
Molecular Cloning and Expression Analysis of a Zinc Finger ... · Molecular Cloning and Expression...
9
Received 27 Oct. 2003 Accepted 9 Mar. 2004 Supported by the Funds from the Science and Technology Commission of Shanxi Province (021034) and Returning Overseas Student Affair Office of Shanxi Province (20011575). * Author for correspondence. Tel :+86 (0)351 7127503 ;Fax:+86 (0)351 7123546 ;E-mail : <[email protected]>. http://www.chineseplantscience.com Acta Botanica Sinica 植 物 学 报 2004, 46 (9): 1091- 1099 Molecular Cloning and Expression Analysis of a Zinc Finger Protein Gene in Apple CAO Qiu-Fen 1* , Masato WADA 2 , MENG Yu-Ping 1 , SUN Yi 1 , CUI Gui-Mei 1 (1. Agricultural Biotechnology Research Center of Shanxi Province, Taiyuan 030031, China; 2. Department of Apple Research, National Institute of Fruit Tree Science, National Agricultural Research Organization Shimokuriyagawa, Morioka, Iwate 020-0123, Japan) Abstract: A cDNA library was created from stem apex tissue from Jonathan apples (Malus domestica Borkh.), harvested in June to August, during which the plant transitions from vegetative growth to reproductive growth. From this library, we isolated an expressed sequence tag (EST) sequence containing a zinc finger motif; using this sequence, a 779 bp cDNA fragment was obtained by using 5' RACE, and a final full-length cDNA encoding an apple zinc finger protein (named MdZF1; GenBank accession number AB116545) was obtained by further PCR. This zinc finger motif of MdZF1 has high homology with INDETERMINATE 1 (ID1) gene from maize which seemed to be involved in the transition to flowering. Northern blot and RT-PCR analyses showed that the MdZF1 expressed in the root, stem, leaves, shoot apex and floral organs of the apple, with expression levels higher in root, stem, leaves and floral shoot apex than that in floral organs (sepals, petals, stamens and pistils). Genomic Southern analysis showed that there was a single copy gene in apple genome. Key words: Malus domestica zinc finger protein (MdZF1); cloning; expression The zinc finger proteins belong to a family of regulatory transcription factors that contain “ finger-like” motifs; they are found in most living organisms, from yeasts to plants as well as mammals. The zinc finger motif can bind to DNA, RNA and DNA-RNA hybrids, as well as to other zinc finger proteins, and zinc finger-containing proteins normally have two or more motifs and are responsible for controlling the transcription and translation in an organism (Huang and Liu, 1998). The classical zinc finger protein is the TFⅢA type, also known as the Cys2/His2 or C2H2 type, which has been extensively studied at a structural level (Sakamoto et al., 2000). In plants, some Cys2/His2 zinc finger cDNAs have been associated with stress resistance (Huang et al., 2002; Wang and Yang, 2002) and growth (Colasanti et al., 1998). Colasanti et al. (1998) cloned the INDETERMI- NATE 1 (ID1) gene, which encoded a maize zinc finger pro- tein associated with the transition to flowering, suggesting that the ID1 gene product functions as a transcription regula- tor of floral development. The phenotype of id1 mutant and the expression analysis of ID1 suggested that the gene prod- uct act as a floral initiation signal like the florigen hypothesis. The shoot apex of higher plants contains an undifferen- tiated cluster of cells, called the shoot apical meristem, which maintains the propensity for vegetative growth and development during the initial stages of plant growth, and then switches to reproductive growth and development when external and internal conditions trigger reproductive signals. To investigate the molecular mechanisms respon- sible for the transition of apple shoot apex from vegetative to reproductive growth, we built a cDNA library from apple shoot apex at the flower bud during differentiation stage (June to August), and obtained a series of cDNA fragments. By comparing the sequences from database in DDBJ/ EMBL/GenBank, we found several expressed sequence tag (EST)sequences that contained a zinc finger motif. One sequence showed high homology with maize ID1 gene , and we were able to assemble a full-length cDNA for this zinc finger protein with 5′RACE and PCR techniques, which we designated MdZF1 (Malus domestica ZINC FINGER PROTEIN 1). Northern blot and RT-PCR methods were used to confirm and analyze the expression of MdZF1 in differ- ent apple tissues and floral organs. Southern blot was used to analyze the copy number of MdZF1 in apple genomes. 1 Materials and Methods 1.1 Plant materials Jonathan apple (Malus domestica Borkh.) tissues were harvested between June and August. Tissue samples
Transcript of Molecular Cloning and Expression Analysis of a Zinc Finger ... · Molecular Cloning and Expression...

Received 27 Oct. 2003 Accepted 9 Mar. 2004Supported by the Funds from the Science and Technology Commission of Shanxi Province (021034) and Returning Overseas Student AffairOffice of Shanxi Province (20011575).
* Author for correspondence. Tel:+86 (0)351 7127503;Fax:+86 (0)351 7123546;E-mail:<[email protected]>.
http://www.chineseplantscience.com
Acta Botanica Sinica植 物 学 报 2004, 46 (9): 1091-1099
Molecular Cloning and Expression Analysis of a ZincFinger Protein Gene in Apple
CAO Qiu-Fen1*, Masato WADA2, MENG Yu-Ping1, SUN Yi1, CUI Gui-Mei1
(1. Agricultural Biotechnology Research Center of Shanxi Province, Taiyuan 030031, China;2. Department of Apple Research, National Institute of Fruit Tree Science, National Agricultural Research Organization
Shimokuriyagawa, Morioka, Iwate 020-0123, Japan)
Abstract: A cDNA library was created from stem apex tissue from Jonathan apples (Malus domesticaBorkh.), harvested in June to August, during which the plant transitions from vegetative growth toreproductive growth. From this library, we isolated an expressed sequence tag (EST) sequence containinga zinc finger motif; using this sequence, a 779 bp cDNA fragment was obtained by using 5' RACE, and a finalfull-length cDNA encoding an apple zinc finger protein (named MdZF1; GenBank accession number AB116545)was obtained by further PCR. This zinc finger motif of MdZF1 has high homology with INDETERMINATE 1(ID1) gene from maize which seemed to be involved in the transition to flowering. Northern blot andRT-PCR analyses showed that the MdZF1 expressed in the root, stem, leaves, shoot apex and floral organsof the apple, with expression levels higher in root, stem, leaves and floral shoot apex than that in floralorgans (sepals, petals, stamens and pistils). Genomic Southern analysis showed that there was a singlecopy gene in apple genome.Key words: Malus domestica zinc finger protein (MdZF1); cloning; expression
The zinc finger proteins belong to a family of regulatorytranscription factors that contain “finger-like” motifs; theyare found in most living organisms, from yeasts to plantsas well as mammals. The zinc finger motif can bind to DNA,RNA and DNA-RNA hybrids, as well as to other zinc fingerproteins, and zinc finger-containing proteins normally havetwo or more motifs and are responsible for controlling thetranscription and translation in an organism (Huang andLiu, 1998). The classical zinc finger protein is the TFⅢAtype, also known as the Cys2/His2 or C2H2 type, whichhas been extensively studied at a structural level (Sakamotoet al., 2000). In plants, some Cys2/His2 zinc finger cDNAshave been associated with stress resistance (Huang et al.,2002; Wang and Yang, 2002) and growth (Colasanti et al.,1998). Colasanti et al. (1998) cloned the INDETERMI-NATE 1 (ID1) gene, which encoded a maize zinc finger pro-tein associated with the transition to flowering, suggestingthat the ID1 gene product functions as a transcription regula-tor of floral development. The phenotype of id1 mutant andthe expression analysis of ID1 suggested that the gene prod-uct act as a floral initiation signal like the florigen hypothesis.
The shoot apex of higher plants contains an undifferen-tiated cluster of cells, called the shoot apical meristem, whichmaintains the propensity for vegetative growth and
development during the initial stages of plant growth, andthen switches to reproductive growth and developmentwhen external and internal conditions trigger reproductivesignals. To investigate the molecular mechanisms respon-sible for the transition of apple shoot apex from vegetativeto reproductive growth, we built a cDNA library from appleshoot apex at the flower bud during differentiation stage(June to August), and obtained a series of cDNA fragments.By comparing the sequences from database in DDBJ/EMBL/GenBank, we found several expressed sequence tag
(EST)sequences that contained a zinc finger motif. Onesequence showed high homology with maize ID1 gene ,and we were able to assemble a full-length cDNA for thiszinc finger protein with 5′RACE and PCR techniques, whichwe designated MdZF1 (Malus domestica ZINC FINGERPROTEIN 1). Northern blot and RT-PCR methods were usedto confirm and analyze the expression of MdZF1 in differ-ent apple tissues and floral organs. Southern blot was usedto analyze the copy number of MdZF1 in apple genomes.
1 Materials and Methods1.1 Plant materials
Jonathan apple (Malus domestica Borkh.) tissues wereharvested between June and August. Tissue samples

Acta Botanica Sinica 植物学报 Vol.46 No.9 20041092
(0.5-1.0 cm) were taken from the shoot apices every twoweeks, frozen in liquid nitrogen and stored at –80 ℃ forconstruction of the cDNA library. Immature leaves, leaveson the middle of newly-growing shoots, mature leaves, post-germination cotyledons, shoot apices, sepals, petals,stamen, pistils, leaves, roots, and stems from seedlings (onemonth old) were harvested, frozen in liquid nitrogen andstored at –80 ℃ for total RNA extraction, Northern blottingand RT-PCR analysis.1.2 Isolation of the EST and full-length cDNA
Total RNA was extracted from shoot apex tissues ofJonathan apples by a modified CTAB method as previouslydescribed (Cheng et al., 1993), reverse transcribed to cDNAwith Oligo(dT), and ligated with EcoRⅠ, NotⅠ, andBamHⅠ adaptors for constructing the cDNA library. Theligated cDNAs were cloned into the EcoRⅠ restriction siteof pBluescript Ⅱ SK+ vectors (Stratagene, La Jolla,California, USA). The recombinant plasmids were trans-fected into Escherichia coli strain DH5α, and plated onLB/ampiciline/IPTG/X-Gal. White clones were chosen foramplification, followed by plasmid extraction. Forward andreverse sequencing was carried out on a Hitachi SQ5500sequencer (Hitachi, Tokyo, Japan) with a Thermo Sequenasepremixed cycle sequence kit (Amersham Biosciences, NewJersey, USA), followed by analysis with DNA reading soft-ware (Software Development Co., Tokyo, Japan). The se-quences of the cloned cDNA fragments were comparedwith those in the GenBank. A 1.35 kb EST, designated asCS029, and was found containing a zinc finger motif.1.3 5' rapid amplification of cDNA ends (5' RACE)
S p e c i f i c p r i m e r s 5 R C S 2 9 1 ( 5 ' -GAAGATGGTTCCACAGTCAC-3') and 5RCS292 (5'-GCACTTGTCACATTTCCAGG-3') were designed accord-ing to the sequence of the 1.35 kb EST fragment. For RACE,PCR was performed using the above specific primers alongwith cassette primers C1 and C2, and LA-Taq DNA poly-merase (TaKaRa Biomedicals, Shiga, Japan). Primers C1 and5RCS291 were used in the first amplification reaction toamplify fragments from 1 µg template DNA from the cDNAlibrary. The PCR conditions were as follows: 30 cycles of 94℃ for 30 s, 56 ℃ for 30 s, and 72 ℃ for 3 min. Primers C2 and5RCS292 were used for RACE PCR as above, with an an-nealing temperature of 54 ℃.
The PCR products were subcloned into the EcoRⅤ re-striction site of pBluescriptⅡ SK+ vectors, and transformedinto E. coli strain DH5α. The clones with recombinant plas-mids were detected, the plasmids were prepared and se-quenced the same as described above. A 779 bp cDNA,containing sequences of another zinc finger motif upstream
of CS029, was obtained and designated CS029-4.1.4 Full-length cDNA amplification
Sense (5CS294(5'-CTCTTCACCACAATTGGAGG-3'))and antisense ( 3CS29(5'-GTCACCGGAGATCATAACAT-G-3')) primers were designed according to the sequencesof fragments CS029 and CS029-4 and used for PCR amplifi-cation as follows: 95 ℃ for 10 min, followed by 35 cycles of94 ℃ for 30 s, 54 ℃ for 30 s, and 72 ℃ for 3 min. The PCRproducts were confirmed by electrophoresis, and subclonedinto the EcoRⅤ restriction site of the pBluescriptⅡ SK+
vector, a positive clone was picked and sequenced in bothforward and reverse directions. The sequence of the insertwas compared with those in the DNA database (DDBJ/EMBL/GenBank).1.5 Northern blotting analysis
Total RNA was extracted from various tissues ofJonathan apple as described above. A 779 bp digoxigenin(DIG)-labeled RNA probe was prepared with DNA fragmentCS029-4, using the DIG RNA Labeling Mix (BoehringerMannheim, Germany) according to manufacturer’sinstructions. Ten µg of total RNA was denatured at 65 ℃for 10 min, then electrophoresed on 1.0% agarose formal-dehyde-denaturing gels, stained with ethidium bromideandtransferred to a Hybond N+ (Amersham Biosciences, NewJersey, USA). The membrane was hybridized overnight inDIG EASY HYB (Roche Molecular Biochemicals, Germany)solution containing 66 ng/mL of the probe. The hybridiza-tion was following Kotoda et al.(2002). CDP-Star(Boehringer Mannheim, Germany) visualization of the bandswas performed according to the manufacturer’sinstructions, and bands were analyzed with an LAS-1000plus Luminescent Image Analysis System (Fuji Film, Tokyo,Japan).1.6 RT-PCR analysis
Total RNA from various tissues was prepared as de-scribed above. cDNA was synthesized in 20 µL reactionmixtures from 1 µg of total RNA using the RT-PCR HIGHKit (Toyobo Co., Ltd. Osaka, Japan) including 5×reversetranscriptase buffer (4 µL), random primer (1 µL), dNTPs(2 µL), RNase inhibitor (1 µL) and M-MLV reverse tran-scriptase (2 µL). The mixture was incubated at 30 ℃ for 10min, 42 ℃ for 60 min, 99 ℃ for 5 min. PCR was performedusing 1 µg of the reverse transcription product as a template,primers 5CS294 and 5RCS291, and the following amplifica-tion conditions: an initial denaturing at 95 ℃for 12 min,followed by amplification cycles of 94 ℃ for 1 min, 56 ℃ for1 min and 72 ℃ for 2 min. PCR was performed for 30, 35 and40 cycles, respectively. Five µL of the PCR products wereseparated by electrophoresis in a 1.0% agarose gel and

CAO Qiu-Fen et al.: Molecular Cloning and Expression Analysis of a Zinc Finger Protein Gene in Apple 1093
Fig.1. Nucleotide and deduced amino acid sequences of Malus demestica ZINC FINGER PROTEIN 1 (MdZF1) cDNA. The zinc fingermotifs #1 and #2 are underlined. Bold letters represent Cystein (C) and Histidine (H). Primers site for PCR reaction are shown by thearrows. Initiation and stop codons for translation are marked by boxed letters, respectively.

Acta Botanica Sinica 植物学报 Vol.46 No.9 20041094
Fig.1. (continued)

CAO Qiu-Fen et al.: Molecular Cloning and Expression Analysis of a Zinc Finger Protein Gene in Apple 1095
visualized by staining with 1 µg/mL ethidium bromide.1.7 Southern blotting analysis
Genomic DNA was extracted by the method reportedpreviously (Wada et al., 2002). The Southern analysis wascarried out using 779 bp DIG-labeled CS029-4 PCR frag-ment amplified between 5CS294 and 5RCS292 primers(Fig.1). Genomic DNA was digested by restriction enzymeEcoRⅠ, BamHⅠ or HindⅢ, respectively and blotted ontonylon membrane (Hybond N+). Hybridization was performedin DIG EASY HYB solution with 20 ng/mL probe overnight
at 42 ℃, the membrane washing and detection by chemilu-minescence were the same as that in Northern blottinganalysis.
2 Results2.1 Cloning of MdZF1
Based on BLAST searching in the DDBJ/EMBL/GenBank database, the EST sequence from apple shootapex during the transition stage of vegetative to reproduc-tive phase was examined, we identified a novel zinc finger
Fig.1. (continued)

Acta Botanica Sinica 植物学报 Vol.46 No.9 20041096
gene with the 1.35 kb cDNA fragment by random sequenc-ing analysis and BLAST comparison with those in theGenBank. Based on the location of this motif, we hypoth-esized that the putative protein should have another zincfinger motif upstream at the 5' end of the sequence.Accordingly, we used 5' RACE to clone 779 bp fragmentupstream of the original EST and found another zinc fingermotif within it. Based on these sequences, we designedadditional primers to allow PCR amplification of the full-length 2.0 kb cDNA fragment, the sequence of the insertwas compared with those in the DNA database (DDBJ/
EMBL/GenBank) and found to be homologues to those ofzinc finger motifs of Arabidopsis, maize, potato, tomatoand rice, which we designated MdZF1 .2.2 Sequence and structural analysis of MdZF1
The nucleotide and deduced amino acid sequences ofMdZF1 are shown in Fig.1. The full-length cDNA was 1 974bp long, and contained a 246 bp 5'-noncoding region, a 159bp 3'-noncoding region, and an ORF encoding a 522-amino-acid polypeptide that contained a typical Cys2/His2 zincfinger motif (underlined in Fig.1). Database comparisonusing the MdZF1 sequence as bait is shown in Fig.2. The
Fig.2. Alignments between Malus demestica ZINC FINGER PROTEIN 1 (MdZF1) and zinc finger proteins from other crops. TheseAlignments show the comparison of sequences including zinc finger motif #1 and #2 from each plants. Identical amino acids among thesequences are shown in shadows. AT, Arabidopsis thaliana; LE, Lycopersicon esculentum (tomato); MD, Malus domestica (apple); OS,Oryza sativa (ssp. japonica (rice)); ST, Solanum tuberosum (potato); ZM, Zea mays (maize). *,the skipped sequence, 130-155(LVVPS SSAAA GSGGR QQQQQ GEAAP).
CAO Qiu-Fen et al.: Molecular Cloning and Expression Analysis of a Zinc Finger Protein Gene in Apple 1097
C-end and N-end portions of the MdZF1 amino acid se-quence were not homologous to other known plant proteins.However, the MdZF1 zinc finger region exhibited high ho-mology with that of other zinc finger motif genes, includinggenes found in Arabidopsis thaliana (AT) (88.05%identity), rice (OS) (83.55% identity), tomato (LE) (85.43%identity), potato (ST) (84.37% identity), and maize (ZM)(80.50% identity) . Interestingly, when compared to MdZF1,ID1 (ZM) contains an additional 25 amino acids betweenthe first and second zinc finger motif regions.2.3 Expression of the MdZF1 gene
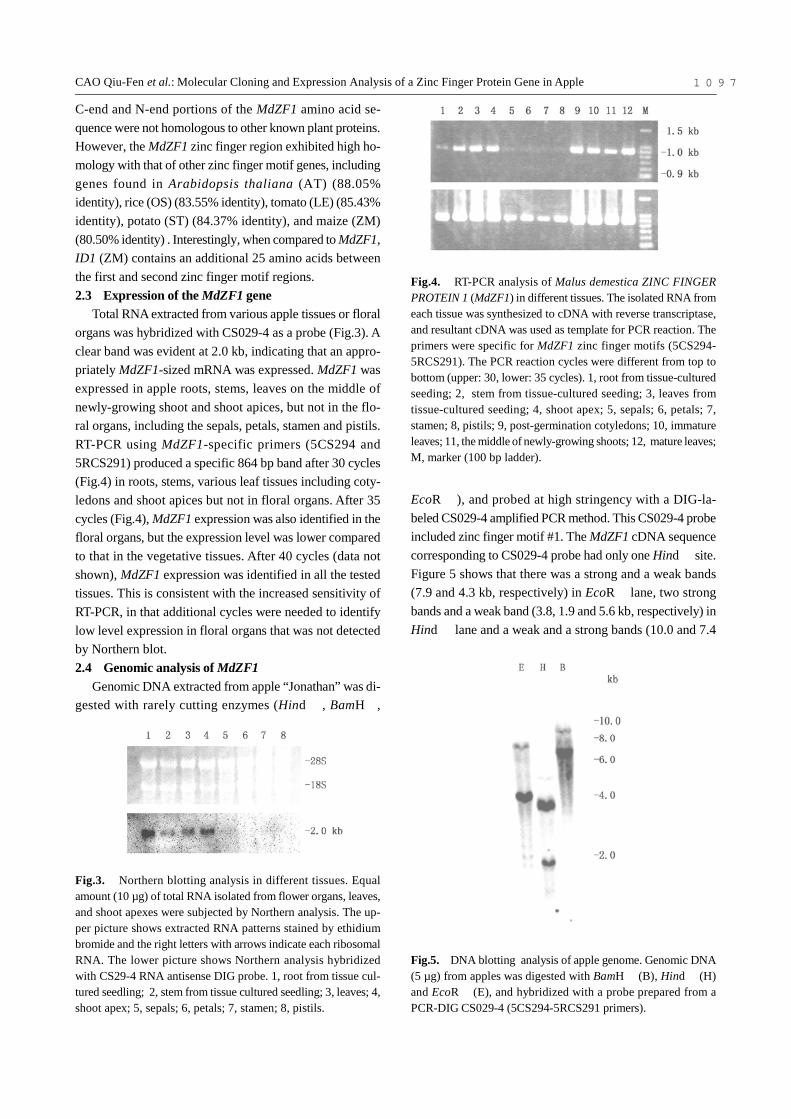
Total RNA extracted from various apple tissues or floralorgans was hybridized with CS029-4 as a probe (Fig.3). Aclear band was evident at 2.0 kb, indicating that an appro-priately MdZF1-sized mRNA was expressed. MdZF1 wasexpressed in apple roots, stems, leaves on the middle ofnewly-growing shoot and shoot apices, but not in the flo-ral organs, including the sepals, petals, stamen and pistils.RT-PCR using MdZF1-specific primers (5CS294 and5RCS291) produced a specific 864 bp band after 30 cycles(Fig.4) in roots, stems, various leaf tissues including coty-ledons and shoot apices but not in floral organs. After 35cycles (Fig.4), MdZF1 expression was also identified in thefloral organs, but the expression level was lower comparedto that in the vegetative tissues. After 40 cycles (data notshown), MdZF1 expression was identified in all the testedtissues. This is consistent with the increased sensitivity ofRT-PCR, in that additional cycles were needed to identifylow level expression in floral organs that was not detectedby Northern blot.2.4 Genomic analysis of MdZF1
Genomic DNA extracted from apple “Jonathan” was di-gested with rarely cutting enzymes (Hind Ⅲ, BamHⅠ,
EcoRⅠ ), and probed at high stringency with a DIG-la-beled CS029-4 amplified PCR method. This CS029-4 probeincluded zinc finger motif #1. The MdZF1 cDNA sequencecorresponding to CS029-4 probe had only one HindⅢ site.Figure 5 shows that there was a strong and a weak bands(7.9 and 4.3 kb, respectively) in EcoRⅠ lane, two strongbands and a weak band (3.8, 1.9 and 5.6 kb, respectively) inHindⅢ lane and a weak and a strong bands (10.0 and 7.4
Fig.3. Northern blotting analysis in different tissues. Equalamount (10 µg) of total RNA isolated from flower organs, leaves,and shoot apexes were subjected by Northern analysis. The up-per picture shows extracted RNA patterns stained by ethidiumbromide and the right letters with arrows indicate each ribosomalRNA. The lower picture shows Northern analysis hybridizedwith CS29-4 RNA antisense DIG probe. 1, root from tissue cul-tured seedling; 2, stem from tissue cultured seedling; 3, leaves; 4,shoot apex; 5, sepals; 6, petals; 7, stamen; 8, pistils.
Fig.4. RT-PCR analysis of Malus demestica ZINC FINGERPROTEIN 1 (MdZF1) in different tissues. The isolated RNA fromeach tissue was synthesized to cDNA with reverse transcriptase,and resultant cDNA was used as template for PCR reaction. Theprimers were specific for MdZF1 zinc finger motifs (5CS294-5RCS291). The PCR reaction cycles were different from top tobottom (upper: 30, lower: 35 cycles). 1, root from tissue-culturedseeding; 2, stem from tissue-cultured seeding; 3, leaves fromtissue-cultured seeding; 4, shoot apex; 5, sepals; 6, petals; 7,stamen; 8, pistils; 9, post-germination cotyledons; 10, immatureleaves; 11, the middle of newly-growing shoots; 12, mature leaves;M, marker (100 bp ladder).
Fig.5. DNA blotting analysis of apple genome. Genomic DNA(5 µg) from apples was digested with BamH Ⅰ(B), Hind Ⅲ(H)and EcoR Ⅰ(E), and hybridized with a probe prepared from aPCR-DIG CS029-4 (5CS294-5RCS291 primers).

Acta Botanica Sinica 植物学报 Vol.46 No.9 20041098
kb, respectively) in BamHⅠ lane. The results indicatedthat the MdZF1was a single copy gene and may have an-other slightly homologous one.
3 DiscussionZinc finger proteins participate in numerous physiologi-
cal activities in plants, including stress resistance and vari-ous developmental processes (Sakamoto et al., 2000 ) . TheArabidopsis thaliana STZ (salt tolerance zinc finger protein)gene, which encodes a zinc finger protein, was expressedin a salt-sensitive yeast strain and led to increased salttolerance, indicating that the zinc finger protein might di-rectly participate in activating the expression of salt-toler-ance genes (Lippuner et al., 1996). Similarly, typical C2H2zinc finger proteins, as well as salt-tolerance-related geneshave been isolated from cotton and rice (Huang et al., 2002;Wang et al., 2002). Some other zinc finger proteins playimportant regulatory roles in plant growth.
We obtained a 2 kb cDNA fragment MdZF1, from a cDNAlibrary of apple stem apex tissue. It had two typical zincfinger motifs. The zinc finger structural regions and thespacer region between them had above 80% homology withdicotyledonous Arabidopsis and potato, and monocotyle-donous rice and maize, implying that they were evolvedfrom one ancestral gene. The maize ID1 and ID1-like gene“P1” genes reported by Colasanti (1998) shared highly ho-mologous zinc finger motifs with MdZF1 gene identified inthis study; these motifs were of the C2-H2 and C2-HCpatterns. The addition of 25 amino acids between the twozinc finger regions in ID1, compared with those in otherplants, indicated that distinctive regions have been evolvedin the homologous genes in each species. ID1 mRNA wasexpressed in immature leaves, but not in the shoot apex andfloral organs. ID1-deficient maize is unable to flowernormally; the mutant maintains a prolonged vegetativegrowth period, and may produce floral inflorescences withvegetative growth characteristics. These observations sug-gest that ID1 products play as a transcription regulator forthe floral initiation or transport of various substances tothe shoot apex. The P1 gene, which is similar in structure toID1, was expressed in both mature and immature leavesand shoot apex. The MdZF1 gene identified in this studyhas zinc finger motifs similar to those encoded by the ID1,P1 and AT genes from maize and Arabidopsis thaliana, aswell as the PCP1 gene from Solanum tuberosum (Kuhnand Frommer, 1995); however, MdZF1 was expressed inroots, stems, cotyledons, young leaves, mature leaves,shoot apex, and various leaf tissues (leaves on middle ofnewly growing shoot and leaves from seedlings). In
contrast, the ID1 and P1 genes were expressed in leaves,shoot apices and stems, but not in roots. MdZF1 was notdetected in floral organs by Northern blotting analysis, butafter the RT-PCR cycles were increased, MdZF1 expressionwas detected in them. Thus, the result implied that MdZF1is expressed at very low levels in floral organs, and it mainlyfunctions in roots, stems, leaves and stem apex.
The genomic Southern analysis with CS029-4 probe,which contained the zinc finger motif #1, showed MdZF1was a single copy gene and might be another slight ho-mologous gene in Jonathan genome (Fig.5). We had al-ready checked “Fuji” genome with the same probe, the re-sultant patterns were the same (data not shown). If the zincfinger motif binds the specific DNA sites, the MdZF1 andID1 showed a similar regulatory manner because their zincfinger parts showed high homology (Fig.2). In Zea mays,the id1 mutant had apparent phenotypic effects on flowertransition. It means clearly that the ID1 gene plays a keyrole in flower development. Then the MdZF1 from applewas also expected the similar function for flowering. Wehave made MdZF1-transformed Arabidopsis, which ex-pressed excess MDZF1 under 35S promoter. But the result-ant transformants were never distinguished from wild typeplants in flower formation or flowering time on photoperi-ods (data not shown). It indicated that apple MdZF1 couldnot be involved in flowering events of Arabidopsis. Forfurther research, the complementation experiment of Z. maysid1 mutant with apple MdZF1 will be needed for investiga-tion of the physiological role.
Based on the expression patterns of MdZF1 and theanalysis of ID1 and P1 functions, we propose that MdZF1may be an apple growth-related regulatory gene. With re-spect to the study of regulatory genes involved in appleflowering, we have already isolated two homologues ofFLORICAULA/LEAFY, AFL1 and AFL2, from apple treeflower buds (Wada et al., 2002). Kotoda et al. (2002) alsoobtained the AP1-homogenous gene MdMADS5 fromapple. The function of MdZF1 and its relationship withthese flowering regulatory genes are needed to investigatein future work.
References:
Cheng S, Puryear J, Cairney J. 1993. A simple and efficient method
for isolation RNA from pine trees. Plant Mol Biol Rep, 11:
113-116.
Colasanti J, Yuan Z, Sunderesan V. 1998. The indeterminate gene
encodes a zinc finger protein and regulates a leaf-generated
signal required for the transition to flowering in maize. Cell,
93: 593-603.

CAO Qiu-Fen et al.: Molecular Cloning and Expression Analysis of a Zinc Finger Protein Gene in Apple 1099
Huang Z-X, Liu Y-Q. 1998. Progress in zinc finger protein studies.
Chem Chin Univ, 19: 1904-1914. (in Chinese with English
abstract)
Huang J , Zhang H-S, Cao Y-J, Wang D, Wang J-F, Yang J-S. 2002.
Cloning and sequence analysis of a new novel C2H2 zinc
finger cDNA from rice (Oryza sativa L.). J Nanjing Agr Univ,
25: 110-112. (in Chinese with English abstract)
Kuhn C, Frommer W. 1995. A novel zinc finger protein encoded
by a couch potato homologue from Solanum tuberosum en-
ables a sucrose transport-deficient yeast strain to grow on
sucrose. Mol Gen Genet, 247: 759-763.
Kotoda K, Wada M, Kusaba S, Kano-Murakami Y, Masuda T,
Soejima J. 2002. Overexpression of MdMADS5, an APETALA1-
like gene of apple, causes early flowering in transgenic
Arabidopsis. Plant Sci, 162: 679-687.
Lippuner V, Martha S Cyert, Charles S Gasser. 1996. Two classes(Managing editor: ZHAO Li-Hui)
of plant cDNA clones differentially complement yeast
calcineurin mutants and increase salt tolerance of wild-type
yeast. J Biol Chem, 271: 12859-12866.
Sakamoto H, Araki T, Meshi T, Iwabuti M. 2000. Expression of
a subset of the Arabidopsis Cys2/His2-type zinc-finger pro-
tein gene family under water stress. Gene, 248: 23-32.
Wada M, Cao Q F, Kotoda N, Soejima N, Masuda T. 2002.
Apple has two orthologues of FLORICAULA/LEAFY in-
volved in flowering. Plant Mol Biol, 49: 567-577.
Wang D, Yang J-S . 2002.Cloning and characterization of cDNA
encoding cotton STZ-like protein. J Fudan Univ (Nat Sci) ,
41: 42-45. (in Chinese with English abstract)


















