although theformer is androgen independent. S. baicalensis was more ef-fective in inhibition of cell growth compared with PC-SPES(IC50 = 0.38 mg/ml for PC-3 cells). Significant reduction ofPGE2 synthesis in both cells after treatment with S. baicalen-sis resulted from direct inhibition of COX-2 activity ratherthan COX-2 protein suppression. S. baicalensis also inhib-ited prostate-specific antigen production in LNCaP cells.Finally, S. baicalensis suppressed expression of cyclin D1in LNCaP cells, resulting in a G1 phase arrest, while in-hibiting cdk1 expression and kinase activity in PC-3 cells,ultimately leading to a G2/M cell cycle arrest. Animal stud-ies showed a 50% reduction in tumor volume after a 7-wktreatment period. This study demonstrated that S. baicalensismay be a novel anticancer agent for the treatment of prostatecancer.
Introduction
Prostate cancer is the most frequently diagnosed malig-nancy in males and the second leading cause of cancer-relateddeath in men after lung cancer. In 2006, the American Can-cer Society reported an expected total of 234,460 newly di-agnosed prostate cancers and 27,350 prostate cancer–related
All authors are affiliated with the Department of Pathology, Mount Sinai School of Medicine, New York University, New York, NY 10029.
deaths in the United States (1). Despite significant advancesin prostate cancer treatment, the survival rate for advancedstages of this particular cancer has not significantly improvedduring the past decade (2). The high mortality rate observedin advanced prostate cancer patients is due to loss of an-drogen dependency for cancer cell growth that results inresistance to androgen ablation therapy (3). Furthermore,standard treatment options for localized prostate cancer (sur-gical, radiation, and hormonal therapy) are associated withmorbidities that often impair patient quality of life, such asurinary incontinence and sexual dysfunction (4). Therefore,prostate cancer patients seek complementary and alternativemedicine (CAM) modalities in hopes of halting or slowingdisease progression to improve the general quality of life(5). Recent surveys demonstrated that approximately 40% ofprostate cancer patients utilize various CAM modalities as acomponent of therapy (6).
PC-SPES, a proprietary herbal mixture, recently receivedsignificant attention in the general public as well as scien-tific and medical communities. PC-SPES contains Scutel-laria baicalensis and seven other herbal extracts (7). PC-SPES has been shown to reduce serum prostate-specific anti-gen (PSA) levels and proven effective in patients with bothandrogen-sensitive and androgen-resistant types of prostatecancer (7–9). Importantly, all studies reported a >50% re-duction of PSA levels in more than half of patients with ad-vanced androgen-refractory prostate cancer. Unfortunately,PC-SPES was voluntarily removed from the consumer mar-ket by its manufacturers due to undeclared substance contam-ination (that is, warfarin and diethylstilbestrol; http://www.meb.unibonn.de/cancer.gov/CDR0000404384.html). In June2002, four National Institutes of Health–sponsored PC-SPES–associated studies, one of which included a clinicaltrial, were placed on hold. Since then, three laboratory stud-ies have resumed their investigations (http://nccam.nih.gov/health/alerts/spes/).
S. baicalensis Georgi, also known as huang qin, or Chi-nese skullcap, has been used for thousands of years in tra-ditional Chinese medicine as an antiinflammatory, antibac-terial, and anticancer remedy to treat respiratory infections,gastrointestinal disorders, hepatitis, and cancer. Our recent

study demonstrated that the raw extract of S. baicalensisinhibits cancer cell growth in different cell lines, includingprostate, squamous cell, and breast cancers (10,11). We fur-ther demonstrated that the S. baicalensis extract decreasedPGE2 production via the suppression of cyclooxygenase-2(COX-2) activity, which is often elevated in various types ofcancers (10,12,13).
Ample evidence supports involvement of the COX-2 path-way in prostate carcinogenesis via several mechanisms, in-cluding stimulation of tumor invasion factors, such as vascu-lar endothelial growth factor (VEGF) expression (14), releaseof matrix metalloproteinases (15), facilitation of inflamma-tion (16), and possible contribution of the COX-2 productPGE2 to enhanced cell proliferation (16,17). Uncontrolledcell proliferation that leads to cancer formation is caused bydisruption of proper cell cycle regulation, a pivotal character-istic of cancer cells. Increased levels of the G0/G1 checkpointcontrolling cyclins D1 and E, and their associated proteinkinases (cdks), concurrently with decreased levels of cdk in-hibitors (Cip/Kip family), were reported in prostate cancerLNCaP cells (18) and in animal models (19).
The purpose of this study is to evaluate S. baicalensisas an anti-prostate cancer agent both in vitro (in androgen-dependent and androgen-independent cell lines) and in vivo(in nude mice xenografted with prostate cancer cells) and toexplore its ability to affect both COX-2 and cell cycle regu-lation pathways to further understand the molecular mecha-nism of its anti-prostate cancer activity. We hope the knowl-edge gained by our research will significantly contribute tothe development of new and effective therapy for this deadlydisease.
Materials and Methods
Chemicals and Drugs
The raw extract of S. baicalensis was prepared by boilingthe dried root of the plant in water, followed by a spray-drying process of the resulting water extract. The powderform of the extract was obtained from E-Fang Pharmaceu-tical Company (Guangdong, China) and contains baicalin,wogonoside, and oroxylin A as major components. The ex-tract was dissolved in culture medium to 20 mg/ml, vor-texed at room temperature for 1 min, and incubated at 37◦Cfor 1 h while rotating before use. This solution was cen-trifuged at 5,000 rpm for 10 min to remove any insolubleingredients. The supernatant was passed through a 0.22-µmfilter for sterilization and diluted with culture medium tofinal concentrations of 0.04–0.8 mg/ml of S. baicalensis ex-tract. The quality and authenticity of S. baicalensis were as-sessed using high-performance liquid chromatography (20).For result consistency, the same batch was used through-out the entire study. A 10-mM stock solution of celecoxib{SC-58635; 4-[5-(4-methylphenyl)-3-(trifluoromethyl)-1H-pyrazol-1]-benzene-sulfonamide, Searle Research and De-velopment, St. Louis, MO}, a selective COX-2 inhibitor, was
prepared with dimethyl sulfoxide (DMSO, Sigma, St. Louis,MO). PC-SPES (Lot# 5431060) in capsule form (320 mg percapsule) was purchased from Botanic Lab (Brea, CA). Be-cause most published studies on PC-SPES were performedusing ethanol as a solvent vehicle to dissolve the extract(21,22), we used both media as well as ethanol to dissolve S.baicalensis and PC-SPES to final concentrations of 0.0015–1.5 mg/ml, following the same preparation steps as describedpreviously.
Cell Lines and Cell Culture
Two human prostate cancer cell lines, LNCaP (andro-gen dependent) and PC-3 (androgen independent), were pur-chased from American Type Culture Collection (Rockville,MD). These two cell lines represent the two majortypes of prostate cancer: androgen-dependent/sensitive andandrogen-independent/insensitive prostate cancer cell popu-lations, the latter more typical of advanced prostate cancer(23). Both cell lines were maintained in RPMI 1640 mediumcontaining 10% GIBCOTM fetal bovine serum (Invitrogen,Carlsbad, CA) at 37◦C in a humidified atmosphere of 5%CO2.
Cell Viability Assay
The percentage of growth inhibition was determined byemploying a 3,4,5-dimethylthiazol-2,5-diphenyltetrazoliumbromide (MTT; Sigma) assay to measure viable cells. Cells(4 × 103cells per well) were seeded onto a 96-well plate for24 h, treated with various concentrations of S. baicalensis,and incubated for an additional 3 or 5 days at 37◦C. Subse-quently, 10 µl of MTT at a concentration of 5 mg/ml wasadded to each well, and cells were incubated for an additional4–6 h. The supernatant was aspirated, and 100 µl of DMSOwas added to the wells to dissolve any precipitate present.The optical density was then measured at a wavelength of570 nm using an ELX800 plate reader (Bio-Tek Instruments,Winooski, VT).
Cell Cycle Analysis
Cells (1.5 × 105 cells per well) were plated onto six-wellplates and incubated for 24 h at 37◦C. Various concentrationsof S. baicalensis were added to the wells and incubated foran additional 3 days. Cells were then washed, pelleted, fixedwith cold 70% ethyl alcohol for at least 30 min at –20◦C, andincubated with 100 µg/ml RNase A and 50 µg/ml propidiumiodide in phosphate-buffered saline at room temperature for30 min. Samples were immediately analyzed using a FAC-Scan flow cytometer (Becton Dickinson, San Jose, CA). Thepercentage of cell cycle distribution was determined usingCell Quest Pro software (Becton Dickinson, Franklin Lakes,NJ).
Vol. 57, No. 1 101

Quantitation of Prostate-Specific Antigen
LNCaP cells, known to produce high levels of PSA(24), were plated onto a six-well plate at a density of 1.5× 105 cells per well. After a 24-h incubation period, themedium was replaced with fresh medium containing 50%growth inhibition concentration (IC50) doses of S. baicalensis(0.15 mg/ml) or celecoxib (25 µM).After 72 h of treatment,the medium was collected and centrifuged to eliminate de-tached cells. PSA level in 150 µl supernatant was deter-mined using an AxSYM PSA Kit (Abbot Laboratories, Ab-bot Park, IL) according to manufacturer’s instructions. PSAisexpressed as nanograms per 106 cells.
PGE2 Enzyme Immunoassay
To quantify the level of PGE2 released into culture me-dia, a competitive PGE2 enzyme immunoassay was per-formed according to manufacturer’s instructions (AmershamPharmacia Biotech, Piscataway, NJ). Supernatants of con-trol and S. baicalensis (0.075–0.3 mg/ml)-treated cells werecollected, centrifuged at 2,500 rpm for 2 min, and stored at–70◦C until further analysis. Aliquots (50 µl) of each samplewere assayed in triplicate. Optical density was measured at450 nm on an ELX800 plate reader (Bio-Tek Instruments).The minimal detectable concentration of PGE2 by the assayis 50 pg/ml. PGE2is expressed as picograms per 106 cells.The effect of celecoxib and S. baicalensis on COX-2 activ-ity was further examined using an intracellular arachidonicacid conversion assay (11,25). After a 24-h incubation ofLNCaP or PC-3 cells, the medium was aspirated and replacedwith fresh medium containing 0.15 mg/ml S. baicalensis or25 µM celecoxib. Cells were incubated for additional peri-ods of time, and the medium was aspirated. After washingthe cells with medium, fresh medium containing 100 µMarachidonic acid (Cayman Chemical, Ann Arbor, MI) wasadded and incubated for an additional 30 min. PGE2in themedium was measured as described previously and expressedas picograms per milliliter of medium.
Western Blot Analysis
To assess dose-dependent protein expression change,LNCaP or PC-3 cells were treated for 72 h with variousconcentrations of S. baicalensis extract (0.05–0.8 mg/ml forCOX-2 protein or 0.1–0.8 mg/ml for cyclin D1, cyclin B1,and cdk1 kinase). To assess time-dependent protein expres-sion change, both cell lines were treated with a single doseof the S. baicalensis extract (0.4 mg/ml) for various times(12, 24, 48, and 72 h). Cellular proteins were extracted us-ing a buffer containing 50 mM Tris-HCl (pH 8.0), 150 mMNaCl, 0.1% sodium dodecyl sulfate (SDS), 1% NP-40, and1X proteinase inhibitor cocktail (Roche Applied Science,Indianapolis, IN) (11). Fifteen micrograms of protein wasfractionated by electrophoresis through a 10% SDS poly-
acrylamide gel, and the proteins were transferred onto anitrocellulose membrane. The membrane was blocked for1 h in a blocking buffer containing 5% dry milk, 20 mMTris-HCl (pH 7.5), 100 mM NaCl, and 0.1% Tween-20 andthen incubated with a polyclonal rabbit immunoglobulin G(IgG) anti-COX-2 antibody (1:400 dilution, Cayman Chem-ical) in the blocking buffer at 4◦C overnight. The membranewas further incubated with a secondary anti-rabbit antibodyconjugated with horseradish peroxidase (Amersham, Arling-ton Heights, IL), and the specific protein was detected usinga chemiluminescent method (Amersham) followed by au-toradiography according to manufacturer’s instructions. Thisprocedure was repeated for the detection of cyclin D1 andcyclin B1 proteins, using polyclonal rabbit IgG antibodiesat a 1:200 dilution (Santa Cruz Biotechnology, Santa Cruz,CA). For detection of cdk1 kinase (p34Cdc2), a monoclonalmouse IgG2aanti-cdk1 antibody (Santa Cruz Biotechnology)at a 1:200 dilution was used, followed by incubation withperoxidase-labeled anti-mouse antibody (Amersham) at a1:10,000 dilution. The same membrane was then used todetect β-actin protein, using a monoclonal mouse IgG2aantiβ-actin antibody (Sigma) at a 1:10,000 dilution as describedpreviously.
cdk1 Kinase Activity Assay
PC-3 cells were treated for 72 h with various concentra-tions of S. baicalensis (0.1–0.8 mg/ml) in a dose-dependencystudy or with a single concentration of S. baicalensis(0.4 mg/ml) in a time-course study. Cellular proteins wereextracted as described previously. One milligram of proteinwas further incubated with 2 µg of anti-cdk1 antibody (SantaCruz Biotechnology) for 2 h at 4◦C, followed by incubationwith 25 µl (25% agarose) of protein A-agarose beads (SantaCruz Biotechnology) at 4◦C on a rotator overnight. Aftertwo sequential wash cycles with a lysis buffer and kinasebuffer (50 mM TrisHCl, 10 mM MgCl2, and 1 mM dithiothre-itol), the immunoprecipitate was resuspended in 30 µl kinasebuffer containing 4 µg Histone H1 (cdk1 kinase substrate),50 µM of unlabeled adenosine triphosphate (ATP), and 5 µCiof [γ -32P]ATP at 30◦C for 30 min. After the addition of 2XSDS loading buffer and heating at 100◦C for 5 min, the reac-tion products were separated on 12% SDS–polyacrylamidegel electrophoresis and visualized by autoradiography.
Animals and Treatment Protocol
Six-week-old male nude mice (NCR-nu/nu) were pur-chased from Taconic (Cincinnati, OH). Each animal receiveda single-bolus subcutaneous injection of 3 × 106 PC-3 cells tothe right flank. After 1 wk of inoculation to allow for adequatetumor cell engraftment (average tumor mass ≈150 mm3), themice were randomized into two experimental groups (n =5). The treatment group was given a 200-mg/kg dose ofS. baicalensis extract dissolved in water by oral gavage once
102 Nutrition and Cancer 2007
a day, five times per week for 7 wk. Mice were weighed, andtumor volume was assessed on a weekly basis. Tumor vol-ume was measured by two perpendicular dimensions (longand short) using a caliper and calculated using the formula(a × b2)/2, where a is the larger and b is the smaller di-mension of the tumor. Treatment and care of experimentalanimals were in accordance with the institutional guidelinesof the Mount Sinai School of Medicine.
Results
Growth Inhibition Effect of Scutellaria Baicalensis onProstate Cancer Cells
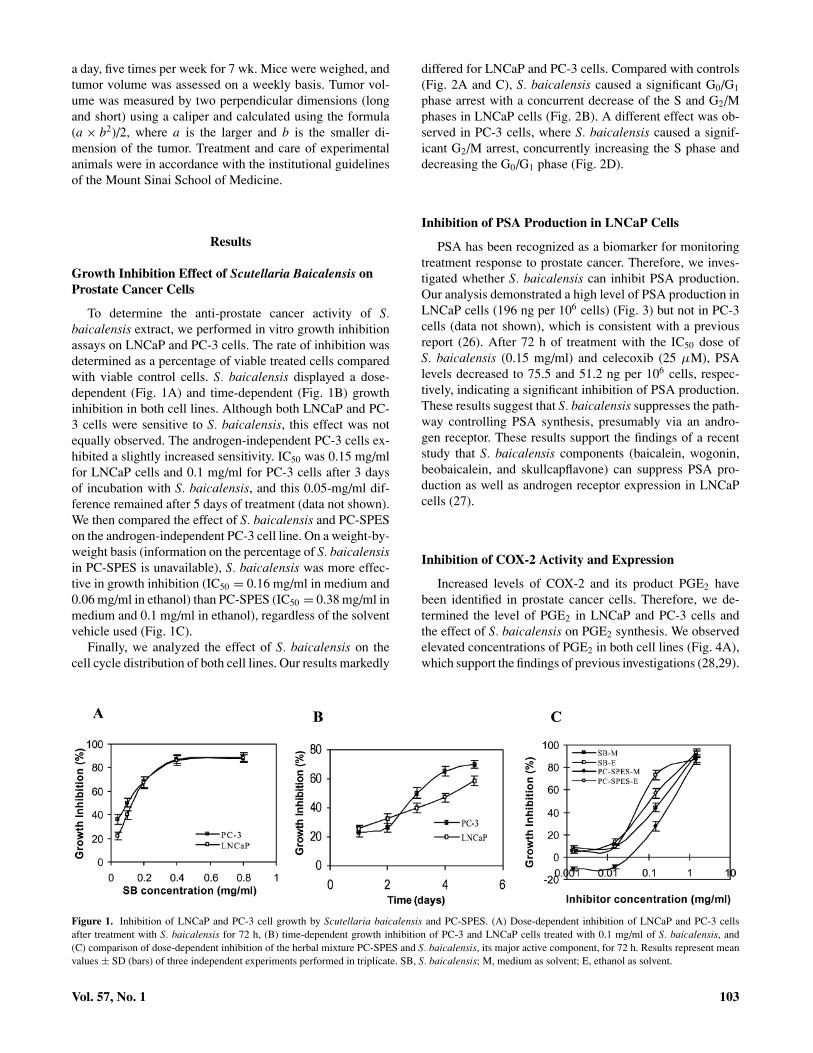
To determine the anti-prostate cancer activity of S.baicalensis extract, we performed in vitro growth inhibitionassays on LNCaP and PC-3 cells. The rate of inhibition wasdetermined as a percentage of viable treated cells comparedwith viable control cells. S. baicalensis displayed a dose-dependent (Fig. 1A) and time-dependent (Fig. 1B) growthinhibition in both cell lines. Although both LNCaP and PC-3 cells were sensitive to S. baicalensis, this effect was notequally observed. The androgen-independent PC-3 cells ex-hibited a slightly increased sensitivity. IC50 was 0.15 mg/mlfor LNCaP cells and 0.1 mg/ml for PC-3 cells after 3 daysof incubation with S. baicalensis, and this 0.05-mg/ml dif-ference remained after 5 days of treatment (data not shown).We then compared the effect of S. baicalensis and PC-SPESon the androgen-independent PC-3 cell line. On a weight-by-weight basis (information on the percentage of S. baicalensisin PC-SPES is unavailable), S. baicalensis was more effec-tive in growth inhibition (IC50 = 0.16 mg/ml in medium and0.06 mg/ml in ethanol) than PC-SPES (IC50 = 0.38 mg/ml inmedium and 0.1 mg/ml in ethanol), regardless of the solventvehicle used (Fig. 1C).
Finally, we analyzed the effect of S. baicalensis on thecell cycle distribution of both cell lines. Our results markedly
differed for LNCaP and PC-3 cells. Compared with controls(Fig. 2A and C), S. baicalensis caused a significant G0/G1
phase arrest with a concurrent decrease of the S and G2/Mphases in LNCaP cells (Fig. 2B). A different effect was ob-served in PC-3 cells, where S. baicalensis caused a signif-icant G2/M arrest, concurrently increasing the S phase anddecreasing the G0/G1 phase (Fig. 2D).
Inhibition of PSA Production in LNCaP Cells
PSA has been recognized as a biomarker for monitoringtreatment response to prostate cancer. Therefore, we inves-tigated whether S. baicalensis can inhibit PSA production.Our analysis demonstrated a high level of PSA production inLNCaP cells (196 ng per 106 cells) (Fig. 3) but not in PC-3cells (data not shown), which is consistent with a previousreport (26). After 72 h of treatment with the IC50 dose ofS. baicalensis (0.15 mg/ml) and celecoxib (25 µM), PSAlevels decreased to 75.5 and 51.2 ng per 106 cells, respec-tively, indicating a significant inhibition of PSA production.These results suggest that S. baicalensis suppresses the path-way controlling PSA synthesis, presumably via an andro-gen receptor. These results support the findings of a recentstudy that S. baicalensis components (baicalein, wogonin,beobaicalein, and skullcapflavone) can suppress PSA pro-duction as well as androgen receptor expression in LNCaPcells (27).
Inhibition of COX-2 Activity and Expression
Increased levels of COX-2 and its product PGE2 havebeen identified in prostate cancer cells. Therefore, we de-termined the level of PGE2 in LNCaP and PC-3 cells andthe effect of S. baicalensis on PGE2 synthesis. We observedelevated concentrations of PGE2 in both cell lines (Fig. 4A),which support the findings of previous investigations (28,29).
Figure 1. Inhibition of LNCaP and PC-3 cell growth by Scutellaria baicalensis and PC-SPES. (A) Dose-dependent inhibition of LNCaP and PC-3 cellsafter treatment with S. baicalensis for 72 h, (B) time-dependent growth inhibition of PC-3 and LNCaP cells treated with 0.1 mg/ml of S. baicalensis, and(C) comparison of dose-dependent inhibition of the herbal mixture PC-SPES and S. baicalensis, its major active component, for 72 h. Results represent meanvalues ± SD (bars) of three independent experiments performed in triplicate. SB, S. baicalensis; M, medium as solvent; E, ethanol as solvent.
Vol. 57, No. 1 103

Figure 2. Cell cycle distribution after treatment with Scutellaria baicalensis. LNCaP (A and B) and PC-3 (C and D) cells treated with S. baicalensis at0.15 mg/ml and 0.2 mg/ml, respectively, for 72 h and subjected to flow cytometry analysis. Results represent mean values of three independent experiments.
However, LNCaP cells produced a notably higher level ofPGE2 (39.1 pg per 106 cells) compared with PC-3 cells (26.9pg per 106 cells) after 12 h of incubation. S. baicalensis sig-nificantly inhibited PGE2 production in LNCaP cells starting
Figure 3. Suppression of prostate-specific antigen (PSA) production inLNCaP cells. PSA levels were measured in the supernatant of androgen-dependent LNCaP cells after treatment with the IC50 doses of Scutellariabaicalensis (0.15 mg/ml) and celecoxib (25 µM). Results represent meanvalues ± SD (bars) of three independent experiments. Data were analyzedusing the Student’s t-test.∗∗P < 0.01 compared with the control group.
at a concentration of 0.075 mg/ml and in PC-3 cells startingat a concentration of 0.15 mg/ml. PGE2production continuedto decline in both cell lines as S. baicalensis concentrationsincreased (up to 0.3 mg/ml).
To further investigate the mechanism by which S.baicalensis inhibits PGE2 production, we examined its effecton COX-2 enzymatic activity using an intracellular arachi-donic acid conversion assay that measures PGE2 produc-tion after the addition of exogenous arachidonic acid to thenewly replenished medium of treated cells. For comparison,a known COX-2 inhibitor, celecoxib, was included in thisassay. By providing exogenous arachidonic acid, a COX-2substrate, we sought to eliminate the effect of endogenousarachidonic acid release from the cell membrane due to in-tracellular protein kinase and/or phospholipase A2 activities.Our results demonstrate a significant decrease of PGE2 levelin the presence of both S. baicalensis and celecoxib at IC50
doses in both PC-3 and LNCaP cells during a 24-h timeperiod (Fig. 4B and C). For each cell line, the pattern ofPGE2 inhibition was similar for celecoxib and S. baicalen-sis, although a slight delay of inhibition by S. baicalensisin PC-3 cells (2-h lag period for S. baicalensis vs. imme-diate for celecoxib) was observed (Fig. 4C). The level ofdecreased PGE2 production was similar in both cell lines,that is, a drop of approximately 200 pg/ml (from 1,048 to853 pg/ml for LNCaP cells and from 482 to 264 pg/ml
104 Nutrition and Cancer 2007

Figure 4. Inhibition of PGE2 synthesis and cyclooxygenase-2 activity by Scutellaria baicalensis and celecoxib. (A) Dose-dependent suppression of PGE2
synthesis after 12-h incubation with S. baicalensis. Suppression of arachidonic acid conversion to PGE2 by S. baicalensis (0.15 mg/ml) and celecoxib (25 µM)in (B) LNCaP and (C) PC-3 cells. Results represent mean values ± SD (bars) of three independent experiments performed in triplicate. Data were analyzedusing the Student’s t-test. *P < 0.05 and **P < 0.01 compared with the control groups.
for PC-3 cells). However, the inhibitory effect in LNCaPdid not occur until 12 h after the addition of arachidonicacid (Fig. 4B). These results strongly support our hypothesisthat S. baicalensis inhibits PGE2 synthesis through COX-2activity.
We also investigated the effect of S. baicalensis on COX-2expression using Western blotting (Fig. 5). The effect of S.baicalensis treatment on COX-2 expression differed in twocell lines, for both time-course and dose-dependency studies.In PC-3 cells, which had a lower baseline COX-2 expression
Figure 5. Effect of Scutellaria baicalensis cyclooxygenase-2 (COX-2) protein expression. PC-3 and LNCaP cells were treated with either S. baicalensis(0.4 mg/ml) for various periods of time (A and C) or with various concentrations of S. baicalensis for 72 h (B and D). The level of protein expression wasdetermined by Western blot. β-Actin was used as an internal control. Three independent experiments were performed, and similar results were obtained.
Vol. 57, No. 1 105

level than LNCaP cells, treatment with S. baicalensis inducedCOX-2 expression in a time- and dose-dependent manner(Fig. 5A and B). In contrast, treatment of LNCaP cells withS. baicalensis slightly decreased COX-2 expression (Fig. 5Cand D). These results further suggested that decreased PGE2
synthesis induced by S. baicalensis is likely due to the directinhibition of enzymatic activity of COX-2 rather than itsexpression.
Effect on Cell Cycle–Regulating Molecules
Cell cycle analysis revealed that S. baicalensis treatmentinduced G1 arrest for LNCaP cells and G2 for PC-3. There-fore, we further evaluated the effect of S. baicalensis oncyclin D1 and cyclin B1 expression, two factors importantfor cell progression from the G1 to S and the G2 to M phases,respectively (Fig. 6). When LNCaP cells were treated withvarious concentrations ofS. baicalensis, cyclin D1 expres-sion decreased in a dose-dependent manner (Fig. 6A). Wealso observed a steady decrease in cyclin D1 expression inthe course of 72 h, which at the end dropped to undetectablelevels (data not shown). No change in cyclin D1 expression
was observed in PC-3 cells, as expected (Fig. 6C). Theseresults indicate that S. baicalensis suppresses the expressionof cyclin D1 in LNCaP cells, resulting in G1-phase arrest.
When cyclin B1 was analyzed, its level of expression didnot change in both cell lines (Fig. 6B and D), although amarked G2/M-phase arrest in PC-3 cells was observed aftertreatment with S. baicalensis. Therefore, we further exam-ined the expression of cdk1 protein kinase (p34Cdc2), thecatalytic subunit of the cyclin B1/cdk1 complex. Our resultsshowed that treatment with different doses of S. baicalen-sis for 72 h resulted in a slight decrease of cdk1 expressionin PC-3 cells (Fig. 7A). Time-course studies confirmed theefficacy of this dose after 72 h of treatment (Fig. 7B). Asexpected, no change in cdk1 expression in LNCaP cells wasobserved after treatment with S. baicalensis (Fig. 7E). Wethen determined the effect of S. baicalensis on cdk1 kinaseactivity by analyzing the level of histone H1 phosphorylation,the substrate of the cyclin B1/cdk1 complex, using a kinaseactivity assay. We observed both a dose- and time-dependentdecrease of cdk1 kinase activity in PC-3 cells after treatmentwith S. baicalensis (Fig. 7C and D). These results indicatethat S. baicalensis inhibits primarily cdk1 kinase activity inPC-3 cells leading to the G2/M-phase arrest.
Figure 6. Effect of Scutellaria baicalensis on cyclin D1 and cyclin B1 protein expression. LNCaP and PC-3 cells were treated with various concentrations ofS. baicalensis for 72 h, and the expression levels of cyclin D1 (A and C) and cyclin B1 (B and D) were determined by Western blot. β-Actin was used as aninternal control. Three independent experiments were performed, each with similar results.
106 Nutrition and Cancer 2007
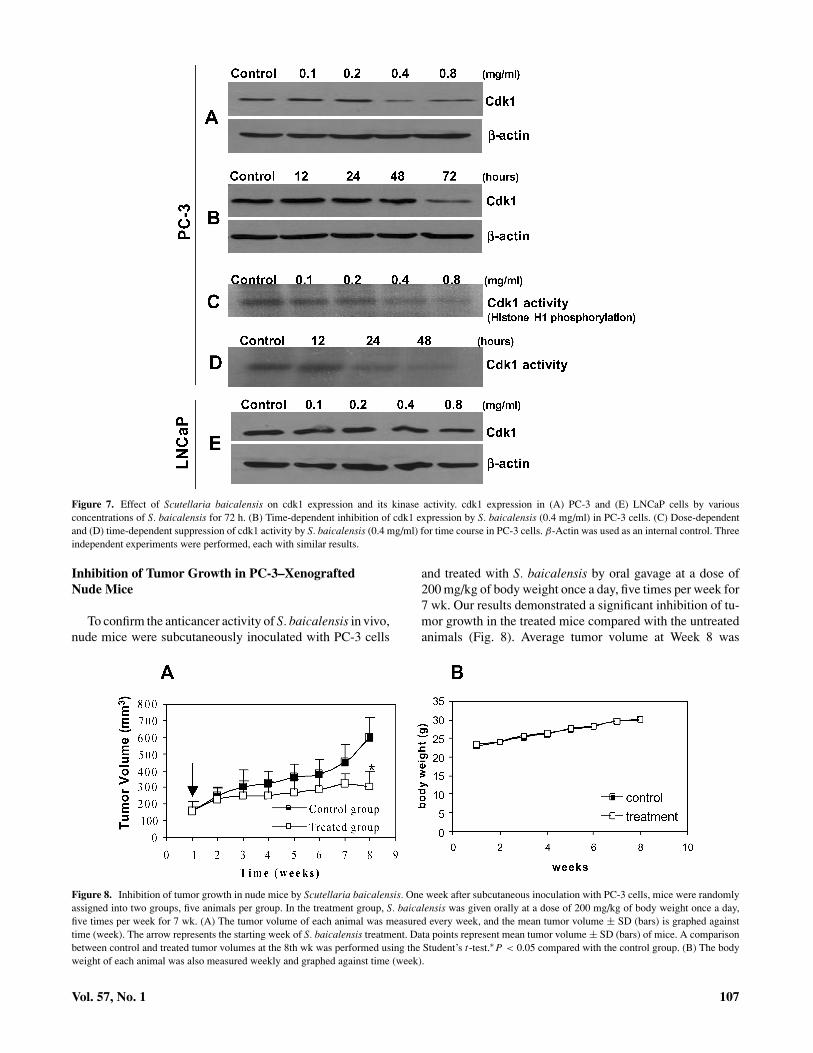
Figure 7. Effect of Scutellaria baicalensis on cdk1 expression and its kinase activity. cdk1 expression in (A) PC-3 and (E) LNCaP cells by variousconcentrations of S. baicalensis for 72 h. (B) Time-dependent inhibition of cdk1 expression by S. baicalensis (0.4 mg/ml) in PC-3 cells. (C) Dose-dependentand (D) time-dependent suppression of cdk1 activity by S. baicalensis (0.4 mg/ml) for time course in PC-3 cells. β-Actin was used as an internal control. Threeindependent experiments were performed, each with similar results.
Inhibition of Tumor Growth in PC-3–XenograftedNude Mice
To confirm the anticancer activity of S. baicalensis in vivo,nude mice were subcutaneously inoculated with PC-3 cells
and treated with S. baicalensis by oral gavage at a dose of200 mg/kg of body weight once a day, five times per week for7 wk. Our results demonstrated a significant inhibition of tu-mor growth in the treated mice compared with the untreatedanimals (Fig. 8). Average tumor volume at Week 8 was
Figure 8. Inhibition of tumor growth in nude mice by Scutellaria baicalensis. One week after subcutaneous inoculation with PC-3 cells, mice were randomlyassigned into two groups, five animals per group. In the treatment group, S. baicalensis was given orally at a dose of 200 mg/kg of body weight once a day,five times per week for 7 wk. (A) The tumor volume of each animal was measured every week, and the mean tumor volume ± SD (bars) is graphed againsttime (week). The arrow represents the starting week of S. baicalensis treatment. Data points represent mean tumor volume ± SD (bars) of mice. A comparisonbetween control and treated tumor volumes at the 8th wk was performed using the Student’s t-test.∗P < 0.05 compared with the control group. (B) The bodyweight of each animal was also measured weekly and graphed against time (week).
Vol. 57, No. 1 107

306 mm3 (95% confidence interval, CI = 166–448 mm3)for treated mice and 600 mm3 (95% CI = 406–794 mm3) forcontrol mice, a 50% reduction of tumor mass (P < 0.05). Ata dose of 200 mg/kg of S. baicalensis, there was no evidenceof drug-related toxicity, suggesting that S. baicalensis is welltolerated by the animals.
Discussion
S. baicalensis has been shown to have a broad spec-trum of biological activities, including antiinflammatory (30)and anticancer (10). The present study confirmed that S.baicalensis has a strong dose-dependent inhibitory activityagainst both androgen-dependent (LNCaP) and androgen-independent (PC-3) types of prostate cancer, with estab-lished IC50 doses at 0.15 mg/ml for LNCaP cells and0.1 mg/ml for PC-3 cells (Fig. 1A). The ability to inhibitprostate cancer growth was further confirmed in our ani-mal study (Fig. 8). The ability of S. baicalensis extract toinhibit both androgen-dependent and androgen-independentprostate cancer cells is clinically relevant and important. It isclear that prostate cancers frequently develop resistance to aconventional therapeutic regimen including androgen abla-tion. Once the prostate cancer cells become androgen inde-pendent, clinical failure often occurs, with a median survivalof 9–12 mo. Therefore, S. baicalensis may be effective forthe treatment of both androgen-independent and androgen-dependent prostate cancers. These promising results supportthe future clinical trial of this herbal extract to determine itsefficacy as an anti-prostate cancer agent.
It is worthy to note S. baicalensis is more effective thanPC-SPES (Fig. 1C), a popular herbal mixture for the treat-ment of prostate cancer that contains eight herbs includingS. baicalensis. In support of our finding, a recent studydemonstrated that S. baicalensis is one of the herbal com-ponents that inhibit prostate cancer cell growth (LNCaP) invitro, and S. baicalensis is the only herbal component withantiandrogenic activity (27). These results also demonstratedthat S. baicalensis may be a viable alternative for PC-SPES,given the quality-control and adulteration concerns. The useof a single herb reduces the side effects and possible antago-nistic interactions of multiple herbal mixtures and eliminatesthe quality-control issues, such as contamination, adulter-ation, and/or poor quality, that are associated with multipleherbal mixtures (20,31).
The anticancer activity of S. baicalensis may be derivedfrom its ability to inhibit cancer cell proliferation. We did notobserve increased apoptosis in both cell lines after treatmentwith S. baicalensis (data not shown). However, S. baicalensiscaused arrest at different cell cycle phases in these two celllines, that is, G1 arrest for androgen-dependent LNCaP cellsand G2 arrest for androgen-independent PC-3 cells (Fig. 2).At the molecular level, we demonstrated that S. baicalen-sis reduced expression of cyclin D1 in androgen-dependentLNCaP cells, which is responsible for the G1- to S-phase tran-
sition of the cell cycle (Fig. 6A). In androgen-independentPC-3 cells, a significant change in cyclin D1 and B1 ex-pression was not observed; however, a significant decreasein cdk1 kinase expression and activity after treatment withS. baicalensis was noted (Figs. 6 and 7). These findings areconsistent with a recent study showing that the major compo-nents of S. baicalensis (baicalein, wogonin, beobaicalein, andskullcapflavone) induce G1 arrest for LNCaP and G2 arrestfor PC-3 cells (27). However, it is unclear why S. baicalensisdifferentially affects these two prostate cancer cell lines. Thismay be due to different regulatory mechanisms of cell prolif-eration between these two cell lines. It has been shown thatLNCaP and PC-3 cells exhibit significant difference in theirgene expression patterns, particularly the androgen-relatedsignal transduction pathways. Karan et al., using a microar-ray method, identified 34 up-regulated and 8 down-regulatedgenes in androgen-dependent cells compared with androgen-independent cells (32). Therefore, although S. baicalensisacts on the same signal transduction pathway (that is, COX-2), the possibility exists that the downstream effect of COX-2is different in these two cell lines (that is, G1 vs. G2 arrest).
The ability of S. baicalensis to affect cell cycle–regulatingmolecules is an important finding because elevated levels ofcyclin D1 promote prostate cancer development by reducingits dependence on androgen stimulation (18). Additionally,there is evidence that androgens may induce G1/S transitionand thus contribute to cancer cell proliferation (19). Increasedexpression of G1-phase kinases cdk2 and cdk4 and increasedactivity of cdk2 kinase were observed as a result of androgenstimulation as well as down-regulation of cdk4 inhibitor p16(33) and cdk2 inhibitor p21 (34). Changes in the expression ofcell cycle–regulating genes, such as cyclins A and D, cdk2,and p21, were also observed in PC-SPES–treated prostatecancer cells (35). Due to the critical role of these cell cycle–regulating molecules in cancer development, these moleculeshave become important targets for drug development. Forexample, flavopiridol, a semisynthetic flavonoid derived fromthe bark of the Dysoxylum binectariferum plant indigenous toIndia, specifically targets cdks to block cell cycle progressionat the G1/S and G2/M checkpoints (36). S. baicalensis mayprove to be a good candidate for this class of agent.
Although it is possible that S. baicalensis acts directlyon cell cycle–regulating molecules, such as cyclin D1 andcdk1, it is also possible that S. baicalensis may act on otherupstream cell cycle–regulating molecules. It has been shownthat COX-2 and its product PGE2 are important molecules inregulating prostate cancer cell development and progression,particularly during the early stages (37). For example, in PC-3 cells, PGE2 induced the immediate-early gene c-fos via theprotein kinase A pathway by activating prostaglandin EP-2/4receptors, whereas COX-2 stimulates VEGF expression, re-sulting in increased proliferation (38,39). Furthermore, COX-2–specific inhibitors celecoxib and nimesulide dramaticallyreduced the expression of androgen-inducible genes, such asPSA, as well as androgen receptor expression via suppressionof their promoter activity (40). Our results showed that bothprostate cancer cell lines had increased COX-2 expression
108 Nutrition and Cancer 2007

(Fig. 5) and increased PGE2 production (Fig. 4) (more so inandrogen-sensitive LNCaP cells), consistent with previousreports (28,29). S. baicalensis suppressed PGE2 synthesis inboth cell lines (Fig. 4A), and the pattern of PGE2 inhibitionwas similar for both celecoxib, a COX-2–specific inhibitor,and S. baicalensis (Fig. 4C), indicating that S. baicalensisacts on COX-2 enzymatic activity directly. However, it is un-clear why a delay of inhibition of PGE2 synthesis in LNCaPcells was observed (Fig. 4B). Analysis of COX-2 expressionin both cell lines revealed either slightly decreased (LNCaP)or increased (PC-3) COX-2 expression (Fig. 5), further sup-porting the notion that decreased PGE2 synthesis resultedfrom direct inhibition of COX-2 activity. Although the cross-talk between COX-2/PGE2 and cell cycle regulation (thatis, cyclin D1 and cdk1) remains unclear, it is plausible thatCOX-2 and/or PGE2 acts on cell cycle regulation through in-termediate kinase pathways such as mitogen activated proteinkinase (MAPK)/extracellular signal-regulated kinase (ERK)(41). It is interesting to note that both S. baicalensis andcelecoxib can inhibit PSA production in LNCaP cells (Fig.3), suggesting that COX-2 also interacts with the androgensignal transduction pathway. In a phase II clinical trial, 92%of prostate cancer patients who received celecoxib treatmentfor 3 mo showed a significant reduction of PSA level, fur-ther confirming the regulatory relationship of COX-2 andPSA (42). It is worthy to note that, although S. baicalen-sis inhibits androgen receptor expression (27), the andro-gen signal pathway alone cannot explain its anti-prostatecancer activity because both androgen-dependent andandrogen-independent prostate cancer cells are sensitive toS. baicalensis.
In summary, this study, together with others (27), demon-strated that S. baicalensis, the major component of the mixedherbal formula PC-SPES, is responsible for its anti-prostatecancer activity. More importantly, it inhibits androgen-independent prostate cancer cells (PC-3) more strongly thanit inhibits androgen-dependent cells (LNCaP), which couldbe a viable alternative to prostate cancer patients who failto respond to conventional hormone-based treatment. Inhi-bition of PSA production in LNCaP cells not only provesthat S. baicalensis can inhibit the androgen-dependent sig-nal transduction pathway but also suggests that PSA can beused as an intermediate marker for monitoring the responseto S. baicalensis treatment for future clinical trials. Mech-anistically, S. baicalensis targets multiple cell proliferationpathways, including COX-2/PGE2, cyclin D1, and cdk1, re-sulting in the suppression of prostate cancer growth at the G1
or G2 phases. Future clinical study is necessary to determineits efficacy in prostate cancer patients.
Acknowledgments and Notes
This work was partially supported by an NIH/NCCAM grant (AT00739)to D.Y.Z.
Address correspondence to D. Y. Zhang, Director, Molecular PathologyLaboratory, Mount Sinai School of Medicine, Box 1122, One Gustave L.
Levy Place, New York, New York 10029. Phone: 212-659-8173. FAX: 212-427-2082. E-mail: [email protected].
Submitted 25 January 2006; accepted in final form 14 September 2006.
References
1. Jemal A, Siegel R, Ward E, Murray T, Xu J, et al.: Cancer statistics,2006. CA Cancer J Clin 56, 106–130, 2006.
2. Chu KC, Tarone RE, and Freeman HP: Trends in prostate cancer mor-tality among black men and white men in the United States. Cancer97, 1507–1516, 2003.
3. Cho D, Di Blasio C, Rhee A, and Kattan M: Prognostic factors forsurvival in patients with hormone-refractory prostate cancer (HRPC)after initial androgen deprivation therapy (ADT). Urol Oncol 21, 282–291, 2003.
4. Eton D and Lepore S: Prostate cancer and health-related quality of life:a review of the literature. Psychooncology 11, 307–326, 2002.
5. Eisenberg D, Davis R, Ettner S, Appel S, Wilkey S, et al.: Trends inalternative medicine use in the United States, 1990–1997: results of afollow-up national survey. JAMA 280, 1569–1575, 1998.
6. Zimmerman R: Prevalence of complementary medicine in urologicpractice: a review of recent studies with emphasis on use among prostatecancer patients. Urol Clin North Am 29, 1–9, 2002.
7. Pfeifer B, Pirani SR, Hamann S, and Klippel K: PC-SPES, a dietarysupplement for the treatment of hormone-refractory prostate cancer.BJU Int 85, 481–485, 2000.
8. de la Taille A, Buttyan R, Hayek O, Bagiella E, Shabsigh A, et al.:Herbal therapy PC-SPES: In vitro effects and evaluation of its efficacyin 69 patients with prostate cancer. J Urol 164, 1229–1234, 2000.
9. Small E, Frohlich M, Bok R, Shinohara K, Grossfeld G, et al.: Prospec-tive trial of the herbal supplement PC-SPES in patients with progressiveprostate cancer. J Clin Oncol 18, 3595–3603, 2000.
10. Ye F, Xui L, Yi J, Zhang W, and Zhang DY: Anticancer activity ofScutellaria baicalensis and its potential mechanism. J Altern Comple-ment Med 8, 567–572, 2002.
11. Zhang DY, Wu J, Ye F, Xue L, Jiang S, et al.: Inhibition of cancer cellproliferation and prostaglandin E2 synthesis by Scutellaria baicalensis.Cancer Res 63, 4037–4043, 2003.
12. Kase Y, Saitoh K, Ishige A, and Komatsu Y: Mechanisms by whichhange-shashin-to reduces prostaglandin E2 levels. Biol Pharm Bull 21,1277–1281, 1998.
13. Chi YS, Lim H, Park H, and Kim HY: Effects of wogonin, a plantflavone from Scutellaria radix, on skin inflammation: in vivo regulationof inflammation-associated gene expression. Biochem Pharmacol 66,1271–1278, 2003.
14. Fujita H, Koshida K, Keller E, Takahashi Y, Yoshimito T, et al.:Cyclooxygenase-2 promotes prostate cancer progression. Prostate 53,232–240, 2002.
15. Attiga F, Fernandez P, Weeraratna A, Manyak M, and Patierno S: In-hibitors of prostaglandin synthesis inhibit human prostate tumor cellinvasiveness and reduce the release of matrix metalloproteinases. Can-cer Res 60, 4629–4637, 2000.
16. Liu X, Kirschenbaum A, Lu M, Yao S, Klausner A, et al.: ProstaglandinE2 stimulates prostatic intraepithelial neoplasia cell growth throughactivation of the interleukin-6/gp130/stat-3 signaling pathway. BiochemBiophys Res Commun 290, 249–255, 2002.
17. Tjandrawinata R and Hughes-Fulford M: Up-regulation ofcyclooxyenase-2 by product-prostaglandin E2. Adv Exp Med Biol 407,163–170, 1997.
18. Chen Y, Martinez L, LaCava M, Coghlan L, and Conti C: Increasedcell growth and tumorigenicity in human prostate LNCaP cells byoverexpression to cyclin D1. Oncogene 16, 1913–1920, 1998.
19. Chen Y, Robles A, Martinez L, Liu F, Gimenez-Conti I, et al.: Expres-sion of G1 cyclins, cyclin-dependent kinases, and cyclin-dependent ki-nase inhibitors in androgen-induced prostate proliferation in castratedrats. Cell Growth Differ 7, 1571–1578, 1996.
Vol. 57, No. 1 109

20. Ye F, Wang H, Jiang S, Wu J, Shao J, et al.: Quality evaluation ofcommercial extracts of Scutellaria baicalensis. Nutr Cancer 49, 217–222, 2004.
21. Schwarz R, Donohue C, Sadava D, and Kane S: Pancreatic cancer invitro toxicity mediated by Chinese herbs SPES and PC-SPES: impli-cations for monotherapy and combination treatment. Cancer Lett 189,59–68, 2003.
22. Lu X, Guo J, and Hsieh T: PC-SPES inhibits cell prolifera-tion by modulating p21, cyclins D, E and B and multiple cellcycle-related genes in prostate cancer cells. Cell Cycle 2, 59–63,2003.
23. Navarro D, Luzardo O, Fernandez L, Chesa N, and Diaz-Chico B: Tran-sition to androgen-independence in prostate cancer. J Steroid BiochemMol Biol 81, 191–201, 2002.
24. Gau J, Salter R, Krill D, Grove M, and Becich M: The biosynthesis andsecretion of prostate-specific antigen in LNCaP cells. Cancer Res 57,3830–3834, 1997.
25. Chen Y, Shen S, Chen L, Lee T, and Yang L: Wogonin, baicalin,and baicalein inhibition of inducible nitric oxide synthase andcyclooxygenase-2 gene expressions induced by nitric oxide synthaseinhibitors and lipopolysaccharide. Biochem Pharmacol 61, 1417–1427,2001.
26. Pecher S, Pflug B, Brink A, and Nelson J: Endothelin A receptorblockade does not alter PSA secretion in prostate cancer cell lines.Prostate 60, 175–177, 2004.
27. Bonham M, Posakony J, Coleman I, Montgomery B, Simon J, et al.:Characterization of chemical constituents in Scutellaria baicalensiswith antiandrogenic and growth-inhibitory activities toward prostatecarcinoma. Clin Cancer Res 11, 3905–3914, 2005.
28. Hsu A, Ching T, Wang D, Song X, Rangnekar V, et al.: Thecyclooxygenase-2 inhibitor celecoxib induces apoptosis by blockingakt activation in human prostate cancer cells independently of bcl-2.J Biol Chem 275, 11397–11403, 2000.
29. Song X, Lin H, Johnson A, Tseng P, Yang Y, et al.: Cyclooxygenase-2,player or spectator in cyclooxygenase-2 inhibitor-induced apoptosis inprostate cancer cells. JNCI 94, 585–591, 2002.
30. Schinella GR, Tournier HA, Prieto JM, Mordujovich de Buschiazzo P,et al.: Antioxidant activity of anti-inflammatory plant extracts. Life Sci70, 1023–1033, 2002.
31. Chung V, Tattersall M, and Cheung H: Interactions of a herbal combi-nation that inhibits growth of prostate cancer cells. Cancer ChemotherPharmacol 53, 384–390, 2004.
32. Karan D, Kelly D, Rizzino A, Lin M, and Batra S: Expression pro-file of differentially-regulated genes during progression of androgen-independent growth in human prostate cancer cells. Carcinogenesis 23,967–975, 2002.
33. Lu S, Tsai S, and Tsai M: Regulation of androgen-dependent prostaticcancer cell growth: androgen regulation of CDK2, CDK4, and CKI p16genes. Cancer Res 57, 4511–4516, 1997.
34. Lu S, Liu M, Epner D, Tsai S, and Tsai M: Androgen regulation ofthe cyclin-dependent kinase inhibitor p21 gene through an androgenresponse element in the proximal promoter. Mol Endocrinol 13, 376–384, 1999.
35. Bonham M, Arnold H, Montgomery B, and Nelson P: Molecular effectsof the herbal compound PC-SPES: identification of activity pathwaysin prostate carcinoma. Cancer Res 62, 3920–3924, 2002.
36. Shapiro G: Preclinical and clinical development of the cyclin-dependentkinase inhibitor flavopiridol. Clin Cancer Res 10, 4270–4275, 2004.
37. Dawson N and Slovin S: Novel approaches to treat asymptomatic,hormone-naive patients with rising prostate-specific antigen after pri-mary treatment for prostate cancer. Urology 62, 102–118, 2003.
38. Chen Y and Hughes-Fulford M: Prostaglandin E2 and the protein ki-nase A pathway mediate arachidonic acid induction of c-fos in humanprostate cancer cells. Br J Cancer 82, 2000–2006, 2000.
39. Hughes-Fulford M, Chen Y, and Tjandrawinata R: Fatty acid regu-lates gene expression and growth of human prostate cancer PC-3 cells.Carcinogenesis 22, 701–707, 2001.
40. Pan Y, Zhang J, Gazi M, and Young C: The cyclooxygenase 2-specificnonsteroidal anti-inflammatory drugs celecoxib and nimesulide inhibitandrogen receptor activity via induction of c-jun in prostate cancercells. Cancer Epidemiol Biomarkers Prev 12, 769–774, 2003.
41. Guo C, Luttrell L, and Price D: Mitogenic signaling in androgen sensi-tive and insensitive prostate cancer cell lines. J Urol 163, 1027–1032,2000.
42. Pruthi R, Derksen J, and Moore D: A pilot study of use of thecyclooxygenase-2 inhibitor celecoxib in recurrent prostate cancer af-ter definitive radiation therapy or radical prostatectomy. Br J Urol 93,275–278, 2004.
110 Nutrition and Cancer 2007



















