
MobilizationofBoneMarrowStemCellsbyGranulocyte...
10
Mobilization of Bone Marrow Stem Cells by Granulocyte Colony-Stimulating Factor Ameliorates Radiation-Induced Damage to Salivary Glands Isabelle M.A. Lombaert, 1,2 Pieter K. Wierenga, 2 Tineke Kok, 1 Harm H. Kampinga, 1 Gerald deHaan, 2 and Robert P. Coppes 1,3 Abstract Purpose: One of the major reasons for failure of radiotherapeutic cancer treatment is the limita- tion in dose that can be applied to the tumor because of coirradiation of the normal healthy tissue. Late radiation-induced damage reduces the quality of life of the patient and may even be life threatening. Replacement of the radiation-sterilized stem cells with unirradiated autologous stem cells may restore the tissue function. Here, we assessed the potential of granulocyte colony- stimulating factor (G-CSF) ^ mobilized bone marrow ^ derived cells (BMC) to regenerate and functionally restore irradiated salivary glands used as a model for normal tissue damage. Experimental Design: Male-eGFP + bone marrow chimeric female C57BL/6 mice were treated with G-CSF, 10 to 60 days after local salivary gland irradiation. Four months after irradiation, sali- vary gland morphology and flow rate were assessed. Results: G-CSF treatment induced homing of large number of labeled BMCs to the submandib- ular glands after irradiation. These animals showed significant increased gland weight, number of acinar cells, and salivary flow rates. Donor cells expressed surface markers specific for hemato- poietic or endothelial/mesenchymal cells. However, salivary gland acinar cells neither express the G-CSF receptor nor contained the GFP/Ychromosome donor cell label. Conclusions: The results show that BMCs home to damaged salivary glands after mobilization and induce repair processes, which improve function and morphology. This process does not involve transdifferentiation of BMCs to salivary gland cells. Mobilization of BMCs could become a promising modality to ameliorate radiation-induced complications after radiotherapy. During radiotherapy, the most important dose-limiting factor is sensitivity of the normal tissue lying in the radiation field. Even with the most optimal radiation schedule, damage will still occur in normal tissues. For radiotherapeutic treatment of head and neck tumors, the salivary glands are one of the tissues at risk. Exposure of the salivary gland to radiation often results in a progressive loss of gland function within the first weeks of radiotherapy (1). The reduction in saliva flow rate and alteration of salivary composition may persist during the rest of the patient’s life, leading to complications, including oral dryness, nocturnal oral discomfort, increased risk for oral infections, dental caries, and difficulties in speech, which severely hamper the quality of life (2, 3). Radiation damage to normal tissue is often due to reduced functioning of the tissue stem cells that can no longer replace differentiated functional cells, resulting in loss of homeostasis. In the salivary gland, early gland dysfunction (0-10 days after irradiation) is evoked by disturbance of intracellular receptor- mediated signaling in differentiated functional acinar cells without significant cell depletion (4). At later time points (10-60 days after irradiation), progressive loss of acinar cells is observed, which is associated with further decrease in saliva production. The lack of replenishment of these functional cells is thought to be due to the radiation-induced sterilization of endogenous stem cells. Replacement of salivary gland stem cells by stem cell therapy could potentially restore tissue homeosta- sis after radiation. At present, there are no reliable methods available to obtain sufficient numbers of well-characterized salivary gland stem cells that could be applicable in such a stem cell therapy approach. Recently, however, bone marrow–derived cells (BMC) were suggested to serve as a valuable source for the regeneration of damaged tissues (5). Homing and engraftment of BMCs in damaged nonhematopoietic organs, such as vascular tissue (6), myocardium (7–15) brain (16–18), liver (19–22), kidney (23–25), lung (26–30), and skin (31), have been observed and were suggested to contribute to the wound-healing process. In some tissues like myocardium (9, 12–15, 32), kidney (24), and Cancer Therapy: Preclinical Authors’ Affiliations: Sections of 1 Radiation and Stress Cell Biology and 2 Stem Cell Biology, Department of Cell Biology and 3 Department of Radiation Oncology, University Medical Center Groningen, University of Groningen, Groningen the Netherlands Received 11/1/05; revised 12/23/05; accepted 1/11/06. Grant support: Dutch Society for Cancer Research grant RUG2003-2909, the European Union FP-6 contract 503436, and Amgen, Inc. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. Requests for reprints: Robert P. Coppes, University Medical Center Groningen, Antonius Deusinglaan 1, 9713 AV Groningen, the Netherlands. Phone: 31-50-363- 2709; Fax: 31-50-363-2913; E-mail: r.p.coppes@med.umcg.nl. F 2006 American Association for Cancer Research. doi:10.1158/1078-0432.CCR-05-2381 www.aacrjournals.org Clin Cancer Res 2006;12(6) March 15, 2006 1804 Research. on May 17, 2018. © 2006 American Association for Cancer clincancerres.aacrjournals.org Downloaded from
-
Upload
phungkhuong -
Category
Documents
-
view
216 -
download
2
Transcript of MobilizationofBoneMarrowStemCellsbyGranulocyte...

Mobilization of Bone Marrow Stem Cells by GranulocyteColony-Stimulating FactorAmeliorates Radiation-InducedDamage to Salivary GlandsIsabelle M.A. Lombaert,1,2 Pieter K. Wierenga,2 Tineke Kok,1Harm H. Kampinga,1
Gerald deHaan,2 and Robert P. Coppes1,3
Abstract Purpose: One of the major reasons for failure of radiotherapeutic cancer treatment is the limita-tion in dose that canbe applied to the tumor because of coirradiationof the normal healthy tissue.Late radiation-induced damage reduces the quality of life of the patient and may even be lifethreatening. Replacement of the radiation-sterilized stem cells with unirradiated autologous stemcells may restore the tissue function. Here, we assessed the potential of granulocyte colony-stimulating factor (G-CSF)^ mobilized bone marrow ^ derived cells (BMC) to regenerate andfunctionally restore irradiated salivary glands used as amodel for normal tissue damage.Experimental Design:Male-eGFP+ bone marrow chimeric female C57BL/6 mice were treatedwith G-CSF,10 to 60 days after local salivary gland irradiation. Four months after irradiation, sali-vary glandmorphology and flow rate were assessed.Results: G-CSF treatment induced homing of large number of labeled BMCs to the submandib-ular glands after irradiation. These animals showed significant increased gland weight, numberof acinar cells, and salivary flow rates. Donor cells expressed surfacemarkers specific forhemato-poietic or endothelial/mesenchymal cells. However, salivary gland acinar cells neither express theG-CSF receptor nor contained the GFP/Ychromosome donor cell label.Conclusions:The results show that BMCs home to damaged salivary glands after mobilizationand induce repair processes, which improve function and morphology. This process does notinvolve transdifferentiation of BMCs to salivary gland cells. Mobilization of BMCs could becomea promisingmodality to ameliorate radiation-induced complications after radiotherapy.
During radiotherapy, the most important dose-limiting factoris sensitivity of the normal tissue lying in the radiation field.Even with the most optimal radiation schedule, damage willstill occur in normal tissues. For radiotherapeutic treatment ofhead and neck tumors, the salivary glands are one of thetissues at risk. Exposure of the salivary gland to radiationoften results in a progressive loss of gland function withinthe first weeks of radiotherapy (1). The reduction in salivaflow rate and alteration of salivary composition may persistduring the rest of the patient’s life, leading to complications,including oral dryness, nocturnal oral discomfort, increased
risk for oral infections, dental caries, and difficulties inspeech, which severely hamper the quality of life (2, 3).
Radiation damage to normal tissue is often due to reducedfunctioning of the tissue stem cells that can no longer replacedifferentiated functional cells, resulting in loss of homeostasis.In the salivary gland, early gland dysfunction (0-10 days afterirradiation) is evoked by disturbance of intracellular receptor-mediated signaling in differentiated functional acinar cellswithout significant cell depletion (4). At later time points(10-60 days after irradiation), progressive loss of acinar cells isobserved, which is associated with further decrease in salivaproduction. The lack of replenishment of these functional cellsis thought to be due to the radiation-induced sterilization ofendogenous stem cells. Replacement of salivary gland stem cellsby stem cell therapy could potentially restore tissue homeosta-sis after radiation.
At present, there are no reliable methods available to obtainsufficient numbers of well-characterized salivary gland stem cellsthat could be applicable in such a stem cell therapy approach.Recently, however, bone marrow–derived cells (BMC) weresuggested to serve as a valuable source for the regeneration ofdamaged tissues (5). Homing and engraftment of BMCs indamaged nonhematopoietic organs, such as vascular tissue (6),myocardium (7–15) brain (16–18), liver (19–22), kidney(23–25), lung (26–30), and skin (31), have been observed andwere suggested to contribute to the wound-healing process. Insome tissues like myocardium (9, 12–15, 32), kidney (24), and
Cancer Therapy: Preclinical
Authors’Affiliations: Sections of 1Radiation and Stress Cell Biology and 2StemCell Biology, Department of Cell Biology and 3Department of Radiation Oncology,University Medical Center Groningen, University of Groningen, Groningen theNetherlandsReceived11/1/05; revised12/23/05; accepted1/11/06.Grant support: Dutch Society for Cancer Research grant RUG2003-2909, theEuropean Union FP-6 contract 503436, and Amgen, Inc.The costs of publication of this article were defrayed in part by the payment of pagecharges.This article must therefore be hereby marked advertisement in accordancewith18 U.S.C. Section1734 solely to indicate this fact.Requests for reprints: Robert P. Coppes, University Medical Center Groningen,Antonius Deusinglaan1, 9713 AVGroningen, the Netherlands. Phone: 31-50-363-2709; Fax: 31-50-363-2913; E-mail: [email protected].
F2006 American Association for Cancer Research.doi:10.1158/1078-0432.CCR-05-2381
www.aacrjournals.orgClin Cancer Res 2006;12(6) March15, 2006 1804
Research. on May 17, 2018. © 2006 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

liver (19), even improved function has been observed. Thesestudies have provided the proof of principle of damage repair bythe application of BMCs. A clinically attractive approach is to usegranulocyte colony-stimulating factor (G-CSF) to mobilize bonemarrow cells to the circulation. It has been reported that G-CSF isassociated with improved cardiac function and survival aftermyocardial infarction in mice (9, 12, 32, 33).
Many investigators use radiation as a conditioning regimenfor stem cell transplantation. However, the prevention/repair oftherapeutic radiation-induced damage to organs has not beenassessed. The salivary gland not only provides a unique modelbut is also clinically very relevant to prevent normal tissuetoxicity after radiotherapeutic treatment of head and neckcancers. Because these side effects are mainly caused by damageto the stem cells, a proof of principle in this area may also shedlight on the general therapeutic use of stem cells. Thearchitecture of the salivary gland is well known: cells can bereadily identified morphologically, and precise function meas-urements in time have been developed and extensively used inour laboratory for both rats and mice. Thus, the exact timepoints of loss of function, the accompanying morphologicchanges, and the incapability of regeneration after stem celldepletion are well known (2–4, 34–39).
In the current study, we assessed the capability of G-CSF–mobilized BMCs to regenerate and functionally restore irradi-ated salivary glands.
Materials andMethods
Animals. Female C57BL/6 mice, 6 to 8 weeks old, were purchasedfrom The Jackson Laboratory (Bar Harbor, ME) and used in a sex-mismatched transplantation setting. Enhanced GFP male C57BL/6-TgN(ActbeGFP) mice, were bred in the animal facility of the UniversityMedical Center Groningen and used as donor mice for GFP+ wholebone marrow. All mice were kept under clean conventional conditionsand fed ad libitum with food pellets (RMH-B, Hope Farms B.V.,Woerden, the Netherlands) and acidified tap water (pH 2.8). Allexperiments were approved by the animal Ethical Committee onanimal testing of the University of Groningen.
Transplantation protocol. Mice were splenectomized under halo-thane/O2 anesthesia and allowed to recover for at least 2 weeks.Splenectomized female acceptor mice were given 9.5 Gy total bodyirradiation of X-rays (Philips CMG 41 X, 200 kV, 10 mA, 5 Gy/min)shielding the salivary glands (12 � 50 � 3 mm lead plate) and weretransplanted with 3 � 106 male bone marrow cells obtained fromfemurs from GFP+ transgenic mice by orbital injection. Bone marrowcells were obtained by flushing the femoral content with Iscove’smodified Dulbecco’s medium (Life Technologies, Paisley, Scotland).Chimerism in peripheral blood was documented 8 weeks later byanalysis on a FACSCalibur flow cytometer (Becton Dickinson,Erembodegem, Germany). Only mice with >60% chimerism were usedfor the experiments.
Irradiation and mobilization protocol. One month after transplanta-tion, the salivary glands of the chimeric mice were locally irradiatedwith a single dose of 15 Gy (as described above), shielding the rest ofthe body with 3-mm lead. This dose is known to induce sufficientdamage without compromising the general health of the animals. Atdays 10, 30, or 60 after gland irradiation, bone marrow cells weremobilized by s.c. injections of 2 � 25 Ag PEG-rHu-G-CSF (Amgen, Inc.,Thousand Oaks, CA) given 3 days apart.
Colony-forming cell assays. Progenitor cells were assayed asdescribed earlier (40). Briefly, cells were plated out in a-medium(StemCell Technologies, Inc., Vancouver, British Columbia, Canada)containing 0.8% methylcellulose (Fluka, Bachs S.G., Switzerland), 30%
FCS (Life Technologies), and 10�4 mol/L 2-mercapthoethanol (Merck,Darmstadt, Germany) at concentrations varying from 104 to 5 � 105
nucleated cells/mL. Colony growth is stimulated by granulocytemacrophage colony-stimulating factor and stem cell factor. Cultureswere plated in 35-mm polystyrene culture dishes (Falcon, BectonDickinson, Lincoln Park, NJ) and grown at 37jC in a 5% CO2
humidified atmosphere. Colonies (>50 cells) were scored after 7 days ofculture.
The cobblestone area-forming cell assay allows the assessment of
primitive hematopoietic stem cells (40). The cobblestone area-forming
cell assay was done by establishing confluent FBMD-1 cell cultures in
96-well plates (Costar, Corning, NY). The confluent cell cultures were
overlaid with mobilized peripheral blood cells in a limiting dilution
setup. Eight dilutions 2-fold apart were used with 10 replicate wells per
dilution. The cells were cultured in Iscove’s modified Dulbecco’s
medium supplemented with 20% horse serum (Life Technologies) at
33jC in a 10% CO2 humidified atmosphere with a half-volume
medium change every week. The percentage of wells with at least one
phase-dark hematopoietic clone of at least five cells beneath the stromal
layer was determined 4 to 5 weeks after initiating the culture.
Cobblestone area frequencies were calculated using Poisson statistics.Saliva collection. Before and 90 days after irradiation, whole saliva
flow rate was determined. The animals were placed in a restrainingdevice (41) after pilocarpine injection (2 mg/kg, s.c.). Saliva wascollected for 15 minutes and determined gravimetrically, assuming adensity of 1 g/mL for saliva.
Immunohistochemistry. At 130 days after irradiation of the salivaryglands, animals were sacrificed for immunohistochemistry. The
submandibular glands were extirpated and incubated for 29 hours at4jC in 4% buffered formaldehyde. Following dehydration, the tissue
was embedded in paraffin. Five-micrometer sections were analyzed forthe presence of eGFP using a confocal scanning laser microscopy (Leica
TCS SP2). Counterstaining was done with 4V,6-diamidino-2-phenyl-indole (DAPI).
Overnight staining with anti-eGFP antibody (Chemicon, Temecula,CA; MAB3580; 1:500) was visualized with light microscopy bydiaminobenzidine chromogen. Avidin-biotin-horseradish peroxidasecomplex (Vector Elite Avidin-Biotin Complex kit) and anti-mouserabbit F(abV)2 biotine (DAKO, Carpinteria, CA; E 0413; 1:300) wereused after a 0.01 mol/L citrate antigen retrieval, following dewaxingand blocking the endogenous peroxidase with 0.3% H2O2 for 30minutes.
Anti-G-CSF receptor (G-CSFR; Santa Cruz Biotechnology, Santa Cruz,CA; clone H-176; 1:100) overnight labeling was done using 0.01 mol/Lcitrate antigen retrieval and anti-rabbit F(abV)2 biotine (DAKO; E 0431;1:300).
In addition, anti–a-smooth muscle actin (Sigma, St. Louis, MO;clone 1A4; 1:100) staining was done using anti-mouse rabbit F(abV)2
biotin for 1 hour at room temperature each. Visualization was made byusing streptavidin-Cy3 (Sigma; S 6402) for 30 minutes. Counterstainingwas done using DAPI.
Overnight CD31 (BD PharMingen, San Diego, CA; 550274; 1:100)labeling was done following 0.05% trypsin pretreatment at 37jC.Visualization was made by using anti-rat rabbit biotine (DAKO; E 0468;1:300) and streptavidin-Cy3. In addition, anti-CD45 (RDI-MCD45-F11; 1:100) labeling required the anti-rat biotine and streptavidin-Cy3in combination with a double labeling for anti-eGFP using avidin-FITC(Sigma; A 2050).
Double labeling of G-CSFR and anti–smooth muscle actin was doneusing avidin-FITC and streptavidin-Cy3, respectively.
The Y chromosome was stained by fluorescence in situ hybridization(Cambio, Cambridge, United Kingdom; 1187-cMB) accordingto manufacturer’s instructions. The following changes were made:5 minutes of 1 mol/L sodium thiocyanate and 0.4% pepsin instead of10 minutes. Nuclear staining was done with methyl green. Negativecontrols were included in the protocols. Gland morphology wasvisualized by routine histologic techniques with H&E. Acini were
Amelioration of Radiation-Induced Damage
www.aacrjournals.org Clin Cancer Res 2006;12(6)March15, 20061805
Research. on May 17, 2018. © 2006 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
detected by Periodic Acid Schiff’s base and Alcian Blue staining bywhich mucin and mucopolysacharides are detected, respectively.
Quantification of eGFP+ cells and acini. Tissue sections were analyzedusing the confocal scanning laser microscopy at a magnification of�630. The number of eGFP+DAPI+ cells in 200 counted cells wasenumerated. The amount of eGFP+ cells in a salivary gland of C57BL/6eGFP+ transgenic mice, was set at 100%, as not all salivary gland cellsseem to express the eGFP protein. The percentage of surface areaoccupied by acinar cells was counted by light microscopy (OlympusCX40, Germany) under �400 magnification using 100 squares of0.25 mm2 each. Of each gland, five squares in three sections (top-middle-bottom) were scored, and subsequently the data were pooled.
RNA isolation and PCR methods. From mice, the submandibularglands were excised, cut into small pieces, snap frozen in liquidnitrogen, and stored at �80jC until used. Total RNA was isolated fromfrozen submandibular gland tissue using TRIzol Reagent (Invitrogen,Carlsbad, CA) according to the manufacturer’s instructions. Total RNAwas isolated from fresh mouse whole bone marrow using the RNeasyMini kit (Qiagen, Hilden, Germany) according to the manufacturer’sinstructions. RNA was checked on an agarose gel for integrity, and RNAconcentration was measured spectrophotometrically. Reverse transcrip-tion was done on 2.5 Ag of total RNA using random primers in a finalvolume of 38 AL (Reverse Transcription System, Promega, Madison,WI) for 10 minutes at 25jC followed by 1 hour at 45jC. Sampleswere subsequently heated for 5 minutes at 95jC to terminate thereverse transcription reaction. The cDNA was amplified using mouseg-csfr (Csf3r) primers with the following sequences: sense primer,5V-GTACTCTTGTCCACTACCTGT-3V and antisense primer, 5V-CAAGA-TACAAGGACCCCCAA-3V (accession no. NM_007782). The PCR wasdone under the following conditions: an initial denaturation at 94jCfor 2 minutes followed by a cycle of denaturation at 94jC for 1 minute,annealing at 58jC for 1 minute, and extension at 72jC for 1 minute.The samples were subjected to 40 cycles, and the PCR products wereanalyzed on an ethidium bromide stained agarose gel.
Real-time quantitative PCR was done on the (irradiated) subman-dibular gland and bone marrow cDNA samples on a Bio-Rad iCyler iQReal-time Detection System. The following primer sequences were usedfor b-actin : sense primer, 5V-AGACCTCTATGCCAACACAG-3V andantisense primer, 5V-TAGGAGCCAGAGCAGTAATC-3V (accession no.NM_007393) and for g-csfr (Csf3r): sense primer, 5V-ATTGGCCCT-GATGTAGTCTC-3V and antisense primer, 5V-CACTCCTGTTCCATG-
TACTC-3V. Real-time PCR was conducted by amplifying the cDNAwith the iQ SYBR Green Supermix (Bio-Rad, Hercules, CA). For each setof primers, the PCR efficiency was determined. Melting curve analysis ofamplification products was done at the end of the PCR reaction toconfirm that a single PCR product was detected. For every PCR reaction,b-actin was used as the internal control. Quantification of the sampleswas calculated from the threshold cycle (C t) by interpolation from thestandard curve.
Statistical analysis. The results were analyzed using an unpairedStudent’s t tests and the Mann-Whitney test. Statistical significance wasdefined as P < 0.05 using SPSS.
Fig. 1. Schematic representation of experimental setup. Mice were total bodyirradiated excluding the salivary glands and transplantedwithwhole bonemarrowofeGFP+ transgenic mice (middle left clockwise). Hereafter, the salivary glands werelocally irradiated. At different time points, G-CSF was given, and several variableswere determined 90 to130 days after irradiation.
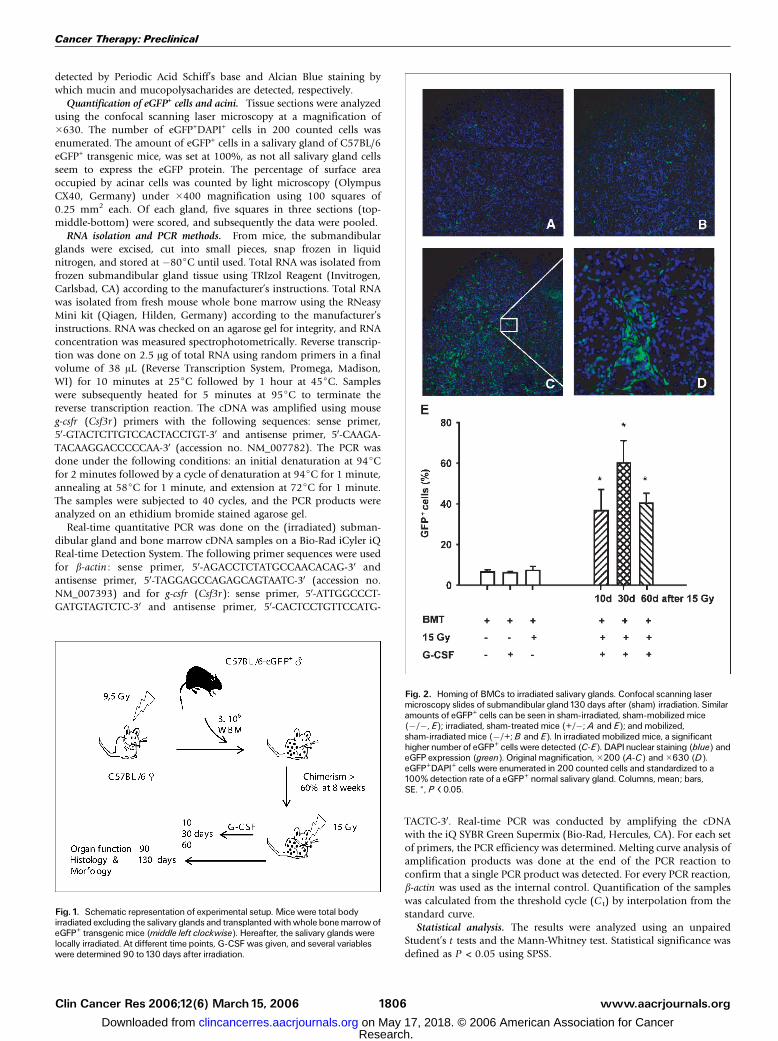
Fig. 2. Homing of BMCs to irradiated salivary glands. Confocal scanning lasermicroscopy slides of submandibular gland130 days after (sham) irradiation. Similaramounts of eGFP+ cells can be seen in sham-irradiated, sham-mobilized mice(�/�, E); irradiated, sham-treated mice (+/�; A and E); and mobilized,sham-irradiated mice (�/+;B and E). In irradiated mobilized mice, a significanthigher number of eGFP+ cells were detected (C-E). DAPInuclear staining (blue) andeGFP expression (green). Original magnification,�200 (A-C) and�630 (D).eGFP+DAPI+ cells were enumerated in 200 counted cells and standardized to a100% detection rate of a eGFP+ normal salivary gland. Columns, mean; bars,SE. *, P < 0.05.
Cancer Therapy: Preclinical
www.aacrjournals.orgClin Cancer Res 2006;12(6) March15, 2006 1806
Research. on May 17, 2018. © 2006 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

Results
Mobilized BMCs home to irradiated salivary glands. To beable to identify BMCs after mobilization, the endogenousbone marrow of wild-type female mice was destroyed by atotal body irradiation of 9.5 Gy (salivary glands wereshielded) and replaced by male bone marrow isolated from
eGFP transgenic mice (Fig. 1). Eight weeks later, mice withperipheral blood chimerism levels >60% were selected andreceived an additional irradiation on the salivary glands. At10, 30, or 60 days after gland irradiation, two doses ofG-CSF given 3 days apart were used to induce circulation ofBMCs in the blood. A clonogenic assay revealed that thismobilization protocol induced circulating levels of 10,000
Fig. 3. Improved morphology after gland irradiation andmobilization of BMCs. Salivary glands were photographed (A) weighted (B) at130 days after irradiation. Animproved gland color and a significant increase in glandweight were observed after G-CSF treatment comparedwith irradiated gland.The salivary glandmorphology (C) wasvisualized with a H&E (a, d, and g), Periodic Acid Schiff (b, e, and h), and Alcian Blue staining (c, f, and i), indicating tissue structure, the presence of mucin andmucopolysaccharides containing acini, respectively. Loss of acinar cells (*)with remaining duct cells (D) appeared after irradiation (d-f) comparedwith a normal salivary gland(a-c). Irradiated G-CSF ^ mobilized mice revealed more acinar groups, whichwere normal sized, as visualized by (g), (h), and (i). Ruler, mm. MeanF SE. *, P < 0.05. Originalmagnification,�200.
Amelioration of Radiation-Induced Damage
www.aacrjournals.org Clin Cancer Res 2006;12(6)March15, 20061807
Research. on May 17, 2018. © 2006 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

granulocyte macrophage colony-forming units and about1,000 hematopoietic stem cells per mL blood for at least1 week.
One hundred thirty days after gland irradiation, the numberof eGFP+ cells in the salivary glands was established. Figure 2shows that the irradiated salivary gland in non–G-CSF-treatedanimals (Fig. 2A) contained few eGFP+ cells. About the samenumber of eGFP+ cells were found in unirradiated salivaryglands of mice after G-CSF treatment (Fig. 2B). The highestlevel of eGFP+ cells was observed in salivary glands of micetreated with G-CSF 30 days after irradiation (Fig. 2C-D).Quantification of the % eGFP+ cells (Fig. 2E) revealed a lowlevel of eGFP in all glands of animals, which were not subjectedto the combination of G-CSF treatment and salivary glandirradiation (Fig. 2E, left). However, a significant 5- to 8-fold
increase in eGFP+ cells was observed in irradiated salivaryglands of G-CSF–treated animals, irrespective of the time pointof mobilization (10, 30, or 60 days after irradiation). Forfurther analysis, these groups were all pooled and treated as onegroup.
These data show that BMCs home to the salivary glandspecifically after irradiation. This effect can be stronglyenhanced by mobilizing BMCs by G-CSF.
G-CSF induced changes in morphology of irradiated salivarygland. Macromorphologic examination of glands, extirpated130 days after radiation, revealed a 60% reduction in glandweight (Fig. 3B). In addition, the color of the gland waschanged after radiation (Fig. 3A). When mice were treatedwith G-CSF after irradiation, gland weights were significantlyincreased and gland color was normalized. Although nosignificant difference in endothelial area was found betweenthe irradiated/G-CSF – treated group and the non – G-CSF-treated or nonirradiated mice, morphologic differenceswere observed. Blood vessels seemed somewhat largerafter irradiation and G-CSF treatment, possibly resultingin an improved blood flow, observed as an improved glandcolor.
Histologic analysis revealed that 130 days after irradiation,the number of acinar cells with remaining ducts wassubstantially reduced compared with normal gland tissue(Fig. 3C, a), and inflammatory aggregations were prevalent,consistent with previous studies (refs. 35, 39; Fig. 3C, d).Periodic Acid Schiff (Fig. 3C, b versus e) and Alcian Blue (Fig.3C, c versus f) staining for mucins and mucopolysaccharides,respectively, showed the reduced presence of acinar cells. Whenpresent, these cells were often enlarged and disorganized andsurrounded by ductal cells and fibrotic tissue. In animals thatwere irradiated and subsequently mobilized, however, thereduction in acinar cell number was less pronounced and theglands overall showed improved morphology (Fig. 3C, g, h,and i). Detailed quantitative analysis (Fig. 4A) confirmed asignificantly higher acinar cell number in the pooled G-CSF–treated group.
These results imply that G-CSF treatment after irradiationimproves micromorphology and macromorphology of thesalivary glands, which coincides with homing of BMCs intothe glands.
G-CSF adminstration increases salivary gland function. Toexamine whether the improved morphology after G-CSFtreatment is associated with restored function, total salivaryflow rates were measured. As expected, irradiation with a singledose of 15 Gy results in a severe drop of stimulated salivasecretion (3.7 F 2.3 AL/15 minutes, Fig. 4B) when comparedwith control nonirradiated animals (194.2 F 10.4 AL/15minutes). Secretion increased significantly after gland irradia-tion and G-CSF treatment (21.1 F 6.2 AL/15 minutes, P < 0.05),when compared with irradiated non G-CSF–treated animals.Only two irradiated but otherwise untreated mice had a salivayield of >10 AL/15 minutes. In contrast, 17 of the 28 mice ofthe G-CSF treated group responded with a level higher than10 AL/15 minutes of saliva secretion. These results indicate thatG-CSF treatment increases the function of irradiated salivaryglands of most animals.
Enduring acinar cells are nearly all not derived from the bonemarrow. Next, we investigated whether the increasednumber of acinar cells responsible for the morphologic and
Fig. 4. Increase in acinar cells and augmentation of saliva production after glandirradiation andmobilization of BMCs. A, histology slides were scored for acinar cellsas described in Materials andMethods. G-CSF treatment induced an increase inacinar cells in irradiated submandibular glands (w ) when compared with irradiatedglands (5) fromuntreated (o) orG-CSF ^ treatedmice (5). Points, mean; bars, SE.*, P < 0.05, significant different from irradiation only. B, saliva flow ratewasmeasured 90 days after irradiation. G-CSF treatment significantly increased thesaliva production after irradiation (5) comparedwith irradiated, sham-treated mice(5). An arbitrarily chosen responder line of10 AL/15 minutes saliva productionrevealed that 61% (17 of 28) of the mobilized mice respondedwith a higher organfunction. n = 5,12, and 28 for the normal (o); irradiated, sham-treated; andirradiated G-CSF ^ treated mice, respectively. Line, mean. *, P < 0.05.
Cancer Therapy: Preclinical
www.aacrjournals.orgClin Cancer Res 2006;12(6) March15, 2006 1808
Research. on May 17, 2018. © 2006 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

functional improvement of the irradiated salivary glands afterG-CSF treatment were derived from BMCs. We hardlyobserved any acinar cells expressing the eGFP protein.However, even not all acini of eGFP transgenic miceexpressed the eGFP protein to a detectable level (data notshown). It may be that the newly formed cells, althoughBMCs do not express the eGFP protein. Therefore, Ychromosome in situ hybridization was also used to detectmale cells originating from the donor bone marrow. Ascontrol, positive staining (Y+) was revealed in the majority ofcells of the male salivary glands (Fig. 5A), whereas nopositive staining was observed in glands of female mice(Fig. 5A, inset). In agreement with the eGFP data shownearlier, irradiated/sham-treated mice showed few Y+ cells inopen spaces or in close vicinity of duct and acinar cells(Fig. 5B). As expected, more Y+ cells were observed in theirradiated/G-CSF–treated mice, occurring either as clusters ofinfiltrating cells in open spaced nodules and arranged aroundcentral veins or as single cell connected to duct or acinar cells(Fig. 5C). However, only very few were detectable aspotential acinar or ductal cell (Fig. 5D). Anti-eGFP stainingconfirmed these results showing rare if any overlap withmucopolysaccharide containing acini (Fig. 6A and B).
Because many eGFP+ cells were found to reside in closevicinity with duct and acinar cells but not being acinar cells,an immunohistochemical staining (Cy3, red) for a-smoothmuscle actin was done. Figure 6C shows colocalization,although no overlap of the two signals. Both signals werealso found as myofibroblasts (Fig. 6D) known to be involvedin the tissue reaction to irradiation. Thus, it is still possiblethat BMCs are involved in the formation of myoepithelial cellssurrounding ducts and acini. Although a significant part ofthe interstitial eGFP+ cells were CD45+ (Fig. 6E), most wereunrelated to hematopoietic cells. CD31 staining revealed thatthese cells at least in part are likely to be mesenchymal cells
derived from the bone marrow (Fig. 6F). These resultsindicates that at best a very limited number of cells potentiallytransdifferentiate from BMCs to salivary gland cells. Themajority of the eGFP+ seem to be hematopoietic ormesenchymal cells.
To elucidate whether the morphologic and functionalimprovements were due to a direct protective or stimulatoryeffect of the cytokine G-CSF, we examined the expression ofthe G-CSF-receptor (G-CSFR) in the mouse salivary glandsusing reverse transcription-PCR, real-time quantitative PCR,and immunohistochemistry. Reverse transcription-PCRrevealed very low, barely detectable amount of g-csfr expres-sion in the salivary glands (Fig. 7A, lane 3). However,measured with a more sensitive method real-time quantitativePCR, some g-csfr expression was detectable although muchlower than the expression in bone marrow (data not shown).Furthermore, irradiation of the salivary glands with 15 Gy didnot increase the expression of g-csfr measured at different timepoints after the irradiation (data not shown). To indicatewhich cells in the salivary glands expressed the G-CSFR,immunohistochemistry was used. Figure 7B indicates that theexpression of the G-CSFR is solely detectable in endotheliumand myoepithelial cells surrounding ducts and acini. This isconfirmed with double labeling for G-CSFR and a-smoothmuscle actin (Fig. 7C and D). At the time of G-CSF treatment(10, 30, or 60 days after irradiation), no changes in G-CSFRexpression were observed.
These results indicate that it is unlikely that G-CSF directlystimulates the morphologic (i.e., formation of new acinar cells)and functional improvements.
In summary, a significant higher number of acinar cellsassociated with improved salivary function were observedwhen BMCs were mobilized with G-CSF after irradiation. Most,if not all, newly formed cells, however, were not derived fromBMCs.
Fig. 5. Donor origin of salivary gland cell: in situhybridization. Untreatedmale salivary gland (A) showedY+ chromosome expression compared with female ones(A, inset).The amount of male BMCs in the irradiatedgland (B, arrows) was enhanced by G-CSF treatment(C, arrows).Y+ BMCs infiltrate in open spaces in thenodules and surrounding the salivary gland cells inirradiated glands of G-CSF ^ treated mice (C).Occasionally, Y+ staining was seen in an acinar cell(*, D). Original magnification,�200 (A-C),�400 (D).Nucleus staining is seen by methyl green (B-D).
Amelioration of Radiation-Induced Damage
www.aacrjournals.org Clin Cancer Res 2006;12(6)March15, 20061809
Research. on May 17, 2018. © 2006 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

Discussion
Coirradiation-induced hyposalivation after cancer therapyfor head and neck cancers remains a major clinical problem.The depletion of resident viable stem cells after treatment hasdirected efforts to a search for alternative stem cell sources.Although bone marrow cells have been suggested to take partin the regeneration of damaged organs after irradiation, nostudy has as yet shown improved organ function afterradiation with clinically relevant doses. In the current study,we have assessed the ability of G-CSF to stimulate salivarygland regeneration by promoting mobilization of bonemarrow cells to the irradiated tissue. The presence of eGFPand male Y chromosome–positive cells in irradiated glandswas clearly enhanced by G-CSF treatment. The accompanyingmorphologic and functional improvements, however, areunlikely to be related to bone marrow–derived stem celltransdifferentiation.
The present study provides the first evidence that mobiliza-tion of bone marrow cells after salivary gland irradiation hasbeneficial effects in preventing/reducing radiation-induceddamage. Yet, many researchers have observed that bonemarrow cells transplanted in lethally irradiated subjects weredetectable in tissues, such as heart, lung, liver, and skin. Most ofthese studies used radiation doses (V10 Gy) that are necessary
to allow formation of a chimeric hematopoietic system.Although these doses induce damage in numerous tissues, theyare well below the ED50s as described for organ dysfunction inrelation to curative radiotherapy (35). This is of importance asthe number of bone marrow cells that home to the irradiatedorgan is likely to be dependent on the extent of damageinflicted.
In our experimental setup, we used a dose that inflictsdamage to a similar level as what is observed in clinicalpractice (35). We observed a high number of BMCs homingto the salivary glands as well as morphologic and functionalimprovements. Potentially, G-CSF could directly protectsalivary gland stem cells and stimulate surviving stem cellsto divide, resulting in the abundance of acinar cells. It hasbeen shown that G-CSF is able to directly bind to G-CSFRspresent on cardiomyocytes to promote their survival aftermyocardial infarction (42). In addition, Schneider et al. (43)revealed that G-CSF directly protects against programmed celldeath in neurons after ischemia. We did detect the presenceof G-CSF receptors on myoepithelial and endothelial cells.These cells are probably responsible for the very lowexpression of the G-CSF receptor in the gland as determinedby real-time quantitative PCR. However, we were not able todetect the expression of G-CSFR on salivary gland acini orducts, which are suggested to contain the salivary gland stem
Fig. 6. Donor origin of salivary gland cell andcharacterization of BMCs by immunolabeling. Anti-eGFPlabeling (brown) showed BMCs lying around or in closevicinity with Alcian Blue ^ stained mucopolysacharidescontaining acinar cells (A-B). C, a-smooth muscle actin(red) costaining with eGFPon salivary myoepithelialcells (arrow).D, a-smoothmuscle actin (red) costainingwith eGFPonmyofibroblast (arrow). E, CD45 (red)costainingwith eGFPon interstitial cells (arrow). F, CD31(red) costaining with eGFPonmesenchymal andendothelial cells (arrow). Original magnification,�200(A),�400 (B),�1,200 (C, E , and F), and�2,400 (D).Nucleus staining is seen by DAPI (C-F).
Cancer Therapy: Preclinical
www.aacrjournals.orgClin Cancer Res 2006;12(6) March15, 2006 1810
Research. on May 17, 2018. © 2006 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

cell (44) and thus normally will not be able to respond toG-CSF.
Possibly, irradiation induces up-regulation of the G-CSFreceptor as has been shown to occur on neurons in the brainafter ischemia (43). Irradiation, however, did not enhance theexpression of G-CSF receptors in the salivary glands. Further-more, our treatment was done rather long after irradiationdamage was inflicted (4), and, if anything, we observed a ratherenhanced beneficial effect of G-CSF after delayed application(10 versus 30 days after irradiation). This is in contrast to theobserved reduction in beneficial effect on cardiac function bydelayed start of G-CSFR stimulation (42). Taken these findingstogether, it seems unlikely that G-CSF directly acts on acinarcells in our experimental setup.
The question remains as to what is responsible for theimprovement of salivary glands function after irradiation andG-CSF treatment. In view of the very low (if any) numbers ofsalivary gland cells found to be potentially derived from thebone marrow (mostly myoepithelial and fibroblasts), it seemsthat ‘‘transdifferentiation’’ of bone marrow cells to acinarsalivary gland cells is not observed.
However, still many BMCs resided in the salivary gland longafter irradiation and mobilization. Most of these cells wereinterstitial and CD45�, indicating that they lost epitopescharacteristics of hematopoietic cells. This is in agreement withresults described in a study using an ischemia/reperfusionmodel of kidney damage (45). Moreover, like us, they alsoshowed the presence of bone marrow–derived myofibroblastscells in the interstitium, which is also known to occur after aradiation insult (27).
The finding that some of the interstitial GFP+ cells were alsoCD31+ indicates that these cells are of mesenchymal cell origin.Thus, apparently, G-CSF also induces the mobilization of these
cells from the bone marrow in the irradiated salivary glands.Indeed, it is known that next to hematopoietic stem cells, G-CSF also induces the mobilization of a number of mesenchy-mal progenitor cells (46), which may even home to salivaryglands in healthy mice (47). Therefore, one can postulate thatmesenchymal cells are involved in the amelioration ofradiation-induced damage to the salivary gland.
Because the percentage of donor-derived bone marrow cellstransdifferentiated in salivary gland cells is at the best verylow, we must assume that G-CSF–induced mobilization andhoming of hematopoietic and/or mesenchymal cells somehowstimulates the recovery of the salivary gland cells. The mostlikely hypothesis is that this occurs through cytokine secretionand subsequent induction of proliferation of the endogenoustissue progenitor/stem cell. A study with long-term follow-upwill need to be done to reveal whether the effects observed aretransient or long lasting.
In summary, our data indicate that G-CSF treatment iseffective in ameliorating radiation-induced salivary glanddysfunction after radiotherapeutic treatment of head and neckcancer, likely via homing of bone marrow–derived cells, whichsubsequently secretion of stimulatory or protective factors. Thisstudy indicates a clinical applicable protocol for the use of BMCmobilization to ameliorate radiation-induced damage. To beable to exploit this treatment most effectively, the molecularmechanism behind the observed protection and duration needsto be further explored.
Acknowledgments
We thank Jane Briggs of the Pathology Department of the University MedicalCenter Groningen for advice on in situ hybridization techniques and Bert Dontje ofthe Stem Cell Department for performing the splenectomy.
Fig. 7. Expression of g-csfr in mouse submandibulargland by reverse transcription-PCR andimmunohistochemistry. A, expression ofg-csfr wasdetected by reverse transcription-PCR inmouse bonemarrow (lane 2) as a positive control. Lane 3, g-csfr canonly slightly be detected with reverse transcription-PCRin the submandibular glandofnormalmice. Lane 4,wateris used as a negative control to exclude the possibilityfrom contamination; lane1, marker. B, anti-G-CSFRlabeling (brown) detected in the salivary gland, 30 daysafter irradiation.The receptor is present onmyoepithelial(C, arrow) and endothelial cells (D, arrow), visualizedby costaining of G-CSFR and a-smoothmuscle actin.Nuclei (blue). Original magnification,�200 (B) and�630 (C-D).
Amelioration of Radiation-Induced Damage
www.aacrjournals.org Clin Cancer Res 2006;12(6)March15, 20061811
Research. on May 17, 2018. © 2006 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

Cancer Therapy: Preclinical
www.aacrjournals.orgClin Cancer Res 2006;12(6) March15, 2006 1812
References1. Konings AW, Coppes RP,Vissink A. On the mecha-nism of salivary gland radiosensitivity. Int J RadiatOncol Biol Phys 2005;62:1187^94.2. Vissink A, Jansma J, Spijkervet FK, Burlage FR,Coppes RP. Oral sequelae of head and neck radiother-apy. Crit Rev Oral Biol Med 2003;14:199^212.3. Vissink A, Burlage FR, Spijkervet FK, Jansma J,Coppes RP. Prevention and treatment of the conse-quences of head and neck radiotherapy. Crit Rev OralBiol Med 2003;14:213^25.4. Coppes RP, Zeilstra LJ, Kampinga HH, Konings AW.Early to late sparing of radiation damage to the parotidgland by adrenergic and muscarinic receptor agonists.Br JCancer 2001;85:1055^63.5. Kuehnle I, Goodell MA. The therapeutic potential ofstem cells from adults. BMJ 2002;325:372^6.6. Cho SW, Lim SH, Kim IK, et al. Small-diameter bloodvessels engineered with bone marrow-derived cells.Ann Surg 2005;241:506^15.7. Adachi Y, Imagawa J, SuzukiY, et al. G-CSF treatmentincreases side population cell infiltration after myocard-ial infractioninmice.JMolCellCardiol2004;36:707^10.8. Deten A,Volz HC, Clamors S, et al. Hematopoieticstem cells donot repair the infarctedmouseheart.Car-diovasc Res 2005;65:52^63.9. Kuethe F, Figulla HR,Voth M, et al. [Mobilization ofstem cells by granulocyte colony-stimulating factorfor the regenerationofmyocardial tissue aftermyocar-dial infarction]. Dtsch MedWochenschr 2004;129:424^8.10.MurryCE,SoonpaaMH,ReineckeH, et al.Haemato-poietic stem cells do not transdifferentiate into cardiacmyocytes in myocardial infarcts. Nature 2004;428:664^8.11. Norol F, Merlet P, Isnard R, et al. Influence of mobi-lized stem cells onmyocardial infarct repair in anonhu-man primatemodel. Blood 2003;102:4361^8.12. Ohtsuka M,Takano H, ZouY, et al. Cytokine therapyprevents left ventricular remodeling and dysfunctionafter myocardial infarction throughneovascularization.FASEB J 2004;18:851^3.13. Orlic D, KajsturaJ, Chimenti S, et al. Mobilized bonemarrow cells repair the infarcted heart, improvingfunction and survival. Proc Natl Acad Sci US A 2001;98:10344^9.14. Orlic D, Kajstura J, Chimenti S, et al. Bone marrowcells regenerate infarcted myocardium. Nature 2001;410:701^5.15. Stamm C,Westphal B, Kleine HD, et al. Autologousbone-marrow stem-cell transplantation for myocardialregeneration. Lancet 2003;361:45^6.16. BrazeltonTR, Rossi FM, Keshet GI, Blau HM. Frommarrow to brain: expression of neuronal phenotypesin adult mice. Science 2000;290:1775^9.17. Eglitis MA, Mezey E. Hematopoietic cells differen-tiate into both microglia and macroglia in the brainsof adult mice. Proc Natl Acad Sci U S A 1997;94:4080^5.18. PrillerJ. Robert FeulgenPrize Lecture. Grenzganger:
adult bonemarrowcells populate the brain. HistochemCell Biol 2003;120:85^91.19. Lagasse E, Connors H, Al Dhalimy M, et al. Purifiedhematopoietic stem cells can differentiate intohepato-cytes in vivo. Nat Med 2000;6:1229^34.20. Yannaki E, Athanasiou E, Xagorari A, et al. G-CSF-primed hematopoietic stem cells or G-CSF per se ac-celerate recovery and improve survival after liver injury,predominantly by promoting endogenous repair pro-grams. Exp Hematol 2005;33:108^19.21.TheiseND,NimmakayaluM,GardnerR,etal.Liverfrombonemarrow inhumans.Hepatology2000;32:11^6.22. Theise ND, Badve S, Saxena R, et al. Derivation ofhepatocytes frombonemarrow cells inmice after radi-ation-induced myeloablation. Hepatology 2000;31:235^40.23. Herrera MB, Bussolati B, Bruno S, Fonsato V,Romanazzi GM, Camussi G. Mesenchymal stem cellscontribute to the renal repair of acute tubular epithelialinjury. IntJMol Med 2004;14:1035^41.24.NishidaM, Fujimoto S,Toiyama K, Sato H, HamaokaK. Effect of hematopoietic cytokines on renal functionin cisplatin-induced ARF in mice. Biochem BiophysRes Commun 2004;324:341^7.25. PoulsomR, Forbes SJ, Hodivala-Dilke K, et al. Bonemarrow contributes to renal parenchymal turnover andregeneration. JPathol 2001;195:229^35.26. Abe S, Boyer C, Liu X, et al. Cells derived from thecirculation contribute to the repair of lung injury. AmJRespir Crit Care Med 2004;170:1158^63.27. Epperly MW, Guo H, Gretton JE, Greenberger JS.Bone marrow origin of myofibroblasts in irradiationpulmonary fibrosis. Am J Respir Cell Mol Biol 2003;29:213^24.28. Ishizawa K, Kubo H, Yamada M, et al. Bonemarrow-derived cells contribute to lung regenerationafter elastase-induced pulmonary emphysema. FEBSLett 2004;556:249^52.29. Kotton DN, Ma BY, Cardoso WV, et al. Bonemarrow-derived cells as progenitors of lung alveolarepithelium. Development 2001;128:5181^8.30. Yamada M, Kubo H, Kobayashi S, et al. Bonemarrow-derived progenitor cells are important for lungrepair after lipopolysaccharide-induced lung injury.J Immunol 2004;172:1266^72.31. Borue X, Lee S, GroveJ, et al. Bonemarrow-derivedcells contribute to epithelial engraftment duringwound healing. AmJPathol 2004;165:1767^72.32. Minatoguchi S, Takemura G, Chen XH, et al.Acceleration of the healing process and myocardialregeneration may be important as a mechanism ofimprovement of cardiac function and remodeling bypostinfarction granulocyte colony-stimulating factortreatment. Circulation 2004;109:2572^80.33. KawadaH, FujitaJ, Kinjo K, et al. Nonhematopoieticmesenchymal stem cells can be mobilized and differ-entiate into cardiomyocytes after myocardial infarc-tion. Blood 2004;104:3581^7.34. Coppes RP, Vissink A, Zeilstra LJ, Konings AW.
Muscarinic receptor stimulation increases tolerance ofrat salivary gland function to radiation damage. IntJRadiat Biol1997;72:615^25.35. Coppes RP,Vissink A, Konings AW. Comparisonof radiosensitivity of rat parotid and submandibularglands after different radiation schedules. RadiotherOncol 2002;63:321^8.36.Vissink A, ’s-Gravenmade EJ, Ligeon EE, KoningsWT. A functional and chemical study of radiationeffects on rat parotid and submandibular/sublingualglands. Radiat Res1990;124:259^65.37.Vissink A, Konings AW, Ligeon EE, ’s-GravenmadeEJ. Irradiation-induced changes in secretion andcomposition of rat saliva. J Biol Buccale 1990;18:3^8.38. Coppes RP, Zeilstra LJ, Vissink A, Konings AW.Sialogogue-related radioprotection of salivary glandfunction: the degranulation concept revisited. RadiatRes1997;148:240^7.39. Zeilstra LJ, Vissink A, Konings AW, Coppes RP.Radiation induced cell loss in rat submandibular glandand its relation to gland function. Int J Radiat Biol2000;76:419^29.40.Wierenga PK, Setroikromo R, Vellenga E, KampingaHH. Purging of acute myeloid leukaemia cells fromstem cell grafts by hyperthermia: enhancement of thetherapeutic index by the tetrapeptideAcSDKP and thealkyl-lysophospholipid ET-18-OCH(3). Br JHaematol2000;111:1145^52.41. Lin AL, Johnson DA,WuY,Wong G, Ebersole JL,Yeh CK. Measuring short-term gamma-irradiationeffects on mouse salivary gland function using a newsaliva collection device. Arch Oral Biol 2001;46:1085^9.42. Harada M, QinY,Takano H, et al. G-CSF preventscardiac remodeling after myocardial infarction by acti-vating the Jak-Stat pathway in cardiomyocytes. NatMed 2005;11:305^11.43. Schneider A, Kruger C, Steigleder T, et al. Thehematopoietic factor G-CSF is a neuronal ligand thatcounteracts programmed cell death and drives neuro-genesis. JClin Invest 2005;115:2083^98.44. Denny PC, Denny PA. Dynamics of parenchymalcell division, differentiation, and apoptosis in theyoung adult female mouse submandibular gland. AnatRec1999;254:408^17.45. Lin F, Moran A, Igarashi P. Intrarenal cells, not bonemarrow-derived cells, are the major source for regen-eration in postischemic kidney. JClin Invest 2005;115:1756^64.46. Lapidot T, Petit I. Current understanding of stemcell mobilization: the roles of chemokines, proteolyticenzymes, adhesion molecules, cytokines, and stromalcells. Exp Hematol 2002;30:973^81.47. Luttichaux IV, Notohamiprodjo M,WechselbergerA, et al. Human adult CD34- Progenitor Cells Func-tionally Express the Chemokine Receptors CCR1,CCR4, CCR7, CXCR5, and CCR10 but Not CXCR4.Stem Cells Dev 2005;14:329^36.
Research. on May 17, 2018. © 2006 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

2006;12:1804-1812. Clin Cancer Res Isabelle M.A. Lombaert, Pieter K. Wierenga, Tineke Kok, et al. Damage to Salivary GlandsColony-Stimulating Factor Ameliorates Radiation-Induced Mobilization of Bone Marrow Stem Cells by Granulocyte
Updated version
http://clincancerres.aacrjournals.org/content/12/6/1804
Access the most recent version of this article at:
Cited articles
http://clincancerres.aacrjournals.org/content/12/6/1804.full#ref-list-1
This article cites 47 articles, 9 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/12/6/1804.full#related-urls
This article has been cited by 5 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://clincancerres.aacrjournals.org/content/12/6/1804To request permission to re-use all or part of this article, use this link
Research. on May 17, 2018. © 2006 American Association for Cancerclincancerres.aacrjournals.org Downloaded from


















