
Mitteilungen - vdhk.de · types and especially skull shape morphology in combination with bone...
40
Mitteilungen des Verbandes der deutschen Höhlen- und Karstforscher e.V. München ISSN 0505-2211 H 20075 Nr. 1/2013 Jahrgang 59 1. Quartal
Transcript of Mitteilungen - vdhk.de · types and especially skull shape morphology in combination with bone...

Mitteilungendes Verbandes
der deutschen Höhlen- undKarstforscher e.V. München
ISSN 0505-2211H 20075 Nr. 1/2013 Jahrgang 59
1. Quartal

Verband der deutschen Vom Finanzamt München für Körperschaften als gemeinnützig anerkannt gem. Bescheid v. 21.8.2007) (Steuer-Nr. 9143/000223305542
Höhlen- und Karstforscher e.V.
Sitz in München
München, 10. November 2012 An alle Mitglieder des VERBANDES DER DEUTSCHEN HÖHLEN- UND KARSTFORSCHER E.V., München
Einladung
zur 58. Jahreshauptversammlung 2013 in Giengen-Hürben, Schwäbische Alb
Werte Mitglieder, liebe Höhlenfreunde,
der VERBAND DER DEUTSCHEN HÖHLEN- UND KARSTFORSCHER E.V. lädt zur 58. ordentlichen Jahreshauptver-sammlung am Samstag, den 1. Juni 2013 in die Hürbetalhalle, Adalbert-Stifter-Str.1, 89537 Giengen-Hürben, herzlich ein. Die Hauptversammlung beginnt um 8.30 Uhr. Ausgabe der Stimmzettel um 8 Uhr. Tagesordnung
1. Eröffnung der Versammlung; Feststellung der Beschlussfähigkeit; Wahl der Versammlungsleitung, der Protokollführung und Bestimmung der Stimmenzähler; Genehmigung des Protokolls der HV 2012
2. Bericht der Vorsitzenden, des stellvertretenden Vorsitzenden, des Geschäftsführers und des Verwal-ters des Bergungskosten-Solidaritätsfonds
3. Bericht des Schatzmeisters über das Geschäftsjahr 2012 4. Bericht der Rechnungsprüfer 5. Entlastung des Vorstandes 6. Neuwahl der Rechnungsprüfer für das Geschäftsjahr 2012 7. Berichte der Referenten und der Landesverbände 8. Neuwahl der Referenten und des Vertreters der Einzelmitglieder 9. Beschlussfassung über Anträge an die Hauptversammlung 10. Beratung über Ort und Termin der Hauptversammlung 2014 11. Festlegung der Jahresbeiträge 2014 12. Sonstiges 13. Mitteilungen, Vorschläge, Kritik und Schluss der Hauptversammlung.
Die diesjährige Hauptversammlung findet im Rahmen der 53. Jahrestagung des VdHK in Giengen-Hürben statt. Am Freitag, den 31. Mai 2013 findet um 20 Uhr die Sitzung des Beratenden Ausschusses statt. Da-zu gehören lt. § 18 der Satzung die Vorstandsmitglieder und deren Stellvertreter, alle Referenten, je ein Mitglied der Landesverbände und der Vertreter der Einzelmitglieder. Der Tagungsort wird per Aushang am Tagungsbüro bekannt gemacht. Alle Ausschussmitglieder sind zur zahlreichen Teilnahme herzlich einge-laden. Zur Sitzung des Ausschusses sind laut HV-Beschluss interessierte Zuhörer (ohne Rederecht) will-kommen.
Mit freundlichen Grüßen
Die Vorsitzende Der Geschäftsführer
Bärbel Vogel Stephan Marks

3
Mitteilungendes Verbandes der deutschenHöhlen- und Karstforscher e.V.München
ISSN 0505-2211, Jahrgang 59, Nr. 1München, 15.3.2013
Inhalt
Copy- Verband der deutschen Höhlen- und Karstforscherright: e.V. München (VdHK)
Schrift- Michael Krause, Jägerhalde 118, 70327 Stuttgartleitung: Tel 0711 / 47 25 98, Fax 032 22 37 28 155,
E-Mail [email protected] (mk)
Dr. Friedhart Knolle, Grummetwiese 16, 38640Goslar, Tel 05321 / 20 281, [email protected] (fk)
Druck u. Oberharzer Druckerei Fischer & Thielbar GmbH,Versand: Alte Fuhrherrenstr. 5, 38678 Clausthal-Zellerfeld.
Der Verband der deutschen Höhlen- und Karstforscher e.V. istals gemeinnützig anerkannt (Finanzamt für KörperschaftenMünchen, Steuernummer 143/223/30554 gem. Bescheid vom6.7.2011).
Bankkonto (auch Volksbank Laichingenfür Spenden): (BLZ 630 913 00) Kto. 1 492 004
Nachdruck oder Veröffentlichung und Verbreitung in elek-tronischen Medien, auch auszugsweise, nur mit schriftlicherGenehmigung der Schriftleitung.
Erscheinungsweise: 4 x jährlich
Bezugspreis: im Mitgliedsbeitrag inbegriffen; Abo: 20 Euro/Jahr
Zugelassen zum Postzustellungsdienst für die Versendung alsStreifbandzeitung (Vertriebskennzeichen H 20075 F).
Die Redaktion behält sich Kürzung und Bearbeitung vonBeiträgen vor. Durch Einsendung von Fotografien und Grafikenstellen die Autoren den VdHK von Ansprüchen Dritter frei.
Der Verband im Internethttp://www.vdhk.deBitte lesen Sie regelmäßig die dort bekanntgegebenen Veran-staltungstermine.
Abo der VerbandsmitteilungenAbonnements der Verbandsmitteilungen – auch als Geschenk!– für 20 Euro/Jahr (incl. Porto/Verpackung) über: LeonhardMährlein, Idealweg. 11, 90530 Wendelstein, Tel. 09129/8428,[email protected]. Das Abonnement gilt jeweilsfür Heft 1-4 eines jeden Jahrganges.
CAJUS G. DIEDRICH:Evolution, Horste, Taphonomie und Prädatoren derRübeländer Höhlenbären, Harz (Sachsen-Anhalt) .................... 4TätigkeitsberichteJahresbericht 2012 von AK Biospeläologie undReferat Biospeläologie ................................................................... 30Von Veranstaltungen ..................................................................... 32Filmtipp ........................................................................................... 32JHV Giengen-Hürben 2013Anmeldeformular .......................................................................... 33Programm ....................................................................................... 34HöhlenfotografieInteressantes für Höhlenfotografen: Funkkoppler stattOptokoppler ................................................................................... 38PersonaliaDr. Klaus Eberhard Bleich 80 ...................................................... 38Gesamtinhaltsverzeichnis 2012 .................................................... 39
Titelbild: Höhlenbären-Komposit aus Originalknochen derRübeländer Höhlen nach neuer Restaurierung in der Daueraus-stellung des Staatlichen Naturhistorischen Museums Braun-schweig, siehe Abb. 2 B im nachfolgenden Hauptbeitrag dieserAusgabe; Foto: Cajus Diedrich 2012Rückseite: Höhlentier des Jahres 2013
Editorial
Mit dem vorliegenden Heft gehen wir in den 59. Jahrgang undnähern uns damit dem 60. Verbandsgeburtstag am 13. März 2015.Zeit, schon einmal zurückzublicken auf die Geschichte unsererVorstandsmitglieder. Eine entsprechende Liste erstellt uns derzeitHans-Martin Luz – sie erscheint in einer der nächsten Ausga-ben.Schwerpunktthema des vorliegenden Hefts ist die Paläontologieder klassischen Rübeländer Höhlen – die Ergebnisse der Unter-suchungen von Dr. Cajus Diedrich sind so interessant, dass wiruns entschlossen, sie hiermit als erste Zeitschrift in Form einesetwas längeren Beitrags zu veröffentlichen.2013 ist ein Jahr interessanter Tagungen – vom 30.5.-2.6. findetunsere 58. Jahreshauptversammlung im Rahmen der 53. Jahres-tagung in Giengen-Hürben auf der Schwäbischen Alb statt, sie-he Programm und Anmeldeformular in diesem Heft. Im Juli folgtvom 21.-28.7. der 16. Internationale Kongress für Höhlen-forschung der UIS in Brno (Brünn), Tschechische Republik, sie-he auch www.speleo2013.com. Weitere Tagungen finden sich imTerminkalender auf www.vdhk.de.fk
Corrigenda
Heft 2/2012: Das Foto von Klaus Cramer auf S. 59 stammt vonAndreas Wolf.Heft 3/2012: Auf S. 84 sollte es im Fledermausbeitrag heißen:Myotis bechsteinii.Heft 4/2012: In der Einladung zur JHV sollte es heißen: 58.Jahreshauptversammlung; im Zitat auf S. 104 muss es statt LESLIE
et al. (2011) heißen: MELIM et al. (2011).

4
Mitt. Verb. dt. Höhlen- u. Karstforscher 59 (1) 4-29 München 2013
KurzfassungNeue Feldstudien und die vorläufige Aufarbeitung von 8.222Knochen aus den beiden großen Bärenhorst-Höhlen Baumanns-höhle und Hermannshöhle in Rübeland (Harz, Sachsen-Anhalt)erlauben die Rekonstruktion der Höhlennutzungen unterschied-licher Höhlenteile und von vier verschiedenen Höhlenbären-arten/-unterarten zwischen der mittleren Saale- bis mittlerenWeichsel-Kaltzeit. Diese datieren relativ-chronologisch mit denPrämolar-Zahnmorphologie-Typen und besonders der Schädel-morphologie in Kombination mit der Knochenerhaltung undProfilaufnahmen die Nutzung der Baumanns- und Hermanns-höhle in Rübeland auf unterschiedlichen Höhlenniveaus. DieLangknochenproportionen unterstützen die Separierung in un-terschiedliche Arten bzw. Geschlechter innerhalb einer Art. Dievier Haupt-Ponorhöhlen-Etagen wurden zwischen der (?Cromer)Elster-Eiszeit und Holozän gebildet, Bären mit Ursus deningerinutzen diese erst ab der mittleren Saale-Eiszeit, wobei der ältes-te Horstbereich in der Baumannshöhle im Bereich des Goethe-saals lag. Die Knochen wurden aber noch in der Saale-Eiszeitfluvial (Einfluss des Vorfluters Bode) in tiefere Vertikalschächtedes östlichen Höhlensystems („Alte Höhle“) verfrachtet. GegenEnde der Saale-Eiszeit wurde ein heute noch unbekannter östli-cher Höhlenteil („Neue Höhle“-Bereich) als Horst genutzt, indem Höhlenbären Ursus spelaeus eremus/spelaeus bis in die mittlereWeichsel-Kaltzeit datieren. Auch diese wurden fluvial, aber nurmit lokalen Kames-/Seitenmoränen-Kalkkies/Löss-Sandab-lagerungen, ebenfalls in tiefere Höhlenbereiche verfrachtet, aberim Westen der Baumannshöhle („Neue Höhle“). Dies geschahdurch Gletscher-Oberflächenwasser des Bodetal-Gletschers, derzur Hauptvereisung (= LGM, Spät-Weichsel) die Höhlen pas-sierte. In der Früh-Mittel-Weichsel-Kaltzeit nutzen die selbenHöhlenbären-Unterarten auch die Hermannshöhle. Der Bären-horst lag dort in der saale-eiszeitlich gebildeten Ponorhöhle derOberen Etage (bzw. in den darüberliegenden älteren Relikt-etagen). Höhlenbärenschliff und teilartikulierte Skelette sowieautochthone Knochenlager beweisen die nicht-fluviale Um-lagerung, da der aktive Weichsel-Höhlenfluss 8-10 m tiefer ent-wässerte. Nur in der Hermannshöhle nutzten die größten Höh-lenbären Ursus ingressus diese als Horst, da die Baumannshöhleoffensichtlich bereits auch im zweiten Eingang zum Ende dermittleren Weichsel-Kaltzeit verbrochen war. Zwei neonate (=ungeborene) Höhlenbärenskelette aus der Hermannshöhle undviele Einzelknochen von Totgeburten sind in allen Bärenhorst-Schichten keine Seltenheit. Auch finden sich Pathologien an ver-schiedenen Bärenarten. Die Großprädatoren der Bären datierenebenfalls zwischen mittlere Saale bis Mittel-Weichsel. Die größ-ten Löwen aller Zeiten Panthera leo fossilis (Saale) stammen nuraus der Baumannshöhle. Spätpleistozäne Panthera leo spelaea-Res-te sind vertreten durch kleinere Eem-warmzeitliche und größereWeichsel-kaltzeitliche Formen in der Baumannshöhle (Knochen-feld/Ochsenhang-Bereich). In der Hermannshöhle fanden sichhingegen Reste zweier adulter und seniler, teilweise pathologi-scher Löwenskelette 90 m tief in der Saal-Halle. Diese Eiszeit-
Löwen sind immer mit 2-4,5 % in Bärenhorsten vertreten. Siegingen nur zur Bärenjagd tief in Höhlen, lebten dort aber nicht.Weitere Saale- (bis Früh-Weichsel)-Eiszeit-Katzen sind die Res-te des Leoparden Panthera pardus subsp., die wiederum nur ausdem Knochenfeld/Ochsenhang der Baumannshöhle stammen.Dieser hatte anscheinend einen Horst in noch unentdecktenHöhlenteilen. Von dort stammen auch etliche Knochen des Wild-hundes Cuon alpinus fossilis, der ebenfalls in die Saale-Eiszeit da-tiert, aber dort keinen Horst hatte und als weiterer sporadischerHöhlenbären-Restkadaververwerter angesehen wird. In der Her-mannshöhle hingegen finden sich die Reste des großen Eiszeit-wolfes Canis lupus spelaeus besonders im vorderen Bereich (Bären-höhlen-Halle) zum Eingang hin. Dort sind auch Reste der eis-zeitlichen Fleckenhyäne Crocuta crocuta spelaea bekannt, die of-fensichtlich zusammen mit Wölfen den ehemaligen Eingang auchsporadisch als Horst nutzte und einige Beutetierreste von Mam-mut, Wollnashorn, Rentier und sogar Steinbock einschleppte.Diese beiden Raubtiere sind die Haupt-Bissspuren-Verursacheran Bärenknochen (weniger Löwen), wobei nur die Hyänen er-folgreich auch die Langknochen der Höhlenbären zerknackenkonnten. Über 80 % der Jungbärenknochen (bis 1 JahrIndividuenalter) sind an den Gelenken abgefressen oder verbis-sen, aber weniger zerknackt (noch zu weich). Knochen adulterHöhlenbären sind mit bis zu 20 % in der Hermannshöhle be-schädigt. Gleichartige Fraßreste finden sich aber in allen Bären-horsten zu allen unterschiedlichen Zeiten. Höhlenbären warenin den Borealwäldern der Mittelgebirge in der Saale- und Weich-sel-Kaltzeit mitunter die Hauptbeute von Hyänen, Wölfen undLöwen, da die Mammutsteppen-Großwildfauna dort abwesendwar oder nur gelegentlich die Täler aus dem Harzvorland her-aufzog, wie auch innerhalb des Bodetals (besonders Rentiere).Zur „Zeit der Höhlenbärenhorste“ müssen im Harz gemischteBorealwald/Alpen-Landschaften vorhanden gewesen sein, wor-auf z.B. die Präsenz von Steinböcken und Gemsen hindeutet.
AbstractNew field work and preliminary studies on 8.222 bones fromthe two large cave bear dens Baumannshöhle and Hermanns-höhle of Rübeland (Harz Mts., Saxony-Anhalt, Germany) allowthe reconstruction of the cave uses in different cave areas byfour different cave bear species/subspecies between the middleSaalian to middle Weichselian Glacial. Those allow to date rela-tive-chronologically the use of the Baumannshöhle and Her-mannshöhle in different cave levels by premolar teeth morpho-types and especially skull shape morphology in combination withbone preservation and new section studies. The long-boneproportions support the separation between different species,or sexes within one species. The four main ponor cave elevationswere formed between the (?Cromerian) Elsterian and Holocene.Cave bears with Ursus deningeri started to use since the middleSaalian first the Goethesaal Hall area of the Baumannshöhle.Their bones were fluvially transported (Bode River impact) alsoin deeper vertical shafts within the cave during the Saalian already,
Evolution, Horste, Taphonomie und Prädatoren der Rübeländer Höhlenbären,Harz (Sachsen-Anhalt)
vonCAJUS G. DIEDRICH

5
but only in the eastern cave area („Alte Höhle“ area). At the endof the Saalian Glacial a new opened but today undiscovered cavepart („Neue Höhle“ area) was used as cave bear den by Ursusspelaeus eremus/spelaeus until the middle Weichselian. Also thosebones were fluvially transported together with local kames/late-ral moraine-limestone gravels/loess sand again into deeper caveareas, but in the eastern part of the Baumannshöhle („NeueHöhle“ area) by surface waters of the Bode Valley Glacier, thatpassed the caves during the maximal glaciation (= LGM, LateWeichselian). Within the early/middle Weichselian Glacial, thesame cave bear subspecies used the Hermannshöhle. The cavebear den was situated in the Saalian-formed ponor cave elevationof the upper level (and the above older relict levels). Cave bearpolish and partly articulated skeleton remains, such asautochthonous bonebeds, indicate non-fluvial transport, becausethe active Weichselian cave river drained 8-10 m below. Only inthe Hermannshöhle the largest cave bears Ursus ingressus used itas den, because the Baumannshöhle was obviously alreadyblocked also in the second entrance at the end of the middleWeichselian. Two neonate (= unborn) cave bear skeletons ofthe Hermannshöhle and many reworked single bones of deadlyborn are not rare in all cave bear den sediments. Also pathologiesare found on several different cave bear species. The largepredators of cave bears date again between middle Saalian tomiddle Weichselian. The largest lions Panthera leo fossilis (Saalian)were found only in the Baumannshöhle. Late Pleistocene Pantheraleo spelaea remains are from small Eemian Interglacial but alsolarger Weichselian Glacial forms and deposits of the Knochen-feld/Ochsenhang in the Baumannshöhle. In the Hermannshöhle,only two skeleton remains of adult to senile pathological lionshave been discovered 90 m deep within the Saal Hall. The IceAge lions always take 2-4.5 % of the bone amount in cave beardens. They went only for cave bear hunting purposes deep intothe caves, but never lived therein. Further Saalian up to earlyWeichselian Ice Age cats are the Leopard Panthera pardus subsp.which remains again come only from the Knochenfeld/Ochsen-hang of the Baumannshöhle. Those obviously had a lair in anundiscovered cave part. Found here were also several wild dogCuon alpinus fossilis remains dating again into the Saalian Glacial.Those had no sporadic den in the unknown cave part, and areseen as further sporadic bear scavengers. In the Hermannshöhleinstead, remains of the large Ice Age wolf Canis lupus spelaeushave been found especially in the anterior part (Bärenhöhle Hall)closer to the former entrance. There, additionally, few Ice Agespotted hyena Crocuta crocuta spelaea bones are known. Those andthe wolf remains indicate a den use in the entrance area by bothfor short term periods. They must have imported remains ofmammoth, woolly rhinoceros, reindeer and ibex. Those twocarnivores are the main bite mark producers (less lions) on cavebear bones. Only hyenas were able to crush adult cave barlongbones. About 80 % of the cub bear bones (up to one year inindividual age) have chewed joints or expose bite damage, butare much less cracked (too soft). Bones of adults (also Hermanns-höhle) are up to 20 % damaged. Similar chewed/damaged bonesare found in all cave bear dens of different time periods. Cavebears were in boreal forests of the middle high elevated mountainregions during the Saalian and Weichselian the main prey ofhyenas, wolves and lions, because of absence of the mammothsteppe game. Those were only sporadic migrating from the HarzMountain Fore Lowlands upstream within the river valleys, suchas the Bode Valley (especially reindeers). At the „time of thecave bear dens“ mixed boreal forest/alpine landscapes must have
been present in the Harz Mountain indicated by ibex and cha-mois remains.Keywords: Höhlenbären-Arten/Unterarten, „Höhlenbären-Uhr“, Bärenhorste, Datierungen, Pathologien, neonate Skelette,Bissspuren, große Raubtiere (Panthera, Crocuta, Canis, Cuon),Faunenkomplexe, Rübeländer Höhlen, Harz-Gebirge, Sachsen-Anhalt.
EinleitungTopographie der Rübeländer HöhlenDie beiden größten norddeutschen eiszeitlichen Höhlenbären-horste, die Baumannshöhle und Hermannshöhle, befinden sichinmitten des Harz-Gebirges (180 x 30 km Ausdehnung) ca. 15 kmBodetal-abwärts vom Brocken (1.141 m), der aus paläozoischenGraniten aufgebaut ist (KNOLLE et al. 1997). Die Höhlen selbstbefinden sich im Bereich des unter-devonischen Massenkalks(Stromatoporen-Riffkalke), dem Elbingeröder Komplex (WEL-LER 1991, 2011). Beide Schauhöhlen liegen mit ihren damaligenund heutigen Eingängen jeweils gegenüber im Bodetal.Die Baumannshöhle, mit den ersten entdeckten Teilen (= „AlteHöhle“) und neu entdeckten Bereichen (= „Neue Höhle“), hateine Gesamtganglänge von 1.980 m und ist in vier Haupt-stockwerken in Höhlenlagen zwischen 384-443 m NHN ange-legt. Der alte Eingang zur ältesten Höhlenbärenhorst-Zeit (=Mittel-Pleistozän) liegt auf 435 m NHN, wobei der teilweisekünstliche heutige Ausgang (parallel zum tatsächlichen und ver-schütteten jüngeren der Spät-Saale/bis Mittel-Weichsel) auf423 m NHN liegt. Der höchste Punkt oberhalb der Höhle liegtbei 445-450 m NHN (FRICKE & KNOLLE 1999 und HöhlenkatasterArGeKH).Die Hermannshöhle ist über 3.300 m lang (z.Zt. noch laufendeNeuvermessung durch ArGeKH und HFG Leipzig) und hatebenfalls vier Hauptetagen (zwischen 378-420 m NHN), vondenen die beiden untersten aber eher kurz und reliktartig ausge-bildet sind. Auch hier ist die saaleeiszeitlich angelegte Etageebenfalls die größte und längste. Auch in dieser Höhle liegt deraktive Ponor nur in der untersten Etage (FRICKE & KNOLLE 1999und Höhlenkataster ArGeKH).
Historie der Grabungen in den HöhlenBaumannshöhle – Der Goethesaal (früher = Tanzsaal) undangrenzende Bereiche der „Alten Höhle“ (älterer entdeckter Teil)scheinen bereits schon seit dem 16. Jh. erwähnt und stark durch-gegraben worden zu sein – man war auf der Suche nach „Dra-chen- oder Einhornknochen“ (KEMPE 2006). Erste Höhlenbären-fundabbildungen, die damals noch nicht als solche erkannt wur-den, stammen von KUNDMANN (1737), Abb. 1. Die „Neue Höh-le“ als Fortsetzung wurde erst 1888 entdeckt und dann verbun-den. Genau im Schuttkegel, der im Zuge des Wegebaus durch-geschachtet wurde, kam die „Rentierfauna“ mit den Vielfraß-resten von BLASIUS (1890, 1901a) zu Tage (Abb. 1). Die Höhlen-bären, Löwen, Leoparden, Wildhunde, Hyänen und Pferdesprin-ger sowie andere Tierreste und mittelpaläolithische Artefakte(Neandertaler) stammen aus der Knochenfeld-Halle und demzum Ausgang hin nachfolgenden Schuttkegel der Ochsenhang-Halle (BLASIUS 1901a, Abb. 3). Nach KLOOS und BLASIUS fandenkeine bekannten Knochengrabungen mehr statt, wobei bei denBefahrungen 2012 festgestellt wurde, dass auch im Tiefen Gangoder oberhalb des Schuttkegels neben der Knochenfeld-Halleintensiver gegraben worden ist.Hermannshöhle – Hermann GROTRIAN begann erste Ausgra-bungen in der Hermannshöhle nach der Entdeckung 1866

6
besonders in der Bärenhöhlen-Halle, die später als „Bären-friedhof“ bezeichnet wurde (GROTRIAN 1878). 1887 setzte KLOOS
sehr umfangreiche Ausgrabungen in diesem vorderen Höhlen-bereich (Bärenhöhlen-Halle bis heutiger Ausgang) fort (KLOOS
& MÜLLER 1889). BLASIUS (1901b) durchgrub auf der Suche nachVogelknochen den Schuttkegel des damaligen Eingangs (ca. 8 mvom Eingang entfernt) im Bereich der ersten kleinen Halle, diehier als „Kloos-Raum“ bezeichnet wird (heute liegt der künstli-che Ausgangstunnel daneben, Abb. 4). 1961 öffnete STEINER dreiGrabungsstellen, zwei Schnitte im südlichen Seitengang derBärenhöhlen-Halle und eine kleine Fläche in der Saal-Halle(STEINER & STEINER 1969), deren Positionen daher genau be-kannt sind. Schließlich grub VÖLKER in der Saal-Halle neben derGrabung von STEINER (VÖLKER 1984/85). 2012 wurden mehre-re lokal zusammengetragene Knochenhaufen an verschiedenenStellen der Hermannshöhle entnommen, gewaschen und be-stimmt. Aus dem oberflächigen Schutt der damaligen Grabun-gen wurden ebenfalls Knochen vor dem Zertreten gerettet. Die-ses Material von 1.430 genauer bestimmten und ca. 1.000 Frag-menten ist Hauptgegenstand dieser ersten paläontologischenArbeit, die zusammen mit den 2.280 Knochen der VÖLKER -Gra-bung der Saal-Halle und 1.323 Knochen (inkl. 8 Schädel) derSTEINER-Grabung sowie unbekannter Menge der KLOOS-Grabun-gen (ca. 1.500 Knochen, davon über 20 Schädel) eine solide Ba-sis zu einer vorläufigen Übersichts-/Revisions-Auswertung bil-den. Die Befahrung und Bestandsaufnahme in 2012 hat außer-dem ergeben, dass an mehreren Stellen undokumentierte Schür-fe vorhanden sind und dass viele der oberflächennahen Kno-chen (weiß gefärbt und schlecht erhalten) zu Staub zerfallen oderzertrampelt sind.
Abb. 1: A. Portrait von Wilhelm Blasius (1845 - 1912): Professorfür Zoologie und Botanik an der Technischen Hochschule undLeiter des Naturhistorischen Museums in Braunschweig (ehem.Schloss), Pionier der Erforschung der Rübeländer Höhlen, Harz(Foto aus BRANDES 2010). B. Erste Abbildungen eines Höhlen-bären-Zahns aus der Alten Baumannshöhle (aus KUNDMANN 1737,dort identifiziert als „Pferdezahn“).
Paläontologische Erforschung und BemerkungenDie erste Abbildung eines Höhlenbärenzahns aus der „AltenHöhle“ der Baumannshöhle findet sich in KUNDMANN (1737)(Abb. 1). Paläontologische Pionierarbeiten gehen besonders aufden Biologen BLASIUS mit der vorläufigen Auflistung der Vogel-reste aus den Rübeländer Höhlen, primär Hermannshöhle(BLASIUS 1901b), zurück, wie auch grobe Bestimmungen vonNeufunden (BLASIUS 1890), die er letztendlich nicht weiter aus-führte. KLOOS & MÜLLER (1889) erwähnten zwar Höhlenbären,aber letztendlich ging der Geologe nicht auf die Paläontologie
im Detail ein, bildete aber erstmals einen Höhlenbärenschädelund ein „montiertes Skelett“ ab (Abb. 2). Einen Teil seiner Samm-lung gab er nach Braunschweig. Auch dort montierte man einSkelett (Abb. 2 und Titelbild).Alle vorausgehenden Arbeiten haben zwei grundlegende Pro-bleme: Erstens sind zwar Faunenunterschiede grob erkannt undmit damaligen systematischen Namen oft korrekt bestimmt undgelistet worden (besonders gut durch BLASIUS), besonders dieder „Schuttkegel“ und der „Bärenfundstellenbereiche“ (BLASIUS
1901a). Ziel war es, „Kalt-“ von „Warmzeitfaunen“ zu unter-scheiden, nicht aber den Hintergrund, warum Knochen vonwelchen Tieren überhaupt in Höhlen angereichert worden sind.Auch ist bereits grob ihr unterschiedliches Alter zweifach disku-tiert worden wie auch die klimatische Interpretation (BLASIUS
1901a, SCHÜTT 1969a), wobei „nur“ zwei „Faunenkomplexe“ un-terschieden wurden, hier werden es aber mindestens vierKnochenansammlungen („Bone-Assemblages“). Keine der Ar-beiten an Vögeln, Höhlenbären oder anderen Mikro- undMakromammaliern hat quantitative Analysen geliefert (= NISP-Analysen), die viel wichtiger sind als „qualitative Faunenlisten“,da die Knochenanreicherungen (Taphonomie, Höhlenbiologie)damit überhaupt nicht verstanden werden. Diese aber sind derSchlüssel zum Verständnis der Nutzung der Höhlen durch dieTiere in unterschiedlichen Epochen.Selbst die Leoparden wurden von SCHÜTT (1969a) nur gelistetund metrisch analysiert (nicht abgebildet), wobei eine Monogra-phie mit zusätzlichen Neufunden aus 2012 gesondert erscheint(DIEDRICH 2013e).Die Nachbestimmung der Herbivoren aus der Bärenhöhlen-Halle(Hermannshöhle) lassen die von BLASIUS (1901a) und SCHÜTT
(1969a) gelisteten wenigen Reste vom „Reh (Capreolus capreolus)“zu Capra ibex und Rupicapra rupicapra vorläufig bestimmen, wasdamit einen „Eem-Horizont“ dort derzeit ausschließen lässt.Auch die zwei Capra ibex-Reste aus der Baumannshöhle (Ochsen-hang) müssen zeitlich korrigiert werden und sind nicht der LastGlacial Maximum-Fauna (= LGM-Fauna) angehörig (Tab. 1+2).Capra/Rupicapra sind in beiden Faunenvergesellschaftungen mitjeweils wenigen Resten vorhanden.Auch die „Wölfe“, die von SCHÜTT (1969a) als „sehr kleine Ar-ten“ beschrieben wurden, müssen hier größtenteils (alle Fundeaus der Baumannshöhle: Kochenfeld/Ochsenhang) zum Wild-hund Cuon alpinus fossilis revidiert werden. Immerhin hat SCHÜTT
(1969a) die meisten Leopardenreste erkannt (einige sind abervon Cuon) und auf ein mögliches „Interglazial/Interstadial“ hin-gewiesen. Ebenso verwies sie bereits auf die Anwesenheit desmittelpleistozänen Löwen Panthera leo fossilis in der Baumanns-höhle (nannte ihren Titel aber trotz dessen „SpätpleistozäneFauna“), der auch in der Einhornhöhle von ihr im dortigenMaterial erkannt wurde (SCHÜTT 1969b). Von daher verwundertes, dass bis jüngst „alle Knochenfunde der Rübeländer Höhlen“immer noch in das Spätpleistozän gestellt wurden (ROSENDAHL
et al. 2005), was letztendlich hiermit korrigiert werden kann.Die jüngeren Arbeiten von FISCHER über die „Höhlenbären“ derGrabung VÖLKER (FISCHER 1995) und der meisten Löwenknochen(FISCHER 1994) zeigen das weitere Problem der korrekten osteo-logischen Identifizierung vieler Löwenknochen. STEINER hat sogarkeinen der etlichen Löwenknochen aus ihrer Grabung erkannt(STEINER & STEINER 1969). Diese 76 übersehenen Löwenknochenwurden im Zuge der neuen Arbeiten in 2012 erstmals aus den„Bärenknochen“ aus ihrer Sammlung aussortiert. Einige Kno-chen der VÖLKER-Grabung wurden auch von FISCHER (1994,1995) nicht erkannt (4 Rippen, Sternum, Patella), andere falsch

7
beschrieben (Patella als „Bären-Patella Spitze-Form“), noch wur-de der zertrümmerte Schädel geklebt oder gar „zwei Skelettreste“als solche verstanden. Die Löwenskelette werden mit allen bisherbekannten und in der Kürze der Zeit ermittelbaren Löwenrestender Rübeländer Höhlen monographisch aufgenommen und inden taphonomischen Kontext der „Bärenprädatoren“ gestellt(DIEDRICH in Vorber.).Die einzigen vorausgegangenen populationsstatistischen Analy-sen beschränken sich auch nur auf das Rentier der Baumanns-höhle (SCHÜTT 1969a), wobei sie die Fundstellen beider Höhlenmischt und unvollständig angibt (nur 78 Knochen von über 520).Dieses Problem gibt es auch bei den anderen Spezies, da sie sel-ber niemals eine Höhlenbefahrung oder Höhlenstudien durch-geführt hatte. Daher wurde das Material der LGM/Pleniglazial-Zeit (z.B. 520 Rentierknochen der Baumannshöhle) in einer se-paraten Arbeit über die „Weichsel-Vergletscherung des Harzes“neu ausgewertet (DIEDRICH 2013c). Diese quantitative Analyse,die beide ehem. west-/ost-deutsche Sammlungen erstmals ver-einigt auswertete (was weder STEINER & STEINER 1969 nochSCHÜTT 1969a konnten), legte auch einen Fokus auf die Kno-chen-Taphonomie. Die Historie der Rübeländer Höhlen ist dasbeste bzw. schlechteste Beispiel, was an Wirbeltierfundstellenpassieren kann – verursacht durch wissenschaftsferne Entschei-dungen. Diese verhinderten die Zusammenführung (bis heute)von ganzen Individuenskeletten (Löwen) und einem korrektenGesamtbild zur Paläontologie, da jeder „ein paar Knochen mit-nahm“ – besonders von den Löwenskeletten. Nach neuestenpositiven Entwicklungen in Rübeland/Sachsen-Anhalt konnteaber nun in 2012 an den 1.490 Knochen (LGM-Fauna) somitein Vielfraß-/Eisfuchshorst in der Doline oberhalb der Bau-mannshöhle (eingebrochen in Vertikalschacht am Fotostand nachAuftauen des Permafrostbodens) und ein Eisfuchs-/Zobel-Horstmehrere Meter tief vom damaligen Eingang der Hermannshöhlefestgestellt werden sowie menschlicher Eintrag (angenommenvon BLASIUS 1901a, SCHÜTT 1969a) eindeutig ausgeschlossenwerden (DIEDRICH 2013c).Neuere Absolut-Datierungen an Höhlenbärenknochen (ROSEN-DAHL et al. 2005) sind leider höchst problematisch, da sie an Bären-knochen der „Knochenhaufen in beiden Höhlen“ entstanden,wobei die Knochenhaufen-Aufarbeitung gezeigt hat, dass über50 % der Bärenknochen des Haufens am Knochenfeld der Bau-mannshöhle (fast alle großen Knochen, Schädel und ehemalsmontiertes Skelett) tatsächlich aber aus Material der Hermanns-höhle zur „Dekoration“ stammen. Nochmals wurden pauschal„alle Höhlenbären- und Knochenfunde der Rübeländer Höh-len“ in das mittlere Spät-Pleistozän datiert (ROSENDAHL et al.2005), was nach den hier präsentierten Details und neusten Feld-/Sammlungsstudien keineswegs haltbar ist, weder für die Bau-manns- noch die Hermannshöhle, weil die höchst komplexenNutzungen durch Tiere seit der Saale-Eiszeit mehr als die letz-ten ca. 250.00 Jahre umfassen.
Archäologische Funde des Mittel-/Jung-Paläolithikumsund NeolithikumsDie Neandertaler-Steinartefakte aus dem Knochenfeld der Bau-mannshöhle wurden schon früh richtig interpretiert, aber nichtdatiert (BLASIUS 1898), und später nochmals ausführlicher be-schrieben (TÖPFER 1954) und separat neu bearbeitet. Ob dieseNeandertaler-Artefakte mit dem Sommer-Jagdlager flussabwärtsan der Bode in Westeregeln auch zeitlich zusammenhängen(DIEDRICH & WEBER 2013), ist noch ungeklärt.In der Hermannshöhle wurden zusätzlich nur jungpaläolithische
Abb. 2: A. Schädel (nach Restaurierung in 2012, rechteSchädelabbildung, linke Schädelabbildung aus KLOOS & MÜLLER
1889). B. Skelett 2 montiert von KLOOS nach neuer Restaurierungim LMNB. C. Skelett 1 montiert von Kloos (aus KLOOS & MÜLLER
1889) im damaligen Museum in Rübeland. D. Dessen heutigeErhaltung im RH. Das Skelett ist ein Kompositum aus unter-schiedlichen Arten/ Unterarten und Geschlechtern sowie Indivi-duen. Einige Knochen fehlen durch die mehrfache Demontage/Aufbau im alten Museum und der Höhle, wobei einige anato-mische Fehler vorhanden sind (ein Lendenwirbel zu viel, Rippenaus nicht ganz korrekten Positionen sowie teilweise nicht korrekteFußknochen). Erläuterung der Abkürzungen siehe Seite 10.

8
Funde mehrfach getätigt (TÖPFER 1956, STEINER 1964, STEINER
& STEINER 1969), die dann als Aurignacien-Kulturreste identifi-ziert (DÖHLE & WEBER 2004) und mit dem Knochenprojektil (=Lautscher Spitze) auch absolut in das frühe Aurignacien mit ei-nem AMS-Alter von 29.210 ± 210 BP datiert wurden (GRÜN-BERG 2006). Die neuesten Untersuchungen und Review aller Fun-de sowie neueste Schnittspuren-Entdeckungen an Höhlenbären-knochen in 2012 beweisen – in Europa in dieser Form einmalig– die Höhlenbärenjagd in der Hermannshöhle 90 m tief im Be-reich der Saal-Halle und das Schlachten im Bereich der Bären-höhlen-Halle (DIEDRICH 2013d).Missinterpretationen in Bezug auf „zerschlagene oder beschnit-tene Knochen“ der LGM-Fauna (LGM = Last Glacial Maxi-mum, vgl. CLARK et al. 2009) (wäre = Spät-Magdalénian-Zeit;vgl. BLASIUS 1901a, SCHÜTT 1969a), sind nicht zu halten (allesamtRaubtier-Bissspuren cf. DIEDRICH 2013c, zerknackte Knochenverglichen mit generellen Raubtier-Bissspuren-Analysen in z.B.KÜCHELMANN 1997) und haben daher zu Fehlinterpretationender Höhlennutzung verleitet.Die neolithischen Funde (Michelsberger Kultur) wurden am Ein-gangsportal der Baumannshöhle (= historischer Führungswegs-Eingang) ergraben (TÖPFER 1967). Diese bestehen nach Ansichtder Originale aus mehreren Keramikgefäßen (z. B. MELLER 2008)und einigen Scherben, einem Backteller, Feuerstein-Klingen undFeuersteinpfeilspitzen, einem Klopfstein und Unterlieger-Mühl-stein, zerschlagenen Haustierknochen (Rind, Schwein, Schaf/Ziege) und einem doppelringartigen Knochen-Kunstobjekt.Auch dieses Material sollte neu bearbeitet werden.
Geologie und DatierungIn beiden Höhlen wurden bereits historisch angeschnittene Pro-file aufgenommen und mit den Profilabbildungen von STEINER
& STEINER (1969) sowie FISCHER (1995) verglichen. Keine derpublizierten Profile waren „datiert“ bzw. es wurden nicht halt-bare „Alter“ angenommen. Die Datierung wurde mit verschie-denen Methoden angegangen. Außerhalb der Höhlen wurdendie Flussterrassen/Gletscherausdehnungen bereits parallel un-tersucht und letztendlich mit den in den Höhlen datierbarenSedimenten/Ponorhöhlenetagen korreliert (DIEDRICH 2013c).Hier ist zusätzlich ausführlich die „Bärenuhr“ erläutert, die eineindirekte Datierung anhand der P4-Zahnmorphologie und derSchädelmorphologie an Höhlenbären-Arten/Unterarten erlaubt(Abb. 5), die letztendlich auf einem Review der jüngerenHöhlenbärenforschungsarbeiten (DNA, C14, P4-Morphologie,Schädelmorphologie) der letzten 10 Jahre basiert (HOFREITER
2002, HOFREITER & RABEDER 2004, PACHER & STUART 2008, STIL-LER et al. 2010, MÜNZEL et al. 2012, DIEDRICH 2013b, h). DieProfile und Ponorhöhlenetagen können hier erstmals mit Hilfeder Höhlenbären in ein relativchronologisches Schema einge-ordnet werden, wobei absolute Datierungen in einem weiterenSchritt erfolgen müssen, um genaue Alter zu ermitteln. Weiterhinsind die Korrekturen/Ergänzungen der Megafaunen der Höh-len sehr bedeutsam, da manche Spezies wie Panthera leo fossilis,Cuon alpinus fossilis (teilweise auch Panthera pardus subsp.) in dasMittel-Pleistozän datieren (z.B. THENIUS 1954, SCHÜTT 1969a,ARGANT 1991, EHRLINGER & ZENGER 1999). Die Neufunde vonUrsus deningeri, Frühhöhlenbären-Formen, aus dem Tiefen Gangder Baumannshöhle (Abb. 5) sowie dortigen P. l. fossilis-Restensind höchst wichtig zum Verständnis der Höhlennutzung unter-schiedlicher Bereiche der Höhle und deren Datierungen. Weite-re Datierungshilfen waren auch die „Knochenerhaltungen“ (sie-he Material und Methoden) sowie ein Cuon alpinus fossilis-Kno-
chen (Calcaneus) der Altsammlungen (LMNB) aus der „AltenHöhle“.Viele Sedimente sind nicht „eingeschwemmt“ (besonders Her-mannshöhle) und in ihrer Genese falsch oder teilweise inkorrektinterpretiert worden, andere hingegen sind von allen, wie auchhier, als „fluvialen Urpsprungs“ bereits früh korrekt interpre-tiert worden – wie z.B. die Kiese der Unteren Schwemmhöhle(KLOOS & MÜLLER 1889, STEINER & STEINER 1969), wobei neuereArbeiten diese nun genauer datieren (DIEDRICH 2013c). Frost-Brekzien hingegen sind nicht, wie SCHÜTT (1969) beschrieb, „ein-geschwemmt“ worden, was zum Verständnis der Knochenan-reicherungen sehr bedeutsam ist, die in der Hermannshöhle nachden neuen Modellen (DIEDRICH 2013c, Abb. 3-4) in den oberenEtagen rein autochthon (bis solifluktial in Vertikalschächten, odergar durch Gletschereis talseitig hineingeschoben) sind, wie auchdie Bären-Bonebeds. Hingegen sind alle allochthonen Bonebedsin der Baumannshöhle nach derzeitigem Stand fluvial in zweiPhasen und unterschiedlichen Zeiten umgelagert worden, of-fenbar durch die höhere Lage der Bodeterrasse in der Saale-Eis-zeit und durch LGM-Gletscheroberflächenwässer (DIEDRICH
2013c).Hier werden nur die neu datierten Profile zu den Arbeiten überdie Höhlengenese während der „Eiszeit“ (DIEDRICH 2013c)nochmals wiedergegeben.
Material und MethodenInterdisziplinäres HöhlensurveyDurch ein interdisziplinäres Studium der Höhlengenese, Geo-logie/Sedimentologie sowie seit historischer Zeit zusammen-getragener Funde verschiedener Ost/West-Sammlungen müssendie Präsentationen von BLASIUS (1901a, b) über KLOOS & MÜLLER
(1989) und STEINER & STEINER (1969) oder SCHÜTT (1969a) bishin zu neusten Datierungen (ROSENDAHL et al. 2005, NORDHOFF
2005) stark revidiert oder kritisch gesehen werden. All diesevorausgegangenen Studien separierten meist nicht sauber dievöllig unterschiedlichen Höhlen, deren Höhlenbereiche und ihresehr unterschiedlich artlich zusammengesetzten Faunen, die wieschon mehrfach andiskutiert aus verschiedenen Zeitepochenstammen. Trotzdem wurden sie schließlich vermischt (auch ineinigen Sammlungen wie der LMNB, besonders SCHÜTT 1969a)bzw. Funde zeitlich falsch den Höhlen oder auch artlich falschzugeordnet. Klassische „Faunenlisten“ der damaligen Publika-tionen helfen daher nur wenig zum Gesamtverständnis und derHöhlennutzung in Raum und Zeit.
Historische Quellen und FeldarbeitAuf diese Literatur als Primärquelle (daher Neustudium amOriginalmaterial) wird daher mit Vorbehalt zurückgegriffen.Diese vorausgehende historische und recht genau beschreibendeLiteratur (besonders BLASIUS 1901a,b, KLOOS & MÜLLER 1989)wurde aber zur „Relokalisierung“ der Funde in den jeweiligenbeiden Höhlen und Grabungsstellen benutzt, was von großerBedeutung war. Damit und mit den Neustudien (Abb. 5) wurdenzunächst Altfunde der Museen (besonders LMNB) wieder ihremFundort zugeordnet. Hierzu wurden dann genehmigte Höhlen-befahrungen vorgenommen sowie Knochenproben und beson-ders Zahnmaterial aus „anthropogen verursachten Knochen-haufen“ in den Höhlen entnommen (siehe Genehmigung inDanksagung). Einige offene Profile (Höhlenausbau, damaligeGrabungen) konnten neu geputzt und neu stratigraphisch/sedimentologisch aufgenommen werden, in denen auch Bären-reste steckten bzw. stecken (siehe Bärenschädel im Profil Seiten-

9
Abb. 3: Die Bärenhorst-Bereiche und deren zeitliche Unterschiede in der Baumannshöhle nach den neuen Ergebnissen. In dermittleren Saale-Eiszeit lag der erste Höhlenbärenhorst (Ursus deningeri) im Bereich des Goethesaals, wobei die Knochenlager (=Bonebeds) bis in tiefere Höhlenteile verschwemmt wurden. Der Alte Eingang wurde verblockt, ein anderer öffnete sich, sodass zumEnde der Saale-Eiszeit bis zur mittleren Weichsel-Kaltzeit Höhlenbären von der anderen Seite aus einen noch heute unbekanntenHöhlenteil als Horst nutzten, der auch von Leoparden als Horst genutzt wurde. Dort waren es aber bereits höher entwickelte Ursusspelaeus eremus/spelaeus-Höhlenbären. Ebenso wurden dort die Knochenschichten allesamt aufgearbeitet und durch Gletscher-Oberflächenwässer zur Zeit des Hochglazials (= LGM) ebenfalls in tiefere Bereiche verfrachtet (Kartengrundlage nach FRICKE 1998).
gang Bärenhöhlen-Halle, Abb. 5). Verschiedene Etagen beiderHöhlensysteme wurden befahren sowie wenige wichtige Sedi-mentproben zwecks Bestimmung des Fossilinhaltes (besondersKleinsäuger-Präsenz, neonate Bärenreste) und einer vorläufigenKieskomponenten-Analyse (lokale devonische Massenkalke odereingeschwemmte Bodetal-Kiese) entnommen.
Knochenerhaltung und Separierung historischer FundeDie Knochen wurden in der Knochenerhaltung (= Färbung,Einlagerung von Eisen-Mangan-Mineralen, Abb. 5) in Samm-lungen bzw. im digitalen Fotokatalog separiert. Danach wurdendie noch anstehenden Profile, aus denen unterschiedlich gefärbte/
erhaltene Knochen stammen, sedimentologisch und stratigra-phisch abgeglichen, wobei wenige Knochenproben aus den Pro-filen entnommen wurden. Dadurch können z.B. die im LMNBvermischten „Rübeland-Höhlen-Funde“ in Zukunft zu 99 %korrekt auseinander dividiert werden. Dieses ist mit dem hierpublizierten Material bereits erfolgt, was daher ein völlig anderesBild zur Taphonomie, Paläobiologie sowie Evolution undDatierung der Höhlenbären ergibt. Durch die Separierung derFaunen entstehen letztendlich völlig neue „Höhlengenese-/Nutzungsmodelle“, wobei hier nun auch viel ältere mittel-pleistozäne saale-eiszeitliche Faunen erstmals präsentiert werdenkönnen, sowie eine LGM-Fauna (spätes Spät-Pleistozän) an

10
beiden Höhlen. Moderne Vielfraß-, Polarfuchs-, Zobel-, Bären-und Hyänenhorst-Taphonomieanalysen, die gesondert in derKorrelation zur Harz-Vergletscherung erscheinen (DIEDRICH
2013c), sind der Schlüssel zum Verständnis zu diesen Faunen-unterschieden, die letztendlich von der Höhlengenese (Öffnung,Einschwemmungen und Versturz der Eingänge) und besondersklimatologisch gesteuert wurden.
Abb. 4: Der Bärenhorst mit der Höhlenbärenhorst-Fauna: Borealwald/alpine Fauna in der Hermannshöhle nach den neuenErgebnissen. Durch Hyänen und Wölfe wurden im vorderen Bereich bis zur Bärenhöhlen-Halle deren Beutetiere (Ren, Steinbock,Gams) eingeschleppt. Primär fraßen die Raubtiere aber an den Höhlenbärenkadavern. Löwen, von denen zwei noch unbeschriebeneausgewachsene (teils pathologische) Skelette gefunden wurden, gingen sehr tief auf Höhlenbären-Jungtier-Jagd in der Zeit ihrerWinterruhe. Der Bärenhorst war im gesamten Spät-Pleistozän (115.000-25.000 BP) immer in den beiden oberen trockenen Etagen,während in derselben Zeit der Höhlenfluss in der mittleren Etage (= Untere Schwemmhöhle) entwässerte. Erst war es Ursus spelaeuseremus (kleine Form), Ursus spelaeus spelaeus (mittlere Form mit steil abfallendem Scheitelkamm), und zuletzt Ursus ingressus(mit horizontalem Scheitelkamm, größter Höhlenbär), der die Höhle als Horst nutzte. Hyänen und Wölfe konnten den Eingangsbereichnur sporadisch nutzen. Löwen Panthera leo spelaea gingen tief in die Höhle auf Bärenjagd.
Studierte SammlungenDas studierte Artefakt-/Knochenmaterial (Grabungen BLASIUS,KLOOS, STEINER, VÖLKER, Knochenhaufen-Rettung in 2012) be-findet sich in folgenden Museen: Staatliches NaturhistorischesMuseum Braunschweig (= LMNB), Naturkundemuseum derHumboldt-Universität Berlin (= MB), Martin Luther-Universi-
tät Halle/Saale, Institut für Geologie (= MLU.IFG), Landes-amt für Denkmalpflege und Archäologie, Landesmuseum fürVorgeschichte (= LDA), Rübeländer Höhlen (= RH), MuseumQuedlinburg (= MQ), Museum Bernburg (= MBE).
Ergebnisse und DiskussionGeologie, Stratigraphie und HöhlengeneseDie Ergebnisse zur Nutzung der Höhlen durch Höhlenbären inder Saale-Weichsel-Eiszeit sind in den beiden thematischenKarten (hier nur Bärenhorstdarstellungen, Abb. 3-4) zusammen-gefasst. Diese enthalten weder die archäologischen Funde nochdie LGM-Faunen. Eine ausführlichere Darstellung der Sedi-mentologie und Argumente für die LGM-Vergletscherung desHarzes sowie die Anwesenheit des Bodetal-Gletschers und dessen

11
Abb. 5: Bärenevolution und Datierungen mit Höhlenbärenknochen, -schädeln und -zähnen (= Höhlenbärenuhr“) sowieHöhlenfaunenunterschiede anhand der Funde aus den Rübeländer Höhlen (Absolutdaten der Sinter aus NORDHOFF 2005).
Kartierung und Stadien der Harz-Vereisung, besonders derWeichsel-Kaltzeit, sowie die LGM-Faunen finden sich inDIEDRICH (2013c). Die wenigen offenen Profile beider Höhlenwurden hier teilweise mit abgebildet (Abb. 3-5), da sie dieAllochthonie oder Autochthonie der Knochenschichten belegen/widerlegen.
Höhlenbären Evolutions-Forschungsstand und ProblemeDie Evolution der eurasischen Höhlenbären ist hier als Initial-arbeit in Anlehnung an die nicht absolut kalibrierte Zahnmor-
phologieänderung sensu RABEDER (1999) angelehnt, die es zuersterlaubt, „ältere und jüngere“ Höhlenbärenformen zu unter-scheiden. Erst durch neuere Arbeiten über die DNA undAbsolutdatierungen (HOFREITER 2002, RABEDER & HOFREITER
2004, STILLER et al. 2010, MÜNZEL et al. 2011) können diese Datenauch hier im Material von Rübeland weiter kalibriert (genauerdatiert) werden, wobei eine zusammenfassende Arbeit zurHöhlenbärenevolution immer noch aussteht, die eine Bestim-mung der vier spätpleistozänen Höhlenbären-Arten/Unterartenauch ohne DNA in Europa möglich macht.

12
Das Fehlen einer Revisionsarbeit liegt daran, dass man wedergenau die Zahnmorphotypen per DNA analysiert (andere Pro-ben) noch die Schädeltypen damit abgeglichen hat. Hätte manSchädel beprobt, die unterschiedliche Morphologien und P4-Zahnmorphologien aufweisen, hätte man die Bärensystematikin den Griff bekommen können. Ein großes Manko der DNA-Studien ist die Nicht-Integration des verloren geglaubten „Ursusspelaeus Rosenmüller 1794“-Holotypen-Schädels der Zoolithen-höhle, der 2009 wiederentdeckt wurde. Dieser wurde vorläufigmorphologisch als frühadulter „U. spelaeus spelaeus“ angespro-chen (DIEDRICH 2009), müsste aber schädelmorphologisch, wiehier neu diskutiert, als „Ursus ingressus“ identifiziert werden (nachhorizontalem Scheitelkamm und Zahnmorphologie). Dafürmüsste jedoch die Höhlenbärensystematik revidiert werden.Dieser Versuch der Korrelationen der Ergebnisse der Höhlen-bärenforschungen wird hier an dem umfangreichen Höhlenbä-renmaterial der Rübeländer Höhlen in einem ersten Schritt alsModell versucht (Abb. 5). Das Material ist hierfür sehr geeignet,da es längere Zeitabschnitte zwischen Saale-Weichsel-Kaltzeitabdeckt (mindestens 250.000 Jahre), die beiden Höhlen-Abfol-gen der Horstnutzung in verschiedenen Höhlenteilen und Zei-ten zeigt, das Schädel- und P4-Zahnmaterial umfangreich ist underste Absolutdatierungen das Modell untermauern. Was fehlt,sind aber DNA-Analysen und weitere U/Th-Absolutdatierungen,um die „Höhlenbären-Uhr“ auch zeitlich zu eichen.
Die Höhlenbärenzahn-Schädelmorphologie-UhrUnter Nutzung aller bisher bekannten DNA-, Zahnmorphologie-Studien und Absolutdatierungen (RABEDER 1999, RABEDER et al.2002, HOFREITER, 2002, RABEDER & HOFREITER 2004, PACHER &STUART 2008, MÜNZEL et al. 2011, STILLER et al. 2011) könnendas Höhlenbärenzahnmaterial (P4) und die zahlreichen Schädelaus den Rübeländer Höhlen zuzüglich der Stratigraphie (vierwichtiger Profile) in ein relativchronologisches Höhlenbären-Evolutionsmodell gestellt werden (Abb. 5). Hier kann die Schädel-morphologie erstmals deutlicher herausgearbeitet werden, daSchädel aller drei weichselzeitlichen Arten/Unterarten vorliegen,leider aber noch kein saale-eiszeitlicher deningeri-Schädel(Abb. 5-7). Interessant ist auch, dass alle kompletten Schädel nuraus der Hermannshöhle stammen. Bei den Untersuchungen in2012 wurde zwar ein erster Schädel im Knochenfeld der Bau-mannshöhle entdeckt, dieser ist aber eingesintert und derzeit insitu und von einem nicht voll ausgewachsenen Individuum.Durch einige Absolutdatierungen von Ursus ingressus in Süd-deutschland (MÜNZEL et al. 2011) können die großen Höhlenbä-ren aus der Hermannshöhle vorläufig mit diesen Absolut-Datenkorreliert werden, die älteren hingegen bleiben in den C14-Datie-rungen durch ROSENDAHL et al. (2005) derzeit ungewiss, da we-der bekannt wurde, an welchen Arten/Unterarten diese vorge-nommen wurden, noch die ursprüngliche Herkunft der Kno-chen klar ist (Knochenhaufen-Proben). Weitere Absolut-
Abb. 6: Ursus ingressus-Höhlenbärenschädel aus der Hermannshöhle mit vielhöckrigem P4 und fast horizontalem Scheitelkamm inunterschiedlichen Altersstadien von spätjuvenil (noch mit abfallendem Scheitelkamm) bis senil (horizontaler Scheitelkamm). A.Schädel aus der Saal-Halle (Grabung STEINER 1961, LDA). B. Schädel eines senilen Bären aus der „Hermannshöhle“ (Grabung KLOOS
& MÜLLER 1889, LMNB). C. Schädel eines hochadulten Bären aus der Bärenhöhlen-Halle (vom Skelett, Abb. 3C-D, RH). D. Schädeleines hochadulten/senilen Bären aus der „Hermannshöhle“ (LMNB). E. Schädel eines hochadulten Bären aus der Bärenhöhlen-Halle (Funddatum: 1892, Grabung KLOOS & MÜLLER, LMNB). F. Schädel eines frühadulten Bären aus der Saal-Halle (GrabungVÖLKER 1984/85, RH). G. Schädel eines hochadulten Bären aus der Bärenhöhlen-Halle (Grabung STEINER 1961, LDA). H. Schädeleines spätjuvenilen Bären aus der Bärenhöhlen-Halle (Grabung KLOOS & MÜLLER 1889, RH).

13
datierungen an Tropfsteinschichten werden hier auch unter Vor-behalt eingefügt (Abb. 5) und müssten nun gezielt in den be-kannten Profilen neu ermittelt werden. Die einzigen hilfreichenDaten stammen aus der Baumannshöhle mit zwei mittel-saale-eiszeitlichen Daten (NORDHOFF 2005). Diese (?alte) dicke Sinter-schicht scheint in einem hier aufgenommenen Profil (Abb. 5)der Baumannshöhle vorhanden zu sein, was die angenommenenMittel-Saale-Alter der Ursus deningeri-Bären untermauern würde.Diese Sinter fallen nach den Absolutdaten dann in die MIS-Zo-nen 9 und 7, wobei hier die jüngsten Daten vorläufig eingesetztwurden (= jüngstes Alter der deningeri-Bären). Die deningeri-Bä-ren können daher zunächst nur an den primitiven dreihöckrigenP4-Zahn-Morphotypen sensu RABEDER (1999) relativ-chronologisch erfasst und grob in das mittlere Mittel-Pleistozän(Mittel-Saale) anhand der Speläothemdaten (siehe NORDHOFF
2005) datiert werden (Abb. 5). Immerhin ist das eine viel ältereDatierung von Höhlenbären als bisher für die „Rübeländer Höh-len“ angenommen (z.B. ROSENDAHL et al. 2005). Ggf. reichen dieBären sogar in die Cromer-Eiszeit zurück und sind vielleicht„gleichalt“ mit frühesten Bärenfunden (?Cromer nach SCHÜTT
1968, oder wie hier „nur“ Saale) der Einhornhöhle im Harz. Dieseältesten Höhlenbärenpopulationen müssten in Zukunft weiterdatiert und korreliert werden, denn sie belegen die Höhlen-nutzung von zwei großen Höhlensystemen seit der Cromer/Saa-le-Eiszeit, was von anderen Autoren „nicht bestätigt wurde“(NIELBOCK 1998). Dieses ist offensichtlich dadurch zu erklären,dass auch in der Einhornhöhle – wie hier im Detail aufgezeigt inden Rübeländer Höhlen – unterschiedliche Höhlenteile zu un-terschiedlichen Zeiten als Bärenhorste genutzt wurden – abhän-gig von der Eingangsöffnung und Verblockung von Eingängen,die mehrfach passierte (siehe hier Rübeländer Höhlen Abb. 3-
4). Dieses wurde dort noch nicht erkannt und neuere Höhlen-bärenfunde, die in die Weichsel-Kaltzeit datieren, als Argumentgenutzt, ältere Bärentypen als nicht vorhanden/fragwürdig zudeklarieren (NIELBOCK 1998). Nach einer Ansicht der Original-funde (Grabung SCHÜTT 1968) im GZG sind die Bärenreste undFauna sicherlich mindestens saale-eiszeitlich und stark fossil,ähnlich wie die Funde aus dem Tiefen Gang der Baumannshöhle(= erster Horstbereich, siehe Abb. 3). Auf jeden Fall müssen inbeiden Höhlen neue U/Th-Datierungen sowohl an Sinter alsauch den Zähnen erfolgen.Neue Aufsammlungen von Zähnen (besonders P4) in denRübeländer Höhlen und die vorbehaltliche Nutzung derAltsammlungen (hier aber gezielt die P4 in Schädeln und Kiefernwegen der möglichen genauen Relokalisierung der Fundorte)zeigen schon jetzt an dem wenigen Material (ca. 60 Zähne)deutliche Zahnmorphologie-Typenunterschiede (Abb. 5). Materialaus dem Tiefen Gang des Goethesaals und dem Profil in der„Alten Höhle“ sind allesamt dreihöckrige primitive Formen, wobeinur ein vierhöckriger (ggf. sogar pathologischer) Zahn vorliegt.Die Zähne sind auch klein in den Proportionen und wären sensuRABEDER (1999) als primitivster Höhlenbärenzahn Typ A für dieRübeländer Höhlensysteme zu bezeichnen. Solche sind älter alsweichsel-kaltzeitlich, aber das Problem der deningeri-Forschung hatnoch keine exakten Modelle und Datierungen der Morphotypenin der mittleren/späten Saale-Eiszeit geliefert. Die Höhlenbären-Zahntypen im Bereich der „Neuen Höhle“ der Baumannshöhle(Bereich Knochenfeld, Ochsenhang) sind höher entwickelteFormen (Typ B-D sensu RABEDER 1999) mit bis zu sechs Höckern,Abb. 5. Diese datieren die Fauna dort indirekt in das späte Saale-bis Mittel-Weichsel. Anders herum datieren Löwenreste (P. l. fossilis:cf. SCHÜTT 1969), Wildhunde (C. alpinus fossilis: cf. THENIUS 1954)
Abb. 7: A-G. Ursus spelaeus spelaeus mit vier- bis fünfhöckrigem P4 und steil abfallendem Scheitelkamm aus der Hermannshöhle.A. Schädel eines hochadulten Bären vom Bärenfriedhof (Funddatum: 1892, LMNB). B. Schädel eines frühadulten Bären aus derSaal-Halle (Funddatum: 1984/85, Grabung VÖLKER). C. Schädel eines adulten Bären (Grabung KLOOS, MQ). D. Schädel eines adultenBären (Grabung BLASIUS oder KLOOS, LMNB). E. Schädel einer adulten Bärin (Funddatum: 1961, Grabung STEINER, LDA). F. Schädeleiner adulten Bärin (RH). G. Schädel, fragmentiert (MBE). H-I. U. spelaeus eremus-Höhlenbärenschädel aus der Hermannshöhle.H. Schädel (RH). I. Schädel (ggf. sogar Knochenfeld, Baumannshöhle, LMNB).
14
und Leoparden (P. pardus, DIEDRICH 2013e) sowie auch diefrühesten Höhlenbären dieses Höhlenbereiches zurück bis in dieSpät-Saale-Eiszeit (vgl. z.B. SCHÜTT 1968). Dieses zeigt dieVermischung der Faunen, die umgelagert und vermengt wurden(= „Kalkkies-Bonebeds“). Vier- bis sechshöckrige Zahnformenfinden sich auch in der Hermannshöhle, die in das Früh-/Mittel-Weichsel mit Hilfe der Schädel, in denen sie sich befinden, datieren(vgl. U. ingressus-Schädel und Datierung in MÜNZEL et al. 2011).Nur aus der Hermannshöhle sind derzeit die vielhöckrigen Zähnebekannt (Typ E sensu RABEDER 1999), die hier in den Schädelnvon Ursus ingressus stecken bzw. auch in der Bärenhöhlen-Halleeinzeln gefunden wurden. Da man diese nun hier dem Schädeltypzuordnen kann und diese Schädeltypen absolut datiert sind (cf.MÜNZEL et al. 2011), kann man diese zwischen 32-25.000 BP
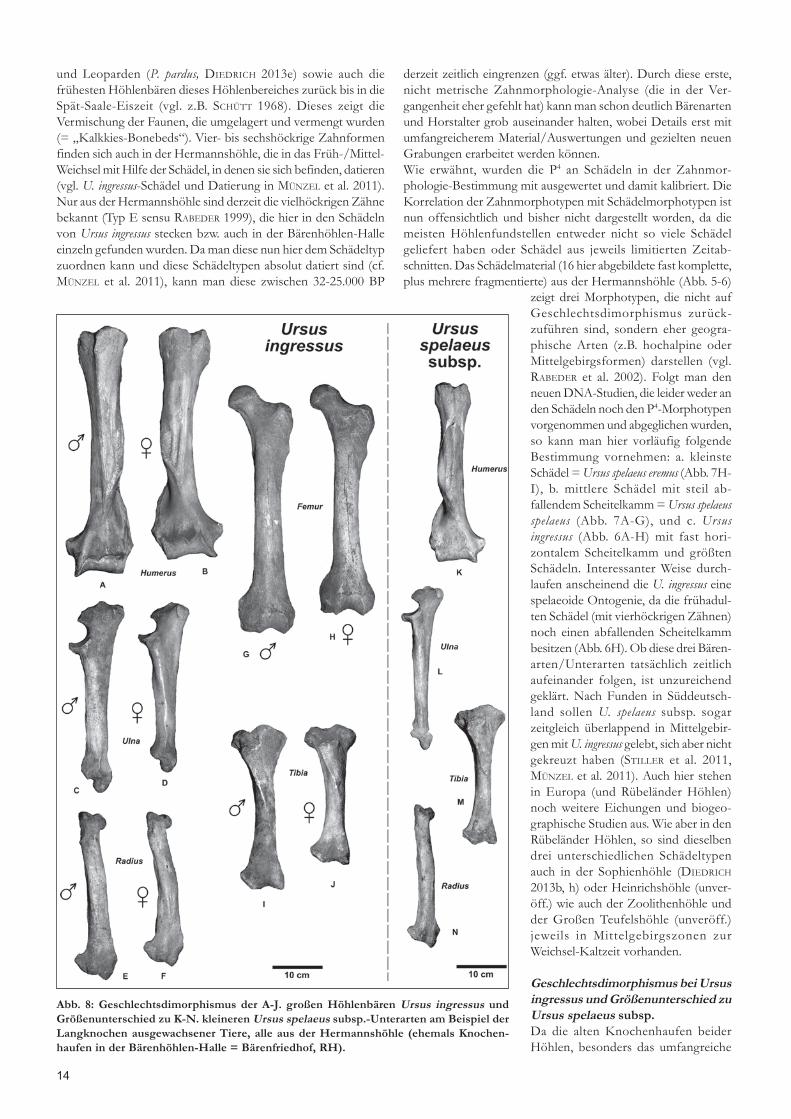
Abb. 8: Geschlechtsdimorphismus der A-J. großen Höhlenbären Ursus ingressus undGrößenunterschied zu K-N. kleineren Ursus spelaeus subsp.-Unterarten am Beispiel derLangknochen ausgewachsener Tiere, alle aus der Hermannshöhle (ehemals Knochen-haufen in der Bärenhöhlen-Halle = Bärenfriedhof, RH).
derzeit zeitlich eingrenzen (ggf. etwas älter). Durch diese erste,nicht metrische Zahnmorphologie-Analyse (die in der Ver-gangenheit eher gefehlt hat) kann man schon deutlich Bärenartenund Horstalter grob auseinander halten, wobei Details erst mitumfangreicherem Material/Auswertungen und gezielten neuenGrabungen erarbeitet werden können.Wie erwähnt, wurden die P4 an Schädeln in der Zahnmor-phologie-Bestimmung mit ausgewertet und damit kalibriert. DieKorrelation der Zahnmorphotypen mit Schädelmorphotypen istnun offensichtlich und bisher nicht dargestellt worden, da diemeisten Höhlenfundstellen entweder nicht so viele Schädelgeliefert haben oder Schädel aus jeweils limitierten Zeitab-schnitten. Das Schädelmaterial (16 hier abgebildete fast komplette,plus mehrere fragmentierte) aus der Hermannshöhle (Abb. 5-6)
zeigt drei Morphotypen, die nicht aufGeschlechtsdimorphismus zurück-zuführen sind, sondern eher geogra-phische Arten (z.B. hochalpine oderMittelgebirgsformen) darstellen (vgl.RABEDER et al. 2002). Folgt man denneuen DNA-Studien, die leider weder anden Schädeln noch den P4-Morphotypenvorgenommen und abgeglichen wurden,so kann man hier vorläufig folgendeBestimmung vornehmen: a. kleinsteSchädel = Ursus spelaeus eremus (Abb. 7H-I), b. mittlere Schädel mit steil ab-fallendem Scheitelkamm = Ursus spelaeusspelaeus (Abb. 7A-G), und c. Ursusingressus (Abb. 6A-H) mit fast hori-zontalem Scheitelkamm und größtenSchädeln. Interessanter Weise durch-laufen anscheinend die U. ingressus einespelaeoide Ontogenie, da die frühadul-ten Schädel (mit vierhöckrigen Zähnen)noch einen abfallenden Scheitelkammbesitzen (Abb. 6H). Ob diese drei Bären-arten/Unterarten tatsächlich zeitlichaufeinander folgen, ist unzureichendgeklärt. Nach Funden in Süddeutsch-land sollen U. spelaeus subsp. sogarzeitgleich überlappend in Mittelgebir-gen mit U. ingressus gelebt, sich aber nichtgekreuzt haben (STILLER et al. 2011,MÜNZEL et al. 2011). Auch hier stehenin Europa (und Rübeländer Höhlen)noch weitere Eichungen und biogeo-graphische Studien aus. Wie aber in denRübeländer Höhlen, so sind dieselbendrei unterschiedlichen Schädeltypenauch in der Sophienhöhle (DIEDRICH
2013b, h) oder Heinrichshöhle (unver-öff.) wie auch der Zoolithenhöhle undder Großen Teufelshöhle (unveröff.)jeweils in Mittelgebirgszonen zurWeichsel-Kaltzeit vorhanden.
Geschlechtsdimorphismus bei Ursusingressus und Größenunterschied zuUrsus spelaeus subsp.Da die alten Knochenhaufen beiderHöhlen, besonders das umfangreiche

15
Material der Hermanshöhle, oft einfach aufgestapelt wurden, istderen Analyse ebenfalls hilfreich zur ersten Auswertung derHöhlenbären-Spezies-Vorkommen. Es werden hier zunächst keinemetrischen Analysen präsentiert, denn letztendlich sind auch dieseproblematisch, wenn man nicht die verschiedenen Arten/Unterarten cranial separieren kann. Daher sind auch die altenArbeiten wie z.B. von KURTÉN (1955) nicht mehr nutzbar, da dieseeben heute nach DNA bekannte unterschiedliche Unterarten als„Geschlechtsdimorphismus“ falsch gedeutet haben. Es scheint,dass das Langknochenmaterial Geschlechtsunterschiede an dengroßen Höhlenbären Ursus ingressus zeigt, mit etwas kleinerproportionierten Bärinnen (Abb. 8). Hier wurden die größten undkleinsten Langknochen ausgewählt, die höchstwahrscheinlichdiesen Geschlechtsdimorphismus widerspiegeln. Dieses korreliertmit neuen Ergebnissen in der Sophienhöhle, in der auch großeU. ingressus von kleineren U. spelaeus subsp.-Formen neben denSchädeln auch am postcranialen Material sehr gut unterschiedenwerden konnten (DIEDRICH 2013h). Selbst wenn hier nur Varianzenvorliegen sollten, so ist der Größenunterschied ganz offensichtlichzu den deutlich kleineren Höhlenbären Ursus spelaeus subsp., vondenen leider kein kompletter Femur zum Vergleich vorlag (Abb. 8).Das gesamte Knochenmaterial müsste metrisch erfasst werden,wobei die wichtigen Größenunterschiede mit den Ergebnissenzur Schädelmorphologie (Abb. 6-7) passen und korrelieren.Vielleicht können in Zukunft die Geschlechtsunterschiede derkleineren Höhlenbärenformen statistisch besser herausgearbeitetwerden. Hierzu kann man auch andere Knochen, besondersMetapodien, heranziehen.
Neonate Höhlenbären-Skelette
Abb. 9: Zwei unterschiedlich alte neonate (= ungeborene) Höhlenbären-Teilskelette ausder Saal-Halle, Hermannshöhle (Grabung STEINER 1961, LDA)
Isolierte Knochen und Milchzähneneonater bis mehrwöchiger Höhlen-bären finden sich in allen Höhlen-bärenhorst-Bereichen beider Rübe-länder Höhlen. Es ist aber nur wenigMaterial in den meisten Sammlungenvorhanden, da die Sedimente in allenFällen nicht durchgesiebt wurden unddaher Kleinfunde in der Regel fehlen.Nach ersten Siebproben (über 1 mm)finden sich pro 10 l-Eimer mehrerekleinste Bärenknochen sowohl in denSchichten des Knochenfeldes (Bau-mannshöhle) als auch in den Sedi-menten der Bärenhöhlen-Halle (Her-mannshöhle). Da leider auch nicht dasSediment im Falle zweier neonaterBärenskelette aus der Saal-Halle (Her-mannshöhle, Abb. 9) gesiebt wurde,fehlen die sicherlich vorhanden gewe-senen Kleinstknochen. Solche neo-naten Skelette wurden aus deutschenHöhlen bisher nur aus der Dechen-höhle (DREYER & GRAW 2002) und derPetershöhle (KAULICH & ROSENDAHL
2000) abgebildet, wobei dieses keine„Höhlenbärenbabies“ waren, sondernUngeborene oder Totgeburten. Derhohe Anteil an solchen, auch in denRübeländer Höhlen, deutet darauf hin,dass recht häufig die Bärin nicht „genugWinterspeck angefressen“ hatte, um
ihre Jungen lebend auf die Welt zu bringen. Gute Statistiken zudiesem Thema fehlen, da die meisten historischen selektivenSammlungen dieses Kleinmaterial nicht beinhalten.
PathologienPathologien an Höhlenbären sind schon früh erkannt (z.B.BREUER 1931a, b) und an verschiedenen Knochen aus Höhlen inEuropa abgebildet worden (z. B. DREYER & GRAW 2002, DIEDRICH
2006). Auch aus der Heinrichshöhle wurden von FISCHER (1995)bereits einige pathologische Stücke abgebildet. Die alten Inter-pretationen, besonders von „rheumatischen Erkrankungen“,müssen zukünftig revidiert werden. Durch die neuen Erkennt-nisse über die Höhlenbären-Prädation durch Löwen und Hyä-nen, wie nun auch Wölfen, sind Schädelverletzungen an Höh-lenbären der Zoolithenhöhle kürzlich revidiert worden (DIEDRICH
2013a). Pathologien sind in vielen Fällen nicht „rheumatischenUrsprunges“ wegen des kalt-feuchten Höhlenklimas, sonderngehen in vielen Fällen eher auf Bissverletzungen durch Artge-nossen bei innerartlichen Auseinandersetzungen oder Konflik-ten mit den Prädatoren zurück (DIEDRICH 2011a, b, 2013d,ROTHSCHILD & DIEDRICH 2012). In vielen Fällen sind es„Knochenwucherungen auf den Knochenoberflächen“ (=Periostreaktion), wie sie auch bei Menschen vorkommen (RANAL
et al. 2009), die aber unterschiedliche Ursachen haben können.Hinzu kommen die neuen Erkenntnisse über die Höhlenbären-jagd, wie sie in der Hermannshöhle nun belegt ist (DIEDRICH
2013d). Auch durch jungpaläolithische Menschen scheint die einoder andere Verletzung verursacht worden zu sein. Eine Revi-

16

17
Abb. 10 (linke Seite): Pathologien an Höhlenbärenknochen derRübeländer Höhlenbären. A. Caninus (U. ingressus) mit Wur-zelauflösung, Ende Untere Schwemmhöhle, Versturzkonus,Hermannshöhle (RH). B-C. Zu schlanke und am Zahnschmelzdeformierte canini (U. ingressus) eines Jungtieres, Bärenhöhlen-Halle, Hermannshöhle (RH). D. Zahn (U. spelaeus subsp.) mitstarker Deformation, Knochenfeld, Baumannshöhle (LMNB). E.Oberkiefer P4 (U. ingressus) mit Zahnschmelz-Unregelmäßigkeit,Bärenhöhlen-Halle, Hermannshöhle (LMNB). F. Unterkiefereines Jungtieres mit nur halb entwickeltem M3, Saal-Halle,Hermannshöhle (LDA). G. Unterkiefer eines adulten Tieres mitausgefallenen M
1 und M
3 (Alveolarentzündung bei M
1), Saal-
Halle, Hermannshöhle (LDA). H. Unterkiefer eines senilen Tieresmit abgerundeter Caninuswurzel und fast zugewachsenen P
4/M
1-
Alveolen, Ende Untere Schwemmhöhle, Versturzkonus, Her-mannshöhle (RH). I. Unterkiefer eines Jungtieres mit aus-gefallenen M1-3
und teils verschlossenen Alveolen sowie
Kieferdeformierung, Saal-Halle, Hermannshöhle (LDA). J.Unterkiefer (U. spelaeus subsp.) eines sehr jungen Tieres mitKnochenwucherungen im vorderen Kieferast, Knochenfeld, Bau-mannshöhle (LMNB). K. Sternalknochen mit Knochen-wucherung (Zusammenwachsen), Saal-Halle, Hermannshöhle(LDA). L. Zweite Rippe (Rippenköpfe) mit Knochenwucherungam distalen Gelenkkopf, Bärenhöhlen-Halle, Hermannshöhle(RH). M. Mittlerer Halswirbel mit Syndesmophyt am unterenRande der hinteren Zentrumscheibe, Bärenhöhlen-Halle,Hermannshöhle (RH). N. Cervical 6-7 und Thoracalwirbel 1 (U.spelaeus subsp.) mit Syndesmophyten an den Zentrumscheiben-Unterseiten, Knochenfeld, Baumannshöhle (RH). O. Syndes-mophyt am mittleren Brustwirbel der Zentrumscheiben-Unterseite (U. spelaeus subsp.), Knochenfeld, Baumannshöhle(LMNB). P. Syndesmophyt am hinteren Brustwirbel der vorderenZentrumscheiben-Unterseite (U. ingressus), Bärenhöhlen-Halle,Hermannshöhle (RH). Q. Distaler Humerus (U. spelaeus subsp.)mit Loch-Anomalie, Hermannshöhle (RH). R. Distaler Humerus(U. spelaeus subsp.) mit Loch-Anomalie, Hermannshöhle (RH).S. Distaler Humerus (U. spelaeus subsp.) mit lateraler Steg-bildung, Hermannshöhle (RH). T. Ulna (U. ingressus) mitPeriostreaktion, Hermannshöhle (RH). U. Ulna (U. ingressus)mit Bissschramm-Verletzung eines subadulten Tieres,Hermannshöhle (RH). V. Distal gebrochener Ulna-Schaft einesJungtieres (U. spelaeus subsp.), Knochenfeld, Baumannshöhle(LMNB). W. Ulna (U. spelaeus subsp.) mit Periostreaktion,Knochenfeld, Baumannshöhle (LMNB). X. Ulna (U. ingressus)mit Periostreaktion, Ende Untere Schwemmhöhle, Versturzkonus,Hermannshöhle (RH). Y. Radius (U. ingressus) mit Periost-reaktion, Bärenhöhlen-Halle, Hermannshöhle (RH). Z. Radius(U. ingressus) mit Periostreaktion, Saal-Halle, Hermannshöhle(RH). Ä. Ulna (U. ingressus) mit Lochbildung, Bärenhöhlen-Halle, Hermannshöhle (RH). Ö. Tibia (U. ingressus) mitPeriostreaktion, Bärenhöhlen-Halle, Hermannshöhle (RH). Ü.Fibula (U. spelaeus subsp.) mit ?Exostose (?Bruch), Knochenfeld,Baumannshöhle (LMNB). AA. Calcaneus (U. ingressus) mitPeriostreaktion, Hermannshöhle (LMNB). BB. Calcaneus (U.spelaeus subsp.) mit Periostreaktion, Knochenfeld, Bau-mannshöhle (LMNB). CC. Phalanx I (U. spelaeus subsp.) mitGelenkdeformierung, Knochenfeld, Baumannshöhle (LMNB).DD. Phalanx II (U. ingressus) mit Gelenkdeformierung,Bärenhöhlen-Halle, Hermannshöhle (RH).
sion der „Höhlenbären-Pathologien europäischer Höhlenbären“steht ebenfalls aus. Hier werden vorläufig einige bisher unpub-lizierte Pathologien aus beiden Rübeländer Höhlen an Ursusingressus und Ursus spelaeus subsp. präsentiert (Abb. 10). In eini-gen Fällen kann man zumindest die Art der Verletzung ersehen,in anderen die Krankheit erahnen, aber auch hier braucht es mehrVergleiche, um die genaue Ursache der Verletzung/Krankheit
zu ermitteln. Einzelne Zähne zeigen Zahnwurzelabsorptionen(Wurzelentzündung) oder Deformationen selbst des Zahn-schmelzes oder abnorme Formen (Abb. 10A-F). Zahn-pathologien sind auch an einigen Kiefern zu erkennen, wobeidie Alveolen vereitert waren, sich erweiterten und der Zahnschließlich ausfiel (Abb. 10G-I). Ein voll deformierter Unterkie-fer (?Bruch und Zahnausfall) ist eine Seltenheit (Abb. 10I). Meistsind es Knochenwucherungen (= Exostose, Periostreaktion, Abb.10K-DD), die meist zwei Hauptursachen haben: a. Biss-verletzungen und b. Arthritis. Knochenwucherungen an Wirbel-körpern, sind immer an den Scheibenunterseiten zu finden, ty-pische Syndesmophyten (Abb. 10M-P). Andere Wucherungenfindet man an Knochenbrüchen (Abb. 10V und Ü).
Raubtier-Bissspuren an HöhlenbärenknochenErste Groß-Raubtier-Verbissspuren an den „Rübeländer Höh-lenbärenknochen“ bildeten schon STEINER & STEINER (1969) undFISCHER (1995) zumindest mit einigen Exemplaren ab, wobeideren Verursacher weder genauer identifiziert wurden noch dieTaphonomie der angefressenen/zerknackten Bärenknochenuntersucht wurde.Umfangreichere Untersuchungen erfolgten erstmals an einemHöhlenbärenhorst (Heinrichshöhle, Sauerland, DIEDRICH 2005b),die aufzeigten, dass nicht wie vorher geglaubt Höhlenbären als„Kannibalen“ für die Knochenzerstörungen verantwortlich wa-ren (RABEDER et al. 2000), sondern primär Eiszeit-Hyänen(DIEDRICH 2005b, 2013C). Nach weiteren Studien an zusätzlichenHöhlen (Keppler-Höhle, Bilstein-Höhle: Sauerland, Zoolithen-höhle, Sophienhöhle: Oberfranken) wurde dann deutlich, dassauch der Eiszeitlöwe zumindest Knochenschäfte und Bissspurenhinterlassen haben muss (DIEDRICH 2011c). Solche Knochen-schäfte, deren Gelenkköpfe abgefressen sind, liegen besondershäufig bei den Jungbärenknochen (bis 1 Jahr Individuenalter)vor, wobei das Material aus der Hermannshöhle etwa 80 % Biss-schäden aufweist. Natürlich bleibt das Problem an dem Materi-al, wie auch wenigen Funden der Baumannshöhle, dass es selek-tives Altfundmaterial ist. Aber genau hier wurden die angefres-senen und beschädigten Knochen, die besonders als unvollstän-dige Knochen auf die Knochenhaufen geworfen wurden, mitden besser erhaltenen der Sammlungen (besonders LMNB) kom-biniert, was ein recht gutes Bild ermöglicht. Während im LMNB(Sammlungen KLOOS/BLASIUS) z.B. Höhlenbärenknochen-fragmente fast fehlen, wurden diese in den Grabungen von STEI-NER/VÖLKER mitgenommen. Abgebildet werden hier exempla-risch Serien unterschiedlich alter Höhlenbären und unterschied-licher Langknochen (Humerus, Ulna, Radius, Femur und Tibia,Abb. 11). Identische Fraßreste (Knochenschäfte) sind aus ande-ren Höhlen in Deutschland abgebildet worden (DIEDRICH 2005b,2011c, 2012b, c). An allen hier abgebildeten Höhlenbärenknochensind an den Schaftenden meist 2-4 mm kleine trianguläre,bumerangförmige oder kleine ovale Bissmarken oder 1 mm dün-ne Bissschrammen vorhanden, die von den Brechscherenzähnender Großraubtiere verursacht wurden (vgl. Bissspuren undVerursacher in DIEDRICH 2011b).Typisch sind auch die zickzack-förmigen Schaftränder, die nurdurch die Brechscherenzähne entstehen können (kein natürli-cher Bruch durch Transport). Zickzack-Ränder an Knochen an-derer Eiszeit-Großsäuger-Langknochen sind ebenfalls zahlreichin Zusammenhang mit Hyänenfraß abgebildet worden (sieheHyänenhorst-Knochentaphonomie: DIEDRICH 2012a, 2013g).Neben den Langknochen finden sich befressene Beckenreste,besonders häufig wurde nur noch das Acetabulum (Gelenk-
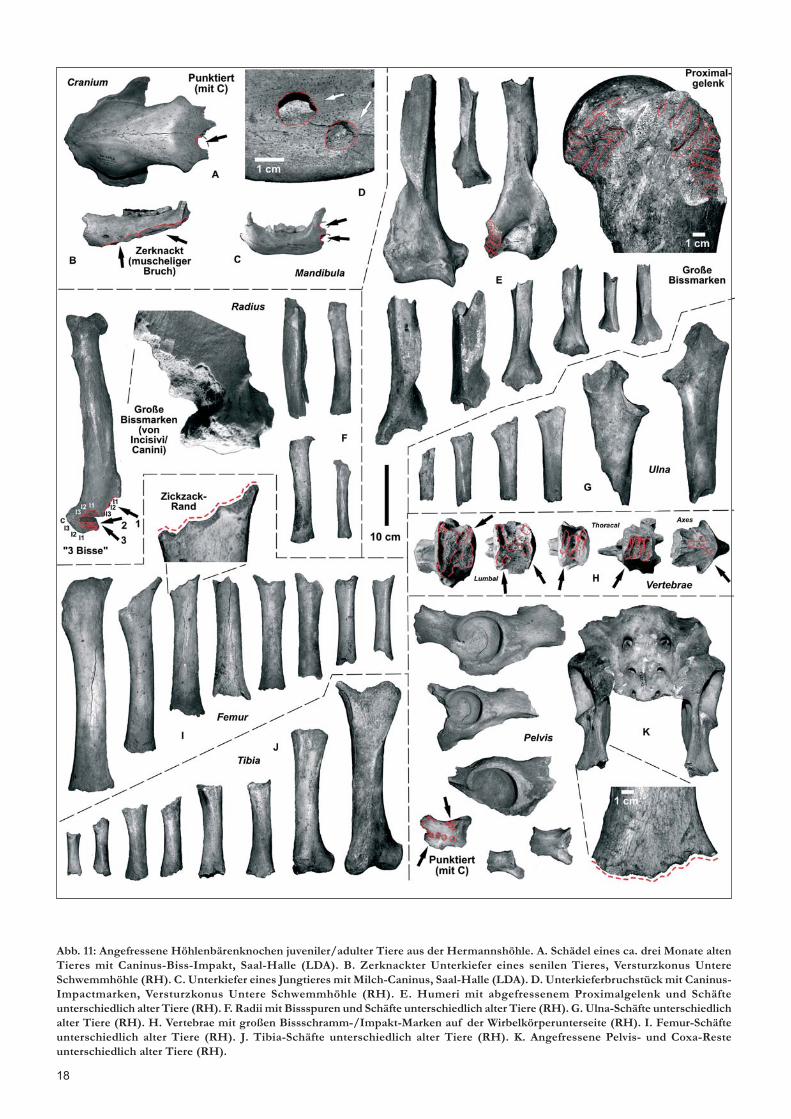
18
Abb. 11: Angefressene Höhlenbärenknochen juveniler/adulter Tiere aus der Hermannshöhle. A. Schädel eines ca. drei Monate altenTieres mit Caninus-Biss-Impakt, Saal-Halle (LDA). B. Zerknackter Unterkiefer eines senilen Tieres, Versturzkonus UntereSchwemmhöhle (RH). C. Unterkiefer eines Jungtieres mit Milch-Caninus, Saal-Halle (LDA). D. Unterkieferbruchstück mit Caninus-Impactmarken, Versturzkonus Untere Schwemmhöhle (RH). E. Humeri mit abgefressenem Proximalgelenk und Schäfteunterschiedlich alter Tiere (RH). F. Radii mit Bissspuren und Schäfte unterschiedlich alter Tiere (RH). G. Ulna-Schäfte unterschiedlichalter Tiere (RH). H. Vertebrae mit großen Bissschramm-/Impakt-Marken auf der Wirbelkörperunterseite (RH). I. Femur-Schäfteunterschiedlich alter Tiere (RH). J. Tibia-Schäfte unterschiedlich alter Tiere (RH). K. Angefressene Pelvis- und Coxa-Resteunterschiedlich alter Tiere (RH).

19
bereich) übrig gelassen (Abb. 11K) – gleichartiges ist vonHyänenfrassresten an Wollnashorn-Becken bekannt (DIEDRICH
2012b). Letztendlich finden sich tatsächlich Caninus-Bissspurenan Höhlenbärenknochen, aber nur in weichen Spongiosa-Berei-chen, ganz besonders am Becken, den Schulterblättern, und –viel wichtiger – an den Unterseiten der Wirbel. Hier sind einigesolcher Wirbel mit 5-8 mm breiten Bissschrammen abgebildet.Identische sind an mehreren Wirbelsäulen von Skelettresten ausder Sophienhöhle untersucht worden.Dort fallen die Caninus-Bissschrammen-Distanzen (= Zahnab-stände) in die Variabilität der Hyänen und Löwen, nicht aber der
Wölfe (DIEDRICH 2013h). Auch die mit drei Bissen versehene Ulnaist recht einmalig im Rübeländer Material, was die C bis I1-3Bissmarken unterscheiden lässt (Abb. 11F), die wiederum in dieGrößenvariabilität von Löwe und Hyäne fallen. Wie in derSophienhöhle angenommen und hier wieder an gleichartigbeschädigten Lenden- bis mittleren Brustwirbeln bestätigt (Abb.11H), haben Löwen und Hyänen der Eiszeit die Eingeweide/inneren Organe der Leibeshöhle herausgefressen. BesondersLöwen fressen diese zuerst, Hyänen meist ebenso, bevor sie andereKörperteile verwerten (DIEDRICH 2012a,b, 2013g). Mit denneuesten Entdeckungen eines Wolfhorstes in der Sophienhöhle
Abb. 12: Knackversuche von Eiszeit-Hyänen an Jungbären-Langknochen (A. Sehr junges Tier, Tiefer Gang, B. ÄlteresJungtier, Saal-Halle) (beide RH) und C. Höhlenbären-Langknochensplitter aus der Hermannshöhle (RH, LDA).

20
(Oberfranken), in dem in Wolfkoprolithen Bärenknochen-fragmente gefunden wurden, auch im Magen angelöste Bären-knochen innerhalb eines Wolf-Fäkalplatzes, müssen auch etlicheBissspuren oder befressene Knochen den Wolfaktivitäten zuge-sprochen werden (DIEDRICH 2013b), was eine exakte Zuordnungeinzelner Bissspuren (auch am Rübeländer Material) meistunmöglich macht. Große rund-ovale Bisseindrücke in Jungbären-Langknochen (Abb. 12) können aber nur der Hyäne zugesprochenwerden und sind nicht wie früher angenommen von den„Fangzähnen der Höhlenbären“ verursacht worden (RABEDER etal. 2000). Kein Raubtier der Welt (Höhlenbären sind ja keine„Raubtiere im klassischen Sinne“) benutzt seine Fangzähne zum„Langknochenknacken“, da diese dabei abbrechen würden. Wiebereits mehrfach dargestellt, sind es die Hyänen-Prämolar-Knackzähne (Abb. 12), die solche Löcher verursachen. Ein ganztypisches Muster der Zerknacktechnik, die eben bei Jungbären-langknochen nicht funktionierte (nicht genug phosphatischverkalkte und daher zu weiche Knochen), ist hier an einem Femuraus der Hermannshöhle demonstriert (Abb. 12A). An einemzweiten doppelt so großen Schaft eines bereits älteren Jungbärenfunktionierte das Zernkacken bereits teilweise (Abb. 12B). Bisheute „glauben“ Archäologen, dass solche „durchlöcherten“ Jung-
bären-Femora „Neandertaler-Knochenflöten“ sein sollen (TURK
et al. 2001) – tatsächlich sind es zweifelsfrei, mehrfach diskutiertund bewiesen lediglich Fraßreste (oval-runde Löcher verursachtdurch Hyänen-Knackzähne = Prämolaren, Abb. 12, 13), die inallen Bärenhorsthöhlen Europas gefunden wurden, mit einemoder vielen Löchern im Schaft (ausführliche Diskussion inDIEDRICH 2011c, 2013h). Diese sind sogar die „Anzeiger für diePräsenz von Hyänen“ (= Indirektnachweis) besonders inBärenhorst-Höhlen, in denen keine Hyänenknochen gefundenwurden.
HyänenAus beiden Höhlen liegen nur 11 Hyänenknochen vor. Auch dieserein historischen Funde lassen sich anhand der Knochenerhaltungbeiden Höhlen und den Fundorten eindeutig zuordnen. Der ersteabgebildete und verschollene Fund von KUNDMANN 1737 (Abb.13A) stammt aus der „Alten Höhle“, also aus dem deningeri-Bärenhorst-Bereich. Dieser sowie drei andere, ähnlich erhalteneund mit Mangandendriten übersäte, orangefarbene und starkfossilisierte Knochen (Abb. 13B-D, Tab. 1) stammen von einerHyäne, die in der Saale-Eiszeit als Crocuta crocuta praespelaea zubezeichnen wäre. Aus dem Knochenfeld oder Ochsenhang fehlen
Abb. 13: Hyänenreste aus der Hermanns- (Bärenhöhlen-Halle) und Baumannshöhle („Alte Höhle“). A-D. Crocuta crocuta aff.praespelaea. A. Unterkieferfragment mit M1 (aus KUNDMANN 1737). B. Humerus mit abgefressenem Gelenkkopf (LMNB). C. RechterMt IV (LMNB). D. Rechte halbe distale Tibia, zerknackt durch Hyänen (Kannibalismus) (LMNB). E-L. Crocuta crocuta spelaea. E.Linkes Maxillare mit P4 aus der Bärenhöhlen-Halle (MLUIFG). F. Linker pathologischer Calcaneus. G. Rechter Femur-Schaft einesJungtieres (LMNB). H. Rechter Mc II (LMNB). I. Rechter Mc IV (LMNB). J. Rechter Mc IV (LMNB). K. Linker Mt II (LMNB). L.Rechter Mt IV (LMNB).

21
interessanterweise Hyänenreste. Jüngere Formen mit Crocuta crocutaspelaea fanden sich nur in der Hermannshöhle. Die rotbraungefärbten Knochen stammen, wie auch KLOOS & MÜLLER (1889)berichten, aus der Bärenhöhlen-Halle. Diese bestehen ausKnochen subadulter (2 x) und ausgewachsener Tiere (6 x), wobeiein Fersenbeinknochen (Abb. 13F) pathologische Wucherungenauf der Knochenoberfläche zeigt. Diese Reste sind aus zweiGründen bedeutsam. Erstens beweisen sie neben den vielenVerbissspuren an Höhlenbärenknochen deren tatsächliche Präsenzüber 40 m tief in der Höhle. Zweitens deuten sie auf einenHyänenhorst im verschütteten Eingangsbereich hin und erklärendamit auch eingeschleppte Nicht-Bärenknochen wie vomMammut (Mammuthus primigenius, 1 x), Wollnashorn (Coelodontaantiquitatis, 2 x), Rentier (Rangifer tarandus, 10 x) oder Steinbock(Capra ibex, 2 x). Solche Nicht-Bärenfaunenreste (Tab. 3) sind inden ersten 10 m häufiger zu erwarten, sofern dieser Bereichtatsächlich als Hyänenhorst (ggf. auch Aufzuchthorst für Jungtiere)genutzt worden ist. Dieses könnten nur Grabungen im ehemaligenEingang verifizieren. Im Vergleich zu anderen Bärenhorsthöhlenin Europa wäre das aber der typische Hyänenhorst amEingangsbereich eines großen Bärenhorstes (z.B. DIEDRICH 2005a,2011a, 2013h, DIEDRICH & ZAK 2006). Die wenigen Reste aus derBaumannshöhle hingegen, die auch noch auf sekundärerLagerstätte gefunden wurden, erlauben keine so deutliche Aussage,aber auch dort wird wahrscheinlich zur deningeri-Höh-lenbärenhorst-Zeit (mittlere Saale-Eiszeit) ein Hyänenhorst imdamaligen Eingang gelegen haben, wobei Knochen mit typischenBissmarken bis in den Tiefen Gang eingeschwemmt gefundenwurden (Abb. 12A).Dass die eiszeitlichen Fleckenhyänen (C. c. spelaea) in der Regionim nördlichen Harzvorland im Eem bis Mittel-Weichsel deutlichmit Clans anwesend waren, beweisen die kürzlich neu inter-pretierten Knochen-Akkumulationsfreilandfundstellen, derHyänenhorst Westeregeln und der Hyänenhorst Quedlinburg-Seweckenberge. Beide liegen ebenfalls entlang der Bodeflussabwärts, wobei dort Hyänen keine Höhlenbären, sonderndie Mammutsteppen-Megafauna erbeuteten und besonders Pferdeoder Bisons jagten (DIEDRICH 2012a, 2013g), aber auch Mammutund Wollnashorn-Kadaverreste zum Horst schleppten – ähnlichwie in der Hermannshöhle im Eingangsbereich (Zahnfragmentvon M. primigenius, Scapula und Zahn von C. antiquitatis).
WölfeDas eiszeitliche Wolfmaterial stammt nur aus der Hermanns-höhle und wurde von dem durch SCHÜTT (1969a) nicht identifi-zierten Wildhundmaterial der Baumannshöhle aussortiert (Fär-bung und alte Etiketten). Ob ggf. auch kleinere Wölfe daruntersind, ist ungewiss. Das hier zusammengestellte historisch gefun-dene und mit Funden aus dem Knochenhaufen 2012 ergänzteMaterial (Abb. 14) ist in den Proportionen deutlich größer alsdas Material vom Wildhund (Cuon alpinus fossilis). Verglichenwurden diese Wolfreste mit noch unpublizierten umfangreichenFunden der Zoolithenhöhle, Oberfranken, Bayern (DIEDRICH inVorber.). Auch diese sind große Wölfe, die mit weiteren Verglei-chen an rezenten Timberwölfen in British Columbia (Kanada)sogar diese größten arktischen Wölfe in den Proportionen nochetwas überschreiten. Gleichartig große Wolfreste wurden auchmit den Sauerland-Höhlen verglichen (Martins-Höhle, Bilstein-Höhle), die einen sehr großen Eiszeitwolf in Mitteleuropa bele-gen, den GOLDFUSS (1823) bereits als Canis lupus spelaeus an ei-nem juvenilen Schädel aus der Zoolithenhöhle beschrieben hat-te. Dieser nach in Höhlen aufgefundenen weichselzeitlichen
Wölfen benannte Name wird derzeit beibehalten. Das überwie-gende Material ist von ausgewachsenen bis senilen Wölfen (19 x),zwei Femurschäfte von Jungtieren sind zerknackt (durch Hyä-nen oder ?Wölfe selbst) in der Saal-Halle aufgefunden worden(Abb. 14O-P). Ein abgefressener Unterschenkel (Tibia, Abb.14R), stammt aus dem Bereich der Bärenhöhlen-Halle.
WildhundeBemerkenswert ist, dass auch hier die Baumannshöhle die um-fangreichsten Cuon-Reste einer europäischen Höhle aufweist. Ca.72 Knochen (einige Rippen und Phalangen unklar) aus der Bau-mannshöhle (Knochenfeld/Ochsenhang, Abb. 15), durch SCHÜTT
(1969a) bestimmt als „Wolf“, werden hier als Cuon alpinus fossilisnach den Publikationen von THENIUS (1954), MALEZ & TURK
(1990), BARYSHNIKOV (1996) oder EHRLINGER & ZENGER (1999)neu bestimmt. Ein ganz bedeutendes Merkmal ist das Fehlendes Unterkiefer-M3 (in Canis vorhanden) sowie deren deutlichkleinere Proportionen zum Wolf (vgl. EHRLINGER & ZENGER
1999). Diese kleinen Proportionen hatte SCHÜTT (1969a) bereitsbei den metrischen Analysen ausgearbeitet (siehe dortOsteometrie), aber nicht die korrekten artlichen Schlussfolge-rungen gezogen. Diese Neubestimmung ist insofern sehr wich-tig, da auch diese überwiegend in das Mittel-Pleistozän (Saale-Eiszeit, zumindest in Westeuropa) datieren (THENIUS 1954,ARGANT 1991, TESTU 2006), wobei osteuropäische Funde (MALEZ
& TURK 1990, BARYSHNIKOV 1996) noch im Spät-Pleistozän bisHolozän vorhanden sind und moderne Vertreter (andere Unter-art) noch heute in Asien leben. Auch in der Zoolithenhöhle sollein „Cuon cf. priscus“ (vermutlich alle Spät-Saale-Formen = C.alpinus fossilis) gefunden worden sein (EHRLINGER & ZENGER 1999),wobei Faunen aus der späten Saale-Eiszeit nach den neustentaphonomischen Studien auch dort vermischt sind (DIEDRICH
2013a). Der von dort abgebildete Unterkiefer ist aber anatomischund in den Zahnmorphologien identisch zu den drei hier abge-bildeten Kiefern aus der Baumannshöhle, auch fällt er in derenGrößenvariabilität (= Geschlechtsdimorphismus). Die Restestammen von mindestens drei Individuen hochadulter bis seni-ler Tiere, was auf sporadische Höhlenbesucher hindeutet undwomöglich nicht auf einen Aufzuchtsort für Jungtiere. StärkererZahnabschliff ist beim überwiegenden Teil des Kiefermaterialsersichtlich (Abb. 15), wobei auch diese Wildhunde nun erstmalsals Kandidaten der Höhlenbärenkadaververwertung in Europaidentifiziert werden müssen, aber zu deren Zeiten deningeri-For-men, die kleiner waren. Damit kommt ein viertes „Großraubtier“als Höhlenbärenkadaververwerter im Pleistozän Europas hinzu.Es ist anzunehmen, dass dieser eine ähnliche Rolle wie späterder Wolf spielte, vielleicht sogar in der Saale-Eiszeit dessen Ni-sche dort vorher einnahm, wo der Wolf nicht vorhanden war(wie hier in Rübeland). Die geringmächtigere Bezahnung lag zwi-schen einem Wolf- und Fuchsgebiss, daher kommt auch dieserHundeartige nicht als „Langknochenknacker“, sondern ebenfallsals nur Restkadaververwerter in Betracht.
LöwenAus der Baumannshöhle stammen 150 Löwenknochen aus demBereich des Knochenfeldes und des Ochsenhanges, die ebenfallsmonographisch gesondert publiziert wurden (DIEDRICH 2013f).Besonders dieses Material zeigt die heterogene Vermischung vonsehr großen saale-eiszeitlichen Panthera leo fossilis (bereits teilweiseerkannt durch SCHÜTT 1969a), sehr kleinen Eem-warmzeitlichenPanthera leo spelaea und typisch mittelgroßen Weichsel-kaltzeit-lichen Panthera leo spelaea-Formen (Abb. 16A, DIEDRICH 2013f).

22
Abb. 14 (oben): Wolf Canis lupus spelaeus, Hermannshöhle (alleBärenhöhlen-Halle, bis auf L. Versturz-Halle, C, P und Q. Saal-Halle, K. Versturzkonus, Untere Schwemmhöhle). A. LinkesMaxillare (LMNB). B. Linker Unterkiefer-I2. C. Rechte Scapula(RH). D. Rechte Ulna (LMNB). E. Rechter Radius (LMNB). F.Rechter Mc III (LMNB). G. Axes (LMNB). H. Cervicalwirbel(LMNB). I. Lumbalwirbel (LMNB). J. Coxa (LMNB). K. Ischium(RH). L. Linker Femur (RH). M. Linker Femur (LMNB). N.Linker distaler Femur (LMNB). O. Linker angefressener Femur(LDA). P. Rechter zerknackter Femur eines Jungtieres (LDA). Q.Rechte Tibia (LDA). R. Linke distale Tibia mit abgefressenemGelenk (RH). S. Rechter Astragalus (LMNB). T. Rechter Calca-neus (LMNB). U. Rechter Mt IV (LMNB).
Abb. 15 (rechte Seite): Wildhund Cuon alpinus fossilis aus demKnochenfeld/Ochsenhang der Baumannshöhle (alle GrabungenBLASIUS, LMNB). A. Maxillare mit M1-3. B. Praemaxillare mit I2-3.C. Maxillare mit M2-3. D. Maxillare mit M1-2. E. Maxillare mit M2.F. Maxillare. G. Zwei Oberkiefer M1. H. Oberkiefer I3. I.Oberkiefer I2. J. Occipital-Condylus. K. Zygomaticum. L. DreiCanini. M. Mandibula. N. Mandibula. O. Mandibula-Fragmentohne Zähne. P. Mandibula. Q. Mandibula. R. Scapula. S.Kompletter und zwei distale Humeri. T. Zwei Ulnae. U. FünfMetacarpi. V. Zwei Radius-Fragmente. W. Vier Halswirbel (Atlas,Axes und C (?3-4)). X. Eine komplette und zwei proximal abge-fressene Tibiae. Y. Zwei Calcanei. Z. Drei Metatarsi.

23

24
Diese nehmen mit 4,8 % einen recht hohen Anteil an der Gesamt-fauna ein. Auch aus dem deningeri-Horstbereich wurden 2012 zweineue Knochen von P. l. fossilis im Tiefen Gang entdeckt (2 % derdortigen Funde), die wiederum in ein typisches Höhlenbären-horst-Borealwald-Faunenspektrum passen. Auch in anderenHöhlenbärenhorsten der Weichsel-Kaltzeit nehmen die Löwenin der Regel um 2 % der Gesamtfaunenknochenreste ein(DIEDRICH 2011b, C). Es ist bekannt, dass die Löwen in den Bären-horsten auf Jagd gingen, was auch andere Löwenskelettfunde ineiner rumänischen großen Bärenhorst-Höhle bewiesen haben(DIEDRICH 2011c). Diese Bärenjagd wird hier erstmals in Deutsch-land und in Europa an einer zweiten Lokalität mit der Hermanns-höhle untermauert. Das Löwenmaterial zweier Löwenskelette ausder Saal-Halle der Hermannshöhle, die übersehen wurden(DIEDRICH 2013f), sind über 90 m tief in der Höhle aufgefundenworden. Löwen waren auch an der Beschädigung von Knochender Höhlenbären beteiligt, aber ihr Gebiss konnte keine massi-ven Langknochen ausgewachsener Höhlenbären in Stücke zer-knacken (DIEDRICH 2011C). Bissschrammen auf den Untersei-ten der Wirbelsäulen (Wirbelkörper) hingegen wurden bereitsfür mehrere Bärenskelette der Sophienhöhle, Oberfranken, Bay-ern, als Beleg für das Ausweiden der inneren Organe/Eingewei-de durch Löwen oder Hyänen angesehen (DIEDRICH 2012b). Ausder Hermannshöhle gibt es sechs solcher Wirbel aus dem Len-den-Brustwirbelbereich (Abb. 11), die identisch mit Bissschädenan Wirbeln der Höhlenbären der Sophienhöhle sind. Auch hiersind die 6-10 mm breiten und tiefen Bissmarken (Bissschrammender Canini/Incisivi) in der Knochenspongiosa der Großraubtierehinterlassen worden.
LeopardenCa. 70 Knochen des eiszeitlichen Leoparden inklusive Schädel-reste (Abb. 16B) und Unterkiefer sind allesamt nur aus demKnochenfeld der Baumannshöhle bekannt (2 % der Faunenreste;DIEDRICH 2013e). Diese stammen von weiblichen und größerenmännlichen und fast ausschließlich ausgewachsenen Tieren, dieim heute noch unbekannten zugeschütteten Höhlenbereich pe-riodisch oder für einige Jahre diesen als Horst genutzt habenmüssen (DIEDRICH 2013e). Auch diese sind offenbar Jäger vonHöhlenbären-Jungtieren gewesen oder haben zumindest Höh-lenbärenkadaver verwertet, was auch für den schweizerischenWildkirchli-Höhlenbärenhorst angenommen wurde (DIEDRICH
2013e). Das Material ist schwer zu datieren und kann vom Ende
der Saale-Eiszeit bis frühen Weichsel-Kaltzeit reichen, woraufdie begleitenden Höhlenbären (Abb. 5) und Vergleiche mit etli-chen anderen europäischen Fundstellen (ARGANT 1991, DIEDRICH
2013e) hindeuten.
Abb. 16: Großkatzenreste aus den Rübeländer Höhlen. A. Schädel und zugehöriger Unterkieferast eines senilen pathologischenLöwen Panthera leo spelaea eines der Skelette aus der Saal-Halle (MB). B. Schädelrest und Unterkiefer (verschiedene Individuenrestekombiniert und teilweise gespiegelt) vom Eiszeit-Leopard Panthera pardus subsp. aus dem Knochenfeld der Baumannshöhle (LMNB).
Ergebnisse
Faunenkomplexe, Datierung, Paläoökologie und Höhlen-nutzung (Horst-Abfolgen)1. Baumannshöhle – Mittel-Saale-Eiszeit, U. deningeri-Höhlenbären-HorstKartiert ist dieser Horstbereich nur in der Baumannshöhle. VomEingang über den Saal bis zur Palmengrotte muss der eigentli-che Horstbereich gelegen haben. Gleichartige Sedimente undgleichartig erhaltene Höhlenbärenknochen (gelb-orange mit den-dritischem Mangan auf der Oberfläche, Abb. 5) identifizierendiesen Bereich als den ältesten Rübeländer Höhlenbären-Horst,der voraussichtlich z.B. mit den deningeri-Bären/Übergangsformenzu U spelaeus subsp. der Saale-Eiszeit der Dechenhöhle (DREYER
& GRAW 2002) zeitlich zu korrelieren wäre. Die P4-Analysen vonZähnen aus dem Tiefen Gang zeigen für U. deningeri typisch pri-mitive dreihöckrige Morphotypen (Abb. 5). Noch deutlicher wirddie Datierung unterstützt durch die wenigen Reste des größtenEiszeitlöwen, Panthera leo fossilis (DIEDRICH 2013f), der nur in derSaale-Eiszeit auftrat. Die Bärenhorstbereiche wurden leiderbereits historisch früh ausgegraben und zerstört, aber die Fundeim Tiefen Gang beweisen fluvialen Transport und Verfrachtungder Knochen in tiefere Höhlenbereiche (Vertikalschächte), genauwie es für die Zoolithenhöhle im gleichen Stil berichtet wurde(DIEDRICH 2013a). Das Material, gewonnen aus dem Tiefen Gang,umfasst etwa 250 Knochen, davon nur 2 Löwenknochen. DieseFaunenkomposition von ca. 99 % Bären- und ca. 1 % Löwen-knochen ist typisch für reine Höhlenbärenhorste in Borealwald-Mittelgebirgszonen der Kaltzeiten Europas generell (DIEDRICH
2013b), wobei dieses nur eine vorläufige Analyse darstellen kann.Mehr Material muss gewonnen und auch absolut datiert werden.Erste Cuon alpinus fossilis-Knochen (ein Calcaneus) gehören ein-deutig in diese Zeit. Nach den Datierungen der vorausgehendenHöhlenbärenhorste mit der P4-Methode müssen die wenigenHyänenreste von Crocuta crocuta praespelaea, die ausschließlich ausder Alten Höhle der Baumannshöhle stammen (Abb. 12),ebenfalls in die Saale-Eiszeit zeitlich datiert werden. Die Faunaist derzeit nur limitiert bekannt (Tab. 1).

25
2. Baumanns- und Hermannshöhle – Spät-Saale- bis Mit-tel-Weichsel-Eiszeit, U. spelaeus subsp.-Höhlenbären-,Leoparden- und ?Wildhund-HorsteIn diesem Zeitabschnitt waren beide Höhlen offen (über die nuntrockene Saale-Ponor-Höhlenetage), wobei die Baumannshöhleoffenbar im älteren Eingang (= historischer touristischer Ein-gang) verblockt war, hingegen aber ein noch unbekannter neuerHöhlen-Hallenteil zur Bode hin geöffnet war (vgl. Abb. 3). Die-ser wurde von kleineren Höhlenbären-Formen der Ursus spelaeuseremus/spelaeus subsp. (Abb. 3-7) als Horst genutzt, nicht abertiefere Höhlenbereiche, nicht einmal der Knochenfeld-Höhlen-teil selbst. Nach der Zahnmorphologie sensu RABEDER (1999)sind aus dem Knochenfeld und angrenzenden „Schwemm-fächern“ vorwiegend P4-Zahnmorphotypen C-D vorhanden, wassomit in das Früh/Mittel-Weichsel datiert (Abb. 5). Für dieseDatierung spricht auch ein Skelett des Pferdespringers Alactagasaliens, welches man in einer „Lösssandlinse“ im Bereich desKnochenfeldes fand (BLASIUS 1901a). Nicht einmal kompletteHöhlenbärenschädel sind derzeit aus dem Knochenfeld in denhistorischen Sammlungen bekannt, die diese Datierungen un-terstützen würden. Hingegen ist bei den Prospektionen ein Schä-del in situ entdeckt und belassen worden, der sich der Morpho-logie der typischen „spelaeus-Gruppe“ zuordnen lässt. Schädeldieser kleineren Höhlenbären sind von den großen U. ingressus(vgl. absolut datierte Schädel aus Süddeutschland in MÜNZEL etal. 2011) zu unterscheiden, die mit etlichen Schädeln in der Her-mannshöhle gefunden wurden und hier abgebildet werden(Abb. 6). Der kleinere U. spelaeus subsp.-Schädel ist nicht als „weib-liche kleinere Form“ zu verwechseln. Er hat neben der kleinerenGröße zwei markante Merkmale: a. fällt der Scheitelkamm ineinem großen Winkel zum Hinterhaupt ab (Abb. 7), und b. sinddie Zähne vom P4-Morphotyp B-D (Abb. 5). Solche Schädel/Zähne sind in der Hermannshöhle offensichtlich besonders ausden „unteren Schichten“ in der Saal-Halle gewonnen worden(besonders Slg. STEINER 1961, VÖLKER 1984/85). Lesefunde ausdem Ende der Unteren Schwemmhöhle der Arbeiten in 2012zeigen von „unteren Schichten“ eingebrochene Stücke, dieebenfalls auf kleinere „spelaeus-Höhlenbären“ auch in anderenHöhlenbereichen der Hermannshöhle hinweisen. In der Arbeitüber „Ursus spelaeus“ aus der Saal-Halle (Material VÖLKER-Gra-bung) konnte FISCHER (1995) aufgrund des damaligen Kennt-nisstandes noch nicht die Höhlenbären-Arten/Unterarten tren-nen, hätte aber anhand der Morphotypen bereits verschiedeneFormen unterscheiden können. Nach den neuen Prospektionenwird deutlich, dass der hier erstmals auskartierte Höhlenbären-horst (Abb. 4) seit dem Früh- und besonders im Mittel-Weichselverstärkt als Bärenhorst fungierte. Dieses erklärt auch die Bären-schliffe an vielen Stellen, die im vorderen Bereich sehr deutlichan zahlreichen Stellen zu sehen sind bzw. 2012 erstmals entdecktwurden (besonders Bereich Bärenhöhlen-Halle). In diese Zeitdes Bärenhorstes fallen auch Knochen von Steppenlöwen(Panthera leo spelaea), die in beiden Höhlen gefunden wurden. Inder Baumannshöhle sind sie im Knochenfeld/Ochsenhang mitEinzelknochen vorhanden, die auf artikulierte Skelette hindeu-ten, die verschwemmt wurden (DIEDRICH 2013f). Hingegen sindin der Hermannshöhle nur in der Saal-Halle zwei von STEINER/VÖLKER völlig übersehene (von FISCHER 1994 auch nur teil-publizierte „Löwenknochen“) komplette männliche Löwen-skelette (zumindest eines stark pathologisch mit Rippenbruchund abgebrochenen Eckzähnen) vorhanden, deren Knochennoch 2012 aus „Knochenhaufen“ im Saal mit acht Knochengerettet wurden (DIEDRICH 2013f). Etliche der Knochen aus dem
Knochenfeld der Baumannshöhle und der Saal-Halle zeigenGroß-Raubtier-Verbiss, der auch schon von STEINER & STEINER
(1969) und FISCHER (1995) zumindest mit einigen Exemplarenerkannt und abgebildet wurde, wobei beide keine Statistiken zumRaubtierverbiss an Bärenknochen lieferten. Der Bärenhorst Her-mannshöhle entspricht nach der Faunenstatistik einem typischenHöhlenbärenhorst des borealen Nadelwaldes der Mittelgebirgs-zonen, in denen 98 % Höhlenbärenknochen und nur 2 %Raubtierknochen vorhanden sind (vgl. DIEDRICH 2013b). DieHöhlenbärenhorst-Knochenstatistik der Baumannshöhlen-Knochenfeld/Ochsenhang-Bereiche hingegen hat ca. 5 %„Fremdknochen“ (Bos/Bison, Megaloceros, Capra, Panthera pardussubsp., Cuon alpinus fossilis, Tab. 2), die auf folgenden zusätzli-chen Ursprung und Überlagerung hindeuten: A. Leoparden-Horst, aus dem die zahlreichen Leopardenreste nur aus diesemBereich der Baumannshöhle stammen (DIEDRICH 2013e), die aberzeitlich auch bis in die späte Saale-Eiszeit reichen könnten. B.Wildhund-Horst mit zahlreichen Knochenfunden nur ausge-wachsener Tiere, die ebenfalls eher in die Saale-Eiszeit datieren.
3. Hermannshöhle – spätes Mittel-Weichsel, U. ingressus-Höhlenbären-?Hyänen-Wolf-HorstDas Hyänen-Material der Hermannshöhle ist zu wenig umfang-reich und stammt nur aus dem vorderen Höhlenbereich bis 30 mhorizontale Tiefe (wenige Funde aus Bärenhöhlen-Halle). Daunter dem Material auch Jungtierreste sind, deutet es auch aufeinen „Aufzuchtshorst“ sensu DIEDRICH (2010) hin. Wie bekannt,werden an solche Höhlenhorste auch die Beutetierstücke ver-schleppt (z.B. DIEDRICH 2005a, 2011a), sodass die wenigen„Nicht-Bärenreste“ vermutlich von Hyänen eingeschleppt wur-den, oder gar Rentierreste von Wölfen. Auch hier sind Jungwölfevorhanden, die wiederum auf einen periodischen Aufzuchts-Horst im verschütteten damaligen „Eingangsbereich“ (vgl.Abb. 4) hindeuten.Nach den P4-Typen und den Schädel-Morphotypen (Abb. 5-7)zu urteilen, sind die letzten großen Höhlenbären nur in den obers-ten Schichten der Hermannshöhle anzutreffen. In dieser Zeitscheinen beide damaligen Eingänge der Baumannshöhle bereitsverblockt zu sein. Durch die Verschließung kamen weder Höh-lenbären noch moderne Menschen (Aurignacien) in das Höhlen-system, hingegen aber in das offene Hermannshöhle-System.Auch hier handelt es sich wiederum um einen reinen Bärenhorst,in dem nur wenige Löwen- (DIEDRICH 2013f) und Wolfreste (Abb.13) gefunden wurden, wobei Hyänen nur indirekt anhand dervielen Bissspuren an den Höhlenbärenknochen (Abb. 10-11)nachzuweisen sind. Wiederum fällt das NISP-Spektrum (98 %Höhlenbär, 2 % andere Karnivoren wie Löwe und Wolf) in einetypische Borealwald-Höhlenbärenhorst-Fauna europäischer Mit-telgebirge (Tab. 3), in der Hyänen und Löwen nur sporadischeBesucher (Kadaververwerter, Höhlenbärenkiller) sind (DIEDRICH
2011c).Neben der Prädation durch Raubtiere ist die Höhlenbärenjagddurch den Cromagnon-Menschen (= Aurignacien) jüngst bewie-sen und das beste Beispiel für Europa überhaupt (DIEDRICH
2013d).
AusblickBeide Rübeländer Höhlensysteme spiegeln das europäischeModell der Höhlengenese mit Nutzung durch Mensch und Tie-re seit der Saale-Eiszeit in einem eiszeitlich-alpinen (bis über1.140 m) und teilweise vergletscherten Mittelgebirgsraum wider.Die Genese ist letztendlich klimatisch und teilweise in sehr kur-

26
zen Zeiträumen gesteuert. Die Rübeländer Höhlen – Baumanns-und Hermannshöhle – weisen gleichartige speläogenetische Ur-sprünge (Ponorhöhlen) auf, haben sich später aber unterschied-lich weiterentwickelt. Sie lassen sich anhand der „Höhlenbären-Uhr“ und der regionalen Vergletscherungsfolgen (Flussterrassen,Talgletscher-Höhenlagen, Höhlenetagen und -sedimente) datie-ren. Die Rübeländer Höhlen haben vier Haupt-Etagen, die wäh-rend der ?Cromer (oder älter), Elster, Saale, Weichsel-Kaltzeitennach unten gestaffelt als Ponorhöhlen und als Untergrund-Ent-wässerung der Bode talseitig entstanden. Die unterste aktivePonorhöhle bildete sich Post-LGM mit Abschmelzen des Bode-Talgletschers, der die Rübeländer Höhlen passierte, und ist derim Holozän aktive Ponor. Die Nutzung der Baumannshöhle istanhand der zusätzlichen Datierung großer Löwen-Unterarten(Panthera leo fossilis) und primitiver dreihöckriger Höhlenbären-zähne aus dem Tiefen Gang über Goethesaal bis damaliger Ein-gang (= historischer Eingang) deutlich älter und in die mittlereSaale-Eiszeit (Mittel-Pleistozän) zu datieren, in der frühe U.deningeri-Höhlenbären nur Teile dieses Systems nutzen konnten.In die mittlere Saale-Eiszeit fallen die größeren, massiverenSpeläothem-Genesen, deren Datierungen grob zwischen 250.00-350.00 (= MIS 7-9) liegen. Diese Höhlenbärenreste wurden nochin der Saale-Eiszeit fluvial verschwemmt (Bode-Flussterrassen-Einfluss) und finden sich daher heute in Vertikalschächten undtiefsten Höhlenbereichen wie dem Tiefen Gang unterhalb desGoethesaals in Sedimenten, die auch einige Bode-Kiese (z.B.Kulmgrauwacken, selten auch vulkanische Gesteine) enthalten.Im Früh-/Mittel-Weichsel (Spät-Pleistozän) waren hingegen beideHöhlen offen, aber die Baumannshöhle nun von einer anderenSeite (östlich, Neue Höhle-Bereich), einem heute wiederum ver-schütteten damaligen Eingang, der zu einer noch unentdecktenHalle führte. Diese war von Neandertalern, Leoparden und Wild-hunden als Horst genutzt worden, aber primär von kleinerenHöhlenbären-Unterarten U. spelaeus eremus/spelaeus, die mit ihrenP4-Zahnmorphotypen besonders im Häufigkeitsmaximum in dasmittlere Weichsel datieren. Wildhundreste von Cuon alpinus fossilisdatieren wiederum in die späte Saale-Eiszeit und zeigen ein vier-tes größeres Raubtier, welches in den Höhlen Bärenkadaver ver-wertete. Diese Faunen-/Artefakt-Reste finden sich aber alle aufsekundärer Lagerstätte tiefer in untere Etagen post-LGM ein-geschwemmt und auch umgelagerte Kalkkies-Randmoränen/Kamesterassen-Sedimente scheinen hier umgelagert und ein-geschwemmt worden zu sein, aber nicht Bodeterrassenkiese, dieeine völlig andere Zusammensetzung und keine Kalkgerölle ent-halten. Auch die Höhlenbären der „unteren Schichten“ desHöhlenbärenhorstes Hermannshöhle datieren mit ihren P4-Zahntypen und kleineren, zum Hinterhaupt stark abfallendenScheitelkamm-Schädeltypen in das frühe/mittlere Weichsel. Indieser Zeit war dort der reine Bärenhorst nur sporadisch vonKarnivoren (Hyänen, Löwen und Wölfen) aufgesucht worden.Besonders Hyänen hinterließen runde Löcher in Jungbären-Lang-knochen und Knochensplitter zerknackter Langknochen, Wölfeund Löwen hingegen nur abgefressene weiche Knochengelenk-enden oder Bissspuren in weichen Knochenteilen. Besondershoch ist der Anteil an beschädigten Jungbärenknochen (ca. 80 %,Hermannshöhle). Hyänenknochen und Wolfreste sind mit we-nigen Knochen bekannt, außerdem zwei komplette (teilweisenoch nicht vollständig ausgegrabene) Löwenskelette von Pantheraleo spelaea (nur aus der Saal-Halle, Hermannshöhle). Diese warenaber nur Höhlen-Besucher, wobei die Löwen Bärenjungtierjägerwaren, Wölfe eher Aasfresser wie auch Hyänen. Neben den Raub-tieren ging auch der moderne Cromagnon-Mensch in der Her-
mannshöhle auf Höhlenbärenjagd auf den zeitgleich lebendengroßen U. ingressus-Höhlenbär. Dieser bildet dort die Oberflä-chen-Bonebeds und ist an den vielhöckrigen P4-Zähnen, seinengroßen treppenartig und im Scheitelkamm fast horizontalenFormen von den kleineren Bärenarten gut zu unterscheiden, wieauch an den größeren Langknochen. Projektile (z.B. LautscherSpitze), Feuerstein-Schneide und -Schabwerkzeuge deuten aufdie Bärenjagd im Aurignacien vor ca. 29.210 BP hin. Da sich dasKlima bereits im Aurignacien verschlechterte, wuchsen auch dieHarz-Gletscher, von denen der Bode-Gletscher sich aus verschie-denen Ästen vom Brocken herabfließend vereinigte und bis zurZeit des „Gravettien“ (Prä-LGM) bereits den Verschluss derRübeländer Höhleneingänge verursachte. In dieser Zeit der ma-ximalen Vereisung scheinen Gletscher-Oberflächenwässer umetwa 24.000 BP eine neue Tropfstein-Generation verursacht zuhaben (besonders Hermannshöhle). Durch die kälteren Klimateentstanden auch die Verschüttungen/ Verblockungen aller Rübe-länder Höhleneingänge, wobei der Bode-Gletscher im LGM amEingang der Hermannhöhle sich sogar auf kalkulierter Höhevon max. 413 m Höhenlage und 30-40 m Mächtigkeit vorbeischob und ggf. sogar Frostbrekzien bis zu 8 m tief in den dama-ligen Eingang hineinpresste. Auch verblockten die Höhlen durchInkasion – große Deckenverstürze deckten die Bären-Bonebedsab, besonders in der Hermannshöhle. Jüngste Speläotheme bil-deten sich dann sogar auf diesen Versturzblöcken um 14-12.000BP (Meiendorf-Alleröd).
DankDie Arbeiten in der Höhle und die Knochenreinigung in der Rübe-länder Höhlenforscherhütte wurden dankenswerterweise von derStadt Oberharz am Brocken und dem Landkreis Harz, UntereNaturschutzbehörde (Nr. 96705-2012/kkr), genehmigt, wofür sehrherzlich gedankt sei. Ein besonderer Dank geht auch an den Bür-germeister F. Damsch der Stadt Oberharz am Brocken und seinenTourismusbetrieb mit den Rübeländer Höhlen und ihrer Geschäfts-führerin C. Hensel, die die Arbeiten stets nach Kräften unterstütz-ten und auch einen finanziellen Zuschuss gaben. Der Arbeitsgemein-schaft für Karstkunde Harz e.V. gilt ein Dank für den Zuschuss unddie unermüdliche inhaltliche und logistische Unterstützung. Dr. F.Knolle half bei der Literaturrecherche, den Höhlenbefahrungen undder gesamten Logistik, E. und M. Schuhose sowie G. Volmer und B.Baumbach (Rübeländer Höhlenforschergruppe) sei Dank für derenEinsatz in der Höhle und die akribische Säuberungsaktion der ca.2.500 Knochen sowie auch für die Unterkunft in der Hütte. Ich dankeweiterhin dem Landesamt für Denkmalpflege und ArchäologieLandesmuseum für Vorgeschichte, hier besonders Dr. R. Mischkaund Dr. B. Stoll-Tucker, für die Unterstützungen in deren Samm-lungen. Prof. Dr. U. Joger erlaubte die mehrfache Bemusterung derRübeländer Höhlenfunde im Staatlichen Naturhistorischen Muse-um Braunschweig, die durch Dr. R. Kosma begleitet wurden. FreieEinsicht in die Sammlungen des Museums für Naturkunde derHumboldt-Universität Berlin wurde durch PD Dr. O. Hampe sehrunterstützt, der auch Fotos bereitstellte. Schließlich danke ich Prof.Dr. B. Rothschild für vorläufige Bestimmungen der Bärenpathologie-Arten.
Literatur
ARGANT A. (1991): Carnivores quaternaires de Bourgogne. – Documentsdes Laboratoires de Géologie Lyon 115: 1-301
BARYSHNIKOV, G. (1996): The dhole, Cuon alpinus (Carnivora, Canidae), fromthe Upper Pleistocene of the Caucasus. – Acta Zoologica Cracoviensia39: 67-73

27
BLASIUS, W. (1890): Neue Knochenfunde in den Höhlen bei Rübeland.– Sitzungsbericht des Vereins für Naturwissenschaft zu Braun-schweig 1890: 1-8
BLASIUS, W. (1898): Spuren paläolithischer Menschen in den Diluvial-Ablagerungen der Rübeländer Höhlen. – Festschrift zur 29. Ver-sammlung der deutschen Anthropologischen Gesellschaft zuBraunschweig 1898: 1-37
BLASIUS, W. (1901a): Bericht über die fortgesetzten Ausgrabungen inden neuen Theilen der Baumannshöhle bei Rübeland im Harzwährend der Jahre 1898 und 1899. – Jahresberichte des Vereinsder Naturwissenschaften Braunschweig 12: 83-86
BLASIUS, W. (1901b): Die Vogelfaunen in den diluvialen Ablagerungender Rübeländer Höhlen. – Journal für Ornithologie 122: 57-60
BRANDES, D. (2009): Blasius: eine Gelehrtenfamilie in Braunschweig. –Einführungsvortrag zur gleichnamigen Ausstellung der Universi-tätsbibliothek Braunschweig (4.11.2009-10.4.2010) Braunschweig.– Universitäts-Bibliothek, http://www.digibib.tu-bs.de/?docid=00031118
BREUER, R. (1931a): Zur Anatomie, Pathologie und Histologie der Zähneund Kiefer von Ursus spelaeus. – Spelaeographische Monographien7(8): 581–610
BREUER, R. (1931b): Pathologisch-anatomische Befunde am Skelettedes Höhlenbären. – Spelaeologische Monographien 7, 8: 611-623
CLARK, P.U., DYKE, A., SHAKUN, J.D., CARLSON, A.E., CLARK, J.,WOHLFARTH, B., MITROVICA, J.X., HOSTETLER, S.W. & MCCABE, A.M.(2009): The Last Glacial Maximum. – Science 325 (5941): 710-714
DIEDRICH, C. (2005a): Eine oberpleistozäne Population von Crocutacrocuta spelaea (GOLDFUSS 1823) aus dem eiszeitlichen Flecken-hyänenhorst Perick-Höhlen von Hemer (Sauerland, NW Deutsch-land) und ihr Kannibalismus. – Philippia 12(2): 93-115
DIEDRICH, C. (2005b): Cracking and nibbling marks as indicators forthe Upper Pleistocene spotted hyaena as a scavenger of cave bear(Ursus spelaeus ROSENMÜLLER 1794) carcasses in the Perick Cavesden of Northwest Germany. – Abhandlungen der Naturhistori-schen Gesellschaft Nürnberg 45: 73-90
DIEDRICH, C. (2006): Die oberpleistozäne Population von Ursus spelaeusROSENMÜLLER 1794 aus dem eiszeitlichen FleckenhyänenhorstPerick-Höhlen von Hemer (Sauerland, NW Deutschland). –Philippia 12(4): 275-346
DIEDRICH, C. (2009): The rediscovered cave bear „Ursus spelaeus Rosen-müller, 1794“ holotype of the Zoolithen Cave (Germany) fromthe historic Rosenmüller collection. – Acta Carsologica Slovacia47(1): 25-32
DIEDRICH, C. (2010): A diseased Panthera leo spelaea (GOLDFUSS 1810)lioness from a forest elephant graveyard in the Late Pleistocene(Eemian) interglacial lake at Neumark-Nord, central Germany. –Historical Biology 23(2-3): 195-217
DIEDRICH, C. (2011a): The Late Pleistocene spotted hyena Crocuta crocutaspelaea (GOLDFUSS 1823) population from the Zoolithen Cave atGailenreuth (Bavaria, South Germany) – a hyena cub rising denof specialized cave bear scavengers in Boreal Forest environmentsof Central Europe. – Historical Biology 23(4): 335-367
DIEDRICH, C. (2011b): The largest European lion Panthera leo spelaea(GOLDFUSS) population from the Zoolithen Cave, Germany –specialized cave bear predators of Europe. – Historical Biology23(2-3): 271-311
DIEDRICH, C. (2011c): Cave bear killers and scavengers from the last iceage of central Europe: Feeding specializations in response to theabsence of mammoth steppe fauna from mountainous regions. –Quaternary International 255: 59-78
DIEDRICH, C. (2012a): Late Pleistocene Crocuta crocuta spelaea (GOLDFUSS
1823) clans as prezewalski horse hunters and woolly rhinocerosscavengers at the open air commuting den and contemporaryNeanderthal camp site Westeregeln (central Germany). – Journalof Archaeological Science 39(6): 1749-1767
DIEDRICH, C. (2012b): The Late Pleistocene Crocuta crocuta spelaea(GOLDFUSS 1823) population from the Emscher River terrace hyenaopen air den Bottrop and other sites in NW-Germany – woollyrhinoceros scavengers and their bone accumulations along riversin lowland mammoth steppe environments. – Quaternary Inter-national 276/277: 93-119
DIEDRICH, C. (2013a): Pleistocene Stratigraphy and bone taphonomy inthe Zoolithen Cave – and new theory about Esper’s „biblic flood“with a first geomorphological Wiesent Valley and river terraceevolution model – ?small glaciers and high/postglacial flood eventsin Upper Franconia. – Eiszeitalter und Gegenwart, QuaternaryScience Journal 62(2) (angenommen)
DIEDRICH, C. (2013b): Ice Age geomorphological Ahorn Valley andAilsbach River terrace evolution– and its importance for the caveuse possibilities by cave bears, top predators and Palaeolithichumans in the Franconian Karst – case studies in the Sophie’sCave, Bavaria. – Eiszeitalter und Gegenwart, Quaternary ScienceJournal 62(2) (angenommen)
DIEDRICH, C. (2013c): Impact of the German Harz MountainWeichselian ice-shield and valley glacier development ontoPalaeolithics and megafauna disappearance. – Geomorphology (imReview)
DIEDRICH, C. (2013d): Oldest most northern Late Palaeolithic Cavebear hunters in Europe. – Im Review
DIEDRICH, C. (2013e): A review of Pleistocene leopards across Europe– most northern European population, highest elevated recordsin the Alps, complete skeletons in the Dinarids and comparison tothe Ice Age cave art. – Im Review
DIEDRICH, C. (2013f): Middle and Late Pleistocene lion remains andskeletons from the Rübeland Caves, northern Germany – newproves for their cave bear hunting deep in cave bear dens. – InVorber.
DIEDRICH, C. (2013g): The Unicornu fossile VALENTINI 1714 „holotype“skeleton from the Ice Age spotted hyena commuting den at theopen air gypsum karst Quedlinburg Sewecken-Berge (CentralGermany). – Historical Biology (im Review)
DIEDRICH, C. (2013h): Extinctions of Late Ice Age Cave Bears as aResult of Climate/Habitat Change and Large Carnivore Lion/Hyena/Wolf Predation Stress in Europe. – ISRN Zoology 2013,Artikel 138319, 25 S., http://dx.doi.org/10.1155/2013/138319
DIEDRICH, C. (in Vorber.): The wolf population from the ZoolithenCave (Bavaria, Germany) – and taphonomic contributions to theextinct European large Ice Age wolf subspecies Canis lupus spelaeus(GOLDFUSS 1823) – hyena prey imported to dens or wolf cub raisingcaves?
DIEDRICH, C. & ZAK, K. (2006): Prey deposits and den sites of theUpper Pleistocene hyena Crocuta crocuta spelaea (GOLDFUSS 1823) inhorizontal and vertical caves of the Bohemian Karst (CzechRepublic). – Bulletin of Geosciences 81 (4): 237-276
DIEDRICH, C. & WEBER, T. (2013): A Middle Palaeolithic Neanderthalcamp and contemporaneous hyena open air den site on a gypsumkarst hill at Westeregeln (north-central Germany) within amammoth steppe environment. – Journal of World Prehistory (imReview)
DÖHLE, H.J. & WEBER, T. (2004): Höhlenbewohner in Mitteldeutsch-land? Die Funde aus Rübeland. – Katalog zur Dauerausstellungim Landesmuseum für Vor- und Frühgeschichte Halle 1 (1): 197-200
DREYER, R. & GRAW, R. (2002): Die Bären im Sauerland: Knochen-funde aus der Dechenhöhle bei Iserlohn. – Abhandlungen zurKarst- und Höhlenkunde 34: 30-35
EHRLINGER, S. & ZENGER, K. (1999): Ein Cuon-Fund aus der Zoolithen-höhle, morphologische und biostratigraphische Studien unter Ver-wendung einer ACCESS-Datenbank. – Quartär 49/50: 55-86

28
FISCHER, K. (1994): Neufunde von jungpleistozänen HöhlenlöwenPanthera leo spelaea (GOLDFUSS 1810) in Rübeland (Harz). – Braun-schweiger Naturkundliche Schriften 4(3): 455-471
FISCHER, K. (1995): Der Höhlenbär Ursus spelaeus Rosenmüller &HEINROTH, 1794 von Rübeland (Harz). – Braunschweiger natur-kundliche Schriften 4(4): 709-742
FRICKE, U. (1998): Die Neuvermessung der Baumannshöhle in Rübe-land. – Mitteilungen des Verbandes der deutschen Höhlen- undKarstforscher 44(3): 100-115
FRICKE, U. & KNOLLE, F. (1999): Naturpark Harz. Die Rübeländer Tropf-steinhöhlen. – Studio V. Schadach, Goslar, 80 S.
GOLDFUSS, G.A. (1823): Osteologische Beiträge zur Kenntnis verschie-dener Säugethiere der Vorwelt. VI. Ueber die Hölen-Hyäne (Hyaenaspelaea). – Nova Acta Physico-Medica Academiea CaesaraeLeopoldino-Carolinae Naturae Curiosorum 3(2): 456-490
GROSS, C. (1992): Das Skelett des Höhlenlöwen (Panthera leo spelaeaGOLDFUSS 1810) aus Siegsdorf/Ldkr. Traunstein im Vergleich mitanderen Funden aus Deutschland und den Niederlanden. –Unveröff. Dissertation, Tierärztliche Fakultät der Maximilians-Universität München, 129 S.
GROTRIAN, H. (1878): Über das Vorkommen von Knochenresten derquartären Säugethier-Fauna in den Höhlen und Spalten des Devon-kalkes von Rübeland am Harz. – Zeitschrift der deutschen Geolo-gischen Gesellschaft 30: 552-553
GRÜNBERG, J. (2006): New AMS dates for Palaeolithic and Mesolithiccamp sites and single finds in Saxony-Anhalt and Thuringia(Germany). – Proceedings of the Prehistoric Society 72: 95-112
HOFREITER, M. (2002): Genetic stability and replacement in latePleistocene cave bear populations. – Abhandlungen zur Karst-und Höhlenkunde 34: 64-67
KAULICH, B. & ROSENDAHL, W. (2002): The Neonate cave bear skeletonfrom the Petershöhle near Velden (Franconian Alb, Germany). –Abhandlungen zur Karst- und Höhlenkunde 34: 12-16
KEMPE, S. (2006): The Baumann’s Cave at Rübeland, Harz, Germany,one of the caves noted in early science history for its cave bearand cave hyena bone deposits. – Scientific Annales, School ofGeology Aristotele University of Thessaloniki Greece SpecialVolume 98: 213-220
KLOOS, J.H. & MÜLLER, M. (1889): Die Hermannshöhle bei Rübeland.– K. Schwier-Verlag, Weimar, 76 S.
KNOLLE, F., OESTERREICH, B., SCHULZ, R. & WREDE, V. (1997): Der Harz– Geologische Exkursionen - Perthes-Exkursionsführer. – JustusPerthes Verlag, Gotha, 230 S.
KÜCHELMANN, H.C. (1997): Bißspuren von Säugetieren an rezenten undsubfossilen Knochen. – Unveröff. Diplom-Arbeit, Carl vonOssietzky-Universität Oldenburg, 127 S.
KUNDMANN, J.C. (1737): Rariora Naturae et Artis item in Re Medica,oder Seltenheiten der Natur und Kunst des KundmannischenNaturalien-Kabinetts wie auch in der Arzney-Wissenschaft. –Michael Hubert, Breslau und Leipzig, 368 + 312 Spalten + 18 S.,Index
KURTÉN, B. (1955): Sex dimorphism and size trends in cave bear, Ursusspelaeus ROSENMÜLLER und HEINROTH. – Acta Zoologica Fennica90: 1-48
MALEZ, M. & TURK, I. (1990): Cuon alpinus europaem BOURGUIGNAT
(Carnivora, Mammalia) from the Upper Pleistocene in the CaveApnarjeva jama et Celje. – Geologija 33: 215-231
MELLER, H. (2008): Lebenswandel. Früh- und Mittelneolithikum. –Begleithefte zur Dauerausstellung im Landesmuseum für Vorge-schichte Sachsen-Anhalt Band 3. Halle/Saale, 234 S.
MÜNZEL, S.C., STILLER, M., HOFREITER, M. MITTNIK, A., CONARD, N.J.& BOCHERENS, H. (2011): Pleistocene bears in the Swabian Jura
(Germany): Genetic replacement, ecological displacement,extinctions and survival. – Quaternary International (im Druck)
NIELBOCK, R. (1998): Faunen des Eiszeitalters. Funde und Grabungenin Schlotten und Höhlen des Südharzes. – NNA-Berichte 11(2):61-70
NORDHOFF, P. (2005): Stable isotope investigations on speleothems fromdifferent cave systems in Germany. – Unveröff. Dissertation, GeorgAugust-Universität Göttingen, 149 S.
PACHER, M. & STUART, A.J. (2008): Extinction chronology andpalaeobiology of the cave bear (Ursus spelaeus). – Boreas 38(2):189-206
RABEDER, G. (1999): Die Evolution des Höhlenbärgebisses. – Mittei-lung der Kommission für Quartärforschung der ÖsterreichischenAkademie der Wissenschaften 11: 1-102
RABEDER, G., NAGEL, D. & PACHER, M. (2000): Der Höhlenbär. –Stuttgart, Thorbecke, 111 S.
RABEDER, G., HOFREITER, M., NAGEL, D., PAABO, S. & WITHALM, G.(2002): Die neue Taxonomie der Höhlenbären. – Abhandlungenzur Karst- und Höhlenkunde 34: 68-69
RABEDER, G. & HOFREITER, M. (2004): Der neue Stammbaum der Höh-lenbären. – Die Höhle 55: 1-19
RANAL, R.S., WU, J.S. & EISENBERG, R.L. (2009): Periosteal Reaction. –American Journal of Roentgenology 193(4): 259-272
ROSENDAHL, W. DÖPPES, D., JOGER, U., LASKOWSKI, R., LÓPEZ CORREA,M., NIELBOCK, R. & WREDE, V. (2005): New radiometric datingsof different cave bear sites in Germany – results and interpre-tations. – Bulletin de la Société Histoire Naturelle Toulouse 141(1):39-46
ROTHSCHILD, B.M. & DIEDRICH, C. (2012): Comparison of pathologiesin the extinct Pleistocene Eurasian steppe lion Pantherea leo spelaea(GOLDFUSS 1810) to those in the modern lion, Panthera leo – Resultsof fights with hyenas, bears and lions and other ecological stress.– International Journal of Paleopathology (im Druck)
SCHÜTT, G. (1968): Die cromerzeitlichen Bären aus der Einhornhöhlebei Scharzfeld. – Mitteilungen aus dem Geologischen Institut derTechnischen Hochschule Hannover 7: 1-21
SCHÜTT, G. (1969a): Die jungpleistozäne Fauna der Höhlen bei Rübe-land im Harz. – Quartär 20: 79-125
SCHÜTT, G. (1969b): Untersuchungen am Gebiß von Panthera leo fossilis(REICHENAU 1906) und Panthera leo spelaea (Goldfuss 1810). – NeuesJahrbuch für Geologie und Paläontologie Abhandlungen 134: 192-220
STEINER, U. (1964): Zwei neue paläolithische Artefakte aus der Her-mannshöhle bei Rübeland /Harz. – Ausgrabungen und Funde 9:17-18
STEINER, U. & STEINER, W. (1969): Zur Genese der Rübeländer Höhlen.– Geologie 18: 198-203
STILLER, M., BARYSHNIKOV, G., BOCHERENS, H., GRANDAL D’ANGLADE,A., HILPERT, B., MÜNZEL, S.C., PINHASI, R., RABEDER, G.,ROSENDAHL, W., TRINKAUS, E., HOFREITER, M. & KNAPP, M. (2010):Withering away – 25,000 years of genetic decline preceded cave bearextinction. – Molecular Biology and Evolution 27(5): 975-978
TESTU, A. (2006): Etude paléontologique et biostratigraphique desFelidae et Hyaenidae pléistocènes de l’Europe méditerranéenne. –Unveröff. Dissertation Université Perpignan, 359 S.
THENIUS, E. (1954). Zur Abstammung der Rotwölfe (Gattung CuonHODGSON). – Österreichische Zoologische Zeitschrift 5: 377-388
TURK, I., DIRJEC, J., BASTIANI, G., PFLAUM, M., LAUKO, T., CIMERMAN, F.& KOSEL, F. (2001): Grum C. Nove analize piscali iz Divji bab I(Slovenija). – Archeoloski vestnik 52: 25-79
TÖPFER, V. (1954): Stand und Aufgaben der urgeschichtlichen Erfor-

29
Anschrift des Autors: Dr. Cajus Diedrich, PaleoLogic, PetraBezruce 96, CZ-26751 Zdice, Tschechien, [email protected],www.paleologic.de
schung der Harzhöhlen bei Rübeland. – Jahresschrift für Mittel-deutsche Vorgeschichte 38: 1-36
TÖPFER, V. (1956): Ein neues Feuersteingerät aus der Altsteinzeit ausder Hermannshöhle bei Rübeland in Harz. – VorgeschichtlicheMuseumsarbeit und Bodendenkmalpflege 3: 9-11
TÖPFER, V. (1967): Steinzeitliche Funde aus den Rübeländer Höhlen imHarz. – Ausgrabungen und Funde 12: 1-3
VÖLKER, R. (1985): Die Grabungen auf Bärenknochen im Frühjahr1984 in der Hermannshöhle Rübeland. – Berichte und Mitteilun-gen Zentraler Fachausschuss für Höhlen- und Karstforschung85(1): 5-7
Spezies Anzahl/Fundort Sammlung
Ursus deningeri 260 (Tiefer Gang) RHPanthera leo fossilis 2 (Tiefer Gang) RHCrocuta crocuta aff. praespelaea 3 („Alte Höhle“) LMNBCuon alpinus fossilis 1 („Alte Höhle“) LMNB
Tab. 1: Knochen per Spezies (NISP = 266) der mittleren Saale-Bärenhorst-Schichten (allochthon) der „Alten Baumannshöhle“ (TieferGang und Goethesaal-Bereiche), Rübeland
Spezies Anzahl/Fundort Sammlung
Mammuthus primigenius 1 (Bärenhöhlen-Halle) LMNBCoelodonta antiquitatis 2 (Bärenhöhlen-Halle) LMNBRangifer tarandus 10 (Bärenhöhlen-Halle) LMNB, RHCapra ibex 2 (Bärenhöhlen-Halle) LMNB, RHRupicapra rupicapra 2 (Bärenhöhlen-Halle) LMNB, RHUrsus spelaeus subsp./U. ingressus 1.250 (Grabung Steiner 1961), 2.230
(Grabung Völker 1984/85), 1.430(Knochenhaufen 2012)= 4.910 (gesamte Höhle) LMNB, LDA, RH, MB und in Höhle
Crocuta crocuta spelaea 8 (Bärenhöhlen-Halle) LMNBPanthera leo spelaea 72 (Grabung Steiner 1961),
56 (Grabung Völker 1984/85),10 (Knochenhaufen 2012)= 132 (Saal-Halle, 2 Skelette) LDA, RH, MB
Canis lupus spelaeus 22 (Saal-Halle, Versturz-Halle,Bärenhöhlen-Halle) LMNB, LDA, RH, MB
Vulpes lagopus 1 (Bärenhöhlen-Halle) RH
Tab. 3: Knochen per Spezies (NISP = 5.090) aus den Früh-/Mittel-Weichsel-Bärenhorst-Schichten (autochthon) der Hermannshöhle,Rübeland
Spezies Anzahl/Fundort Sammlung
Bison/Bos sp. 6 (Knochenfeld/Ochsenhang) LMNBMegaloceros giganteus 3 (Knochenfeld/Ochsenhang) LMNBCervus elaphus 1 (Knochenfeld) LMNBCapra ibex 2 (Knochenfeld/Ochsenhang) LMNBUrsus deningeri/spelaeus subsp. 2.640 (Knochenfeld/Ochsenhang) LMNB, RH, und in HöhlePanthera leo spelaea/fossilis 151 (Knochenfeld/Ochsenhang) LMNB, RHPanthera pardus subsp. 75 (Knochenfeld/Ochsenhang) LMNB, RHCuon alpinus fossilis 68 (Knochenfeld/Ochsenhang) LMNB
Tab. 2. Knochen per Spezies (NISP = 2.946) der Spät-Saale- bis mittleren Weisel-Bärenhorst-Schichten (allochthon) der Baumannshöhle(Knochenfeld/Ochsenhang-Bereiche), Rübeland
WELLER, H. (1991): Facies and development of the Devonian (Givetian/Frasnian) Elbingerode reef complex in the Harz Area (Germany).– Facies 25: 1-50
WELLER, H. (2011): Eine neue Geologische Karte des ElbingeröderKomplexes/Harz – GK EK 2011. – Hallesches Jahrbuch Beiheft28: 57-62

30
Jahresbericht 2012 von AK Biospeläologie und Referat Bio-speläologieRheinland-Pfalz (WEBER)Insgesamt wurden 25 (Vorjahr 63) biospeläologische Exkursio-nen durchgeführt, davon 5 (10) in Naturhöhlen und 20 (49) inkünstliche Hohlräume mit insgesamt 30 (128) Ethan-diol-1,2-Fallen. Insgesamt wurden rund 2.700 (10.000) Tiere gesammelt.Für 2012 ist besonders die Erfassung und Determination derAraneae, Dermaptera, Formicidae und Trichoptera hervorzuhe-ben. Die Funddaten der Jahre 2001-2005 wurden Anfang 2012publiziert (WEBER 2012a). Die Erfassung wird fortgeführt.Disziplinübergreifend wurde in Zusammenarbeit mit derGeneraldirektion kulturelles Erbe Rheinland-Pfalz, DirektionLandesarchäologie, seit 2010 die Fauna der unterirdischen Gän-ge der Zitadelle Mainz erforscht. Die Ergebnisse wurden als klei-nes Heft veröffentlicht (WEBER 2012b), das in Form einer Prä-sentation bei der gemeinsamen Jahrestagung von VdHK undVÖH Speleo-Austria in Bad Mitterndorf sowie in Mainz in ei-nem öffentlichen Vortrag vorgestellt wurde.
Saarland (WEBER)Insgesamt wurden 25 (28) biospeläologische Exkursionen durch-geführt, davon in 1 Naturhöhle (0) und 24 (23) künstliche Hohl-räume mit insgesamt 0 (185) Ethan-diol-1,2-Fallen. Dabei wur-den rund 1.000 (3.000) Tiere gesammelt. Eine der Exkursionenfand in Kooperation mit der Delattinia Naturforschenden Ge-sellschaft des Saarlandes statt (WEBER 2012c).Für 2012 ist besonders die Erfassung und Determination derGastropoda, Formicidae und Trichoptera hervorzuheben. DieFunddaten der Jahre 2001-2005 wurden Anfang 2012 publiziert(WEBER 2012a). Die Erfassung wird fortgeführt.Die Funde der letzten Jahre wurden bei der Delattinia Natur-forschenden Gesellschaft des Saarlandes in Form eines Abend-vortrags vorgestellt.
Hessen (ZAENKER)Mit Stand vom 31.12.2012 waren im Bearbeitungsgebiet des Bio-speläologischen Katasters von Hessen 3.036 (2.965) Taxa nach-gewiesen, die mindestens bis zur Gattung bestimmt wurden. Inder Datenbank sind derzeit 126.022 (118.315) Einzelfunde aus6.249 (5.983) Objekten erfasst. An den biospeläologischenForschungsarbeiten waren wie schon in den vergangenen Jahrensämtliche Mitgliedsorganisationen des Landesverbands für Höh-len- und Karstforschung Hessen e.V. beteiligt.Im Bereich des FFH-Gebiets „Gipskarst bei Berneburg“ wurdeauch der letzte Felsenkeller mit einem fledermausgerechten Ver-schluss versehen. Damit sind die Sicherungsarbeiten dort vorerstabgeschlossen.Im Jahr 2012 lag der Schwerpunkt der Quellenkartierung wiederim Nationalpark Kellerwald-Edersee und im BiosphärenreservatRhön. Im Biosphärenreservat sind derzeit 1.882 (1.741) Quellenbearbeitet. Für die hessische und die thüringische Verwaltungs-stelle des Biosphärenreservats Rhön wurden drei umfangreicheGutachten zur Fauna und zur Gefährdung der Quellen imSchwarzwald bei Wüstensachsen, im Bereich Oberweid undKaltenwestheim sowie im Schmerbachquellgebiet westlich vonAndenhausen erstellt. Im Nationalpark Kellerwald-Edersee sindzwischenzeitlich 585 (562) Quellen erfasst und zoologisch kar-tiert.
Im Bereich der Öffentlichkeitsarbeit des BiospeläologischenKatasters wurden mehrere Presse- und Fachartikel zur Quellen-kartierung publiziert. Zu diesem Thema und zu fleder-mauskundlichen Themen wurden verschiedene Führungen undVorträge für die Öffentlichkeit angeboten. Die Erfassung imLiteraturverwaltungsprogramm Citavi wurde fortgesetzt.Zwischenzeitlich kann auf über 38.000 (27.000) Literaturdaten-sätze zugegriffen werden.Zusammen mit Dorian Dörge (Universität Frankfurt am Main)wurde im Rahmen seiner Masterarbeit ein Führer zur Fauna desHohlsteins zwischen Hilgershausen und Kammerbach entwor-fen, der 2013 publiziert werden soll. Ein weiterer Teil der Master-arbeit beschäftigte sich mit dem DNA-Barcoding der Höhlen-pilzmücke (Speolepta leptogaster). Das Biospeläologische Katastervon Hessen bildet den Grundstock für die DNA-Erfassung derdeutschen Höhlentiere, die derzeit anläuft.Unterstützt wurde der Landesverband für Höhlen- und Karst-forschung Hessen e.V. bei seiner biospeläologischen Tätigkeitvom Nationalparkamt Kellerwald-Edersee, den Verwaltungs-stellen des Biosphärenreservats Rhön, den Unteren und OberenNaturschutzbehörden und dem Hessischen Ministerium fürUmwelt, ländlichen Raum und Verbraucherschutz (Hessen-ForstFENA, Abteilung „Naturschutzdaten“). Mit dieser Behördewurde auch ein fünfjähriger Vertrag zur „Freiwilligen Arbeit imNaturschutz“ abgeschlossen, der den Mitgliedern des hessischenLandesverbands umfangreiche Betretungsrechte für Untertage-quartiere einräumt. Im Gegenzug erhält das Land Hessen in re-gelmäßigen Abständen die erhobenen biospeläologischen Art-daten.Bei verschiedenen Tagungen der Arbeitsgemeinschaft Arten-schutz im Biosphärenreservat Rhön waren Mitarbeiter des bio-speläologischen Katasters vertreten. Die Interessen der Bio-speläologie sowie des Fledermaus- und Quellenschutzes werdenweiterhin im Naturschutzbeirat des Landkreises Fulda, im Bei-rat des Biosphärenreservats Rhön, im Verein RhönNatur e.V.und im Forum „Naturschutz und Kulturlandschaft“ des VereinsNatur- und Lebensraum Rhön vertreten. Auf verschiedenenFachtagungen konnten die biospeläologischen Forschungen imRahmen von Vorträgen vorgestellt werden.Auch 2012 konnten zahlreiche Archivproben von ehrenamtli-chen Bestimmern aus ganz Europa bearbeitet werden, denenauf diesem Wege für ihre hervorragende Arbeit gedankt werdensoll. Informationen zum Biospeläologischen Kataster und dieaktuellen Forschungsergebnisse finden sich im Internet unterwww.hoehlenkataster-hessen.de.
Tätigkeitsberichte
Abb. 1: Teilnehmer des biospeläologischen Seminars; Foto:Zaenker

31
Bayern (ZAENKER)Am Wendelstein (Bayerische Alpen) wurde für den VdHK eindreitägiges biospeläologisches Seminar ausgerichtet, an dem 16interessierte Höhlenforscher aus ganz Deutschland teilnahmen.Die umfassende Untersuchung der Wendelsteinhöhle brachte alsErgebnis gleich zwei Erstnachweise für Deutschland – die Scheu-fliege Gymnomus soosi und den Springschwanz Schaefferia sexoculata.
Sachsen (ARNHOLD)Zwei Schwerpunkte kennzeichneten die Arbeit. Im Rahmen derErgänzung des biospeläologischen Katasters wurden die Sand-steinhöhlen im Bielatal um die Felsgruppe der Bennohöhle un-tersucht. Die Flora und Fauna ist in diesem Bereich stark vonKletterinteressen sowie Abenteuertouristik geprägt und birgt einSpannungsfeld gegenüber Interessen des Naturschutzes.Im Auftrag des Sächsischen Landesamts für Umwelt, Landwirt-schaft und Geologie erfolgten in Fortsetzung die naturschutz-fachlichen Bewertungen von Höhlen. Dabei wurde die Alten-dorfer Dorfbachhöhle im Kirnitzschtal untersucht. Sie zählt zuden wenigen regionalen, von einem Bach durchflossenen Höh-len mit einer vielfältigen Fauna.Exkursionen zu künstlichen Hohlräumen galten der Erfassungcavernicoler Vorkommen. Hier sind Funde zu Amphibien undInsekten hervorzuheben.Die regionalen Beobachtungen zur Großen Höhlenspinne (Metamenardi) wurden fortgesetzt. Eine Veröffentlichung in den Mit-teilungen der Höhlen- und Karstforschung Dresden e.V. beschäf-tigt sich mit dem Höhlentier des Jahres.
Tag der Artenvielfalt (WEBER, ARNHOLD)Beim von der Zeitschrift GEO veranstalteten und von der HeinzSielmann-Stiftung geförderten Tag der Artenvielfalt, dessenHauptveranstaltung 2012 ins Grenzgebiet Pfalz/Elsass führte,konnten in Zusammenarbeit mit lokalen Fledermausforschernzwei Wehrmachtsstollen befahren und biologisch bearbeitetwerden. Der Bericht hat es immerhin bis in GEO Online ge-schafft (BROSCHART 2012). Einige der gesammelten Tiergruppen,allen voran die Spinnen, konnten direkt vor Ort von Expertendeterminiert werden.In einer lokalen Aktion zum Tag der Artenvielfalt wurden dieSandsteinhöhlen im Bielatal (Sächsische Schweiz) um die Fels-gruppe der Bennohöhle untersucht (ARNHOLD 2012b). Das Er-gebnis der Erfassung belegt unsere Erfahrung, dass touristischinteressante Gebiete nur wenig Raum für besondere Funde bie-ten. Trotzdem hält sich ein fester Stamm von Höhlenbewoh-nern, wie auch Langzeitbeobachtungen belegen. Hierzu zählenu.a. Vorkommen von Streckerspinnen (Tetragnathidae) undKöcherfliegen (Trichoptera).
Bibliographie (WEBER)Die in der letzten Version 2000 veröffentlichte Bibliographiewurde auch 2011 weitergeführt, um zu gegebener Zeit eine Ver-sion 3 veröffentlichen zu können.
Höhlentier des Jahres (ZAENKER, WEBER)Das Höhlentier des Jahres 2012 ist die Spinne Meta menardi (VO-GEL et al. 2012). Es ist uns gelungen, sie gleichzeitig von derArachnologischen Gesellschaft und der European Society ofArachnology zur Spinne des Jahres (HÖRWEG 2012) ausrufen zulassen, was unser „Höhlentier des Jahres“ auf europäische Ebe-ne hebt. Das Tier wurde in zahlreichen regionalen und überregi-onalen Presseberichten und Vorträgen vorgestellt.
FFH-Richtlinie (ZAENKER, WEBER)Der Schwerpunkt der Arbeiten lag auch im Jahr 2012 auf derbiospeläologischen Erfassung der im bundesweiten FFH-Monitoring zu untersuchenden Naturhöhlen. Nachdem Hessenbereits abgearbeitet war, wurden 2012 in Rheinland-Pfalzinsgesamt vier Höhlen bearbeitet und ein entsprechendes Gut-achten erstellt. Damit ist das FFH-Monitoring für Hessen undRheinland-Pfalz nach den gleichen einheitlichen Bewertungs-richtlinien und Untersuchungsstandards erfolgt. Ferner wurdeeine Höhle in Unterfranken für das bayerische FFH-Monitoringuntersucht.
Literatur
ARNHOLD, M. (2012a): GEO-Tag der Artenvielfalt 2012, Exkursions-bericht. – Mitteilung Höhlen- und Karstforschung Dresden 2012
ARNHOLD, M. (2012b) Ergebnisse und Artenlisten zum GEO-Tag derArtenvielfalt. – www.geoartenvielfalt.de/aktionen/2012
ARNHOLD, M. (2012c) Die Große Höhlenspinne / Meta menardi im Elb-sandsteingebirge. Das Höhlentier 2012. – Mitteilung Höhlen- undKarstforschung Dresden 2012
BROSCHART, J. (2012): Artenvielfalt im Pfälzerwald. – www.geo.de/GEO/natur/oekologie/tag_der_artenvielfalt/geo-tag-der-artenvielfalt-artenvielfalt-im-pfaelzerwald-71936.html
HÖRWEG, C., BLICK, T. & ZAENKER, S. (2011): Die Große Höhlenspinne,Meta menardi (Araneae: Tetragnathidae), Spinne des Jahres 2012. –Arachnologische Mitteilungen 42: 62-64
HÖRWEG, C., BLICK, T. & ZAENKER, S. (2011): Die Große HöhlenspinneMeta menardi (LATREILLE, 1804) – Höhlentier des Jahres und Euro-päische Spinne des Jahres 2012. – Mitt. Verb. dt. Höhlen- u. Karst-forscher 57(4): 108-109
HÖRWEG, C., BLICK, T. & ZAENKER, S. (2012): Meta menardi. Die GroßeHöhlenspinne begeistert Arachnologen und Höhlenforschergleichermaßen. – Senckenberg natur - forschung - museum. DasSenckenberg-Wissenschaftsmagazin 142(3/4): 134-135
SCHNEIDER, N., ZAENKER, S. & DOROW, W. H. O. (2012): Staubläuse(Psocodea, ‚Psocoptera‘) aus zoologischen Untersuchungen inHessen und den angrenzenden Gebieten. – Hessische Faunisti-sche Briefe 31(1/2): 1-30
STEINKE, D., ZAENKER, S., VOGEL, B. & WEIGAND, A.M. (2012): Vonkryptischen Arten und Lebensräumen – DNA-Barcoding trifft auf
Abb. 2: Meta menardi, Benno-Höhle, Sächsische Schweiz; Foto:Arnhold 2012

32
die subterrane Fauna Deutschlands. – Mitt. Verb. dt. Höhlen- u.Karstforscher 58(2): 49-50
VOGEL, B., WEBER, D. & ZAENKER, S. (2012): Höhlentier des Jahres2012. – www.hoehlentier.de
WEBER, D. (2012a): Die Höhlenfauna und -flora des Höhlenkataster-gebietes Rheinland-Pfalz/Saarland, 5. Teil. – Abh. Karst- u. Höh-lenkunde 36: 1-2367
WEBER, D. (2012b): Die subterrane Fauna der Bastion Drusus, Zitadel-le, Mainz. – 29 S., Mainz
WEBER, D. (2012c): Exkursion in Ursulas Keller (Merzig-Brotdorf) am8.9.2012. – www.delattinia.de/Sektion-Hoehlenfauna.htm
Autoren: Dieter Weber, Im Wachtelschlag 33, 67454 Hassloch,[email protected]; Stefan Zaenker, Königswarter Str. 2a,36039 Fulda, [email protected];Matthias Arnhold, HFG Dresden e.V., Finkenweg 6, 01237 Dres-den, [email protected]
Dritter Workshop „Bauen im Karst“Am diesjährigen Arbeitstreffen zu ingenieurgeologischen Fra-gestellungen im Karst trafen sich am 17. und 18. November 2012sechzehn Teilnehmer aus der Schweiz und Deutschland inHayingen, Landkreis Reutlingen. Gastgeber war das ortsansässi-ge Bauunternehmen MTS, auch und im Besonderen in der Per-son Uli Nohlen, das für einen reibungslosen Ablauf, idealeRahmenbedingungen und ein angenehmes Ambiente sorgte.Themenschwerpunkt in 2012 war der Untertagebau im Karst.Marco Filipponi gelang es dazu neben Höhlenforschern auchFachleute aus dem Tunnelbau als Referenten hochkarätiger Vor-träge und Teilnehmer zu gewinnen.Der besondere Charakter des Workshops zeichnet sich dadurchaus, dass den Vortragenden kein strenges Vortragskorsett ange-legt und damit eine stetige und offene Diskussion möglich wird.Dies gelang in diesem Jahr in herausragender Weise.Der Samstag stand im Zeichen methodischer Beiträge und wur-de durch Praxisbeispiele untermalt. Es kamen Beiträge zur Karst-ALEA-Methode für die Beurteilung potenzieller Karst-gefährdungen im Tunnelbau, Modellierungsansätze zur Karst-prognose, geophysikalische Vorauserkundung im Tunnelbau unddie Möglichkeiten des Einsatzes von Laserscans in Höhlen so-wie ein Diskussionsbeitrag zur Standsicherheitsbewertung vonKarsthohlräumen zum Vortrag. Weiter wurde ein beeindrucken-des Beispiel einer Schrägschachtauffahrung in einem Karstgebirgevorgestellt. Genug Stoff, um beim gemeinsamen Abendessen denErfahrungsaustausch fortzuführen. Am Sonntagvormittag be-richteten zwei weitere Referenten über konkrete Fälle, in denendie spezifische Karstexpertise half, komplexe Fragestellungen zulösen. Ein dritter Beitrag demonstrierte am Beispiel des thürin-gischen Bleßbergtunnels und der dort angefahrenen Großhöhle,zu welchen Konsequenzen fehlender Sachverstand hinsichtlichBauwerkssicherung führen kann und welche Auswirkungen dasfür die Karsthydrologie und letztendlich auch den Schutz derHöhle hat.Mehrere der Beiträge und ausgearbeitete Methoden werden aufwww.bauen-im-karst.info publiziert.Abgerundet wurde das zweitägige Kolloquium durch eine kleineKarstwanderung zur nahegelegenen Wimsener Höhle.Der länderübergreifende Austausch von praktischen Erfahrun-gen sowie theoretischen und organisatorischen Ansätzen zwi-schen den schweizer und den deutschen Kollegen wurde von
allen Workshop-Teilnehmern als überaus fruchtbar empfundenund weckte den Wunsch, den Kreis der Teilnehmer für die ge-plante Folgeveranstaltung 2013 zu erweitern und mittelfristig eineVeranstaltungsreihe für den deutschsprachigen Raum zu etab-lieren.Sven Bauer
Von Veranstaltungen
„No Place on Earth – Kein Platz zum Leben“ – Ein Höhlen-film von Janet TobiasJede Höhle hat ein Geheimnis. Doch was HöhlenforscherChristopher Nicola 1993 auf seiner Expedition in das weit-verzweigte Höhlensystem der Ukraine entdeckt, ist eine unglaub-liche, erschütternde, aber vor allem wahre Geschichte. Sie führttief ins Herz der europäischen Vergangenheit. Unter der Erde,fern vom Nachwende-Taumel, in den Europa versunken war,stößt der Höhlenforscher auf menschliche Spuren. Stück fürStück setzt er ihre ungeheuerliche Geschichte zusammen – es istdie Geschichte von Esther Stermer und ihrer jüdischen Familie.In die Enge getrieben vom heraufziehenden Faschismus, ver-kauft Esther Stermer 1939 Haus und Hof. Doch kurz bevor dieFlucht ins rettende Ausland gelingt, bricht der Krieg aus undlässt die Stermers ausweglos zurück. Esther Stermer trifft eineunglaubliche Entscheidung: Im Oktober 1942 führt sie ihrenEhemann, ihre Töchter, Söhne und Enkelkinder unter die Erdein Höhlen. Von hier aus gibt es kein Weiterkommen. Andert-halb Jahre verbringen sie in der Finsternis am Ende der Welt –bis sie von den Nazis aufgespürt werden...Die Emmy-preisgekrönte Regisseurin Janet Tobias („Life 360“)erzählt von einem Rückzug in die Höhle, die zur letzten Rettungwird. Die ununterbrochene Dunkelheit hat die Überlebendender Familie Stermer bis zum Ende ihres Lebens begleitet. In„No Place on Earth – Kein Platz zum Leben“ spürt Janet Tobiasihrem Schrecken nach, aber auch ihrem Trost. Ihr gelingt dabeinicht nur die spannungsvolle und berührende Geschichte einerwahnsinnigen Flucht, sondern auch das Porträt einer faszinie-renden Frau mit einem unverrückbaren Lebenswillen. Im Spielmit Licht und Schatten, in Interviews und Spielfilm-Sequenzenfindet der Film Bilder für das Unvorstellbare unserer jüngstenVergangenheit und spürt den Schattenseiten ihrer Darstellungnach.Dabei sind die Szenen untertage hervorragend umgesetzt. Ge-dreht wurde in der Baradla-Höhle im Nationalpark Aggtelek undan den Orginalschauplätzen in der Westukraine. Die 7,8 km lan-ge Verteba-Höhle befindet sich in der Nähe des Ortes BilcheZolote. Nur wenige Kilometer entfernt befindet sich das Gips-höhlensystem Priestergrotte, die aufgrund ihres unterirdischen Seesauch „Ozerna“ genannt wird und mit über 127 km Länge diezweitlängste Gipshöhle der Welt ist.Dem Film ist die Mitarbeit erfahrener Höhlenforscher wie GustavStibranyi anzusehen. Der Gründer des Speläoausrüsters„Meander“ war lange Jahre Technikreferent des SlowakischenHöhlenverbandes.Fazit: Nicht nur für Höhlenforscher eine schier unglaublicheStory, bestens in Szene gesetzt und einfühlsam nacherzählt. Sollteman gesehen haben!Daten: 84 Minuten, im Verleih von Senator Film Verleih, Kino-start 9. Mai 2013.Weitere Informationen: www.senator.deBärbel Vogel
Film

33
53. Jahrestagung 2013 des VdHK in Giengen/Hürben 30.5. – 2.6.2013
Anmeldeformular Hiermit melde ich mich zur 53. Verbandstagung der deutschen Höhlen- und Karstforscher vom 30.5. – 2.6.2013 an Verein: ____________________________________________________________________ _____________________________________________________________________ Vor- und Nachname: _________________________________________________________ __________________________________________________________ Anschrift: __________________________________________________________________ ___________________________________________________________________ e-mail-Adresse : _____________________________________________________________ Anreisedatum:_______________________________________________________________ Übernachtung: Zelt:__ Massenquartier:__ Wohnmobil:___ Naturfreunde-Haus (ca. 6 km entfernt, 11.00 €/Nacht) _____ Gasthäuser, Hotels bitte selbst buchen über i-Punkt Giengen Frühstück: Donnerstag ___ Freitag___ Samstag___ Sonntag___ (3,50 €) Lunchpaket: Donnerstag___ Freitag:___ Sonntag:___ (3,00 €: 2 Brötchen, Apfel, Müsliriegel) Mittagessen: Donnerstag____ Freitag____ Samstag____ (Leberkäse mit (Gulaschsuppe mit (Maultaschen Salat, 5,00 €) mit Brötchen, 4,00 €) 4,30 €) Abendessen: Donnerstag_____ Freitag_____ Samstag_____ (Grillwürste (Erbseneintopf (Kaltes Vesper á 2,50 €) mit Würstle, 4,00 €) 4,00 €) Teilnahme an folgenden Exkursionen: _________________________________________ (bitte die E-Nummern eintragen) Interesse an folgenden Vorträgen: _____________________________________________ Bei Anmeldung bis zum 15. April 2013 beträgt die Tagungsgebühr: 25,00 € Ab 16. April 2013 beträgt die Tagungsgebühr: 30,00 € Kinder in Begleitung ihrer Eltern sind bis 14 Jahren frei. Teilnahme nur am Samstag: 5,00 € Die Anmeldung ist verbunden mit der Überweisung auf das Konto Nr. 20 94 74 025 bei der Volksbank Brenztal eV, BLZ 600 695 27, Verwendungszweck: Tagungsgebühr 2013 in Hürben Überweisungsträger bitte gut lesbar und vollständig in Druckbuchstaben ausfüllen! Die Kosten für die Essen sind nicht im Tagungsbetrag enthalten! Bei der Anmeldung bitte mit überweisen. Die Anmeldung kann per e-mail an: [email protected] oder per Post an: Ulrike Brender oder Susanne Prechtl, Höhlen- und Heimatverein e.V., Lonetalstr. 61, 89537 Giengen-Hürben
Achtung – aktuelle Programmversion auf www.vdhk.de beachten

34
ExkursionenDonnerstag 30.5.2013
E1 11:00 Archäopark VogelherdIm Mai 2013 wird der neue ArchäoPark an der Vogel-herdhöhle eröffnet. Sie zählt weltweit zu den bedeutendstenFundstellen der Steinzeitkultur und eröffnet einen tiefenBlick in unsere Urgeschichte. Der Besucher taucht selbstaktiv in die Steinzeit ein und bekommt auf faszinierendeWeise einen umfassenden Eindruck von der Epoche. DerVogelherd ist die Fundstelle des berühmten Vogelherd-pferdchen, des Mammuts sowie zahlreicher anderenPlastiken aus Mammutelfenbein. Die Funde gehören zuden ältesten menschlichen Kunstwerken der Welt.Auch mit kleineren Kindern geeignet ! Weitere Infos unterwww.archaeopark-vogelherd.deLeitung: Führung durch Mitarbeiter des ArchäoparksTeilnehmer: 25 Personen, Dauer: je nach Interesse, Abfahrt:11:00 Uhr Höhlenhaus, Fahrtdauer ca. 5 Minuten, mitPrivat-PKWs.
......Oder kleine Wanderung ca. 1 Std. dann Abmarsch 10:00 Uhr
E2 10:00 Tour Rosenstein Heinrichshöhle, Finsteres Loch,FuchslochErkundung der Heinrichshöhle (Weißer Jura, geol.interessant), des Finsteren Lochs, Fuchslochs (Deckenkolk,Belemniten, Fauna), eng, zum Teil lehmig und nass(Finsteres Loch ist trocken), nur Elektrolicht!Leitung: Höhlenkundliche Arbeitsgemeinschaft Rosen-stein/Heubach.Ausrüstung: Grundausrüstung, Schlaz erforderlich.Die Höhlen sind engräumig, lehmig und zum Teil nass,Fortbewegung: aufrecht, gebückt, schlufen, Schwierigkeit:leicht-mittelschwer. Dauer: ca. 5 Std.Teilnehmer: 12 Personen.Abfahrt: 10:00 Uhr am Höhlenhaus, Fahrtdauer ca. 1 Std.mit Privat-PKWs.
E3 10:00 Tour Scheuelberg, Saal-am-anderen-Höhle, Jakobs-höhleBefahren der Saal-am-anderen-Höhle (reich an Sinter-schmuck) und Jakobshöhle.Achtung: beide Eingänge sind sehr eng! Die Höhlen sindeher trocken. Saal-am-anderen-Höhle: Nach dem Einstieggeht es sehr steil auf lehmigen Boden ca. 8 m abwärts. EinSeil mit Handschlaufen ist eingebaut.Leitung: Höhlenkundliche Arbeitsgemeinschaft Rosen-stein/Heubach.Ausrüstung: Grundausrüstung, Fortbewegung: aufrecht,Schwierigkeit: leicht.Dauer: ca. 5 ½ Std. 2 Std. Wanderung zu den Höhlen, 31/2 Std. Befahrung, Teilnehmer: 14 Personen.Abfahrt: 10:00 Uhr am Höhlenhaus, Fahrtdauer ca. 1 Std.mit Privat-PKWs.
E4 10:00 Wanderexkursion zum Rosenstein (für Begleitpersonen,Familien, Kinder)Gemütliche Wanderung mit Besichtigung von 3 Höhlen(Finsteres Loch: ca. 140 m, Scherbenfunde; Dreieingangs-höhle: ca. 96 m, Knochenfunde; Kleine Scheuer: 26 m,steinzeitliche Höhle, Feuersteinklingen (Mikroliten),Backenzähne vom Höhlenbär. Alle Höhlen sind trockenund leicht begehbar. Viel Wissenswertes über die Besied-lung der Höhlen in der Steinzeit und im Mittelalter wird
vermittelt. Der Rosenstein ist ein herrliches Wandergebietmit toller Aussicht, einer Ruine sowie Grill- und Spielplatz.Auch für kleinere Kinder geeignet, allerdings nichtkinderwagentauglich !Ausrüstung: Taschenlampe.Dauer: ca. 3 Std. Einkehrmöglichkeit, dann längere DauerLeitung: Höhlenkundliche Arbeitsgemeinschaft Rosen-stein/Heubach.Teilnehmer: 20 Personen, Abfahrt: 10:00 Uhr am Höhlen-haus, Fahrtdauer ca. 1 Std. mit Privat-PKWs.
ACHTUNG ! Tour E2 , E3, E4 fahren gemeinsam !
E5 11:00 Laierhöhle Tour BVon der Charakteristik her ist die Laierhöhle ein dreidimen-sionales, abwechslungsreiches Labyrinth mit vielenAbseilstrecken. Immer wieder münden erstaunlich engeVerbindungen in große Hallen mit weiteren Fortsetzungen.Diese Tour führ in die Obere Etage: Amphitheater, Lehm-gang, Verbruchhalle und Pyramidenhalle.Trittsicherheit erforderlich. Keine SRT nötig. Einstiegs-schacht und Durchstieg Schleuse sind mit Steighilfenausgebaut.Leitung: Kahlensteiner Höhlenverein.Ausrüstung: Grundausrüstung, Fortbewegung: aufrecht,gebückt, schlufen, Schwierigkeit: leicht, Dauer: ca. 2,5 Std.Abfahrt: 11:00 Uhr am Höhlenhaus Fahrtdauer ca. 1 Std.mit Privat-PKWs. Treffpunkt Parkplatz Laierhöhle DerParkplatz bei der Laierhöhle befindet sich beim Wasser-hochbehälter direkt am östlichen Ortsausgang von Weilerin Richtung Schalksfetten.Teilnehmer: 8 Personen
E6 10:00 Laierhöhle Tour ADiese Tour führt in den „Wilden Westen“ und „Hindu-kusch“. Trittsicherheit unbedingt erforderlich. Die Abstiegesind mit Steighilfen ausgebaut. Keine SRT-Ausrüstungnötig. Auf dem Rückweg wird in der oberen Etage nochdas Amphitheater besucht.Leitung: Kahlensteiner Höhlenverein.Ausrüstung: Grundausrüstung, Fortbewegung: aufrecht,gebückt, schlufen, abseilen, Schwierigkeit: mittel, Dauer:ca. 4 Std. Abfahrt: 10:00 Uhr am Höhlenhaus Fahrtdauerca. 1 Std. mit Privat-PKWs. Treffpunkt ParkplatzLaierhöhle. Teilnehmer: 8 Personen.
E7 8:30 BärentalhöhleDie Höhle ist eine typische Albhöhle – eng und lehmig.Sie hat eine vermessene Länge von 425 m, größtenteilshorizontal, mehrere Engstellen und Schlufe, kurze Kletter-stelle, die aber ohne Sicherung begangen werden können.In der Höhle wird aktiv gegraben und es ist ein Telemetrie-system eingebaut. Genauere Infos und Plan aufwww.hv.blaubeuren.deLeitung: Höhlenverein Blaubeuren.Ausrüstung: Grundausrüstung, Fortbewegung: meistgebückt, schlufen, Schwierigkeit: leicht-mittelschwer,Dauer: ca. 4 Std. Abfahrt: 8:30 Uhr Höhlenhaus Fahrtdauerca. 1,5 Std. mit Privat-PKWs. Teilnehmer: 6 Personen.
E8 18:00 Sinnesführung durch die CharlottenhöhleAuch Schauhöhlen müssen ihr Angebot erweitern, sobieten wir seit Jahren „Sinnesführungen“ an. Das Höhlen-licht bleibt aus, mit einer Taschenlampe bewaffnet machtman sich auf den Weg um die Höhle im Dunklen zuerkunden. Bei dieser Tour werden auch unsere Sinne getes-tet. Das Konzept ist auf Familien und Kinder ausgerichtet,aber natürlich sind alle Interessierte willkommen.Leitung: Höhlen- und Heimatverein Hürben.
Programm der 53. VdHK-Tagung 2013in Giengen-Hürben
30.5. – 2.6.2013

35
Dauer: ca. 1,5 Std., Beginn: 18:00 Uhr am Eingang derCharlottenhöhle, Ausrüstung: festes Schuhwerk, Jacke,Taschenlampe. Teilnehmer: 20 Personen pro Gruppe.
E21 15:00 Korallen und Klifflinie in GerstettenGerstetten, bekannt für die zahlreichen Korallenfunde ausdem Jurameer und der Klifflinie des Molassemeeres, bietetfür geologisch Interessierte anschauliche Erkenntnisse überdiese lang vergessenen Zeiten. Ein erfahrener Geologe wirduns zu diesen Besonderheiten führen.Leitung: Dr. GregorBeginn: 15:00 Uhr, Dauer: ca. 2 Std., Teilnehmer: 40Personen, Abfahrt am Höhlenhaus: 14:30 Uhr, Fahrtdauerca. 15 Minuten, mit Privat-PKWs.
Freitag 31.5.2013
E2/2 10:00 Tour RosensteinSiehe E2 am Donnerstag
E3/2 10:00 Tour ScheuelbergSiehe E3 am Donnerstag
E4/2 10:00 Wanderexkursion zum RosensteinSiehe E4 am Donnerstag
E9 9:30 Steinzeitkunst im LonetalDie Höhlen im Lonetal gehören europaweit zu denwichtigsten Fundstellen für die mittlere und JüngereAltseinzeit. Hier haben sich in der letzten Eiszeit immerwieder Neandertaler, seit etwa 40.000 Jahren dann frühemoderne Menschen aufgehalten. Letztere hinterließen unsaus Mammutelfenbein geschnitzte Figuren, wie dasVogelherdpferdchen oder den Löwenmenschen. Diesegehören zur ältesten figürlichen Kunst weltweit. Auf einerkleinen Wanderung werden die Fundstellen aufgesucht.Auch mit kleineren Kindern geeignet. Bis auf die Bock-steinhöhle auch kinderwagentauglich !Leitung: Hannes Wiedmann (Urzeitmuseum Blaubeuren)Dauer: ca. 4-5 Std. Abfahrt: 9:30 Uhr am Höhlenhaus,Fahrtdauer ca. 10 Minuten mit Privat-PKWs.Teilnehmer: 30 Personen.
E10 9:30 Besichtigung des Egau-WasserwerksDie Ausläufer der Ostalb sind geprägt durch ein qualitativgutes und hohes Wasseraufkommen. Die mittlere Neckar-region und verschiedene Albflächendörfer werden durchdieses Wasser versorgt. Der „Buchbrunnen“ ist eine dergrößten für die öffentliche Trinkwasserversorgung genutz-ten Quellen des Landes.Leitung: Mitarbeiter des Egau-WasserwerksDauer: Führung ca. 1,5 Std., Teilnehmer: 40 Personen,Abfahrt: 9:30 Uhr Höhlenhaus, Fahrtdauer ca. 30 Minuten,mit Privat-PKWs.
E11 8:30 Meteorkrater-Museum, SteinheimVon allen Meteorkratern auf der Welt hat wohl das Stein-heimer Becken die interessanteste Geschichte. Denn imUnterschied zu größer geformten Kratern besitzt esherausragende Merkmale:- Der am besten erhaltene Krater mit Zentralhügel- Die erste Beschreibung von „Strahlenkalken“- Die Seebildung, Verlandung und Freispülung der Füllung- Die Fundstellen von Fossilien.Eine interessante und landschaftlich schöne Wanderungauf den Zentralberg des Meteorkraters; danach Besuch desMuseums, alles mit Führung. Auch mit kleineren Kinderngeeignet.Leitung: G. Müller vom Museum
Dauer: 4-5 Std., Teilnehmer: 25 Personen, Abfahrt: 8:30Uhr Höhlenhaus, Fahrtdauer ca. 45 Minuten, mit Privat-PKWs.
E12 12:30 Mergelstetter WasserwegDie Ausläufer der Ostalb sind geprägt durch ein qualitativesund hohes Wasseraufkommen. Auf dem MergelstetterWasserweg wird Frau Marquard-Mader verschiedene Quel-len der Karstlandschaft erwandern. Gemütliche Wanderungvorbei an zahlreichen Karstquellen, unter fachkundlicherLeitung. Auch mit Kindern geeignet. Länge: ca. 8 km.Leitung: Frau Marquard-MaderDauer: 4 Std., Teilnehmer: 25 Personen, Abfahrt: 12:30Uhr Höhlenhaus, Fahrtdauer ca. 10 Minuten, mit Privat-PKWs.
E13 12:30 Tiefer Stollen, WasseralfingenDer Tiefe Stollen ist ein Besucherbergwerk, in dem bis1930 Eisenerz abgebaut wurde. Er wird seit 1989 auch fürdie Behandlung von Patienten mit Atemwegserkrankungengenutzt. Die Heilstollentherapie ist besonders gut geeignetfür Patienten mit Asthma bronchiale, chronischer Bronchi-tis, allergischen Nebenhöhlenerkrankungen, Heuschnupfenund Neurodermitis. Besichtigung der Therapieräume mitVortrag.Leitung: Führung durch Mitarbeiter des Tiefen StollensDauer: ca. 2 Std., Teilnehmer: 30 Personen, Abfahrt: 12:30Uhr Höhlenhaus, Fahrtdauer ca.1 Std., mit Privat-PKWs.
E14 10:00 Hessenloch bei KönigsbronnDas Hessenloch ist eine Horizontalhöhle mit einer leichtenKletterpassage im Eingangsbereich. Während dieser Exkur-sion werden auch der Brenztopf und die Pfeffer-Quellebesucht.Leitung: Herbert Jantschke, Höhlen- und Interessen-gemeinschaft Ostalb-KirchheimAusrüstung: Grundausrüstung, Fortbewegung: gebückt,schlufen, Schwierigkeit: leicht-mittel, Teilnehmer: 8 Perso-nen, Dauer: ca. 3-4 Std., Abfahrt: 10:00 Uhr Höhlenhaus,Fahrtdauer 30 Minuten mit Privat-PKWs.
E15 8:30 Mordloch bei EybachDas Mordloch ist eine aktive Wasserhöhle mit einem 10 m-Siphon im Eingangsbereich. Daher sollten die TeilnehmerTaucherfahrungen mit kleineren Siphons und Wasser-höhlen vorweisen. Für die Tauchausrüstung, wie z.B.Tauchflaschen etc., ist jeder Teilnehmer selbst verant-wortlich und muss diese zur Verfügung stellen.Leitung: Werner Gieswein, Höhlen- und Interessenge-meinschaft Ostalb-KirchheimGrundausrüstung + Tauchausrüstung, Schwierigkeit:schwer mit Tauchstecken, Dauer: ganztägig, Teilnehmer:5 Personen, Abfahrt: 8:30 Uhr Höhlenhaus, Fahrtdauerca. 1 Std. mit Privat-PKWs.
E16 8:30 Hessenhauhöhle bei BerghülenDie Hessenhauhöhle ist die tiefste Höhle der SchwäbischenAlb (144 m) und führt nach einem 135 m tiefen Abstieg(viele Schachtengstellen) in einen aktiven Flusslauf.ACHTUNG ! Nur für erfahrene Höhlenforscher, sehrsportiv, gute SRT-Technik erforderlich !!!Leitung: Jürgen Bohnert HöhleninteressengemeinschaftOstalb-Kichheim, Arge BlaukarstGrundausrüstung + SRT, Teilnehmer: 5 Personen, Dauer:ganztägig, Abfahrt: 8:30 Uhr Höhlenhaus, Fahrtdauer ca.1 Std. mit Privat-PKWs.
E17 13:00 Besichtigung Landeswasserversorgung LangenauDie Ausläufer der Ostalb sind geprägt durch ein qualitativ

36
gutes und hohes Wasseraufkommen. Die mittlereNeckarregion und verschiedene Albflächendörfer werdendurch dieses Wasser versorgt. Langenau ist eine der quellen-reichsten Städte in Deutschland. Hier ist die Landes-wasserversorgung, eine der größten Wassergewinnungs-anlagen in Europa, angesiedelt. Dort kann auch diehervorragend konzipierte Ausstellung „Erlebnis Grund-wasser“ besichtigt werden.Dauer: 2 Std., Teilnehmer: 40 Personen, Abfahrt: 13:00Uhr Höhlenhaus, Fahrzeit ca. 20 Minuten, mit Privat-PKWs.
E18 18:00 Historische Führung in der CharlottenhöhleDie Charlottenhöhle wurde vor 120 Jahren entdeckt. ZurEröffnung der Schauhöhle kam die Königin Charlotte vonWürttemberg als Patin. Der damalige Entdecker, Oberförs-ter Sihler wird Königin Charlotte durch die Höhle führen.Leitung: Höhlen- und Heimatverein HürbenDauer: ca. 1,5 Std. Teilnehmer: 25 Personen, Treffpunkt:18:00 Uhr am Eingang der Charlottenhöhle.
Samstag, 1. Juni 2013
E1/2 14:00 ArchäoparkSiehe E1 Donnerstag, Abfahrt: 14:00 Uhr Höhlenhaus
E19 14:00 Bolheimer Geologenpfad mit Lindacher HöhleDie besondere Lage am Albrand, der Klifflinie desMolassemeers und die Befahrung der Lindacher Höhle wirdmit Herrn Ziemann ein besonderes Erlebnis sein.Leitung: Herr ZiemannAusrüstung: Grundausrüstung, Schwierigkeit: leicht,Dauer: ca. 3,5 Std., Teilnehmer: 25 Personen, Abfahrt: 14:00Uhr Höhlenhaus, Fahrtdauer ca. 10 Minuten, mit Privat-PKWs.
E22 17:00 Geschichten aus dem Leben eines HöhlenführersBei der Arbeit als Höhlenführer erlebt man so manchSkurriles und Witziges. Nach jahrelangen Erfahrungen indieser Tätigkeit gehen einem die Geschichten nie aus. EineHöhlenführung durch die Charlottenhöhle der eigenen Art.Führer und Referent: Eckhard Baschin, Höhlen- undHeimatverein HürbenDauer: ca. 1 Std., Teilnehmer: 25 Personen, Beginn: 17:00Uhr am Eingang der Charlottenhöhle.
Sonntag 2. Juni 2013
E7/2 10:00 BärentalhöhleSiehe E7 am Donnerstag, Abfahrt: 10:00 Uhr Höhlenhaus,Fahrdauer ca. 1,5 Std. mit Privat-PKWs.
E19/2 10:00 Bolheimer Geologenpfad mit Lindacher HöhleSiehe E19 am Samstag, Abfahrt 10:00 Uhr Höhlenhaus.
E1/3 11:00 ArchäoparkSiehe E1 am Donnerstag.
E20 10:00 Himmelsfelsenschacht bei EybachDer Himmelsfelsenschacht ist eine Schachthöhle mit2 Schächten (25 m, 15 m)Leitung: Michael KlöbleAusrüstung: Grundausrüstung + SRT-Ausrüstung,Schwierigkeit: mittelschwer, SRT-Erfahrung erforderlich.Teilnehmer: 8 Personen, Abfahrt: 10:00 Uhr Höhlenhaus,Fahrtdauer ca. 45 Minuten, mit Privat-PKWs.
Während der Tagung ist der Besuch der Schauhöhle „Charlottenhöhle“und des dazugehörige interaktiven Museums „Faszination Höhle, Mensch,
Natur“ für alle Teilnehmer, nach Vorzeigen des Tagungsausweises, kosten-los. Die Charlottenhöhle kann nur im Rahmen einer Führung besichtigtwerden!! Da am Donnerstag bei uns Feiertag ist, raten wir ab, an diesemTag die Charlottenhöhle zu besichtigen, da voraussichtlich an diesemTag die Höhle stark von Besuchern frequentiert ist.
Teilnahme an allen Höhentourennur mit Elektro-Licht möglich!
Keine Haftung des Veranstalters – Teilnahme auf eigene GefahrVersicherung ist Sache der Teilnehmer.
Bei allen Exkursionen ist die Teilnehmerzahl begrenzt!Die Einteilung erfolgt nach Eingangsdatum der Anmeldung!
Änderungen und Ergänzungen unter www.vdhk.de
VorträgeDonnerstag 30.5.2013
V1 10:00 Besiedlung im Hohle FelsVor 40.000 Jahren wurde die „Venus vom Hohle Fels“geschnitzt. Im September 2008 wurde sie auf derSchwäbischen Alb entdeckt – seither schlagen die Speku-lationen über ihre Entstehung und ihre Bedeutung hoheWellen. Die schwäbische Venus stammt aus der Zeit, alsdie ersten Menschen vom Schwarzen Meer entlang derDonau durch Europa zogen, vom heutigen Rumänien bisin die Chauvet-Höhle in Frankreich. Damals schien es denMenschen so gut zu gehen, dass sie Zeit hatten für dieKunst: Sie schnitzten Plastiken, erfanden Musikinstrumenteund bemalten Höhlenwände.Referent: Herr Frey, Hohle FelsDauer: ca. 1 Std., Beginn: 10:00 Uhr Bärensaal.
V3 15:00 GeoPark Schwäbische AlbDie Schwäbische Alb wurde aufgrund ihres bedeutendengeologischen Erbes im Jahr 2002 zum Nationalen GeoParkernannt. Der Begriff „Geopark“ ist ein Prädikat für Ge-biete, die über ein besonders reichhaltiges geologischesErbe verfügen. Neben dem Schutz dieses Erbes und derVermittlung geologischer Themen im Rahmen der Umwelt-bildung gehört die nachhaltige Regionalentwicklung ebensowie die wissenschaftliche Forschung innerhalb des Parkszu den Aufgaben eines Geoparks.Referent: Ulrike Brender-Nothnick.Dauer: ca. 40 min, Beginn: 15:00 Uhr GeoPark InfoStelleim Höhlenhaus.
V11 16:00 Opas Speere – Erschuf der „Wurf“ den Menschen?Die Evolution des Menschen zum Jäger – und zumSportler. Schimpansen ernähren sich im Allgemeinen durchSammeln – gelegentlich aber auch durch die Jagd auf kleineTiere, die sie „handgreiflich“ erledigen können. Ihr Jagenunterscheidet sich aber fundamental von der Jagd heutigerVölker, die noch Jäger und Sammler sind oder bis vorkurzem waren. Diese jagen bevorzugt Großwild, wenn essolches gibt. Schimpansen nie!Wie vollzog sich der Wandel vom letzten gemeinsamenVorfahren, der sich so wie die heutigen Schimpansenernährte, zum menschlichen Jäger? Verdanken wir diesemWandel, dass wir heute Fußball und Tennis spielen oderkegeln – bzw. begeistert dabei zuschauen?Danach: Jagen wie die Eiszeitmenschen mit Schleuder-speeren auf dem Freigelände.Referent: Hermann HuberDauer: ca. 1 Std., auf dem Freigelände 2-3 Std., Beginn:16:00 Uhr Bärensaal.
V12 17:00 Telemetrie in HöhlenJede ernsthafte Erforschung von Höhlen ist auf grund-legende Daten wie Wasser- und Lufttemperatur, Wasser-

37
stand, Windgeschwindigkeit, CO2-Gehalt etc. angewiesen.Einfache Messungen während einer Befahrung ergebenkeinerlei Trends. Deshalb werden Daten-Logger immerbeliebter. Da die Daten aber nur im Speicher abgelegt undausgelesen werden müssen, sind aktuelle Daten kaumverfügbar. Das vorgestellte Telemetriesystem liefert Datenaus der Höhle durch drahtlose Übertragung in Echtzeit andie Oberfläche, die aktuell im Internet verfolgt werdenkönnen und zu weiteren Auswertung auf einem Serverzur Verfügung stehen. Im Vortrag werden die physikali-schen Hintergründe der drahtlosen Datenübertragungdurch Erdreich und die praktische Realisation dargestellt.Referent: Knut Brenndörfer, Höhlenverein BlaubeurenDauer: ca. 1 Std., Beginn: 17:00 Uhr Bärensaal.
V2 18:00 Wimsener HöhleDer Besuch der Wimsener Höhle am südlichen Rand derSchwäbischen Alb, etwa drei Kilometer nordwestlich vonZwiefalten, ist ein einmaliges Erlebnis, denn es ist dieeinzige mit dem Boot befahrbare Schauhöhle in Deutsch-land. Hier strömt die unterirdische Ach aus dem markantenHöhlenportal hervor. Geheimnisvoll schimmert das Tages-licht ins Halbdunkel der Höhle hinein und zaubert Reflexeauf dem Wasserspiegel des Höhlensees und am Felsgewöl-be. Im Jahr 2007 gelang die sensationelle Entdeckung einesstark versinterten Skeletts hinter der ersten Tauchstrecke.Die neusten Forschungsergebnisse von der WimsenerHöhle werden vorgestellt.Referent: Rainer StraubDauer: ca. 45 Minuten, Beginn: 18.00 Uhr Bärensaal.
V15 19:30 Gerstetter Urmeere und KorallenGerstetten, bekannt für die zahlreichen Korallenfunde ausdem Jurameer und der Klifflinie des Molassemeeres bietetfür geologisch Interessierte anschauliche Erkenntnisse überdiese lang vergessenen Zeiten.Referent: Dr. Gregor, Gerstetter KorallenmuseumDauer: ca. 1 Std., Beginn: 19:30 Uhr.
Freitag 31.5.2013
V14 18:00 Hausener BröllerReferent: Karsten Gessert (HFGOK)Beginn: 18:00 Uhr Bärensaal.
V4 19:00 Humorvolle Unterhaltung über nützliche und nutz-lose HöhlenwerkzeugeEin nicht ganz so ernstzunehmender Vortag über Höh-lenwerkzeuge und ihre Verwendung.Referent: Eckhardt Baschin, Höhlen- und HeimatvereinHürbenDauer: ca. 45 Minuten, Beginn: 19:00 Uhr Bärensaal.
Samstag, 1.6.2013
V10 14:00 Die Höhlenfauna LuxemburgsErgebnisse eines fünfjährigen Projekts. Im Rahmen derPräsentation wird auch das Buch vorgestellt.Referent: Dieter Weber, wissenschaftl. Mitarbeiter amMusée national d‘histoire naturelle du LuxemburgDauer: Beginn: 14:00 Uhr, Mammutsaal.
V5 14:00 Luchs-Land Schwäbische Ostalb – HöhlenfundezuhaufVon der östlichen Schwäbischen Alb kennt man wegenihres Reichtums an Höhlen und aufgrund deren gründlicherErforschung mehr fossile und subfossile Nachweise desLuchses (Lynx lynx) als aus den anderen Teilen Baden-Württembergs. Eine kritische Bestandsaufnahme zeigt, dass
diese faszinierende Katze während klimatisch unterschied-lichster Abschnitte des jüngeren Eiszeitalters und dergeologischen Gegenwart hier geeignete Lebensbedingenvorgefunden hat.Referent: Thomas Rathgeber (Staatliches Museum fürNaturkunde Stuttgart)Dauer: ca. 1 Std., Beginn: 14:00 Uhr Bärensaal.
V13 15:00 Karst und Höhlen in HeubachEin besonders reichhaltiges und vielfältiges Angebot anKarsterscheinungen bietet die Schwäbische Alb bei Heu-bach. Rund um die Rauhe Wiese – die zweitgrößteKarstwanne der Schwäbischen Alb – finden sich anRosenstein und Scheuelberg zahlreiche Höhlen, die bereitsseit der Steinzeit von Mensch und Bär genutzt wurden.Zwischen diesen sagenbehafteten, altbekannten Höhlenfinden sich auch spektakuläre neue Entdeckungen wie derjüngst erforschte Teufelsklingenbröller.Referenten: Gerhard Nowak (Arge Rosenstein), KarstenGessert (HFGOK) und Hannes Köble (KHV)Beginn: 15:00 Uhr Bärensaal.
V8 15:00 Das Blauhöhlensystem bei BlaubeurenReferent: Andreas KüchaDauer: ca. 1 Std., Beginn: 15:00 Uhr Hürbetal-Halle.
V9 16:00 Zur aktuellen Situation der Schauhöhlen Deutsch-landsReferent Michael BrustDauer: ca. 1 Std., Beginn: 16:00 Uhr Bärensaal.
V7 16:00 Die HessenhauhöhleKnapp zwei Jahre erst sind seit der Entdeckung der„Nordblau“ am 26.3.2011 vergangen. Doch die Mitgliederder Arbeitsgemeinschaft Blaukarst konnten in diesemZeitraum der Hessenhauhöhle so manches Geheimnisentreißen und sie auf fast 4 km Ganglänge und 144 mTiefe vermessen. Ein großer unterirdischer Flusstunnel vonbis zu 20 m Breite windet sich durch die unterirdische Alb,Tropfsteine von unglaublicher Schönheit säumen den Weg.Dabei steht die Erforschung der Höhle erst ganz amAnfang. Bis zum bekannten Ende der Blautopfhöhlebeträgt die Entfernung noch 1,5 km Luftlinie, bis nachLaichingen sogar noch fast 7 km! Dass das im Untergrundversickernde Wasser von Laichingen aus den Weg über dieHessenhauhöhle bis zum Blautopf nimmt, konnte erstkürzlich durch Markierungsversuche im Rahmen einesgemeinschaftlichen Forschungsprojektes von Hydrogeo-logen des Karlsruher Institutes für Technologie sowie denForschern der Arbeitsgemeinschaft Blautopf und derArbeitsgemeinschaft Blaukarst gesichert werden.Referent: Jürgen BohnertDauer: ca. 1 Std., Beginn: 16:00 Uhr, Hürbetal-Halle.
Änderungen und Ergänzungen unter www.vdhk.de
Allgemeine InformationenAnreiseMit dem PKW: A7: Ausfahrt Giengen/Herbrechtingen, dannder Beschilderung „Charlottenhöhle“ folgen.Tagungsbüro befindet sich in der Hürbetal-Halle, ca. 200 m nachdem Ortseingang auf der linken Seite.Mit der Bahn: der Giengener Bahnhof liegt ca. 5 km von Hürbenentfernt – an Wochentagen fahren Busse.TagungsräumeDie Jahrestagung findet in der Hürbetal-Halle und in den Räu-men des Höhlen- und Heimatvereins in Hürben statt.

38
Privatzimmer: Pensionen, Ferienwohnungen oder Hotelüber-nachtungen können über die Touristen-Information erfragt wer-den:Tourist-InformationMarktstraße 9, 9537 Giengen an der Brenz, Tel. 07322/952-2920,[email protected], www.baerenland.deParkplätzeParkplätze gibt es im Bereich der Hürbetal-Halle. Campingwagenund Wohnmobile können auf dem Wohnmobil-Stellplatz an derHöhlenErlebnisWelt abgestellt werden. Bitte um baldige Anmel-dung, da der Donnerstag (Fronleichnam) in Baden-Württembergein Feiertag ist. Strommarken für 2,00 €/12 Std. am Höhlen-haus erhältlich.Verkaufsstände, Ausstellungen, PosterFür Verkaufsstände, sowie Ausstellungen und Poster steht aus-reichend Fläche zur Verfügung. Verkaufsstände sind in einemabschließbaren Raum untergebracht. Bei der Anmeldung bittedie gewünschte Anzahl der Tische/Flächen angeben.VerpflegungIm Höhlenhaus gibt es Getränke und Speisen. Die angemelde-ten Essen werden vorrangig bedient. Wir rechnen während dergesamten Zeit mit sehr vielen Gästen, da die HöhlenErlebnisWeltstark frequentiert wird.ÜbernachtungNur angemeldete Personen können eine Gewähr für die ange-botenen Übernachtungsmöglichkeiten auf dem Gelände haben.TagungsheftEs wird ein Tagungsband erscheinen. Termin zum Einreichender Vortragszusammenfassungen und Kurzberichte ist der15. April 2013.ExkursionenFür die Exkursionen wird eine Absicherung über einen Rettungs-fond empfohlen.
Dr. Klaus Eberhard Bleich 80Am 5. Januar 2013 konnte Dr. Klaus Eberhard Bleich seinen 80.Geburtstag feiern. Er ist der Senior der ehemaligen Vorsitzen-den unseres Verbandes – seine Amtszeit umfasste 7 Jahre, von1965 bis 1972. In dieser fand 1969 der 5. Internationale Kon-gress für Speläologie in Stuttgart statt.Die Verdienste von Dr. Klaus Eberhard Bleich um den Verbandsind vielgestaltig und seine wissenschaftlichen Publikationen er-fassen alle Bereiche der Karst- und Höhlenforschung. Im Heft2/1993 der Verbandsmitteilungen sind sie ausführlich gewür-digt. Wir gratulieren und wünschen mit einem herzlichen „GlückAuf“ unserem ehemaligen Vorsitzenden noch viele weitere Jah-re.Prof. Dr. Dr. h.c. Karl-Heinz Pfeffer, Tübingen
Personalia
Interessantes für Höhlenfotografen: Funkkopplerstatt OptokopplerWer in Höhlen fotografiert, kennt das leidige Problem mit denZweitblitzen. Ganz früher wurde mit Stativ und Drahtauslösergearbeitet, in neuerer Zeit dann mit Optokopplern, die auf denHauptblitz reagieren. Vor einigen Jahren kamen nun Funkkopplerauf den Markt, die eine Reihe von Vorteilen bieten.Über das Internet bin ich auf den Yongnuo RF-603-Blitzauslösergestoßen und habe vier Stück gekauft. Sie arbeiten im 2,4 GHz-Band und lösen einige immer wieder ärgerliche Probleme. Zumeinen werden die eigenen Blitze „gern“ von anderen Fotografennebenbei ausgelöst, und zum anderen sorgt die moderne LED-Beleuchtung mit ihrer Pulsweitenmodulation (unsichtbares hoch-frequentes Flackern) dafür, dass auch die besten Optokoppler(z.B. der Firefly) ungewollt auslösen.Mit den Yongnuo RF-603 hat sich das erledigt. Sie haben eineReichweite von ca. 100 m und 16 Funkkanäle. Eine LED zeigtan, ob sich der Zweitblitz in Funkreichweite befindet – sehr prak-tisch. Bei Betrieb mit zwei Stück Micro AAA-Batterien sind 10bis 15 Stunden Betrieb möglich (Stromverbrauch etwa 25 mA).Die Verbindung zum Blitz und Fotoapparat läuft über zwei Blitz-schuhe (oben und unten) oder passendes Kabel. Die Synchroni-sierungszeit liegt mit 1/200 s in sehr akzeptablen Größenord-nungen.
Bisher gab es nur das Problem, dass man dafür eine teuere Nikonoder Canon mit Zusatzkontakten am Blitzschuh besitzen muss.Mit einfacheren Kameras, die nur einen Mittenkontakt haben,funktionierten sie nicht.Die Yongnuo RF-603 in der Nikon-Ausführung lassen sich aberproblemlos so umbauen, dass sie mit „normalen“ Kameras funk-tionieren, die nur einen Mittenkontaktauslöser haben. Bei derCanon-Ausführung geht das nicht – die Kontakte liegen anders!Das Funktionsprinzip bei der Nikon D700 ist folgendes: BeideYongnuo RF-603 sind standardmäßig als Empfänger aktiviert.Durch Betätigen der Fokustaste am Fotoapparat werden 3 Vvom Fotoapparat auf den Pin SP (siehe Foto) gelegt und derYongnuo RF-603 schaltet auf Sendebetrieb um. Dadurch wirdder Funkkoppler am Blitz aktiviert. Nun erfolgt die Auslösungüber den Mittenkontakt und der Zweitblitz zündet.Um den Yongnuo RF-603 für alle Kameras umzubauen, die nureinen Mittenkontakt haben, muss lediglich der gelbe Draht vomSP-Pin abgelötet und mit einem hochohmigen Widerstand (ca.100 k oder mehr) an den Pluspol der Batterieklemme im Yongnuoangelötet werden. Beim Testen ging es auch ohne Widerstand,ich würde ihn aber trotzdem empfehlen. Durch das Umlötenwird der Yongnuo mit dem Einschalten als Sender aktiviert. Erkann dann nur noch am Fotoapparat als Sender verwendet wer-den, also bitte entsprechend kennzeichnen!Bestellen kann man den Yongnuo RF-603 unter www.8df.de/fotozubehoer/komplett-set/rf-603-set/prod_69.html. Die Lie-ferzeit beträgt etwa 4 bis 6 Wochen.Dieter Weiß, Eckstraße 6, 98528 Suhl, [email protected]
Höhlenfotografie

39
Mitteilungendes Verbandes der deutschen Höhlen- und Karstforscher e.V
ISSN 0505-2211, München
Herausgeber: Verband der deutschen Höhlen- und Karstforscher e.V. MünchenSchriftleitung: Michael Krause, Jägerhalde 118, 70327 Stuttgart
Dr. Friedhart Knolle, Grummetwiese 16, 38640 GoslarDruck u. Verlag: Oberharzer Druckerei Fischer & Thielbar GmbH, Alte Fuhrherrenstr. 5, 38678 Clausthal-Zellerfeld
InhaltsverzeichnisJahrgang 58, 2012
OriginalbeiträgeANJA ADLER & DOROTHEE MERTMANN:Fossilien in den Höhlenlehmen der Barbarossahöhle,Kyffhäuser ................................................................................... I/6ECKHARD HINDERER & ANDREAS KÜCHA:Forschungsaktivitäten der Arge Blautopf in derBlauhöhle 2011/2012 ............................................................. III/79ANNE IPSEN:Zwischenbericht zur Renaturierung der ehemaligenTeichfläche in der Segeberger Höhle 2011 .............................. I/4FRIEDHART KNOLLE:Julius Riemer und die Höhlenforschung .......................... IV/110FRIEDHART KNOLLE & CORNELIA GROTE-BICHOEL:Professor Dieter Meischner in memoriam ........................ IV/126STEPHAN MARKS:Phantomkarst im Muschelkalk des Gertrudenbergsvon Osnabrück, Niedersachsen ............................................ III/66JOHANNES MATTES:Von Industriearbeitern, Soldaten und Höhlentouristen –Forschungsgeschichte und Beschreibung der Gassel-Tropf-steinhöhle bei Ebensee (Oberösterreich) ............................. II/40FRITZ REINBOTH:Der Gipskarst des Rösebergs bei Walkenried im Südharz –was ist noch zu retten? .......................................................... III/74FRITZ REINBOTH:Naturschutz nach Forstmeisterart im Karst – das Natur-schutzgebiet Priorteich-Sachsenstein bei Walkenried,Südharzvorland, Niedersachsen .......................................... IV/122MICHAEL SCHOPPER:Minerale und Speläotheme im Blauhöhlensystem(Kat.-Nr. 7524/30, Schwäbische Alb) ............................... IV/102DIRK STEINKE, STEFAN ZAENKER, BÄRBEL VOGEL &ALEXANDER M. WEIGAND:Von kryptischen Arten und Lebensräumen – DNA-Barcoding trifft auf die subterrane Fauna Deutschlands .. II/49
KurzberichteVdHK-Verbandsbibliothek ...................................................... I/22Zugang zur Bleßberghöhle – Erklärung zum Sachstand .. III/73Lahn-Dill-Kreis, Hessen: Verleihung des Naturschutzpreisesan Ingo Dorsten ...................................................................... III/78Neue Software zur Artbestimmung von Fledermäusen ... III/84NSG „Alter Stolberg“, Wettbewerb, Naturwunder 2012,Neudatierung von Höhlenmalereien.................................. IV/128
RubrikenTätigkeitsberichteISAAK 2011, HRVD-Treffen 2011 ........................................ I/17Jahresbericht AK Biospeläologie, Referat Biospeläologie ... I/19Arge Blaukarst, AK Kluterthöhle ......................................... II/52
Aktionstag Karls- und Bärenhöhle ....................................... II/56Nationale HRVD-Höhlenrettungsübung 2011 .................. III/84Technik: Lampenumbau Petzl-Duo zu LED .................... III/86FSEEuroSpeleo Protection, EuroSpeleo Magazine ..................... I/21Speleo-TV ................................................................................. II/62EuroSpeleo Projects ............................................................... III/83Kongress „Millau 2013“ ...................................................... IV/129DNRHandbuch Verbandsbeteiligung ............................................ III/83UISNeue Website ........................................................................... III/83Forschung AktuellFrankreich, Mexiko, Saudi-Arabien, Spanien, Ungarn ......... I/17Laos, USA ................................................................................... I/27Abchasien, Frankreich, Großbritannien, Italien,Riesending (Längste und tiefste Höhlen Deutschlands) .... II/51Spanien, USA, Papua-Neuguinea, Frankreich: Eingangverstürzt, Karbidlampenverbot ............................................. III/82Trou du Vent, Grotte des Chamois (F), La Mine (Tunesien)IV/129Von VeranstaltungenEuroparc Federation, UN-Hauptabteilung Presse undInformation, IUCN-Regionalkonferenz, Biospeläo-logisches Seminar, 2. Workshop „Bauen im Karst“ ..................... I/31PersonaliaAlfred Bögli, Karl Mais, Ronald Greeley ................................ I/28Hannes Dauer, Herbert Weichert, Claus Cramer,Karl Dietrich Adam, Urs Widmer ......................................... II/57Peter Henne ............................................................................. III/84VerbandEinladung zur 57. HV 2012 in Bad Mitterndorf undSpeleo Austria 2012 ................................................................... I/23Workshop Untertagebau im Karst ....................................... III/83Dr. Benno Wolf-Preis 2012 ................................................... III/92Protokoll der 57. HV in Bad Mitterndorf ........................... III/93Einladung zur 58. HV in Giengen-Hürben ...................... IV/100Publikationsliste ......................................................................... I/34Adressenliste der VdHK-Verbandsfunktionäre ............... IV/131Klimakataster, Ausleihe von Messgeräten beim VdHK ... III/96Einige Gedanken zum Referat für Ausbildung .................. III/95Leserbriefe ............................................................. III/85, IV/130Termine ............................................. I/27, II/62, III/96, IV/129Einladung 85 Jahre Drachenhöhle Syrau ............................ III/91Phantomkarst-Symposium in Belgien .................................. III/92Schriftenschau ............................................... I/33, II/63, IV/130



















