
Modulation of nitrate reductase activity in rice seedlings under aluminium toxicity and water...
11
Journal of Plant Physiology 162 (2005) 854—864 Modulation of nitrate reductase activity in rice seedlings under aluminium toxicity and water stress: role of osmolytes as enzyme protectant Pallavi Sharma, Rama Shanker Dubey Department of Biochemistry, Faculty of Science, Banaras Hindu University, Varanasi 221005, India Received 5 August 2004; accepted 24 September 2004 Summary Nitrate reductase (NR) activity in the presence of Mg 2+ (NR act) representing the non- phosphorylated NR state and the activity in the presence of EDTA (NR max) representing maximum NR activity was measured in roots and shoots of 15 d grown aluminium and water stressed rice seedlings to examine changes in NR activation state due to these stresses. Seedlings subjected to a moderate water stress level of 0.5 MPa for 24 h or grown in presence of 80 mM Al 3+ showed decreased level of NR max but resulted in higher NR act and NR activation state. However, seedlings grown in presence of a higher level of 160 mM Al 3+ showed a decline in NR act as well as NR max. With a higher water stress level of 2.0 MPa a marked decline in the levels of both NR act and NR max was observed, whereas NR activation state remained almost unaltered with severe water stress. NR activity appeared to be sensitive to H 2 O 2 , PEG-6000, NaCl and various metal salts. Incorporation of these components in the enzyme assay medium led to decreased affinity of enzyme towards its substrate with increase in K m and decrease in V max values. Addition of each of the osmolytes i.e. 1 mol/L proline, 1 mol/L glycine betaine or 1 mol/L sucrose in the enzyme assay medium caused a considerable protection to the enzyme against the damaging effects of stressful components. An enhanced level of proline and glycine betaine was observed in Al- stressed seedlings and sucrose in Al as well as water stressed seedlings. Results suggest that Al toxicity and water stress decrease total amount of functional NR in rice seedlings and the osmolytes proline, glycine betaine and sucrose appear to have a direct protective action on enzyme NR under stressful conditions & 2005 Elsevier GmbH. All rights reserved. ARTICLE IN PRESS www.elsevier.de/jplph KEYWORDS Aluminium; Metal toxicity; NaCl salinity; Nitrate reductase; Oryza sativa; Osmolytes; Water stress 0176-1617/$ - see front matter & 2005 Elsevier GmbH. All rights reserved. doi:10.1016/j.jplph.2004.09.011 Abbreviations: Non-pp NR, non-phosphorylated nitrate reductase; NR, nitrate reductase; NR act, actual NR activity; NR max, maximum NR activity; PEG-6000, polyethylene glycol-6000 Corresponding author. Tel.: +91-542-2317190; fax: +91-542-2368174. E-mail address: [email protected] (R. Shanker Dubey).
-
Upload
pallavi-sharma -
Category
Documents
-
view
213 -
download
0
Transcript of Modulation of nitrate reductase activity in rice seedlings under aluminium toxicity and water...

ARTICLE IN PRESS
Journal of Plant Physiology 162 (2005) 854—864
KEYWORDAluminiumMetal toxiNaCl salinNitrate reOryza satiOsmolytesWater stre
0176-1617/$ - sdoi:10.1016/j.
Abbreviationmaximum NR a�CorrespondE-mail addr
www.elsevier.de/jplph
Modulation of nitrate reductase activity in riceseedlings under aluminium toxicity and waterstress: role of osmolytes as enzyme protectant
Pallavi Sharma, Rama Shanker Dubey�
Department of Biochemistry, Faculty of Science, Banaras Hindu University, Varanasi 221005, India
Received 5 August 2004; accepted 24 September 2004
S;city;ity;ductase;va;;ss
ee front matter & 200jplph.2004.09.011
s: Non-pp NR, non-pctivity; PEG-6000, poing author. Tel.: +91-5ess: [email protected] (R
SummaryNitrate reductase (NR) activity in the presence of Mg2+ (NR act) representing the non-phosphorylated NR state and the activity in the presence of EDTA (NR max)representing maximum NR activity was measured in roots and shoots of 15 d grownaluminium and water stressed rice seedlings to examine changes in NR activation statedue to these stresses. Seedlings subjected to a moderate water stress level of�0.5MPa for 24 h or grown in presence of 80 mM Al3+showed decreased level of NR maxbut resulted in higher NR act and NR activation state. However, seedlings grown inpresence of a higher level of 160 mM Al3+ showed a decline in NR act as well as NR max.With a higher water stress level of �2.0MPa a marked decline in the levels of both NRact and NR max was observed, whereas NR activation state remained almost unalteredwith severe water stress. NR activity appeared to be sensitive to H2O2, PEG-6000, NaCland various metal salts. Incorporation of these components in the enzyme assaymedium led to decreased affinity of enzyme towards its substrate with increase in Kmand decrease in Vmax values. Addition of each of the osmolytes i.e. 1mol/L proline,1mol/L glycine betaine or 1mol/L sucrose in the enzyme assay medium caused aconsiderable protection to the enzyme against the damaging effects of stressfulcomponents. An enhanced level of proline and glycine betaine was observed in Al-stressed seedlings and sucrose in Al as well as water stressed seedlings. Results suggestthat Al toxicity and water stress decrease total amount of functional NR in riceseedlings and the osmolytes proline, glycine betaine and sucrose appear to have adirect protective action on enzyme NR under stressful conditions& 2005 Elsevier GmbH. All rights reserved.
5 Elsevier GmbH. All rights reserved.
hosphorylated nitrate reductase; NR, nitrate reductase; NR act, actual NR activity; NR max,lyethylene glycol-600042-2317190; fax: +91-542-2368174.. Shanker Dubey).

ARTICLE IN PRESS
Modulation of nitrate reductase under stresses 855
Introduction
Nitrate reductase (NR; EC 1.6.6.1), which cata-lyzes the first enzymatic step in nitrate assimilationin higher plants, involving reduction of nitrate tonitrite, is a highly regulated enzyme. Severalmetabolic and environmental signals regulate NRactivity (Kaiser et al., 1999). At transcriptionallevel NR activity is regulated by the availability ofsubstrate NO3
� and by the end product of the Nassimilation pathway, glutamine. NR activity is alsoregulated post translationally by a phosphoryla-tion–dephosphorylation mechanism. The phos-phorylated and non-phosphorylated NR (non-ppNR) proteins are equally active, but phosphoryla-tion sensitizes the enzyme to inhibition by aninhibitory 14-3-3 protein in the presence of Mg2+
(McKintosh et al., 1995). It has been establishedthat these post-translational changes of NR activityare responsible for plant adaptation to fluctuatingenvironmental conditions (Botrel et al., 1996).Several factors such as nitrate, glutamate andsugar have been shown to modulate the post-translational regulation of NR. Actual NR activity(NR act) determined in tissue extracts in presenceof Mg2+ represents activity due to non-pp NR, whereas the activity measured in presence of EDTArepresents maximum NR capacity (NR max) or totalamount of functional NR in the tissue extract(Kaiser et al., 2000). The activation state of NRthus reflects the percentage of non-pp NR anddetermines rate of nitrate reduction in situ (Kaiseret al., 2000). Due to the central role of NR in plantprimary metabolism and alteration in its activityunder stressful conditions, this enzyme has beenthe focus of study under various harsh environ-mental conditions that affect plant growth andmetabolism (Oji et al., 1988).
Accumulation of compatible solutes is one of theadaptive strategies of plants in response to abioticenvironmental stresses (Wang et al., 1999). Com-patible solutes are low molecular mass substanceswhich accumulate in high amounts and do notinterfere with cellular metabolism (Hagemann andMarin, 1999). Accumulation of these solutes likeproline, glycine betaine and sucrose contribute toosmotic adjustment, prevention of protein dena-turation, preservation of enzyme structure andactivity and protection of membranes from damageby reactive oxygen species (ROS) (Hare et al.,1999). Many osmolytes stabilize protein in vivo andin vitro (Saunders et al., 2000; Sharma and Dubey,2004). Two classes of model have been proposed toexplain the effects of osmolytes on protein stability(Saunders et al., 2000). The first class focuses onthe binding between osmolytes and protein while
the other class focuses on the excluded volumeeffects arising from the increased steric repulsionbetween osmolytes and protein. The actual me-chanism however, has not been properly workedout and could be a combination of two classes.Through natural selection nature has used osmo-lytes to advantage in generically stabilizing proteinagainst denaturing stresses (Saunders et al., 2000).
Rice is an important food crop of the worldpopulation and is often subjected to a variety ofharsh environmental conditions (Dubey and Pessar-akli, 2002). The aim of the present investigationwas to study the effect of in situ Al stress and invitro water stress on NR act, NR max as well as NRactivation state in rice seedlings. In vitro effects ofH2O2, water stress, salinity stress and various metalsalts on the activity of NR were examined. Furtherthe content and putative roles of osmolytes pro-line, glycine betaine and sucrose in protecting NRactivity under various abiotic stresses were deter-mined.
Materials and methods
Plant material and stress conditions
Seedlings of two rice (Oryza sativa L.) cultivarsPant-12 and Malviya-36 were raised in sand culturesusing Hoagland nutrient solution (Hoagland andArnon, 1938) as described earlier (Richharia et al.,1997) and were maintained for 20 d in a greenhouse at a temperature of 2871 1C, relativehumidity of 80%, 12 h light/dark photoperiod andirradiance of 40–50mmol/m2 s. Pots were kept atfield saturation capacity by irrigation. To observe Altoxicity effects on NR act, NR max, NR activationstate and osmolytes content, sand cultures weremaintained at pH 4.5. Hoagland nutrient solutionserved as control and nutrient solution supplemen-ted with 1 and 5mM Al2(SO4)3 served as treatmentsolutions. As the amount of available toxic Alspecies is related to the ionic strength of thenutritive solution the actual levels of toxic Al3+ inthe treatment solution were determined usingaluminon reagent (Hsu, 1963). Treatment solutioncontaining 1 and 5mM Al2(SO4)3 thus correspondedto 80 and 160 mM of active Al3+ respectively.Seedlings were uprooted at 5 d intervals up to20 d and NR act, NR max, NR activation states andcontents of osmolytes were determined in rootsand shoots of growing seedlings. To study waterstress induced changes in NR act, NR max, NRactivation states and osmolytes contents, 10, 15and 20 d grown seedlings were subjected to in vitro

ARTICLE IN PRESS
P. Sharma, R. Shanker Dubey856
water stress for 24 h (Paleg et al., 1984) by placingtwo or three uprooted seedlings in aerated testtubes containing Hoagland nutrient solution whichserved as control and polyethylene glycol (PEG-6000) solutions of 17% and 41.2% concentrations toserve as water stress levels of �0.5 and �2.0MPa,respectively.
Nitrate reductase activity
The activity of NR was assayed following themethod of Botrel et al. (1996) with some modifica-tions. About 1 g root/shoot samples were homo-genized using chilled mortar and pestle in 3mL ofextraction buffer containing 50mM HEPES-KOH (pH7.5), 10mM MgCl2, 10 mM FAD, 1mM DTT, 1% (w/v)insoluble polyvinyl pyrrolidone (PVP), 0.05% (w/v)casein and 1% Triton X-100. Contents were cen-trifuged at 22,000g for 10min at 4 1C. The clearsupernatant obtained was dialyzed in cellophanemembranes in cold for 3–4 h against the extractionbuffer and the desalted enzyme extract was addedto an incubation mixture containing 50mM HEPES-KOH (pH 7.5), 10mM MgCl2, 10 mM FAD, 1mM DTT(for determination of NR act) or preincubated at20 1C for 15min with 50mM HEPES-KOH (pH 7.5),10mM MgCl2, 10 mM FAD, 1mM DTT and 15mM EDTA(for determination of NR max). The reaction wasstarted by the addition of 10mM KNO3 and 0.2mMNADH and incubated at 30 1C for 30min and thenstopped by adding 0.1M Zn-acetate. After 15min,nitrite produced was measured colorimetrically at540 nm by adding 2mL of colour developing reagent(1% sulphanilamide and 0.2% N-(1-naphthyl) ethy-lene diamine dihydrochloride (NEDH) each pre-pared in 1.5mol/L HCl and mixed in equalvolumes). Protein was estimated by the methodof Bradford (1976) using bovine serum albumin(BSA, Sigma) as standard.
Effect of H2O2, PEG-6000, NaCl and metalsalts in vitro on NR activity
Reaction mixture in addition to the normalingredients of the assay medium contained increas-ing concentrations of H2O2 (0–50mM), NaCl(0–600mM) PEG-6000 (0–80%) and metal salts(0–4mM). To observe the possible protective rolesof osmolytes proline, glycine betaine and sucroseon NR activity, 1mol/L of each osmolyte was addedto the reaction mixture in which either H2O2 levelof 1mM, NaCl level of 300mM, PEG level of 40% ormetal concentration of 2mM was maintained.Contents were kept at 40 1C for 45min and there-after NR activities were assayed. Value of Michaelis
constant (Km) and Vmax for NR under all conditionswere determined with respect to nitrate byconstructing Lineweaver–Burk (L–B) plots.
Proline, glycine betaine and sucroseestimation
Proline was estimated in roots and shoots of theseedlings according to the methods of Bates et al.(1973). Fresh samples (500mg) were homogenizedin 10mL of 3% aqueous sulfosalicylic acid andcentrifuged at 22,000g for 5min. To 2mL of thesupernatant 2mL of acid ninhydrin was added.Further, 2mL of glacial acetic acid was added andthe contents were boiled for 1 h at 100 1C in a waterbath. The mixture was further extracted with 10mLof toluene by mixing the two thoroughly in a testtube with vigorous stirring. Absorption of chromo-phore was read at 515 nm in an ELICO CL-24spectrophotometer (Hyderabad, India). L-Proline(Sigma) was used for the preparation of standardcurve. The amount of proline in the sample wascalculated in mg (proline)/g fresh weight. Forglycine betaine estimation roots and shoots sam-ples were oven dried at 80 1C for 4 d. About 500mgdried samples were ground to fine powder andshaken at 25 1C for 24 h with 20mL of water.Contents were filtered and stored in the freezeruntil the analysis was done. Aliquots containing0.5mL of thawed extract were diluted to 1:1 with0.2 N H2SO4 and cooled in ice water for 1 h. Further0.2mL cold KI–I2 reagent (prepared by dissolving15.7 g of iodine and 20.0 g of KI in 100mL water)was added and stirred gently. This was stored at0–4 1C overnight (16 h) and then centrifuged nextday at 16,000 rpm for 15min and supernatant wasaspirated with a fine tipped glass tube. Throughoutthe process cold conditions were maintainedbecause the solubility of complex in the acidreaction mixture increases markedly with tempera-ture. It is important that tubes be kept cold untilthe periodide complex is separated from themedia. Periodide crystals were separated fromthe acid media and then dissolved in 9mL of 1,2-dichloroethane. Vigorous shaking was done forcomplete solubilization of crystals in the develop-ing solvent. After 2 h absorbance was recorded at365 nm. To prepare standard curve glycine betaineobtained from Sigma was used in the range of50–200 mg/mL. Standard curve was prepared in 1NH2SO4. Amount of glycine betaine was expressed asmg/g dry weight of samples (Grattan and Grieve,1985). Sucrose was estimated in roots and shootsusing anthrone reagent (Van Handel, 1968). Root/shoot samples were oven dried at 70 1C for 24 h.
ARTICLE IN PRESS
Modulation of nitrate reductase under stresses 857
About 100mg dried sample was homogenized in10mL 80% ethanol. Hexoses were destroyed byplacing the reaction tubes in boiling water for10min. After cooling 0.1mL 30% aqueous KOH wasadded and kept at 100 1C for 10min. When thetubes were cooled to room temperature, 3mLanthrone reagent (prepared by mixing 76mL ofconcentrated H2SO4, 36mL water and 150mg ofanthrone) was added and incubated at 38 1Cfor 20min and absorbance was recorded at620 nm in a Bausch and Lomb spectrophotometer.Sucrose served as standard. Amount of sucrosewas expressed in terms of mg/g dry weight ofsamples.
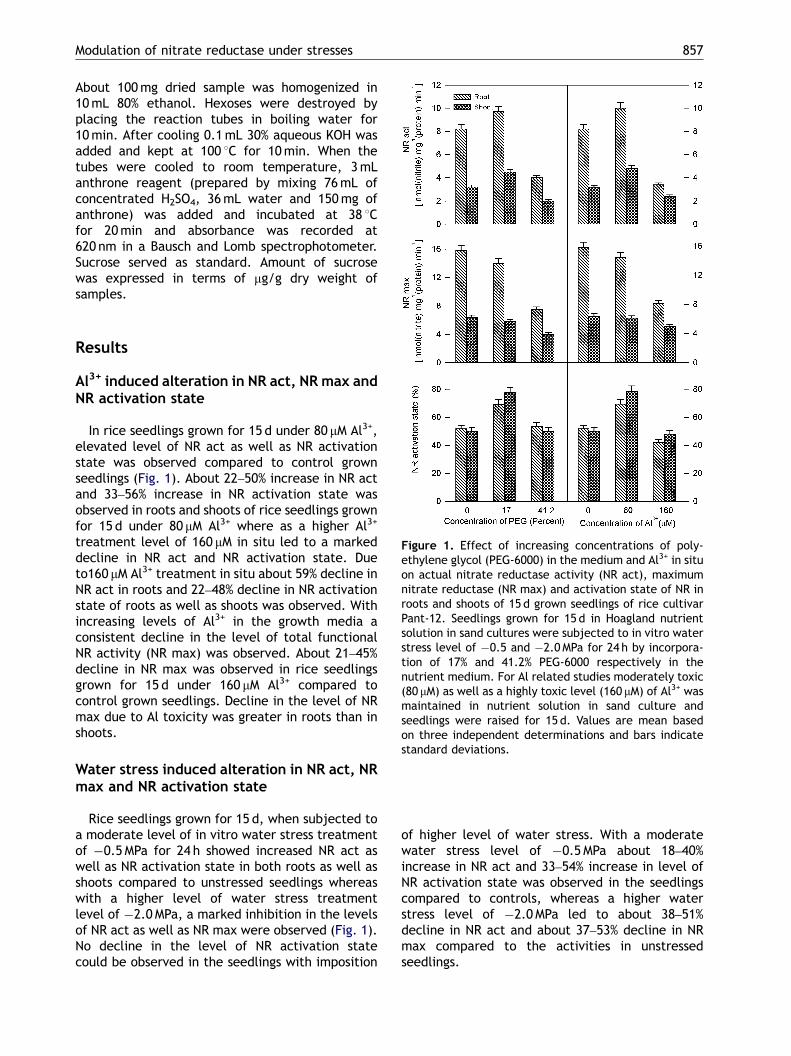
Figure 1. Effect of increasing concentrations of poly-ethylene glycol (PEG-6000) in the medium and Al3+ in situon actual nitrate reductase activity (NR act), maximumnitrate reductase (NR max) and activation state of NR inroots and shoots of 15 d grown seedlings of rice cultivarPant-12. Seedlings grown for 15 d in Hoagland nutrientsolution in sand cultures were subjected to in vitro waterstress level of �0.5 and �2.0MPa for 24 h by incorpora-tion of 17% and 41.2% PEG-6000 respectively in thenutrient medium. For Al related studies moderately toxic(80 mM) as well as a highly toxic level (160 mM) of Al3+ wasmaintained in nutrient solution in sand culture andseedlings were raised for 15 d. Values are mean basedon three independent determinations and bars indicatestandard deviations.
Results
Al3+ induced alteration in NR act, NR max andNR activation state
In rice seedlings grown for 15 d under 80 mM Al3+,elevated level of NR act as well as NR activationstate was observed compared to control grownseedlings (Fig. 1). About 22–50% increase in NR actand 33–56% increase in NR activation state wasobserved in roots and shoots of rice seedlings grownfor 15 d under 80 mM Al3+ where as a higher Al3+
treatment level of 160 mM in situ led to a markeddecline in NR act and NR activation state. Dueto160 mM Al3+ treatment in situ about 59% decline inNR act in roots and 22–48% decline in NR activationstate of roots as well as shoots was observed. Withincreasing levels of Al3+ in the growth media aconsistent decline in the level of total functionalNR activity (NR max) was observed. About 21–45%decline in NR max was observed in rice seedlingsgrown for 15 d under 160 mM Al3+ compared tocontrol grown seedlings. Decline in the level of NRmax due to Al toxicity was greater in roots than inshoots.
Water stress induced alteration in NR act, NRmax and NR activation state
Rice seedlings grown for 15 d, when subjected toa moderate level of in vitro water stress treatmentof �0.5MPa for 24 h showed increased NR act aswell as NR activation state in both roots as well asshoots compared to unstressed seedlings whereaswith a higher level of water stress treatmentlevel of �2.0MPa, a marked inhibition in the levelsof NR act as well as NR max were observed (Fig. 1).No decline in the level of NR activation statecould be observed in the seedlings with imposition
of higher level of water stress. With a moderatewater stress level of �0.5MPa about 18–40%increase in NR act and 33–54% increase in level ofNR activation state was observed in the seedlingscompared to controls, whereas a higher waterstress level of �2.0MPa led to about 38–51%decline in NR act and about 37–53% decline in NRmax compared to the activities in unstressedseedlings.

ARTICLE IN PRESS
P. Sharma, R. Shanker Dubey858
Effect of H2O2, PEG-6000 and NaCl in vitro onNR activity
The effects of increasing concentrations of H2O2,PEG-6000 and NaCl were studied in vitro on theactivity of NR (Fig. 2). It was observed that H2O2 ata lower concentration of 50 mM in the reactionmedium led to increased activity of NR comparedto controls, whereas in a concentration beyond50 mM H2O2 caused gradual inhibition in NR activity.With 1mM H2O2 in the assay medium about 40%inhibition in enzyme activity was observed. Addi-tion of 1mol/L proline, glycine betaine or sucrosein the enzyme assay medium led to considerable
Figure 2. Effect of increasing concentrations of (a) H2O2,(b) PEG-6000 and (c) NaCl in the enzyme assay medium inpresence of 1M of the osmolytes proline, glycine betaineand sucrose on the activity of NR. Values are mean basedon three independent determinations and bars indicatestandard deviations.
protection of the activities of NR, in presence ofH2O2. Similarly PEG-6000 in a concentration beyond20% and NaCl beyond 50mM in the reaction mediumled to significant decline in NR activity. Incorpora-tion of proline, glycine betaine or sucrose in1mol/L concentration in the assay mediumprovided considerable protection to the enzymeagainst inhibition due to PEG-6000 or NaCl. Prolineappeared to be a good protectant for NR followedby glycine betaine and sucrose against the inhibi-tory effects of H2O2, PEG-6000 or NaCl.
Effect of metal salts in vitro on NR activity
Effects of increasing concentration of metal saltsoften present as contaminants in polluted soils likeAl2(SO4)3, Al(NO3)3, Na2HAsO4, As2O3, NiSO4, NiCO3,Pb(CH3COO)2, CdCl2, Cd(NO3)2 were studied in vitroon the activity of NR (Fig. 3). The results indicatedinhibition in the activity of NR even with 500 mM
Figure 3. Effect of increasing concentration of metalsalts Al(NO3)3, Al2(SO4)3, Na2HAsO4, As2O3, Ni(NO3)2,NiSO4, CdCl2, Cd(CH3COO)2, Pb(NO3)2, and Pb(CH3COO)2in vitro along with 1M of each of the osmolytes proline,glycine betaine and sucrose on the activity of NR. Valuesare mean based on three independent determinationsand bars indicate standard deviations.

ARTICLE IN PRESS
Modulation of nitrate reductase under stresses 859
concentrations of metal salts in the assay medium.All metal salts studied led to about 34–59%inhibition in NR activity in 1.5mM concentrationsand about 53–76% inhibition in 4mM concentration.Among metal salts examined As2O3 was most toxicto NR. With 4mM As2O3 in vitro about 76% inhibitionin the activity of NR was observed. Addition of eachof the osmolytes, i.e. 1mol/L proline, 1mol/Lglycine betaine or 1mol/L sucrose in the enzymeassay medium caused a considerable protection tothe enzyme against the damaging effects of metalsalts.
Michaelis constants (Km) of NR with respectto nitrate in presence of H2O2, PEG-6000,NaCl and metal salts in the assay medium
When typical Lineweaver and Burk (L–B) plotswere constructed for determination of Km and Vmax
values of NR with respect to nitrate as substrate,the Km values obtained was 0.7mmol/L KNO3 and4.0 nmol/minmg protein, respectively. Incorpora-tion of 1mM H2O2 in the enzyme assay mediumalmost doubled the enzyme Km value to 1.42mmol/L KNO3 and reduced the Vmax value to 2.2 nmol/minmg protein. But when the compounds proline,glycine betaine or sucrose were incorporated in thereaction medium in 1M concentrations a significantdecrease in Km values compared to Km in presenceof H2O2 were observed (Table 1). Similarly when thecompounds 40% polyethylene glycol, 300mM NaClor 2mM salts of the metals Al, As, Ni, Pb, Cd wereincorporated in the enzyme assay medium andenzyme activities were determined, a significantincrease in Km (1.10–2.04mmol/L KNO3) anddecrease in Vmax (2.5–1.8 nmol/minmg protein)values were obtained compared to Km(0.70mmol/L KNO3) and Vmax (4.0 nmol/min -mg protein) of the enzyme determined in theabsence of these compounds. PEG, NaCl or thestudied metal salts significantly decrease theaffinity of NR towards its substrate NO3
� by possiblydenaturing/inactivating the enzyme but the addi-
Table 1. Effect of the osmoprotectants proline, glycine betconstants (Km) and Vmax of NR extracted from shoots of 15 d
Treatments Km
Enzyme 0.7Enzyme+1mM H2O2 1.4Enzyme+1mM H2O2+1mol/L Proline 0.8Enzyme+1mM H2O2+1mol/L Glycine betaine 1.0Enzyme+1mM H2O2+1mol/L Sucrose 1.1
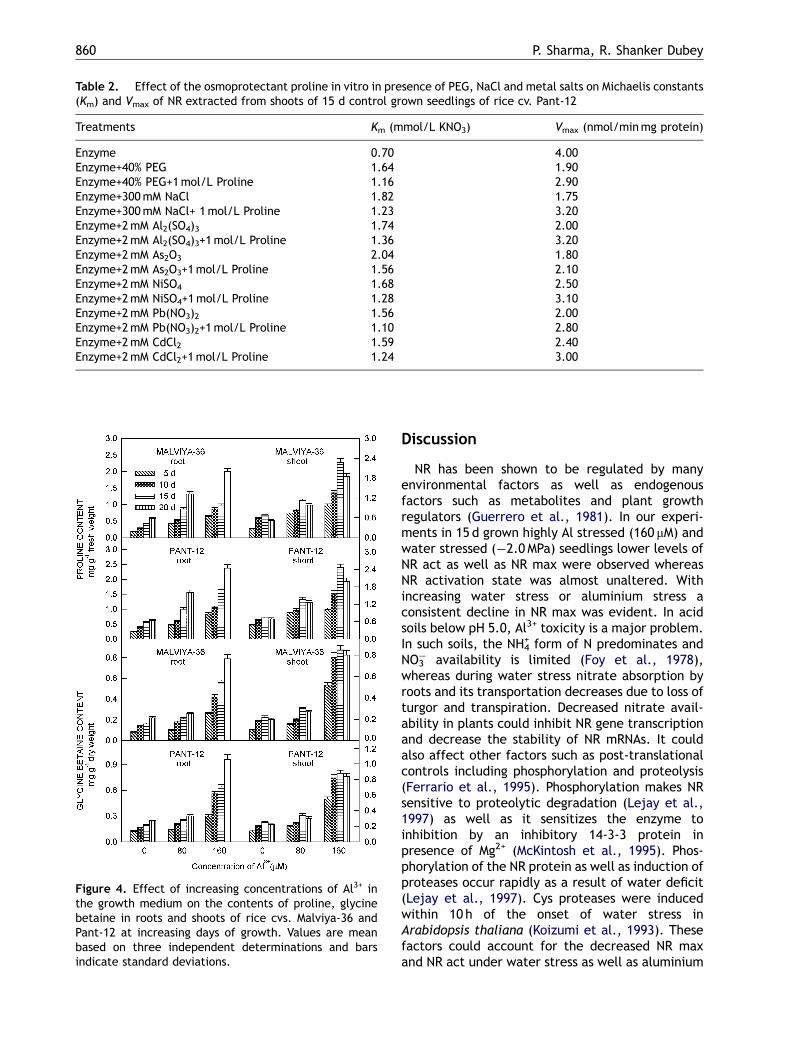
tion of 1M proline led to a significant decrease inKm values and increase in Vmax values therebymaintaining adequate rates of enzymatic catalysis(Table 2).
Effects of aluminium toxicity in situ oncontents of proline, glycine betaine andsucrose in rice seedlings
Experiments were conducted to examine thelevels of the osmolytes proline, glycine betaine andsucrose in the rice seedlings growing in presence of80 and 160 mM Al3+ in sand culture experiments. Inboth the rice cultivars Malviya-36 and Pant-12increasing levels of Al3+ treatment in situ led toincreased accumulation of all the three osmolytesproline, glycine betaine and sucrose in roots as wellas shoots during a 5–20 d growth period (Figs. 4 and5a). With Al treatment level of 160 mM in situ about180–276% increase in proline content, 236–286%increase in glycine betaine content and 25–41%increase in sucrose content was observed in rootsas well as shoots of 15 d grown rice seedlingscompared to the values in controls
Effect of water stress on sucrose content
When rice seedlings grown for 10 and 20 d weresubjected to in vitro water stress treatment of�0.5 and �2.0MPa for 24 h by using varyingconcentrations of polyethylene glycol (PEG-6000),it was observed that with a mild water stress levelof �0.5MPa sucrose content increased in roots aswell as shoots compared to control seedlings,whereas with a higher water stress level of�2.0MPa a decline in sucrose content was observedin seedlings compared to controls (Fig. 5b). Seed-lings subjected to mild water stress showed about19–29% increase in sucrose content whereas with ahigher water stress level of �2.0MPa about 10–21%decline in the content of sucrose was observed.
aine and sucrose in vitro in presence of H2O2 on Michaeliscontrol grown seedlings of rice cv. Pant-12
(mmol/L KNO3) Vmax (nmol/minmg protein)
0 4.02 2.25 3.34 3.02 2.8
ARTICLE IN PRESS
Table 2. Effect of the osmoprotectant proline in vitro in presence of PEG, NaCl and metal salts on Michaelis constants(Km) and Vmax of NR extracted from shoots of 15 d control grown seedlings of rice cv. Pant-12
Treatments Km (mmol/L KNO3) Vmax (nmol/minmg protein)
Enzyme 0.70 4.00Enzyme+40% PEG 1.64 1.90Enzyme+40% PEG+1mol/L Proline 1.16 2.90Enzyme+300mM NaCl 1.82 1.75Enzyme+300mM NaCl+ 1mol/L Proline 1.23 3.20Enzyme+2mM Al2(SO4)3 1.74 2.00Enzyme+2mM Al2(SO4)3+1mol/L Proline 1.36 3.20Enzyme+2mM As2O3 2.04 1.80Enzyme+2mM As2O3+1mol/L Proline 1.56 2.10Enzyme+2mM NiSO4 1.68 2.50Enzyme+2mM NiSO4+1mol/L Proline 1.28 3.10Enzyme+2mM Pb(NO3)2 1.56 2.00Enzyme+2mM Pb(NO3)2+1mol/L Proline 1.10 2.80Enzyme+2mM CdCl2 1.59 2.40Enzyme+2mM CdCl2+1mol/L Proline 1.24 3.00
Figure 4. Effect of increasing concentrations of Al3+ inthe growth medium on the contents of proline, glycinebetaine in roots and shoots of rice cvs. Malviya-36 andPant-12 at increasing days of growth. Values are meanbased on three independent determinations and barsindicate standard deviations.
P. Sharma, R. Shanker Dubey860
Discussion
NR has been shown to be regulated by manyenvironmental factors as well as endogenousfactors such as metabolites and plant growthregulators (Guerrero et al., 1981). In our experi-ments in 15 d grown highly Al stressed (160 mM) andwater stressed (�2.0MPa) seedlings lower levels ofNR act as well as NR max were observed whereasNR activation state was almost unaltered. Withincreasing water stress or aluminium stress aconsistent decline in NR max was evident. In acidsoils below pH 5.0, Al3+ toxicity is a major problem.In such soils, the NH4
+ form of N predominates andNO3
� availability is limited (Foy et al., 1978),whereas during water stress nitrate absorption byroots and its transportation decreases due to loss ofturgor and transpiration. Decreased nitrate avail-ability in plants could inhibit NR gene transcriptionand decrease the stability of NR mRNAs. It couldalso affect other factors such as post-translationalcontrols including phosphorylation and proteolysis(Ferrario et al., 1995). Phosphorylation makes NRsensitive to proteolytic degradation (Lejay et al.,1997) as well as it sensitizes the enzyme toinhibition by an inhibitory 14-3-3 protein inpresence of Mg2+ (McKintosh et al., 1995). Phos-phorylation of the NR protein as well as induction ofproteases occur rapidly as a result of water deficit(Lejay et al., 1997). Cys proteases were inducedwithin 10 h of the onset of water stress inArabidopsis thaliana (Koizumi et al., 1993). Thesefactors could account for the decreased NR maxand NR act under water stress as well as aluminium

ARTICLE IN PRESS
Figure 5. Effect of Al3+ in situ and water stress onsucrose content in roots and shoots of seedlings of ricecvs. Malviya-36 and Pant-12. (a) Seedlings were raised insand cultures containing 80 and 160 mM Al3+ and sucrosecontent was estimated in 5, 10, 15 and 20 d grownseedlings, (b) Ten and 20 d grown seedlings weresubjected to in vitro water stress (17% PEG ¼ �0.5MPa,41.2% PEG ¼ �2.0MPa) for 24 h and sucrose content wasdetermined in roots and shoots. Values are mean basedon three independent determinations and bars indicatestandard deviations.
Modulation of nitrate reductase under stresses 861
toxicity conditions. Decreased NR max under in situAl stress might be due to direct inhibitory effect ofAl metal on NR activity (Campbell, 1999). Mildwater stress level of �0.5MPa or Al treatment of80 mM led to increased NR act and NR activationstate. Similar to our findings of mild water stressand Al toxicity conditions, higher NR activationstate and a lower level of NR max were observed inMo deficient wheat and nitrate deficient spinachplants (Yaneva et al., 2000). It is suggested that atmild stressed condition such a compensatoryresponse involving higher activation state andlower NR max becomes important in insuring high
level of active (dephosphorylated) form of NRhaving a lower amount of functional NR protein orNR max (Yaneva et al., 2000).
Stressful conditions of the soil environment suchas water stress, salinity and excessive metalstrigger production of ROS including hydrogenperoxide (H2O2) that can cause oxidative damageto the biomolecules such as proteins, lipids, andnucleic acids (Foyer and Noctor, 1998). A markedinhibition in the activity of NR was observed whenincreasing concentrations of H2O2 was provided inthe medium. Incorporation of 1M proline, glycinebetaine or sucrose in the assay medium led to aconsiderable protection of enzyme activity inpresence of H2O2. This suggests that these osmo-lytes possibly possess some antioxidant and ROSscavenging properties thereby protecting NRagainst damage due to H2O2 (Hare et al., 1999).
Increasing concentration of NaCl, PEG and metalsalts in vitro showed a marked inhibitory effect onthe activity of NR. Though Na+ and K+ are essentialfor NR, their salts are strong inhibitors (Hewitt,1975). NaCl readily enters the hydration sphere ofproteins, favouring unfolding. Addition of PEG inthe assay medium leads to a considerable pre-cipitation and denaturation due to in vitro osmoticstress (Paleg et al., 1984). It is suggested that thereis a putative metal binding site on NR responsiblefor inhibition of NR activity by metals (Campbell,1999). Inhibition in the activities of many enzymeswith metals like As, Pb, Cd has been attributed tothe interaction of these metals with functional –SHgroup present on enzymes (Shah and Dubey, 1995).Addition of 1M proline, glycine betaine or sucrosein the assay medium provided considerable protec-tion to the enzyme against inhibition due to NaCl,PEG-6000 and metal salts probably by providing anenvironment compatible with macromolecularstructure and function. It has been suggested thatthese organic solutes are preferentially excludedfrom the surface of protein and their immediatehydration sphere. Thus the addition of thesesolutes to a protein suspension creates a thermo-dynamically unfavourable situation, since the che-mical potential of both protein and the additivesare increased. This situation stabilizes the nativeconformation of the protein because denaturationwould lead to a greater contact surface betweenthe protein and the solvent, thus, augmenting theunfavourable effect (Crowe et al., 1988).
Incorporation of 1mM H2O2, 40% PEG-6000,300mM NaCl, 2mM salts of the metal Al, As, Ni,Pb, Cd in the enzyme assay medium led to asignificant increase in Km and decline in Vmax valuesof the enzyme determined in the absence ofthese compounds. This is suggestive that these

ARTICLE IN PRESS
P. Sharma, R. Shanker Dubey862
compounds H2O2, PEG, NaCl or polluting metalssignificantly decrease the affinity of NR towards itssubstrate NO3
� and denature/inactivate NR leadingto its decreased activity. Decreased Km andincreased Vmax values were observed when eitherof the osmolytes proline, glycine betaine or sucrosewas added in presence of H2O2 in the enzyme assaymedium. Similarly when proline was added to theenzyme assay medium containing PEG, NaCl orpolluting metal salts, a decrease in Km and increasein Vmax were observed compared to Km and Vmax inabsence of proline. This suggest that osmolytes inthe vicinity of enzyme help in maintaining ade-quate affinity of enzyme towards its substrate aswell as its catalytic rate.
Increased content of proline and glycine betainewas observed in our experiments in Al treatedrice seedlings and sucrose in Al as well as waterstressed seedlings compared to the levels inuntreated seedlings. Accumulation of proline,glycine betaine and sucrose in response to environ-mental stresses including water stress, salinity andmetal toxicity has been reported by various workers(Shah and Dubey, 1998; Aziz et al., 1998; Hagemannand Marin, 1999; Yang et al., 2000; Xing andRajashekar, 2001; Girija et al., 2002; Gill et al.,2003). Proline accumulation is regarded as acomponent of stress tolerance mechanism (Hareet al., 1999). Proline accumulation in plant tissueunder stressful conditions has been suggested tobe result of a decrease in proline degradation,increase in proline biosynthesis, a decrease inprotein synthesis or proline utilization and in-creased hydrolysis of proteins (Hare et al., 1999).There are different opinions regarding the mechan-isms by which proline can alleviate metal toxicitywithin the cell. It has been shown that free prolineacts as osmoprotectant (Paleg et al., 1984), proteinstabilizer (Sharma and Dubey, 2004), metal chela-tor (Farago and Mullen, 1979) inhibitor of lipidperoxidation (Mehta and Gaur, 1999), free radicalscavenger (Alia et al., 2001), etc. The detailmechanism of protection provided by proline toNR however, remains to be worked out.
The accumulation of glycine betaine occursmainly by transmembrane transport or by synthesisfrom choline (Rontein et al., 2002). Glycine betainecounter effects denaturants and stabilizes nativeprotein structures and membranes. There is alsoevidence that glycine betaine can protect proteinsvia a chaperon-like action on protein folding andthat glycine betaine may act as a signal moleculethat could elicit the expression of genes associatedwith stress tolerance (Rontein et al., 2002). Sucrosecan counteract the unwanted changes induced inthe structure of the ordered water by denaturants
and can stabilize the protein conformation (Chenand Arakawa, 1996). The degree of structuralprotection conferred by sucrose correlates withthe extent of hydrogen bonding potential and thepolarization of the backbone permanent dipole,which induce an increase in hydrogen bondingcharacter of neighboring polar residues (Hammes,1968). The consequences are local rearrangementsof water molecules and stabilization of the or-iented dipole, both of which contribute to thestability of a macromolecule like NR protein. Ourobservations that the osmolytes proline, glycinebetaine and sucrose accumulate in rice plants withimposition of stresses and that under in vitroconditions these osmolytes help in maintainingcatalytic activity of NR suggest that these osmo-lytes have a definite protective role for NR protein.
Our results thus suggest that aluminium toxicityas well as water deficit modulate NR activity bydecreasing total amount of functional NR and thatthe compounds proline, glycine betaine and sucroseaccumulate in stressed rice plants and appear tohave a putative role in protecting NR activity underabiotic stresses.
Acknowledgements
Financial support for this work was provided bythe University Grants Commission, New Delhi, inthe form of a major research project. PS is thankfulto the Council of Scientific and Industrial Research(CSIR), New Delhi for providing a Junior ResearchFellowship.
References
Alia, Mohanty P, Matysik J. Effect of proline on theproduction of singlet oxygen. Amino Acids2001;21:195–200.
Aziz A, Martin-Tanguy J, Larher F. Stress induced changesin polyamine and tyramine levels can regulate prolineaccumulation in tomato leaf discs treated with sodiumchloride. Physiol Plant 1998;104:195–202.
Bates LS, Waldren RP, Teare ID. Rapid determination offree proline for water stress studies. Plant Soil1973;39:205–7.
Botrel A, Magne C, Kaiser WM. Nitrate reduction, nitritereduction and ammonium assimilation in barley rootsin response to anoxia. Plant Physiol Biochem1996;34:645–52.
Bradford MM. A rapid and sensitive method for thequantitation of microgram quantities of protein-dyebinding. Anal Biochem 1976;72:248–54.
Campbell WH. Nitrate reductase structure, function andregulation: bridging the gap between biochemistry

ARTICLE IN PRESS
Modulation of nitrate reductase under stresses 863
and physiology. Annu Rev Plant Physiol Plant Mol Biol1999;50:277–303.
Chen BL, Arakawa T. Stabilization of recombinant humankeratinocyte growth factor by osmolytes and salts. JPharm Sci 1996;85:419–26.
Crowe JH, Crowe LM, Carpenter JF, Rudolph AS, WistromCA, Spargo BJ, Anchordoguy TJ. Interaction of sugarswith membranes. Biochim Biophys Acta1988;947:367–84.
Dubey RS, Pessarakli M. Physiological mechanisms ofnitrogen absorption and assimilation in plants understressful conditions. In: Pessarakli M, editor. Handbookof Plant and Crop Physiology. Marcel Dekker: NewYork; 2002. p. 637–55.
Farago ME, Mullen WA. Plants which accumulate metals.Part IV. A possible copper–proline complex from theroots of Armeria maritime. Inorg Chim Acta1979;32:L93–4.
Ferrario S, Valadier M-H, Morot-Gaudry J-F, Foyer CH.Effects of constitutive expression of nitrate reductasein transgenic Nicotiana plumbaginifolia L. in responseto varying nitrogen supply. Planta 1995;196:288–94.
Foy CD, Chaney RL, White MC. The physiology of metaltoxicity in plants. Annu Rev Plant Physiol1978;29:511–66.
Foyer CH, Noctor G. Ascorbate and glutathione: keepingactive oxygen under control. Annu Rev Plant PhysiolPlant Mol Biol 1998;49:249–79.
Gill PK, Sharma AD, Singh P, Bhullar SS. Changes ingermination, growth and soluble sugar contents ofSorghum bicolor L. Moench seeds under various abioticstresses. Plant Growth Regul 2003;40:157–62.
Girija C, Smith BN, Swamy PM. Interactive effects ofsodium chloride and calcium chloride on the accumu-lation of proline and glycine betaine in peanut(Arachis hypogae L.). Env Exp Bot 2002;47:1–10.
Grattan SR, Grieve CM. Betaine status in wheat inrelation to nitrogen stress and to transient salinitystress. Plant Soil 1985;85:3–9.
Guerrero MG, Vega JM, Losada M. The assimilatory nitratereducing system and its regulation. Annu Rev PlantPhysiol 1981;32:169–204.
Hagemann M, Marin K. Salt-induced sucrose accumulationis mediated by sucrose phosphate synthase in Cyano-bacteria. J Plant Physiol 1999;155:424–30.
Hammes GG. Relaxation spectrometry of biologicalsystems. Adv Protein Chem 1968;23:1–57.
Hare PD, Cress WA, Van Staden J. Proline synthesis anddegradation: a model system for elucidating stress-related signal transduction. J Exp Bot 1999;50:413–34.
Hewitt EJ. Assimilatory nitrate–nitrite reaction. AnnuRev Plant Physiol 1975;26:73–100.
Hoagland DR, Arnon DI. The water culture method forgrowing plants without soil. Cal Agr Exp St Circ1938;1:39.
Hsu PH. Effect of pH, phosphate and silicate on thedetermination of aluminium with aluminon. Soil Sci1963;96:230–8.
Kaiser WM, Weiner H, Huber SC. Nitrate reductase inhigher plants: a case study for transduction of
environmental stimuli into control of catalytic activ-ity. Physiol Plant 1999;105:385–90.
Kaiser WM, Kandlbinder A, Stoimenova M, GlaabJ. Discrepancy between nitrate reduction rates inintact leaves and nitrate reductase activity in leafextracts: What limits nitrate reduction in situ. Planta2000;210:801–7.
Koizumi M, Yamaguchi-Shinozaki K, Tsuji H, ShinozakiK. Structure and expression of two genes that encodedistinct drought-inducible cysteine proteinases inArabidopsis thaliana. Gene 1993;129:175–82.
Lejay L, Quillere I, Roux Y, Tillard P, Cliquet JB, Meyer C,Morot-Gaudry JF, Gojon A. Abolition of posttranscrip-tional regulation of nitrate reductase partially pre-vents the decrease in leaf NO3
– reduction whenphotosynthesis is inhibited by CO2 deprivation, butnot in darkness. Plant Physiol 1997;115:623–30.
McKintosh C, Douglas P, Lillo C. Identification of a proteinthat inhibits the phosphorylated form of nitratereductase from spinach (Spinacia oleracea) leaves.Plant Physiol 1995;107:451–7.
Mehta SK, Gaur JP. Heavy-metal-induced proline accu-mulation and its role in ameliorating metal toxicity inChlorella vulgaris. New Phytol 1999;143:253–9.
Oji Y, Takahashi M, Nagai Y, Wakiuchi N. NADH dependentnitrate reductase from 2-row barley roots: purifica-tion, characterization and comparison with leafenzyme. Physiol Plant 1988;72:311–5.
Paleg LG, Stewart GR, Bradbeer JW. Proline and glycine-betaine influence on protein solvation. Plant Physiol1984;75:974–8.
Richharia A, Shah K, Dubey RS. Nitrate reductase fromrice seedlings: partial purification, characterizationand the effects of in situ and in vitro NaCl salinity. JPlant Physiol 1997;151:316–22.
Rontein D, Nishida I, Tashiro G, Yoshioka K, Wu WI,Voelker DR, Hanson AD. Plant synthesize ethanolamineby direct decarboxylation of serine using a pyridoxalphosphate enzyme. J Biol Chem 2002;276:35523–9.
Saunders AJ, Davis-Searles PR, Allen DL, Pielak GJ, ErieDA. Osmolyte induced changes in protein conforma-tional equilibria. Biopolymers 2000;53:293–307.
Shah K, Dubey RS. Effect of cadmium on RNA level as wellas activity and molecular forms of ribonuclease ingrowing rice seedlings. Plant Physiol Biochem1995;33:577–84.
Shah K, Dubey RS. Effect of cadmium on prolineaccumulation and ribonuclease activity in rice seed-ling: role of proline as a possible enzyme protectant.Biol Plant 1998;40:121–30.
Sharma P, Dubey RS. Ascorbate peroxidase from riceseedlings: properties of enzyme isoforms, effects ofstresses and protective roles of osmolytes. Plant Sci2004;167:541–50.
Van Handel E. Direct microdetermination of sucrose. AnalBiochem 1968;22:280–3.
Wang HL, Lee PD, Liu LF, Su JC. Effect of sorbitol inducedosmotic stress on the changes of carbohydrate andfree amino acid pools in sweet potato cell suspensioncultures. Bot Bull Acad Sin 1999;40:219–25.

ARTICLE IN PRESS
P. Sharma, R. Shanker Dubey864
Xing W, Rajashekar CB. Glycine betaine involvement in
freezing tolerance and water stress in Arabidopsis
thaliana. Environ Exp Bot 2001;46:21–8.
Yaneva IA, Baydonova VD, Vunkova-Radeva RV. Nitrate
reductase activation states in leaves of molybdenum-
deficient winter wheat. J Plant Physiol 2000;157:495–501.
Yang YY, Jung JY, Song WY, Suh SH, Lee Y. Identification ofrice varieties with high tolerance or sensitivity to leadand characterization of the mechanism of tolerance.Plant Physiol 2000;124:1019–26.


















