
Molecular cloning, functional expression and tissue ... · INTRODUCTION Pyridoxal 5’-phosphate...
27
Human Pyridoxal Phosphatase : Molecular cloning, functional expression and tissue distribution By Young Min Jang (1) , Dae Won Kim (2) , Tae-Cheon Kang (3) , Moo Ho Won (3) , Nam-In Baek (4) , Byung Jo Moon (1) , Soo Young Choi (2) a and Oh-Shin Kwon (1) * 1) Department of Biochemistry, Kyungpook National University, Taegu, Korea. 2) Department of Genetic Engineering, Division of Life Sciences, Hallym University, Chunchon 200-702, Korea 3) Department of Anatomy, College of Medicine, Hallym University, Chunchon 200-702, Korea 4) Graduate School of Biotechnology & Plant Metabolism Research Center, Kyunghee University, Suwon 449-701, Korea Running title : human Recombinant Pyridoxal Phosphatase * To whom correspondence should be addressed : Oh-Shin Kwon, Department of Biochemistry, Kyungpook National University, Taegu, 702-701, Korea Tel : 82-53-950-6356, Fax : 82-53-943-2762 e-mail address : [email protected] a Co-corresponding author : [email protected] Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc. JBC Papers in Press. Published on September 30, 2003 as Manuscript M309619200 by guest on August 9, 2018 http://www.jbc.org/ Downloaded from
Transcript of Molecular cloning, functional expression and tissue ... · INTRODUCTION Pyridoxal 5’-phosphate...

Human Pyridoxal Phosphatase :
Molecular cloning, functional expression and tissue distribution
By
Young Min Jang (1), Dae Won Kim (2), Tae-Cheon Kang (3), Moo Ho Won (3), Nam-In Baek (4),
Byung Jo Moon (1), Soo Young Choi (2) a and Oh-Shin Kwon (1) *
1) Department of Biochemistry, Kyungpook National University, Taegu, Korea.
2) Department of Genetic Engineering, Division of Life Sciences, Hallym University,
Chunchon 200-702, Korea
3) Department of Anatomy, College of Medicine, Hallym University, Chunchon 200-702,
Korea
4) Graduate School of Biotechnology & Plant Metabolism Research Center, Kyunghee
University, Suwon 449-701, Korea
Running title : human Recombinant Pyridoxal Phosphatase
* To whom correspondence should be addressed :
Oh-Shin Kwon,
Department of Biochemistry, Kyungpook National University, Taegu, 702-701, Korea
Tel : 82-53-950-6356, Fax : 82-53-943-2762
e-mail address : [email protected]
a Co-corresponding author : [email protected]
Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on September 30, 2003 as Manuscript M309619200 by guest on A
ugust 9, 2018http://w
ww
.jbc.org/D
ownloaded from

Acknowledgements :
"This work was supported by the Grant No. R01-2002-000-00008-0 from the Basic Research
Program of the Korea Science & Engineering Foundation, and Korea Health 21 R&D Project
Grant (02-PJ1-PG10-20706-0002) from Ministry of Health and Welfare, Korea."
The nucleotide sequence(s) reported in this paper has been submitted to the GenBankTM/EBI
Data bank with accession number(s) AY125047, AY366300.
The abbreviation used are: PLP, pyridoxal 5’-phosphate; PNP, pyridoxine 5’-phosphate; PMP,
pyridoxamine 5’-phosphate; PLPP, PLP phosphatase; EST, expressed sequence tag; RACE, rapid
amplification of cDNA ends.
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

SUMMARY
Pyridoxal phosphatase catalyzes the dephosphorylation of pyridoxal 5’-phosphate (PLP) and
pyridoxine 5’-phosphate. A human brain cDNA clone was identified to the PLP phosphatase on
the basis of peptide sequences obtained previously. The cDNA predicts a 296-amino acid protein
with a calculated Mr of 31698. The open reading frame is encoded by two exons located on
human chromosome 22q12.3, and the exon-intron junction contains the GT/AG consensus splice
site. In addition, a full-length mouse PLP phosphatase cDNA of 1978 base pairs was also
isolated. Mouse enzyme encodes a protein of 292 amino acids with Mr of 31512, and it is
localized on chromosome 15.E1. Human and mouse PLP phosphatase share 93% identity in
protein sequence. A BLAST search revealed the existence of putative proteins in organism
ranging from bacteria to mammals. Catalytically active human PLP phosphatase was expressed
in E. coli, and characteristics of the recombinant enzyme were similar to those of erythrocyte
enzyme. The recombinant enzyme displayed Km and kcat values for pyridoxal of 2.5 M and
1.52 s-1, respectively. Human PLP phosphatase mRNA is differentially expressed in a tissue-
specific manner. A single mRNA transcript of 2.1 kb was detected in all human tissues examined,
and was highly abundant in the brain. Obtaining the molecular properties for the human PLP
phosphatase may provide new direction for investigating metabolic pathway involving vitamin
B6.
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

INTRODUCTION
Pyridoxal 5’-phosphate (PLP) is the coenzymatically active form of vitamin B6, and plays
an important role in maintaining the biochemical homeostasis of the body (1,2). Thus, the other
nutritionally available vitamer forms in this family must be converted to PLP. The enzymes that
are conventionally involved in the metabolism are an ATP dependent pyridoxal kinase (EC
2.7.1.35) (3,4) and a flavin mononucleotide (FMN)–dependent pyridoxine 5’-phosphate (PNP)
oxidase (EC 1.4.3.5) (5,6). On the other hand, the preferred degradation route from PLP to 4-
pyridoxic acid involves the dephosphorylation of PLP by the phosphatase (7) followed separately
by the actions of aldehyde oxidase and NAD-dependent dehydrogenase (8,9).
The major pathways of vitamin B6 metabolism have been established for decades, but little
is known how the concentration of PLP is controlled in mammalian tissues. Studies on the
homeostasis have shown that there is not a single mechanism. The factors for the regulation of
PLP may include catabolism of PLP by PLP phosphatase, activities of pyridoxal kinase and PNP
oxidase, degree of protein binding of the synthesized coenzyme and transport of the precursors
(10,11). Although PNP oxidase plays a kinetic role in regulating the level of PLP formation
(12,13), PLP availability is mainly dependant on protein binding and phosphatase action
(10,14,15). Thus, the phosphatase action may be important in the regulation of PLP
concentration.
A PLP specific phosphatase has been purified from human erythrocytes (16). The molecular
properties including the molecular weight, pH optimum, substrate specificity and metal
requirement indicate that the PLP phosphatase distinct from other known phosphatases. Alkaline
phosphatase (EC 3.1.3.1) and acid phosphatase (EC 3.1.3.2) also hydrolyze PLP and
pyridoxamine 5’-phosphate (PMP), but they have broad substrate specificity for
phosphomonoesters (17,18). Alkaline phosphatase is resistant to fluoride and inhibited by L(+)-
tartrate and levamisole, while the PLP specific phosphatase not. Acid phosphatase is not
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

inhibited by fluoride and EDTA. The PLP phosphatase is most active at physiological pH, and
requires Mg2+ for activity (19). It is a dimer with a molecular mass of 64kDa, and has a Km
value of approximately 1 M for PLP, which is at 10-fold lower than that reported with other
phosphatases (16). Data obtained in chemical modification studies revealed that PLP
phosphatase has essential residues such as cysteine, arginine, histidine and a carboxylate group at
or near the active site (20,21,22).
Although the PLP specific phosphatase has been purified and characterized, a cDNA
encoding PLP phosphatase has not yet been identified. Here we cloned cDNAs encoding PLP
phosphatase from human and mouse. In addition, we describe the high efficiency expression of
the protein in E. coli with its kinetic properties and the tissue-specific expression.
EXPERIMENTAL PROCEDURES
Materials
We purchased a human brain cDNA library as well as a dot blot array containing
poly(A)+RNAs from human tissues from CLONTECH (Palo Alto, CA). Restriction
endonuclease and other cloning reagents were purchased from New England Biolabs, Inc.
(Beverly, MA) or Promega (Madison, WI). Double-stranded DNA probes were radiolabeled with
[ -32P]dCTP (3000 Ci/mmol) from Amersham Pharmacia Biotech (Buckinghamshire, United
Kingdom) using a commercial random priming kit (Amersham Pharmacia Biotech, Uppsala,
Sweden). All other reagent-grade chemicals were obtained from standard suppliers.
Identification and Cloning of human brain PLP phosphatase (hPLPP) cDNA
BLAST searches, conducted with PLP phosphatase peptides (AQGVLFDCDGVL and
AVLVGYDEHFSFAK) as query sequences, revealed a full match with the predicted amino acid
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

sequence of a cDNA (GenBankTM/EBI accession number NM020315). This clone was used to
design PCR primers for the cloning of human PLP phosphatase cDNA.
We employed 5’-rapid amplification of cDNA ends (RACE) using hPLPP-specific primer 1
(5’-CGATGCTGAAGTTCTCCGTGATGCAC-3’), AP1 anchor primer, and Marathon Ready
cDNA (human whole brain, CLONTECH) as a template. PCR was carried out with betaine in a
GeneAmp PCR system 2400 (PerkinElmer Life Sciences) for 30 cycles of denaturation (94°C,
30 s annealing; 60°C, 1 min; 72°C, 2 min extension). The resulting PCR product of 737 bp
fragment revealed that the 5’-untranslated region (UTR) is completely matched except 9 bp
shorter than the cDNA from lung carcinoma. The full-length ORF of 891 bp was amplified using
the hPLPP-specific primers (sense, 5’-GGATCCGGCTGCATGGCGCGCTGCGA-3’; antisense,
5’-GGATCCGGGGTGG GCCTGTGGCTGCAG-3’). The PCR product was cloned into the
pGEM-T vector (Promega, Madison, WI) and sequenced (GenBankTM accession number
AY125047).
A multiple protein sequence alignment of the human PLP phosphatase clone, along with the
sequences most closely related to it, was performed using the ClustalW (version 1.74) program
(23), and the resulting aligned sequence were shaded using Boxshade. Identification of the
chromosomal localization of the human PLP phosphatase was performed by a BLASTN search
of the human genome sequences of the National Center for Biotechnology Information (NCBI).
Cloning of mouse brain PLP phosphatase (mPLPP) cDNA
We also identified a mouse expressed sequence tag (EST) clone (GenBankTM accession
number AK043228) with high similarity to human PLP phosphatase. To obtain a full-length
cDNA corresponding to this EST clone, 5’-RACE and 3’-RACE were performed using mouse
brain cDNA (Seegene, Seoul, Korea) to the manufacturer’s protocol. Two consecutive PCR
reactions using Ex Taq polymerase (Takara) were performed as follow: (1) 5’ RACE , 94°C for 3
min followed by 30 cycles of 94°C for 1min and 63°C for 40 s, followed by primer extension at
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

72°C for 5 min with 5’ race primer (5’-GTCTACCAGGCATTCGCTTCAT-3’) and mPLPP-
specific primer 1 (5’-ATCCCAGCCTCCCTTCACT-3’); and (2) 3’ RACE , 94°C for 3 min
followed by 30 cycles of 94°C for 1 min, 56°C for 40 s, and primer extension at 72°C for 5 min
with 3’ race primer (5’-CTGTGAATGCTGCGACTACGAT-3’) and mPLPP-specific primer 2
(5’-ATGGAAGGGAGGCTGGGATCCAGG-3’). The PCR products were cloned and sequenced.
Expression in E. coli and Purification of Human PLP phosphatase
The PLP phosphatase cDNA was cloned between the BamHI of the bacterial expression
vector pQE30 (Qiagen) after PCR amplification. Transformants of E. coli M15/pRER4 with the
resulting pQE30-hPLPP construct were grown at 37°C in 200 ml of LB medium with 100 g/ml
ampicillin and 25 g/ml kanamycin. The plasmid pREP4 constitutively expresses the Lac
repressor protein encoded by the lacI gene to reduce the basal level of expression (Qiagen).
When that culture had grown to an A600 of 0.6, isopropyl -D-thiogalactopyranoside was added
to a final concentration of 1 mM. After inducing the expression of the PLP phosphatase protein
for 16 h at 25°C cells were harvested, washed, and resuspended in 20 ml of 20 mM Tris-HCl
buffer, pH 7.4, containing 200 mM NaCl and 20 mM imidazole.
The cell suspension was sonicated, and the lysate was cleared by centrifugation at 10,000
×g and 4°C for 30 min. The supernatant was then poured into the column loaded with the nickel-
nitrilotriacetic acid (Ni-NTA) agarose (Qiagen), washed with Tris buffer containing 40 mM
imidazole, and eluted protein was eluted with 200 mM imidazole. The purity of the eluted
protein was evaluated by 12% SDS-PAGE using coomassie blue staining to visualize the protein.
Enzyme Assay and Kinetic Characterization
The enzymatic activity of PLP phosphatase was measured at pH 7.4 in 40 mM
triethanolamine-HCl. The rate of production of pyridoxal from PLP was measured by following
the decrease in absorbance at 390 nm for at least 3min. The initial velocity data were fitted by a
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

least-squares method to the Lineweaver-Burk transformation of the equation below:
][]max[ SKmSV
Where [S] represents the concentration of the varied substrate PLP, and Km represents the
Michaelis constant.
A molecular weight of 64,000 for PLP phosphatase with a specific activity of 1.4 molmin-
1mg-1 was used in the calculations of molar enzyme concentrations. One unit of specific activity
is defined as the amount of protein that catalyzes the formation of pyridoxal/min from PLP. PLP
is known to have an extinction coefficient of 4900 cm-1M-1 at pH 7. Protein concentrations were
determined by the Bradford assay using the reagent and micro procedure from Pierce Chemical
Co. Bovine serum albumin was used as a standard.
In addition, hydrolysis of all other substrates was measured in 40 mM triethanolamine-HCl,
pH 7.4, containing 4 mM MgCl2. The release of free phosphate from potential substrates was
measured colorimetrically as the molybdate complex with malachite green (16,24).
Multiple tissue expression array.
A Northern filter containing 8 human tissue-specific poly(A)+ RNAs and a dot blot array
containing human poly(A)+ RNAs from various adult tissues, fetal tissues, and cancer cell lines
were prehybridized at 65°C for 1h in ExpressHybTM Hybridization solution (Clontech, Palo Alto,
CA). The filters were then hybridized at 65°C for 16 h with 32P-labeled cDNA probe containing
the complete open reading frame. After washing as recommended by the manufacturer, blots
were exposed to x-ray films at -70°C with an intensifying screen for the appropriate time period.
For scanning densitometry, the blot was scanned and BioLab Image software was used to
quantify the signals. Blots were reprobed with -actin as a loading control.
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

RESULTS
Identification and Cloning of the PLP Phosphatase cDNA from human and mouse.
Using two tryptic peptide sequences from the erythrocytes PLP phosphatase previously
reported by Gao and Fonda (21), we conducted NCBI BLAST searches of the human EST
database. A cDNA clone encoding both peptide sequences, designated a hypothetical protein
dj37E 16.5 from lung carcinoma (GenBankTM/EBI accession number NM020315) was identified.
When the tryptic peptide sequences were aliened with the deduced amino acid sequence, there
was a perfect match, confirming that the cDNA in the EST GenBank database codes for PLP
phosphatase.
We used the sequence information for designing specific primers for PCR amplification
utilizing a human brain cDNA library. Sequencing of this clone revealed that the brain cDNA is
identical to the hypothetical protein from lung carcinoma. The open reading frame (ORF)
encodes a 296-amino acid protein with a molecular mass of 31698 Da. A computer calculation
reveals that the isoelectric point for the protein is 6.12. ScanProsite software analysis by ExPASy
showed that the deduced human protein has the following putative post-translational
modification sites: 2 N-glycosylation sites, 7 phosphorylation sites, and 4 N-myristoylation sites.
In addition, we cloned a complete mouse homologue by 5’-RACE and 3’-RACE-PCR using
an EST sequence (GenBankTM/EBI accession number AK043228). As shown in Fig. 1, the
longest cDNA contains 1978 bp consisting of a 879-bp ORF, a 13-bp 5’-untranslated region and
a 1086-bp 3’-noncoding region. The 3’-end of the sequence contains a poly (A) stretch, preceded
by a putative polyadenylation signal AATAAA. The open reading frame encodes a 292-amino
acid protein with a predicted molecular mass of 31512 Da and pI of 5.53. The deduced protein
has a putative N-glycosylation site, 5 phosphorylation sites, and 4 amidation sites.
Using BLAST searches of the EST databases, we found putative PLP phosphatases in
various species. Multiple alignment of the protein sequence of PLP phosphatase with other
homologous proteins or putative proteins was performed using the ClustalW program as shown
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

in Fig. 2. All of the PLP phosphatases are highly homologous at the amino acid sequence level.
The mouse PLP phosphatase sequence shows a high degree of similarity (94% identity and 98%
similarity) with the human enzyme. The sequence identities of human PLP phosphatase to the
hypothetical proteins from D. Rerio, D. melanogaster, C. elegans, A. thaliana, S. erevisiae, E.
Coli are 36%, 28%. 26%, 25%, 31% and 27%, respectively, and similarities share 77%, 65%,
68%, 60%, 70% and 66%, respectively. This result suggests that PLP phosphatase is highly
homologous and widely expressed throughout evolution. The tryptic sequences from human
erythrocytes enzyme that are perfectly matched with the deduced amino sequence are indicated
by asterisks (*) in Fig.2.
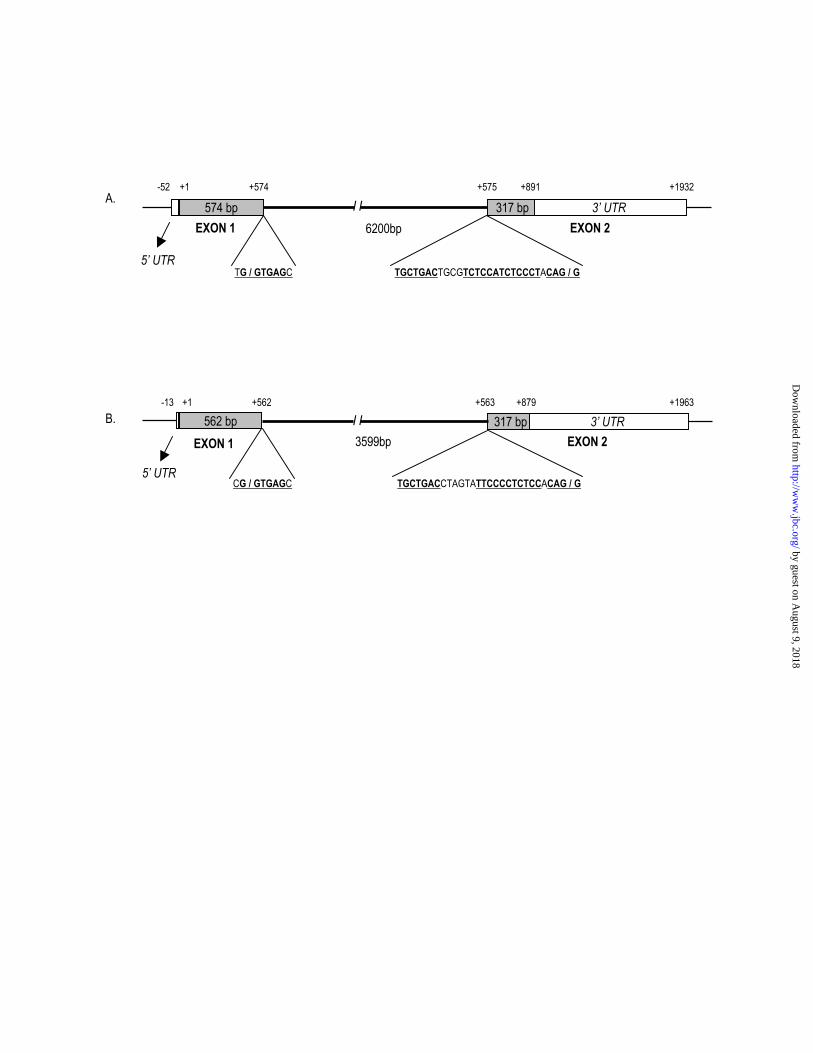
Genomic Organization and Chromosomal Localization
Using PLPP cDNA as a query sequence, a BLAST analysis of the genomic sequence
available through NCBI Web site mapped the PLP phosphatase gene to human chromosome
22q12.3. Mouse PLP phosphatase gene is located on chromosome 15.E1 with a very similar
genomic organization to that observed in human. The ORF of PLP phosphatase is encoded by
two exons as shown in Fig.3. The intron/exon junctions followed the GT/AG rule (25) and were
flanked by conserved sequences. The lariate branch-point in the consensus sequence is located at
23 nucleotides upstream of the 3’ splice site. The possible exon/intron junction site in mouse
cDNA is also indicated by a slash in Fig.1.
Functional expression and Catalytic Properties of the human recombinant enzyme
After E. coli M15/pQE30-hPLPP was induced with isopropyl -D-thiogalactopyranoside,
the 6x His-tagged recombinant PLP phosphatase yielded a major band with an apparent
molecular mass of 32 kDa on an SDS-PAGE gel corresponding to about 15 % of the total applied
protein (Fig. 4). Most of the expressed protein was found in the soluble extract after lysis of the
cells. As shown in Table I, the crude extraction from 200ml of cells contained approximately
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

14.5 mg of total protein and 0.14 units/mg of PLP phosphatase activity. Purification by binding
to Ni2+-nitrilotriacetic acid-agarose resin and elution with buffer containing 200mM imidazole
resulted in a near homogeneous preparation. The purified recombinant protein had a specific
activity of 1.4 units/mg, and appeared as a single band with a molecular mass of 32 kDa agreeing
with the value calculated from the deduced sequence (Fig. 4, lane 6).
Activity of the human PLP phosphatase toward PLP, PNP and PMP was determined (Table
II). Recombinant human enzyme followed Michaelis-Menten kinetics with respect to PLP, and
the Michaelis constant and kcat value for this reactant was 2.5 M and 1.52 s-1, respectively. Km
for PNP and PMP were 43 and 81 M, respectively, and kcat values were 1.25 and 0.45 s-1,
respectively, whereas activity with p-nitrophenyl phosphate was very low. Thus, the specificity
constant of the recombinant PLP phosphatase was highest with PLP followed by PNP. The
recombinant enzyme requires a divalent ion for activity, and Mg+2 is the most effective in
catalyzing the dephosphorylation of PLP.
To investigate the state of aggregation of recombinant human PLP phosphatase, the purified
enzyme was examined by FPLC gel filtration at pH 7.4 in 50 mM triethanolamine-HCl buffer.
The elution profile of the enzyme exhibits a symmetrical peak characterized by a molecular mass
of about 60 kDa when the concentration of the recombinant protein in the fraction is around 10
M (data not shown). These data clearly indicate that the recombinant enzyme is a dimer,
agreeing with previous data for the erythrocyte enzyme.
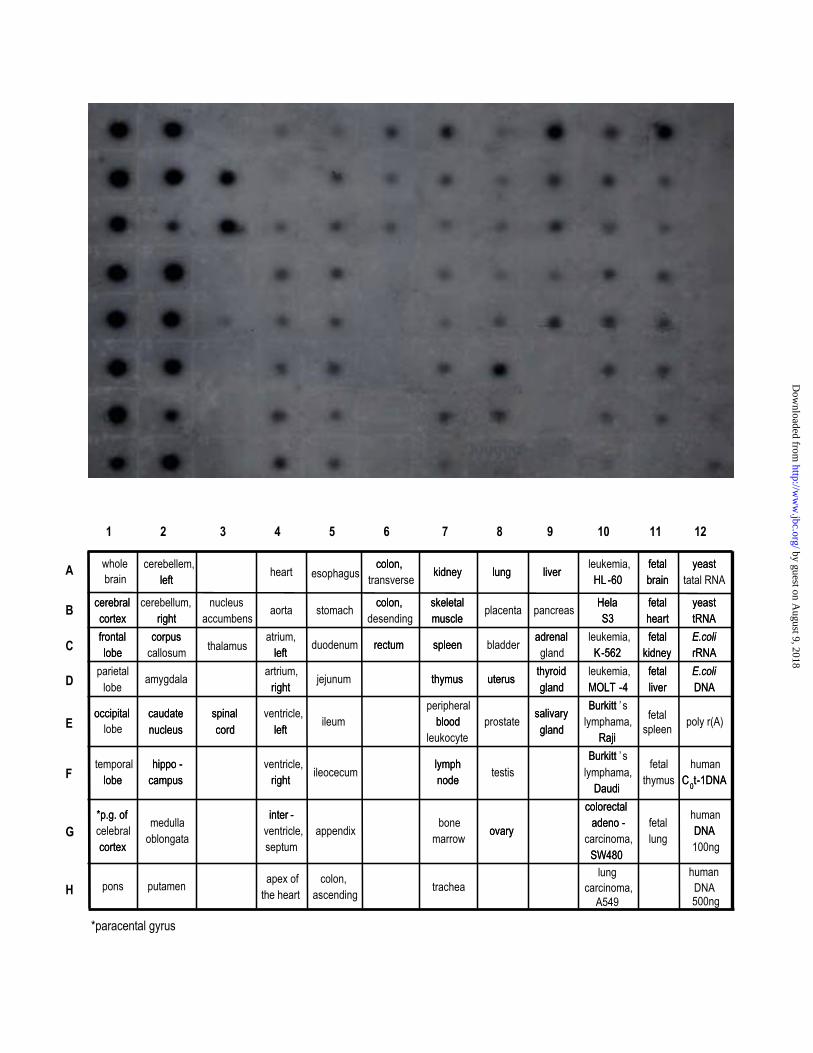
Tissue-specific Expression of human PLP phosphatase.
Dot blot array and Northern blot analyses were used to determine the expression pattern for
PLP phosphatase in human tissues. As evident by multiple tissue expression (MTETM) array
analysis, PLP phosphatase transcripts were detected in most tissues, indicating ubiquitous
expression. However, the level of expression was markedly variable. PLP phosphatase was most
highly represented in all the regions of central nerve system except the spinal cord (Fig. 5). The
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

other major sites of expression were found in liver and testis. In fetus, expression levels of PLP
phosphatase transcript showed a rather even distribution in all organs except brain. Like adult,
fetal brain expressed PLP phosphatase transcript with a very high level. To determine the size of
mRNA transcripts, Northern blots analyses were performed. A single 2.1 kb transcript was
detected with variable intensity in all adult tissues examined: heart, brain, placenta, lung, liver,
skeletal muscle, kidney and pancreas (data not shown).
DISCUSSION
In this report, we describe the first identification and characterization of cDNAs encoding a
PLP phosphatase from human and mouse. A human cDNA clone of PLP phosphatase was
identified by NCBI BLAST search on the basis of the amino acid sequences of two peptides
from the purified human erythrocytes enzyme. Several lines of evidence confirm that the cDNA
is indeed encoding the PLP phosphatase. First, the amino acid sequences of the two peptides of
the purified protein are contained in the amino acid sequence deduced from the ORF (Fig. 2).
Second, the molecular mass of the protein of 32 kDa calculated from the deduced cDNA
sequence is identical with the values from SDS-polyacrylamide gel electrophoresis for the
purified erythrocyte protein and the recombinant protein. Third, the kinetic parameters of the
recombinant protein are similar to those of purified enzyme. The Km values for the substrate PLP
(2.5 M) are practically indistinguishable from those of the erythrocyte (1.5 M) (16).
Human PLP phosphatase cDNA encodes a 296-amino acid protein with a predicted
molecular mass of 31698 Da. The hydropathy profile of PLP phosphatase indicated a rather even
distribution of hydrophobic and hydrophilic regions throughout the molecule (data not shown).
Therefore, no evidence for integral membrane domains in PLP phosphatase was found. The
isolated cDNA contains a short 5’-untranslated segment similar to lung carcinoma cDNA. No
larger product has been generated in repeated RACE experiments using different primer sets.
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Furthermore, the 2.1kb message determined by Northern analysis agrees well with the size of the
full-length cDNA, suggesting that the 5’-end of the cDNA is at or near the transcription start site.
Therefore, the cloned human brain cDNA may be complete. By contrast, analysis the EST clone
for mouse PLP phosphatase shows that it was incomplete. The mouse EST sequence
(GenBankTM/EBI accession number AK043228) contains several unidentified nucleotides and a
stop codon in ORF, but no polyadenylate tail. Thus, we cloned a full-length cDNA of mouse PLP
phosphatase by 5’- and 3’-RACE-PCRs, and the sequence has been deposited in
GenBankTM/EBI as accession number AY366300.
Comparison of the identified cDNA sequences with the genome sequences revealed the
genomic structure and chromosomal localization. The PLP phosphatase gene is divided into two
exons and the splice site sequences are remarkably well conserved as shown in Fig. 3. In
mammals two ends of the intron are conserved by a donor-site consensus sequence
AG/GURAGU and an acceptor-site consensus sequence (Y)nNCAG/G, where R, Y and N
indicate purine, pyrimidine and any nucleotide, respectively (25, 26). A branch-point consensus
sequence, YNCUGAC, is usually about 15-40 nucleotides upstream of the 3’ splice site, and the
adenosine in the sequence forms the 2’-5’ linkage in the lariat structure. In addition, the genomic
sequences were examined for the presence of CpG islands using the CpG plot program in
European Bioinformatics Institute (EBI), as defined by Gardiner-Garden and Frommer (27). The
hPLPP gene contains a CpG island with a CGobs/CGexp ratio in excess of 0.8 and a GC content of
74% spanning a region from 343bp upstream to 614bp downstream of the start codon, while
mPLPP gene has a CpG island extending -282 bp to +643bp with a CG content of 71 %.
We also reported here the functional expression of human PLP phosphatase. The expressed
protein was purified by a one-step affinity chromatographic method, and consequently
endogenous PLP phosphatase produced by E. coli was eliminated. Enzymological analysis of the
recombinant human protein expressed from this cDNA clone showed a high specificity for the
hydrolysis of PLP. Characteristics of the recombinant enzyme were similar to those from the
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

native enzyme purified from human erythrocytes, except kinetic values for PNP (16).
Noteworthy is the relatively high Vmax and Km with PNP as a substrate. As shown in Table II,
the Km values of recombinant enzyme for PNP and PLP were 43 M and 2.5 M, respectively,
whereas the Vmax values for PNP were surprisingly high as 1.2 units/mg, which is almost same
to the value for PLP (1.4 units/mg). Judging from the kinetic results, the catalytic efficiency
(kcat/Km) for PNP was 20-fold lower than that of PLP. On the other hand, the kinetic values for
the native enzyme were somewhat different (16). The enzyme had relatively low Km and Vmax
values with PNP. Km values with respect to PNP and PLP were 5.2 M and 1.5 M, respectively,
while the Vmax were 3.2 units/mg and 0.7 units/mg, respectively. In consequence, catalytic
efficiency value for PNP was 17 fold low than that of PLP, which is practically indistinguishable
from that of the recombinant enzyme.
An interesting aspect of our work is the finding that PLP phosphatase is differentially
expressed in a tissue-specific manner. Based on the levels of mRNA expression, the widespread
distribution of PLP phosphatase in human tissues is consistent with its essential role in cellular
metabolism. In addition to its housekeeping role in PLP metabolism, however, PLP phosphatase
may play a more specific role especially in brain, where its level is substantially higher. In this
connection, it is should be noted that pyridoxal kinase is expressed in essentially all mammalian
organs (4), whereas PNP oxidase is selectively present with the highest activity found in liver
and kidney (28).
In conclusion, the results obtained in this study will provide valuable information on the
action mechanism of human PLP phosphatase and the physiological role of this enzyme. The
availability of the cDNAs encoding PLP phosphatase and the recombinant enzyme will be
helpful to expand our view of metabolic pathways that could involve PLP phosphatase and to
evaluate its functional role in human cellular homeostasis.
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

REFERENCES
1. Snell, E. E. (1990) Ann. N. Y. Acad. Sci. 585, 1-12
2. Meister, A. (1990) Ann. N. Y. Acad. Sci. 585, 13-31
3. McCormick, D. B., Gregory, M. E. and Snell, E. E. (1961) J. Biol. Chem. 236, 2076-2084
4. Hanna, M. C., Turner, A. J. and Kirkness, E. F. (1997) J. Biol. Chem. 272, 10756-10760
5. Kwok, F. and Churchich, J. E. (1992) in Chemistry and Biochemistry of Flavoenzymes
(Muller, F. ed) Vol. 3, pp. 1-20, CRC Press, London
6. Ngo, E. O., LePage, G. R., Thanassi, J. W., Meisler, N. and Netter, L. M. (1998) Biochemistry,
37, 7741-7748
7. . Lumeng, L. and Li, T. K. (1980) in Vitamin V6 metabolism and Role in Growth (Tryfiates, G.
P. ed) pp. 27-51, Food and nutrition Press, CT
8. Schwarts, R. and Kjelgaard, N.O. (1951) Biochem. J. 48, 333-337
9. Stanulovic, M., Jeremic, V., Leskovas, V. and Chaykin, S. (1976) Enzyme 21, 357-369
10. Snell, E. E., and Haskell, B. E. (1971) Comprehensive Biochemistry (Florkin, M., and Stotz,
E. H., eds) Vol. 21, pp. 47-71, Elsevier Scientific Publishing Co., Amsterdam
11. Anderson, B. B., Newmark, P. A. and Rawlins, M. (1974) Nature 250, 502-504
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

12. McCormick, D. B. and Merrill, A. H. (1980) in Vitamin V6 metabolism and Role in Growth
(Tryfiates, G. P. ed) pp. 1-26, Food and nutrition Press, CT
13. Kwon, O-S., Kwok, F. and Churchich, J. E. (1991) J. Biol. Chem. 266, 22136-22140
14. Li, T. K., Lumeng, L. and Veitch, R. L. (1974) Biochem. Biophys. Res. Commun. 61, 677-684
15. Lumeng, L. and Li, T. K. (1975) J. Biol. Chem. 250, 8126-8131
16. Fonda, M. L. (1992) J. Biol. Chem. 267, 15978-15983
17. Harris, H. (1990) Clin. Chim. Acta 186, 133-150
18. Bull, H., Murray, P.G., Thomas, A.M. and Nelson, P.N. (2002) Mol. Pathol. 55, 65-72
19. Fonda, M. L. and Zhang, Y. (1995) Arch. Biochem. Biophys. 320, 345-352
20. Gao, G. and Fonda, M. L. (1994) J. Biol. Chem. 269, 7163-7168
21. Gao, G. and Fonda, M. L. (1994) J. Biol. Chem. 269, 8234-8239
22. Gao, G. and Fonda, M. L. (1994) Arch. Biochem. Biophys. 313, 166-172
23. Thompson, J. D., Hinggins, D. G., and Gibson, T. J. (1994) Nucleic Acids Res. 22, 4673-4680
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

24. Rose, Z. B. (1981) Arch. Biochem. Biophys. 208, 602-609
25. Breathnach, R. and Chambon, P. (1981) Annu. Rev. Biochem. 50, 349-383
26. Padgett, R. A., Grabowski, P. J., Konarska, M. M., Seiler, S., and Sharp, P. A. (1986) Annu.
Rev. Biochem. 55, 1119-1150
27. Gardiner-Garden, M. and Frommer, M. (1987) J. Mol. Biol. 196, 261-282
28. Buss, D. D., Hamm, M. W., Mehansho, H. and Henderson, L. M. (1980) J. Nutr. 110, 1655-
1663
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

FIGURE LEGENDS
Fig. 1. Nucleotide and deduced amino acid sequence of a full-length cDNA encoding mouse
PLP phosphatase (GenBank accession No. AY366300). The predicted amino acid sequence of
PLP phosphatase is shown below the nucleotide sequence. The termination codon is marked by
an asterisk (*) and the polyadenylation signal is double underlined. Putative glycosylation sites
(g), phosphorylation sites (p) and N-myristoylation sites (m) in mouse PLP phosphatase are
indicated. A potential exon/intron junction also indicated by a slash. The numbers on the right
refer to nucleotide and amino acid positions.
Fig. 2. Sequence alignment of human and mouse PLP phosphatase with homologous
proteins. Full-length Homo sapiens hPLPP and Mus musculus mPLPP sequences were aligned
with those of homologous proteins or putative proteins using the Clustal W program (version
1.74). EST sequences (GenBankTM accession number in brackets) are from Danio Rerio
(AAH45860), Drosophila melanogaster (AAF49296), Caenorhabditis elegans (P19881),
Arabidopsis thaliana (BAA98057), S. cerevisiae (NP504509), E. Coli (AAC09327). The
introduced gaps are shown as hyphens, and aligned amino acids are boxed (black for identical
residues and dark gray for similar residues). The tryptic fragment sequences determined from
human erythrocytes enzyme are indicated by asterisk (*).
Fig. 3. Genomic organization of human and mouse PLP phosphatase genes. Schematic
representation of human (A) and mouse PLP phosphatase genes (B) arranged as two exons. Exon
regions are denoted by boxes. Shadowed boxes represent ORF, and open boxes denote the 5’- and
3’-untranslated regions. Black line indicate the introns. The consensus intron/exon junction
sequences are underlined and the intron/exon borders are marked by slashes.
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Fig. 4. Expression and purification of recombinant human PLP phosphatase. Twelve
percent of SDS-PAGE analysis of crude cell extracts of E. coli M15 containing the expression
vector pQE30 without and with the coding sequence. Lane 1, low molecular mass standards
(Bio-Rad Laboratory); lane 2 and lane 3, crude extracts from cultured cells harboring pQE30
uninduced and induced with 1mM IPTG, respectively; lane 4 and 5, cells containing pQE30-
hPLPP in the absence and presence of IPTG, respectively; lane 6, purified recombinant PLP
phosphatase from Ni resin (7 g).
Fig. 5. Tissue-specific expression of the human PLP phosphatase mRNA. The human
multiple tissue expression (MTETM) array was hybridized with a 32P-labeled human PLP
phosphatase -specific cDNA probe. Tissue sources for the RNA are indicated below the blot.
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

GCCAGCCGGCTGC 13
ATGGCGCGCTGCGAGCGGCTGCGCGGCGCGGCCCTGCGCGACGTGCTGGGCCAGGCGCAGGGAGTCCTGTTCGACTGC 91M A R C E R L R G A A L R D V L G Q A Q G V L F D C 26
mGACGGAGTGCTGTGGAACGGCGAACGCATCGTGCCGGGCGCCCCTGAGCTGCTGCAGCGGCTGGCGCGGGCCGGCAAG 169D G V L W N G E R I V P G A P E L L Q R L A R A G K 52
mAACACGCTGTTCGTGAGCAACAACAGCCGGCGCGCGCGGCCCGAGCTGGCGCTGCGCTTCGCCCGCCTGGGCTTCGCG 247N T L F V S N N S R R A R P E L A L R F A R L G F A 78
g pGGGCTGCGCGCCGAGCAGCTCTTCAGCTCCGCGCTGTGTGCCGCGCGCCTCCTGCGCCAGCGGCTGTCCGGGCCGCCC 325G L R A E Q L F S S A L C A A R L L R Q R L S G P P 104
GACGCATCGGGCGCAGTGTTCGTGCTGGGAGGCGAGGGGCTGCGCGCCGAGCTGCGCGCCGCGGGGCTGCGCCTGGCG 403D A S G A V F V L G G E G L R A E L R A A G L R L A 130
mGGGGACCCCGGCGAGGACCCGCGCGTGCGCGCCGTGCTCGTAGGCTACGACGAGCAGTTTTCCTTCTCCAGGTTGACC 481G D P G E D P R V R A V L V G Y D E Q F S F S R L T 156
GAGGCGTGCGCGCACCTGCGCGACCCCGACTGCCTGCTCGTGGCCACCGATCGCGACCCTTGGCACCCGCTCAGCGAC 559E A C A H L R D P D C L L V A T D R D P W H P L S D 182
pGGAAGCCGGACCCCCG/GTACTGGAAGCCTGGCTGCCGCGGTGGAGACAGCCTCGGGACGCCAGGCCCTGGTGGTGGGC 637G S R T P /G T G S L A A A V E T A S G R Q A L V V G 208
m pAAGCCCAGCCCTTACATGTTCCAGTGCATCACAGAAGACTTCAGCGTGGACCCCGCCCGCACGCTGATGGTGGGGGAC 715K P S P Y M F Q C I T E D F S V D P A R T L M V G D 234
CGCCTGGAGACCGACATACTCTTTGGCCACCGCTGCGGCATGACCACCGTGCTCACGCTCACGGGCGTCTCCAGCCTC 793R L E T D I L F G H R C G M T T V L T L T G V S S L 260
pGAAGAAGCCCAGGCCTACCTGACGGCTGGCCAGCGTGACCTTGTGCCTCACTACTATGTGGAGAGCATTGCGGACTTG 871E E A Q A Y L T A G Q R D L V P H Y Y V E S I A D L 286
pATGGAGGGGCTGGAGGACTGAGCCCACCTCCCCTGTTGTTCCCTGATCCCACAGATGGAGGCGACGGGCCATGAGTCA 949M E G L E D * 292
CATTCGGGACCATGAGCTTTCTGGAGTGGGTGTCTGTACCCCAGTGAAGGGAGGCTGGGATCCAGGAAGGTTTGTGGG 1027
CTTTCACACCCCCCTCCTAGCCGTGGCTGGACACTCTGCTCTTCAGAAGCTGTCCCCCCACATCTTGAAGGTTCACCC 1105
TGGCCTGACCCCACCAGGTGGCCTTATTTCCTGCCCTGTCACCTCCCTTCCTTGGGCCCCGAAGCCAAATGAAGATTT 1183
TTTTTTTTCCAACCTTTTAAGTTCCTCCCACTCTCTGGGATCTCTAGTGCTCTGTCCCTAGAGATTGTTGGCCAATCA 1261
GAGATCTAGGGAACTCCAGCCCGTTCCTGGCCTGACCAGGCCTCTGGGAAGAGCTGCTGGCACTGCCGGCCTATGTCT 1339
CAGAGTGGGCCTTTAGTGGAGCAAGTAGGAGCTTAAAGAATGCCTTGCATATCCTGGAACCCCAGAGCACCAGTCAGA 1417
GGCTATGCGTCCATCAGGACAATTAACGATCCCACTGGCCTCTTAGATGCCAATTAGAACAATTTCCAAGTCCAAGAG 1495
ACTTGTGAACTTTTGACAATCCTAGCCTGACTGGATCCTGGGACCAGATGCTGGTAGCACAGGGTGATGAATGCCGGT 1573
ATGTATAGCACTCGGGAGGTTGAAGCAAGAGGATTGCTGAGTCTGGTGACCGGGATGGCAGAGTCAGACCTCATCTCA 1651
GACAGAGGGTCATCCTTCAAGTCACCCCACTCCACAGACTGCTCAGTGGGGCACCTCTGTGGCCCTGGGGTAGTGTGG 1729
AGAGGTTCCCACTCATACCTTAGCACCGTCCAGTGTTGATGGCTTTTTCGGCTTGGAGGGGGCCCTCCTTTTGTCCCC 1807
ACCTGCAGTCAGTGTTTGCAGTGCCACCCTCACCTGCCCATTCCCAGCTATTTATTTATTTATTGTGTGCCAGTGAAG 1885
GTGGGGGTAGGGACCTTCCCACCCCCTCCACCTGTTGTAACCGGTCCTTGAACTTAATAAAATGTGAATTGAAGTGTC 1963
AAAAAAAAAAAAAAA 1978
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

30 60 90H.sapiens ------------------------------ ----MAR-----CE--------RLRGAALR -------------DVLGRAQGVLFDCDGVL 30M.musculus ------------------------------ ----MAR-----CE--------RLRGAALR -------------DVLGQAQGVLFDCDGVL 30D.rerio ------------------------------ --MAASK-----CV--------RLNGALSR -------------QLLDSVDCVLFDCDGVI 32D.melanogaster -----------------MSCNFARFSRQAA INMYKQS-----CTNLL-----ELSSAKVT -------------EWLAGFDSVITDCDGVL 50C.elegans ---------------------------MCI EAGLDPK-----CRSTK-----PLCPDTFA -------------KVMKTIDTFIFDADGVL 40A.thaliana MLSRSVASAVTPVSSSSLLPNSKPIFCLKT LSGYRSSSFCGGCIRKINHKPLRMTSSNIT PRAMATQQLENADQLIDSVETFIFDCDGVI 90S.cerevisiae ------------------------------ --MTAQQG----VP-------IKITNKEIA Q------------EFLDKYDTFLFDCDGVL 35E.coli ------------------------------ ------------------------------ ----------------MTIKNVICDIDGVL 14
************ 120 150 180 H.sapiens WNGERAVPGAPELLERLARAGKAALFVSNN SRRARPELALRFARLGFGG--LRAEQLFSS ALCAARLLRQRLPGPPDAPG—AVFVLGGE 116M.musculus WNGERIVPGAPELLQRLARAGKNTLFVSNN SRRARPELALRFARLGFAG--LRAEQLFSS ALCAARLLRQRLSGPPDASG—AVFVLGGE 116D.rerio WRGDQAIPGAPEVINSLKKHGKQVFFVTNN STKTRQMYADKLGKLGFDA--A-ADEVFGT AYCSAQYLKN----VCKLDG—KVYLIGSK 113 D.melanogaster WIYGQALEGSVDVMNQLKGMGKSIYFCTNN STKTRSELLKKGVELGFHI--K-ENGIIST AHATAAYLKRR-----NFSK—RVFVIGSE 130C.elegans WLGESVMPGSPRLIDYLVKHNKQIIVLTNN ATKSRAVYAKKLAKLGYNSSKMNKNNLVNP AAVVADTLHR-----AGLDGK-RVYLIGEQ 124A.thaliana WKGDKLIEGVPETLDMLRAKGKRLVFVTNN STKSRKQYGKKFETLGLN---VNEEEIFAS SFAAAAYLQS-----INFPKDKKVYVIGEE 172S.cerevisiae WLGSQALPYTLEILDLLKQLGKQLIFVTNN STKSRLAYTKKFASFGID---VKEEQIFTS GYAPAVYIRD---FLKLQPGKDKVWVFGES 119 E.coli MHDNVAVPGAAEFLHGIMDKGLPLVLLTNY PSQTGQDLANRFATAGVD---VPDSVFYTS AMATADFLRR-------QEGK-KAYVVGEG 93
210 240 270H.sapiens GLRAELRAAGLRLAGD-PS----AGDG--A ------------APRVRAVLVGYDEHFSFA KLREACAHLRD-PECLLVATDRDPWHPLSD 186 M.musculus GLRAELRAAGLRLAGD-P--------G--E ------------DPRVRAVLVGYDEQFSFS RLTEACAHLRD-PDCLLVATDRDPWHPLSD 182 D.rerio AMKQELEEVGIQPVGVGPD----LISG--V QIDWAN---VPLDQEVQAVLVGFDEHFSYM KLNRALQYLCD-PDCQFVGTNTDTRLPLEG 193 D.melanogaster GITKELDAVGIQHTEVGPE----PMKGSLA EFMAQH---LKLDTDIGAVVVGFDEHFSFP KMMKAASYLND-PECLFVATNTDERFPMPN 212 C.elegans GLRDEMDELGIEYFGHGPEKKQDEADG--S GAFMYD---IKLEENVGAVVVGYEKHFDYV KMMKASNYLRE-EGVLFVATNEDETCPGPN 208 A.thaliana GILKELELAGFQYLGG-PD------DGK-R QIELKPGFLMEHDHDVGAVVVGFDRYFNYY KIQYGTLCIRENPGCLFIATNRDAVTHLTD 254 S.cerevisiae GIGEELKLMGYESLGGADSRLDTPFDAAKS PFLVNG-----LDKDVSCVIAGLDTKVNYH RLAVTLQYLQK-DSVHFVGTNVDSTFPQKG 203 E.coli ALIHELYKAGFTITDVNPD----------- -----------------FVIVGETRSYNWD MMHKAAYFVAN--GARFIATNPDTHGRG-- 151 ************* *
300 330 360 H.sapiens G-SRTPGTGSLAAAVETASGRQALVVGKPS PYMFECITENFSIDPARTLMVGDRLETDIL FGHRCGMT-TVLTLTGVSRLEEAQAYLAAG 274 M.musculus G-SRTPGTGSLAAAVETASGRQALVVGKPS PYMFQCITEDFSVDPARTLMVGDRLETDIL FGHRCGMT-TVLTLTGVSSLEEAQAYLTAG 270 D.rerio G-KAVPGTGCLLRAVETAAQRQAQVVGKPS NFMFECVASQFNLKPQRCLMVGDRLDTDIM LGSNCGLK-TLLTLTGVSTVAEAEANQKSE 281 D.melanogaster --MIVPGSGSFVRAIQTCAERDPVVIGKPN PAICESLVTEKKIDPSRTLMIGDRANTDIL LGFNCGFQ-TLLVGSGIHQLKDVERWKLSQ 299 C.elegans PEVVIPDAGPIVAAIKCASGRDPLTVGKPC TPAFNYIKRKWNINPSRTMMIGDRTNTDVK FGRDHGMK-TLLVLSGCHQIEDIIENQMNE 297 A.thaliana A-QEWAGGGSMVGALVGSTQREPLVVGKPS TFMMDYLADKFGIQKSQICMVGDRLDTDIL FGQNGGCK-TLLVLSGITNLQHFIHFVFVD 342 S.cerevisiae --YTFPGAGSMIESLAFSSNRRPSYCGKPN QNMLNSIISAFNLDRSKCCMVGDRLNTDMK FGVEGGLGGTLLVLSGIETEERTLKI--S- 288 E.coli ---FYPACGALCAGIEKISGRKPFYVGKPS PWIIRAALNKMQAHSEETVIVGDNLRTDIL AGFQAGLE-TILVLSGVSSLDDIDSMP--- 234
390H.sapiens ----QHDLVPHYYVESIADL-TEGLED--- ----------------- 296 M.musculus ----QRDLVPHYYVESIADL-MEGLED--- ----------------- 292 D.rerio C-PQKQRMVPDYYIDSIADI-LPALQA--- ----------------- 306 D.melanogaster D-PEEKKLIPDVYLPKLGDL-LPSIVNSLT DLK-------------- 330 C.elegans ----RDDMVPDYVAPCLGAL-VPESNQIYY ----------------- 322 A.thaliana LKVLLQSLCWKALRTRYNQISTPARSPIFC LRKPQLYNLVSPASITL 389 S.cerevisiae ----HDYPRPKFYIDKLGDIYALTNNEL-- ----------------- 312 E.coli -------FRPSWIYPSVAEIDVI------- ----------------- 250
by guest on August 9, 2018 http://www.jbc.org/ Downloaded from
-52 +1 +574 +575 +891 +1932A.
5’ UTR
317 bp574 bp6200bpEXON 1 EXON 2
/ /
TG / GTGAGC TGCTGACTGCGTCTCCATCTCCCTACAG / G
3’ UTR
+1963
317 bp/ /3599bp
+563 +879
562 bp
5’ UTR
EXON 2EXON 1
CG / GTGAGC TGCTGACCTAGTATTCCCCTCTCCACAG / G
-13 +1 +562
B. 3’ UTR
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1 2 3 4 5 6
97.4
66.245.0
31.0
21.5
14.4
PLP Phosphatase( 33kDa )
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from
fetallung
fetalthymus
fetalliver
fetalkidney
fetalheart
fetalbrain
humanDNA500ng
lungcarcinoma,
A549
colon,ascending
apex ofthe heart
putamenpons
humanDNA100ng
colorectaladeno -
carcinoma,SW480
ovarymarrow
appendixinter -
ventricle,septum
*p.g. ofcelebralcortex
humanC0t-1DNA
Burkitt slymphama,
Dauditestis
lymphnode
ileocecumventricle,
righthippo -
campustemporal
lobe
poly r(A)Burkitt s
lymphama,Raji
salivarygland
prostateperipheral
bloodleukocyte
ileumventricle,
leftspinalcord
caudatenucleus
occipital
E.coliDNA
leukemia,MOLT -4
thyroidgland
uterusthymusjejunumartrium,
rightamygdala
parietallobe
E.colirRNA
leukemia,K-562
adrenalgland
bladderspleenrectumduodenumatrium,
leftthalamuscorpusfrontal
lobe
yeasttRNA
HelaS3
pancreasplacentaskeletalmuscle
colon,desending
stomachaortanucleus
accumbenscerebellum,
rightcerebralcortex
yeasttatal RNA
leukemia,HL -60
liverlungkidneycolon,
transverseheart
cerebellem,left
1
*paracental gyrus
spleen
fetalliver
fetalkidney
fetalheart
fetalbrain
trachea
DNA
colorectaladeno -
SW480
ovarybone
inter -medulla
oblongata
*p.g. of
cortex
C0t-1DNA
Burkitt ’
Daudi
lymphnoderight
hippo -campuslobe
Burkitt ’
Raji
salivarygland
bloodleft
spinalcord
caudatenucleus
occipital
E.coliDNAMOLT -4
thyroidgland
uterusthymusright
E.colirRNAK-562
adrenalspleenrectum
leftcorpus
callosumfrontallobe
yeasttRNA
HelaS3
skeletalmuscle
colon,right
cerebralcortex
yeastHL -60
liverlungkidneycolon,
left esophagus
fetallobe
wholebrain
2 3 4 5 6 7 8 9 10 11 12
B
C
D
A
E
F
G
H
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from
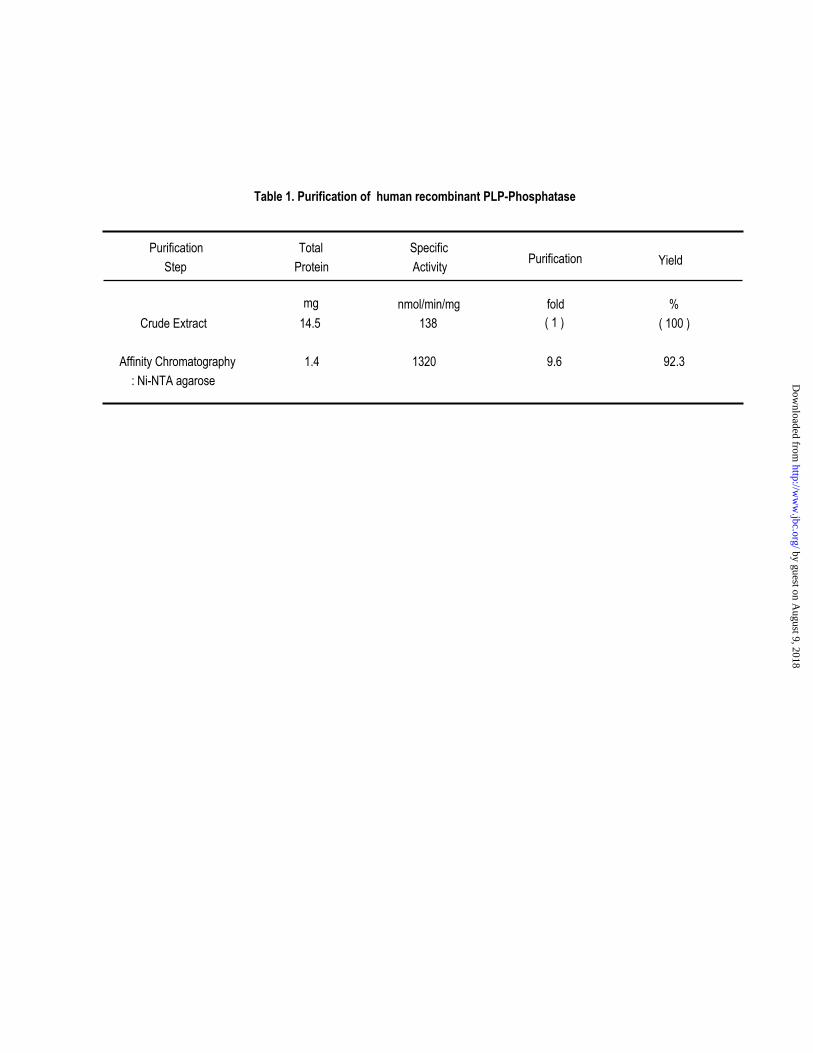
Table 1. Purification of human recombinant PLP-Phosphatase
nmol/min/mg fold %Crude Extract 14.5 138 ( 100 )
Affinity Chromatography 1.4 1320 9.6 92.3: Ni-NTA agarose
Purification Total SpecificStep Protein Activity
( 1 )mg
Purification Yield
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from
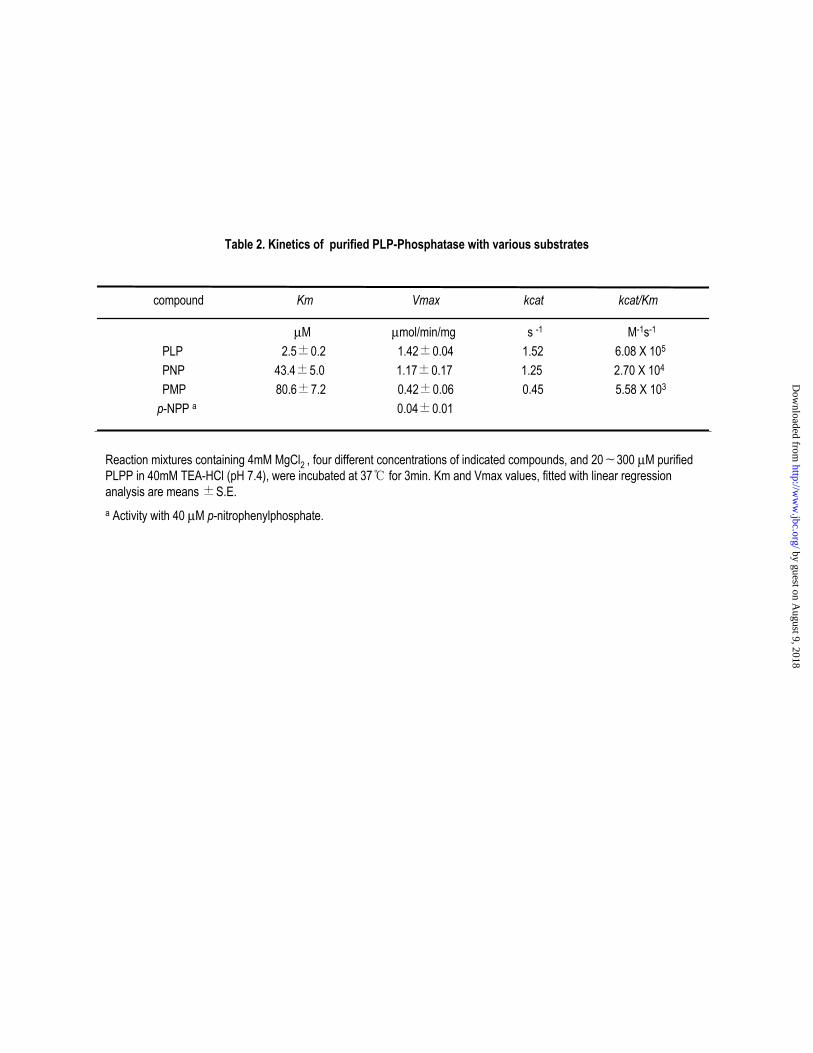
Table 2. Kinetics of purified PLP-Phosphatase with various substrates
compound Km Vmax kcat kcat/Km
µM µmol/min/mg s -1 M-1s-1
PLP 2.5±0.2 1.42±0.04 1.52 6.08 X 105
PNP 43.4±5.0 1.17±0.17 1.25 2.70 X 104
PMP 80.6±7.2 0.42±0.06 0.45 5.58 X 103
p-NPP a 0.04±0.01
Reaction mixtures containing 4mM MgCl2 , four different concentrations of indicated compounds, and 20∼300 µM purifiedPLPP in 40mM TEA-HCl (pH 7.4), were incubated at 37℃ for 3min. Km and Vmax values, fitted with linear regressionanalysis are means±S.E.a Activity with 40 µM p-nitrophenylphosphate.
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Moon, Soo Young Choi and Oh-Shin KwonYoung Min Jang, Dae Won Kim, Tae-Cheon Kang, Moo Ho Won, Nam-In Baek, Byung Jo
distributionHuman pyridoxal phosphatase: Molecular cloning, functional expression and tissue
published online September 30, 2003J. Biol. Chem.
10.1074/jbc.M309619200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on August 9, 2018
http://ww
w.jbc.org/
Dow
nloaded from


















