
Molecular Machines That Control Genes - Ohio University 720/Molecular Machines... · Molecular...
8
Molecular Machines That Control Genes The activities of our genes are tightly regulated by elaborate complexes of proteins that assemble on DNA. Perturbations in the normal operation of these assemblies can lead to disease by Robert Tjian DANA BURNS-PIZER TRANSCRIPTION FACTORS GENE RNA POLYMERASE 1 2 3 Copyright 1995 Scientific American, Inc.
Transcript of Molecular Machines That Control Genes - Ohio University 720/Molecular Machines... · Molecular...

Molecular MachinesThat Control Genes
The activities of our genes are tightly regulated by elaborate complexes of proteins that assemble on DNA. Perturbations in the
normal operation of these assemblies can lead to disease
by Robert Tjian
DA
NA
BU
RN
S-P
IZE
R
TRANSCRIPTIONFACTORS
GENE
RNAPOLYMERASE
1
2
3
Copyright 1995 Scientific American, Inc.
Asthma, cancer, heart disease, im-mune disorders and viral infec-tions are seemingly disparate
conditions. Yet they turn out to share a surprising feature. All arise to a greatextent from overproduction or under-production of one or more proteins, themolecules that carry out most reactionsin the body. This realization has recent-ly lent new urgency to research aimed atunderstanding, and ultimately manipu-lating, the fascinating biochemical ma-chinery that regulates an essential stepin protein synthesis: the transcriptionof genes. For a protein to be generated,the gene that speciÞes its compositionmust be transcribed, or copied, fromDNA into strands of messenger RNA,which later serve as the templates from
which the protein is manufactured.Even before therapy became a goal,
transcription had long captivated sci-entists for another reason: knowledgeof how this process is regulated prom-ises to clarify some central mysteriesof life. Each cell in the body containsthe same genome, the complement ofsome 150,000 genes that form theblueprint for a human being. How is itthat the original cell of an organismÑ
the fertilized eggÑgives rise to a myri-ad of cell types, each using somewhatdiÝerent subsets of those genes to pro-duce diÝerent mixtures of proteins?And how do the cells of a fully formedbody maintain themselves, increasingand decreasing the amounts of pro-teins they manufacture in response totheir own needs and those of the largerorganism?
To answer these questions and design
ROBERT TJIAN, who was born in Hong Kong, is an investigator with the HowardHughes Medical Institute and professor of molecular and cell biology at the Universityof California, Berkeley. He earned his Ph.D. in biochemistry and molecular biology fromHarvard University in 1976 and conducted research at the Cold Spring Harbor Labora-tory in New York State before joining the faculty of Berkeley in 1979.
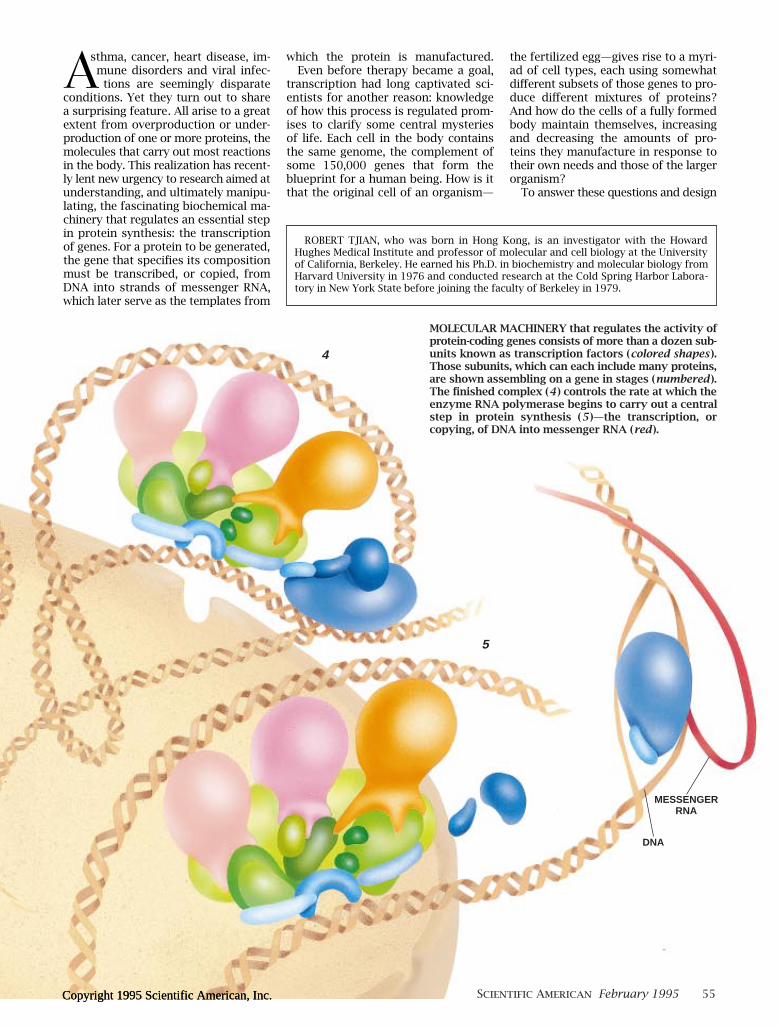
MOLECULAR MACHINERY that regulates the activity ofprotein-coding genes consists of more than a dozen sub-units known as transcription factors (colored shapes ).Those subunits, which can each include many proteins,are shown assembling on a gene in stages (numbered ).The Þnished complex (4 ) controls the rate at which theenzyme RNA polymerase begins to carry out a centralstep in protein synthesis (5 )Ñthe transcription, orcopying, of DNA into messenger RNA (red ).
DNA
MESSENGERRNA
4
5
SCIENTIFIC AMERICAN February 1995 55Copyright 1995 Scientific American, Inc.Copyright 1995 Scientific American, Inc.

drugs able to modulate transcription,investigators need to know somethingabout the makeup of the apparatus thatcontrols reading of the genetic code inhuman cells. After some 25 years of ex-ploration, the overall structure of thatapparatus is becoming clear. Work inmy laboratory at the University of Cali-fornia at Berkeley and at other institu-tions has revealed that one part of theapparatusÑthe engine driving transcrip-tion of most, if not all, human genesÑconsists of some 50 distinct proteins.These proteins must assemble into atight complex on DNA before a specialenzyme, RNA polymerase, can begin tocopy DNA into messenger RNA. Theputative constituents have now beencombined in the test tube to yield a ful-ly operational transcription engine. Stillother proteins essentially plug into re-ceptive sockets on the engine and, inso doing, ÒprogramÓ it, telling it whichgenes should be transcribed and howquickly. Critical details of these interac-tions are emerging as well.
Clues from Bacteria
When my colleagues and I at Berke-ley began focusing on human
genes in the late 1970s, little was knownabout the transcription machinery inour cells. But studies begun early in thatdecade had provided a fairly clear pic-ture of transcription in prokaryotesÑbacteria and other primitive single-celledorganisms that lack a deÞned nucleus.That work eventually lent insight intohuman and other eukaryotic (nucleat-ed) cells and helped to deÞne featuresof transcription that hold true for vir-tually all organisms.
The bacterial research showed thatgenes are essentially divided into twofunctionally distinct regions. The codingregion speciÞes the sequence of aminoacids that must be linked together tomake a particular protein. This sequenceis spelled out by the nucleotides (thebuilding blocks of DNA) in one strandof the DNA double helix; the nucleo-tides are distinguished from one anoth-er by the nitrogen-rich base they car-ryÑadenine (A), thymine (T ), cytosine(C ) or guanine (G ). The other region ofa gene has regulatory duties. It controlsthe rate at which RNA polymerase tran-scribes the coding region of a gene intomessenger RNA.
In bacteria, as in most prokaryotes,the regulatory region, called the promot-er, resides within a stretch of nucleo-tides located a short distanceÑoften asfew as 10 nucleotidesÑin front of (up-stream from) the start of the coding re-gion. For transcription to proceed accu-rately and eÛciently, RNA polymerase
must attach to the promoter. Once it isso positioned, it slides over to the startof the coding region and rides along theDNA, like a train on a track, construct-ing an RNA replica of the coding se-quence. Except in very long genes, thenumber of RNA molecules made at anymoment depends mainly on the rate atwhich molecules of RNA polymeraseattach to the promoter and initiatetranscription.
Interestingly, RNA polymerase is arather promiscuous molecule, unable todistinguish between the promoter andother DNA sequences. To direct the en-zyme to promoters of speciÞc genes,bacteria produce a variety of proteins,known as sigma factors, that bind toRNA polymerase. The resulting com-plexes are able to recognize and attachto selected nucleotide sequences in pro-moters. In this way, sigma factors pro-gram RNA polymerase to bypass allnonpromoter sequences and to lingeronly at designated promoters.
Considering the importance of sigmafactors to the diÝerential activation ofgenes in bacteria, my colleagues and Ibegan our inquiry into the human tran-scription apparatus by searching for sig-malike molecules in human cells. But wehad underestimated the complexity ofthe machinery that had evolved to re-trieve genetic information from ourelaborate genome. It soon became ap-parent that human sigma factors mightnot exist or might not take the sameform as they do in bacteria.
Surprising Complexity
If there were no simple sigma factorsin eukaryotes, how did such cells en-
sure that RNA polymerase transcribedthe right genes at the right time and atthe right rate? We began to see glimmer-ings of an answer once the unusual de-sign of eukaryotic genes was delineated.
By 1983 investigators had establishedthat three kinds of genetic elements,consisting of discrete sequences of nu-cleotides, control the ability of RNApolymerase to initiate transcription inall eukaryotesÑfrom the single-celledyeast to complex multicellular organ-isms. One of these elements, generallylocated close to the coding region, hadbeen found to function much like a bac-terial promoter. Called a core promoter,it is the site from which the polymerasebegins its journey along the coding re-gion. Many genes in a cell have similarcore promoters.
Walter SchaÝner of the University ofZurich and Steven Lanier McKnight ofthe Carnegie Institution of Washington,among others, had additionally iden-tiÞed an unusual set of regulatory ele-
ments called enhancers, which facilitatetranscription. These sequences can belocated thousands of nucleotides up-stream or downstream from the corepromoterÑthat is, incredibly far fromit. And subsequent studies had uncov-ered the existence of silencers, whichhelp to inhibit transcription and, again,can be located a long distance from thecore promoter.
In a somewhat imperfect analogy, if the core promoter were the ignitionswitch of a car engine, enhancers wouldact as the accelerator, and silencers asthe brakes. Eukaryotic genes can includeseveral enhancers and silencers, andtwo genes may contain some identicalenhancer or silencer elements, but notwo genes are precisely alike in the com-bination of enhancers and silencers they
56 SCIENTIFIC AMERICAN February 1995
Anatomy of theTranscription Apparatus
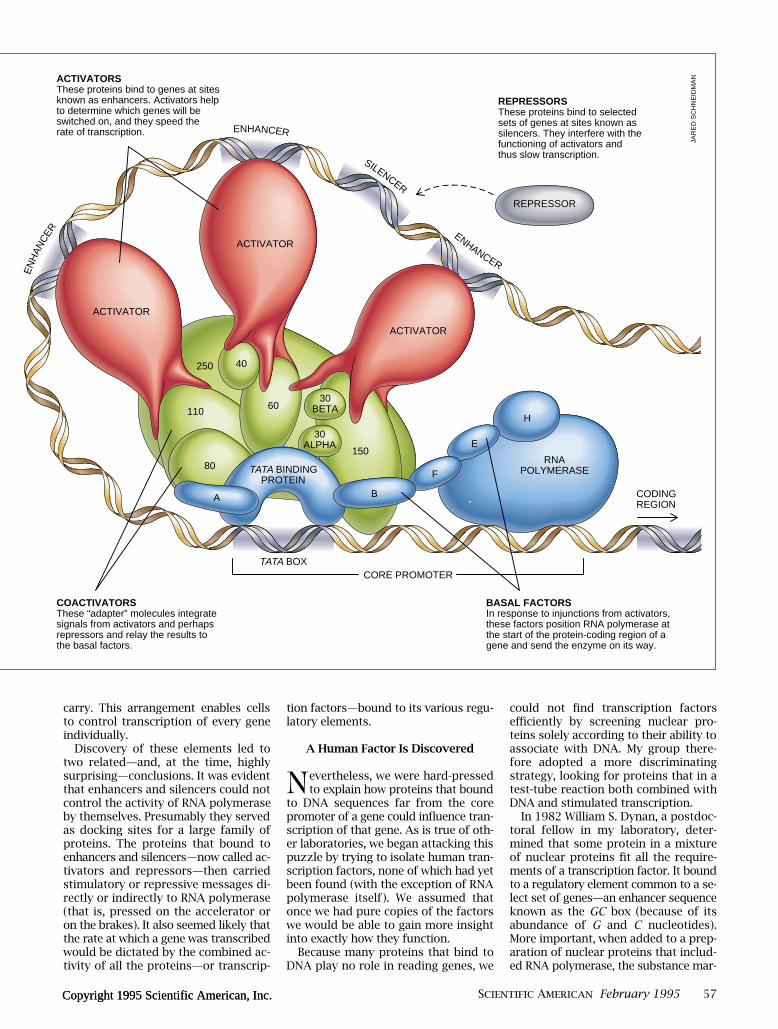
The molecular apparatus controllingtranscription in human cells consists offour kinds of components. Basal fac-tors (blue shapes at bottom), generallynamed by single letters, are essentialfor transcription but cannot by them-selves increase or decrease its rate.That task falls to regulatory moleculesknown as activators (red ) and repres-sors (gray ); these can vary from geneto gene. Activators, and possibly re-pressors, communicate with the basalfactors through coactivators (green)—proteins that are linked in a tight com-plex to the TATA binding protein (TBP),the first of the basal factors to land on aregulatory region of genes known as thecore promoter. Coactivators are namedaccording to their molecular weights(in kilodaltons).
Copyright 1995 Scientific American, Inc.
carry. This arrangement enables cells to control transcription of every geneindividually.
Discovery of these elements led totwo relatedÑand, at the time, highlysurprisingÑconclusions. It was evidentthat enhancers and silencers could notcontrol the activity of RNA polymeraseby themselves. Presumably they servedas docking sites for a large family ofproteins. The proteins that bound toenhancers and silencersÑnow called ac-tivators and repressorsÑthen carriedstimulatory or repressive messages di-rectly or indirectly to RNA polymerase(that is, pressed on the accelerator oron the brakes). It also seemed likely thatthe rate at which a gene was transcribedwould be dictated by the combined ac-tivity of all the proteinsÑor transcrip-
tion factorsÑbound to its various regu-latory elements.
A Human Factor Is Discovered
Nevertheless, we were hard-pressedto explain how proteins that bound
to DNA sequences far from the corepromoter of a gene could inßuence tran-scription of that gene. As is true of oth-er laboratories, we began attacking thispuzzle by trying to isolate human tran-scription factors, none of which had yetbeen found (with the exception of RNApolymerase itself ). We assumed thatonce we had pure copies of the factorswe would be able to gain more insightinto exactly how they function.
Because many proteins that bind toDNA play no role in reading genes, we
could not Þnd transcription factorseÛciently by screening nuclear pro-teins solely according to their ability toassociate with DNA. My group there-fore adopted a more discriminatingstrategy, looking for proteins that in atest-tube reaction both combined withDNA and stimulated transcription.
In 1982 William S. Dynan, a postdoc-toral fellow in my laboratory, deter-mined that some protein in a mixtureof nuclear proteins Þt all the require-ments of a transcription factor. It boundto a regulatory element common to a se-lect set of genesÑan enhancer sequenceknown as the GC box (because of itsabundance of G and C nucleotides).More important, when added to a prep-aration of nuclear proteins that includ-ed RNA polymerase, the substance mar-
SCIENTIFIC AMERICAN February 1995 57
30ALPHA
30BETA
TATA BOX
TATA BINDINGPROTEIN
40
60
80
CODINGREGION
110H
E
250
150
A B
FRNA
POLYMERASE
CORE PROMOTER
REPRESSOR
SILENCER
EN
HA
NC
ER
ENHANCER
ENHANCER
ACTIVATORSThese proteins bind to genes at sites known as enhancers. Activators help to determine which genes will be switched on, and they speed the rate of transcription.
REPRESSORSThese proteins bind to selected sets of genes at sites known as silencers. They interfere with the functioning of activators and thus slow transcription.
COACTIVATORSThese “adapter” molecules integrate signals from activators and perhaps repressors and relay the results to the basal factors.
BASAL FACTORSIn response to injunctions from activators, these factors position RNA polymerase at the start of the protein-coding region of a gene and send the enzyme on its way.
ACTIVATOR
ACTIVATOR
ACTIVATOR
JAR
ED
SC
HN
EID
MA
N
Copyright 1995 Scientific American, Inc.Copyright 1995 Scientific American, Inc.
kedly increased the transcription onlyof genes carrying the GC box. Thus, wehad identiÞed the Þrst human transcrip-tion factor able to recognize a speciÞcregulatory sequence. We called it speci-Þcity protein 1 (Sp1).
We immediately set out to purify themolecule. One daunting aspect of thiswork was the fact that transcription fac-tors tend to appear only in minusculequantities in cells. Typically, less than a thousandth of a percent of the totalprotein content of a human cell consistsof any particular factor. In 1985 JamesT. Kadonaga in my laboratory found away to overcome this substantial tech-nical barrierÑand in the process intro-duced a powerful new tool that hassince been used to purify countlesstranscription factors and other scarceDNA binding proteins.
Because Sp1 selectively recognizedthe GC box, Kadonaga synthesized DNAmolecules composed entirely of thatbox and chemically anchored them tosolid beads. Then he passed a complexmixture of human nuclear proteins overthe DNA, predicting that only Sp1 wouldstick to it. True to plan, when he sepa-rated the bound proteins from the syn-thetic DNA, he had pure Sp1.
From studies carried out by MarkPtashne and his colleagues at HarvardUniversity, we knew that bacterial tran-scription regulators are modular pro-teins, in which separate regions per-form distinct tasks. Once we learnedthe sequence of amino acids in Sp1, wetherefore looked for evidence of dis-tinct modules and noted at least two in-teresting ones.
One end of the molecule contained aregion that obviously folded up intothree Òzinc Þngers.Ó Zinc-Þnger struc-tures, in which parts of a protein foldaround a zinc atom, are now known toact as the ÒhooksÓ that attach many ac-tivator proteins to DNA. But at the timeSp1 was only the second protein foundto use them. Aaron Klug and his col-leagues at the Medical Research Coun-cil in England had discovered zinc Þn-gers, in a frog transcription factor, justa short time before [see ÒZinc Fingers,Óby Daniela Rhodes and Aaron Klug;SCIENTIFIC AMERICAN, February 1993].
The other end of Sp1 contained a do-main consisting of two discrete seg-ments Þlled with a preponderance ofthe amino acid glutamine. We stronglysuspected that this region played animportant role during transcription be-cause of a striking Þnding. In test-tubeexperiments, mutant Sp1 moleculeslacking the domain could bind to DNAperfectly well, but they failed to stimu-late gene transcription. This outcomeindicated that Sp1 did not aÝect tran-scription solely by combining with DNA;it worked by using its glutamine-richsegmentÑnow known as an activationdomainÑto interact with some otherpart of the transcription machinery. Thequestion was, which part?
In 1988 when we began searching for
the target of Sp1, we had some idea ofwhere it lay. Our guess was based on anemerging understanding of the so-calledbasal transcription complex, one partof which seemed to be a likely target.
Closing in on a Target
In the mid-1980s Robert G. Roederand his colleagues at the Rockefeller
University had shown that RNA poly-merase cannot transcribe eukaryoticgenes unless several other transcriptionfactorsÑnow called basal factorsÑalsocollect on the core promoter. And overthe course of the 1980s, RoederÕs labo-ratory and others had identiÞed at leastsix of those essential factors, called A,B, D, E, F and H.
In a test tube, this assembly of factorsenabled RNA polymerase to transcribea bound gene at a basalÑlow and in-variantÑrate, but it could not by itselfmodulate that rate. It was as if someonehad constructed and switched on theengine of a car but had lost use of thesteering wheel, the accelerator and thebrakes. For instance, when my groupmixed the components of the complex(including RNA polymerase) with a genecontaining a GC box, we obtained a low,unchanging level of transcription. Wesaw a marked increase in transcriptiononly when we incorporated Sp1 into themixture.
By the late 1980s it was apparent thathuman cells harbor at least two sep-arate classes of transcription factors.Basal factors are required for initiationof transcription in all genes; other pro-teinsÑactivators and repressorsÑdic-tate the rate at which the basal complexinitiates transcription. DiÝerent genesare controlled by distinct combinations
58 SCIENTIFIC AMERICAN February 1995
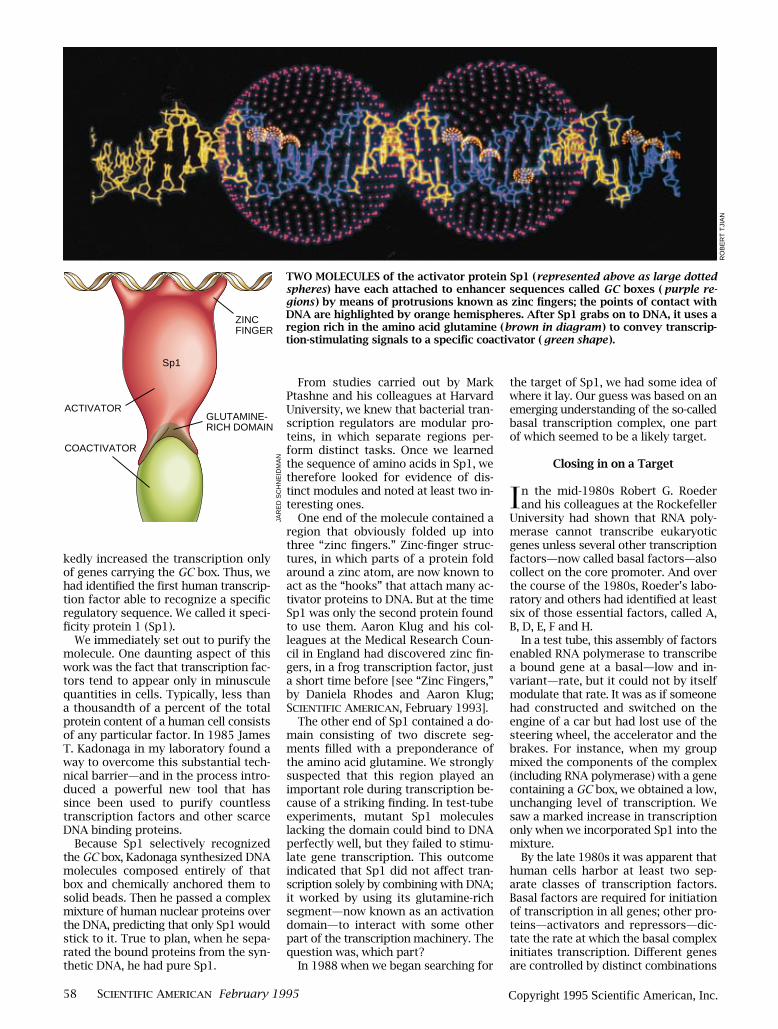
TWO MOLECULES of the activator protein Sp1 (represented above as large dottedspheres) have each attached to enhancer sequences called GC boxes ( purple re-gions ) by means of protrusions known as zinc Þngers; the points of contact withDNA are highlighted by orange hemispheres. After Sp1 grabs on to DNA, it uses aregion rich in the amino acid glutamine (brown in diagram ) to convey transcrip-tion-stimulating signals to a speciÞc coactivator ( green shape ).
RO
BE
RT
TJI
AN
Sp1
ACTIVATOR
COACTIVATOR
ZINCFINGER
GLUTAMINE-RICH DOMAIN
JAR
ED
SC
HN
EID
MA
N
Copyright 1995 Scientific American, Inc.

of activators and repressors. We nowsuspect that in the body the basal com-plex arises spontaneously only rarely;most of the time, cells depend on acti-vators to initiate its construction.
These various discoveries suggestedthat the glutamine-rich domain of Sp1enhanced transcription by contacting abasal factor. More speciÞcally, we sus-pected that Sp1 latched on to factor D,and facilitated its attachment to thepromoter. We focused on this subunitbecause Phillip A. Sharp and StephenBuratowski of the Massachusetts Insti-tute of Technology had shown that itcan land on the core promoter beforeall other basal factors and can facilitateassembly of the complete basal engine.In fact, factor D is the only basal com-ponent able to recognize DNA. It bindsselectively to a sequence called theTATA box, found in the core promot-ers of many eukaryotic genes.
To pursue our hypothesis, we neededto know more about the compositionof factor D, which we assumed was asolitary protein. Other investigators alsowanted to know its makeup, and so therace was on to attain pure copies. Isola-tion from human cells proved morechallenging than anyone anticipated.Consequently, many groups eventuallytried their luck with yeast cells. Finally,in 1989, several laboratories indepen-dently succeeded in isolating a yeastprotein that displayed the expectedproperties of factor D. The protein,named TBP (for TATA binding protein),recognized and bound selectively to theTATA box and led to a low level of tran-scription when it was joined at the corepromoter by RNA polymerase and oth-er constituents of the basal machinery.
Believing that the TBP protein wasfactor D itself, we undertook to testthis idea in additional studies. Once wedid that, we intended to determine ex-actly which regions of TBP were con-tacted by Sp1 and other regulators. Lit-tle did we know that we were about tobe completely thwartedÑand to makea critical discovery.
Unexpected Trouble
When B. Franklin Pugh in our labo-ratory replaced the impure prepa-
rations of factor D previously used inour test-tube reactions with puriÞedmolecules of TBP, he had no troublereplicating the earlier Þnding that suchsubstitution in no way disrupted basaltranscription. To our surprise and con-sternation, though, he found that Sp1was no longer able to inßuence the bas-al machinery. We had to conclude thatfactor D and TBP were not, in fact,equivalent and that factor D actually
consisted of TBP plus other subunits.(It is now known that many transcrip-tion factors consist of more than oneprotein.) Apparently, those subunitswere not needed for operation of thebasal machinery, but they were essen-tial to regulation of that machinery byactivators.
In other words, these additional com-ponents were not themselves activa-tors, for they did not bind to speciÞcsequences in DNA. Nor were they basalfactors, because low, unregulated levelsof transcription could be achieved with-out them. They seemed to constitute athird class of transcription factor, whichwe called coactivators. We further pro-posed that coactivators, not TBP, werethe targets for the protein binding do-mains of activators. We envisioned thatactivators would bind to selected coac-tivators to speed up the rate at whichthe basal complex set molecules of RNApolymerase in motion.
We were attracted to this scenario be-cause we had diÛculty imagining howa single protein, TBP, would have enoughbinding sites to accommodate all theactivators made by human cells. But ifthe coactivators that were tightly linkedto TBP bore multiple binding domains,the coactivators could collectively pro-vide the docking sites needed to relay
messages from hundreds or thousandsof activators to the transcription engine.
It was Pugh who originally proposedthat coactivators might function assuch adapter molecules. His data soonconvinced me he was probably correct,but not everyone in our laboratoryagreed. Indeed, our weekly meetings inearly 1990 were often punctuated byheated discussions. Not surprisingly,when the coactivator concept was pre-sented to other workers in the Þeld,they, too, expressed considerable skep-ticism. This reaction to an unexpectedand complicating result was probablyjustiÞed at that stage, because our datawere only suggestive, not conclusive. Wehad not yet isolated a single coactivator.
Coactivators: The Missing Links
To satisfy ourselves and the scientif-ic community that we were correct,
we had to devise an experimental pro-cedure that would unambiguously es-tablish whether coactivators existed andoperated as the relays we envisioned.For approximately two years after Pughformulated the coactivator hypothesis,we struggled to purify an intact andfunctional complex containing TBP andall the other associated constituents offactor D. I must admit to some dark mo-
SCIENTIFIC AMERICAN February 1995 59
ACTIVATORS CAN COMMUNICATE with one another, not only with DNA and coac-tivators, as is demonstrated in an experiment involving Sp1. Copies of the protein(dark dots) each recognized and bound to GC boxes at the tips of a string of DNA.After binding, they joined togetherÑcausing DNA to loop out in the process.
RO
BE
RT
TJI
AN
Copyright 1995 Scientific American, Inc.Copyright 1995 Scientific American, Inc.
ments when it seemed the rather un-popular coactivator hypothesis mightbe based on some error in our studies.
The breakthrough Þnally came in1991, when Brian D. Dynlacht, TimothyHoey, Naoko Tanese and Robert Wein-zierlÑgraduate students and postdoc-toral fellows in our laboratoryÑfoundan ingenious way to isolate pure copiesof factor D. Subsequent biochemicalanalyses revealed that, aside from TBP,the complete unit included eight previ-ously unknown proteins. Because wedid not yet have proof that these pro-teins could function as coactivators, wereferred to them more generically asTBP-associated factors, or TAFs.
We became convinced that TAFs doindeed convey molecular signals fromactivators to the basal transcription ap-
paratus after we separated the boundproteins from TBP and completed sev-eral more experiments. For instance, wewere able to show that mixing of the ac-tivator Sp1 with basal factors and RNApolymerase enhanced production ofmessenger RNA from a gene containinga GC box only when TAFs were addedas well. Later, Jin-Long Chen, a graduatestudent, combined puriÞed TBP and theeight isolated TAFs in a test tube alongwith a human gene and the rest of thebasal transcription machinery. The var-ious proteins assembled on the geneand proved able to respond to severaldiÝerent types of activator proteins.These activators, we later showed, pro-duced their eÝects by coupling directlywith selected TAFs. Together the coac-tivators in factor D do indeed consti-
tute a kind of central processing unitthat integrates the regulatory signalsissued by DNA-bound activators.
A Universal Theme
The complexes formed by activators,coactivators and the basal machin-
ery appear to be human equivalents ofsigma factors; they, too, draw RNApolymerase to speciÞc genes at speciÞcrates. In a way, the complexes can beviewed as sigma factors that have beenelaborated into many subunits. Gratify-ingly, recent evidence from our groupand others suggests we have uncovereda universal mode of gene regulation ineukaryotes. Those studies conÞrm thatcoactivators also exist in yeast and thatfactor D consists of multiple subunitsin fungi as well as in humans.
As satisfying as these results are,they do not fully explain how bindingof activators to enhancers and to coac-tivators inßuences the rate at whichRNA polymerase transcribes genes inliving cells. It may be that linkage of ac-tivators to enhancers causes DNA tobend in a way that brings the enhanc-ers closer to one another and to the corepromoter. This arrangement may helpactivators (alone or in concert with oneanother) to dock with coactivators andposition factor D on the promoter. Thisstep, in turn, would facilitate assemblyof the complete basal complex. Forma-tion of this complex may distort the un-derlying DNA in a way that enables RNApolymerase to begin its journey alongthe coding region.
Researchers know less about thefunctioning of repressors. Nevertheless,many of us think repressors may alsobind to coactivators at times. This bind-
60 SCIENTIFIC AMERICAN February 1995
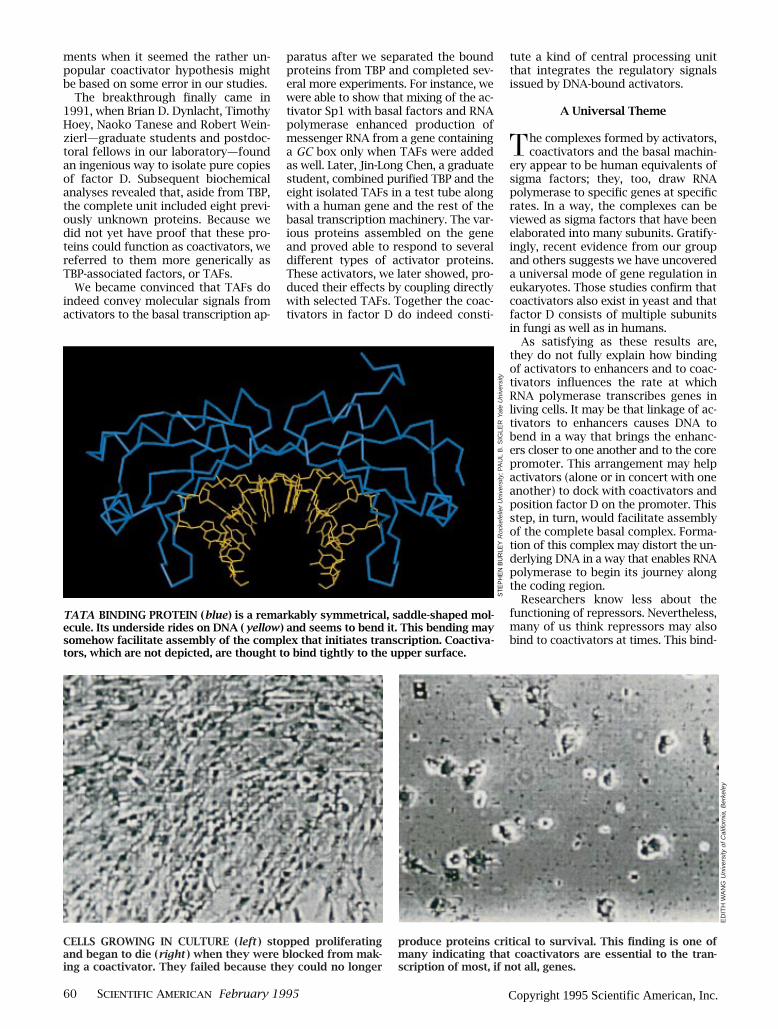
TATA BINDING PROTEIN (blue) is a remarkably symmetrical, saddle-shaped mol-ecule. Its underside rides on DNA ( yellow ) and seems to bend it. This bending maysomehow facilitate assembly of the complex that initiates transcription. Coactiva-tors, which are not depicted, are thought to bind tightly to the upper surface.
STE
PH
EN
BU
RLE
Y R
ocke
felle
r U
nive
rsity
; PA
UL
B. S
IGLE
R Y
ale
Uni
vers
ity
CELLS GROWING IN CULTURE (left ) stopped proliferatingand began to die (right ) when they were blocked from mak-ing a coactivator. They failed because they could no longer
produce proteins critical to survival. This Þnding is one ofmany indicating that coactivators are essential to the tran-scription of most, if not all, genes.
ED
ITH
WA
NG
Uni
vers
ity o
f Cal
iforn
ia, B
erke
ley
Copyright 1995 Scientific American, Inc.

ing could inhibit transcription by pre-venting activators from attaching totheir usual sites on coactivators. Othertimes repressors might bypass the ba-sal machinery, blocking transcriptionby preventing activators from connect-ing with enhancers.
Although there are gaps in our knowl-edge, we can now begin to sketch outan explanation as to why diÝerent cellsmake diÝerent mixtures of proteinsduring embryonic development and inmature organisms. A gene will be tran-scribed at a measurable rate only if thevarious activators it needs are presentand can successfully overcome the in-hibitory eÝects of repressors. Cells varyin the proteins they make because theycontain distinct batteries of activatorsand repressors. Of course, this scenariobegs the question of how cells decidewhich transcription factors to producein the Þrst place, but progress is beingmade on that front as well.
Therapies of Tomorrow
How might investigators use ournewly acquired knowledge of gene
regulation to develop drugs for com-bating life-threatening diseases involv-ing excessive or inadequate transcrip-tion of a gene? In theory, blocking se-lected activators from attaching toenhancers or coactivators should de-press unwanted transcription, and sta-bilizing the transcription machinery ona gene should counteract undesirablyweak transcription.
Blockade could be achieved by Þttinga molecular ÒplugÓ into an activator,thereby preventing its interaction witha coactivator, or by enticing an activatorto attach to a decoy that resembles acoactivator. Stabilization of a complexmight be achieved by deploying mole-cules that would strengthen the inter-action between activators and DNA orbetween activators and coactivators.Such approaches are remote today, butit is exciting to consider a sampling ofthe applications that might eventuallybe possible.
Take, for example, the human immu-nodeÞciency virus (HIV), which causesAIDS. To reproduce itself in humancells, HIV needs the viral transcriptionfactor TAT to enhance transcription ofHIV genes. If TAT could be inhibited bysome agent that recognized TAT butignored human transcription factors,replication of the virus might be haltedwithout aÝecting production of pro-teins needed by the patient.
Conversely, treatment of some disor-dersÑfor instance, hypercholesterole-miaÑmight involve enhancing the tran-scription of selected genes. Hypercho-
lesterolemia increases a personÕs riskfor heart disease. Cholesterol accumu-lates to destructive levels in the bloodwhen low-density lipoprotein (LDL),otherwise known as the bad cholester-ol, is not removed eÛciently. In theory,the disease could be corrected by turn-ing up transcription of the gene for theLDL receptor in liver cells. This recep-tor helps to clear LDL from the blood.This idea may soon be testable, becausestudies by Michael S. Brown and JosephL. Goldstein of the University of TexasHealth Science Center at Dallas are teas-ing apart the speciÞc molecular consti-tuents of the apparatus that regulatestranscription of the receptor gene.
Until recently, no one put much eÝortinto screening small molecules, naturalproducts or other compounds for theirability to modulate transcription. Evenso, a number of drugs already on themarket have been found by chance towork by altering the activity of tran-scription factors. One of these, RU 486(the French ÒabortionÓ pill), repressesthe function of particular steroid re-ceptors, a class of activators that directembryonic development. Similarly, theimmunosuppressants cyclosporine andFK506 suppress transcription of a genewhose protein product is needed by cer-tain cells of the immune system. Thesedrugs act indirectly, however. They ac-tivate an enzyme that impedes the func-tioning of a transcription factor for thegene.
As time goes by, the precise combi-nation of transcription factors that reg-
ulate individual genes is sure to be iden-tiÞed. And drug developers will proba-bly use this information to devise so-phisticated compounds for Þghtingcancer, heart disease, immune disor-ders, viral infections, AlzheimerÕs dis-ease and perhaps even the aging pro-cess. How well these agents will suc-ceed is anybodyÕs guess, but it is likelythat therapies of the future will beneÞtin one way or another from basic re-search into transcriptionÑresearch thatbegan not out of a wish to design drugsbut rather out of a simple desire to getto the heart of the molecular machinerythat controls the activity of our genes.
SCIENTIFIC AMERICAN February 1995 61
FURTHER READING
TRANSCRIPTIONAL SELECTIVITY OF VIRALGENES IN MAMMALIAN CELLS. Steven Mc-Knight and Robert Tjian in Cell, Vol. 46,No. 6, pages 795Ð805; September 12,1986.
TRANSCRIPTIONAL REGULATION IN MAM-MALIAN CELLS BY SEQUENCE-SPECIFICDNA BINDING PROTEINS. Pamela J.Mitchell and Robert Tjian in Science,Vol. 245, pages 371Ð378; July 28, 1989.
EUKARYOTIC COACTIVATORS ASSOCIAT-ED WITH THE TATA BOX BINDING PRO-TEIN. G. Gill and R. Tjian in CurrentOpinion in Genetics and Development,Vol. 2, No. 2, pages 236Ð242; April1992.
TRANSCRIPTIONAL ACTIVATION: A COM-PLEX PUZZLE WITH FEW EASY PIECES. R.Tjian and T. Maniatis in Cell, Vol. 77,No. 1, pages 5Ð8; April 8, 1994.
ONE OF THE WORLDÕS TALLEST WOMEN reportedly reached her height of sevenfeet, seven and a quarter inches because her pituitary gland produced an excess ofgrowth hormone. Investigators hope eventually to treat this and many other disor-ders by increasing or decreasing the transcription of selected genes.
BE
TT
INA
CIR
ON
E P
hoto
Res
earc
hers
, Inc
.
Copyright 1995 Scientific American, Inc.Copyright 1995 Scientific American, Inc.


















