Molecular cloning and expression of four phenylalanine ...
10
DOI: 10.1007/s10535-016-0619-1 BIOLOGIA PLANTARUM 60 (3):459-468, 2016 459 Molecular cloning and expression of four phenylalanine ammonia lyase genes from banana interacting with Fusarium oxysporum Z. WANG 1 , J.-Y. LI 2 , C.-H. JIA 1 , J.-P. LI 1 , B.-Y. XU 1 *, and Z.-Q. JIN 1,2 Key Laboratory of Tropical Crop Biotechnology, Ministry of Agriculture, Institute of Tropical Bioscience and Biotechnology, Chinese Academy of Tropical Agricultural Sciences, Hainan 571101, P.R. China 1 Haikou Experimental Station, Chinese Academy of Tropical Agricultural Sciences, Hainan 570101, P.R. China 2 Abstract Phenylalanine ammonia lyase (PAL) is the first enzyme in the phenylpropanoid pathway. In this study, we describe the molecular characteristics of four PAL genes (MaPAL1, MaPAL2, MaPAL3, and MaPAL4) cloned from banana (Musa acuminata L. AAA group, cv. Cavendish) using a rapid amplification of cDNA ends and real time quantitative PCR. The predicted molecular masses of corresponding PALs ranged from 70.2 to 77.3 kDa and their isoelectric points were acidic. At the amino acid level, they shared a high sequence similarity with PALs in the banana DH-Pahang (AA group) genome. Phylogenetic analysis shows that the deduced amino acid sequences of MaPALs had also a high similarity with PALs of other plant species. Expression analysis by semi-quantitative reverse transcriptase PCR reveals that these genes were differentially expressed in various tissues. Enzyme activity of PAL and expression of MaPALs in bananas were induced after infection with Fusarium oxysporum f. sp cubense Tropical Race 4. Our findings suggest that MaPALs play important roles in banana resistance to F. oxysporum. Additional key words: disease resistance; gene expression; Musa acuminata; real time qPCR. Introduction Bananas are an important food staple for many people in tropical regions (Aurore et al. 2009). There are several devastating diseases that target the Musa crop (Robinson 1996) including Fusarium wilt, which is caused by the Fusarium oxysporum f. sp. cubense (Foc) (Stover 1962). Foc is divided into four races (Koenig et al. 1997), and Race 4 can be further divided into two types: tropical and subtropical (Visser et al. 2010). Tropical Race 4 (TR4) causes a severe destruction of the commercial banana cultivar Cavendish (Hwang and Ko 2004). The resistant cultivar Musa acuminata L. AAA group, cv. Nongke No. 1, was created through somaclonal variation (Hwang and Ko 2004, Li et al. 2012a). In higher plants, phenylalanine ammonia lyase (PAL, EC 4.3.1.5) catalyzes conversion of L-phenylalanine to trans-cinnamic acid, which is at the gateway of primary and secondary phenylpropanoid metabolism in plants (Hahlbrock and Scheel 1989). Many secondary metabolic products in plants, such as anthocyanins, lignins, hormones, flavonoids, and phytoalexins, are derived from phenylpropanoids (Dixon and Paiva 1995). A variety of environmental stresses, such as pathogenic attack, UV radiation, wounding, and excessive radiation, can induce accumulation of PAL mRNA (Jorrin and Dixon 1990, Leyva et al. 1995). In addition, PAL plays an important role in a plant defense system and is also involved in biosynthesis of a signaling molecule salicylic acid, which is required for plant systemic acquired resistance (Nugroho et al. 2002, Chaman et al. 2003). Phenylalanine ammonia lyase is known to be present in many higher plants, some fungi, yeasts, and prokaryotes. Many plant PAL genes have been cloned and sequenced recently in several plant species, including tomato (Chang et al. 2008), cucumber (Shang et al. 2012), Bambusa oldhamii Submitted 21 January 2015, last revision 21 October 2015, accepted 3 November 2015. Abbreviations: CTAB - cetyltrimethyl ammonium bromide; DPI - days post-inoculation; Foc TR4 - Fusarium oxysporum f. sp. cubense Tropical Race 4; GFP - green fluorescent protein; PAL - phenylalanine ammonia lyase; qPCR - quantitative PCR; RACE - rapid amplification of cDNA ends. Acknowledgments: This research was funded by the Ministry of Science and Technology of P.R. China (No. 2011AA10020605) and the Modern Agro-industry Technology Research System (nycytx-33). Z. Wang and J.-Y. Li contributed equally to this work. * Corresponding author; fax: (+86) 898 66890772, e-mail: [email protected]
Transcript of Molecular cloning and expression of four phenylalanine ...

DOI: 10.1007/s10535-016-0619-1 BIOLOGIA PLANTARUM 60 (3):459-468, 2016
459
Molecular cloning and expression of four phenylalanine ammonia lyase genes from banana interacting with Fusarium oxysporum Z. WANG1, J.-Y. LI2, C.-H. JIA1, J.-P. LI1, B.-Y. XU1*, and Z.-Q. JIN1,2
Key Laboratory of Tropical Crop Biotechnology, Ministry of Agriculture, Institute of Tropical Bioscience and Biotechnology, Chinese Academy of Tropical Agricultural Sciences, Hainan 571101, P.R. China1 Haikou Experimental Station, Chinese Academy of Tropical Agricultural Sciences, Hainan 570101, P.R. China2 Abstract Phenylalanine ammonia lyase (PAL) is the first enzyme in the phenylpropanoid pathway. In this study, we describe the molecular characteristics of four PAL genes (MaPAL1, MaPAL2, MaPAL3, and MaPAL4) cloned from banana (Musa acuminata L. AAA group, cv. Cavendish) using a rapid amplification of cDNA ends and real time quantitative PCR. The predicted molecular masses of corresponding PALs ranged from 70.2 to 77.3 kDa and their isoelectric points were acidic. At the amino acid level, they shared a high sequence similarity with PALs in the banana DH-Pahang (AA group) genome. Phylogenetic analysis shows that the deduced amino acid sequences of MaPALs had also a high similarity with PALs of other plant species. Expression analysis by semi-quantitative reverse transcriptase PCR reveals that these genes were differentially expressed in various tissues. Enzyme activity of PAL and expression of MaPALs in bananas were induced after infection with Fusarium oxysporum f. sp cubense Tropical Race 4. Our findings suggest that MaPALs play important roles in banana resistance to F. oxysporum.
Additional key words: disease resistance; gene expression; Musa acuminata; real time qPCR. Introduction Bananas are an important food staple for many people in tropical regions (Aurore et al. 2009). There are several devastating diseases that target the Musa crop (Robinson 1996) including Fusarium wilt, which is caused by the Fusarium oxysporum f. sp. cubense (Foc) (Stover 1962). Foc is divided into four races (Koenig et al. 1997), and Race 4 can be further divided into two types: tropical and subtropical (Visser et al. 2010). Tropical Race 4 (TR4) causes a severe destruction of the commercial banana cultivar Cavendish (Hwang and Ko 2004). The resistant cultivar Musa acuminata L. AAA group, cv. Nongke No. 1, was created through somaclonal variation (Hwang and Ko 2004, Li et al. 2012a). In higher plants, phenylalanine ammonia lyase (PAL, EC 4.3.1.5) catalyzes conversion of L-phenylalanine to trans-cinnamic acid, which is at the gateway of primary and secondary phenylpropanoid metabolism in plants
(Hahlbrock and Scheel 1989). Many secondary metabolic products in plants, such as anthocyanins, lignins, hormones, flavonoids, and phytoalexins, are derived from phenylpropanoids (Dixon and Paiva 1995). A variety of environmental stresses, such as pathogenic attack, UV radiation, wounding, and excessive radiation, can induce accumulation of PAL mRNA (Jorrin and Dixon 1990, Leyva et al. 1995). In addition, PAL plays an important role in a plant defense system and is also involved in biosynthesis of a signaling molecule salicylic acid, which is required for plant systemic acquired resistance (Nugroho et al. 2002, Chaman et al. 2003). Phenylalanine ammonia lyase is known to be present in many higher plants, some fungi, yeasts, and prokaryotes. Many plant PAL genes have been cloned and sequenced recently in several plant species, including tomato (Chang et al. 2008), cucumber (Shang et al. 2012), Bambusa oldhamii
Submitted 21 January 2015, last revision 21 October 2015, accepted 3 November 2015. Abbreviations: CTAB - cetyltrimethyl ammonium bromide; DPI - days post-inoculation; Foc TR4 - Fusarium oxysporum f. sp. cubense Tropical Race 4; GFP - green fluorescent protein; PAL - phenylalanine ammonia lyase; qPCR - quantitative PCR; RACE - rapid amplification of cDNA ends. Acknowledgments: This research was funded by the Ministry of Science and Technology of P.R. China (No. 2011AA10020605) and the Modern Agro-industry Technology Research System (nycytx-33). Z. Wang and J.-Y. Li contributed equally to this work. * Corresponding author; fax: (+86) 898 66890772, e-mail: [email protected]

Z. WANG et al.
460
(Hsieh et al. 2011), Lycoris radiata (Jiang et al. 2011, 2013), Cistanche deserticola (Hu et al. 2011), Jatropha curcas (Gao et al. 2012), Dracaena cambodiana (Wang et al. 2013), Salvia miltiorrhiza (Hou et al. 2013), and Populus trichocarpa (Shi et al. 2010). Phenylalanine ammonia lyase is encoded by a small multigene family in plants (Cramer et al. 1989, Ohl et al. 1990). Four PALs are contained in the genome of Arabidopsis (Cochrane et al. 2004), seven in cucumber (Shang et al. 2012), and five in P. trichocarpa (Shi et al. 2010). In the banana DH-Pahang (AA group) genome, 8 PALs are predicted to distribute in 11 chromosomes (D'Hont et al. 2012). So far, three PAL genes have been cloned from Cavendish bananas (AAA group) including two PALs involved in banana fruit chilling tolerance (Chen et al. 2008), and one PAL involved in banana
resistance to Mycosphaerella fijiensis infection, which causes an important and destructive leaf fungal disease (Alvarez et al. 2013). In general, knowledge regarding these genes in bananas interacting with Foc TR4 is limited, and only PAL enzyme activity during banana infection with Foc TR4 has been reported (Thangavelu et al. 2003, Fishal et al. 2010). Furthermore, there is currently no data published about expression of these genes in relation to the Fusarium wilt of banana. Previously, we assembled a high-quality banana root transcriptome using RNA-sequencing (Wang et al. 2012), and performed further analyses to isolate four partial PAL genes. In this study, we describe molecular cloning and expression of four different PAL genes (MaPAL1, MaPAL2, MaPAL3, and MaPAL4) from two banana cultivars.
Materials and methods Plants and treatments: We used banana (Musa acuminata L. AAA group) cv. Cavendish, which is susceptible to Foc TR4, as well as a resistant mutant cv. Nongke No. 1. The banana cultivars were grown in Hoagland’s solution (Arnon and Hoagland 1939) in a greenhouse with natural irradiance. Maximum and minimum temperatures were 30 and 25 °C, respectively, and a relative humidity oscillated between 55 and 80 %. Our previous study confirmed that the growth characteristics and virulence of green fluorescent protein (GFP)-tagged Foc TR4 did not change, and the fungus could efficiently infect banana plants and induce disease symptoms (Yin et al. 2011). The fungus was grown on a potato dextrose agar medium and incubated at 28 °C for 5 d. The roots of two-month-old susceptible and resistant cultivars of banana were incubated in a Foc TR4 spore suspension of 1.0 × 106 conidia cm-3 for 2 h. The entire root system was harvested at 0, 2, 4, and 6 d post-inoculation (DPI), flash-frozen in liquid nitrogen, and stored at -70 °C. For each time point, we used 10 plants. The experiment was repeated three times. The roots of the uninfected banana plants were harvested at 0 d as described above. To determine tissue-specific expression, samples were collected from roots, pseudostems, corms, leaves, flowers, and fruits of eight-month-old plants. All collected materials were flash-frozen in liquid nitrogen and stored at -70 °C until use. Cloning full-length MaPAL cDNA: The partial sequences of four PAL genes (ID numbers: scaffold15858, scaffold2051, scaffold9618, and scaffold8809) were obtained from the banana root transcriptome using RNA-sequencing (Wang et al. 2012). To obtain the full-length cDNA, four specific primers (Table 1) were designed based on the fragments, and a nested PCR strategy was applied for 5' rapid-amplification of cDNA ends (RACE). These reverse
primers were used in both cloning analyse and expression analyse. The total RNA was extracted from banana roots using a modified cetyltrimethylammonium bromide (CTAB) method (Wan and Wilkins 1994). Synthesis of cDNA, PCR amplification, and PCR product sequencing were performed as previously described (Li et al. 2012b). The obtained PAL sequences were analyzed by the BLASTn algorithm (http://www.ncbi.nlm.nih.gov/blastn), and the deduced amino acid sequences were compared with the banana DH-Pahang (AA group) genome database (http://banana-genome.cirad.fr/greenphyl) and analyzed with the Expert Protein Analysis System (http://www.expasy.org/). The percentages of similarity between MaPAL and PAL proteins from the DH-Pahang genome were calculated using the DNAman 6.0 software (Lynnon Biosoft, Quebec, Canada). A phylogenetic tree was constructed with MEGA 4 (Tamura et al. 2007). Semi-quantitative reverse transcriptase PCR for MaPAL expression analysis: Semi-quantitative RT-PCR was performed using gene-specific forward and reverse primers (Table 1) for different MaPALs. The total RNA was extracted from roots, corms, pseudostems, leaves, flowers, and fruits by a modified CTAB method (Wan and Wilkins 1994). The first strand of cDNA was synthesized from 2 μg of the poly-(A)+ RNA from each sample using AMV reverse transcriptase (Promega, Heidelberg, Germany). The PCR conditions were as follows: 94 °C for 4 min followed by 30 cycles of amplification (94 °C for 10 s, 56 °C for 20 s, and 72 °C for 30 s), and 72 °C for 10 min. As internal controls, MaActin transcripts (Genebank acc. No. EF672732) were amplified using appropriate forward (5΄-CGAGGC TCAATCAAAGA-3΄) and reverse (5΄-ACCAGCAA GGTCCAAAC-3΄) primers. The experiments were repeated at least three times, and similar results were obtained for each.

CHARACTERIZATION OF PAL GENES IN BANANA
461
Real time quantitative PCR analysis: The total RNA was isolated from banana roots before treatment. The first strand of cDNA was synthesized from 2 μg of the poly-(A)+ RNA from each sample using AMV reverse transcriptase. A SYBR® Premix Ex Taq™ (TaKaRa, Dalian, China) was used in 0.025 cm3 of a reaction mixture with 0.5 mm3 of a ROX reference dye. Primers (100 nM each) were mixed with the equivalent of 100 ng of a reverse-transcribed RNA template per reaction. In all experiments, negative controls containing no template RNA were subjected to the same procedure to exclude or detect any possible contamination. Before proceeding with the actual experiments, a series of template dilutions were performed to determine an optimal template concentration necessary to obtain a maximal amplifi-cation of the target. Each real time quantitative PCR (qPCR) was performed with Stratagene Mx3000P (Stratagene, La Jolla, CA, USA). Thermal cycling conditions were as follows: 94 °C for 3 min followed by 40 cycles of 94 °C for 7 s, 55 °C for 10 s, and 72 °C for 15 s. The reactions were performed in triplicate, and data were analyzed using the MxProTM QPCR software (Stratagene). Transcription of MaActin was used as control. Differences in Ct values between the MaPALs and MaActin transcripts were expressed as fold-changes related to MaActin.
Assay for PAL activity: PAL activity in plant extracts was determined spectrophotometrically as described previously (Edwards and Kessmann 1992). Formation of a trans-cinnamic acid product was monitored at 290 nm with a T6 UV spectrophotometer (Beijing Persee General Equipment Co., Beijing, China). In brief, approximately 1 g of banana roots was ground under cooling with liquid nitrogen in a mortar and pestle and suspended in 10 cm3 of an extraction buffer containing 50 mM Tris-HCl buffer (pH 8.5), 5 mM Na2EDTA, 15 mM β-mercaptoethanol, 1 mM PMSF, and 0.15 % (m/v) polyvinylpyrrolidine. The homogenate was centrifuged at 12 000 g and 4 °C for 20 min. The reaction was carried out at 30 °C with 0.5 cm3 of the crude enzyme extract in 50 mM Tris-HCl (pH 8.5) containing 12 mM L-phenylalanine, with a control that contained no substrate. One unit of PAL deaminates 1.0 μmol of L-phenylalanine to trans-cinnamate and NH3 each hour at pH 8.5 and 30 °C using a coefficient of absorbance of 1.0 × 104 M-1 cm-1 for cinnamate. Protein content was determined by Bradford assay (Bradford 1976). All enzyme assays were independently replicated three times. Statistical analysis: The SAS 9.0 software for Windows was used to analyze the results. P-values < 0.05 were considered statistically significant. The experimental results obtained were expressed as means ± standard deviations. All analyses were performed in triplicate.
Table 1. Primers used for cloning and expression analyses of MaPALs.
Gene Forward primer for cloning (5′ to 3′) Forward primer for PCR (5′ to 3′) Reverse primer (5′ to 3′)
MaPAL1 TCCGACACGCTTCAGCGCCTCGACG CCTCCATCGCCTCATAAACC CAGTCAGCCAGGAAATCGC MaPAL2 GTTTCCCCGGCCGGTGAGCAGGCCG GCCCTTGAAGCCGTAGTCC TCCACGAGCAAGACCCACT MaPAL3 ACGAAGGCAATGTGTTTCCCGACTC GGTCCTGCTTTGGCTTCTG TCTCGTCCCGCTGTCTTACA MaPAL4 AAGGGCGCCTCCTTGCTTGGTCCTC AGCCCATTGTTGTAGAAGTCGT TCCAGAAGCCAAAGCAGG
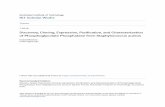
Results Four partial PAL genes were acquired from the banana root transcriptome, and their full-length cDNAs were cloned using RACE. The cDNA and deduced amino acid sequences of MaPALs described in this study were deposited in GenBank under the following accession numbers: MaPAL1 (KF582545), MaPAL2 (KF582546), MaPAL3 (KF594427), and MaPAL4 (KF594428). MaPAL1, 2, 3, and 4 consisted of 703, 652, 654, and 651 amino acids, respectively. MaPAL1 had the highest molecular mass (77.3 kDa) and MaPAL2 had it the lowest (70.2 kDa). Theoretical isoelectric points were calculated to be 5.82, 6.43, 6.12, and 6.56 for MaPAL1, 2, 3, and 4, respectively. All the MaPAL amino acid sequences contained phenylalanine and histidine ammonia lyase domains (GTITASGDLVPLSYIAG)
(Fig. 1). However, there was a low sequence similarity among the four MaPALs identified (74.072 %). They were then annotated using BLASTp in NCBI, and their sequences mapped with respect to the banana DH-Pahang Table 2. Results of BLASTp for MaPALs in the banana DH-Pahang genome.
Gene name
DH-Pahang Gene name
Identities [%]
MaPAL1 GSMUA_Achr9P15990_001 PAL 99.62 MaPAL2 GSMUA_Achr5P18560_001 PAL 1 99.29 MaPAL3 GSMUA_Achr1P09070_001 PAL 99.80 MaPAL4 GSMUA_Achr5P03950_001 PAL 1 99.49
Z. WANG et al.
462
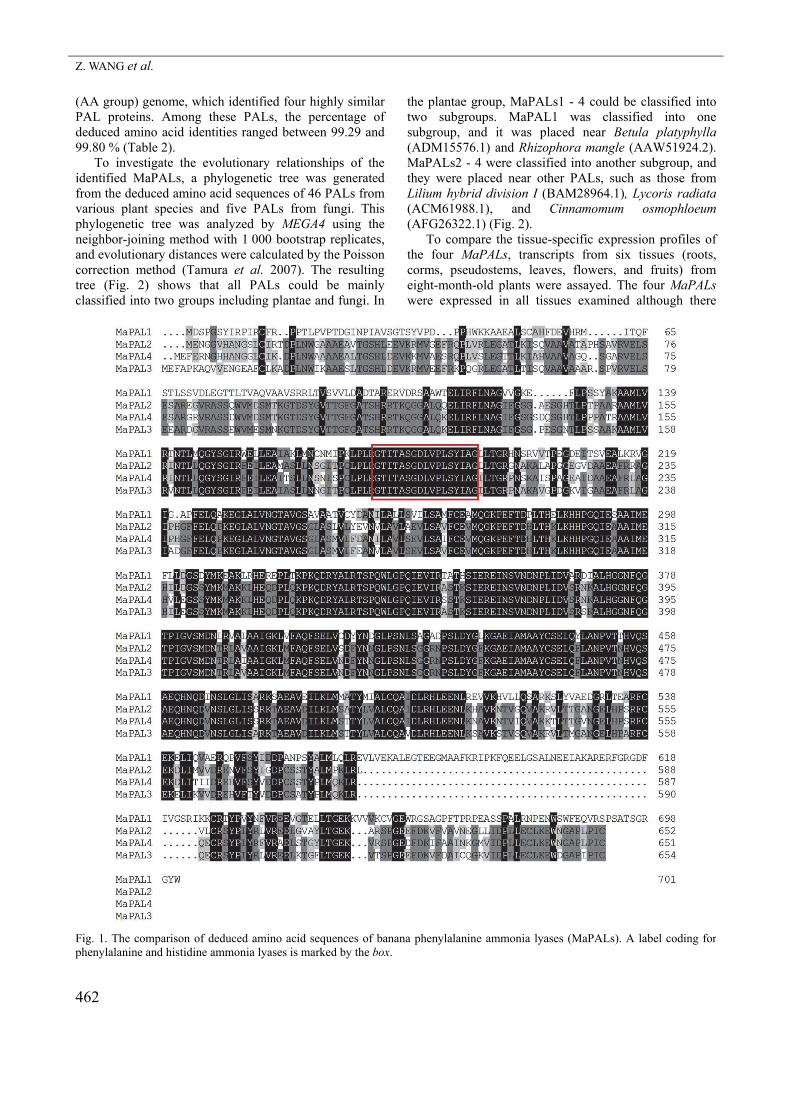
(AA group) genome, which identified four highly similar PAL proteins. Among these PALs, the percentage of deduced amino acid identities ranged between 99.29 and 99.80 % (Table 2). To investigate the evolutionary relationships of the identified MaPALs, a phylogenetic tree was generated from the deduced amino acid sequences of 46 PALs from various plant species and five PALs from fungi. This phylogenetic tree was analyzed by MEGA4 using the neighbor-joining method with 1 000 bootstrap replicates, and evolutionary distances were calculated by the Poisson correction method (Tamura et al. 2007). The resulting tree (Fig. 2) shows that all PALs could be mainly classified into two groups including plantae and fungi. In
the plantae group, MaPALs1 - 4 could be classified into two subgroups. MaPAL1 was classified into one subgroup, and it was placed near Betula platyphylla (ADM15576.1) and Rhizophora mangle (AAW51924.2). MaPALs2 - 4 were classified into another subgroup, and they were placed near other PALs, such as those from Lilium hybrid division I (BAM28964.1), Lycoris radiata (ACM61988.1), and Cinnamomum osmophloeum (AFG26322.1) (Fig. 2). To compare the tissue-specific expression profiles of the four MaPALs, transcripts from six tissues (roots, corms, pseudostems, leaves, flowers, and fruits) from eight-month-old plants were assayed. The four MaPALs were expressed in all tissues examined although there
Fig. 1. The comparison of deduced amino acid sequences of banana phenylalanine ammonia lyases (MaPALs). A label coding forphenylalanine and histidine ammonia lyases is marked by the box.

CHARACTERIZATION OF PAL GENES IN BANANA
463
Fig. 2. The phylogenetic analysis of phenylalanine ammonia lyases (PALs) from banana and other plant species and fungi. Values shown at branch nodes are confidence levels from 1 000 replicate bootstrap samplings. The scale bar indicates the evolutionary distance between the groups. The amino acid sequences used in phylogenetic analysis correspond to the Genbank accession numbers:Plantae (PAL): Ophioglossum reticulatum (AAW80642.1), Betula platyphylla (ADM15576.1), Pellia epiphylla (AAW80645.1), Equisetum arvense (AAW80639.1), Rhizophora mangle (AAW51924.2), Psilotum nudum (AAW80640.1), Blechnum spicant(AAW80643.1), Ginkgo biloba (ABU49842.1), Diphasiastrum tristachyum (AAW80636.1), Cinnamomum osmophloeum (AFG26322.1), Jatropha curcas (ABI33979.1), Malus hybrid cultivar (AFG30054.1), Vitis vinifera (AEX32784.1, ABM67591.1, AEX32790.1), Lycoris radiata (ACM61988.1), Rhus chinensis (AGH13333.1), Prunus salicina (AFP24940.1), Lilium hybrid division I (BAM28964.1), Pyrus communis (AGL50914.1, AGL81344.1), Populus tomentosa (AFZ78651.1), Glycine soja(ACT32033.1), Daucus carota (BAG31930.1, BAC56977.1), Populus trichocarpa (ACC63889.1), Theobroma cacao (EOY07856.1), Morus alba var. multicaulis (AEE81750.1), Manihot esculenta (AAK60275.1), Lactuca sativa (AAL55242.1), Gossypium hirsutum (AER12109.1), Rubus idaeus (AAF40224.1), Hibiscus cannabinus (AFN85669.1), Arabidopsis thaliana (AEC09341.1, AEE79055.1, AED90715.1, AEE74893.1); Fungi (PAL): Aspergillus clavatus NRRL 1 (FGSG_09311), Fusarium graminearum NRRL 31084 (FGSC_10136), Aspergillus fumigatus Af293 (XP_755245.1), Aspergillus niger CBS 513.88 (XP_001401803.1), and Uncinocarpus reesii 1704 (XP_002544702.1).
Z. WANG et al.
464
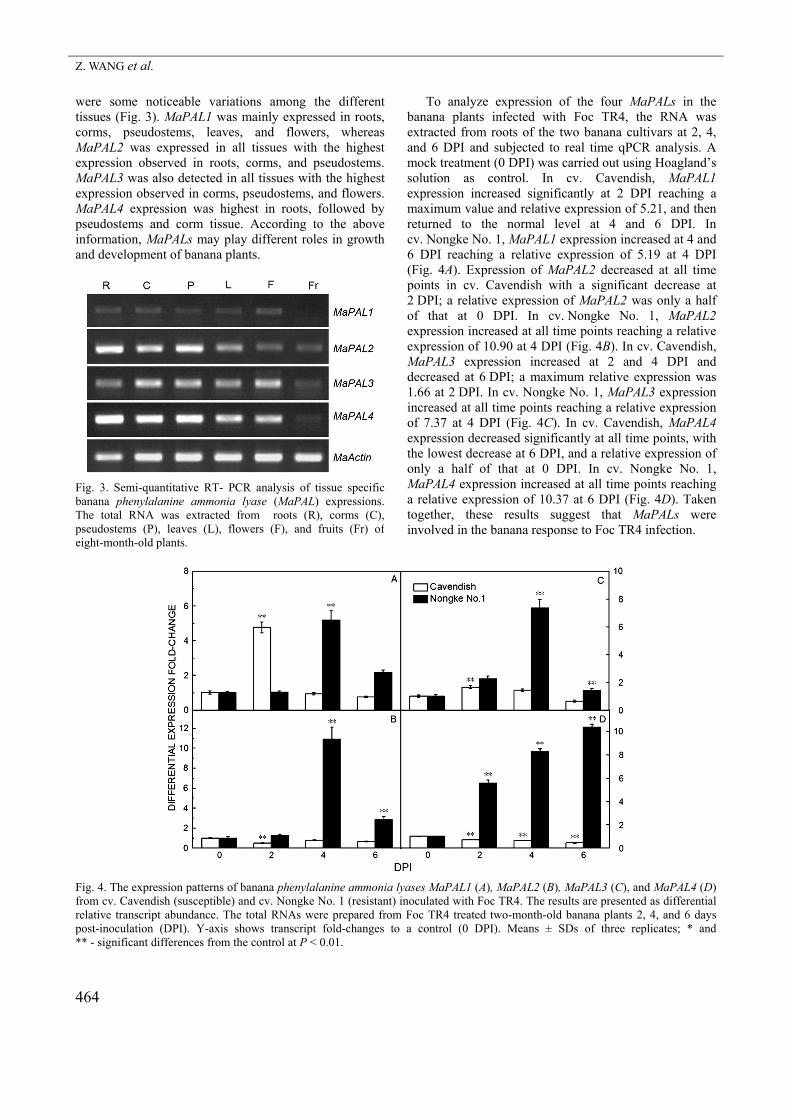
were some noticeable variations among the different tissues (Fig. 3). MaPAL1 was mainly expressed in roots, corms, pseudostems, leaves, and flowers, whereas MaPAL2 was expressed in all tissues with the highest expression observed in roots, corms, and pseudostems. MaPAL3 was also detected in all tissues with the highest expression observed in corms, pseudostems, and flowers. MaPAL4 expression was highest in roots, followed by pseudostems and corm tissue. According to the above information, MaPALs may play different roles in growth and development of banana plants.
Fig. 3. Semi-quantitative RT- PCR analysis of tissue specificbanana phenylalanine ammonia lyase (MaPAL) expressions. The total RNA was extracted from roots (R), corms (C),pseudostems (P), leaves (L), flowers (F), and fruits (Fr) ofeight-month-old plants.
To analyze expression of the four MaPALs in the banana plants infected with Foc TR4, the RNA was extracted from roots of the two banana cultivars at 2, 4, and 6 DPI and subjected to real time qPCR analysis. A mock treatment (0 DPI) was carried out using Hoagland’s solution as control. In cv. Cavendish, MaPAL1 expression increased significantly at 2 DPI reaching a maximum value and relative expression of 5.21, and then returned to the normal level at 4 and 6 DPI. In cv. Nongke No. 1, MaPAL1 expression increased at 4 and 6 DPI reaching a relative expression of 5.19 at 4 DPI (Fig. 4A). Expression of MaPAL2 decreased at all time points in cv. Cavendish with a significant decrease at 2 DPI; a relative expression of MaPAL2 was only a half of that at 0 DPI. In cv. Nongke No. 1, MaPAL2 expression increased at all time points reaching a relative expression of 10.90 at 4 DPI (Fig. 4B). In cv. Cavendish, MaPAL3 expression increased at 2 and 4 DPI and decreased at 6 DPI; a maximum relative expression was 1.66 at 2 DPI. In cv. Nongke No. 1, MaPAL3 expression increased at all time points reaching a relative expression of 7.37 at 4 DPI (Fig. 4C). In cv. Cavendish, MaPAL4 expression decreased significantly at all time points, with the lowest decrease at 6 DPI, and a relative expression of only a half of that at 0 DPI. In cv. Nongke No. 1, MaPAL4 expression increased at all time points reaching a relative expression of 10.37 at 6 DPI (Fig. 4D). Taken together, these results suggest that MaPALs were involved in the banana response to Foc TR4 infection.
Fig. 4. The expression patterns of banana phenylalanine ammonia lyases MaPAL1 (A), MaPAL2 (B), MaPAL3 (C), and MaPAL4 (D)from cv. Cavendish (susceptible) and cv. Nongke No. 1 (resistant) inoculated with Foc TR4. The results are presented as differentialrelative transcript abundance. The total RNAs were prepared from Foc TR4 treated two-month-old banana plants 2, 4, and 6 days post-inoculation (DPI). Y-axis shows transcript fold-changes to a control (0 DPI). Means ± SDs of three replicates; * and ** - significant differences from the control at P < 0.01.
CHARACTERIZATION OF PAL GENES IN BANANA
465
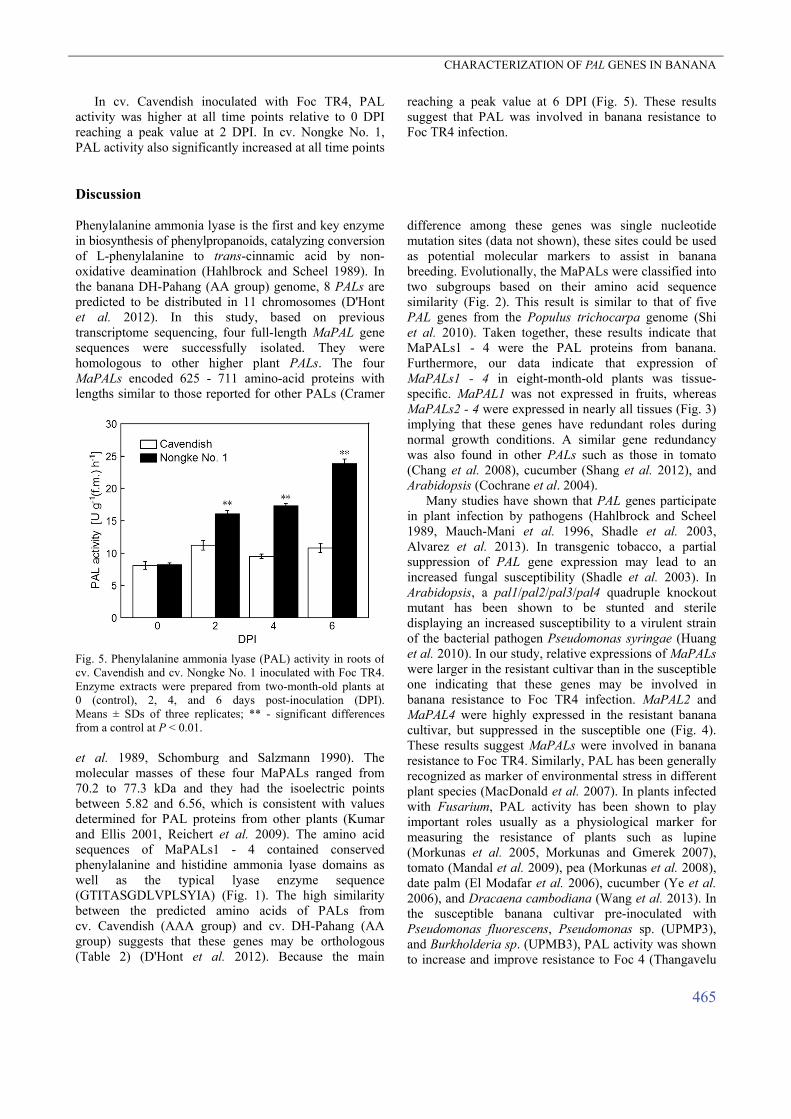
In cv. Cavendish inoculated with Foc TR4, PAL activity was higher at all time points relative to 0 DPI reaching a peak value at 2 DPI. In cv. Nongke No. 1, PAL activity also significantly increased at all time points
reaching a peak value at 6 DPI (Fig. 5). These results suggest that PAL was involved in banana resistance to Foc TR4 infection.
Discussion Phenylalanine ammonia lyase is the first and key enzyme in biosynthesis of phenylpropanoids, catalyzing conversion of L-phenylalanine to trans-cinnamic acid by non-oxidative deamination (Hahlbrock and Scheel 1989). In the banana DH-Pahang (AA group) genome, 8 PALs are predicted to be distributed in 11 chromosomes (D'Hont et al. 2012). In this study, based on previous transcriptome sequencing, four full-length MaPAL gene sequences were successfully isolated. They were homologous to other higher plant PALs. The four MaPALs encoded 625 - 711 amino-acid proteins with lengths similar to those reported for other PALs (Cramer
Fig. 5. Phenylalanine ammonia lyase (PAL) activity in roots of cv. Cavendish and cv. Nongke No. 1 inoculated with Foc TR4.Enzyme extracts were prepared from two-month-old plants at0 (control), 2, 4, and 6 days post-inoculation (DPI). Means ± SDs of three replicates; ** - significant differencesfrom a control at P < 0.01. et al. 1989, Schomburg and Salzmann 1990). The molecular masses of these four MaPALs ranged from 70.2 to 77.3 kDa and they had the isoelectric points between 5.82 and 6.56, which is consistent with values determined for PAL proteins from other plants (Kumar and Ellis 2001, Reichert et al. 2009). The amino acid sequences of MaPALs1 - 4 contained conserved phenylalanine and histidine ammonia lyase domains as well as the typical lyase enzyme sequence (GTITASGDLVPLSYIA) (Fig. 1). The high similarity between the predicted amino acids of PALs from cv. Cavendish (AAA group) and cv. DH-Pahang (AA group) suggests that these genes may be orthologous (Table 2) (D'Hont et al. 2012). Because the main
difference among these genes was single nucleotide mutation sites (data not shown), these sites could be used as potential molecular markers to assist in banana breeding. Evolutionally, the MaPALs were classified into two subgroups based on their amino acid sequence similarity (Fig. 2). This result is similar to that of five PAL genes from the Populus trichocarpa genome (Shi et al. 2010). Taken together, these results indicate that MaPALs1 - 4 were the PAL proteins from banana. Furthermore, our data indicate that expression of MaPALs1 - 4 in eight-month-old plants was tissue-specific. MaPAL1 was not expressed in fruits, whereas MaPALs2 - 4 were expressed in nearly all tissues (Fig. 3) implying that these genes have redundant roles during normal growth conditions. A similar gene redundancy was also found in other PALs such as those in tomato (Chang et al. 2008), cucumber (Shang et al. 2012), and Arabidopsis (Cochrane et al. 2004). Many studies have shown that PAL genes participate in plant infection by pathogens (Hahlbrock and Scheel 1989, Mauch-Mani et al. 1996, Shadle et al. 2003, Alvarez et al. 2013). In transgenic tobacco, a partial suppression of PAL gene expression may lead to an increased fungal susceptibility (Shadle et al. 2003). In Arabidopsis, a pal1/pal2/pal3/pal4 quadruple knockout mutant has been shown to be stunted and sterile displaying an increased susceptibility to a virulent strain of the bacterial pathogen Pseudomonas syringae (Huang et al. 2010). In our study, relative expressions of MaPALs were larger in the resistant cultivar than in the susceptible one indicating that these genes may be involved in banana resistance to Foc TR4 infection. MaPAL2 and MaPAL4 were highly expressed in the resistant banana cultivar, but suppressed in the susceptible one (Fig. 4). These results suggest MaPALs were involved in banana resistance to Foc TR4. Similarly, PAL has been generally recognized as marker of environmental stress in different plant species (MacDonald et al. 2007). In plants infected with Fusarium, PAL activity has been shown to play important roles usually as a physiological marker for measuring the resistance of plants such as lupine (Morkunas et al. 2005, Morkunas and Gmerek 2007), tomato (Mandal et al. 2009), pea (Morkunas et al. 2008), date palm (El Modafar et al. 2006), cucumber (Ye et al. 2006), and Dracaena cambodiana (Wang et al. 2013). In the susceptible banana cultivar pre-inoculated with Pseudomonas fluorescens, Pseudomonas sp. (UPMP3), and Burkholderia sp. (UPMB3), PAL activity was shown to increase and improve resistance to Foc 4 (Thangavelu

Z. WANG et al.
466
et al. 2003, Fishal et al. 2010). In the resistant banana cultivar, PAL activity is higher than that in the susceptible cultivar, and PAL activity in leaves quickly increases following Foc 1 infection (Gunavathi et al. 2003). In our study, the PAL activity in cv. Nongke No. 1 was higher than that in cv. Cavendish at all time points (Fig. 5). These results suggest that PAL activity positively correlated with resistance, which is consistent with results of previous studies (Gunavathi et al. 2003, Damodaran et al. 2009). Comparison of PAL activity and MaPAL gene expression demonstrates that the expression data are more convincing. The elevated relative expressions of MaPALs2 - 4 were higher in the resistant
cultivar than in the susceptible one, and their expression patterns were consistent with the progression of Foc TR4 infection. Therefore, these genes could be used as new markers of Foc TR4 infection. In conclusion, four banana PAL genes (MaPAL1, MaPAL2, MaPAL3, and MaPAL4) were cloned from banana roots. All four MaPALs were expressed in roots, and MaPAL1 was not expressed in fruits. Activity of PAL and MaPAL gene expression patterns were consistent with banana resistance to Foc TR4. All four MaPALs were induced in the resistant banana cultivar after being inoculated with Foc TR4, which indicates that MaPALs were involved in banana resistance to Foc TR4 infection.
References Alvarez, J.C., Rodriguez, H.A., Rodriguez-Arango, E.,
Monsalve, Z.I., Morales, O.J.G., Arango, I.R.E.: Characterization of a differentially expressed phenylalanine ammonia-lyase gene from banana induced during Mycosphaerella fijiensis infection. - J. Plant Studies 2: p35, 2013.
Arnon, D.I., Hoagland D.R.: A comparison of water culture and soil as media for crop production. - Science 89: 512-514, 1939.
Aurore, G., Parfait, B., Fahrasmane, L.: Bananas, raw materials for making processed food products. - Trends Food Sci. Technol. 20: 78-91, 2009.
Bradford, M.M.: A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. - Anal. Biochem. 72: 248-254, 1976.
Chaman, M.E., Copaja, S.V., Argandona, V. H.: Relationships between salicylic acid content, phenylalanine ammonia-lyase (PAL) activity, and resistance of barley to aphid infestation. - J. Agr. Food Chem. 51: 2227-2231, 2003.
Chang, A., Lim, M.H., Lee, S.W., Robb, E.J., Naza,r R.N.: Tomato phenylalanine ammonia-lyase gene family, highly redundant but strongly underutilized. - J. biol. Chem. 283: 33591-33601, 2008.
Chen, J., He, L., Jiang, Y., Wang, Y., Joyce, D., Ji, Z., Lu, W.: Role of phenylalanine ammonia-lyase in heat pretreatment-induced chilling tolerance in banana fruit. - Physiol. Plant. 132: 318-328, 2008.
Cochrane, F.C., Davin, L.B., Lewis, N.G.: The Arabidopsis phenylalanine ammonia lyase gene family: kinetic characterization of the four PAL isoforms. - Phytochemistry 65: 1557-1564, 2004.
Cramer, C.L., Edwards, K., Dron, M., Liang, X., Dildine, S.L., Bolwell, G.P., Dixon, R.A., Lamb, C.J., Schuch, W.: Phenylalanine ammonia-lyase gene organization and structure. - Plant mol. Biol. 12: 367-383, 1989.
Dixon, R.A., Paiva, N.L.: Stress-induced phenylpropanoid metabolism. - Plant Cell 7: 1085, 1995.
Damodaran, T., Kumar, N., Kavino, M.: Breeding and evaluation of Musa hybrids resistant to Fusarium oxysporum f. sp. cubense race 1. - Fruits 64: 3-12, 2009.
D'Hont, A., Denoeud, F., Aury, J.M., Baurens, F.C., Carreel, F., Garsmeur, O., Noel, B., Bocs, S., Droc, G., Rouard, M., Da Silva, C., Jabbari, K., Cardi, C., Poulain, J., Souquet, M.,
Labadie, K., Jourda, C., Lengellé, J., Rodier-Goud, M., Alberti, A., Bernard, M., Correa, M., Ayyampalayam, S., Mckain, M.R., Leebens-Mack, J., Burgess, D., Freeling, M., Mbeguie-A-Mbeguie, D., Chabannes, M., Wicker, T., Panaud, O., Barbosa, J., Hribova, E., Heslop-Harrison, P., Habas, R., Rivallan, R., Francois, P.: The banana (Musa acuminata) genome and the evolution of monocotyledonous plants. - Nature 448: 213-217, 2012.
Edwards, R., Kessmann, H.: Isoflavonoid phytoalexins and their biosynthetic enzymes. - In: Gurr, S.J., McPherson, M.J., Bowles, D.J. (ed.): Molecular Plant Pathology: a Practical Approach. Pp 45-62. Oxford University Press, Oxford 1992.
El Modafar, C., El Boustani, E., Rahioui, B., El Meziane, A., El Alaoui-Talibi, Z.: Suppression of phenylalanine ammonia-lyase activity elicited in date palm by Fusarium oxysporum f. sp. albedinis hyphal wall elicitor. - Biol. Plant. 50: 697-700, 2006.
Fishal, E.M.M, Meon, S., Yun, W.M.: Induction of tolerance to Fusarium wilt and defense-related mechanisms in the cultivar of susceptible berangan banana pre-inoculated with Pseudomonas sp. (UPMP3) and Burkholderia sp. (UPMB3). - Agr. Sci. China 9: 1140-1149, 2010.
Gao, J., Zhang, S., Cai, F., Zheng, X., Lin, N., Qin, X., Ou, Y., Gu, X., Zhu, X., Xu, Y., Chen, F.: Characterization, and expression profile of a phenylalanine ammonia lyase gene from Jatropha curcas L. - Mol. Biol. Rep. 39: 3443-3452, 2012.
Gunavathi, J., Kumar, N., Soorianathasundaram, K., Samiappan, R.: Reaction of synthetic diploids to Fusarium oxysporum f. sp. cubense (Race-1) and to a Fusarium-nematode complex. - In: Proceedings of the 2nd International Symposium on Fusarium Wilt on Banana. P. 22. Saverdor de Bahia 2003.
Hahlbrock, K., Scheel, D.: Physiology and molecular biology of phenylpropanoid metabolism. - Annu. Rev. Plant Biol. 40: 347-369, 1989.
Hou, X., Shao, F., Ma, Y., Lu, S.: The phenylalanine ammonia-lyase gene family in Salvia miltiorrhiza: genome-wide characterization, molecular cloning and expression analysis. - Mol. Biol. Rep. 40: 4301-4310, 2013.
Hsieh, L.S., Hsieh, Y.L., Yeh, C.S., Cheng, C., Yang, C., Lee, P.: Molecular characterization of a phenylalanine ammonia-lyase gene (BoPAL1) from Bambusa oldhamii. - Mol. Biol. Rep. 38: 283-290, 2011.

CHARACTERIZATION OF PAL GENES IN BANANA
467
Hu, G.S., Jia, J.M., Hur, Y.J., Chung, Y., Lee, J., Yun, D., Chung, W., Yi, G., Kim, T., Kim, D.: Molecular characterization of phenylalanine ammonia lyase gene from Cistanche deserticola. - Mol. Biol. Rep. 38: 3741-3750, 2011.
Huang, J., Gu, M., Lai, Z., Fan, B., Shi, K., Zhou, Y.H., Yu, J.Q., Chen, Z.: Functional analysis of the Arabidopsis PAL gene family in plant growth, development, and response to environmental stress. - Plant Physiol. 153: 1526-1538, 2010.
Hwang, S.C., Ko, W.H.: Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan. - Plant Dis. 88: 580-588, 2004.
Jiang, Y., Xia, N., Li, X., Shen, W., Liang, L., Wang, C., Wang, R., Peng, F., Xia, B.: Molecular cloning and characterization of a phenylalanine ammonia-lyase gene (LrPAL) from Lycoris radiata. - Mol. Biol. Rep. 38: 1935-1940, 2011.
Jiang, Y., Xia, B., Liang, L., Li, X., Xu, S., Peng, F., Wang, R.: Molecular and analysis of a phenylalanine ammonia-lyase gene (LrPAL2) from Lycoris radiata. - Mol. Biol. Rep. 40: 2293-2300, 2013.
Jorrin, J., Dixon, R.A.: Stress responses in alfalfa (Medicago sativa L.) II. Purification, characterization, and induction of phenylalanine ammonia-lyase isoforms from elicitor-treated cell suspension cultures. - Plant Physiol. 92: 447-455, 1990.
Koenig, R.L., Ploetz, R.C., Kistler, H.C.: Fusarium oxysporum f. sp. cubense consists of a small number of divergent and globally distributed clonal lineages. - Phytopathology 87: 915-923, 1997.
Kumar, A., Ellis, B.E.: The Phenylalanine ammonia-lyase gene family in raspberry: structure, expression, and evolution. - Plant Physiol. 127: 230-239, 2001.
Leyva, A., Jarillo, J.A., Salinas, J., Martinez-Zapater, J.M.: Low temperature induces the accumulation of phenylalanine ammonia-lyase and chalcone synthase mRNAs of Arabidopsis thaliana in a light-dependent manner. - Plant Physiol. 108: 39-46, 1995.
Li, C., Deng, G., Yang, J., Viljoen, A., Jin, Y., Kuang, R., Zuo, C., Lv, Z., Yang, Q., Sheng, O., Wei, Y., Hu, C., Dong, T., Yi, G.: Transcriptome profiling of resistant and susceptible Cavendish banana roots following inoculation with Fusarium oxysporum f. sp. cubense tropical race 4. - BMC Genomics 13: 374, 2012a.
Li, M., Xu, B., Liu, J., Yang, X., Zhang, J., Jia, C., Ren, L., Jin, Z.: Identification and expression analysis of four 14-3-3 genes during fruit ripening in banana (Musa acuminata L. AAA group, cv. Brazilian). - Plant Cell Rep. 31: 369-378, 2012b.
MacDonald, M.J., D’Cunha, G.B.: A modern view of phenylalanine ammonia lyase. - Biochem. Cell Biol. 85: 273-282, 2007.
Mandal, S., Mallick, N., Mitra, A.: Salicylic acid-induced resistance to Fusarium oxysporum f. sp. lycopersici in tomato. - Plant Physiol. Biochem. 47: 642-649, 2009.
Mauch-Mani, B., Slusarenko, A.J.: Production of salicylic acid precursors is a major function of phenylalanine ammonia-lyase in the resistance of Arabidopsis to Peronospora parasitica. - Plant Cell 8: 203-212, 1996.
Morkunas, I., Bednarski, W., Kopyra, M.: Defense strategies of pea embryo axes with different levels of sucrose to Fusarium oxysporum and Ascochyta pisi. - Physiol. mol. Plant Pathology 72: 167-178, 2008.
Morkunas, I., Gmerek, J.: The possible involvement of
peroxidase in defense of yellow lupine embryo axes against Fusarium oxysporum. - J. Plant Physiol. 164: 185-194, 2007.
Morkunas, I., Marczak, L., Stachowiak, J., Stobiecki, M.: Sucrose-induced lupine defense against Fusarium oxysporum: Sucrose-stimulated accumulation of isoflavonoids as a defense response of lupine to Fusarium oxysporum. - Plant Physiol. Biochem. 43: 363-373, 2005.
Nugroho, L.H., Verberne, M.C., Verpoorte, R.: Activities of enzymes involved in the phenylpropanoid pathway in constitutively salicylic acid-producing tobacco plants. - Plant Physiol. Biochem. 40: 755-760, 2002.
Ohl, S., Hedrick, S.A., Chory, J., Lamb, C.J.: Functional properties of a phenylalanine ammonia-lyase promoter from Arabidopsis. - Plant Cell 2: 837-848, 1990.
Reichert A.I., He X.Z., Dixon R.A.: Phenylalanine ammonia-lyase (PAL) from tobacco (Nicotiana tabacum): characterization of the four tobacco PAL genes and active heterotetrameric enzymes. - Biochem. J. 424: 233-242, 2009.
Robinson, J.C.: Bananas and Plantains. - CAB International, Oxford 1996.
Schomburg, D., Salzmann, M.: Class 4: lyases, phenylalanine ammonia-lyase. - In: Schomburg, D., Salzmann, M. (ed.): Enzyme Handbook. Pp. 1-5. Springer-Verlag, Berlin 1990.
Shadle, G.L., Wesley, S.V., Korth, K.L., Chen, F., Lamb, C., Dixon, R.A.: Phenylpropanoid compounds and disease resistance in transgenic tobacco with altered expression of L-phenylalanine ammonia-lyase. - Phytochemistry 64: 153-161, 2003.
Shang, Q.M., Li, L., Dong, C.J.: Multiple tandem duplication of the phenylalanine ammonia-lyase genes in Cucumis sativus L. - Planta 236: 1093-1105, 2012.
Shi, R., Sun, Y.H., Li, Q., Heber, S., Sederoff, R., Chiang, V.L.: Towards a systems approach for lignin biosynthesis in Populus trichocarpa: transcript abundance and specificity of the monolignol biosynthetic genes. - Plant Cell Physiol. 51: 144-163, 2010.
Stover, R.H.: Fusarial Wilt (Panama Disease) of Bananas and Other Musa Species. - Commonwealth Mycological Institute, Kew - Surrey 1962.
Tamura, K., Dudley, J., Nei, M., Kumar, S.: MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. - Mol. Biol. Evol. 24: 1596e9, 2007.
Thangavelu, R., Palaniswani, A., Doraiswamy, S., Velazhahan, R.: The effect of Pseudomonas fluorescens and Fusarium oxysporum f. sp. cubense on induction of defense enzymes and phenolics in banana. - Biol. Plant. 46: 107-112, 2003.
Visser, M., Gordon, T., Fourie, G., Viljoen, A.: Characterisation of South African isolates of Fusarium oxysporum f. sp. cubense from Cavendish bananas. - South Afr. J. Sci. 106: 1-6, 2010.
Wan, C.Y., Wilkins, T.A.: A modified hot borate method significantly enhance the yield of high-quality RNA from cotton (Gossypium hirsutum L.). - Anal. Biochem. 223: 7-12, 1994.
Wang, X., Gong, M., Tang, L., Zheng, S., Lou, J., Ou, L., Gomes-Laranjo, J., Zhang, C.: Cloning, bioinformatics and the enzyme activity analyses of a phenylalanine ammonia-lyase gene involved in dragon’s blood biosynthesis in Dracaena cambodiana. - Mol. Biol. Rep. 40: 97-107, 2013.
Wang, Z., Zhang, J., Jia, C., Liu, J., Li, Y., Yin, X., Xu, B., Jin, Z.: De Novo characterization of the banana root

Z. WANG et al.
468
transcriptome and analysis of gene expression under Fusarium oxysporum f. sp. cubense tropical race 4 infection. - BMC Genomics 13: 1-9, 2012.
Ye, S., Zhou, Y., Sun, Y., Zou, L., Yu, J.: Cinnamic acid causes oxidative stress in cucumber roots, and promotes incidence of Fusarium wilt. - Environ.exp. Bot. 56: 255-262, 2006.
Yin, X., Xu, B., Zheng, W., Wang, Z., Wang, B., Ha, W., Mao, H., Li, J., Sheng, Z., He, Y., Han, L., Chen, S., Zhang, J., Fu, Y., Wang, J., Jin, Z.: Characterization of early events in banana roots infected with green fluorescent protein-tagged Fusarium oxysporum f. sp. cubense. - Acta Hort. 897: 371-376, 2011.