
Julia Simner- Beyond perception: synaesthesia as a psycholinguistic phenomenon

1
Title of PhD Thesis: Mirror-touch synaesthesia: The role of shared
representations in social cognition
Michael Joseph Banissy
Institute of Cognitive Neuroscience
University College London
PhD in Cognitive Neuroscience
I, Michael Joseph Banissy, confirm that the work presented in this thesis is my own.
Where information has been derived from other sources, I confirm that this has been
indicated in the thesis.

2
TABLE OF CONTENTS
Abstract 5
Publications Arising from Thesis 6
Acknowledgements 7
Chapter 1: Introduction 8
Chapter 2: Prevalence and Characteristics of Mirror-Touch Synaesthesia 36
Chapter 3: Sensory Processing in Synaesthesia 63
Chapter 4: Mirror-Touch Synaesthesia and Empathy 78
Chapter 5: Facial Expression Recognition in Mirror-Touch Synaesthesia 95
Chapter 6: A Methodological Introduction to TMS 112
Chapter 7: The Role of Sensorimotor Simulation in Auditory Emotion 125
Discrimination
Chapter 8: The Role of Sensorimotor Simulation in Facial Expression 140
Recognition
Chapter 9: Conclusions 155
References 175
Appendix 212

3
LIST OF FIGURES AND TABLES
Figure 1.1 fMRI data from a control and grapheme-colour synaesthetic subject when presented with synaesthesia inducing graphemes. 16 Figure 1.2 Summary of Muggleton et al. (2007). 19 Figure 1.3 Activations resulting from the interaction between mirror-touch synaesthete ‘C’ and non-synaesthetic control participants. 30 Figure 1.4 Sepcular and anatomical mappings reported by mirror-touch synaesthetes. 31 Figure 1.5 Summary of Banissy and Ward (2007). 32 Figure 2.1 Summary of task used to confirm potential cases of synaesthesia in experiment 1. 43 Figure 2.2 Mean reaction time performance and percentage of error tyes on human trials for mirror-touch synaesthetes recruited within the prevalence study compared to synaesthetes recruited via self referral. 49 Figure 2.3 Summary of task used for somatotopic cueing experiment. 51 Figure 2.4 Mean reaction time performance and percentage of error types for mirror-touch synaesthetes and non-synaesthetic control subjects observing a light flash on another person’s face or light flash only. 52 Figure 2.5 The ‘Who, What, Where Model of Mirror-Touch Synaesthesia’. 56 Figure 2.6 The influence of perspective on synaesthetic experience. 58 Figure 3.1 Synaesthetes who experience colour outperform individuals who do not experience synaesthetic colour on a measure of colour perception. 70 Figure 3.2 Synaesthetes who experience touch outperform individuals who do not experience synaesthetic touch on a measure of tactile perception. 72 Figure 4.1 Mirror-touch synaesthetes show significantly higher scores than controls on the emotional reactivity componenet, but not other components, of the Empathy Quotient. 83 Figure 4.2. In experiment 2, mirror-touch synaesthetes show significantly higher scores than controls on the emotional reactivity componenet, but not other components, of the Empathy Quotient. 88 Figure 4.3 Mirror-touch synaesthetes show significantly higher scores than controls on the fantasizing componenet, but not other components, of the Interpersonal Reactivity Index. 89 Figure 4.4 Mirror-touch synaesthetes show significantly higher scores than controls on the Openess subscale, but not other components, of the BFI. 89 Figure 5.1 Summary of tasks used. 102 Figure 5.2 Mean accuracy and reaction time performances of synaesthetes and controls on the films facial expression task. 104 Figure 5.3 Mean accuracy performances on the CFMT and CFMT+. 105 Figure 5.4 Mean error score for synaesthetes and controls on upright and inverted trials of the CFPT. 107 Figure 5.5 Mean accuracy performances on the expression matching and identity matching task for synaesthetes and controls. 108 Figure 6.1 The sequence of events for a transcranial magnetic stimulation pulse. 116 Figure 6.2 The effects of fibre orientation and electric field orientation for the application of a TMS pulse. 117 Figure 6.3 TMS-induced electrical fields produced by circurlar and figure-of-eight shaped coils. 117

4
Figure 6.4 The spatial and temporal resolution of TMS compared with other experimental techniques 118Figure 6.5 Functional dissociation method that can be employed using TMS. 121 Figure 7.1 Summary of cTBS and task protocol. 131 Figure 7.2 Summary of TBS sites stimulated. 133 Figure 7.3 Magnitude of disruption or facilitation in milliseconds following cTBS targeted at rSI, rPM and the vertex across task groups. 135 Figure 8.1 Summary of TMS sites stimulated. 146 Figure 8.2 Magnitude of disruption or facilitation across expression types following cTBS targeted at rSI, rIFG, and right V5 / MT. 149 Figure 9.1 The ‘Who, What, Where Model of Mirror-Touch Synaesthesia’. 164 Table 2.1 Reaction time performance and percentage of mirror-touch errors and other error types for potential mirror-touch synaesthetes when observing a human or corresponding object being touched. 48

5
ABSTRACT
Synaesthesia is a condition in which one property of a stimulus results in conscious
experiences of an additional attribute. In mirror-touch synaesthesia, the synaesthete
experiences a tactile sensation on their own body simply when observing touch to
another person. This thesis investigates the prevalence, neurocognitive mechanisms,
and consequences of mirror-touch synaesthesia. Firstly, the prevalence and
neurocognitive mechanisms of synaesthesia were assessed. This revealed that mirror-
touch synaesthesia has a prevalence rate of 1.6%, a finding which places mirror-touch
synaesthesia as one of the most common variants of synaesthesia. It also indicated a
number of characteristics of the condition, which led to the generation of a
neurocognitive model of mirror-touch synaesthesia. An investigation into the
perceptual consequences of synaesthesia revealed that the presence of synaesthesia is
linked with heightened sensory perception - mirror-touch synaesthetes showed
heightened tactile perception and grapheme-colour synaesthetes showed heightened
colour perception. Given that mirror-touch synaesthesia has been shown to be linked
to heightened sensorimotor simulation mechanisms, the impact of facilitated
sensorimotor activity on social cognition was then examined. This revealed that
mirror-touch synaesthetes show heightened emotional sensitivity compared with
control participants. To compliment this, two transcranial magnetic stimulation
(TMS) studies were then conducted to assess the impact of suppressing sensorimotor
activity on the expression recognition abilities of healthy adults. Consistent with the
findings of superior emotion sensitivity in mirror-touch synaesthesia (where there is
facilitated sensorimotor activity), suppressing sensorimotor resources resulted in
impaired expression recognition across modalities. The findings of the thesis are
discussed in relation to neurocognitive models of synaesthesia and of social cognition.

6
PUBLICATIONS ARISING FROM THESIS
Research Articles:
Banissy, M. J., and Ward, J. (2007). Mirror-touch synaesthesia is linked with
empathy. Nature Neuroscience, 10, 815-816.
Banissy, M. J., Walsh, V., and Ward, J (2009). Enhanced sensory perception in
synaesthesia. Experimental Brain Research, 196, 565-571.
Banissy, M. J., Cohen Kadosh, R., Maus, G. W., Walsh, V., & Ward, J. (2009).
Prevalence, characteristics, and a neurocognitive model of mirror-touch
synaesthesia. Experimental Brain Research, 198, 261-272.
Banissy, M. J., Sauter, D. A., Ward, J, Warren, J. E., Walsh, V., and Scott, S.
(Submitted). Suppressing sensorimotor activity modulates the discrimination
of auditory emotions but not speaker identity. Journal of Neuroscience.
Banissy, M. J., Garrido, L., Kusnir, F., Duchaine, B., Walsh, V, and Ward, J.
(Submitted). Superior facial expression but not idenitity recognition in mirror-
touch synaesthesia. Journal of Neuroscience.
Invited Book Chapters and Reviews:
Banissy, M. J., and Ward, J. (2008). On being moved: From mirror neurons to
empathy. Child and Adolescent Mental Health, 13, 50-51.
Ward, J., Banissy, M. J., and Jonas, C. (2008). Haptic perception in synaesthesia. In
Human Haptic Perception: Basics and Applications, Edited by
M.Grunwald. Birkhäuser, Basel.

7
ACKNOWLEDGEMENTS
Firstly, I would like Vincent Walsh for his unparalleled support and guidance. I
would also like to thank Jamie Ward for providing me with the opportunity to
complete this research, for his support, and for his advice. The two of them have
shaped my intellectual development and I will be forever grateful.
I am also thankful to a large number of my friends and colleagues at University
College London and the University of Sussex for their support. Of note, Roi Cohen
Kadosh, Brad Duchaine, Lúcia Garrido, Amir Javadi, Clare Jonas, Ryota Kanai, Neil
Muggleton, David Pitcher, Sophie Scott, and Lilli Tcheang. There are also a number
of friends and family who have provided me with encouragement and support over
the years; Lucy Annett, Edith Cole, Margaret Cole, Stanley Cole, Steven Cole, Claire
Doyle, Martin Family, Stephen Ford, Davina Reynolds, and Mark Taylor.
My sincerest thanks and love goes to Sian Fitzpatrick. Her encouragement, support
and faith in me have been incredible. I am thankful to you for so many reasons and
more grateful than I could ever say.
Finally, deepest thanks go to my parents, for their unconditional support and love
throughout the years. To my sister Jasmine and to my brother Jamie – you are both an
inspiration and I only wish I had the words to explain how much. Thank you for
everything. This is for you all, with love always.

Chapter 1
8
CHAPTER 1: INTRODUCTION
This chapter provides a summary of the motivation to investigate mirror-touch
synaesthesia. It proposes that synaesthesia relies upon similar mechanisms of
multisensory interactions that are present in non-synaesthetic individuals and can be
used to inform normal models of cognitive processing. The condition of synaesthesia
is introduced and the prevalence and characteristics of the condition are discussed.
An overview of current psychological and neurobiological studies is provided which
reveals insights into the neurocognitive mechanisms of synaesthesia and demonstrates
how the condition makes use of neural pathways involved in normal sensory
perception. Recent research demonstrates that developmental mirror-touch
synaesthesia appears to rely upon activation of the same somatosensory
representations within the human mirror-touch system that are activated when non-
synaesthetic individuals observe another person being touched The aims of this thesis
are described. These include i) investigations into the neurocognitive mechanisms of
mirror-touch synaesthesia and ii) investigations into the role of sensorimotor
simulation processes in emotion processing and empathy.
1.1 Origins of synaesthesia
Derived from the Greek roots syn (meaning together) and aisthesis (meaning
sensation), synaesthesia is a condition in which one property of a stimulus (the
inducer) gives rise to a conscious experience of a different attribute (the concurrent;
Grossenbacher and Lovelance, 2001). For example, in tone-colour synaesthesia,
sounds may elicit the experience of colour (Ward, Huckstep, and Tsakanikos, 2006);
in grapheme-colour synaesthesia, the visual presentation of achromatic letters or
numbers results in subjective experiences of colour (Simner et al., 2006); and in
lexical-gustatory synaesthesia, words trigger synaesthetic experiences of taste (Ward,
Simner and Auyeung, 2005).
Accounts of the condition can be traced to the 19th century (c.f. Jewanski, Day,
and Ward, 2009). For example, Sachs (1812) describes synaesthesia involving
colours for music and simple sequences. Later, Galton (1880) described cases of
individuals in whom numbers were consciously visualized in space (spatial number
forms) and of synaesthesia involving colour. While early accounts of the condition

Chapter 1
9
aroused some interest, failure to develop an objective approach to confirm the
phenomenon resulted in a decline in research. It was not until the advent of the
development of new behavioural and neurophysiological measures which could be
used to corroborate self reports that the topic of synaesthesia returned as a topic of
legitimate scientific investigation (Baron-Cohen et al., 1987; Cytowic and Wood,
1982; Marks, 1975; see Ramachandran and Hubbbard, 2001; Rich and Mattingely,
2002; Sagiv, 2004 for reviews).
Since this time, research into the topic of synaesthesia has grown rapidly with
a focus moving beyond exploring the reality of the condition to consider how
synaesthesia can be used to inform models of typical cognition in domains such as
numerical cognition (Cohen Kadosh and Henik, 2007), language (Simner, 2007),
multisensory processing (Sagiv and Ward, 2006), imagery (Barnett and Newell, 2008;
Spiller and Jansari, 2008), and attention (Treisman, 2004). In this introductory
chapter, I will review this literature by describing studies on the prevalence,
authenticity and aetiology of synaesthesia. The focus of this thesis is to investigate a
newly documented type of synaesthesia, mirror-touch synaesthesia (in which
individuals’ experience tactile sensations on their own body simply when observing
touch to others) and to use this condition to examine more general neurocognitive
processes in social cognition. In this chapter, I will introduce research investigating
synaesthesia involving touch and consider the evidence that mirror-touch synaesthesia
relies upon similar mechanisms to multisensory interactions which are shown in non-
synaesthetic individuals. Finally, I will discuss how synaesthesia may be used to
inform typical models of cognition and discuss the role of sensorimotor simulation in
social cognition.

Chapter 1
10
1.2 Prevalence and characteristics of synaesthesia
Synaesthesia is typically considered as having three defining features; 1)
experiences are conscious perceptual or percept-like experiences; 2) experiences are
induced by an attribute not typically associated with the conscious experience; 3)
these experiences occur automatically (Ward and Mattingley, 2007). Further, the
synaesthetic percept tends to co-exist with the percept of the inducing stimulus rather
than over-riding it – for example in lexical-gustatory synaesthesia written or heard
words are recognised but also result in a simultaneous subjective sensation of taste in
the mouth and tongue area (Ward and Simner, 2003). Note that throughout the thesis
the terminology of referring to different types of synaesthesia in terms of inducer-
concurrent pairs separated with a hyphen is used. As such, touch-colour synaesthesia
refers to tactile inducers eliciting a concurrent experience of colour, and vision-touch
synaesthesia refers to a visual inducer eliciting a tactile experience.
Cases of synaesthesia can be either developmental or acquired, with
developmental cases thought to be dependent upon genetic and environmental factors
and acquired cases reflecting synaesthesia following specific environmental
influences (e.g. following brain injury or drug ingestion). Developmental forms of
synaesthesia have been shown to run in families and previous research suggested that
the condition may be more common in women than men, which may reflect an X-
linked dominant mode of inheritance (Baron-Cohen, Burt, Smith-Laittan, Harrison,
and Bolton, 1996). More recent research indicates that synaesthesia may be equally
common within males and females and that previous methodologies may have led to
an over inflated male-female ratio (Ward and Simner, 2005; but see Barnett et al.,
2008). Similarly, reports of twins discordant for synaesthesia (Smilek, Mofatt,
Pasternak, White, Dixon, and Merilke, 2002), as well as evidence that synaesthesia

Chapter 1
11
can skip generations (Hubbard and Ramachandran, 2003), and that the proportion of
sons or daughters born to synaesthete mothers does not significantly differ (Barnett et
al., 2008; Ward and Simner, 2005), suggest that an X-linked dominant mode of
inheritance may be an over simplified account of the genetic mechanisms which
underlie developmental forms of the condition (Asher et al., 2009).
Current estimates on the prevalence of developmental synaesthesia indicate
that the condition has a prevalence rate of at least 4% and a female to male ratio of 1:1
(Simner et al., 2006; Ward and Simner, 2005). Although, depending on whether one
includes cases of ordinal linguistic personification (in which individuals attribute
genders or personalities to letters or numbers; Simner and Holenstein, 2007) or spatial
number forms (Sagiv, Simner, Collins, Butterworth, and Ward, 2006), the prevalence
rate of 4% is likely to be much higher (Simner et al., 2006).
A trend of all studies of the prevalence of synaesthesia is to report a higher
proportion of synaesthetes who experience colour evoked by letters or other linguistic
stimuli (e.g. grapheme-colour / day-colour synaesthesia; Baron-Cohen, Burt, Smith-
Laittan, Harrison, and Bolton, 1996; Rich, Bradshaw, and Mattingley, 2005; Simner
et al., 2006). It is perhaps not surprising that this variant of synaesthesia has been the
topic of much research amongst synaesthesia researchers. Research into this variant
of the condition has highlighted a number of interesting individual differences
between synaesthetes. For example, distinctions have been made between projector
and associator synaesthetes; which distinguishes between synaesthetes whose locus of
experienced colour is projected to a specific spatial location (projector synaesthetes)
and synaesthetes whose concurrent is perceived internally or in the ‘minds eye’
(associator synaesthetes) (Dixon, Smilek, and Merilke, 2004; see also Ward, Li, Salih,
and Sagiv, 2007). Similarly, Ramachandran and Hubbard (2001) have categorised

Chapter 1
12
synaesthetes based upon the level of induction of the synaesthesia, distinguishing
between higher synaesthetes, in whom conceptual properties of a grapheme trigger
colours (e.g. a number name or dice pattern for a particular number), and lower
synaesthetes in whom the physical properties of the grapheme (e.g. shape / form)
trigger synaesthetic experience. Distinctions such as these may also be valid for other
variants of synaesthesia (see below).
1.3 Authenticity and perceptual nature of synaesthesia
1.3.1 Authenticity of synaesthesia
Typically, the authenticity of synaesthesia has been confirmed behaviourally
using tests of consistency of synaesthetic associations over time. Synaesthetes tend to
show greater consistency in inducer-concurrent pairings (synaesthetes are typically
around 90% consistent) compared with non-synaesthetic subjects asked to freely
associate or use a particular strategy (i.e. memory or imagery), even when tested over
longer time periods (Baron-Cohen, Harrison, Goldstein and Wyke, 1993). This
pattern has been shown to be the case in a number of variants of synaesthesia,
including grapheme-colour (Baron-Cohen et al., 1996), emotion-colour (Ward, 2004)
and lexical-gustatory synaesthesia (Ward and Simner, 2003).
A further paradigm used to confirm the automaticity of the synaesthetic
experience has been the modified ‘synaesthetic stroop’ task in which synaesthetic
inducers are paired with either a congruent or incongruent concurrent. For example,
if a grapheme-colour synaesthete perceives the letter A as red, then this grapheme
would be presented in a colour which is either congruent with synaesthetic experience
(e.g. A) or a colour which is incongruent with synaesthetic experience (e.g. A). The
subject is asked to ignore the synaesthetic colour and name the true colour of the
grapheme. Grapheme-colour synaesthetes tend to be faster in the congruent relative

Chapter 1
13
to incongruent condition (Mills, Boteler, and Oliver, 1999), while non-synaesthetes do
not show this pattern. As with tests of consistency, this pattern of performance has
been found for different subtypes of synaesthesia, including not only grapheme-colour
synaesthesia, but also music-colour (Ward et al., 2006), music-taste (Beeli, Esslen,
and Jäncke, 2005), and mirror-touch (Banissy and Ward, 2007; summarised later in
Figure 1.5) variants.
Notably, individuals who have over-learned colour associations may also
behave similar to synaesthetes on the synaesthetic stroop task. For example, Elias and
colleagues report a single case study in which a non-synaesthetic individual with
reliable digit-colour associations, as a result of years of training using coloured
numerical codes in cross-stitching, performed comparably to synaesthetic subjects on
tests of consistency and stroop interference (synaesthetes differed from the control on
functional magnetic resonance imaging [fMRI] measures of synaesthesia in colour
selective regions but not on behavioural measures; Elias, Saucier, Hardie, and Sart,
2003). This is consistent with the findings of MacLeod and Dunbar (1988) who
trained non-synaesthetic subjects to associate black and white geometric shapes with
colour names over thousands of trials. When participants were later presented with a
stroop task, involving the geometric shapes presented in a congruent or incongruent
colour, the typical stroop interference pattern was observed (MacLeod and Dunbar,
1988). In neither study were subjects experiencing synaesthetic colour interactions,
implying that associative (rather than perceptual) components may be able to account
for some of the patterns of performance shown by colour synaesthetes on synaesthetic
stroop and consistency measures. However, more recent findings suggest that, in
colour synaesthetes, the synaesthetic stroop effect may be a consequence of both
perceptual and associative components (Nikolić, Lichti and Singer, 2007). Using

Chapter 1
14
principles of colour-opponency (Hering, 1868/1964), Nikolic and colleagues varied
incongruent stimuli within the synaesthetic stroop by using colour-opponent (i.e. red
changed to green) or non-opponent colours (i.e. red changed to blue). If synaesthetic
stroop relies upon perceptual processes as well as associative components then one
would expect the colour-opponent condition to produce the most interference – this
pattern was observed (Nikolić et al., 2007).
1.3.2 Psychophysical studies
In addition to measures of stroop and consistency, other psychophysical
measures have been used to investigate how closely synaesthetic perception resembles
veridical sensory perception. Again, much of this work has focussed on the
perceptual reality of synaesthetic colours in grapheme-colour synaesthesia. These
findings indicate that synaesthetic and real colours interact under conditions of
binocular rivalry (Kim, Blake, and Palmeri, 2006); that synaesthetic colours can
induce orientation contingent colour adapting after-effects such as a synaesthetic
‘McCollough Effect’ (Blake, Palmeri, Marois, and Kim, 2004; but see Hong and
Blake, 2008); that synaesthetic and real colours can combine to produce apparent
motion (Kim et al., 2006); and that, in projector synaesthetes, synaesthetic experience
can be modulated by background contrast, implying that synaesthesia relies upon
early contrast-dependent visual mechanisms (Hubbard, Manohar, and Ramachandran,
2006; Witthoft and Winawer, 2006).
1.3.3 Neuroimaging studies
Aside from behavioural and psychophysical tests, functional brain imaging
methods have been used to distinguish between synaesthetic and non-synaesthetic
subjects. These have included positron emission tomography (PET) studies of

Chapter 1
15
word/grapheme-colour synaesthesia triggered by speech (Paulesu et al., 1995); fMRI
studies of grapheme-colour (Aleman, Rutten, Sitskoorn, Dautzenberg, and Ramsey,
2001; Hubbard, Arman, Ramachandran, and Boynton, 2005; Weiss, Zilles, and Fink,
2005; Sperling, Prvulovic, Linden, Singer, and Stirn, 2006; Rich et al., 2006), mirror-
touch (Blakemore, Bristow, Bird, Frith, and Ward, 2005), word-colour (Aleman et al.,
2001; Nunn et al., 2002; Gray, Parslow, Brammer, Chopping, Vythelingum, and
ffytche, 2006), digit-colour (Elias et al., 2003), people-colour (Weiss, Shah, Toni,
Zilles, and Fink, 2001), time-colour (Steven, Hansen, and Blakemore, 2006), time-
space (Steven et al., 2006), sound-vision (Stewart, Mulvenna, Griffiths, and Ward, in
prep), and bidirectional synaesthesia (Cohen Kadosh, Cohen Kadosh, and Henik,
2007). In addition, there have been two diffusion tensor imaging studies (DTI) of
grapheme-colour synaesthesia (Rouw and Scholte, 2007; Jäncke, Beeli, Eulig, and
Hänggi, 2009).
While there is some inconsistency between studies, the majority point to
synaesthetic experience being correlated with activations in brain regions involved in
normal perceptual experience. For example, studies investigating synaesthesia
involving colour tend to report activation of colour area V4 / V8 for synaesthesia
inducing stimuli (e.g. Hubbard et al., 2005; Nunn et al., 2002; Sperling et al., 2006),
although not always (i.e. Paulesu et al., 1995; Weiss et al., 2005; Figure 1.1). The
reasons behind this inconsistency remain unclear, although they may be related to
differences in task demands, statistical power, or qualitative differences between
synaesthetic subjects (Hubbard et al., 2005). Moreover, by correlating performance
on different synaesthetic psychophysical paradigms with fMRI activations, Hubbard
and colleagues (2005) show that synaesthetes who show larger effects on

Chapter 1
16
psychophysical tasks show greater activations in colour selective regions of the visual
cortex (V4).
Figure 1.1 fMRI data from a control and grapheme-colour synaesthetic subject when presented with synaesthesia inducing graphemes. Colour area V4 (as per Wade, Brewer, Rieger, and Wandell, 2002) is shown in purple and the grapheme area (Gr) in blue. The synaesthete shows activation in both V4 and Gr, while the control only shows activation in Gr. Taken from Hubbard and Ramachandran (2005).
Recent research making use of DTI techniques is also consistent with the
notion that inter-individual differences within the synaesthetic population may
contribute to different patterns of brain activation. DTI is a neuroimaging technique
which measures the diffusion of water molecules in the living human brain to enable
analysis of the degree of structural connectivity between brain regions (Basser, 1995).
Using this method, Rouw and Scholte (2007) report that grapheme-colour
synaesthesia is linked with increased structural connectivity (as compared with non-
synaesthetes) in the left superior parietal cortex, right inferior temporal cortex
(adjacent to the fusiform gyrus) and in a bilateral cluster located beneath the central
sulcus. Of these four clusters, greater connectivity in the right inferior temporal
cortex was found to be strongest in ‘projector’ synaesthetes who saw their colours in

Chapter 1
17
the outside world (compared to ‘associator’ synaesthetes who saw their colours in
their mind’s eye).
In addition to shared activations in brain regions involved in normal and
synaesthetic perceptual experience (e.g. V4 / V8 in colour), a number of common
brain activations have been reported across different variants of the condition. Two
brain regions of note are the insula and the intraparietal sulcus (IPS). IPS and insula
activations have been reported in studies of both visual and auditory grapheme-colour
synaesthesia (insula activations - Nunn et al., 2002; Paulesu et al., 1995; IPS
activations - Paulesu et al., 1995; Weiss et al., 2005); synaesthesia involving spatial
number forms (Tang, Ward, and Butterworth, 2008); and studies of sound-vision
synaesthesia (Stewart et al., in prep). Insula activations have also been found in
mirror-touch synaesthesia (Blakemore et al., 2005). Both regions have been
implicated in multi-sensory processing and integration (Bushara, Grafman, and
Hallett, 2001; Hadjikhani and Roland, 1998; Olson, Gatenby, and Gore, 2002),
indicating that they may play a role in integrating synaesthesia inducing materials
with experience.
1.3.4 Transcranial magnetic stimulation (TMS) studies
TMS is a non-invasive technique that uses induced current to depolarize the
cell membrane in the cortex leading to a temporary modulation of neural activity in
the stimulated cortex (Walsh and Rushworth, 1999). This method enables
examination of the necessity of stimulated brain structures for given cognitive
functions. To date, two TMS studies have been conducted to investigate the necessity
of parietal cortex activations in grapheme-colour synaesthesia (Esterman, Verstynen,
Ivry, and Robertson, 2006; Muggleton, Tsakanikos, Walsh, and Ward, 2007).
Esterman et al. investigated the magnitude of synaesthetic interference on a

Chapter 1
18
synaesthetic stroop task in two ‘projector’ synaesthetes following TMS to a parieto-
occipital region in the right hemisphere, to the corresponding region in the left
hemisphere, and to area V1. They found that the magnitude of synaesthetic
interference was reduced following TMS to the right parieto-occipital area, but not for
the other two brain regions (Esterman et al., 2006). Extending upon this, Muggleton
et al. contrasted the effects of TMS over four parietal brain regions (right parieto-
occipital, left parieto-occipital, right parietal and left parietal regions) in five
grapheme-colour synaesthetes (comprised of one ‘projector’ and four ‘associators’).
Consistent with the findings of Esterman et al., these authors also report that the
automaticity of synaesthetic experience (as measured using a synaesthetic stroop task)
was disrupted following stimulation of the right parieto-occipital area only
(Muggleton et al., 2007; Figure 1.2). Therefore both studies suggest that the right
parieto-occipital cortex is necessary for the experience of synaesthesia. In non-
synaesthetes this brain region has been shown to participate in visual feature binding
(Freidman-Hill, Robertson, and Treisman, 1995; Donner, Kettermann, Diesch,
Ostendorf, Villringer, and Brandt, 2002) and one explanation for the selective TMS
disruption observed is that the right parieto-occipital area may act in the spatial
binding of graphemes with synaesthetic colour (Esterman et al., 2006; Muggleton et
al., 2007). If so, this suggests that, in grapheme-colour synaesthesia, synaesthetic
experience acts upon the same cortical pathways that exist in the non-synaesthetic
brain (Cohen Kadosh and Walsh, 2008; Cohen Kadosh, Henik, Catena, Walsh, and
Fuentes, 2009). However, even if synaesthetes use common mechanisms of cross-
modal binding, it remains unclear whether synaesthetic binding follows the same
time-course of processing as veridical cross-modal binding or how the parietal lobe

Chapter 1
19
interacts with processing in other cortical areas (i.e. sensory-selective cortical regions)
to elicit synaesthetic experience.
Figure 1.2 Summary of Muggleton et al. (2007). (a) The location of stimulated right parietal-occipital region (RPO; x = 22, y = -71, z = 27) and right parietal region. (b) Interference between real and synaesthetic colours on synaesthetic stroop task. Performance for individual synaesthetes, divided between projectors and associators, following TMS targeted at the RPO region compared to control condition. Synaesthetes EP and CP were reported by Esterman et al. (2006) and are shown for comparison. Adapted from Muggleton et al. (2007) with permission.
1.3.5 Electrophysiological studies
As with TMS studies, to date there have been few studies utilizing
electrophysiological techniques to investigate the time course neural activity in
synaesthetic experience. Schiltz and colleagues (1999) investigated the
electrophysiological correlates of grapheme-colour synaesthesia (n = 17) and reported
an increased positivity at frontal and central scalp sites of synaesthetes (relative to
controls) occurring around 150 msec after stimulus onset which was sustained until
600 msec. A more recent study by Beeli and colleagues, conducted with grapheme-
colour synaesthetes who only experience colours from spoken letters and words (n =
16), revealed differences (reduced amplitude and / or increased latencies) in the
auditory N1, P2, and N2 deflections (Beeli, Esslen, and Jäncke, 2008). Source
a. b.

Chapter 1
20
localization implicated intracerebral sources of these components to lay in inferior
temporal and orbitofrontal brain regions (although few electrode sites were available).
These authors interpret their finding as evidence for increased cortical wiring in
synaesthetes (c.f. Ramachandran and Hubbard, 2001; Bargary and Mitchell, 2008),
but may also be consistent with accounts of synaesthesia which posit differences in
local mechanisms of cortical inhibition (Cohen Kadosh and Walsh, 2008).
In addition to this, two single case studies and one group study have
investigated auditory-visual synaesthesia. In a single case study of acquired auditory-
visual synaesthesia, Rao and colleagues report that synaesthesia inducing sounds
resulted in a modulation of the auditory evoked N1 deflection (Rao, Nobre,
Alexanader and Cowey, 2007). Rizzo and Eslinger (1989) investigated the
electrophysiological correlates of a single case of developmental auditory-visual
synaesthesia, but restricted analysis to three electrode sites (O1/2, or Oz). No
abnormal potentials were found at these three sites, but this does not rule out the
possibility that abnormal potentials may occur at alternative electrode sites. A more
recent group study of tone-colour synaesthesia (n = 10; Goller, Otten, and Ward,
2009) revealed early onset (around 100msec after stimulus onset) differences in
deflections of the auditory evoked potential (auditory N1, N2, and P2). No posterior
difference was observed, implying that synaesthetic experience may be generated
locally (potentially through mechanisms of local differences in cortical inhibition; c.f.
Cohen Kadosh and Walsh, 2008).
1.3.6 Neurocognitive models of synaesthesia
While much research has determined the authenticity of synaesthesia, the
neurocognitive mechanisms which underpin synaesthesia are a subject of uncertainty.
A current area of dispute in the synaesthesia literature is whether synaesthetic

Chapter 1
21
experience is due to additional structural connectivity (i.e. structural differences),
malfunctions in cortical inhibition (i.e. functional but not structural differences) or a
combination of both (Bargary and Mitchell, 2008; Cohen Kadosh and Henik, 2007;
Cohen Kadosh and Walsh, 2008; Grossenbacher and Lovelace, 2001; Hubbard and
Ramachandran, 2005; Rouw and Scholte, 2007; Smilek, Dixon, Cudahy, and Merikle,
2001).
Supporting evidence for structural connectivity accounts is provided by a DTI
study which reports greater white matter coherence in grapheme-colour synaesthetes
compared to non-synaesthetic control subjects - grapheme-colour synaesthetes show
increased structural connectivity in inferior-temporal, parietal and frontal brain
regions when compared to non-synaesthetes (Rouw and Scholte, 2007). Some
authors have interpreted these findings to be consistent with accounts of synaesthesia
which argue in favour of aberrant connectivity between adjacent cortical regions
(Ramachandran and Hubbard, 2001; Hubbard, 2007). For example, given that the
brain regions involved in the visual recognition of graphemes (i.e. the putative visual
word form area; Cohen and Dehaene, 2004) lie adjacent to brain areas involved in
colour perception (i.e. human V4 - Wade et al., 2002), it has been suggested that
grapheme-colour synaesthesia may arise from direct cross-activation between these
regions as a result of either increased connectivity between adjacent brain regions or
reduced inhibition between adjacent regions. This local cross-activation account has
also been extended to explain sequence-space synaesthesia (i.e. number forms), in
terms of cross-activation between adjacent parietal regions (Ramachandran and
Hubbard, 2001), and may also be important for other variants of synaesthesia (e.g.
lexical-gustatory synaesthesia; Ward, Simner and Auyeung 2005). However, it is of
note that the generality of enhanced structural connectivity in grapheme-colour

Chapter 1
22
synaesthesia is debatable (e.g. see Jäncke et al., 2009) and the extent to which these
differences extend to other variants of synaesthesia (e.g. mirror-touch synaesthesia) or
play a causal role in generating synaesthetic experience remains unknown.
In contrast to aberrant cortical connectivity accounts, other authors have
argued in favour of feedback accounts of synaesthesia which explain the condition in
terms of malfunctions in cortical inhibition, either within (Cohen Kadosh and Henik,
2007; Cohen Kadosh and Walsh, 2008) or between brain regions (e.g. from a
multisensory cortical nexus; Grossenbacher and Lovelace, 2001). According to this
view, synaesthesia is mediated by the same cortical pathways that exist in the non-
synaesthetes’ brain (i.e. aberrant connectivity is not necessary to induce synaesthesia),
but unmasking of these pathways due to alterations in cortical inhibition results in
synaesthetic experience (Grossenbacher and Lovelace, 2001; Cohen Kadosh and
Walsh, 2006; Cohen Kadosh and Henik, 2007; Cohen Kadosh and Walsh, 2008;
Cohen Kadosh, Henik, Catena, Walsh, and Fuetnes, 2009). Evidence that TMS
disruption of the parietal lobe impairs synaesthetic stroop performance (Esterman et
al., 2006; Muggleton et al., 2007); that synaesthetic-like experiences can be induced
following hallucinogenic drugs (i.e. in the absence of altered cortical connectivity;
Aghajanian and Marek, 1999); and that colour synaesthesia can be induced in non-
synaesthetes (individuals without aberrant connectivity) using post-hypnotic
suggestion (Cohen Kadosh et al., 2009) are consistent with this.
1.4 Synaesthesia involving touch
Synaesthesia involving touch has been less well documented than other
variants of synaesthesia. Despite this, there are some reports of both developmental
and acquired synaesthesia involving either a tactile inducer or concurrent. These
cases are discussed below.

Chapter 1
23
1.4.1 Synaesthesia involving tactile inducers
To date much research on synaesthesia involving tactile inducers has centred
on cases of touch-vision synaesthesia in which touch results in visualised photisms.
For example, Armel and Ramachandran (2001) report a case of acquired touch-vision
synaesthesia shown by a patient who suffered blindness due to retinitis pigmentosa.
One year after becoming completely blind the patient began to experience
synaesthetic visual photisms from haptic stimuli. Such sensations were projected onto
the spatial location of the body part touched irrespective of the location of the body
part in space (e.g. a touch to the right hand in left space would elicit photisms in left
space). Detailed investigations indicated that the intensity of tactile stimulation
required to induce synaesthesia was lower when body parts were presented in front of
the patient relative to behind the head (i.e. moving the hands from in front of the head
to behind the head), suggesting that despite the patient being blind a preference was
shown for when the inducer was “in view”. This may be indicative of a body-based
spatial reference that incorporates information about gaze and head orientation. Such
findings are consistent with evidence from non-synaesthetes on normal multi-sensory
interactions between touch and vision indicating that the spatial reference frame
which processes current hand position is modulated by gaze direction (Armel and
Ramachandran, 2001).
In addition to this, cases of blind synaesthetes for whom Braille reading elicits
a visual concurrent have been reported (Wheeler and Cutsforth, 1921; Steven and
Blakemore, 2004). In the latter case, synaesthete JF, who suffered from retinitis
pigmentosa leading to blindness, consistently experienced coloured visual photisms
both when reading Braille and when thinking about Braille characters (Steven and
Blakemore, 2004). Similar geometrical arrangements of Braille dots evoked similar

Chapter 1
24
colours, but photisms were not elicited when touching other textures or objects.
Notably, it has been reported that J.F experienced visual synaesthesia from childhood
(i.e. since before losing his sight; Steven, Hansen, and Blakemore, 2006) so it may be
the case that his synaesthesia reflects a different manifestation of grapheme-colour
synaesthesia in which graphemes are processed haptically rather than visually (Ward,
Banissy, and Jonas, 2008).
There have been relatively few documented cases of developmental touch-
vision synaesthesia. While Day (2005) reports that 4% of synaesthetes report
‘coloured-touch’ these figures are based on self reported cases only (a failure to
objectively confirm these self reports with tests of genuiness may mean that this 4%
claim includes false positive cases; c.f. Simner et al., 2006) and no information is
given regarding the nature of these cases (i.e. developmental or acquired cases).
Recently, two cases of developmental touch-vision synaesthesia in which touch
triggers visual sensations of colour (TV and EB) have been investigated more
systematically (Ward et al., 2008). Each case indicated important distinctions
between the spatial representations which underpin synaesthetic experience.
Moreover, for synaesthete TV coloured photisms were projected onto the spatial
location of the body part touched, whereas for EB photisms were perceived in her
mind’s eye. This distinction appears similar to the projector / associator distinction in
grapheme-colour synaesthesia outlined above (Dixon et al., 2004).
It is of note that while synaesthetes TV and EB appeared consistent on a test
of consistency for their synaesthesia, involving 40 different tactile stimuli across two
testing sessions; they were not shown to be significantly more consistent than control
subjects. This is likely to be related to elevated levels of control consistency (c.f.
Kusnir, MSc Thesis, University of London, 2008) indicating that the touch-vision

Chapter 1
25
synaesthesia reported by TV and EB may rely upon similar multi-sensory principles
which underpin non-synaesthetic touch-vision interactions (Ward et al., 2008).
Moreover, cross-modal correspondences between roughness and luminance (rougher
textures associated to darker colours) and pressure and luminance (higher pressure
associated with darker colours) were found for both synaesthetes and control subjects.
Consistent with this, previous reports of touch-vision synaesthesia have indicated a
relationship between pressure and luminance, in which the synaesthete experienced
dark coloured photisms to hard objects (i.e. higher pressure) and lightly coloured
photisms to soft objects (i.e. lower pressure) (Smith, 1905). Thus, it may be the case
that developmental touch-vision synaesthesia relies upon similar mechanisms of cross
modal transfer as observed in non-synaesthetic multi-sensory interactions between
touch and vision. This would be consistent with findings indicating that other variants
of synaesthesia appear to act upon the ‘normal’ architecture for cross-modal
interactions (e.g. Ward, Huckstep, and Tsakanikos, 2006; Blakemore et al., 2005;
Sagiv and Ward, 2006).
1.4.2 Synaesthesia involving a tactile concurrent
As with cases of synaesthesia in which touch acts to induce synaesthetic
experience, cases of synaesthesia involving a tactile concurrent are less well
documented than other more common variants of synaesthesia. Despite this, there are
reports of acquired auditory-tactile synaesthesia in which sounds elicit tactile
sensations (Ro et al., 2007) and of both acquired and developmental mirror-touch
synaesthesia in which observed touch results in tactile experiences on the observer’s
own body (Halligan, Hunt, Marshall, and Wade, 1996; Bradshaw and Mattingley,
2001; Blakemore et al. 2005; Banissy and Ward, 2007). There is also evidence that
the presence of synaesthesia for colour is linked to a greater incidence mitempfindung

Chapter 1
26
(the referral of a tactile sensation away from the stimulation site; Burrack, Knoch, and
Brugger, 2006).
Synaesthetic interactions involving hearing and touch have rarely been
documented, however recently Ro et al. (2007) report a single case of acquired
auditory-tactile synaesthesia in a female patient following a discrete neurological
lesion to the right ventrolateral thalamus. The synaesthesia was first reported 18
months post lesion when the patient began to feel tactile sensations in response to
sounds. The synaesthetic somatosensations were typically experienced on the
patient’s left upper part of the body and a test of consistency (across three testing
sessions separated by 35 and 15 days) indicated that they were generally consistent
over time. Magnetic resonance imaging (MRI) and diffusion tensor imaging (DTI)
conducted at approximately 3 years post lesion indicated disorganised fibre bundles in
the right ventrolateral thalamus (lesion site) - at 3 years post onset DTI tracking from
the unaffected left hemisphere showed direct projections to motor / premotor cortices,
whereas fibre bundles in the lesioned hemisphere were disorganised and smaller
compared to the unaffected hemisphere. DTI conducted at 1.3 years post onset (i.e.
before synaesthetic experiences were reported) indicated no white matter differences
between the right and left ventrolateral thalamus. The authors suggest that this
disorganisation in cortico-callosal pathways may account for synaesthetic experiences
(Ro et al., 2007; see Chapter 9).
In addition to cases of acquired auditory-tactile synaesthesia there are a
number of accounts of acquired synaesthesia involving vision-touch interactions.
For example, patient D.N., suffered paralysis and loss of sensation in the left side of
his body following stroke. This resulted in D.N. being unable to feel any tactile
stimulation presented to the left side when the touch was hidden from view, however

Chapter 1
27
if tactile stimulation was made visible then D.N. was able to feel touch to the left side.
Similarly, when observing previous videos of his arm being touched and informed
that this reflected live video feedback D.N. reported being able to feel his arm being
touched despite the fact that the experimenter was not actually touching him. In this
sense, observed bodily touch attributed to the patient lead to tactile sensations,
indicating that in some conditions vision alone can be sufficient to elicit tactile
stimulation (Halligan et al., 1996). Such findings appear consistent with research in
the non-synaesthetic population which indicates that observing one’s own body can
lead to enhancements in one’s own tactile sensitivity (Taylor-Clarke, Kennett, and
Haggard, 2004) and with evidence provided by Rorden and colleagues (1999) of a
patient whose tactile detection increased when observing a flash of light to a rubber
hand seen in the same orientation and directly above the patient’s concealed hand (i.e.
when the rubber hand was attributed to the participant’s own body). Taken
collectively these findings highlight the important role of vision, and more so of
observing one’s own body, on haptic perception.
In addition to cases of acquired vision-touch synaesthesia involving one’s own
body there is also one case report involving an interaction between observed and
actual pain (“mirror pain”). This anecdotal report, given to clinicians posthumously
by the patient’s wife, describes a man who experienced observed pain to others as
actual pain to himself (Bradshaw and Mattingley, 2001). Here the inducer is observed
touch to another’s body rather than to one’s own body as described above. The
patient was known to have suffered widespread cancer, but as this case was reported
post-mortem no information about the neural circuitry involved was available. More
recently however, evidence for the interpersonal sharing of observed pain has been
provided (Singer, Seymour, O’Doherty, Kaube, Dolan, and Frith, 2004; Morrison,

Chapter 1
28
Lloyd, di Pellegrino, and Roberts, 2004; Avenanti, Beuti, Galati, and Aglitoi, 2005).
For example, Avenanti and colleagues (2005) report that observing pain to another
person results in significant reductions in motor evoked potentials (MEPs). The
modulation of MEP amplitude correlated with subjective ratings of the sensory
aspects of pain attributed by the observer to the actor and was somatotopically
organised such that the reduced amplitude was specific to the muscles observed in a
painful event. These authors suggest that the findings provide evidence for a mirror-
pain resonance system in which observed pain is matched to the observer’s own
sensorimotor representation of pain. Such interpretation builds upon the findings of
mirror neurons within the monkey premotor cortex and inferior parietal lobule, which
respond both when a monkey performs an action and when the monkey watches
another person perform a similar action (Gallese, Fadiga, Fogassi, and Rizzolatti,
1996; Rizzolatti and Craighero, 2004) and evidence for similar mirror systems in the
human brain for not only action (Buccino et al., 2001), but also touch (Keysers,
Wicker, Gazzola, Anton, Fogassi, and Gallese, 2004; Blakemore et al., 2005; Ebisch,
Perucci, Ferretti, Del Gratta, Luca Romani, and Gallese, 2008), pain (Singer et al.,
2004; Aventani et al., 2005), disgust (Wicker, Keysers¸ Plailly, Royet, Gallese, and
Rizzolatti, 2003) and other emotions (Carr, Iacoboni, Dubeau, Mazziotta, and Lenzi,
2003).
Similar to the case of acquired “mirror pain” described above; developmental
cases of vision-touch or “mirror-touch” synaesthesia have also been documented
(Blakemore et al., 2005; Banissy and Ward, 2007). First reported in a single case
fMRI study (Blakemore et al., 2005), mirror-touch synaesthesia refers to cases of
synaesthesia in which observing touch to another person leads to tactile sensations on
the equivalent part of the synaesthete’s own body. In the original study by Blakemore

Chapter 1
29
and colleagues (2005) the case of synaesthete C was described. C reports
experiencing touch on her own body when observing another person being touched,
but not when observing inanimate objects being touched. These experiences have
been described as being automatic, in so far as they occur whenever she observes
another person being touched, and to have occurred throughout her lifetime. Her
experiences mirror observed touch to another person, such that observing touch to
another person’s left facial cheek leads to a sensation of touch on her own right facial
cheek (i.e. as if looking at a mirror reflection of herself). Using fMRI Blakemore and
colleagues investigated the neural systems underlying C’s synaesthetic experience by
contrasting brain activity when watching videos of humans relative to objects being
touched (the latter did not elicit synaesthesia) in the synaesthete and in 12 non-
synaesthetic control subjects. In controls a network of brain regions were activated
during the observation of touch to a human relative to an object, including primary
and secondary somatosensory cortex, premotor regions and the superior temporal
sulcus. Similar brain regions were also activated during actual touch, indicating that
observing touch to another person activates a similar neural circuit as actual tactile
experience – a “mirror touch” system. A comparison between synaesthete C and non-
synaesthetic subjects indicated that the synaesthete showed hyper activity within a
number of regions within this network (including primary somatosensory cortex) and
additional activity in the anterior insula (Figure 1.3). Thus suggesting that mirror-
touch synaesthesia is a consequence of increased neural activity in the same mirror-
touch network that is evoked in non-synaesthetic controls when observing touch to
another person (Blakemore et al., 2005).

Chapter 1
30
Figure 1.3 Activations resulting from the interaction between mirror-touch synaesthete ‘C’ and non-synaesthetic control participants. The subtraction shown indicates the brain regions more active in synaesthete ‘C’ compared to non-synaesthete controls when observing touch to a human relative to an object. The primary (SI) and secondary somatosensory cortex (SII), bilateral anterior insular and the left premotor cortex were significantly more active in C than in the control group (Blakemore et al., 2005).
More recently, Banissy and Ward (2007) report a behavioural study of ten
developmental mirror-touch synaesthetes, including C. Notably, while all
synaesthetes report similar experiences (i.e. tactile sensations when observing touch to
another person) some important individual differences were found between them. It
appears that mirror-touch synaesthetes can be divided into two subgroups based upon
the spatial mapping between observed and felt (synaesthetic) touch (Figure 1.4).
Some synaesthetes report that an observed touch to the left cheek is felt on their right
cheek (as if the other person is a mirror reflection of oneself – hereafter referred to as
the ‘specular’ subtype) whereas others report synaesthetic touch on their left cheek
when observing touch to another person’s left cheek (as if self and other share the
same anatomical body space – hereafter referred to as the ‘anatomical’ subtype). The
automaticity of these experiences was confirmed by the development of a visuo-
tactile synaesthetic stroop experiment. In the task synaesthetes and matched non-
synaesthetic controls were asked to detect a site touched on their own body (either
SI
SI
Premotor Cortex STS
Anterior Insula
SII SII
Anterior Insula

Chapter 1
31
facial cheeks or hands) while observing touch to another person’s cheek/hands or to a
corresponding object. Participants were asked to report the site of actual touch (left,
right, or no touch) and to ignore observed touch. For synaesthetes, but not controls,
observed touch to humans elicited a tactile sensation whose location was either in the
same position as actual touch (congruent condition – as determined by synaesthetic
self reports) or in a different spatial location (incongruent condition). Synaesthetes,
but not control participants, were faster at detecting the location of actual touch during
the congruent condition relative to the incongruent condition (Figure 1.5b). Further,
synaesthetes produced a higher percentage of errors consistent with their synaesthesia
(hereafter referred to as mirror-touch errors; i.e. reporting touch on trials involving no
actual touch) compared to other error types and to control participants (Figure 1.5c).
Figure 1.4 Specular and anatomical spatial mappings reported by mirror-touch synaesthetes (c.f. Banissy and Ward, 2007). Under a specular frame of reference, mirror-touch synaesthetes report synaesthetic touch as if looking in a mirror. Under an anatomical frame of reference synaesthetic experience is as if self and other share the same anatomical body space.

Introduction 32
300
350
400
450
500
550
600
650
700
750
800
850
Mirror touch Controls
Reaction time (ms)
Congruent
Incongruent
b.
*
0
2
4
6
8
10
12
14
16
18
20
22
24
26
28
30
Percentage of errors
Mirror-touch errors
Other errors
c.
*
*
Mirror-Touch Synaesthetes Non-synaesthetes
Figure 1.5 Summary of Banissy and Ward (2007). (a) Task Protocol. Participants were required to report the site upon which they were actually touched (i.e. left cheek, right cheek, both cheeks or no touch) while ignoring observed touch (and the synaesthetic touch induced from it). Note that although the example given is for a specular mirror-touch synaesthete, both subtypes were tested and congruency was determined according to each synaesthetes’ self-reports. (b) Mirror-touch synaesthetes, but not controls, were significantly faster at detecting the site of real touch in the congruent relative to incongruent condition. (c) Mirror-touch synaesthetes also produced significantly more mirror-touch errors than controls (errors consistent with their synaesthesia), but not other error types. * = p <.05.

Chapter 1
33
1.5 Synaesthesia and models of typical cognition
The preceding sections reviewed evidence for the authenticity of synaesthesia.
While this is now well established, there is growing interest in using the condition to
inform us about non-synaesthetic perceptual and cognitive processing. Following the
logic of cognitive neuropsychology, the positive symptoms related to synaesthesia
may be able to constrain theories on multisensory interactions and inform about the
relationship between multisensory processing and other aspects of cognition (Ward
and Mattingley, 2006; Cohen Kadosh and Henik. 2007).
So far, a number of examples have been cited whereby synaesthetic
interactions have been shown to rely upon similar neurocognitive mechanisms as
those observed in non-synaesthetes and therefore may inform us about general
principles of multisensory interactions (e.g. feature binding in grapheme-colour
synaesthesia; cross-modal interactions in touch-colour synaesthesia; heightened
visual-tactile interactions in mirror-touch synaesthesia). Non-random associations,
which are similar to those found in non-synaesthetic subjects, have also been found
between pitch and lightness in tone-colour synaesthetes (individuals who experience
colour sensations in response to tones) – both synaesthetes and non-synaesthetes show
a tendency to associate low pitches with dark colours and high pitches with light
colours, although only synaesthetes experience these colours consciously (Ward et al.,
2007; also see Parise and Spence, 2009). Evidence of non-random associations in
other variants have also been documented, including number and lightness in digit-
colour synaesthesia (Cohen Kadosh and Walsh, 2008); word form properties and
colour associations in linguistic-colour synaesthesia (Barnett, Feeney, Gormley, and
Newell, 2009); and phonology and tastes in lexical-gustatory synaesthesia (Ward and
Simner, 2003).

Chapter 1
34
Further, in the case of feature binding in synaesthesia, it has been suggested
that an integration of synaesthesia and patient-based research may contribute to our
understanding of the binding problem – how two independently processed features are
combined to be perceived as a unified experience (Robertson, 2003). In the
numerical domain, digit-colour synaesthesia and spatial number-form synaesthesia
have been successfully used as models to make inferences about the mental
representation of two-digit numbers (Seron, Pesenti, Nöel, Deloche, and Cornet, 1992;
Sagiv et al., 2006), the spatial representation of number (Sagiv et al., 2006), and
whether numerical representations are compressed or linear (Cohen Kadosh et al.,
2007).
One aim of this thesis is to use mirror-touch synaesthesia as a model to make
inferences about the role of visual-tactile interactions in cognition and perception. As
noted previously, mirror-touch synaesthesia is thought to arise because of hyper-
activation of the same cortical network (the mirror-touch system) which is active in
non-synaesthetes when observing touch to others (Blakemore et al., 2005). In recent
years there has been much interest in the role brain systems with mirror properties
(e.g. the mirror-touch system) may play in social cognition (Gallese, Keysers, and
Rizzolatti, G, 2004; Gallese, 2006; Keysers and Gazzola, 2006). Moreover, it has
been suggested that brain systems with mirror properties (i.e. common brain regions
in the experience and observation of a particular sensation) may act as a
neurophysiological candidate to facilitate sensorimotor simulation of another’s
experience and thereby promote an understanding of another’s emotions / experience
(Gallese, Keysers, and Rizzolatti, G, 2004; Gallese, 2006; Keysers and Gazzola,
2006). Given that mirror-touch synaesthesia has been linked to neural mechanisms
common to us all when observing touch to another person (i.e. hyper activity within

Chapter 1
35
the tactile mirror system), this variant of synaesthesia highlights one means in which
synaesthesia may be used to investigate vision-touch interactions more generally –
namely what is the impact of heightened sensorimotor simulation on affective
processing. Moreover, mirror-touch synaesthesia is currently one of the only forms of
synaesthesia which depends upon interpersonal interaction and therefore offers a
unqiue opportunity to assess mechanisms of social perception.
1.6 Aims of thesis
This thesis has two primary aims. The first is to investigate cases of mirror-
touch synaesthesia and to document neurocognitive and perceptual profiles associated
with the condition. This includes investigations into the prevalence, characteristics,
and perceptual processing of mirror-touch synaesthesia. The second is to investigate
the function of sensorimotor simulation mechanisms (thought to underpin mirror-
touch synaesthesia) in cognition. This aspect of the thesis aims to evaluate the
importance of somatosensory resources for social cognition and examine the
hypothesis that sensorimotor simulation is critical for understanding the emotions and
thoughts of others (Adolphs, 2002; Adolphs, 2003; Gallese, Keysers, and Rizzolatti,
G, 2004; Gallese, 2006; Gallese and Goldman, 1998; Keysers and Gazzola, 2006;
Oberman and Ramachandran, 2007). Studies involving non-synaesthetic individuals
and studies using synaesthetic participants to inform us about the role of sensorimotor
simulation in affective processing shall be presented.

Chapter 2
36
CHAPTER 2: PREVALENCE AND CHARACHTERISTICS OF
MIRROR-TOUCH SYNAESTHESIA
In so-called ‘mirror-touch synaesthesia’, observing touch to another person induces a
subjective tactile sensation on the synaesthete’s own body. It has been suggested that
this type of synaesthesia depends on increased activity in neural systems activated
when observing touch to others. This is the first study on the prevalence of this variant
of synaesthesia. The findings indicate that this type of synaesthesia is just as
common, if not more common than some of the more frequently studied varieties of
synaesthesia such as grapheme-colour synaesthesia. Additionally, behavioural
correlates associated with the condition are examined further. In a second
experiment, it is shown that synaesthetic experiences are not related to somatotopic
cueing - a flash of light on an observed body part does not elicit the behavioural or
subjective characteristics of synaesthesia. Finally, a neurocognitive model to account
for these characteristics is proposed and the implications of the findings are discussed
in relation to general theories of synaesthesia.
2.1 Introduction
As noted in chapter 1, the term synaesthesia is used to describe a condition in
which one property of a stimulus (the inducer) results in conscious experiences of an
additional attribute (the concurrent). This inducer-concurrent relationship can occur
either within or between modalities. For example, in grapheme-colour synaesthesia a
visually or auditorily presented grapheme can result in synaesthetic experiences of
colour (Ramachandran and Hubbbard, 2001; Cohen Kadosh and Henik, 2007; Rich
and Mattingley, 2002), whereas in lexical-gustatory synaesthesia written or heard
words trigger a subjective sensation of taste (Ward and Simner, 2003).
Early research on the prevalence of synaesthesia indicated that the condition
may have a minimum prevalence rate of 1 in 2000 with a female-to-male ratio of 6:1
(Baron-Cohen, Burt, Smith-Laittan, Harrison, and Bolton, 1996; Rich, Bradshaw, and
Mattingley, 2005). These studies assessed the prevalence of the condition based upon
the number of respondents to newspaper advertisements who pass an objective
measure of synaesthesia (relative to newspaper circulation figures). This method of
assessment does not permit inferences about non-responders and may also lead to an

Chapter 2
37
over inflated female to male ratio. More recent studies, which overcome these
difficulties by screening a large population and supplementing this with the use of
objective measures of different variants of synaesthesia, suggest a higher prevalence
rate of 4% and a female to male ratio of 1:1 (Simner et al., 2006; Ward and Simner,
2005). These studies indicate that the most common forms of the condition include
day-colour synaesthesia (estimated to have a prevalence of 2.8%; Simner et al., 2006)
and grapheme-colour synaesthesia (estimated to have a prevalence of 2%; Simner et
al., 2006).
Since these studies, a new variant of synaesthesia has been documented in
which observing touch to another person induces a tactile sensation on the
synaesthete’s own body (mirror-touch synaesthesia). A single case fMRI study
suggests that this variant of synaesthesia is a consequence of increased neural activity
in a network of brain regions which are also activated in non-synaesthetic control
subjects when observing touch to another person (Blakemore, Bristow, Bird, Frith,
and Ward, 2005). In that study, the authors contrasted brain activity in a single
mirror-touch synaesthete with twelve non-synaesthetic control subjects while
observing humans relative to objects being touched. This indicated that while similar
brain regions were active in the observed touch condition as when participants were
touched (a mirror-touch system present in non-synaesthetes), the synaesthete showed
increased activity within bilateral primary somatosensory cortex (SI), secondary
somatosensory cortex (SII), left premotor cortex and additional activity in the anterior
insula relative to non-synaesthetes. In view of this, it was argued that mirror-touch
synaesthesia reflects hyper-activation of normal (i.e. non-synaesthetic) visual-tactile
interactions in the mirror-touch network (i.e. SI, SII, premotor cortex). Notably, the
general role of SI activations in the mirror-touch system in non-synaesthetes remains

Chapter 2
38
to be clarified, with some authors reporting SI activity when non-synaesthetes observe
touch to another’s face (Blakemore et al., 2005) or arm (McCabe, Rolls, Bilderbeck,
and McGlone, 2008), others reporting SII, but not SI, activation following observed
touch to the legs (Keysers, Wicker, Gazzola, Anton, Fogassi, and Gallese, 2004), and
others reporting SI activity when non-synaesthetes observe intentional but not
unintentional touch (Ebisch, Perrucci, Ferretti, Del Gratta, Romani, and Gallese,
2008).
Extending the single case report, a group study of ten mirror-touch
synaesthetes showed that individuals with mirror-touch synaesthesia can be divided
into two subtypes based upon the spatial mapping between observed and
synaesthetically induced touch. Some synaesthetes report a spatial mapping as if
looking in a mirror (i.e. observed touch to another person’s left cheek induces
synaesthetic touch on their right cheek - specular subtype), while others report a
spatial mapping as if self and other share the same anatomical body space (i.e.
experiencing synaesthetic touch on their left cheek when observing touch to another
person’s left cheek – anatomical subtype; (Banissy and Ward, 2007).
Authenticity and characteristics of synaesthesia
When considering the prevalence of mirror-touch synaesthesia it is important
to note what constitutes synaesthesia in general and the methods used to confirm the
authenticity of the condition. Synaesthesia is typically considered as having three
defining features; 1) experiences are conscious perceptual or percept-like experiences;
2) experiences are induced by an attribute not typically associated with that conscious
experience; 3) these experiences occur automatically (Ward and Mattingley, 2006).
In line with this, mirror-touch synaesthesia requires the conscious experience of a
tactile stimulus which occurs automatically following the observation of touch to

Chapter 2
39
another person (or possibly an object; see Banissy and Ward, 2007). There are
several ways to determine the validity of mirror touch synaesthetes, for example, with
regards to automaticity, Banissy and Ward (2007) developed a visuo-tactile congruity
experiment to explore this aspect of synaesthesia (for description see Chapter 1; also
see Blakemore et al., 2005 methods to assess validity mirror-touch synaesthesia).
Synaesthesia has a number of other important characteristics that also appear
to be found in the mirror-touch variety. Synaesthetic experiences tend to be
consistent over time (e.g. if ‘A’ is red at time 1 then it will be at time 2 several weeks
or months later; Baron-Cohen, Wyke, and Binnie, 1987). Mirror-touch synaesthetes
report their experiences to be enduring and an individual’s spatial sub-type (i.e.
whether they belong to the specular or anatomical category) is consistent both across
time and across different body parts. Further, whilst it was once believed that
synaesthetic experiences reflect random but consistent associations this view is no
longer widely held. For example, non-random associations have been found between;
pitch and lightness (Ward, Huckstep, and Tsakanikos, 2006); number and lightness
(Cohen Kadosh, Henik, and Walsh, 2007); grapheme frequencies and colour (Simner
et al., 2005); and phonology and tastes (Ward and Simner, 2003). More overt
semantic links are also found: it is not uncommon for the word “sausage” to taste of
sausage (and similarly for other food names; Ward, Simner and Auyeung 2005) or for
the word “red” to be coloured red (and similarly for other colour names; Gray et al.,
2002; Rich et al., 2005). The mappings in mirror-touch synaesthesia are non-arbitrary
in that somatotopy is generally preserved between the observed and felt touch.
Here two studies investigating the prevalence and the characteristics of mirror-
touch synaesthesia are presented. In Experiment 1, the prevalence of mirror-touch
synaesthesia is investigated by screening a large population and confirming self

Chapter 2
40
reports using a behavioural paradigm designed to test for the authenticity of the
condition. Then potential factors which may contribute to the behavioural correlates
observed are addressed. Experiment 2 examines the nature of the synaesthetic inducer
and considers the role of somatotopic cueing on synaesthetic experience. Finally, the
factors which may underpin synaesthetic experience are discussed and a
neurocognitive model of mirror-touch synaesthesia is outlined.
2.2 Experiment 1: Prevalence of mirror-touch synaesthesia
This study investigates the prevalence of mirror-touch synaesthesia and
compares new cases with previously studied cases of mirror-touch synaesthesia
(Banissy and Ward, 2007) to ascertain the main cognitive characteristics of the
condition.
Method
All participants (n = 567) were recruited from the University College London
and University of Sussex undergraduate communities. Each participant was given a
written and verbal description of synaesthesia including examples of what did and did
not constitute synaesthesia. Participants were then administered a questionnaire
asking about different variants of synaesthesia with one question specifically related
to mirror-touch synaesthesia (Appendix 1). Participants were asked to indicate on a
five point scale the extent to which they agreed with the question “Do you experience
touch sensations on your own body when you see them on another person’s body?”
Following initial screening, all participants who gave positive responses to the above
question (n = 61; approximately 10.8% of all subjects) were contacted and
interviewed about their experiences. This included them being shown a series of
online videos showing another person, object, or cartoon face being touched.

Chapter 2
41
Participants were asked to indicate the location (if any) in which they experienced a
tactile stimulus and the type of experience. Typical responses of potential mirror-
touch synaesthetes (n = 14; approximately 2.5% of all subjects) included reports that
observing touch elicits a tingling somatic sensation in the corresponding location on
their own body, and that a more intense and qualitatively different sensation is felt for
painful stimuli (i.e. videos of a pin pricking a hand rather than observed touch to the
hand).
In an attempt to investigate reports of mirror-touch synaesthesia the
performance of each potential synaesthete was compared to ten age and gender
matched non-synaesthetic control subjects on the paradigm developed by Banissy and
Ward (2007). In the task, participants were required to detect a site touched on their
own face (left, right, both or none) while observing touch to another person’s face or
to a corresponding object (a lamp). For synaesthetes, but not for controls, observed
touch elicited a synaesthetic sensation in a congruent or incongruent location as actual
touch (Figure 2.1). The tactile stimuli were administered via two miniature solenoid
tappers attached to the face with a Velcro strap. Each tapper was controlled using a
Dual Solenoid Tapper Controller (MSTC3-2, M and E Solve) and the intensity of taps
was filtrated to account for individual sensitivity of the participant (as in Banissy and
Ward, 2007). The visual stimuli were presented on a 17” CRT monitor with a refresh
rate of 100Hz, were sized to fit the screen, and consisted of two presentations of 100
ms each followed by a third stimulus which remained on the screen until the
participant responded. The first two stimuli showed the approach of the hand towards
the face and the third showed contact with the face. After a 10 ms presentation of the
final slide participants received a tap to either their left, right or both cheeks. The
location of the felt touch (left, right, both or none) was indicated with a button press

Chapter 2
42
and the need for both speed and accuracy was emphasised. Following this, there was
a gap of 1500 ms with a fixation cross before the start of the next trial. A train of
white noise was presented via headphones for the duration of each trial in order to
prevent participants from using auditory cues (i.e. the sound of the taps) to determine
the location of actual touch (c.f. Banissy and Ward, 2007 for more details on task
methodology) .
A total of 80 congruent trials, 80 incongruent trials and 80 trials involving no
actual touch were completed. For each potential synaesthete, congruency was
determined according to self reports when observing videos showing another person
being touched. Within each condition, 60 trials involved observed touch to either a
female or male actor, with the remaining 20 trials involving observed touch to a
corresponding object. The order of trials was randomised over 3 blocks of 80 trials
(preceded by 5 practice trials). Reaction times and error rates were measured. Based
upon previous findings synaesthetes were expected to be faster at identifying a site
touched in the congruent compared to incongruent condition and / or to show a higher
proportion of mirror-touch errors compared to non-synaesthetic controls. The control
data were scored according to the reported sub-type of the corresponding mirror-touch
synaesthete (i.e. anatomical versus specular congruency).

Chapter 2
43
Figure 2.1 (a) Summary of the task used to confirm potential cases of mirror-touch synaesthesia in experiment 1. Participants were asked to detect the site of real touch while observing another person being touched. For mirror-touch synaesthetes observed touch elicited a tactile sensation which could either be in a congruent or incongruent location as the site real touch. (b) Example of congruent and incongruent trials including error types for a specular mirror-touch synaesthete. On a congruent trial, real touch was applied to the same side of the face as synaesthetic experience. On an incongruent trial real touch was applied to the side of the face which was opposite to synaesthetic experience. Participants were asked to report the location of real touch and to ignore synaesthetic touch. ‘Mirror-touch’ errors could be produced on incongruent trials if the subject was to report real touch to both cheeks (despite real touch being applied to one cheek only) or if the subject was to report synaesthetic rather than real touch. All other error types were classified as ‘Other’ error types.

Chapter 2
44
Results and Discussion
Behavioural performance of each potential synaesthete was compared to an
age and gender matched non-synaesthetic control group using Crawford’s modified t-
test (Crawford and Garthwaite, 2002). Reaction time performance (filtered prior to
analysis, ± 3 s.d. and all errors removed) and the percentage of error types on human
and object trials were compared separately (Table 2.1). For reaction time
performance, the size of congruency effect (incongruent minus congruent reaction
time) was used as an index of synaesthetic experience. For errors, the percentage of
mirror-touch errors (errors consistent with synaesthetic experience) and other error
types were compared. Subjects who showed either significantly larger reaction time
differences or significantly more mirror-touch errors relative to controls were counted
as synaesthetes. Using this method nine cases (seven female)1 of mirror-touch
synaesthesia were confirmed on reaction time performance, the percentage of mirror-
touch errors produced, or both (Table 2.1). This indicates a prevalence rate of 1.6%.
In comparison to previous prevalence estimates of other types of synaesthesia this
places mirror-touch synaesthesia as one of the most common forms of synaesthesia
along with grapheme-colour synaesthesia (1.4% prevalence) and day-colour
synaesthesia (2.8% prevalence; Simner et al., 2006).
Comparison of the prevalence group with previously reported cases
In order to ensure that these cases were consistent with previously reported
cases of mirror-touch synaesthesia, these characteristics of synaesthetic experience
were considered further by contrasting synaesthetes recruited through the prevalence
study (n = 9) with mirror-touch synaesthetes recruited via self referral including some
1 Participants came from courses with a higher female-male ratio and gender information for non-respondants was not available, so no empirical claims about female-male ratio of mirror-touch synaesthetes are made.

Chapter 2
45
previously reported cases (n = 12). Reaction time (Congruency x Group) and the
percentage of error types (Error Type x Group) were compared separately using a 2 x
2 ANOVA (Figure 2.2a, 2.2b – for comparison non-synaesthetic control data, n = 20,
is also shown, but not included in analysis). One participant from the self referral
group was withdrawn from analysis of reaction times due to an insufficient number of
correct responses (< 25% correct responses in any one condition).
Analysis of reaction time data revealed a significant main effect of
congruency, with subjects performing faster overall on trials which were congruent
with their synaesthesia compared to incongruent trials [F(1, 18) = 13.98, p = < .01].
Analysis of error type data revealed a significant main effect of error type, which was
due to a higher proportion of mirror-touch errors being produced relative to other
error types [F(1, 19) = 11.18, p = < .01]. No significant interaction or main effect of
group was found for reaction time (Group: [F(1, 18) = .048, p = .829]; Group x Cong:
[F(1. 18) = .095, p = .761]) or error type analysis (Group: [ F(1, 19) = 2.77, p = .113];
Group x Cong: [F(1. 19) = 2.75, p = .114]). This indicates that the prevalence and
self-referred mirror-touch synaesthete group come from the same population with
regard to congruency effects. Therefore, both prevalence and self-referred cases are
combined to consider additional cognitive characteristics of mirror-touch
synaesthesia.
For the majority of cases, the effects of spatial congruity are found for bodies
but not objects and this corresponds well with their phenomenological reports. There
are, however, a minority of synaesthetes who do report tactile experiences when
watching objects being touched (4 out of 21). For some of these synaesthetes, this
experience is reported in the finger tip that is touching the objects, but for others
synaesthetic touch is mapped onto particular body locations which are thought to

Chapter 2
46
spatially correspond to the object being touched (e.g. when looking directly at a
monitor the experience maps onto the face, but when standing in front of the monitor
the experience maps onto the trunk). In addition, another minority of synaesthetes (6
out of 21, including I., Z. and H.G. in Table 2.1) show an effect of spatial congruity
for both bodies and objects despite initially claiming to experience synaesthesia for
touched bodies alone. One possibility is that this reflects the fact that object trials are
interleaved with the more frequent human trials and this leads to objects being treated
more like human bodies than expected. In the normal population, fMRI studies
suggest that the tactile mirror system does respond to objects under some
circumstances (Ebisch et al., 2008; Keysers et al., 2004).
Of the 21 cases of mirror-touch synaesthesia reported to date, seventeen (nine
of which come from the prevalence sample) report a specular frame of reference and
four report an anatomical frame of reference. This finding is consistent with studies
on imitation behaviour which demonstrate that both adults and children tend to imitate
in a specular mode (Schofield, 1976; Franz, Ford, and Werner, 2007). The relative
bias in synaesthetes could be due to the fact that one’s own head is only ever seen
from a mirror-reflected perspective and this regularity may drive the choice of spatial
frame. However, it is to be noted that those synaesthetes who adopt a specular frame
for the head also do so with the hands (Banissy and Ward, 2007) even though this part
of one’s own body is not normally viewed from a reflected perspective.
A general characteristic of synaesthesia is that different variants of
synaesthesia tend to co-occur (Simner et al., 2006). Some preliminary evidence based
upon self reports suggests that this may also be the case with mirror-touch
synaesthetes. While, some mirror-touch synaesthetes only report mirror-touch
synaesthesia (implying that the overall prevalence of synaesthesia may be higher than

Chapter 2
47
the previously assumed 4% estimate), nine of the twenty-one mirror-touch
synaesthetes sampled also report genders or personalities for graphemes and/or certain
other linguistic stimuli (e.g. 3 is a bossy male; Simner and Holenstein, 2007; Smilek,
Malcolmson, Carriere, Eller, Kwan, and Reynolds, 2007). Five of these cases have
been confirmed using behavioural tests for this phenomenon (N. Sagiv, personal
communication). Additionally, seven report synaesthetic experiences of colour for
linguistic stimuli. Notably, this data is preliminary because the sample contains a
mixture of randomly (i.e. prevalence group) and non-randomly sampled participants
(i.e. self-referred group), and because members of the prevalence group were not
systematically tested for other variants of synaesthesia.
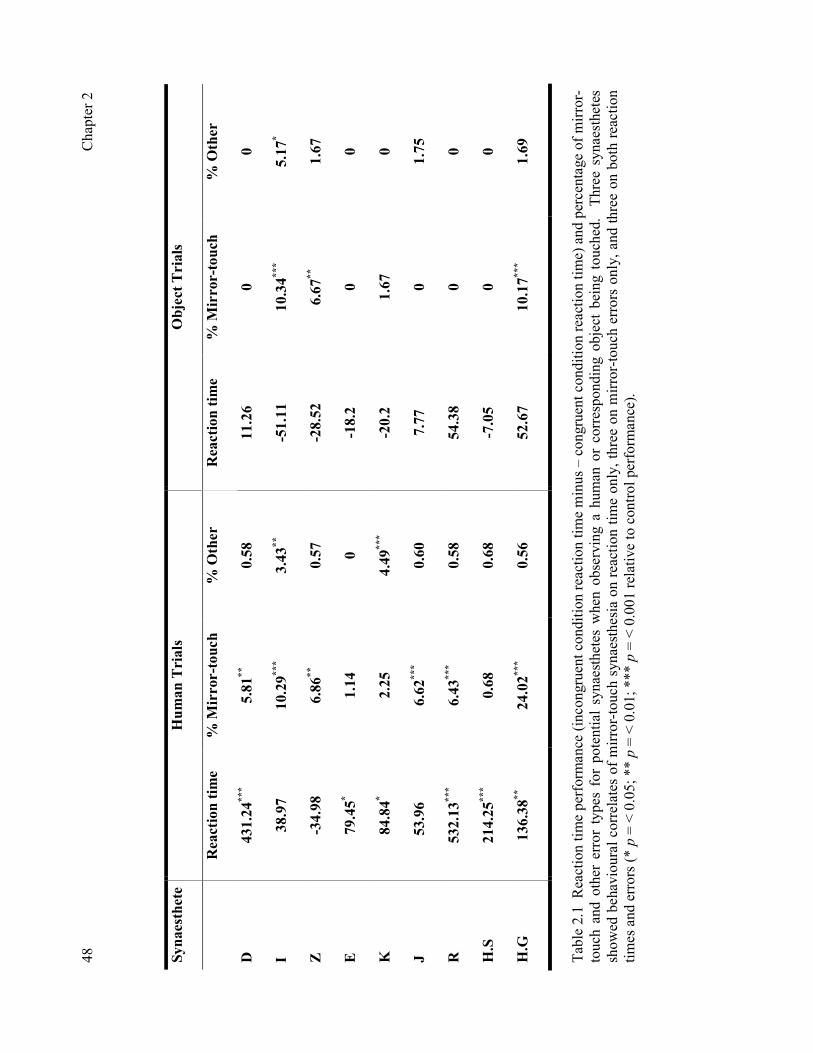
C
hapt
er 2
48 Synaesthete
Human Trials
Object Trials
Reaction time
% Mirror-touch
% Other
Reaction time
% Mirror-touch
% Other
D
431.24***
5.81**
0.58
11.26
0
0
I 38.97
10.29***
3.43**
-51.11
10.34***
5.17*
Z
-34.98
6.86**
0.57
-28.52
6.67**
1.67
E
79.45*
1.14
0
-18.2
0
0
K
84.84*
2.25
4.49***
-20.2
1.67
0
J
53.96
6.62***
0.60
7.77
0
1.75
R
532.13***
6.43***
0.58
54.38
0
0
H.S
214.25***
0.68
0.68
-7.05
0
0
H.G
136.38**
24.02***
0.56
52.67
10.17***
1.69
Tabl
e 2.
1 R
eact
ion
time
perf
orm
ance
(inc
ongr
uent
con
ditio
n re
actio
n tim
e m
inus
– c
ongr
uent
con
ditio
n re
actio
n tim
e) a
nd p
erce
ntag
e of
mir
ror-
touc
h an
d ot
her
erro
r ty
pes
for
pote
ntia
l sy
naes
thet
es w
hen
obse
rvin
g a
hum
an o
r co
rres
pond
ing
obje
ct b
eing
tou
ched
. T
hree
syn
aest
hete
s sh
owed
beh
avio
ural
cor
rela
tes
of m
irror
-touc
h sy
naes
thes
ia o
n re
actio
n tim
e on
ly, t
hree
on
mirr
or-to
uch
erro
rs o
nly,
and
thre
e on
bot
h re
actio
n tim
es a
nd e
rror
s (*
p = < 0
.05;
** p =
< 0
.01;
***
p = < 0
.001
rela
tive
to c
ontr
ol p
erfo
rman
ce).

C
hapt
er 2
49 a.
b.
550
600
650
700
750
800
850
900
950
1000
1050
1100
1150
Reaction Time (msec)
Congruent
Incongruent
Prevalence
Self referral
Control
**
05
10
15
20
25
30
% Error
Mirror-touch
Other
Prevalence
Self referral
Control
*
*
Figu
re 2
.2 M
ean
reac
tion
time
perf
orm
ance
(a)
and
per
cent
age
of e
rror
type
s (b
) on
hum
an tr
ials
for
mir
ror-
touc
h sy
naes
thet
es re
crui
ted
with
in
the
prev
alen
ce s
tudy
com
pare
d to
syn
aest
hete
s re
crui
ted
via
self
-ref
erra
l (co
ntro
l gro
up p
erfo
rman
ce is
sho
wn
for c
ompa
rison
). ±
s.e
.m (*
p =
<
0.05
).

Chapter 2 50
2.3 Experiment 2: Behavioural correlates and somatotopic cueing While the results from experiment 1 establish evidence for the authenticity of
mirror-touch synaesthesia and suggest that behavioural correlates are related to
‘observed bodily touch’, it remains unclear if the behavioural data could also be
consistent with ‘observed bodily cueing’ – whereby an observed visual event cues a
particular location on the body. There is growing evidence from research
investigating visual-tactile interactions that non-informative vision associated with
one’s own body can influence tactile processing (e.g. Johnson, Burton, and Ro, 2006).
In order to establish whether the findings could be related to somatotopic cueing, the
performance of mirror-touch synaesthetes and non-synaesthetic subjects was
compared on a condition in which a human face is observed but is accompanied by a
flash of light on the cheek rather than a touch. As these stimuli did not induce
synaesthesia it was expected that the pattern of effects shown by synaesthetes on
experiment 1 would be related specifically to ‘observed bodily touch’ and that
differences between synaesthetes and non-synaesthete controls would not be found for
‘observed bodily cueing’.
Method
Ten mirror-touch synaesthetes (7 females and 3 male, mean age ± Std. Error =
30.1 ± 11.17 years; 1 anatomical and 9 specular) and ten non-synaesthetic controls
matched for age and gender (7 females and 3 males, mean age ± Std. Error = 31 ±
13.23 years) took part. Congruency was determined according to synaesthetes’ self
reports when observing touch to another person. Controls were randomly allocated to
either a specular or anatomical congruency group to match the synaesthetic group.
The experimental task and procedure was the same as Experiment 1, with the
exception of the stimuli presented (Figure 2.3). For the human trials, rather than

Chapter 2
51
observing touch to the cheek(s), a flash of light appeared on the observed person’s
cheek(s). As before, the visual stimuli consisted of 3 frames which were sized to fit
the screen. The first stimulus, lasting 100 ms, depicted a male or female face. The
second stimulus, also lasting 100 ms, was the same as the first except that a patch of
white light appeared on the person’s left/right/both cheek(s). The flash was then
removed for the third stimulus which remained on the screen until the participant
responded. The tactile stimulus was applied 10 ms after the flash, i.e. at the onset of
the third stimulus. For the control trials, all pictures of the person were replaced by a
blank screen with a 100ms flash of light on the left, right or both sides of space
immediately before the tactile event. A total of 306 trials were completed, of which
180 involved human stimuli and 126 involved control stimuli.
Figure 2.3 Summary of the task used for somatotopic cueing experiment. Participants observed a flash of light on the left/right/or both cheek(s) of another person. Immediately following the light flash, subjects were touched on their own facial cheeks (either left, right or both cheeks). Participants were asked to report the site of real touch.
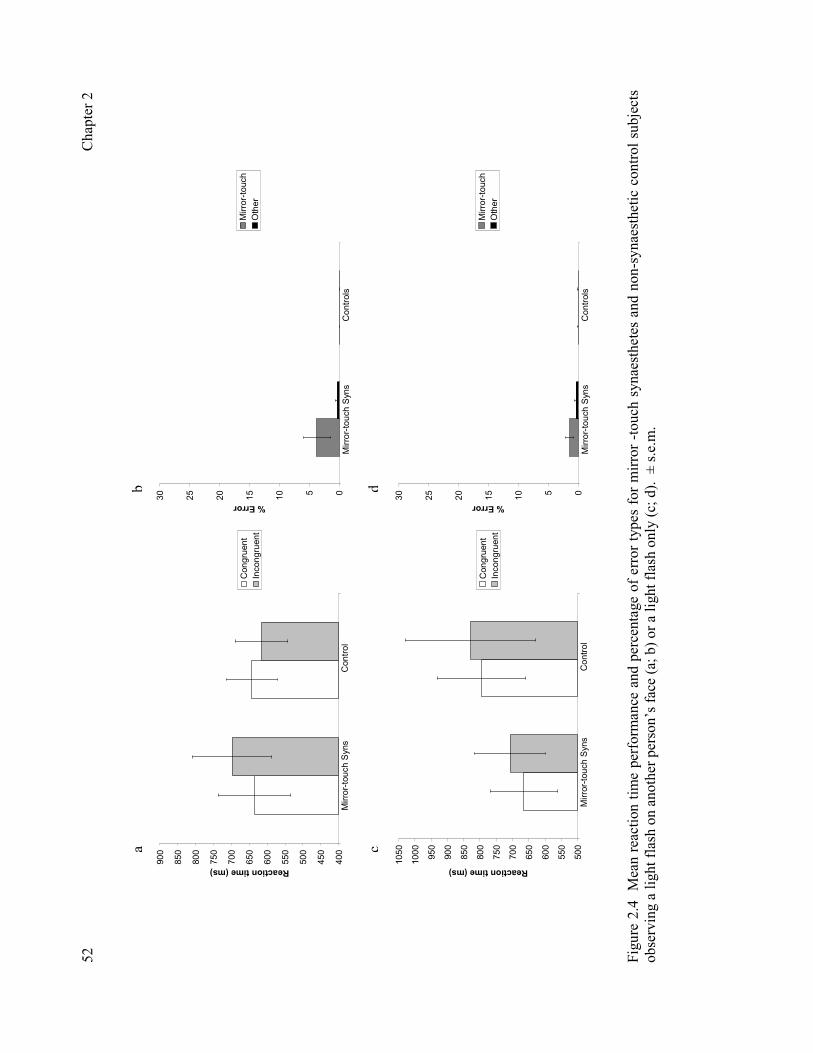
C
hapt
er 2
52
400
450
500
550
600
650
700
750
800
850
900
Reaction time (ms)
Congruent
Incongruent
Mirror-touch Syns
Control
05
10
15
20
25
30
% Error
Mirror-touch
Other
Controls
Mirror-touch Syns
500
550
600
650
700
750
800
850
900
950
1000
1050
Reaction time (ms)
Congruent
Incongruent
Mirror-touch Syns
Control
05
10
15
20
25
30
% Error
Mirror-touch
Other
Controls
Mirror-touch Syns
Figu
re 2
.4
Mea
n re
actio
n tim
e pe
rfor
man
ce a
nd p
erce
ntag
e of
err
or ty
pes
for
mirr
or -
touc
h sy
naes
thet
es a
nd n
on-s
ynae
sthe
tic c
ontro
l sub
ject
s ob
serv
ing
a lig
ht fl
ash
on a
noth
er p
erso
n’s
face
(a; b
) or a
ligh
t fla
sh o
nly
(c; d
). ±
s.e
.m.
a b
c d

Chapter 2 53
Results and Discussion
The results are summarised in Figure 2.4. Reaction times and error rates were
measured. Reaction time data were filtered prior to analysis (± 3 s.d. and all errors
removed). A 2 (Group) x 2 (Congruency) ANOVA conducted on reaction times
revealed no significant main effects or interactions (p = > .05 in all cases; Figure
2.4a). Although the direction of the effect was the same as in Experiment 1 the effect
was not significant. Analysis of the percentage of error types made by participants on
human trials also revealed no significant main effects or interactions (p = > .05 in all
cases; Figure 2.4b). Similarly, no significant differences were observed on control
trials (Figure 2.4c, d). These findings are unlikely to be due to the fact that the flash
of light is less salient than the hand, because the synaesthetes also failed to show an
effect in Experiment 1 when a hand was used on a non-human object.
To further validate that the performance of mirror-touch synaesthetes
significantly differed between experiment 1 and 2 a within-group comparison on the
size of congruency effect (incongruent minus congruent trial reaction time) shown by
synaesthetes across each task was conducted. This revealed that synaesthetes showed
a significantly greater effect of congruency on trials involving observed touch to a
human face in experiment 1 (mean ± s.e.m = 208.24 ± 52.32 msec) compared to a
flash of light shown on a human face in experiment 2 (mean ± s.e.m = 51.49 ± 34.18
msec), [t(9) = 2.98, p = < .02]. Thus the findings from Experiment 1 are related
specifically to ‘observed bodily touch’ and cannot be attributed to somatotopic
cueing.

Chapter 2
54
2.4 General Discussion Taken together, these measures detail the prevalence and characteristics of
mirror-touch synaesthesia. In relation to prevalence, the findings suggest that:
• mirror-touch synaesthesia is one of the more common forms of
synaesthesia
• there are two sub-types (specular and anatomical) depending on the
visuo-tactile spatial transformation used
• the specular (mirror-reflected) sub-type is the more common
• the effects are quite specific to observed touch to a human body.
In many respects, mirror-touch synaesthesia shares common ground with other
types of synaesthesia; for instance, with regards to phenomenology, automaticity,
consistency (of the spatial mapping), reliability over time, and possibly with regards
to associated traits (e.g. attributing personalities and genders to graphemes).
However, when one turns to consider its neural basis the similarities are less apparent.
A current area of debate in the synaesthesia literature is whether synaesthetic
experience is due to cross-activation between brain regions or cortical disinhibition
(Bargary and Mitchell, 2008; Cohen Kadosh, Henik, Catena, Walsh, and Fuetnes,
2009; Cohen Kadosh and Walsh, 2008; Grossenbacher and Lovelace, 2001; Hubbard
and Ramachandran, 2005; Rouw and Scholte, 2007). Thus far, accounts of
synaesthesia in terms of cross-activation have mainly focussed on grapheme-colour
synaesthesia and highlight the role of adjacency between visual grapheme and colour
processing areas in the fusiform gyrus (Ramachandran and Hubbard, 2001). It is
possible that adjacency is one of several biasing principles that influence which forms
of synaesthesia will, and will not, be found. Another biasing principle may be the
‘normal’ architecture for multi-sensory interactions. As noted before, there is now

Chapter 2
55
good evidence for a visuo-tactile mirror system in humans (Blakemore et al., 2005;
Ebisch et al., 2008; Keysers et al., 2004) and mirror-touch synaesthesia could be
construed as hyper-activity within this network (either as a result of cortical
disinhibition or cross-activation).
Below I propose a model of this type of synaesthesia.
A Neurocognitive Model of Mirror-Touch Synaesthesia: What, Who, Where.
In this model, mechanisms thought to underpin synaesthetic experience are
divided into processes involved in identifying the visual stimulus touched (“what”
mechanism – shown in red boxes), discriminating between self and other (“who”
mechanism – shown in blue boxes), and locating where on the body and in space
observed touch occurs (“where” mechanism – shown in green boxes). Connections
between processes common to all subjects are shown in black; connections between
processes necessary for an anatomical reference frame in purple; connections between
processes contributing to a specular reference frame are shown in orange (Figure 2.5).
Visual Encoding: “What” Mechanisms
With regards to the tactile mirror system, the putative “what” mechanisms are
needed to implement several discriminations. Is this a human or object? Is this a face
or body? One potential brain region which may be crucial to human body perception
in mirror-touch synaesthesia is the extrastriate body area (EBA; Downing, Jiang,
Shuman and Kanwisher, 2001). The EBA is a body-selective visual region which
responds more to bodies and body parts, than faces, objects and object parts (Downing
et al., 2001). This is in contrast to the fusiform body area (FBA; Peelen and
Downing, 2005), a further body selective visual region, which appears more important
for processing body parts into wholes (Taylor, Wiggett and Downing, 2007).

Chapter 2
56
Figure 2.5 The ‘What, Who, Where Model of Mirror-Touch Synaesthesia’. ‘What’ mechanisms are shown in red boxes and are involved in defining the stimulus touched. ‘Who’ mechanisms implement discriminations between self and other, and are shown in blue boxes. ‘Where’ mechanisms are shown in green boxes and are involved in locating where on the body and in space observed touch occurs. Processes necessary for all subjects are shown with black arrows, necessary for specular mirror-touch synaesthetes with orange arrows, and for anatomical mirror-touch synaesthetes with purple arrows. Brain regions represented are considered with regard to importance for mirror-touch synaesthesia. AI = Anterior Insula; EBA = Extrastriate Body Area; FBA = Fusiform Body Area; FFA = Fusiform face area; IFG = Inferior Frontal Gyrus; IPL = Inferior Parietal Lobule; IPS = Intraparietal Sulcus; LO = Lateral Occipital Cortex; SI = Primary Somatosensory Cortex; SII = Secondary Somatosensory Cortex; STS = Superior Temporal Sulcus; TPJ = Temporoparietal Junction.
In addition to the EBA, object selective visual regions and their interactions
along higher-order visual systems may then be crucial for distinguishing between
those synaesthetes for whom observing touch to objects elicit synaesthetic interactions
and for those in which no synaesthetic interaction is experienced. In cases where
observing touch to objects evokes synaesthesia, the processing of object information

Chapter 2
57
via the dorsal stream to areas along the medial bank of the intraparietal sulcus (IPS;
Konen and Kaster, 2008) may be particularly important. The IPS forms part of the
tactile mirror system (Blakemore et al., 2005) and is known to contain visual-tactile
body maps which are important for dynamic multisensory body representations
(Bremmer et al., 2001; Duhamel, Colby and Goldberg, 1998; Iriki, Tanaka and
Iwamura., 1996; Macaluso and Driver, 2003; also see Colby, 1998; Maravita and
Iriki, 2002 for review). Therefore the degree to which observing touch to an object is
able to elicit visual-tactile synaesthetic interactions may depend upon the extent to
which the object is incorporated into visual-tactile representations of the body,
potentially within the IPS.
Visual Encoding: “Who” Mechanisms
The most crucial distinction to be made by the putative “who” mechanism is
that between self and other. Is it my body/face that is seen?
One can consider mirror-touch synaesthesia as a breakdown in the
mechanisms that normally distinguish self from other. A dedicated module to
distinguish between self and other is not proposed; rather, this discrimination will
emerge out of other processes involved in linking visual representations with internal
representations of bodies. Namely, there may be a tendency to over-incorporate
viewed bodies within the observer’s current body schema (Coslett, 1998; Gallagher,
1995; Head and Holmes, 1911-1912; Sirigu, Grafman, Bressler, and Sunderland,
1991). This process is likely to depend on a variety of factors: the perspective of the
viewed body part; the current posture of the mirror-touch synaesthete; and the
similarity (facial or otherwise) between the perceiver and perceived.

Chapter 2
58
Figure 2.6 The influence of perspective on synaesthetic experience. (a) Observing touch to another person from one’s own perspective induces touch on an anatomically corresponding hand for both the anatomical and specular subtypes (i.e. observing touch to another’s left hand evokes synaesthesia on the synaesthete’s left hand). (b) For anatomical mirror-touch synaesthetes, synaesthetic touch is still evoked on the anatomically corresponding hand when observing touch to another person’s hand from another’s perspective (i.e. observing touch to another’s left hand evokes synaesthesia on the synaesthete’s left hand). (c) For specular mirror-touch synaesthetes, this not the case. When observing touch to another person’s hand from another’s perspective, synaesthesia is evoked on the mirrored right hand (i.e. observing touch to another’s left hand evokes synaesthesia on the synaesthete’s right hand). See Banissy and Ward (2007). Blue dots correspond to the location of the synaesthetic sensation evoked.
The perspective of the seen body part provides one way of discriminating
between self and other. The importance of discriminations between first-person and
third-person perspectives (Figure 2.6) also varies between synaesthetic subtypes when
observing touch to body parts (excluding the face) and this may require more
computations for specular compared to anatomical synaesthetes. For specular

Chapter 2
59
synaesthetes, touch to the hands from a first-person perspective induces synaesthetic
touch to the anatomically corresponding hand (i.e. right hand to right hand), but from
a third-person perspective induces synaesthetic touch to the mirrored hand (i.e. right
hand to left hand). In contrast, for anatomical synaesthetes, observed touch from
either perspective elicits synaesthetic touch to the anatomically corresponding hand
(c.f. Kusnir, MSc Thesis, University of London, 2008). The response of the right
EBA is greater for body parts in the third-person than first-person perspective (Saxe,
Jamal and Powell, 2006) and this brain region may contribute to this distinction. .
With regards to faces, viewing one’s own face activates a different network of
brain regions from other faces including famous or personally familiar ones (Uddin,
Iacoboni, Lange, and Keenan, 2007). FMRI research has highlighted the role of a
right-fronto-parietal network in this process, including the right inferior parietal lobule
(IPL) and right inferior frontal gyrus (IFG; Sugiura, Watanabe, Maeda, Matsue,
Fukuda, and Kawashima, 2005; Uddin, Kaplan, Molnar-Szakacs, Zaidel, and
Iacoboni, 2005). These two regions form part of the classical mirror neuron system in
humans (Rizzolatti and Craighero, 2004) and it has been suggested that they may be
necessary to not only establish shared representations, but also to implement
mechanisms to distinguish between self and other (Uddin, Molnar-Szakacs, Zaidel,
and Iacoboni, 2006). It may be the case that this same sensorimotor network is over-
active in mirror-touch synaesthetes when viewing faces other than their own, causing
the body part to be incorporated into the observer’s own body representations. One
prediction is that mirror-touch synaesthetes (at least the specular sub-type) will show
little behavioural or phenomenological differences on the spatial congruity task used

Chapter 2
60
here if the unfamiliar faces were replaced with images of their own faces2. However,
controls may begin to show similar behavioural performance to the synaesthetes if
images of their own face are displayed. In accordance with this, Serino and
colleagues (2008) report that, for non-synaesthetes, observing touch to one’s own or
another’s face increases tactile sensitivity on the observers own face (also see
Haggard, 2006 for similar evidence of interpersonal enhancements of touch). This
visual-tactile enhancement was maximised when observing touch to one’s own face
rather than another’s face, indicating that self-similarity can modulate the extent of
visuo-tactile resonance (Serino, Pizzoferrato, and Làdavas, 2008).
Perspective Taking: “Where” Mechanisms
The third class of mechanism that is considered to be relevant involves linking
visual representations of body with tactile representations based on proprioception
and somatosensation. One distinction that has been made in the literature is between
“embodied” and “disembodied” representations of body (Giummarra, Gibson,
Georgiou-Karistianis, and Bradshaw, 2007; also see Brugger, 2002 for a discussion of
similar spatial aspects of autoscopic phenomena). Evoked potential mapping
indicates that the right temporoparietal junction (TPJ) is related to disembodied
perspective taking (judging left/right from someone else’s perspective), while left
EBA activation is linked with embodied perspective taking (judging left/right from
own perspective; Arzy, Thut, Mohr, Michel and Blanke, 2006). Moreover,
stimulation of the TPJ has been shown to lead to disembodied experiences in
neurological patients (Blanke, Landis, Spinelli and Seeck, 2004; Blanke, Ortigue,
Landis and Seeck, 2002).
2 The predictions for synaesthetes with the anatomical sub-type are unclear because their usual synaesthetic phenomenology would contradict their own prior experiences of observing their own face in a mirror (e.g. when shaving or putting on make-up).

Chapter 2
61
This distinction is similar to the specular-anatomical division between mirror-
touch synaesthetes. For the specular sub-type, the visual representation of the other
body is spatially processed as if it is a mirror-image of one’s own (embodied) body.
For the anatomical sub-type, the spatial mapping is more disembodied in that one’s
own body is placed in the perspective of the other person (or one’s own body and that
of the other person are copied into some other shared bodily template). If this is the
case, it makes a specific and testable prediction - namely, that the anatomical sub-type
will be associated with greater activity in the TPJ than the specular sub-type.
Somatosensory Processes
A final component within the model is the role of somatosensation in mirror-
touch synaesthesia. Previous fMRI findings indicate that the condition is linked with
increased activations in SI, SII and additional activations in bilateral anterior insula
(Blakemore et al., 2005). The specific role of these regions in the experience of
synaesthetic touch remains unclear. For example, the anterior insula is connected
with both somesthetic cortex and visual association areas (Mesulam and Mufson,
1982; Mufson and Mesulam, 1982) which may make this brain region a potential
candidate for accounts of mirror-touch synaesthesia in terms of mechanisms of
disinhibition or hyper-connectivity. This brain region also contains tactile receptive
fields in the absence of activations of primary somatosensory cortices (Olausson et al.,
2002) and is important in processing the affective consequences of touch (Craig,
2002). In this sense, anterior insula activations observed in mirror-touch synaesthesia
may reflect processing of tactile and affective consequences of synaesthetic
experience; self reports indicate that the synaesthetic tactile sensation varies with the
type of touch observed (i.e. pain versus touch) and has differing affective
consequences accordingly. Alternatively, the anterior insula is also important in

Chapter 2
62
distinguishing between self and other (Fink, Markowitsch, Reinkemeier, Bruckbauer,
Kessler, and Heiss, 1996; Kircher et al., 2001; Ruby and Decety, 2001) and this
region could be involved in misattributing observed touch to oneself through
mechanisms of self-other discrimination (Blakemore et al., 2005). The use of brain
imaging to investigate more closely the interactions between activations in the
anterior insula and primary somatosensory cortices observed in mirror-touch
synaesthesia may shed light on these issues.
Summary
In summary, by investigating the prevalence and characteristics of mirror-
touch synaesthesia it has been shown that this variant of the condition may be one of
the most common forms of synaesthesia. Furthermore, there are a number of
important characteristics which indicate that the condition goes beyond a simple one
to one mapping between observed and synaesthetic touch. A neurocognitive model is
proposed (Figure 2.5), which distinguishes between subtypes of mirror-touch
synaesthesia and suggest potential neural mechanisms to account for how differences
in the interpersonal body maps adopted may lead to different cognitive processes
related to synaesthetic experience.

Chapter 3
63
CHAPTER 3: SENSORY PROCESSING IN SYNAESTHESIA
The studies presented in chapter 2 explored the prevalence and characteristics of
mirror-touch synaesthesia. Here I investigate the perceptual characteristics
associated with the presence of mirror-touch synaesthesia. Previous findings imply
that synaesthetic experience may have consequences for sensory processing of stimuli
that do not themselves trigger synaesthesia. For example, synaesthetes who
experience colour show enhanced perceptual processing of colour compared to non-
synaesthetes. This study aimed to investigate whether enhanced perceptual
processing was a core property of synaesthesia by contrasting tactile and colour
sensitivity in synaesthetes who experience either colour, touch, or both touch and
colour as evoked sensations. For comparison the performance of non-synaesthetic
control subjects was also assessed. There was a relationship between the modality of
synaesthetic experience and the modality of sensory enhancement. Synaesthetes who
experience colour have enhanced colour sensitivity and synaesthetes who experience
touch have enhanced tactile sensitivity. These findings suggest the possibility that a
hyper-sensitive concurrent perceptual system is a general property of synaesthesia
and are discussed in relation to theories of the condition.
3.1 Introduction
As noted previously, synaesthesia is a developmental condition in which one
property of a stimulus results in conscious perceptual or ‘percept like’ experiences of
an additional attribute. The authenticity of the condition is now well established (for
reviews see Cohen Kadosh and Henik, 2007; Hubbard and Ramachandran, 2005; Rich
and Mattingley, 2002) and a number of psychophysical studies indicate that
synaesthetic experience resembles veridical sensory perception (but see Hong and
Blake, 2008), e.g. in grapheme-colour synaesthetes, synaesthetic and real colours
interact in binocular rivalry (Kim, Blake, and Palmeri, 2006); can induce orientation
contingent colour adaption after-effects such as a synaesthetic ‘McCollough Effect’
(Blake, Palmeri, Marois, and Kim, 2004); and can combine to produce apparent
motion (Kim et al., 2006). Under certain circumstances, synaesthetic experience may
also aid veridical sensory perception. Vision-sound synaesthetes (synaesthetes for
whom seeing visual motion triggers auditory perception) show an advantage at

Chapter 3
64
perceiving visually presented rhythmic patterns compared to non-synaesthetes.
Typically, non-synaesthetes are superior at recognising auditory compared to
equivalent visual rhythmic patterns, but vision-sound synaesthetes recode the visual
information aurally leading to superior visual rhythm perception (Saenz and Koch,
2008).
Recent ERP evidence indicates that the presence of synaesthesia may also
exert a wider influence over veridical sensory processing (Barnett et al., 2008; Goller,
Otten, and Ward, 2009; Yaro and Ward, 2007). Barnett and colleagues (2008) report
that, compared to non-synaesthetes, linguistic-colour synaesthetes show differences in
early components of the visual evoked potential (VEP) when presented with simple
visual stimuli which do not evoke synaesthesia. VEP differences were observed
following the presentation of high spatial frequency Gabor patches which
preferentially activate the parvocellular pathways (pathways highly responsive to
colour; Kaplan, 1991) of the visual system (Barnett et al., 2008). Goller et al. (2009)
report similar early VEP differences between tone-colour synaesthetes and non-
synaesthetic controls following the presentation of coloured visual stimuli. These
findings indicate electrophysiological differences in two groups of synaesthetes for
stimuli which do not themselves trigger synaesthetic experience, implying that
synaesthesia may be linked with general differences in veridical sensory perception.
Behavioural correlates of this may include enhanced perceptual processing for stimuli
related to synaesthetic experience. For example, Yaro and Ward (2007) report that
synaesthetes who experience colour show better perceptual discrimination of colour
relative to non-synaesthetic control subjects.
Although synaesthetic experiences are nearly always unidirectional in terms of
conscious experience (e.g. graphemes or sounds trigger colour but not vice versa)

Chapter 3
65
there is good evidence of implicit bidirectionality (Cohen Kadosh and Henik, 2006;
Cohen Kadosh, Cohen Kadosh and Henik, 2007). It is possible that previously
reported sensory enhancements in colour processing (Yaro and Ward, 2007) are not
localised within the colour domain but reflect back-coding, e.g. into the verbal
domain. As such it is important to test other varieties of synaesthesia in which this
possibility is less likely. For example, ‘mirror-touch’ synaesthetes experience tactile
sensations on their own body when observing touch to another person (Banissy and
Ward, 2007; Blakemore, Bristow, Bird, Frith, and Ward, 2005). To address this, the
current study investigates tactile and colour perception in three groups of synaesthetes
and a group of non-synaesthetic control subjects. The experiment was a 2x2 between
subjects design in which I contrasted presence/absence of mirror-touch synaesthesia
with presence/absence of synaesthetic colour experiences (including but not limited to
grapheme-colour synaesthesia). The group reporting an absence of both types of
synaesthesia are termed ‘normal’ control group. The three synaesthetic groups either
have both types (hereafter referred to as dual-synaesthetes), or only one of these types.
Based on previous research synaesthetes were expected to show enhanced perceptual
sensitivity, but it remains to be shown whether this is specific to the modality (or
modalities) that participate in the synaesthesia.
3.2 Methods
Participants
The touch-synaesthete group were comprised of six mirror-touch synaesthetes
who do not experience any other forms of synaesthesia (4 female and 2 male; mean
age ± s.e.m = 35.5 ± 3.93 years). Eight mirror-touch synaesthetes who also experience
some form of colour synaesthesia (i.e. grapheme-colour, digit-colour, tone-colour)
formed the dual-synaesthete group (6 female and 2 male; mean age ± s.e.m = 39.14 ±

Chapter 3
66
3.74 years). Eight synaesthetes who experience synaesthetic perceptions of colour
only (7 digit-colour synaesthetes and 1 letter-colour synaesthete who also reports
digit-colour) were recruited for the colour-synaesthete group (7 female and 1 male;
mean age ± s.e.m = 30.83 ± 3.01 years). None of the colour synaesthetes reported that
colour elicited synaesthesia (e.g. colour-to-sound). Additionally, 20 non-synaesthetic
control subjects were recruited for the experiment (16 female and 4 male; mean age ±
s.e.m = 31.4 ± 3.66 years).
Cases of mirror-touch synaesthesia were confirmed on a visual-tactile spatial
congruity paradigm designed to provide evidence for the authenticity of the condition
(Banissy and Ward, 2007; described in Chapter 1 and 2). All cases of synaesthesia
involving colour were confirmed using tests of consistency over time, with subjects
demonstrating test-retest consistency of 85% (for letters, numbers, or other verbal
stimuli) or a score of ≤ 0.75 on the Eagleman Synaesthesia Test Battery (Eagleman,
Kagan, Nelson, Sagaram, and Sarma, 2007).
Materials and Procedure
Subjects completed two tests of sensory perception in a counterbalanced order:
The Farnsworth-Munsell Colour Hue Test and the Gratings Orientation Test (Van
Boven and Johnsen, 1994).
The Farnsworth-Munsell Colour Hue Test is a test of colour discrimination.
The apparatus is a palette of different colour hues with identical luminance and
saturation. Each hue is presented as a coloured cap and when arranged correctly the
set forms a regular colour series transforming from one hue to another. Four colour
series are presented in different trays, each containing 23 or 24 colours showing a
distinct colour transformation. The procedure for the task is as follows. For each tray,
the coloured caps are removed and arranged on the table in front of the participants.

Chapter 3
67
Two coloured caps remain in the tray, which represent the two end points of the
colour sequence (e.g. a purple and pink cap). The participant is given 2 minutes to
arrange the remaining caps into an ordered colour series from one hue to another (e.g.
purple through violet through pink). The correct order for the hues can be identified
by the experimenter from the numeric coding on the underside of each cap. A
deviation score is calculated by considering how far each colour cap deviates from the
correct location in the sequence. For example, in a correct ordering such as 4–5–6;
colour number “5” has a score of 2 because it is 1 unit from 4 and 1 unit from 6. An
incorrect ordering such as 2–5–9 would yield a score of 7 for colour “5” because it is
3 units from “2” and 4 units from “9”. The error score is the difference between the
actual score obtained and the expected score based on flawless ordering. The same
procedure was used for each of the four trays and the order of trays was randomized
across participants.
In order to investigate tactile discrimination, the Gratings Orientation Test
(GOT) was used to measure tactile acuity on the index finger tip. The GOT is a well
established method for measuring the spatial resolution of touch. It consists of a
series of square wave gratings with varying ridge widths (0.35mm – 3mm; Van Boven
and Johnsen, 1994). Each grating is applied to the finger tip in one of two orthogonal
orientations (across or along the axis of the finger tip). The task is to report the
orientation of the probe. Typically as ridge width decreases, accuracy decreases. The
GOT is thought to reflect cortical representations of the finger tip in SI (Van Boven
and Johnsen, 1994) and in this vein provides a method for accurate threshold
estimates of sensory function.
Six spatial grating probes (0.35mm, 0.5mm, 0.75mm, 1mm, 1.25mm, 1.5mm)
were used to investigate each participant’s tactile sensory threshold. Using a blocked

Chapter 3
68
design (20 trials per block; Van Boven and Johnsen, 1994) each probe was applied
manually to the participant’s right index finger tip in one of two different orientations
(across or along the axis of the finger tip). Manual application was chosen because
performance in spatial resolution is generally insensitive to the force of application
(Johnsen and Philips, 1981) and the receptive fields of the afferent population
involved in grating orientation detection are relatively independent of skin indentation
(Vega-Bermudez and Johnsen, 1999). Participants were asked to indicate the
orientation of each probe by giving a verbal response (i.e. “across” or “along”). In
total participants completed 20 trials per probe (120 trials in total) which were
randomised across blocks. For each probe half of the trials were orientated across the
finger tip and the remainder were applied along the finger tip. Participants were
blindfolded during the task to prevent any visual cues to orientation. Prior to
threshold measurement, participants completed two practice blocks, using 2mm and
3mm gratings, in which feedback was given on participants’ responses. No feedback
was given on trials involved in threshold detection.
3.3 Results
Farnsworth-Munsell Colour Hue Test The results from the Farnsworth-Munsell Colour Hue Test are shown in figure
3.1. The test is measured according to a total error score (TES) based on the deviation
from the expected ordering (it is not a percentage error). Superior performance is
reflected by a lower TES.
To investigate if the presence of synaesthesia for colour or touch was linked
with superior performance, a 2 (presence/absence of colour synaesthesia) x 2
(presence/absence of touch synaesthesia) ANOVA was conducted. It was predicted
that synaesthetes who experience colour would show superior performance on the

Chapter 3
69
colour perception task compared to those who do not. This was found to be the case,
a significant main effect of Colour Group was found [F(1,38) = 4.61, p = <.05]. The
presence of colour synaesthesia was linked with a lower TES (mean ± s.e.m = 48.22 ±
7.7), and therefore better colour discrimination, than the absence of colour
synaesthesia (mean ± s.e.m = 75.54 ± 10.1). This replicates previous reports (Yaro
and Ward, 2007). No main effect of touch group or any interaction between groups
was found. Therefore, the presence of colour synaesthesia, but not touch
synaesthesia, was linked with enhanced colour discrimination.
To further delineate the contribution of different variants of synaesthesia to
performance, the performance between each subgroup of subjects (colour-
synaesthetes; touch-synaesthetes; dual-synaesthetes; non-synaesthete controls) was
compared using a one-way ANOVA (Figure 3.1). The main effect of group
approached significance [F(3,38) = 2.45, p = .078]. In order to examine the basis of
this strong trend, and to test the a priori assumption that synaesthetes would show
enhanced colour sensitivity relative to control subjects, a series of planned t-tests were
carried out (cf. Howell, 2002, pg. 372-373). This revealed that synaesthetes who
experience colour (but not touch) as their induced experience showed superior colour
discrimination relative to non-synaesthetic control subjects [t(26) = 2.04, p = <.05].
This was also true of dual-synaesthetes – synaesthetes who experience touch and
colour significantly outperformed non-synaesthetic control subjects [t(25.90) = 2.47, p =
<.05]. No significant differences were found between the colour discrimination
abilities of synaesthetes who only experience touch-synaesthesia and non-synaesthetic
control subjects [t(24) = .62, nsig]. This indicates that the presence of ‘colour’
synaesthetic experience (rather than synaesthesia per se) is related to enhancements in
the perceptual processing of colour relative to non-synaesthetic control subjects.

Chapter 3
70
Figure 3.1 Synaesthetes who experience colour outperformed individuals who do not experience synaesthetic colour on a measure of colour perception (a). This pattern of performance was shown by synaesthetes who experience colour only relative to non-synaesthetic controls and by synaesthetes who experience both colour and touch relative to non-synaesthetic controls, but not by synaesthetes who experience touch only relative to controls (b). Error scores are based on the deviation from the expected ordering of hues. Superior colour performance is indicated by a lower error score. * = p < .05. Gratings Orientation Test Figure 3.2 shows the average tactile discrimination thresholds for all subjects
on the gratings orientation test. Enhanced tactile discrimination is reflected in a lower
threshold value (in millimetres). Thresholds were calculated using the following
formula and provide an estimate of the grating level which would lead to a 75%
response level:
g75 = glow + ((0.75 -plow)/(phigh - plow)) (ghigh - glow) g = grating spacing
20
30
40
50
60
70
80
90
100
Group
Colour Sensitivity: Total Error Score
Touch-Synaesthetes
Dual-Synaesthetes
Colour-Synaesthetes
Non-synaesthetes
*

Chapter 3
71
p = trials correct / n n = number of trials high=the grating spacing on the lowest grating spacing on which the participant responded correctly better than 75% of the time. low=the grating spacing on the highest grating spacing on which the participant responded correctly less than 75% of the time. g75 =the interpolated grating spacing on which the subject would have scored 75% had it been present. A 2 (presence/absence of colour synaesthesia) x 2 (presence/absence of touch
synaesthesia) ANOVA revealed a significant main effect of Touch Group (F(1,40) =
13.44, p = <.01). This was because synaesthetes who experience touch (either touch
only or dual-synaesthetes) showed heightened tactile sensitivity (mean ± s.e.m = 0.79
± 0.05 mm) compared to participants who do not experience synaesthetic touch (mean
± s.e.m = 1.25 ± 0.08 mm). No significant main effect of Colour Group or interaction
was observed, indicating that the presence of synaesthetic touch was linked with
heightened tactile sensitivity but not the presence of synaesthesia in general.
As with colour discrimination performance, tactile discrimination performance
between all four groups (colour-synaesthetes; touch-synaesthetes; dual-synaesthetes;
non-synaesthete controls) was compared using a one-way ANOVA. A significant
main effect of group was observed [F(3,38) = 4.50, p = .008]. Post-hoc comparisons
(corrected using Fisher’s LSD) revealed that this was because synaesthetes who only
experienced mirror-touch synaesthesia showed superior tactile discrimination relative
to both colour-synaesthetes (p = < .05) and non-synaesthetic control subjects (p = <
.05). Dual-synaesthetes also significantly outperformed both colour-synaesthetes (p =
< .05) and non-synaesthetic control subjects (p = < .01). No significant differences
were found between ‘colour-only’ synaesthetes and non-synaesthetic (p = .694)
controls or between touch-synaesthetes and dual-synaesthetes (p = .891). Therefore
the presence of synaesthesia for touch was related to enhancements in the perceptual
processing of touch relative to the absence of synaesthesia for touch (Figure 3.2).

Chapter 3
72
Figure 3.2 Synaesthetes who experience touch outperformed individuals who do not experience synaesthetic touch on a measure of tactile perception (a). This pattern of performance is shown by synaesthetes who experience touch only relative to non-synaesthetic and synaesthetic control subjects, and by synaesthetes who experience touch and colour relative to non-synaesthetic and synaesthetic control subjects. No differences in tactile perception were found between synaesthetes who experience colour only and non-synaesthete control subjects (b). Superior tactile sensitivity is indicated by a lower tactile threshold (mm). * = p < .05. 3.4 Discussion
This study extends previous reports of enhanced perceptual processing in
grapheme-colour synaesthesia (Yaro and Ward, 2007) and aimed to clarify whether
synaesthesia in other modalities has similar repercussions for perceptual processing of
stimuli in that modality. Using a sample of synaesthetes who experience either colour,
touch, or both touch and colour as evoked sensations, the findings first replicate
previous reports that synaesthetes who experience colour show superior perceptual
0.25
0.5
0.75
1
1.25
1.5
1.75
Group
Tactile Threshold (mm)
Touch-Synaesthetes
Dual-Synaesthetes
Colour-Synaesthetes
Non-synaesthetes
*

Chapter 3
73
discrimination of colour relative to non-synaesthetic subjects. They extend this by
showing:
i) synaesthetes who experience only touch show enhanced perceptual
discrimination of touch but not of colour,
ii) synaesthetes who experience both a tactile and visual concurrent show
enhanced perceptual processing of both touch and colour (although the
robustness of differences in colour processing was less strong than for tactile
processing),
iii) synaesthetes who experience only colour do not show enhanced sensory
processing in modalities outside of vision.
These findings suggest that enhanced perceptual processing is a core property
of synaesthesia, which is not limited to colour but occurs in each affected sensory
modality.
There are two possible accounts for why synaesthetes should demonstrate an
oversensitive concurrent perceptual system: 1) enhanced perceptual processing is a
consequence of the additional synaesthetic percepts which are experienced in
everyday life (i.e. enriched perceptual experience leads to enhanced perceptual
processing) or 2) enhanced perceptual processing is related to differences in brain
development as a function of synaesthesia (which may be either a cause or
consequence of synaesthesia; i.e. widespread differences in cortical connectivity or
mechanisms of cortical unmasking; Rouw and Scholte, 2007; Cohen Kadosh et al.,
2009).
Under the first account, an oversensitive concurrent perceptual system would
be explained as a proximal consequence of synaesthetic experience. For example, the
presence of stable synaesthetic associations may impact on the internal structure of

Chapter 3
74
sensory representations. In the case of colour, it has been suggested that linguistic
labels are necessary to categorize colours across a perceptual continuum (Davidoff,
2001) and cross-cultural differences in the number of colour labels has been shown to
influence colour perception and memory (Robertson, Davies, and Davidoff, 2000). It
may be the case that for some colour synaesthetes (i.e. grapheme-colour
synaesthesia), the presence of stable associations with colour increases the number of
colour terms and thereby impacts on the internal structure of colour space (Yaro and
Ward, 2007). In accordance with this, Simner and colleagues (2005) report that
grapheme-colour synaesthetes produce a greater depth of colour descriptions and use
more colour terms than non-synaesthete control participants. It is unclear however,
how this would extend to other variants and concurrent perceptual systems such as
enhanced tactile acuity in mirror-touch synaesthesia.
Additionally, insights into the influence of enriched perceptual experience on
sensory enhancement in the deprived brain would suggest that it is unlikely that extra
synaesthetic percepts are the cause of superior perceptual processing (Pascual-Leone,
Amedi, Fregni, and Merabet, 2005). For example, blind subjects have been shown to
be superior to sighted subjects on the grating orientations test, however this
superiority does not correlate with Braille reading experience or differ between Braille
readers and blind non-readers (Goldreich and Kanics, 2003; Van Boven, Hamilton,
Kauffman, Keenan, and Pascual-Leone, 2000). This indicates that mechanisms of
cross-modal plasticity following visual deprivation (rather than increased tactile
experiences) drives tactile acuity enhancement in the blind (Goldreich and Kanics,
2003). Consistent with this, in sighted-subjects visual deprivation induced by long-
term blindfolding results in temporary enhancements of passive tactile acuity, but
enriched tactile experience on the same finger does not (Kauffman, Théoret, Pascual-

Chapter 3
75
Leone, 2002) – further implying that elevated sensory sensitivity may be linked to
mechanisms of cross-modal plasticity rather than additional sensory experience.
Two mechanisms of cross-modal plasticity have been suggested to account for
compensatory changes in the deprived brain: cortical unmasking of pre-existing
connections and cortical reorganisation (e.g. Pascual-Leone et al., 2005; Wittenberg et
al., 2004). Mechanisms of cortical unmasking (through processes such as
disinhibition) involve the strengthening of existing within and between region
anatomical pathways (i.e. functional but not structural differences), while mechanisms
of cortical reorganisation involve the establishment of new local and widespread
anatomical connections (i.e. structural differences). The role of these mechanisms on
compensatory change is thought to reflect differences in the speed of plasticity, with
unmasking representing a form of rapid change which if sustained leads to long
lasting cortical changes such as the establishment of new anatomical connections
(slow acting mechanism; Pascual-Leone et al., 2005). For example, in the case of
temporary enhancements in tactile acuity following blindfolding, unmasking of
existing connections may offer a fast-acting mechanism of cross-modal plasticity to
maintain functional behaviour (i.e. perception of the environment through rapid
enhancements in tactile processing). In comparison, in the case of enhanced tactile
processing in the blind, sustained unmasking of existing connections may lead to new
local and widespread anatomical pathways resulting in long lasting enhancements in
tactile acuity which aid in daily life (Pascual-Leone et al., 2005).
There is growing evidence that synaesthesia may act upon the ‘normal’
architecture for cross-modal interactions (see Sagiv and Ward, 2006 for review) and
parallels may be drawn between mechanisms of cross-modal plasticity following
compensatory changes in the deprived brain and those which have been suggested to

Chapter 3
76
underlie synaesthetic experience (Cohen Kadosh and Walsh, 2006). A current area of
debate is whether synaesthetic experience is a consequence of additional structural
connectivity between brain regions (i.e. structural differences akin to cortical
reorganisation following sensory deprivation), malfunctions in cortical inhibition (i.e.
functional but not structural differences akin to cortical unmasking following sensory
deprivation), or a combination of both (Bargary and Mitchell, 2008; Cohen Kadosh
and Henik, 2007; Cohen Kadosh and Walsh, 2008; Grossenbacher and Lovelace,
2001; Hubbard and Ramachandran, 2005; Rouw and Scholte, 2007; Smilek et al.,
2001). Supporting evidence for structural connectivity accounts is provided by
diffusion tensor imaging findings that grapheme-colour synaesthetes show increased
structural connectivity in inferior-temporal, parietal and frontal brain regions when
compared to non-synaesthetes (Rouw and Scholte, 2007). Evidence for inhibition
accounts is provided by findings that synaesthetic-like experiences can be induced
following hallucinogenic drugs (Aghajanian and Marek, 1999) and that grapheme-
colour synaesthesia can be induced in non-synaesthetes (individuals without aberrant
connectivity) using post-hypnotic suggestion (Cohen Kadosh et al., 2009). It is
plausible that enhanced sensory perception in synaesthesia may reflect a combination
of these mechanisms. Mechanisms of reduced inhibition may act by unmasking local
anatomical pathways (akin to visual deprivation studies in sighted subjects; Cohen
Kadosh and Henik, 2007; Cohen Kadosh and Walsh, 2008), while altered cortical
connectivity may lead to enhanced perceptual sensitivity through aberrant circuitry
within the concurrent perceptual system (akin to sensory sensitivity enhancements in
the blind). It will be interesting to determine if altered connectivity in synaesthesia
(Rouw and Scholte, 2007) may reflect sustained unmasking of existing connections

Chapter 3
77
(e.g. Cohen Kadosh et al., 2009) and what implications this may have for veridical
sensory processing.
In summary, this study extends previous findings that grapheme-colour
synaesthetes show enhanced perceptual processing of colour (Yaro and Ward, 2007)
to suggest that an oversensitive concurrent perceptual system is a core property of
synaesthesia. Mirror-touch synaesthetes were shown to have enhanced tactile
sensitivity only, synaesthetes who experience both mirror-touch and a form of colour
synaesthesia were shown to demonstrate enhanced tactile and colour perception, and
synaesthetes who only experience colour were shown to have enhanced perceptual
processing of colour only. These findings imply that the presence of synaesthesia has
repercussions for sensory processing for stimuli which do not themselves induce
synaesthetic experience. It remains to be determined whether an oversensitive
concurrent perceptual system is a cause or consequence of synaesthesia.

Chapter 4
78
CHAPTER 4: MIRROR-TOUCH SYNAESTHESIA AND
EMPATHY
In the preceding chapters I investigated the behavioural correlates and perceptual
consequences of mirror-touch synaesthesia. Here I consider the implications of
mirror-touch synaesthesia for general cognitive processing. Previous fMRI findings
link mirror-touch synaesthesia to heightened activations in the mirror-touch system
(the same neural system activated in non-synaesthetes when observing touch to
others). It has been suggested that components of the mirror-touch system may act to
facilitate processes such as empathy and emotion recognition because they provide
the perceiver with a neurophysiological mechanism to simulate what it would “feel
like” to be in the same situation. To examine this possibility, two experiments were
conducted to investigate the influence of heightened sensorimotor simulation in
mirror-touch synaesthesia on empathy. Experiment 1, ‘Mirror-touch synaesthesia
and empathy’, demonstrates that mirror-touch synaesthesia, but not other variants of
synaesthesia, is linked with heightened empathic abilities for specific components of
empathy. Experiment 2, ‘Empathy and personality’, extends the findings from
experiment 1 by demonstrating that differences in empathy are ‘other’ rather than
‘self’ orientated reactions.
4.1 Introduction
Empathy is a higher order psychological construct and is considered to have
two main strands: (i) cognitive empathy – predicting and understanding another’s
mental state by using cognitive processes (i.e. role / perspective taking), and (ii)
affective empathy – experiencing an appropriate emotional response as a consequence
of another’s state (Baron-Cohen and Wheelwright, 2004; Decety and Jackson, 2004;
Preston and de Waal, 2002). Evolutionary perspectives suggest that there are several
possible systems which mediate this division, including phylogentically early
emotional contagion systems and more recently evolved cognitive perspective taking
mechanisms (De Waal, 2007), with the former thought to play a crucial role in
supporting the ability to empathize emotionally (e.g. I feel sad when I see someone
else sad) and the later considered to be linked to more complex empathic cognitions
including perspective taking and mentalizing.

Chapter 4
79
Recent research has suggested that one neurophysiological mechanism which
may mediate people’s abilities to empathize and understand the emotions of others is
shared affective neural systems in which common brain areas are activated during
both experience and passive observation of other’s experiences. Moreover, building
on the discovery of mirror neurons in the monkey brain (Gallese, Fadiga, Fogassi, and
Rizzolatti, 1996; Rizzolatti and Craighero, 2004), functional brain imaging has
suggested the existence of mirror systems in humans not only for actions (e.g.
Buccino et al., 2001), but also for sensations and emotions (e.g. disgust: Jabbi, Swart
and Keysers, 2006; Wicker, Keysers¸ Plailly, Royet, Gallese, and Rizzolatti, 2003;
touch: Blakemore, Bristow, Bird, Frith, and Ward, 2005; Ebisch, Perucci, Ferretti, Del
Gratta, Luca Romani, and Gallese, 2008; Keysers, Wicker, Gazzola, Anton, Fogassi,
and Gallese, 2004; pain: Avenanti, Beuti, Galati, and Aglitoi, 2005; Bufalari, Aprile,
Avenanti, Di Russo, and Aglioti, 2007; Morrison, Lloyd, di Pellegrino, and Roberts,
2004; Singer, Seymour, O’Doherty, Kaube, Dolan, and Frith, 2004; emotion: Carr,
Iacoboni, Dubeau, Mazziotta, and Lenzi, 2003). These systems may be crucial for
empathy because they enable the observer to simulate another’s experience by
activating the same brain areas that are active when the observer experiences the same
emotion or state (Gallese, 2006; Gallese and Goldman, 1998; Keysers and Gazzola,
2006; Oberman and Ramachandran, 2007). Consistent with this, is evidence that
increased activations in the auditory mirror system are correlated with high self
reported empathy (Gazzola, Aziz-Zadeh, and Keysers, 2006); that increases in trait-
cognitive empathy are correlated with increases in sensorimotor simulation when
viewing others’ pain (Avenanti, Minio-Paluello, Bufalari, and Aglioti, 2009); and that
participants self reported empathy skills are positively correlated with levels of
cortical mirroring of when witnessing disgust (Jabbi et al., 2006). Furthermore, there

Chapter 4
80
is a growing body of evidence suggesting that individuals with autistic spectrum
disorder (ASD) have impaired activity in the action mirror system (Dapretto, Davies,
Pfeifer, Scott, Sigman, Bookheimer, and Iacoboni, 2006; Oberman, Hubbard,
McCleery, Altschuler, Ramachandran, and Pineda, 2005), which may lead to the
deficits in imitation and empathy observed in ASD (Iacoboni and Dapretto, 2006;
Oberman and Ramachandran, 2007; but see Southgate and Hamilton, 2008).
As discussed in preceding chapters, previous functional magnetic resonance
imaging findings indicate that synaesthetic tactile experiences in mirror-touch
synaesthesia are associated with hyperactivity in the same mirror-touch network that
is evoked by observed touch in non-synaesthete controls in which no overt tactile
experience is elicited (Blakemore et al., 2005). As such, mirror-touch synaesthesia
may offer a unique opportunity to explore the role that the tactile mirror system has in
empathy because it enables investigations into the relationship between heightened
sensorimotor simulation in the mirror-touch system and empathic sensitivity.
To address this possibility two experiments were conducted. In experiment 1,
the empathic abilities of ten mirror-touch synaesthetes were compared to a
synaesthetic and non-synaesthetic control group. In experiment 2, potential factors
which may contribute to heightened empathy were investigated by contrasting mirror-
touch synaesthetes with non-synaesthetic participants on empathy and personality
measures.
4.2 Experiment 1: Mirror-touch synaesthesia and empathy
Participants
Ten mirror-touch synaesthetes (6 females and 4 males, mean age ± Std. Error
= 37.6 ± 5.59 years) and twenty non-synaesthetic controls matched for age and gender
(12 females and 8 males, mean age ± Std. Error = 32.95 ± 3.24 years) took part in the

Chapter 4
81
study. All cases of mirror-touch synaesthesia were confirmed on the visual-tactile
spatial congruity paradigm described previously (Banissy and Ward, 2007; also see
Chapter 1 and Chapter 2 of this thesis for a description of the task).
In addition to this, twenty-five synaesthetes (22 females and 3 males, mean
age ± Std. Error = 43.96 ± 3.38 years) experiencing other forms of synaesthesia
(minimally grapheme-colour synaesthesia) but not mirror-touch synaesthesia took
part. These synaesthetes acted as a synaesthetic control group and demonstrated test-
retest consistency of ≥ 85% for letters, numbers and other verbal stimuli. The
synaesthete control group were included to ensure that any differences in empathy
were not due to a general feature of synaesthesia.
Materials and Procedure
All participants completed the Empathy Quotient (Baron-Cohen, Richler,
Bisarya, Gurunathan, and Wheelwright, 2003; Baron-Cohen and Wheelwright, 2004).
The EQ is a self report scale designed to empirically measure empathy. As noted
previously, empathy is a higher order construct and has been theorised as having two
main strands: (i) cognitive empathy – predicting and understanding another’s mental
state by using cognitive processes (i.e. role / perspective taking), and (ii) affective
empathy – experiencing an appropriate emotional response as a consequence of
another’s state (Baron-Cohen and Wheelwright, 2004; Preston and de Waal, 2002).
The EQ was developed to measure both cognitive and affective components of
empathy, has been validated on both clinical and control groups (Baron-Cohen et al.,
2003), and it has been shown to distinguish between these groups (Baron-Cohen et al.,
2003; Baron-Cohen and Wheelwright, 2004). In addition to this, the EQ has been
validated on measures of concurrent validity (Lawrence, Shaw, Baker, Baron-Cohen,
and David, 2004) and has been shown to have high test-retest reliability over 12

Chapter 4
82
months (Baron-Cohen et al., 2003). The scale is comprised of 40 test items and 20
filler items. All items are a series of statements (e.g. ‘I can tune into how someone
feels rapidly and intuitively) and responses are given on a 4 point scale ranging from
‘strongly agree’ to ‘strongly disagree’. Responses score 1 or 2 points for an empathic
response and 0 points for all other responses. Principal component analysis has
indicated that the EQ is comprised of three main factors (i) cognitive empathy, (ii)
emotional reactivity and (iii) social skills (Lawrence et al., 2004; Muncer and Ling,
2006). Confirmatory factor analysis has indicated that the EQ may be better
conceived as comprising of this three factor structure rather than a 40 item unifactorial
scale (Lawrence et al., 2004; Muncer and Ling, 2006).
Results and Discussion
The empathic ability of mirror-touch synaesthetes was compared with non-
synaesthetic control participants and controls that report other types of synaesthesia
but do not report mirror-touch synaesthesia. Results from non-synaesthetic controls
and synaesthetes lacking mirror touch did not differ and were therefore combined.
Empathy scores for each component of the EQ are summarised in Figure 4.1. Mirror-
touch synaesthetes showed significantly higher scores on the emotional reactivity
subscale of the EQ relative to controls [t(53) = 2.15, p = < .05]. This subscale is
thought to reflect affective components of empathy, and instinctive empathic
responses to others (Baron-Cohen and Wheelwright, 2004; Lawrence et al., 2004).
There was also a non-significant trend for mirror-touch synaesthetes to outperform
control subjects on the cognitive empathy (i.e. mentalizing / perspective taking)
subscale [t(53) = 1.92, p = .061]. Scores on the social skills subscale did not approach
significance [t(53) = 1.22, p = .227]. Therefore, mirror-touch synaesthetes showed
heightened empathy on some, but not all aspects of empathy. This supports the notion

Chapter 4
83
that empathy is multi-faceted (Baron-Cohen and Wheelwright, 2004; Decety and
Jackson, 2004; Lawrence et al., 2004; Muncer and Ling, 2006; Preston and de Waal,
2002) and implies that sensorimotor simulation may modulate some, but not all,
aspects of this ability. Further, the evidence that enhanced empathy is not found in
other types of synaesthesia suggests that heightened empathy relates specifically to
mirror-touch synaesthesia (and the neural system which underpins this condition).
6
8
10
12
14
16
18
20
22
24
EQ Score
Mirror-Touch Synaesthetes
Controls*
Cognitive Empathy Emotional Reactivity Social Skills
Figure 4.1 Mirror-touch synaesthetes showed significantly higher scores than controls on the emotional reactivity component, but not other components, of the Empathy Quotient (mean ± s.e.m). * = p < .05.
Previous functional magnetic imaging findings indicate that, in healthy adults,
emotional empathy engages the cortical sensorimotor network (including the premotor
cortex, primary somatosensory cortex and motor cortex) more than cognitive empathy
(Nummenmaa, Hirvonen, Parkkola, and Hietanen, 2008). Further,
neuropsychological findings have demonstrated a functional and anatomical double
dissociation between deficits in cognitive empathy and emotional empathy, with

Chapter 4
84
emotional empathy being linked to lesions to the human mirror system and cognitive
empathy being associated to lesions to the ventromedial prefrontal cortices (Shamay-
Tsoory, Aharon-Peretz, and Perry, 2009). This functional coupling between
emotional and cognitive empathy suggests that emotional empathy may be linked
more closely to sensorimotor simulation of another’s state and the evidence that
mirror-touch synaesthetes only significantly differ from controls on levels of
emotional reactivity is consistent with this.
Two qualifications are apposite: (a) it remains unclear whether heightened
emotional reactivity reflects differences in the nature of the individual (i.e. more
emotional or more distressed by emotional scenes as opposed to more empathic /
concerned), and (b) evidence of a borderline significant difference on cognitive
empathy suggests that differences in empathy may not be limited to emotional
empathy per se. To further address these issues a second experiment was conducted.
4.3 Experiment 2: Empathy and personality in mirror-touch synaesthesia
While findings from experiment 1 indicate that mirror-touch synaesthesia is
related to heightened emotional empathy, it remains unclear if the emotional reactivity
component of the EQ reflects the nature of the individual (i.e. more emotional / more
distressed) rather than emotional empathy per se (Muncer and Ling, 2006).
Moreover, the emotional reactivity subscale of the EQ fails to consider the role of
personal distress (an emotionally specific response to one’s own state rather than an
emotional response related to emotional empathy) and thus it is difficult to confirm
whether responses on the emotional reactivity component are indeed other (e.g.
feeling compassion or sorrow towards to another person) rather than self-oriented
(e.g. feeling distress from an unpleasant scene rather than feeling sorrow or concern;
c.f. Batson, 1991; Davis, 1994; Lamm, Batson, and Decety, 2007; Saarela,

Chapter 4
85
Hlushchuk, Williams, Schürmann, Kalso, and Hari, 2007). The implications of this
distinction for behaviour are that other-oriented empathy may promote altruistic
motivations to help another person, while self-orientated empathy may lead to egoistic
motivations to reduce the personal distress felt by the observer and ultimately
counteract positive empathic behaviours (e.g. helping behaviour; Batson, 1991;
Eisenberg, 2000).
This study aimed to investigate this, by contrasting mirror-touch synaesthetes
with age and gender matched non-synaesthetes on the EQ and an additional measure
of empathy - the Interpersonal Reactivity Index (IRI; Davis, 1980). The IRI contains
a component relating to ‘personal distress’ that is regarded as being self-oriented
emotional reactivity and so is able to clarify if heightened emotional reactivity in
mirror-touch synaesthetes is other rather than self-oriented. In addition, the
relationship between individual differences in emotional reactivity (in both the
synaesthetes and controls) and personality traits were investigated using the ‘Big Five
Inventory (BFI; John, Donahue, and Kentle, 1991). If emotional reactivity is self-
oriented then a correlation with the neuroticism (or emotional stability) trait is
expected, but not if it is other-oriented. A previous study found no relationship
between empathy and emotional stability, but it did not divide empathy into
component factors (Del Barrio, Aluja, and Garcia, 2004), thus my study is the first to
consider the importance of personality on different facets of empathy as indexed by
the EQ and IRI.
4.3.1 Methods
Participants
In the first part of the study, sixteen mirror-touch synaesthetes (mean age ±
s.e.m = 35.5 ± 3.58 years) and sixteen age and gender matched non-synaesthete

Chapter 4
86
controls (mean age ± s.e.m = 35.69 ± 3.45 years) were compared. All cases of mirror-
touch synaesthesia were confirmed on the visual-tactile spatial congruity paradigm
designed to provide evidence for the authenticity of the condition (cf. Banissy and
Ward, 2007; Chapter 1 and Chapter 2 this thesis). Of the sixteen synaesthetes, five
took part in experiment 1.
For analysis of the relationship between empathy and personality, an
additional 88 non-synaesthete control subjects were recruited. These controls were
combined with mirror-touch synaesthetes and non-synaesthete controls from the first
part of the study to provide a sample of 120 subjects (mean age ± s.e.m = 27.98 ± 2.78
years). The additional controls were included to provide a range of scores on the
empathy and personality measures, with mirror-touch synaesthetes thought to reflect
the top end of a spectrum on empathy scores.
Materials and procedure
All subjects completed the EQ, IRI, and BFI.
The EQ is a self report measure designed to empirically measure empathy and
is described in experiment 1 of this chapter (pp. 78-79).
The IRI is a 28 item self-report empathy measure (Davis, 1980). It is
comprised of four subscales; perspective taking, fantasizing, empathic concern and
personal distress. Each subscale contains seven items which are measured on a five
point Likert scale ranging from 0 (“Does not describe me well”) to 4 (“Describes me
very well”). For each subscale, a minimum score of 0 or maximum score of 35 is
possible. For the perspective taking, fantasizing and empathic concern subscales
higher scores reflect heightened empathy. For the personal distress subscale higher
scores are reflective of self-orientated emotional reactivity (Davis, 1994).

Chapter 4
87
The BFI is a 44-item scale designed to measure components of the Big Five
personality traits (extraversion, agreeableness, conscientiousness, neuroticism, and
openness; John et al., 1991). Respondents are asked to indicate on a five point Likert
scale the extent to which a series of statements related to each personality trait best
describe their own characteristics. Responses are given from 1 (“Disagree strongly”)
to 5 (“Agree strongly”). Analysis of the reliability of the scale (John and Srivastava,
1999) indicates a coefficient alpha of 0.83 and the BFI shows good convergent
validity with TDA (Goldberg, 1992) and NEO-PI personality measures (Costa and
McCrae, 1985).
4.3.2 Results
Mirror-touch synaesthetes compared to non-synaesthete controls
The empathic abilities of mirror-touch synaesthetes and controls on each
component of the EQ and IRI were compared. On the EQ (Figure 4.2), mirror-touch
synaesthetes scored significantly higher than non-synaesthete controls on the
emotional reactivity subscale [t (30) = 2.29, p = < .05]. Despite synaesthetes scoring
higher than controls on the cognitive empathy (CE) and social skills (SS) subscales
these did not approach significance [CE: t(30) = 1.20, p = .239; SS: t(30) = 1.37, p =
.181]. This is consistent with findings from experiment 1 where mirror-touch
synaesthetes differed significantly from controls on the emotional reactivity subscale,
but not cognitive empathy or social skills subscale of the EQ.

Chapter 4
88
4
6
8
10
12
14
16
18
20
22
EQ Score
Mirror-Touch Synaesthetes
Controls
*
Cognitive Empathy Emotional Reactivity Social Skills
Figure 4.2 In experiment 2, mirror-touch synaesthetes showed significantly higher scores than controls on the emotional reactivity component, but not other components, of the Empathy Quotient (mean ± s.e.m). * = p < .05.
In addition, on the IRI mirror-touch synaesthetes showed significantly
elevated scores on the fantasizing subscale [t(30) = 2.35, p = < .05], but not on the
alternative subscales (Figure 4.3). Of note, is the comparable performance of
synaesthetes and controls on the perspective taking subscale and higher scores of
controls relative to synaesthetes on the personal distress subscale, which indicates that
differences were not simply due to a tendency for synaesthetes to provided higher
self-reported values overall (also see Figure 4.4).

Chapter 4
89
8
10
12
14
16
18
20
22
24
26
28
IRI Score
Mirror-Touch Synaesthetes
Controls*
PT F PD EC
Figure 4.3 Mirror-touch synaesthetes showed significantly higher scores than controls on the fantasizing component, but not other components, of the Interpersonal Reactivity Index (mean ± s.e.m). * = p < .05. PT = Perspective Taking, F = Fantasizing, PD = Personal Distress, EC = Empathic Concern.
18
20
22
24
26
28
30
32
34
36
38
40
42
44
BFI Score
Mirror-Touch Synaesthetes
Controls
*
O A C N E
Figure 4.4 Mirror-touch synaesthetes showed significantly higher scores than controls on the Openness subscale, but not other components, of the BFI (mean ± s.e.m). * = p < .05. E = Extraversion, A = Agreeableness, C = Conscientiousness, N = Neuroticism, O = Openness.

Chapter 4
90
Synaesthetes also differed from controls on the openness to experience trait of
the BFI [t(30) = 2.62, p = .014], with synaesthetes demonstrating higher scores on this
personality trait. No significant differences were found on the other BFI subscales
(Figure 4.4).
Empathy and personality
Pearson’s correlations were conducted to investigate the relationship between
five components of personality (extraversion, agreeableness, conscientiousness,
neuroticism and openness) with empathic abilities on each component of the EQ
(cognitive empathy, emotional reactivity, social skills). This analysis revealed that
the cognitive empathy subscale of the EQ showed a moderate correlation with
extraversion [n = 120, r = .401, p = < .001], agreeableness [n = 120, r = .463, p = <
.001], and conscientiousness [n = 120, r = .383, p = < .001]; but not neuroticism or
openness traits. Similar associations were observed for the emotional reactivity
subscale, which was associated with extraversion [n = 120, r = .292, p = < .01],
agreeableness [n = 120, r = .455, p = < .001], conscientiousness [n = 120, r = .248, p
= < .01] and openness [n = 120, r = .180, p = < .05]. Importantly, emotional reactivity
was not correlated with neuroticism / emotional stability [n = 120, r = .016, p = .862].
The social skills subscale displayed a positive relationship with extraversion [n = 120,
r = .337, p = < .001], agreeableness [n = 120, r = .336, p = < .001], and
conscientiousness [n = 120, r = .324, p = < .001]; a negative relationship with
neuroticism [n = 120, r = -.311, p = < .01]; but no relationship with openness.
Additionally, the relationship between IRI scores and personality were
explored. This revealed a positive association between the perspective taking
subscale and agreeableness [n = 120, r = .237, p = < .01]; the fantasizing subscale and
openness [n = 120, r = .319, p = < .001]; the empathic concern subscale and

Chapter 4
91
extraversion [n = 120, r = .185, p = < .05], agreeableness [n = 120, r = .524, p = <
.001], and conscientiousness [n = 120, r = .201, p = < .05]. The personal distress
subscale was found to correlate negatively with extraversion [n = 120, r = -.256, p = <
.01] and conscientiousness [n = 120, r = -.283, p = < .01], but positively with the
neuroticism trait [n = 120, r = .532, p = < .001].
4.4 Discussion
Experiment 2 sought to examine findings of heightened emotional reactivity in
mirror-touch synaesthesia documented in experiment 1. The study aimed to clarify if
enhanced emotional reactivity was specific to enhanced other-orientated emotional
empathy or to self related processes, and to investigate previous trends towards
significantly higher levels of cognitive empathy in mirror-touch synaesthetes. Using a
larger sample of mirror-touch synaesthetes and a new control group, the findings first
replicate previous reports of heightened emotional reactivity, but not other
components of empathy, in mirror-touch synaesthesia. They then confirm that
heightened emotional reactivity in mirror-touch synaesthesia is not linked with
heightened personal distress, indicating that heightened empathy in mirror-touch
synaesthesia is indeed other rather than self-oriented. Further, the findings indicate
that emotional reactivity does not reflect a less emotionally stable personality type –
scores on emotional reactivity component of the EQ were not related to neuroticism /
emotional stability personality trait.
Mirror-touch synaesthetes also differed from non-synaesthete controls on the
fantasizing scale of the IRI. This subscale reflects an individual’s ability to match
another’s feelings and behaviours onto their own. Previous findings indicate that
increased scores on the fantasizing subscale of the IRI are related with heightened
activations within the anterior insula and frontal operculum when witnessing others’

Chapter 4
92
gustatory emotions (Jabbi et al., 2006). In a previous brain imaging study on mirror-
touch synaesthesia, the only brain region to differ between mirror-touch synaesthetes
and non-synaesthetes when observing touch to others was the anterior insula
(Blakemore et al., 2005). Shared representations within this brain region may be
important for this component of empathy.
Finally, analysis of correlations between personality and measures of empathy
provide important insights into the EQ and IRI measures. Evidence that heightened
extraversion, agreeableness and conscientiousness are related with higher responses
on all components of the EQ is consistent with findings implicating these personality
traits in social cognition more generally; extraversion has been suggested to be a
measure of social skills (John and Srivastava, 1999); agreeableness has been linked to
altruistic behaviour (Barrick and Mount, 1991); and conscientiousness correlates
negatively with psychoticism (Aluja, Garcia, and Garcia, 2002). Of note, is the lack
of association between emotional reactivity and neuroticism, but presence of a strong
positive correlation between the personal distress subscale of the IRI and neuroticism.
This is consistent with the notion that the neuroticism personality trait would indicate
self rather than other-oriented processes when correlated with empathy. Further to
this, the openness trait appears specifically related to one’s ability to match another’s
emotional state with one’s own (as indicated by the positive relationship between the
openness subscale and emotional reactivity subscale of the EQ; and between the
openness subscale and fantasizing subscale of the IRI) and this was the only
personality trait where synaesthetes significantly differed from controls. Notably, it is
difficult to determine the nature of the higher levels of openness to experience
observed in mirror-touch synaesthetes relative to controls because the mirror-touch
group included self-referred cases who have already demonstrated openness to

Chapter 4
93
experience by contacting unknown researchers for a study. Therefore, it is difficult to
determine whether differences in the levels of openness to experience would extend to
larger randomly recruited populations of mirror-touch synaesthetes (i.e. whether
higher levels of openness to experience are more prominent in self-referred rather
than randomly sampled populations of synaesthetes).
General Summary
In sum, the studies presented in this chapter indicate that mirror-touch
synaesthetes show heightened levels of emotional, but not other components, of
empathy. In experiment 1, mirror-touch synaesthetes scored significantly higher on
emotional reactivity components of empathy, but not on social skills or cognitive
empathy. A control group of synaesthetes who experience other types of synaesthesia
but not mirror-touch did not differ from non-synaesthete controls, indicating that
differences in empathy were specific to this subtype of synaesthete. Experiment 2
extended findings in experiment 1 to demonstrate that the heightened emotional
reactivity observed in mirror-touch synaesthetes reflects other, rather than self-
orientated, emotional reactions. Mirror-touch synaesthetes were also shown to differ
from non-synaesthetes on an alternative measure to that used in experiment 1. On
both measures synaesthetes showed heightened affective empathy (but not cognitive
empathy), implying that sensorimotor simulation is important for some, but not all
components of empathy. Given that mirror-touch synaesthesia has been linked to
heightened sensorimotor simulation (Blakemore et al., 2005), these findings appear
consistent with accounts of empathy that posit a role for sensorimotor simulation
mechanisms (Gallese, 2006; Gallese and Goldman, 1998; Keysers and Gazzola, 2006;
Oberman and Ramachandran, 2007) and are consistent with functional brain imaging

Chapter 4
94
(Nummenmaa et al., 2008) and neuropsychological findings (Shamay-Tsoory et al.,
2009) which indicate that emotional empathy may be linked more closely to
sensorimotor simulation of another’s state.

Chapter 5
95
CHAPTER 5: FACIAL EXPRESSION RECOGNITION IN MIRROR-TOUCH SYNAESTHESIA
The findings from chapter 4 indicate that mirror-touch synaesthesia is linked with
heightened affective empathy. Simulation models of expression recognition contend
that in order to understand another’s facial expressions individuals map the
perceived expression onto the same sensorimotor representations which are active
during the experience of the perceived emotion. To investigate this view, the present
study examines facial expression and identity recognition abilities in mirror-touch
synaesthesia. Mirror-touch synaesthetes outperformed non-synaesthetic controls on
measures of facial expression recognition, but not on control measures of face
memory or face perception. These findings imply a role for sensorimotor simulation
in the recognition of facial affect, but not facial identity.
5.1 Introduction
The ability to perceive a face is one of the most highly developed visual skills
in humans, important not only for our ability to recognise the identity of others but
also to facilitate social interaction. Neurocognitive models of face perception
highlight the role of a number of face-specific and domain-general mechanisms in this
process, and distinguish between those involved in the recognition of facial identity
and those involved in the recognition of expressions at different stages of cortical
processing (Bruce and Young, 1986; Calder and Young, 2005; Haxby, Hoffman, and
Gobbini, 2000).
Simulation accounts of expression recognition contend that to understand
another’s facial emotion the observer simulates the sensorimotor response associated
with generating the perceived facial expression (Adolphs, 2002; Adolphs, 2003;
Gallese, Keysers, and Rizzolatti, 2004; Goldman, and Sripada, 2005; Keysers and
Gazzola, 2006). This is supported by evidence that electromyographic responses in
expression relevant facial muscles are increased during subliminal exposure to
emotional expressions (Dimberg, Thunberg, and Elmehed, 2000); that preventing the

Chapter 5
96
activation of expression relevant muscles impairs expression recognition (Oberman,
Winkielman, and Ramachandran, 2007); and that perceiving another person’s facial
expressions recruits similar premotor and somatosensory representations as when the
perceiver generates the same emotion or expression (Carr, Iacoboni, Dubeau,
Mazziotta, and Lenzi, 2003; Hennenlotter et al., 2005; Montgomery and Haxby, 2008;
van der Gaag, Minderaa, and Keysers, 2007; Winston, O’Doherty, and Dolan, 2003).
Further, neuropsychological findings indicate that focal brain damage to right
somatosensory cortices is associated with expression recognition deficits (Adolphs,
Damasio, Tranel, Cooper, and Damasio, 2000), and transcranial magnetic stimulation
findings demonstrate the necessity of the right somatosensory cortex for facial
expression recognition abilities in healthy adults but not face identity recognition
(Pitcher, Garrido, Walsh and Duchaine, 2008). These findings imply that purely
visual face-processing mechanisms interact with sensorimotor representations to
facilitate expression recognition. This thought to differ to facial identity recognition,
in which there is no clear indication of how one could simulate another’s identity
(Calder and Young, 2005).
While much has been learnt from studies involving a disruption of simulation
mechanisms, an alternative approach is to consider whether facilitation of these
mechanisms promotes expression recognition. One example of facilitated
sensorimotor simulation is the case of mirror-touch synaesthesia (Blakemore, Bristow,
Bird, Frith, and Ward, 2005). As noted previously, in mirror-touch synaesthesia,
simply observing touch to others elicits a conscious tactile sensation on the
synaesthete’s own body. Functional brain imaging indicates that this variant of
synaesthesia is linked to heightened neural activity in a network of brain regions
which are also activated in non-synaesthetic control subjects when observing touch to

Chapter 5
97
others (the mirror-touch system; Blakemore et al., 2005). The mirror-touch system is
comprised of brain areas active during both the observation and passive experience of
touch (including primary and secondary somatosensory cortices, and premotor cortex;
Blakemore et al., 2005; Ebisch, Perucci, Ferretti, Del Gratta, Luca Romani, and
Gallese, 2008; Keysers, Wicker, Gazzola, Anton, Fogassi, and Gallese, 2004). It has
been suggested that brain systems with mirror properties may be crucial for social
perception because they provide a probable neural mechanism to facilitate
sensorimotor simulation of another’s perceived state (Gallese, Keysers, and Rizzolatti,
2004; Keysers and Gazzola, 2006). In this sense, mirror-touch synaesthesia can be
viewed as a case of heightened sensorimotor simulation, which may be able to inform
on the role of sensorimotor simulation mechanisms in social cognition. Consistent
with this, in chapter 4 I report that mirror-touch synaesthetes show heightened
emotional empathy compared to control subjects. Enhanced empathy was not found in
other types of synaesthesia, suggesting that it relates specifically to this variety of
synaesthesia.
This study sought to establish whether this type of synaesthetes differed in
another aspect of social perception, namely facial expression recognition. To do so,
the performance of mirror-touch synaesthetes and non-synaesthetic control subjects
on tasks of facial expression recognition, identity recognition and identity perception
were compared. Based upon the hypothesis that mirror-touch synaesthetes have
heightened sensorimotor simulations mechanisms it was predicted that synaesthetes
would show superior performance on expression recognition tasks due to heightened
sensorimotor simulation mechanisms, but not on the identity recognition or face
perception control tasks that are less dependent on simulation.

Chapter 5
98
5.2 Method
Participants
Eight mirror-touch synaesthetes (6 female and 2 male; mean age ± s.d = 45.6 ±
11.7 years) and twenty non-synaesthetic control subjects (15 female and 5 male; mean
age ± s.d = 35.6 ± 13.6 years) took part in the study. All cases of mirror-touch
synaesthesia were confirmed using a previously developed visual-tactile congruity
paradigm designed to provide evidence for the authenticity of the condition (Banissy
and Ward, 2007; also see Chapters 1 and 2 of this thesis for a description of the task
used).
Materials and Procedure
Participants completed four tasks in a counterbalanced order. These tasks are
detailed below.
Films Facial Expression Recognition
This task investigated participants’ abilities to recognize the emotional
expressions of others. In each trial participants were presented with an adjective
describing an emotional state followed by three images (each image shown for 500
msec, with a 500msec ISI) of the same actor or actress displaying different facial
expressions. Participants were asked to indicate which of the three images best
portrayed the target emotional adjective. There was no fixed inter-trial interval as
participants began each trial with a key press (i.e. the task was self-paced).
In order to portray subtle facial expressions, expression stimuli were captured
from films (Figure 5.1a). Fifty-eight target images (preceded by three practice trials)
from 15 films were used. All films were from a non-English speaking country to
decrease the probability that participants had seen them or were familiar with the

Chapter 5
99
actors. Target and distracter stimuli were selected based on four pilot studies (see
Garrido et al., in press for description). Each stimulus was shown once during the test
and trials were presented in a pseudo-random order over two blocks (29 trials per
block).
Cambridge Face Memory Test Long Form (CFMT+)
To test face recognition the performance of synaesthetes and non-synaesthetes
on the CFMT+ was compared (Russell, Duchaine, and Nakayama, 2009). The task is
an adapted version of the Cambridge Face Memory Test (Duchaine and Nakayama,
2006; Figure 5.1b) and was designed to distinguish normal from super-normal ability
at recognising faces (Russell et al., 2009). During the task subjects learn to recognize
six unfamiliar male faces from three different views (left 1/3 profile, frontal, right 1/3
profile) and are then tested on their ability to recognise these faces in a three-
alternative forced-choice task. The test is comprised of four sections, each more
difficult than the previous. The first three sections are taken from the original CFMT
(Duchaine and Nakayama, 2006) and the final section forms the longer CFMT+
(Russell et al., 2009).
The test begins by testing recognition with the same images that were used
during training (i.e. participants are are asked to memorise an unfamiliar male face
from three different views and are then tested on their memory for the trained face).
In the first phase each training image is shown for three seconds and immediately
followed by three trials per face, resulting in eighteen trials overall. This relatively
easy introduction is followed by a further training phase in which participants are
shown six frontal views of the faces for twenty seconds. The participants’ memory
for each face is then assessed using novel images that show the target faces from un-
trained views and lighting conditions (thirty trials). Following a further twenty

Chapter 5
100
second training phase in which participants are shown six frontal views of the face, a
third section consisting of novel images with visual noise added (twenty-four trials) is
completed. In the CFMT+, this is followed by a final section containing thirty very
difficult trials in which distracter images repeat much more frequently, targets and
distracters contain more visual noise than the images in the third section, cropped (i.e.
only showing internal features) and uncropped images (i.e. showing hair, ears, and
necks) are used, and images showing the targets and distracters making emotional
expressions are included. An inter-trial interval of 1 second is used throughout. The
percentage of correct responses for each section and overall are measured. Feedback
is not provided during the test.
Cambridge Face Perception Test (CFPT)
To investigate face perception the CFPT was administered (Duchaine,
Germine, and Nakayama, 2007). This test assesses the ability to perceive differences
between facial identities. Memory demands are minimal because faces are presented
simultaneously. During the task, subjects are shown a target face (from a ¾
viewpoint) and six faces (from a frontal view) morphed between the target and
distracter in varying proportions (88%, 76%, 64%, 52%, 40%, 28%) so that they vary
systematically in their similarity to the target face (Figure 5.1c). Subjects are asked to
sort the six faces by similarity to the target face and are given one minute to do so.
Participants sorted the faces by clicking on the face which they wished to move and
then indicating where the face should be by clicking in the area between two faces.
The desired face was then moved to the chosen location by the program. At the end
of each trial participants then clicked an option on screen to begin the next trial (i.e.
the task was self-paced). The task involves eight upright and eight inverted trials that
alternate in a fixed pseudo-random order. This allows investigation of the inversion

Chapter 5
101
effect for face perception. Performance is measured by an error score. This is
calculated by summing the deviations from the correct position for each face, with
one error reflecting each position that a face must be moved in order to be in the
correct location. For example, if a face was one position from the correct location
than this leads to an error score of one. If it is three positions away this is an error
score of three.
Same-Different Expression and Identity Matching Task
This experiment investigated participants’ abilities to match another’s facial
identity or facial expressions under identical experimental conditions.
In the expression matching task, participants were presented with a “sample”
face (250 msec) followed by a fixation cross (1000 msec), and a “target” face (250
msec). Participants were asked to indicate whether the target facial expression
matched, or was different to, the sample facial expression. On half of the trials, the
target and sample face expressed the same emotion and half the sample-target pairs
showed different emotions (Figure 5.1d). A total of 72 trials (split between 2 blocks)
were completed. Each image showed one of six female models making one of six
basic facial expressions: anger, disgust, fear, happiness, sadness or surprise. Each
stimulus was a greyscale image taken from the Ekman and Friesen (1976) facial affect
series. Stimuli were cropped with the same contour to cover the hair and neck using
Adobe Photoshop. In the expression task, identity always changed between sample-
target pairs and each expression was presented an equal number of times.
In the identity matching task, the same stimuli and procedure were used.
Participants were asked to indicate whether the sample and target face were the same
or a different person. Half of the trials showed pairs with the same identity and half

Chapter 5
102
with a different identity. Expression always changed between the sample and target
face, and the six models were presented an equal number of times.
Figure 5.1 Summary of the tasks used. (a) Films Facial Expression Task. This task investigated participants’ abilities to categorize the emotional expressions of others. Participants were presented with a target adjective describing an emotional state followed by three images shown consecutively for 500 msec each. Participants were asked which of the three images best portrayed the target emotion. In the actual task colour stimuli were used. (b) Cambridge Face Memory Test Long Form. This task investigated participants’ abilities to memorize facial identity and was derived from the Cambridge Face Memory Test (shown in figure). During the task participants memorized six unfamiliar male faces. They were then tested on their ability to recognize the faces in a three-alternative-forced-choice paradigm. The task involves four sections (for stimuli from the final section see Russell et al., 2009), each more difficult than the preceding section. (c) Cambridge Face Perception Test. This task investigated participants’ abilities to perceive faces in the absence of memory. Participants were shown a target face and six faces morphed between the target and a distracter face. Participants sorted the six faces by similarity to the target face. Faces were presented upright and inverted in a fixed pseudo-random order. (d) Same-Different Expression-Identity Matching Task. This task investigated participants’ abilities to match another’s facial identity or facial expressions. Participants were presented with a sample face followed by a fixation cross, and then a target face. In the expression matching task participants indicated whether the expression in the target face matched the expression in the sample face. In the identity matching task participants indicated whether the identity of the target face and the prime face matched.

Chapter 5
103
5.3 Results
Films Facial Expression Recognition
Accuracy and reaction time performance were compared separately using a
one way between subjects ANCOVA. Participant age was used a covariate on all
analyses because of slight trend for synaesthetes to differ from controls on age [t26 =
1.84, p = .078]. One control subject was withdrawn from analysis due to difficulties in
understanding the meaning of expression adjectives and performing more than three
standard deviations below the control group mean on accuracy and reaction time
measures.
Synaesthetes showed superior abilities at recognizing the emotional
expressions of others (Figure 5.2). Analysis of accuracy performance revealed that
mirror-touch synaesthetes outperformed control subjects on expression recognition
[F1,24 = 16.38, p = < .001] (Figure 5.2a). This difference was not due to a speed-
accuracy trade off as no significant effect of group (synaesthete or control) was found
for reaction time performance, and in fact synaesthetes tended to perform faster than
controls [F1,24 = .962, p = .336] (Figure 5.2b). These findings suggest that mirror-
touch synaesthetes show superior facial expression recognition, which may be due to
heightened sensorimotor simulation mechanisms.

C
hapt
er 5
104
Fi
gure
5.2
Mea
n ac
cura
cy (
a) a
nd r
eact
ion
time
perf
orm
ance
s (b
) of
syn
aest
hete
s an
d co
ntro
ls o
n th
e fi
lms
faci
al e
xpre
ssio
n ta
sk (
± s.
e.m
).
Syna
esth
etes
wer
e si
gnif
ican
tly m
ore
accu
rate
at
cate
goriz
ing
the
emot
iona
l fa
cial
exp
ress
ions
of
othe
rs (
a).
Thi
s w
as n
ot d
ue t
o a
spee
d ac
cura
cy tr
ade
off
as n
o si
gnif
ican
t diff
eren
ces
wer
e fo
und
for
reac
tion
time
and
syna
esth
etes
sho
wed
a tr
end
to b
e fa
ster
than
con
trol s
ubje
cts
(b).
75
80
85
90
95
Percentage Correct
Synaesthetes
Controls
*a
400
500
600
700
800
Reaction Time (msec)
Synaesthete
Control
b

C
hapt
er 5
105
Figu
re 5
.3 M
ean
accu
racy
per
form
ance
s (±
s.e
.m)
on t
he C
FMT
(a)
and
CFM
T+
(b)
for
syna
esth
etes
and
con
trols
. T
he p
erfo
rman
ce o
f sy
naes
thet
es a
nd c
ontr
ols
did
not s
igni
fica
ntly
dif
fer o
n th
e C
FMT
(fir
st th
ree
sect
ions
of t
ask)
or C
FMT+
.
70
75
80
85
Percentage Correct
Synaesthete
Controls
a
60
65
70
75
Percentage Correct
Synaesthete
Control
b

Chapter 5
106
Cambridge Face Memory Test Long Form
Accuracy performance from the Cambridge Face Memory Test (first three
sections) and Cambridge Face Memory Test Long Form are shown in figure 5.3. No
significant differences were observed between synaesthetes and controls on either the
CFMT [F1,25 = .023, p = .880] (Figure 5.3a) or the CFMT + [F1,25 = .095, p = .761]
(Figure 5.3b). Therefore unlike facial expression recognition, synaesthetes and
controls did not differ in their ability to memorize facial identity.
Cambridge Face Perception Test
Error scores on the eight upright and eight inverted trials were summed to
determine the total number of upright and inverted errors. A 2 (Group) x 2 (Trial
Type) ANCOVA revealed a significant effect of trial type [F1,25 = 5.81, p = .024]
which was due to an inversion effect, whereby overall participants were less accurate
on inverted (mean ± s.e.m = 70 ± 3) compared with upright trials (mean ± s.e.m =
41.5 ± 3.21). Importantly, this effect did not interact with group [F1,25 = .37, p = .549]
and no main effect of group was found [F1,25 = .253, p = .619] (Figure 5.4).
Therefore, unlike expression recognition, synaesthetes and controls did not
significantly differ in their abilities to detect another’s facial identity.

C
hapt
er 5
107
Figu
re 5
.4 M
ean
erro
r sco
re (±
s.e
.m) f
or s
ynae
sthe
tes
and
cont
rols
on
upri
ght (
a) a
nd in
verte
d (b
) tria
ls o
f the
CFP
T.
No
sign
ific
ant d
iffe
renc
es
wer
e fo
und
betw
een
the
perf
orm
ance
of
syna
esth
etes
and
con
trol
s on
a m
easu
re o
f fa
cial
ide
ntity
per
cept
ion.
Er
ror
scor
es a
re b
ased
on
the
devi
atio
n fr
om th
e ex
pect
ed c
orre
ct o
rder
ing
of th
e co
rrec
t loc
atio
n of
the
targ
et fa
ce.
Supe
rior p
erfo
rman
ce is
refl
ecte
d in
a lo
wer
err
or s
core
.
30
35
40
45
Upright Errors
Synaesthete
Control
a
65
70
75
80
Inverted Errors
Synaesthete
Control
b

C
hapt
er 5
108
Figu
re 5
.5 M
ean
accu
racy
per
form
ance
s (±
s.e
.m) o
n th
e ex
pres
sion
mat
chin
g (a
) and
iden
tity
mat
chin
g (b
) tas
k fo
r syn
aest
hete
s an
d co
ntro
ls.
A
2 (T
ask)
x 2
(G
roup
) A
NC
OV
A r
evea
led
a si
gnif
ican
t ta
sk x
gro
up i
nter
actio
n. O
n th
e ex
pres
sion
tas
k, t
here
was
a t
rend
for
syn
aest
hete
s to
ou
tper
form
con
trols
whe
reas
on
the
iden
tity
task
, the
re w
as a
tren
d fo
r con
trols
to o
utpe
rfor
m s
ynae
sthe
tes.
With
in-g
roup
com
pari
sons
bet
wee
n th
e ta
sks
reve
aled
that
con
trols
wer
e si
gnif
ican
tly m
ore
accu
rate
in th
e id
entit
y co
mpa
red
to th
e ex
pres
sion
task
. Sy
naes
thet
es d
id n
ot s
how
this
bi
as to
war
ds id
entit
y m
atch
ing
- exp
ress
ion
and
iden
tity
mat
chin
g pe
rfor
man
ces
wer
e co
mpa
rabl
e.
70
75
80
85
Percentage Correct
Synaesthete
Controls
aExpression matching
75
80
85
90
Percentage Correct
Synaesthete
Control
bIdentity matching

Chapter 5
109
Same-Different Expression and Identity Matching Task
A 2 (Group) x 2 (Task) mixed ANCOVA was conducted. Participant age was
used as a covariate. No main effect of task or group was found. No relationship
between task and age was observed. There was however a significant interaction
between task and group [F1,25 = 4.507, p = .044]. Controls were more accurate, and
therefore showed an advantage, on the identity matching task relative to the emotion
matching task [F1,18 = 5.10, p = .037]. Synaesthetes did not show this pattern -
analysis of within-subject effects revealed no significant difference between the two
tasks for the synaesthetic group [F1,6 = .759, p = .417]. There was also a non-
significant trend for synaesthetes to outperform controls on the expression matching
task (Figure 5.5a), but for controls to outperform synaesthetes on the identity
matching task (Figure 5.5b).
5.4 GENERAL DISCUSSION
This study investigated expression and identity face processing in mirror-touch
synaesthetes and non-synaesthete control participants. It was predicted that
heightened sensorimotor simulation mechanisms would result in superior expression
recognition, but would not affect the identity recognition abilities of mirror-touch
synaesthetes. Consistent with these predictions, mirror-touch synaesthetes were
superior when recognizing the facial expressions, but not facial identities of others.
These findings are consistent with simulation accounts of expression recognition
which suggest that in order to understand another’s emotional expressions individuals
must simulate the sensorimotor response associated with generating the perceived
facial expression (Adolphs, 2002; Adolphs, 2003; Gallese, Keysers, and Rizzolatti, G,
2004; Goldman, and Sripada, 2005; Keysers and Gazzola, 2006).

Chapter 5
110
A variety of sources indicate that recognizing another’s identity and
expressions relies upon multiple stages of representation, including purely visual,
multimodal, expression-general and expression-specific mechanisms (e.g. Adolphs et
al., 2000; Anderson, Spencer, Fulbright, and Phelps, 2000; Calder, Lawrence, and
Young, 2001; Calder, Keane, Lawrence, and Manes, 2004; Lawrence, Calder,
McGowan, and Grasby, 2002; Lewis et al., 2003; Keane, Calder, Hodges, and Young,
2002; Pitcher et al., 2008; Sprengelmeyer et al., 1996). Simulation accounts of
expression recognition contend that one mechanism involved in expression, but not
identity, recognition is an internal sensorimotor re-enactment of the perceived
expression (Adolphs, 2002; Adolphs, 2003; Gallese, Keysers, and Rizzolatti, 2004;
Goldman, and Sripada, 2005; Keysers and Gazzola, 2006). Functional brain imaging
(Carr et al., 2003; Hennenlotter et al., 2005; Montgomery and Haxby, 2008; van der
Gaag, et al., 2007; Winston et al., 2003), neuropsychological (Adolphs et al., 2000),
and transcranial magnetic stimulation studies (Pitcher et al., 2008) suggest a key role
for somatosensory resources in expression recognition. The findings that individuals
who show increased levels of somatosensory simulation (mirror-touch synaesthetes)
demonstrate superior expression, but not identity perception, are consistent with this
view. The task specific nature of the findings also indicate that the superior
performance shown by mirror-touch synaesthetes on the expression recognition tasks
are not linked to heightened motivation on the part of the synaesthetic subjects.
The experiments in chapter 4 documented that mirror-touch synaesthetes show
heightened emotional reactivity compared to controls, but do not differ on other
components of empathy. The findings from the current investigation indicate that
mirror-touch synaesthesia is not only linked with some components of empathy, but
also with superior emotion recognition. It remains to be established whether

Chapter 5
111
heightened emotion sensitivity displayed by mirror-touch synaesthetes is a cause or
consequence of this type of synaesthetic experience. While it is assumed that mirror-
touch synaesthetes form part of the synaesthetic population, and are therefore a unique
group of subjects, the principles which bias what type of synaesthesia will or will not
be developed are a matter of debate (Bargary and Mitchell, 2008; Cohen Kadosh,
Henik, and Walsh, 2009; Cohen Kadosh and Walsh, 2008; Grossenbacher and
Lovelace, 2001; Hubbard and Ramachandran, 2005; Ramachandran and Hubbard,
2001; Rouw and Scholte, 2007; Sagiv and Ward, 2006). Conceptually there are two
possibilities: i) mirror-touch synaesthetes reflect the top end of a spectrum along
which emotion sensitivity ranges (e.g. the ‘normal’ architecture for multi-sensory
interactions) and this biases them towards interpersonal synaesthetic experience, or ii)
mirror-touch synaesthetes are a unique population whose extra sensory experiences
predispose superior emotion sensitivity.
In sum, this study demonstrates that mirror-touch synaesthesia is associated
with superior facial expression recognition abilities. The observed superiority in face
processing is restricted to expression recognition. Mirror-touch synaesthetes show
enhanced emotional expression abilities. They did not differ from controls on identity
perception measures. Given that mirror-touch synaesthesia has been linked to
heightened somatosensory simulation these findings are consistent with simulation
based accounts of expression recognition and indicate that somatosensory resources
are an important facet in our ability to recognise the emotions of others.

Chapter 6
112
CHAPTER 6: METHODOLOGICAL INTRODUCTION TO TMS
This chapter outlines the methodological principles for using transcranial magnetic
stimulation (TMS) to disrupt normal cognitive functioning. The main principles,
ethical considerations, spatial and temporal constraints, and types of TMS are
discussed. In chapters 7 and 8 continuous theta burst TMS was performed to
investigate the role of sensorimotor simulation in expression recognition. This TMS
paradigm is introduced here and it’s spatial and temporal effects are discussed.
6.1 INTRODUCTION
Transcranial magnetic stimulation (TMS) is a non-invasive experimental
technique that is capable of suppressing or facilitating activity in the brain. The effect
of this modulation of neural activity can be measured using a variety of standard
behavioural (e.g. reaction time, accuracy, thresholds) and physiological (e.g. evoked
potentials, functional brain imaging) measures; is temporally discrete (c.f. Walsh and
Cowey, 2000); and shows good spatial specificity (e.g. Pitcher, Charles, Devlin,
Walsh, and Duchaine, 2009). The technique provides a unique tool to the cognitive
neuroscientist because it permits the opportunity to assess the causality of a particular
brain region to a given cognitive task. For example, one can use TMS to disorganize
neural activity in a given brain region and investigate the effects of this disruption on
functionally specific cognitive tasks (e.g. motion priming and human V5 / MT
stimulation – Campana, Cowey, and Walsh, 2002; face processing and Occipital Face
Area stimulation – Pitcher, Walsh, Yovel, and Duchaine, 2007). In this sense, TMS is
similar to both human (e.g. Milner, 1966; Shallice, 1988) and animal (e.g. Cowey and
Gross, 1970; Walsh and Butler, 1996) lesion studies in which one is able to make
inferences about the necessity of specific brain areas based upon impairment to
cognitive functioning.

Chapter 6
113
There are also important differences between lesion and TMS studies, which
lead to a number of benefits over patient-based research. Firstly, the nature of lesion
studies means that the experimenter is required to make inferences about the normal
architecture based on an abnormal system. This has a number of caveats including the
influence of mechanisms of compensatory plasticity in the abnormal system, which
may lead to changes in function or task performance. For example, it is often months
after brain injury that the experimenter is able to examine patient performance
systematically and it is difficult to disentangle whether one is measuring the removal
of a region or the ability of other brain regions to compensate the function being
investigated (Lomber, 1999; Robertson and Murre, 1999). Further, brain lesions are
rarely spatially discrete and removal of a brain area may also incur damage at distal
sites (e.g. via severed vessels, ablated white matter; Robertson and Murre, 1999;
Walsh and Pascual-Leone, 2003). In contrast to this, TMS permits investigation of
spatially specific brain regions in normal subjects and overcomes problems of neural
compensation / reorganization because the main effects occur in a discrete temporal
window (lasting a few tens of milliseconds to minutes depending on the type of
stimulation used). In addition, because behavioural performance can be measured
within-subjects (during both the application and the absence of stimulation), it is
possible for subjects to act as their own control group, thereby strengthening the
validity of the conclusions that can be drawn from a TMS experiment.
To consider the method further, this chapter discusses the principles, ethical
aspects, spatial resolution, temporal constraints, and alternative TMS paradigms.

Chapter 6
114
6.2 What is TMS?
Attempts to modulate human brain function using magnetic fields can be
traced back to the late 19th century and developed over the next 100 years (c.f. Walsh
and Pascual-Leone, 2003). However, these attempts rarely systematically measured
the effects of magnetic stimulation and it was not until the 1980s that TMS (as we
know it) was first introduced as a tool for cognitive neuroscience - Barker and
colleagues (1985) reported the first successful attempts to disrupt normal cortical
functioning when they applied magnetic stimulation over the motor cortex in human
subjects and recorded the resulting muscle twitches via motor evoked potentials
(MEPs).
The technique builds upon Faraday’s observations of electromagnetic
induction (1831), in which electric current passed along one wire coil (coil A)
generates a magnetic field that induces electrical current in another wire coil (coil B).
In TMS, rapidly changing electrical current is passed through a coil (i.e. coil A in
Faraday example) located on the participant’s scalp to generate a brief magnetic field
which passes through the skull and induces electrical fields in the underlying cortex
(i.e. coil B in Faraday example). The induced current alters the electrical state inside
and outside of nerve axons, leading to membrane depolarisation and the initiation of
action potentials (Nagarjan, Durand, and Warman, 1993). Thus, the electrical field
induced by the TMS coil causes changes in the resting potential of the underlying
neurons and effectively disorganises neural processing in a stimulated cortical region.
The duration of this disorganisation of neural processing is linked to the size
of the induced current, which is related to the amplitude and the rate of change of the
current passing through the TMS coil. Typically the current in the coil is large, up to 8
kiloamperes (kA), with a swift rise time of roughly 200 milliseconds (ms) and an

Chapter 6
115
overall duration of roughly 1 ms (Figure 6.1). The extent to which the resulting TMS
pulse disrupts neural processing in the targeted area depends on the orientation of the
coil and the orientation of the underlying nerve fibres (Amassian, Eberle, Maccabee,
and Cracco, 1992). If the induced field is uniform across the cell membrane then no
current will be induced. The TMS effects are optimised when the electric field is
tangential to the orientation of the nerve fibre either due to the electric field
orientation being perpendicular to a straight axon or an axon bending relative to the
orientation of the induced field (Figure 6.2).
Two types of coils are typically used in TMS studies. They are the figure-of-
eight and circular coils (Figure 6.3). All of the studies reported in this thesis use a
figure-of-eight coil, which has been shown to produce the most focal effects of TMS
(Ueno, Tashiro, and Harada, 1988). In the figure-of-eight coil, current flows in
opposite directions around each of the windings and converges on the centre point of
the coil where the electrical currents summate. This leads to focal neural stimulation
with the largest effect occurring in the cortex situated directly under the centre-point
of the coil. Because the outer-windings of the coil are away from the surface of the
scalp they are unlikely to induce an additional disruptive magnetic field. The
stimulation effects dissipate gradually as distance from the maximal point increases
(Figure 6.3).

Chapter 6
116
Figure 6.1 The sequences of events for a transcranial magnetic stimulation pulse (taken with permission from Walsh and Cowey, 2000). (a) A capacitor generates an electric current (up to 8kA). (b) This discharges into the TMS coil generating a magnetic pulse of up to 2 T. (c) The pulse has a rise time of roughly 200ms and lasts for 1ms, which changes rapidly due to its intense and brief nature. (d) The magnetic field generates an electrical field. (e) The magnetic field causes neural activity or changes in the resting potential of the underlying neurons. Note that monophasic pulses are shown in the figure (but biphasic pulses are used for repetitive TMS).

Chapter 6
117
Figure 6.2 The effects of fibre orientation and electric field orientation for the application of a TMS pulse.
Figure 6.3 TMS-induced electrical fields produced by circular (top left) and figure-of-eight (top right) shaped coils. Maximal intensity with a circular coil is located directly under the winding, but with a figure-of-eight coil it is at the intersection of the two windings. The intensity of the induced current dissipates with a radial distance from the area of maximum intensity (diagram taken from the Magstim Guide to Magnetic Stimulation).

Chapter 6
118
6.3 The spatial resolution of TMS
Figure 6.4 illustrates the spatial and temporal specificity of TMS in relation to
other experiment methodologies.
Figure 6.4 The spatial and temporal resolution of TMS compared with other experimental techniques. TMS benefits from high spatial and temporal resolution and is capable of interfering with brain function (taken with permission from Walsh and Cowey, 2000).
As noted in the last section, the magnetic field induced by TMS dissipates
gradually as distance from the maximal point of stimulation decreases. This raises a
concern of how confident one can be in the spatial and functional specificity of TMS.
Put another way, how can one be sure that the effects observed in a TMS study are
due to stimulation of the target region or due to more widespread dissipating effects of
the induced magnetic field. The answer to this lies in a number of converging lines of
evidence which demonstrate that while in theory, the magnetic field induced by TMS

Chapter 6
119
is infinite (with the induced electrical field decreasing from the centre of the
stimulation focal point), in practice the size of the electrical field capable of disrupting
normal neuronal activity is limited (i.e. it is anatomically and functionally specific).
For example, TMS of the motor cortex results in visible and measurable (via MEPs)
motor twitches in the muscles. This effect however is neither random nor non-
specific – TMS to the motor cortex has been shown to evoke muscular twitches from
selective muscle groups in a manner compatible with the functional layout of the
motor homunculus, with stimulation at target sites varying from 0.5 to 1 centimetres
apart leading to selective activation of each muscle type (Singh, Hamdy, Aziz, and
Thompson, 1997; Wassermann, McShane, Hallet, and Cohen, 1992). The functional
focality of the method is further demonstrated in the visual domain, where a similar
spatial resolution (less than 1 cm on the scalp) has been reported through the study of
the physiological effects induced by TMS stimulation of the primary visual cortex
(V1; c.f. Walsh and Cowey, 2000). Moreover, stimulation to V1 leads to the
generation of phosphenes, which are spatially distributed in a manner that corresponds
with the retinotopic organisation of V1 (Kammer, 1999).
Outside of the primary motor and sensory areas, the effective spatial
resolution3 of TMS has regularly been demonstrated by functional dissociations
following TMS to spatially discrete regions of the cortex. For example, in the same
subjects, TMS targeted at the right occipital face area (thought to be functionally
specialised for face processing) has been shown to impair face but not body or object
recognition, while stimulation at a region of lateral occipital cortex (LO; thought to be
functionally specialised for object processing) has been shown to impair object but
not face or body recognition, whilst stimulation at the right extra-striate body area 3 It is of note, that TMS does not only stimulate the neuron in a 1cm region, rather, it is that this represents the physiologically effective resolution of TMS.

Chapter 6
120
(EBA; thought to be functionally specialised for body processing) has been shown to
impair body but not face or object recognition (Pitcher et al., 2009) – this dissociation
is particularly striking given the anatomical adjacency between these regions.
Functional dissociations such as the triple dissociation described above demonstrate
the functional focality of TMS and have been regularly observed across a variety of
domains (e.g. Ashbridge, Walsh, and Cowey, 1997; Stewart, Walsh, Frith, and
Rothwell, 2001).
A further approach to assess the spatial specificity of TMS is to combine the
approach with methods such as fMRI, MEG or PET. To date, studies that have
combined these methodologies have demonstrated a good correspondence between
TMS defined functional regions and the areas revealed with high spatial resolution
brain imaging techniques (Bestmann, Baudewig, Siebner, Rothwell, and Frahm, 2004;
Bohning, Shastri, McConnell, Nahas, Lorberbaum, and Roberts, 1999; Ruff et al.,
2006; Siebner et al., 1998; Terao et al., 1998).
In sum, while the effects of TMS will dissipate from the targeted region to
other cortical areas, the functionally effective resolution is much more discrete
(approximately 1cm). This has been demonstrated across a variety of studies by
systematically measuring the effect on behaviour as the coil is moved away from an
optimal stimulation site (e.g. by stimulating adjacent areas of cortex [Figure 6.5]
which demonstrate functionally different characteristics or examining direct
physiological effects).

Chapter 6
121
Figure 6.5 Functional dissociation method that can be employed using TMS (taken with permission from Walsh and Cowey, 2000). Using TMS one can infer the importance of a targeted region of stimulation for a particular function. Stimulation to an adjacent or functionally alternative region enables refined inferences to be made, including functional-anatomical attributions. The ‘coils’ and induced fields in this figure are illustrative of the methodological rationale and do not represent real configurations and effects. 6.4 Offline and online TMS: The temporal resolution of TMS
When considering the temporal effects of TMS a distinction needs to be drawn
between offline and online paradigms. Online paradigms involve stimulation
concurrent with behavioural performance (i.e. when a participant is doing a task) and
have short-lasting effects (e.g. 1 msec per pulse), while offline paradigms take place
prior to task performance and have more long-lasting effects during an experimental
session (e.g. 20-60 minutes of altered brain activity).
For online TMS experiments, a typical single pulse of TMS is very brief,
around 1ms. An important consideration for online TMS experiments is when is the
most appropriate time window for neural disruption to produce behavioural
impairment. A number of approaches have been developed to address this, including
delivering single (Amassian, Cracco, Maccabee, Cracco, Rudell, and Eberle, 1993) or
paired-pulses (Juan and Walsh, 2003; O’Shea, Muggleton, Cowey, and Walsh, 2004)
of TMS to the target region at different time points after stimulus onset or the start of

Chapter 6
122
trial. The later has the advantage of reducing the number of temporal conditions in a
TMS timing experiment.
It is also possible that the disruption induced by multiple pulses of TMS will
summate, therefore inducing larger behavioural impairments. The potential for
summation of the disruptive effect has been further exploited by repetitive TMS
(rTMS) protocols. For example, Rushworth and colleagues (2001) delivered TMS at
a frequency of 10 Hz for 500 ms and showed dissociations between parietal regions
(supramarginal gyrus and angular gyrus) for mediating modality-specific attentional
processes. To date there is no corroborating physiological evidence that the five
pulses of TMS actually do summate, but despite this the approach has proven to be a
robust online TMS protocol for demonstrating function-specific involvement of a
wide variety of cortical areas across a number of domains (e.g. Beck, Muggleton,
Walsh, and Lavie, 2006; Bjoertomt, Cowey, and Walsh, 2002; Campana et al., 2002;
Lavidor and Walsh, 2003; Pitcher et al., 2008; Pitcher et al., 2009; Wig, Grafton,
Demos, and Kelley, 2005).
In contrast to online TMS paradigms which rely upon observing effects in
very short-lasting temporal windows, a recently introduced paradigm of offline
continuous theta-burst stimulation (cTBS) provides a more long lasting window for
one to examine changes in cortical function on behaviour. cTBS is a form of rTMS
based on the burst patterns used to induce long lasting changes in synaptic
effectiveness in animal experiments. The approach uses high frequency stimulation
bursts (3 pulses at 50Hz), which are repeated at intervals of 200 milliseconds (i.e.
5Hz). In the motor system cTBS to M1 suppresses the excitability of motor cortical
circuits for 20-60 minutes depending on the cTBS parameters used (Di Lazzoro et al.,
2005; Di Lazzoro et al., 2008; Huang, Edwards, Rounis, Bhatia, and Rothwell, 2005;

Chapter 6
123
Huang et al., 2009; Stefan, Gentner, Zeler, Dang, and Classen, 2008). In paradigms in
which 20 seconds of cTBS (300 TMS pulses) is applied to the motor cortex, MEP
amplitude is reduced for approximately 20-30 minutes; in paradigms in which 40
seconds of cTBS (600 TMS pulses) is applied MEP amplitude is reduced for
considerably longer (up to one hour; Huang et al., 2005). This rapid method of
suppressing cortical activity offers a unique opportunity to examine the functional role
of a given brain region on behaviour in a relatively large time window. It also
overcomes a number of potential confounds related to online stimulation, including
the peripheral and proprioceptive effects of online TMS (e.g. muscular twitching;
Terao et al. 1997) that may impact on task performance. The potential for this
paradigm to be used to study cognitive processing has recently been fulfilled in
several domains. Firstly, Vallesi and colleagues (2007) used cTBS to show a critical
role for the right dorsolateral prefrontal cortex, but not left dorsolateral prefrontal
cortex or right angular gyrus, in temporal processing. More recently, Kalla and
colleagues used cTBS targeted at the dorsolateral prefrontal cortex to demonstrate the
necessity of this brain region, but not the vertex or MT/V5 (TMS to MT / V5
facilitated processing), in conjunction visual search (Kalla, Muggleton, Cowey, and
Walsh, 2009). The studies reported in chapters 7 and 8 used cTBS to investigate the
role of sensorimotor cortices in expression recognition from visual and auditory cues.
6.5 The safety of TMS as an experimental tool
The primary concern in any TMS experiment is the health and safety of the
subjects (c.f. Wasserrmann, 1998 for detailed safety guidelines). The main ethical
concern in the use of TMS is the possible side effects of this method. TMS is
sometimes associated with minor discomfort, muscular pain, and occasionally mild
headache. These are all treatable with simple pain killers such as aspirin, and any

Chapter 6
124
discomfort can mostly be alleviated by repositioning the TMS coil. All volunteers
should be informed of these possible effects and they should be minimised in each
individual. As with all studies, participants should be informed that they can withdraw
from the experiment at any time, without having to give a reason why. Sessions in
which subjects are perceived to be uncomfortable but do not report this should also be
terminated by the experimenter. The concern most commonly associated with TMS is
that in rare circumstances TMS has induced seizures. These are most likely to occur in
individuals already susceptible to seizures (i.e. with a history of epilepsy) and those
taking neuroleptic medication (Stewart, Ellison, Walsh, and Cowey, 2001). For the
studies reported in this PhD, only healthy adult subjects (aged from 18-50 years), with
no psychiatric or neurological history and no family history of seizures took part in
the proposed research. All safety guidelines regarding the use of TMS (Wassermann,
1998) were followed and all the experiments reported in the thesis were approved by
the local ethics committee at University College London.

Chapter 7
125
CHAPTER 7: THE ROLE OF SENSORIMOTOR SIMULATION
IN AUDITORY EMOTION DISCRIMINATION
Functional neuroimaging studies indicate that activity in primary somatosensory and
premotor cortex is evoked during the perception of emotion. In the visual domain,
right somatosensory cortex activity has been shown to be critical for facial emotion
recognition. However, the importance of these representations in modalities outside
of vision remains unknown. This study used continuous theta-burst transcranial
magnetic stimulation (cTBS) to investigate whether neural activity in the right
primary somatosensory cortex (rSI) and right lateral premotor cortex (rPM) is
central to non-verbal auditory emotion recognition. Two groups of participants
completed same-different tasks on auditory stimuli, discriminating between either the
emotion expressed or the speakers’ identities, prior to and following cTBS targeted at
rSI, rPM or the vertex (control site). A task-selective deficit in auditory emotion
discrimination was observed. Stimulation to rSI and rPM resulted in a disruption of
participants’ abilities to discriminate emotion, but not identity, from vocal signals.
7.1 Introduction
Our ability to recognise the emotions of others is a crucial feature of human
social cognition. The neurocognitive processes which underpin this have recently
been described as being achieved through simulation processes (Adolphs, 2002;
Adolphs, 2003; Damasio, 1990; Gallese, Keysers, and Rizzolatti, 2004; Goldman and
Sripada, 2005; Keysers and Gazzola, 2006). These models suggest that understanding
another’s emotions requires individuals to map the observed state onto their own
representations which are active during the experience of the perceived emotion. The
discovery of mirror neurons in the primate brain (Gallese, Fadiga, Fogassi, and
Rizzolatti, 1996), and evidence of not only a ‘classical’ action mirror system (Buccino
et al., 2001; Fadiga, Fogassi, Pavesi, and Rizzolatti, 1996; Gazzola, Aziz-Zadeh, and
Keysers, 2006), but also ‘extended’ mirror systems in the human brain (involved in
mirroring sensation and emotion; Avenanti, Bueti, Galati, and Aglioti, 2005;
Blakemore, Bristow, Bird, Frith, and Ward, 2005; Carr, Iacoboni, Dubeau, Mazziotta,
and Lenzi, 2003; Keysers, Wicker, Gazzola, Anton, Fogassi, and Gallese, 2004;

Chapter 7
126
Singer, Seymour, O’Doherty, Kaube, Dolan, and Frith, 2004; Wicker, Keysers¸
Plailly, Royet, Gallese, and Rizzolatti, 2003) has provided a candidate
neurophysiological mechanism for such shared representations in emotion
recognition. These brain regions may, in part, aid emotion recognition because they
enable the observer to match the observed emotion within cortical areas active during
the observer’s own experience of the perceived emotion (Carr et al., 2003;
Hennenlotter et al., 2005; Jabbi, Swart and Keysers, 2007; Leslie, Johnsen-Frey, and
Grafton, 2004; van der Gaag, Minderaa, and Keysers, 2007). Consistent with this,
functional brain imaging studies indicate that components of classical and extended
mirror systems (including premotor cortex and primary somatosensory cortex) are
recruited when perceiving others’ facial emotions (Hennenlotter et al., 2005; Leslie,
Johnsen-Frey, and Grafton, 2004; Montgomery and Haxby, 2008; van der Gaag,
Minderaa, and Keysers, 2007); that primary somatosensory cortex is activated when
judging another’s facial emotion (Winston, O’Doherty, and Dolan, 2003); and that the
auditory-motor mirror system is activated during the perception of non-vocal emotion
expressions (e.g. hearing somebody laughing; Warren et al., 2006). Further, in
chapters 4 and 5 I show that facilitated sensorimotor simulation (in mirror-touch
synaesthesia) is linked to heightened emotional empathy and emotional expression
recognition. In attempt to assess what impact suppressing sensorimotor activity has
on the expression recognition abilities of healthy adults, here (and in chapter 8) I use
TMS in non-synaesthetes to assess whether sensorimotor activity plays a central role
in our ability to recognize the emotions of others.
In the visual domain, there is growing evidence that sensorimotor activity
plays a causal role in facial emotion recognition. Neuropsychological findings
indicate that deficits in the recognition of facial affect are related to damage within

Chapter 7
127
right hemisphere somatosensory-related cortices (Adolphs, Damasio, Tranel, Cooper,
and Damasio, 2000) and transcranial magnetic stimulation (TMS) findings in healthy
adults are consistent with this (Pitcher, Garrido, Walsh and Duchaine, 2008). In the
study by Pitcher and colleagues (2008), rTMS targeted at right primary
somatosensory cortex resulted in a disruption of participants’ abilities to discriminate
the facial expressions, but not facial identities, of others. It remains unclear if neural
activity within these systems is necessary for the recognition of affect from alternative
modalities. Moreover, if sensorimotor resources are vital for global processing of
emotion then these resources should also be central for emotion recognition abilities
of healthy adults in modalities outside of visual perception. An example of this would
be in the auditory domain. Primates are highly sensitive to vocal cues and the
affective contents of vocal signals are reliably recognised by humans (Bryant and
Barett, 2007; Sauter and Scott, 2007; Schröder, 2003). An fMRI study indicates that
adult human listeners activate the sensorimotor cortices when listening to emotional
vocalisations of others (Warren et al., 2006), however whether this activity is
necessary for affect recognition remains unknown. To address this, the studies
presented in this chapter use continuous theta burst TMS (cTBS; Di Lazzaro et al.,
2005; Huang, Edwards, Rounis, Bhatia, and Rothwell, 2005; Vallesi, Shallice, and
Walsh, 2007; Kalla, Muggleton, Cowey, and Walsh, 2009), an offline (i.e. conducted
while the participant is at rest) TMS paradigm following which neural activity may be
suppressed for several minutes (Di Lazzaro et al., 2005; Huang et al., 2005), to
examine whether neural activity in the right lateral premotor (rPM) and right primary
somatosensory cortex (rSI) is involved in discriminating affect from vocal signals.
Right hemisphere representations were selected based on previous fMRI,
neuropsychological and TMS findings demonstrating the importance of right

Chapter 7
128
hemisphere activity in affect recognition (e.g. Adolphs et al., 2000; Mitchell and
Crow, 2005; Pitcher et al., 2008; Pourtois et al., 2004; Van Lancker and Fromkin,
1973).
Two experiments were conducted. Experiment 1 sought to establish the
effects of cTBS targeted at rPM, rSI, or the vertex (cTBS control site) on participants’
abilities to complete a same-different auditory emotion recognition task (Figure 7.1;
Figure 7.2). Non-verbal emotional vocalisations (such as laughter or screams) were
used. These vocalisations are reliably recognised by human listeners (Sauter and
Scott, 2007; see also Meyer, Zysset, von Cramon, and Alter, K, 2005; Schröder, 2003)
and can be considered to be closer to emotional facial expressions than emotional
speech because they do not contain the segmental structure of emotionally inflected or
nonsense speech (Dietrich, Szameitat, Ackermann, and Alter, 2006; Scott, Sauter, and
McGettigan, in press; Scott, Young, Calder, Hellawell, Aggleton, and Johnsen, 1997).
In experiment 2, identical stimuli and cTBS parameters were used, but a new group of
participants were instructed to complete a same-different auditory identity
discrimination task. This enabled examination of any non-specific effects of cTBS
and whether the effects observed in experiment 1 were selective to affective
processing. Based on simulation accounts of emotion recognition it was predicted
that cTBS targeted at rPM and rSI would result in a disruption of participants’ ability
to discriminate the auditory emotions, but not identity, of others.
7.2 Methods
Participants
Twenty healthy naïve adult participants, 11 female and 9 male (aged 20 to
35years), took part in the study. All were right handed, had normal or corrected-to-
normal vision, and gave informed consent in accordance with the ethics committee of

Chapter 7
129
University College London. Ten participants took part in each experiment
(Experiment 1: 6 female and 4 male aged 20 to 30 years; Experiment 2: 5 female and
5 male aged 20 to 35 years).
Materials
Identical stimuli were used in experiments 1 and 2. Stimuli were one of four
categories of non-verbal auditory emotions (amusement, sadness, fear, or disgust).
These stimuli were adapted from a previously validated set of non-verbal
vocalisations (Sauter, PhD thesis, University of London, 2006; Sauter and Scott,
2007; Sauter and Eimer, in press; Warren et al., 2006) and two of these emotions
(amusement and fear) were adapted (trimmed to 500msec) from stimuli used in a
previous fMRI study investigating the role of sensorimotor resources in non-verbal
auditory emotion perception (Warren et al., 2006). Ten stimuli, produced by four
different actors (two male / two female), per emotion type were used. All were 500
milliseconds in duration and were presented aurally via headphones.
Procedure
Both experiments consisted of three testing sessions conducted over three non-
consecutive days. At each testing session one of the three brain regions was
stimulated (rSI, rPM or Vertex). The order of site of stimulation was pseudo-
randomised between participants in an ABC-BCA-CAB fashion. Participants
completed the experimental task twice within each session, one run prior to cTBS
(baseline performance) and the other following cTBS.
In experiment 1, the task comprised of 120 trials (preceded by 20 practice
trials) divided between two blocks of 60 trials. Each trial began with the presentation
of a fixation cross (1500 ms) followed by the presentation of the prime stimulus. 500

Chapter 7
130
milliseconds after the offset of the prime stimulus a second emotion was presented
aurally. Concurrent with the presentation of the second emotion, participants were
asked to indicate if the second non-verbal emotion was the same or different from the
first using a key press (Figure 7.1). The need for speed and accuracy were
emphasised. Each block lasted approximately 10-15 minutes.
In experiment 2, the same stimuli and paradigm were used, but participants
were instructed to indicate if the prime and target emotions were expressed by the
same or different person.

Chapter 7
131
Figure 7.1 Summary of cTBS and task protocol. Participants completed a same-different auditory emotion (Experiment 1) or identity (Experiment 2) matching task. Both experiments consisted of three testing sessions conducted over three non-consecutive days. At each testing session one of the three brain regions was stimulated (rSI, rPM or the vertex). In each session, the task was completed prior to and 5 minutes following cTBS to each site. The 5 minute rest period was based on the observed time course of effects seen in the motor cortex. During the task, trials began with a fixation cross followed by the presentation of a prime emotion via headphones. Non-verbal auditory emotions of amusement, sadness, fear or disgust were used. After 500 milliseconds a second emotional expression was presented which was either the same or different from the target. In experiment 1, participants were asked to indicate whether the emotional expression was the same or different to the prime using a key press. In experiment 2, a new group of participants were asked to indicate whether the emotional expressions were produced by the same or different person. The need for speed and accuracy were emphasised.
TMS parameters and coregistration
TMS was delivered via a figure of eight coil with a 70mm diameter using a
Magstim Super Rapid Stimulator (Magstim, UK). An offline cTBS paradigm was
used, which consisted of a burst of 3 pulses at 50Hz repeated at intervals of 200ms for
20 seconds, resulting in a total of 300 pulses. This paradigm was used to prevent any
influence of online auditory and proprioceptive effects of TMS on task performance

Chapter 7
132
(Terao et al., 1997). Based upon previous findings (Di Lazzaro et al., 2005; Huang et
al., 2005; ; Kalla et al., 2009; Valessi et al., 2007) the time window of reduced
excitability following theta burst stimulation was expected to last between 20-30
minutes and a 5 minute rest period after stimulation offset was implemented for each
site stimulated.
TMS machine output was set to 80% of each participant’s motor threshold
with an upper limit of 50% of machine output. Motor threshold was defined using
visible motor twitch of the contralateral first dorsal interosseus following single pulse
TMS delivered to the best scalp position over motor cortex. Motor threshold was
calculated using a modified binary search paradigm (MOBS [Tyrell and Owens, 1988;
see also Thilo, Santoro, Walsh and Blakemore, 2004 for example use). For each
subject, motor threshold was calculated following pre-cTBS baseline and prior to
coregistration.
Locations for cTBS were identified using Brainsight TMS-magnetic resonance
coregistration system (Rogue Research, Montreal, Canada). FSL software (FMRIB,
Oxford) was used to transform coordinates for each site to each subject’s individual
MRI scan (Figure 7.2). Coordinates for rSI (27, -27, 69) were taken from Blakemore
and colleagues (2005) and were averages for twelve neurologically normal
participants in an fMRI study following touch to their own face. The coordinates for
rPM (54, -2, 44) were the averages of neurologically normal participants in an fMRI
study of non-verbal auditory emotion processing (Warren et al., 2006). The vertex
was identified as the point midway between the inion and the nasion, equidistant from
the left and right intertragal notches.

Chapter 7
133
Figure 7.2 Summary of TBS sites stimulated, rSI (a), rPM (b). Locations of TMS were determined using the Brainsight coregistration system. The co-ordinates for rSI (27, -27, 69) were taken from a study on the tactile mirror-system (Blakemore et al., 2005), the co-ordinates for rPM (54 -2 44) were taken from a study on the role of sensorimotor resources in auditory emotion recognition (Warren et al., 2006). cTBS parameters were used to stimulate each site. To ensure that any differences observed were not due to non-specific effects of cTBS, the vertex was stimulated as a TMS control site.
7.3 Results
Reaction times were trimmed prior to analysis (± 3 standard deviations and all
errors removed) and were corrected for accuracy (mean RT divided by percentage of
correct).
The role of r SI and rPM in recognizing emotions and identity from auditory cues
Baseline performance did not differ significantly across sites in either task
(Emotion task group: F(2,18) = 1.64, n.sig; Identity task group: F(2,18) = .815, n.sig).
To assess the effects across tasks and across sites, the difference between the post

Chapter 7
134
cTBS and pre-cTBS baseline reaction times (i.e. baseline RT corrected for accuracy
minus post cTBS RT corrected for accuracy) was compared for each site stimulated.
A 2 (Task Group) x 3 (TMS Site) mixed ANOVA was then conducted to determine
the effects of cTBS on participants’ abilities to recognize identity and emotion from
auditory signals. The overall main effect of TMS Site was not significant [F(2,36) =
.361, p = .699], however a significant Task Group x TMS Site interaction was found
[F(2, 36) = 3.43, p = < .05]. This was because the effects of cTBS significantly
differed across sites on the emotion [F(2, 18) = 4.78, p = < .05], but not on the
identity task [F(2, 18) = .574, p = .573] (Figure 7.3). The main effect of TMS Site on
the emotion task was due to significant impairments following cTBS targeted at rPM
compared to the vertex (p = < .05) and following cTBS targeted at rSI relative to the
vertex (p = < .05). Therefore, cTBS stimulation of rSI and rPM disrupted
participants’ abilities to discriminate between the auditory emotions, but not the vocal
identities, of others - indicating that neural activity within these brain regions is
important for the emotion discrimination abilities of healthy adults.
Between-group comparisons also revealed a main effect of Task Group [F(1,
18) = 12.81, p = < .005]. This task-specific dissociation was modulated by site of
stimulation, with cTBS targeted at rSI (p = < .01) and rPM (p = < .01) resulting in a
different pattern of performance between the emotion and identity tasks (Figure 7.3).
This pattern of effects was not found following stimulation at the vertex (cTBS
control site), where there was a trend for facilitation in both tasks (p = .841). Thus,
the cTBS impairments observed at rSI and rPM in the emotion task are not due to
general impairments in processing following cTBS, but reflect a task specific
impairment on emotion discrimination performance.

C
hapt
er 7
135
-200
-150
-100
-500
50
100
150
200
250
300
Baseline minus cTBS RT (msec)
rSI
rPM
Vertex
*
Emotion Discrimination
a
-200
-150
-100
-500
50
100
150
200
250
300
Baseline minus cTBS RT (msec)
rSI
rPM
Vertex
Identity Discrimination
b
Figu
re 7
.3 M
agni
tude
of d
isru
ptio
n or
faci
litat
ion
(mea
n ±
s.e.
m) i
n m
illis
econ
ds f
ollo
win
g cT
BS
targ
eted
at r
SI, r
PM a
nd th
e ve
rtex
acro
ss ta
sk
grou
ps. I
n or
der
to d
eter
min
e if
the
mag
nitu
de o
f im
pairm
ent
follo
win
g cT
BS
stim
ulat
ion
diff
ered
acr
oss
the
site
s an
d ta
sks,
the
diff
eren
ce
betw
een
the
post
cT
BS
and
pre-
cTB
S ba
selin
e re
actio
n tim
es (
± 3
stan
dard
dev
iatio
ns a
nd a
ll er
rors
rem
oved
; and
cor
rect
ed f
or a
ccur
acy)
wer
e ca
lcul
ated
for
eac
h co
nditi
on (
i.e. b
asel
ine
RT/
Acc
urac
y m
inus
pos
t cT
BS
RT/
Acc
urac
y fo
r ea
ch s
ite s
timul
ated
acr
oss
task
s). A
dis
rupt
ion
in
reac
tion
times
fol
low
ing
stim
ulat
ion
is s
how
n by
a n
egat
ive
valu
e an
d fa
cilit
atio
n by
a p
ositi
ve v
alue
. (a
) Fo
r th
e em
otio
n di
scri
min
atio
n ta
sk,
parti
cipa
nts
(n =
10)
wer
e im
pair
ed i
n th
eir
abili
ties
to d
iscr
imin
ate
betw
een
the
audi
tory
em
otio
ns o
f ot
hers
fol
low
ing
stim
ulat
ion
to r
SI a
nd
rPM
com
pare
d to
stim
ulat
ion
at t
he v
erte
x (c
TBS
cont
rol
site
). (
b) T
his
was
not
fou
nd t
o be
the
cas
e w
hen
parti
cipa
nts
(n =
10)
had
to
disc
rimin
ate
audi
tory
iden
tity
in th
e ex
perim
ent -
the
effe
cts
of c
TBS
targ
eted
at r
SI, r
PM a
nd th
e ve
rtex
did
not s
igni
fica
ntly
dif
fer b
etw
een
the
site
s st
imul
ated
, and
ther
e w
as a
tren
d fo
r fa
cilit
atio
n at
all
site
s. B
etw
een-
grou
p co
mpa
riso
ns a
lso
reve
aled
that
the
disr
uptio
n in
per
form
ance
on
the
emot
ion-
disc
rim
inat
ion
task
fol
low
ing
cTB
S to
rSI
and
rPM
was
sig
nifi
cant
ly d
iffe
rent
to th
e fa
cilit
atio
n sh
own
in th
e id
entit
y ta
sk.
No
sign
ific
ant d
iffer
ence
bet
wee
n th
e em
otio
n di
scrim
inat
ion
and
iden
tity
disc
rimin
atio
n ta
sk p
erfo
rman
ce w
as fo
und
follo
win
g cT
BS
at th
e ve
rtex
.
* = P
<.0
5.

Chapter 7
136
Are some emotions affected more than others?
To clarify whether the overall disruption of auditory emotion discrimination
observed at rSI and rPM in experiment 1 was linked to a greater impairment for
specific emotions or was expression-general, the effects of cTBS (corrected baseline
RT minus corrected post cTBS RT) for each emotion type (amusement, disgust, fear,
sadness) in the emotion task group were compared using a 3 (TMS Site) x 4
(Emotion-Type) repeated measures ANOVA. This revealed a main effect of TMS
Site, [F(2, 18) = 4.97, p = <.05], due to the overall impairment in auditory expression
matching following cTBS targeted at rSI and rPM relative to the vertex noted
previously, but no significant interaction [F(6, 54) = 1.32, p = .266] or main effect of
emotion-type [F(3, 27) = .812, p = .498]. Therefore the overall impairment in
emotion discrimination following cTBS to rSI and rPM was not modulated by
emotion-type.
7.4 Discussion
The current study investigated whether neural activity in rSI and rPM was
important for discriminating affect from non-verbal vocal signals. Using
neuronavigation procedures to co-register targeted sites onto each participant’s
structural MRI scan, stimulation targeted at rSI and rPM led to a significant disruption
in the ability to discriminate the auditory emotions, but not identities, of others
(Figure 7.3). This pattern was not found following cTBS at the vertex, indicating that
the differences observed were not due to non-specific effects of cTBS. Therefore
consistent with predictions, rSI and rPM stimulation reduced the ability to
discriminate the auditory emotions, but not identities, of others.
These findings extend upon research demonstrating the involvement of right
somatosensory cortex representations in facial affect recognition (Adolphs et al.,

Chapter 7
137
2000; Pitcher et al., 2008) to suggest that activity in rSI is central to the perception of
emotion across different modalities. Further, in recent years a number of functional
brain imaging studies have documented the role of premotor cortex activity in the
mirroring of actions and emotions of others (Dapretto et al., 2006; Hennenlotter et al.,
2005; Leslie, Johnsen-Frey, and Grafton, 2004; Montgomery and Haxby, 2008; van
der Gaag, Minderaa, and Keysers, 2007; Warren et al., 2006). Using stimuli adapted
from one such study (Warren et al., 2006), the findings show that neural activity in
rPM plays a central role in non-verbal auditory emotion discrimination in healthy
adults. These findings are consistent with simulation-based accounts of emotion
processing, which contend that perceived emotions are mapped onto an individual’s
own somatosensory and motor representations to facilitate emotion recognition
(Adolphs, 2002; Adolphs, 2003; Damasio, 1990; Gallese, Keysers, and Rizzolatti,
2004; Goldman and Sripada, 2005; Keysers and Gazzola, 2006).
The task specific nature of the findings further supports the role of rSI and
rPM activity as a substrate for a mechanism that facilitates emotion processing.
Under equivalent conditions to experiment 1, cTBS targeted at rSI and rPM did not
impair participants’ ability to discriminate another’s identity, indicating that the
changes in reaction time are not simply due to a general reduction in reaction times
following cTBS stimulation of these regions or more widespread suppression of
neural activity. In contrast to a disruption in emotion discrimination abilities, there
was a trend for facilitation when participants were asked to discriminate the identity
of others. This facilitation is non-specific because it is seen over all sites stimulated in
the identity task, and does not differ significantly between sites. The nature of the
effect may reflect practice in the post-cTBS blocks or intersensory facilitation
following cTBS (Marzi et al., 1998; Walsh and Pascual-Leone, 2003).

Chapter 7
138
The findings are also compatible with recent TMS findings documenting the
necessity of the right fronto-parietal operculum in emotional prosody (van Rijn,
Aleman, van Diessen, Berckmoes, Vingerhoets, and Kahn, 2005; Hoekert, Bais,
Kahn, and Aleman, 2008). They extend upon them by examining the role of
somatosensory and motor cortices in non-verbal auditory emotion processing. These
kind of auditory signals differ from emotionally inflected speech because they do not
have the segmented structure of speech and provide relatively “pure” vocal
expressions of emotion (Scott et al., 1997; Scott, Sauter, and McGettigan, in press;
Dietrich, Szameitat, Ackermann, and Alter, 2006). This enables a closer parallel to
previous studies examining the necessity of cortical resources in the processing of
emotional facial expressions (Adolphs et al., 2000; Pitcher et al., 2008). In addition,
the findings demonstrate the importance of motor resources in auditory emotion
discrimination and show a functional dissociation for the role of sensorimotor
simulation in discriminating speaker emotion, but not speaker identity, from vocal
signals.
There is growing evidence that the ability to detect affect from voice relies
upon similar neural mechanisms which are recruited for visual social signals. For
example, in the visual domain, event related potential (ERP) studies have
demonstrated enhanced frontal positivity for emotional compared to neutral faces 150
msec after stimulus onset (Ashley, Vuilleumier, and Swick, 2004; Eimer and Holmes,
2002; Eimer and Holmes, 2007; Eimer, Holmes, and McGlone, 2003; Holmes,
Vuilleumier, and Eimer, 2003). This mechanism also extends to the auditory domain,
in which non-verbal auditory emotions compared with spectrally rotated neutral
sounds result in an early fronto-central positivity which is similar in timing, polarity
and scalp distribution to ERP markers of emotional face processing (Sauter and

Chapter 7
139
Eimer, in press). The findings presented here add to this by demonstrating that
activity in rSI is implicated in not only facial (Adolphs et al., 2000; Pitcher et al.,
2008), but also auditory emotion perception and imply that sensorimotor resources
may sub-serve an emotion-general processing mechanism in healthy adults (Adolphs,
2002; Adolphs, 2003; Damasio, 1990; Gallese, Keysers, and Rizzolatti, 2004;
Goldman and Sripada, 2005; Keysers and Gazzola, 2006).
In the current study I focussed on right hemisphere representations based on
previous fMRI, neuropsychological and TMS findings demonstrating the importance
of right hemisphere activity in affect recognition (Adolphs et al., 2000; Mitchell and
Crow, 2005; Pitcher et al., 2008; Pourtois et al., 2004; Van Lancker and Fromkin,
1973). There is some fMRI evidence that viewing static and dynamic facial
expressions evokes activity in bilateral primary somatosensory cortex and premotor
cortex (Montgomery and Haxby, 2008; van der Gaag et al., 2007). Further, in the
auditory domain, listening to non-vocal emotional expressions leads to bilateral
activations of the lateral premotor cortex (Warren et al., 2006). The lateralization of
these effects shall be addressed with further studies.
In sum, this study extends previous findings that rSI activity is important in
facial emotion recognition (Adolphs et al., 2000; Pitcher et al., 2008), by
demonstrating that neural activity in rSI is involved in emotion processing across
modalities. The findings also demonstrate that rPM activity reported in previous
fMRI studies is central to non-verbal auditory emotion discrimination. These
resources are not specifically required for discriminating the identity of others and
appear to play a specific role in facilitating emotion discrimination in healthy adults.

Chapter 8
140
CHAPTER 8: THE ROLE OF SENSORIMOTOR SIMULATION
IN FACIAL EXPRESSION RECOGNITION
The findings in chapter 7, demonstrated that the right primary somatosensory cortex
and right premotor cortex play a critical role in discriminating between the non-
verbal auditory emotions of others. Recent findings indicate that neural activity in
right primary somatosensory cortex is also necessary for the recognition of facial
expressions in healthy adults, but it remains unclear whether neural activity in
cortical regions involved in other aspects of sensorimotor simulation (e.g. simulation
of motor as opposed to somatic consequences of the perceived emotion) are also
central to the facial expression recognition abilities of healthy adults. Further, in the
face processing literature, whether neural activity in different components of the
sensorimotor simulation network are central for recognizing all expressions (i.e. an
expression-general mechanism) or for subsets of expressions remains a point of
debate (i.e. an expression-specific mechanism). Using continuous theta burst
transcranial magnetic stimulation (cTBS) in neurologically normal subjects, this
study sought to establish whether sensorimotor neural activity is critical for the facial
expression recognition abilities for some, or all, basic facial expressions. cTBS was
targeted at right primary somatosensory cortex (rSI), right inferior frontal gyrus
(rIFG) or right V5 / MT (control site) while participants completed a four-forced-
choice expression categorization task. cTBS to rSI, but not rIFG or right V5 / MT
(control site), significantly disrupted participants abilities to correctly categorize
happy and sad facial expressions, but not disgust or neutral facial expressions. These
findings are consistent with sensorimotor simulation models of expression recognition
which suggest that in order to understand another’s facial expressions individuals
must map the perceived expression onto the same sensorimotor representations which
are active during the experience of the perceived emotion.
8.1 Introduction
As noted previously, perceiving and correctly interpreting the expressions of
others is a vital component of social interaction. The processes which facilitate this
skill have often been described through mechanisms of simulation (Adolphs, 2002;
Adolphs, 2003; Damasio, 1990; Gallese, Keysers, and Rizzolatti, G, 2004; Goldman,
and Sripada, 2005; Keysers and Gazzola, 2006). These simulation-models of
expression recognition contend that in order to understand another’s expression one
must match the perceived state onto the sensorimotor responses associated with
experiencing the expression. Supporting this contention, in the visual domain,

Chapter 8
141
subliminal exposure to emotional facial expressions leads to increased responses in
expression relevant facial muscles of the observer (Dimberg, Thunberg, and Elmehed,
2000); blocking expression relevant facial muscles results in deficits in the observer’s
ability to correctly categorize the expressions of others (Oberman, Winkielman, and
Ramachandran, 2007); perceiving another person’s facial expressions correlates with
increased activity in similar motor (e.g. inferior frontal gyrus and premotor cortex of
the human mirror system) and somatosensory representations (e.g. primary and
secondary somatosensory cortex) as when the perceiver generates the same emotion
or expression (Hennenlotter et al., 2005; Leslie, Johnsen-Frey, and Grafton, 2004;
Montgomery and Haxby, 2008; van der Gaag, Minderaa, and Keysers, 2007);
transiently disrupting neural activity in the somatosensory cortex disrupts the
observer’s expression recognition abilities (Pitcher, Garrido, Walsh and Duchaine,
2008; Pourtois et al., 2004); and brain damage to somatosensory-related cortices
results in facial expression recognition deficits (Adolphs, Damasio, Tranel, Cooper,
and Damasio, 2000).
While these findings converge on a key role for sensorimotor simulation in
facial expression recognition, a number of unanswered questions remain. For
example, recent findings indicate that right somatosensory-related cortices play a
pivotal role in facial expression recognition (Adolphs et al., 2000; Pitcher et al., 2008;
Pourtois et al., 2004), but the extent to which neural activity in cortical regions
involved in other aspects of sensorimotor simulation (e.g. simulation of motor as
opposed to somatic consequences of the perceived emotion) are also critical for facial
expression recognition remains unclear. Functional brain imaging indicates a role of
neural activity in both somatosensory and motor regions of the cortex in expression
recognition (Carr, Iacoboni, Dubeau, Mazziotta, and Lenzi, 2003; Hennenlotter et al.,

Chapter 8
142
2005; Leslie, Johnsen-Frey, and Grafton, 2004; Montgomery and Haxby, 2008; van
der Gaag, Minderaa, and Keysers, 2007; Winston, O’Doherty, and Dolan, 2003), but
these data alone cannot imply causation about the role of motor resources for
expression recognition. Motor mirror system activation involving the inferior frontal
gyrus (IFG; BAs 44, 45) has been shown to occur in a number of studies investigating
facial expression recognition or evaluation (Carr et al., 2001; Dapretto et al., 2006;
Hennenlotter et al., 2006; Kesler-West et al., 2001; Seitz et al., 2008); emotional
empathy (Jabbi, Swart, and Keysers, 2007; Schulte-Ruther, Markowitsch, Fink and
Piefke, 2007); and emotion recognition more generally (Wildgruber et al., 2005).
Neuropsychological findings indicate that the IFG is necessary for recognizing
emotions from the eyes (Shamay-Tsoory, Aharon-Peretz, and Perry, 2009), but the
ability to recognize expressions from the whole-face was not tested. Adolphs and
colleagues (2002) also report deficits in facial emotion recognition following damage
to the frontal operculum (including BA44), but the lack of region specificity limits the
conclusions which one can draw on the role of the IFG in facial expression
recognition in healthy adults. Therefore whether the IFG is critical to facial
expression recognition in healthy adults remains unknown.
There is also a discrepancy in the literature on whether sensorimotor resources
are critical for recognizing all or only some distinct facial expressions. fMRI
indicates that changes in neural activity in both motor (including premotor cortex and
IFG) and somatosensory (including SI and SII) cortices are related to perceiving a
range of facial expressions (Montgomery and Haxby, 2008; van der Gaag, Minderaa,
and Keysers, 2007). Neuropsychological findings are consistent with a role for
somatosensory-related cortices in expression general processing (Adolphs et al.,
2000), but whether neural activity in the somatosensory cortex of healthy adults is

Chapter 8
143
necessary for all or only some distinct facial expressions is a matter of debate. To
date only two transcranial magnetic stimulation studies have addressed the necessity
of the right somatosensory cortex in healthy adults. In one study, two emotional
expressions (fear and happiness) and single pulse TMS over right somatosensory
cortex were used to investigate the necessity of this brain region for emotion
recognition in healthy adults. These authors observed an expression-selective TMS-
related interference following stimulation of right somatosensory cortex during the
recognition of fearful, but not happy expressions (Pourtois et al., 2004). In contrast, a
more recent repetitive TMS study (Pitcher et al., 2008), using six emotional
expressions (anger, disgust, fear, happiness, sadness and surprise), found an
expression-general impairment following stimulation of the right somatosensory
cortex. Further study is needed to clarify this discrepancy.
Furthermore, in both previous TMS studies a same-different matching task
was used to assess participants’ expression recognition abilities. The nature of these
tasks requires some degree of working memory in which the participant must not only
recognize a sample expression but store the information in memory to match it to a
sequential expression. Therefore it is difficult to disentangle whether TMS
impairment results from a disruption of fronto-parietal working memory networks (cf.
Harris, Harris, and Diamond, 2001; Mottaghy, Gangitano, Sparing, Krause, and
Pascual-Leone, 2002; Oliveri et al., 2001) or sensorimotor simulation mechanisms per
se4.
To address this, this study sought to establish: i) whether neural activity in the
right primary somatosensory cortex (rSI) and right inferior frontal gyrus (rIFG) is
central to recognizing the facial expressions of others and ii) whether, at the cortical
4 Note that this is controlled for in Chapter 7 because of the task-specific dissociation observed.

Chapter 8
144
level, sensorimotor simulation is expression-general or expression-selective. To do
so, participants performed a four-forced-choice expression categorization task
(disgust, happy, neutral, sadness) following continuous theta burst TMS (cTBS)
targeted at the rSI, rIFG, and right V5 / MT (visual control TMS site). Facial
expressions of disgust, happiness, sadness, and neutral (expression control) were used.
All expressions were selected from the Karolinska Directed Emotional Faces set
(Lundqvist, Flykt, and İhman, 1998). Based on previous TMS findings (Pitcher et
al., 2008) it was predicted that cTBS targeted at rSI would result in an impairment of
participants’ abilities to correctly categorize expression types, but not neutral (control
expression). If rIFG activity plays a causal role in the emotional expression
recognition abilities of healthy adults then an impairment for categorizing emotional
expression types (but not neutral) following cTBS targeted at the rIFG site was
expected. Whether this impairment would be expression specific or expression
selective remains to be determined. No cTBS disruption was expected following
stimulation at right V5 / MT, which acted as a visual control site.
8.2 Method
Participants
Eleven healthy adult participants, 3 female and 8 male (aged 24 to 45 years),
took part in the study. All participants were right handed and had normal or corrected
to normal vision. Each participant gave written informed consent in accordance with
the ethics committee of University College London and was naïve to the hypothesis of
the experiment.
Materials and Procedure
The experiment consisted of three testing sessions conducted over three non-
consecutive days. At each testing session one of the three brain regions were

Chapter 8
145
stimulated: rSI, rIFG and right V5 / MT (TBS control site; Figure 8.1). The order of
site of stimulation was randomized between subjects. Each session consisted of two
blocks, one run prior to TBS (baseline performance) and the other following TBS.
During each block, participants completed a four-forced-choice emotion
recognition task, comprised of 140 trials (preceded by 20 practice trials) in which
participants had to indicate the emotional expression of a target face. Each trial began
with the presentation of a fixation cross (1500 ms) followed by the presentation of the
target stimuli. Target stimuli were presented in the centre of the screen for 250 ms.
Following the offset of the target stimuli participants were asked to indicate the
emotion expressed in the target emotion (either happy, sad, neutral or disgust) using a
key press (Figure 8.1d). Each block lasted approximately ten minutes. Stimuli were
displayed on an SVGA 17 inch monitor with a refresh rate of 100Hz. Thirty-five
grey-scale standardised images per emotion were used. Stimuli were selected from the
Karolinska Directed Emotional Faces set (Lundqvist, Flykt, and İhman, 1998).

Chapter 8
146
Figure 8.1 Summary of TMS sites simulated, right SI (a), rIFG (b), V5 / MT (c), and single trial protocol (d). (a, b, c) Locations of TMS were determined using the Brainsight Coregistration System. In addition to coregistration, the location of area V5 / MT was confirmed functionally via phosphenes. Continuous TBS parameters were used to stimulate each site. (d) Participants completed pre- and post-TBS blocks. Within each block, trials began with a fixation cross followed by the presentation of a target face displaying emotional expressions of happy, sad, neutral or disgust. Participants were asked to indicate the expression of the target face using a key press.

Chapter 8
147
TMS Protocol and Site Localisation
TMS was delivered via a figure of eight coil with a 70mm diameter using a
Magstim Super Rapid Stimulator (Magstim, UK). An offline continuous theta burst
TMS paradigm was used to prevent any influence of proprioceptive effects of TMS on
reaction time performance (Terao et al., 1997). The parameters were identical to
those used in Chapter 7. TMS machine output was set to 80% of each participant’s
motor threshold with an upper limit of 50% of machine output. Motor threshold was
defined using visible motor twitch of the contralateral first dorsal interosseus
following single pulse TMS delivered to the best scalp position over motor cortex and
was calculated using a modified binary search paradigm (MOBS; Tyrell and Owens,
1988). For each subject, motor threshold was calculated following pre-TMS baseline
and prior to coregistration.
Following the pre-TMS block, locations for TMS were identified using
BrainSight TMS-magnetic resonance coregistration system (Rogue Research,
Montreal, Canada). FSL software (FMRIB, Oxford) was used to transform
coordinates for each site to each subject’s individual MRI scan (Figure 1a, b, c).
Talairach coordinates for rSI (27, -27, 69) were taken from Blakemore and colleagues
(2005) and were averages for twelve neurologically normal subjects in an fMRI study
following touch to their own face (the same rSI site as that used in Chapter 7). The
MNI coordinates for rIFG (60, 8, 6) were the averages of twelve neurologically
normal subjects in an fMRI study of facial emotion (Hennenlotter et al., 2005) and the
region broadly corresponding to BA 44 of the inferior frontal gyrus. Coordinates for
V5 / MT (44, -67, 0) were taken from Dumoulin and colleagues (2000). In addition to
BrainSight coregistration, V5 / MT was also confirmed functionally using
phosphenes.

Chapter 8
148
8.3 Results
To assess the influence of speed and accuracy, reaction times were corrected
for accuracy in each condition. This was achieved by dividing reaction time (± 3
standard deviations and all errors removed) by accuracy in each condition.
The role of rSI and rIFG in recognizing different facial expressions of emotion
Preliminary analysis confirmed that baseline performance for each expression-
type did not significantly differ across the sites stimulated [Disgust - F(2,20) = 1.4 ,
nsig; Happy – F(2,20) = .280, nsig; Neutral – F(2,20) = .913, nsig; Sad – F(2,20) =
1.01, nsig].
To assess the effects across expression-types and across sites, the difference
between the post cTBS and pre-cTBS baseline reaction times for each expression (i.e.
baseline RT corrected for accuracy minus post cTBS RT corrected for accuracy) was
compared for each site stimulated (as per Chapter 7). A 3 (TMS Site) x 4
(Expression-Type) repeated measures ANOVA showed that neither the main effect of
TMS Site [F(2,20) = 1.55, p = .237] nor the main effect of Expression-Type reached
significance [F(3,30) = 1.39, p = .265]. There was however a significant TMS Site x
Expression-Type interaction [F(6,60) = 2.82, p = .017]. This was because cTBS at rSI
resulted in a significantly different pattern of effects across expression types [F(3,30)
= 4.34, p = .012], whereby cTBS impaired performance on trials involving
expressions of happiness relative to neutral facial expressions (p = < .05), and on trials
involving sadness relative to neutral (p = < .01) and disgusted facial expressions (p =
< .05). This was not the case at rIFG [F(3,30) = .977, p = .417] or right V5 / MT
[F(1.863,18.629) = .489, p = .608], where the effects of cTBS did not significantly
differ between the expression-types (Figure 7.2).

Chapter 8
149
-250
-200
-150
-100
-50
0
50
100
150
200
250
Baseline minus cTBS RT (msec)
rSI
rIFG
rV5
*
*
Disgust Happy Neutral Sad
* *
*
Figure 8.2 Magnitude of disruption or facilitation across expression-types (mean ± s.e.m) following cTBS targeted at rSI, rIFG and right V5 / MT. In order to determine if the magnitude of impairment following cTBS stimulation differed across the sites and expression the difference between the post cTBS and pre-cTBS baseline reaction times (± 3 standard deviations and all errors removed; and corrected for accuracy) were calculated for each expression (i.e. baseline RT/Accuracy minus post cTBS RT/Accuracy for each site stimulated across tasks) and compared across conditions. A disruption in reaction times following stimulation is shown by a negative value and an improvement by a positive value. Stimulation to rSI disrupted performance at recognizing happy and sad facial expressions compared to rIFG and right V5 / MT. The disruption on happy and sad trials at rSI significantly differed from the improvement seen on neutral trials. The pattern of performance on sad trials at rSI also differed significantly from that seen on disgust trials. * = P < .05.
Comparisons for each expression-type across TMS sites revealed that the
disruption caused by cTBS to rSI on trials involving sadness was significantly
different to the effects of cTBS at rIFG and right V5 / MT – a one way repeated
measures ANOVA comparing performance on sadness recognition across the three
sites revealed dissociable effects of site stimulated [F(2,20) = 4.35, p = .027], and
pair-wise comparisons revealed that this was due to participants’ showing a
significant disruption at recognising sadness following stimulation at rSI compared to
rIFG (p = <.01) and right V5 / MT (p = < .05). Similarly, a comparison on happiness

Chapter 8
150
recognition across the three sites, revealed a trend towards significance [F(2,20) =
3.04, p = .07]. Paired-comparisons conducted on the basis of this strong trend,
revealed that this was due to a significantly larger disruption on participants’ abilities
to recognize happiness following stimulation at rSI compared to rIFG [t(10) = 2.30, p
= < .05] and right V5 / MT [t(10) = 2.41, p = < .05] (Figure 7.2).
8.4 Discussion
This study sought to establish whether i) neural activity in rSI and rIFG is
critical for recognizing the facial expressions of others and ii) whether these
mechanisms are expression-general or expression selective. The findings first
demonstrate that neural activity in rSI, but not rIFG, is critical for the ability to
recognize others’ facial expressions. They further indicate that neural activity in rSI
plays a more crucial role in the recognition of emotional compared with neutral facial
expressions.
Previous neuropsychological findings indicate that lesions to right
somatosensory-related cortices lead to expression-general face recognition
impairments (Adolphs et al., 2000). In healthy adults, focal transcranial magnetic
stimulation of rSI has been shown to a) impair fearful, but not happiness, facial
expression recognition, implying that rSI is expression selective (Pourtois et al., 2004
- note that only 2 expression types were used and differences only found when
participants discriminated between two fearful expressions), and b) to impair general
facial expression recognition abilities, including happiness – implying that rSI is
expression-general (Pitcher et al., 2008 – note only 12 trials per expression type used).
Using neuronavigation procedures, the findings from the current study show that
magnetic stimulation of rSI results in selective impairments of happiness and sadness

Chapter 8
151
recognition, but not disgust or neutral recognition, and indicate that neural activity in
rSI plays a crucial role in the recognition of facial expressions other than fear per se
(c.f. Pourtois et al., 2004). The study is also the first to show a facial expression
recognition deficit on an emotion categorisation task (as opposed to same-different
matching tasks used previously), which further clarifies that previously reported
impairments in expression recognition following stimulation to rSI are not linked to a
disruption of fronto-parietal working memory networks.
Models of face processing posit a number of expression-selective, expression-
general, face-selective and multimodal mechanisms which support our ability to
perceive another’s emotions (Bruce and Young, 1986; Calder and Young, 2005;
Haxby, Hoffman, and Gobbini, 2000). Sensorimotor simulation is likely to be one of
these mechanisms, with neural activity in rSI providing a possible candidate to
facilitate this process. While the evidence that cTBS stimulation resulted in a
disruption of participants’ abilities to recognise sadness and happiness, but not
disgust, may imply a degree of expression selectivity within this region, it remains
possible that rSI is involved in recognizing alternative expression types. Moreover,
findings from chapter 7, of a general impairment in auditory expression recognition
following cTBS to rSI, and from Pitcher et al. (2008), of general impairments in the
facial expression recognition following TMS to rSI would argue against rSI acting as
an expression-specific mechanism. Outside of rSI, there is evidence for emotion-
specific neuropsychological deficits for expressions of disgust (Calder, Keane, Manes,
Antoun, and Young, 2000; Sprengelmeyer et al., 1996, 2003), fear (Calder, Lawrence,
and Young, 2001) and anger (Calder, Keane, Lawrence, and Manes, 2004; Lawrence,
Calder, McGowan, and Grasby, 2002). This disproportionate role of different brain
regions in processing specific expressions indicates that facial expressions are not

Chapter 8
152
processed by a single “expression system” and that expression-general and
expression-specific representations may interact at varying levels of processing. The
lack of impairment on disgust recognition found here may reflect the fact that the
neural noise introduced in rSI by TMS is better compensated for by emotions which
have more alternative mechanisms / expression-specific representations elsewhere in
the brain. For example, there is a good degree of evidence from functional brain
imaging (Phillips et al., 1997, 1998; Sprengelmeyer, Rausch, Eysel, and Przuntek,
1998), intracerebral recording (Krolak-Salmon et al., 2003), and neuropsychological
studies (Calder et al., 2000; Kipps, Duggins, McCusker, and Calder, 2007) which
indicate that the anteroventral insula acts as an expression-specific mechanism for
disgust recognition. The anteroventral section of the insula is connected to number of
regions which are thought to be involved in emotion processing across modalities
(including the primary somatosensory cortex, basal ganglia, amygdala, orbitofrontal
cortex and superior temporal cortex; Augustine, 1996; Flynn, Benson, and Ardila,
1999; Mesulam and Mufson, 1982) and has been suggested to act as a point of
convergence for sources involved in the processing of disgust recognition to varying
degrees (Kipps et al., 2007). It is feasible that suppressing rSI with cTBS reduces
one, of the multiple sources, of information which contributes to disgust processing in
this section of insula, and therefore does not result in impairment. It is also possible
that with alternative paradigms (e.g. same-different expression matching paradigms as
opposed to a forced-choice paradigm which could bias responses of disgust and
thereby facilitate performance) and stimuli (e.g. dynamic stimuli as opposed to static),
stimulation to rSI may lead to a disruption of participants’ abilities to recognise
disgusted facial expressions (c.f. Chapter 7; Pitcher et al., 2008).

Chapter 8
153
The findings that cTBS targeted at rIFG did not result in an impairment of
participants’ abilities to correctly recognise the other’s facial emotions are also
intriguing, especially in light of evidence that lesions to the IFG have been linked to
deficits in self-reported emotional empathy and in the ability to recognise emotions
from the eyes (Shamay-Tsoory et al., 2009). These differences are likely to reflect
variations in the processes involved in recognising emotional expressions from the
whole-face compared with emotional empathy per se, and imply that the IFG may not
be critical for all emotion-general tasks. The IFG has been reported as a
cytoarchitectonic homologue to monkey F5 (Petrides, Cadoret, and Mackey, 2005)
and highlighted as a core component of the classical human mirror system (Rizzolatti
and Craighero, 2004). A number of authors have suggested that the human mirror
system (including IFG, IPL, and STS) may be pivotal to social cognition (Gallese,
Keysers, and Rizzolatti, G, 2004; Keysers and Gazzola, 2006; Oberman and
Ramachandran, 2007). While caution is urged in interpreting a null result, the
evidence that magnetic stimulation targeted at rIFG does not impair the ability to
recognise other’s facial emotions stands in contrast to this hypothesis. It is of note
that this may not imply that motor simulation play no role in emotion processing,
because while the human mirror system is one neurophysiological candidate to
facilitate this process it need not be the only mechanism and other regions of the
motor system may be crucial for facial expression recognition (e.g. the human
premotor cortex – c.f. Chapter 7).
In summary, the findings from the current study indicate that rSI is critical for
the recognition of emotional (across multiple expression types) compared with neutral
facial expressions. They also indicate the rIFG is not critical to facial expression
recognition. This adds to the evidence that somatosensory activity may provide a

Chapter 8
154
general expression recognition mechanism (Chapter 7; Adolphs et al., 2000; Pitcher et
al., 2008). However, the evidence that rSI stimulation disproportionately affected
happy and sad expressions but not disgust, indicates that while somatosensory cortex
activity may be involved in the processing of a variety of facial expressions, in some
cases (e.g. with disgust recognition) alternative facial expression recognition
mechanisms (e.g. expression selective) may be sufficient to support facial expression
recognition when rSI activity is suppressed.

Chapter 9
155
CHAPTER 9: CONCLUSIONS
In this chapter, the empirical findings from chapters 2-5 and 7-8 of this thesis are
discussed in a wider context. Chapters 2-5 addressed the prevalence, neurocognitive
mechanisms, and consequences of mirror-touch synaesthesia for perception and
social cognition. These findings are discussed in relation to previous research on
synaesthesia and future studies on the neurocognitive mechanisms of mirror-touch
synaesthesia are also described. Chapters 4 and 5 also used mirror-touch
synaesthesia to inform us about the role of sensorimotor resources in social
cognition. These findings were complemented by studies in chapters 7 and 8, which
investigated the impact of suppressing sensorimotor representations on the expression
recognition abilities of healthy adults. The findings from chapters 4, 5, 7 and 8 are
discussed in the context of research on social cognition and sensorimotor accounts of
social cognition.
9.1 Introduction
As noted in the introduction to this thesis, synaesthesia is a condition in which
one attribute of a stimulus (the inducer) triggers a conscious experience of another
attribute (the concurrent) not typically associated with the inducer. For example, in
grapheme-colour synaesthesia the letter ‘a’ may trigger synaesthetic experiences of
colours. A large body of synaesthesia research has focussed on grapheme-colour
synaesthesia, which is often reported as being one of the most common forms of the
condition (Baron-Cohen, Burt, Smith-Lailtan, Harrison, and Bolton, 1996; Rich,
Bradshaw, and Mattingley, 2005; Simner et al., 2006). More recently, a newly
documented form of synaesthesia has been described in which individuals experience
tactile sensations on their own body simply when observing touch to another’s body
(mirror-touch synaesthesia; Banissy and Ward, 2007; Blakemore, Bristow, Bird, Frith,
and Ward, 2005). The studies in the first five chapters of this thesis investigated the
neurocognitive and perceptual profiles of mirror-touch synaesthesia. In addition, the
role of sensorimotor simulation mechanisms in social cognition was examined by
using principles of neuropsychology (in the case of mirror-touch synaesthesia) and

Chapter 9
156
transcranial magnetic stimulation (TMS). Specifically the following questions were
addressed:
1. What is the prevalence of mirror-touch synaesthesia and what
characteristics identify the condition (Chapter 2)?
2. What neurocognitive mechanisms give rise to mirror-touch synaesthesia
(Chapter 2)?
3. Does the presence of synaesthesia have implications for perceptual
processing (Chapter 3)?
4. What are the implications of heightened sensorimotor simulation in mirror-
touch synaesthesia for social cognition (Chapters 4 and 5)?
5. What are the implications of suppressing sensorimotor resources on
expression recognition abilities in healthy adults (Chapters 7 and 8)?
9.2 What is the prevalence of mirror-touch synaesthesia and what
characteristics underpin the condition?
As noted previously, in mirror-touch synaesthesia individuals experience
tactile sensations on their own body simply when observing touch to another person
(Banissy and Ward, 2007; Banissy, Cohen Kadosh, Maus, Walsh, and Ward, 2009;
Blakemore et al., 2005). The mapping between synaesthetic experience (i.e. location
on the synaesthete’s body) and observed-touch (i.e. the location where touch is
perceived on another person’s body) has been shown to vary between mirror-touch
synaesthetes (Banissy and Ward, 2007), with some synaesthetes reporting
synaesthetic experiences as if looking in a mirror (e.g. observed-touch to the left face
elicits a synaesthetic experience on the left cheek of the mirror-touch synaesthete;
specular subtype) and others as if they share the same anatomical body space (e.g.
observed-touch to the left face elicits a synaesthetic experience on the left cheek of

Chapter 9
157
the mirror-touch synaesthete; anatomical subtype). The first reported case of the
condition was provided in a single case fMRI study, which linked the condition to
heightened neural activity in a network of brain regions which are also activated in
non-synaesthetic control subjects when observing touch to others (the mirror-touch
system, comprised of the primary and secondary somatosensory cortex, premotor
cortex, intraparietal sulcus and superior temporal sulcus; Blakemore et al., 2005).
Chapter 2 examined the prevalence and characteristics of the condition.
In the first experiment reported in chapter 2 the prevalence of mirror-touch
synaesthesia was assessed by screening a large population of undergraduate students
for the presence of mirror-touch synaesthesia and determining the validity of reported
cases with a synaesthetic stroop task (Banissy and Ward, 2007). In the task,
participants were asked to indicate the site of touch on their own body while
observing touch to another person. Participants were asked to report the site of
veridical touch and ignore any synaesthetic tactile experience induced. For
synaesthetes, but not for controls, veridical touch could be in the same (congruent) or
different (incongruent) location to observed / synaesthetic touch (congruency was
determined according to each synaesthete’s self reports). Synaesthetes performed
slower in the incongruent condition and produced more errors linked to their
synaesthesia. Nine mirror-touch synaesthetes (from 567 participants screened) were
confirmed, which provides an estimated prevalence rate of 1.6%. This places mirror-
touch synaesthesia as one of the most common variants of synaesthesia, alongside
grapheme-colour synaesthesia (estimated prevalence rate of 2%; Simner et al., 2006)
and day-colour synaesthesia (estimated prevalence rate of 2.8%; Simner et al., 2006).
By combining cases of mirror-touch synaesthesia from the prevalence study
with cases of mirror-touch synaesthesia from self-referrals, the findings from chapter

Chapter 9
158
2 also indicate a number of features linked to the characteristics of the condition. The
findings from experiment 2 indicate that the inducer for synaesthetic experience is not
linked to spatial cueing, but is related to bodily touch (and in some cases touch to
objects). The findings also indicate the specular subtype is the more common frame
of reference adopted by mirror-touch synaesthetes and the relative frequencies
(approximately 81% show a specular frame of reference) are similar to those reported
in studies investigating the preferred spatial frame adopted when imitating another’s
behaviour - both adults and children tend to imitate in a specular mode (Schofield,
1976; Franz, Ford and Werner, 2007). Further characteristics indicate commonalities
between mirror-touch synaesthesia and other variants of synaesthesia. For example, a
general characteristic of synaesthesia is that different variants of synaesthesia tend to
co-occur (Simner et al., 2006). This also appears to be the case in mirror-touch
synaesthesia. Synaesthetic experiences also tend to be consistent over time (Baron-
Cohen, Wyke, and Binnie, 1987) and the mirror-touch synaesthete’s spatial sub-type
(i.e. whether they belong to the specular or anatomical category) appears to be
consistent across time (Chapter 2) and across different body parts (Banissy and Ward,
2007).
While these findings indicate that mirror-touch synaesthesia shares common
ground with other types of synaesthesia, possible similarities in the neural basis of the
condition are less apparent. A point of dispute in the synaesthesia literature is
whether synaesthetic experience is due to cross-activation between brain regions
(either through increased structural connectivity or malfunctions in cortical inhibition)
or disinhibition of the same cortical networks found in non-synaesthetes (Bargary and
Mitchell, 2008; Cohen Kadosh, Henik, Catena, Walsh, and Fuetnes, 2009; Cohen
Kadosh and Walsh, 2008; Grossenbacher and Lovelace, 2001; Hubbard and

Chapter 9
159
Ramachandran, 2005; Rouw and Scholte, 2007). Cross-activation accounts have
tended to focus on adjacent brain regions (e.g. in the case of grapheme-colour
synaesthesia - between visual grapheme and colour processing areas in the fusiform
gyrus) and suggest that activation in the region responsible for processing the
synaesthetic inducer (e.g. the grapheme in grapheme-colour synaesthesia) leads to
activation in the adjacent region for processing the synaesthetic concurrent (e.g.
colour in grapheme-colour synaesthesia). It is not entirely clear how the principle of
adjacency can be applied to mirror-touch synaesthesia, and an alternative mechanism
which may bias individuals to this type of synaesthesia is the normal architecture for
multi-sensory interactions (Sagiv and Ward, 2006). For example, there is good
evidence for an observed-touch mirror system in non-synaesthetes (Keysers, Wicker,
Gazzola, Anton, Fogassi, and Gallese, 2004; Blakemore et al., 2005; Ebisch, Perrucci,
Ferretti, Del Gratta, Romani, and Gallese, 2008) and mirror-touch synaesthesia has
been suggested to reflect over-activity within this network (Blakemore et al., 2005).
Future studies will aim to address the similarities and differences in the neural
basis of different subtypes of synaesthesia by investigating structural and functional
correlates of different variants of synaesthesia (e.g. grapheme-colour, tone-colour,
mirror-touch, and number-space synaesthesia). For example, previous DTI findings
indicate that grapheme-colour synaesthesia is linked with increased structural
connectivity in right inferior-temporal, right parietal, and bilateral frontal regions
(Rouw and Scholte, 2007), and research in progress indicates that tone-colour
synaesthesia is linked to increased cortical thickness (a marker of cortical morphology
and neurodevelopment; MacDonald, Kabani, Avis, and Evans, 2000; Shaw et al.,
2006) in similar right inferior temporal regions (Banissy, Stewart, Ward, Walsh, and
Kanai, in prep). I am also starting a combined fMRI-DTI study of mirror-touch

Chapter 9
160
synaesthesia, which will investigate functional and structural correlates of mirror-
touch synaesthesia at the group level. This will permit further assessment of
similarities and differences in the neural substrates of different variants of
synaesthesia and across subtypes of mirror-touch synaesthetes.
9.3 What neurocognitive mechanisms may underpin mirror-touch
synaesthesia?
In addition to the studies reported in chapter 2, I proposed a neurocognitive
model to account for the processes which may underpin mirror-touch synaesthesia
(Figure 9.1). Three key mechanisms were highlighted: identifying the type of visual
stimulus touched (‘What’ mechanism), discriminating between self and other (‘Who’
mechanism), and locating where on the body and in space observed touch occurs
(‘Where’ mechanism).
The ‘What’ mechanism is considered to be involved in several
discriminations, including: ‘is this a human or object?’ ‘Is this a face or body?’ One
intriguing characteristic shown by some mirror-touch synaesthetes is that observing
touch to objects can elicit synaesthetic interactions in some, but not all, synaesthetes
(Chapter 2; Banissy and Ward, 2007). One brain region of the observed-touch
network (Blakemore et al., 2005) which may be central to this is the intraparietal
sulcus (IPS). Recent findings indicate that visual object information is processed
along the dorsal stream to areas along the medial bank of the intraparietal sulcus (IPS;
including IPS1 and IPS2, Konen and Kaster, 2008). For mirror-touch synaesthetes,
this pathway may be particularly important when considered in relationship to visual-
tactile body maps within the intraparietal cortex. Single-cell recording in primates has
identified bimodal neurons in the intraparietal cortex which fire in response to not
only passive somatosensory stimulation, but also to a visual stimulus presented in

Chapter 9
161
close proximity to the touched body part (Duhamel, Colby and Goldberg, 1998).
Intriguingly, the visual spatial reference frames of such bimodal neurons are dynamic,
such that if the monkey is trained to use a tool the visual receptive field extends to
incorporate the tool into the representation of the body - potentially as an extension of
the body schema (Iriki, Tanaka and Iwamura., 1996). Similar evidence of dynamic
multisensory body representations in the parietal cortex have been reported in human
subjects (Bremmer et al., 2001; Macaluso and Driver, 2003; also see Colby, 1998;
Maravita and Iriki, 2002; Berlucchi and Aglioti, 1997 for review). One hypothesis
generated by the model is that the degree to which observing touch to an object is able
to elicit visual-tactile synaesthetic interactions depends upon the extent to which the
object is incorporated into visual-tactile representations of the body, potentially within
the intraparietal cortex. A potential approach to investigate this would be to
investigate if extending the body-schema of a mirror-touch synaesthete through tool
use can result in a synaesthete who does not normally experience synaesthetic touch
for objects showing synaesthetic interactions for observed object-touch.
The key process instigated by the ‘Who’ mechanism is to distinguish between
the self and other. I suggest that mirror-touch synaesthesia reflects a breakdown in
the mechanisms which normally distinguish between self and other (i.e. processes
involved in linking visual representations with internal representations of bodies).
One prediction of this is that mirror-touch synaesthetes will show a tendency to over-
incorporate viewed bodies onto their own body schema. In accordance with this,
research in progress indicates that mirror-touch synaesthetes show a greater degree of
the rubber hand illusion (RHI) compared to matched non-synaesthete controls
(Banissy, MacDonald, Ward, Walsh, Haggard and Longo, in prep). The RHI is a
body schema illusion in which an observer is touched on their own hand while

Chapter 9
162
observing a rubber hand being touched. When observed and veridical touch are
synchronous the perceived location of the observer’s hand drifts towards the location
of the rubber hand (Longo, Schuur, Kammers, Tsakiris, and Haggard, 2008; Tsakiris
and Haggard, 2005) and participants begin to attribute the rubber hand to their own
body representation (Botvinick and Cohen, 1998). This effect is abolished with
asynchronous stroking and when participants view a non-hand object rather than a
rubber hand (Tsakiris and Haggard, 2005). A comparison of the size of the perceived
drift towards the rubber hand (in centimetres) indicates the mirror-touch synaesthetes
show a greater incidence of the illusion compared to matched non-synaesthetes when
observed and veridical touch are synchronous (i.e. under normal RHI conditions), but
no differences are found in conditions in which the illusion is not expected to take
place (e.g. non-hand object control conditions; Banissy et al., in prep). Future studies
will investigate whether modulations of the RHI differ between mirror-touch
synaesthetes and non-synaesthetes. For example, in non-synaesthetes rotating the
RHI 180° (i.e. as if from another’s perspective) abolishes the illusion (Tsakiris and
Haggard, 2005), however given that mirror-touch synaesthetes experience tactile
sensations when viewing touch from another’s perspective it will be interesting to
determine whether they show a similar or different pattern to non-synaesthetes.
The final class of mechanism described in the model involves linking visual
representations of body with tactile representations based on spatial frames of
reference. For this I draw a distinction between embodied (the sense of being
localised within one’s own body) and disembodied representations of the body (e.g.
autoscopic phenomena in which individuals experience the location of the self outside
of one’s own body – Brugger 2002; Blanke, Landis, Spinelli and Seeck, 2004; Blanke,
Ortigue, Landis and Seeck, 2002). It is postulated that a similar division can be made

Chapter 9
163
for the specular-anatomical division in mirror-touch synaesthetes. The specular
subtype appear to process the visual representation of the other body in an embodied
manner (i.e. as a mirror-image of oneself), while for the anatomical subtype the
spatial mapping between self and other could be considered disembodied in that the
synaesthete’s own body appears to share the same bodily template as the others
person (i.e. the synaesthete’s body is placed in the perspective of the other person).
This difference may suggest that anatomical mirror-touch synaesthesia can be
considered to be similar to types of autoscopic phenomena in which individuals
experience the location of the self outside of one’s own body (Brugger 2002; Blanke,
Landis, Spinelli and Seeck, 2004; Blanke, Ortigue, Landis and Seeck, 2002),
however, no mirror-touch synaesthetes report typical phenomena of autoscopy – that
of seeing one’s own body and the world from a location outside of their own physical
body5 (Bünning and Blanke, 2005). Therefore rather than classifying anatomical
mirror-touch synaesthesia within the bracket of autoscopic phenomena I would
suggest that some mechanisms which give rise to the spatial frames of adopted by
mirror-touch synaesthetes are modulated by similar mechanisms as those observed in
autoscopy. For example, disembodied experiences have been suggested to arise from
functional disintegration of low-level multisensory processing mechanisms (Bünning
and Blanke, 2005; Blanke and Mohr, 2005) and abnormal activity at the temporal
parietal junction (TPJ; Arzy, Thut, Mohr, Michel and Blanke, 2006; Blanke, Landis,
Spinelli and Seeck, 2004; Blanke, Ortigue, Landis and Seeck, 2002) and one may
suggest the anatomical sub-type will be associated with these neurocognitive
mechanisms.
5 It is of note that while synaesthetes may not have overtly reported autoscopy they may still experience this if tested systematically (e.g. Terhune, 2009).

Chapter 9
164
Figure 9.1 The ‘What, Who, Where Model of Mirror-Touch Synaesthesia’. ‘What’ mechanisms are shown in red boxes and are involved in defining the stimulus touched. ‘Who’ mechanisms implement discriminations between self and other, and are shown in blue boxes. ‘Where’ mechanisms are shown in green boxes and are involved in locating where on the body and in space observed touch occurs. Processes necessary for all subjects are shown with black arrows, necessary for specular mirror-touch synaesthetes with orange arrows, and for anatomical mirror-touch synaesthetes with purple arrows. Brain regions represented are considered with regard to importance for mirror-touch synaesthesia. AI = Anterior Insula; EBA = Extrastriate Body Area; FBA = Fusiform Body Area; FFA = Fusiform face area; IFG = Inferior Frontal Gyrus; IPL = Inferior Parietal Lobule; IPS = Intraparietal Sulcus; LO = Lateral Occipital Cortex; SI = Primary Somatosensory Cortex; SII = Secondary Somatosensory Cortex; STS = Superior Temporal Sulcus; TPJ = Temporoparietal Junction.
9.4 Does the presence of synaesthesia have implications for perceptual
processing?
Chapter 3 investigated whether the presence of synaesthesia has implications
for perceptual processing. Previous ERP findings indicated that the presence of
synaesthesia may exert a wider influence over sensory processing and impact on the
veridical sensory processing of synaesthetes (Barnett et al., 2008; Goller, Otten, and
Ward, 2009). For example, Barnett and colleagues (2008) report that, compared to

Chapter 9
165
non-synaesthetes, linguistic-colour synaesthetes show differences in early components
of the visual evoked potential (VEP) when presented with simple visual stimuli which
do not evoke synaesthesia. Further to this, Yaro and Ward (2007) report that
synaesthetes who experience colour show superior colour discrimination abilities
compared to non-synaesthetic control subjects. To assess whether enhanced
perceptual processing was a core property of synaesthesia, colour and tactile
sensitivity was contrasted between mirror-touch synaesthetes, synaesthetes who
experience colour as evoked sensations (colour synaesthetes), synaesthetes who
experience mirror-touch synaesthesia and colour synaesthesia (dual synaesthetes), and
a group of non-synaesthetic controls. The findings indicate a relationship between the
modality of synaesthetic experience and the modality of sensory enhancement. On a
test of tactile discrimination, mirror-touch synaesthetes showed superior tactile
discrimination compared to colour synaesthetes and non-synaesthetes. On a test of
colour perception, colour synaesthetes outperformed non-synaesthetes. Dual
synaesthetes (synaesthetes who experience both touch and colour as evoked
sensations) outperformed the non-synaesthetic control group on both tasks, and
outperformed colour synaesthetes on the tactile perception task. These findings imply
that sensory enhancement in the concurrent perceptual system may be a general
property of synaesthesia and show that the presence of synaesthesia exerts a wider
influence over sensory processing.
The mechanisms which underpin sensory enhancement in synaesthesia are
likely to reflect differences in brain development as a function of synaesthesia (which
may be either a cause or consequence of synaesthesia). As noted previously, the
neural mechanisms which underpin synaesthesia are a subject of uncertainty, with
some authors suggesting that the condition may be due to additional structural

Chapter 9
166
connectivity (i.e. structural differences; Bargary and Mitchell, 2008; Rouw and
Scholte, 2007), others in favour of malfunctions in cortical inhibition (i.e. functional
but not structural differences; Cohen Kadosh and Henik, 2007; Cohen Kadosh and
Walsh, 2008; Grossenbacher and Lovelace, 2001), and others for a combination of
both (Smilek, Dixon, Cudahy, and Merikle, 2001). In principle, the findings of
enhanced sensory perception in synaesthesia could be accounted for by any of these
approaches (e.g. mechanisms of inhibition may unmask local anatomical pathways,
while altered connectivity may result in alternative local and widespread anatomical
pathways which could facilitate performance). Moreover, compatible findings of
sensory-enhancement in the deprived brain would suggest that both aberrant
connectivity and malfunctions in cortical inhibition could play a role in the sensory-
enhancement found in synaesthetes. For example, temporary enhancements in tactile
acuity can occur following blindfolding, and are thought to be due to fast-acting
unmasking of existing connections to maintain functional behaviour. In comparison,
tactile acuity can also be enhanced in blindness, which is though to reflect sustained
unmasking of existing connections leading to new local and widespread anatomical
pathways (a slow-acting mechanism; Pascual-Leone, Amedi, Fregni, and Merabet,
2005). Future studies should investigate how and whether mechanisms of cortical
inhibition and connectivity interact in synaesthesia, and assess the possibility that
increased structural connectivity in synaesthesia (Rouw and Scholte, 2007) may
reflect sustained unmasking of existing connections (Cohen Kadosh et al., 2009). A
further intriguing possibility would be to examine the interaction between fast-acting
cortical unmasking mechanisms in sensory-enhancement following deprivation and
synaesthesia. For example, if temporary enhancements in tactile acuity following
blindfolding are linked to perceptual unmasking, and synaesthesia is linked to reduced

Chapter 9
167
cortical inhibition (and therefore increased unmasking; c.f. Cohen Kadosh and Walsh,
2006), then one may suspect that grapheme-colour synaesthetes will show more rapid
enhancements in tactile acuity following sensory deprivation compared to non-
synaesthetes.
9.5 What are the implications of heightened sensorimotor simulation in
mirror-touch synaesthesia for social cognition?
In addition to studies investigating the neurocognitive basis of synaesthesia,
there is a growing interest in using the condition to inform us about normal models of
cognitive processing (Mattingley and Ward, 2006). This approach rests on the
assumption of neuropsychology, where one is able to use a symptom affecting the
normal system to inform us about the function of the normal system. In the case of
synaesthesia the symptom is a positive one and in the case of mirror-touch
synaesthesia one is assessing the impact of facilitated sensorimotor simulation
mechanisms on cognition. Moreover, functional brain imaging has linked mirror-
touch synaesthesia to heightened neural activity in a network of brain regions which
are also activated in non-synaesthetic control subjects when observing touch to others
(the mirror-touch system; Blakemore et al., 2005). The mirror-touch system is
comprised of brain areas active during both the observation and passive experience of
touch (including primary and secondary somatosensory cortices, and premotor cortex;
Blakemore et al., 2005; Ebisch et al., 2008; Keyers et al., 2004) and has been
suggested to be a candidate neural mechanism to aid social cognition through
sensorimotor simulation (Gallese, 2006; Gallese and Goldman, 1998; Keysers and
Gazzola, 2006; Oberman and Ramachandran, 2007). Accounts of social cognition
involving sensorimotor simulation contend that, in order to understand another’s
emotions and physical states, the perceiver must map the bodily state of the observer

Chapter 9
168
onto the same representations involved in experiencing the perceived state (Adolphs,
2002; Adolphs, 2003; Damasio, 1990; Gallese, Keysers, and Rizzolatti, G, 2004;
Gallese, 2006; Gallese and Goldman, 1998; Goldman, and Sripada, 2005; Keysers and
Gazzola, 2006; Oberman and Ramachandran, 2007). There is now a good degree of
evidence from functional brain imaging, neuropsychological, transcranial magnetic
stimulation, and electrophysiological studies to indicate a role for sensorimotor
resources in this process (Adolphs, Damasio, Tranel, Cooper, and Damasio, 2000;
Carr, Iacoboni, Dubeau, Mazziotta, and Lenzi, 2003; Dapretto et al., 2006;
Hennenlotter et al., 2006; Jabbi, Swart, and Keysers, 2007; Kesler-West et al., 2001;
Leslie, Johnsen-Frey, and Grafton, 2004; Montgomery and Haxby, 2008;
Nummenmaa, Hirvonen, Parkkola, and Hietanen, 2008; Oberman, Winkielman, and
Ramachandran, 2007; Pitcher, Garrido, Walsh, and Duchaine., 2008; Schulte-Ruther,
Markowitsch, Fink, and Piefke, 2007; Seitz et al., 2008; van der Gaag, Minderaa, and
Keysers, 2007; Warren et al., 2006; Wildgruber et al., 2005; Winston, O’Doherty, and
Dolan, 2003). The studies reported in chapters 4 and 5 attempted to use mirror-touch
synaesthesia as a model to inform us about the impact of facilitated sensorimotor
simulation on empathy and facial expression recognition.
In chapter 4, the empathic abilities of mirror-touch synaesthetes were
compared to control participants in two studies. The first study used a self-report
empathy questionnaire (the empathy quotient – Baron-Cohen and Wheelwright, 2004)
and showed that mirror-touch synaesthetes have higher levels of emotional reactive
empathy (i.e. instinctive responses to others emotions), but do not have higher levels
of cognitive or social components of empathy compared with control participants.
Higher levels of empathy were not observed in other-variants of synaesthesia,
indicating that it relates specifically to mirror-touch synaesthesia (and the mechanisms

Chapter 9
169
which underpin it). In the second study, these findings were replicated using the same
measure of empathy and on another measure of empathy (Davis, 1980). The findings
from the second study also confirmed that differences observed in empathy between
mirror-touch synaesthetes and non-synaesthetes were linked to ‘other’ rather than
‘self’ orientated reactions, and found a significant relationship between the openness
to experience personality trait and emotional empathy. Mirror-touch synaesthetes
were also shown to significantly differ from non-synaesthetes on the openness to
experience personality trait. These findings appear consistent with accounts of
empathy that posit a role for sensorimotor simulation mechanisms (Gallese, 2006;
Gallese and Goldman, 1998; Keysers and Gazzola, 2006; Oberman and
Ramachandran, 2007) and with functional brain imaging (Nummenmaa et al., 2008)
and neuropsychological findings (Shamay-Tsoory, Aharon-Peretz, and Perry, 2009)
which indicate that emotional empathy is linked more closely to sensorimotor
simulation of another’s state than cognitive empathy.
In addition to the differences in emotional empathy reported in chapter 4, the
study presented in chapter 5 sought to establish whether mirror-touch synaesthetes
differed in another aspect of social perception, namely facial expression recognition.
The findings from this study showed that mirror-touch synaesthetes outperformed
non-synaesthete control participants on tasks of facial expression recognition, but not
control tasks involving identity recognition and identity perception. These findings
are consistent with functional brain imaging (Carr et al., 2001; Hennenlotter et al.,
2005; Leslie, Johnsen-Frey, and Grafton, 2004; Montgomery and Haxby, 2008; van
der Gaag, Minderaa, and Keysers, 2007; Winston, O’Doherty, and Dolan, 2003),
neuropsychological (Adolphs et al., 2000; Adolphs, Baron-Cohen, and Tranel, 2002)
and TMS findings (Pitcher et al., 2008; Pourtois et al., 2004) which indicate a central

Chapter 9
170
role for sensorimotor resources in facial expression recognition, and suggest that
facilitated sensorimotor simulation appears to be linked with heightened facial
expression sensitivity and emotional empathy (Chapter 4).
A number of predictions can also be drawn from the evidence of heightened
emotion sensitivity in mirror-touch synaesthesia. For example, given the evidence
that mirror-touch synaesthesia has been reported to be linked to heightened activity in
the mirror-touch system activated by us all when observing touch to others, one may
predict that the extent of activity in this system may correlate with levels of emotional
reactive empathy (but not cognitive empathy – where mirror-touch synaesthetes did
not significantly differ from controls). Further, one may suspect that the level of
activity in sensorimotor cortices when perceiving touch to others should also correlate
with an individual subject’s facial expression recognition abilities. These possibilities
are to be addressed with future studies.
Another interesting point for consideration is whether mirror-touch
synaesthesia represents a distinct population or the tail-end of a distribution of how
much we simulate / empathize with of others6. As noted previously, mirror-touch
synaesthesia shares a number of similarities with other types of developmental
synaesthesia. For example, there is a tendency for mirror-touch synaesthetes to have
other family members with synaesthesia and multiple types of synaesthesia, indicating
a genetic component to the condition. Yet, the principles which bias what type of
synaesthesia will or will not be developed are largely unclear. One could envisage
that if individual differences in emotional sensitivity are in someway heritable /
6 I would like to thank Prof. Christian Keysers for raising and corresponding with me on this issue.

Chapter 9
171
separate from a ‘specific allele’ for synaesthesia7 then heightened emotional
sensitivity may bias these individuals to a form of interpersonal synaesthesia.
9.6 What are the implications of suppressing sensorimotor resources on
expression recognition abilities in healthy adults?
While the findings from chapter 5 assessed the influence of facilitated
sensorimotor simulation on expression recognition, the studies presented in chapters 7
and 8 assessed what impact suppressing sensorimotor resources has on the expression
recognition abilities of healthy adults.
In chapter 7, a continuous theta burst (cTBS) TMS paradigm was used to
suppress cortical activity in the right primary somatosensory cortex (rSI), right lateral
premotor cortex (rPM), and the vertex. Participants completed two tasks. In
experiment 1, participants were asked to complete a same-different auditory
expression recognition task. In experiment 2, a new group of participants were asked
to complete a same-different auditory identity task. Stimuli were non-verbal auditory
emotions (amusement, disgust, fear and sadness), adapted from a previous study
documenting the role of sensorimotor resources in non-verbal auditory emotion
recognition (Warren et al., 2006), and were identical in each task. A comparison
across tasks and sites, revealed that cTBS targeted at rSI and rPM impaired
participants’ abilities to discriminate between the auditory emotions, but not
identities, of others. This was not found to be the case following cTBS to the vertex
(cTBS control site). Therefore consistent with simulation accounts of expression
recognition (Adolphs, 2002; Adolphs, 2003; Damasio, 1990; Gallese, Keysers, and
Rizzolatti, G, 2004; Goldman, and Sripada, 2005; Keysers and Gazzola, 2006),
7 The genetic mechanisms for synaesthesia are of course likely to be more complex than this (e.g. see Asher et al., 2009).

Chapter 9
172
suppressing sensorimotor activity resulted in a task-specific impairment on
participants’ abilities to discriminate emotions but not identities from vocal signals.
These findings add to previous studies documenting a pivotal role for sensorimotor
cortices in facial expression recognition (Adolphs et al., 2000; Pitcher et al., 2008;
Pourtois et al., 2004) and suggest that they are central to the discrimination of emotion
across modalities.
In chapter 8, I again used a cTBS paradigm, but investigated the role of neural
activity in rSI and the right inferior frontal gyrus (rIFG) on facial expression
recognition. Previous findings had indicated that rSI activity is central to facial
expression discrimination (Adolphs et al., 2000; Pitcher et al., 2008; Pourtois et al.,
2004; Winston et al. 2003), but whether this was expression-general or expression-
specific remained a point of dispute. Further, rIFG activity had been reported in a
number of brain imaging experiments investigating the neural correlates of facial
expression recognition or evaluation (Carr et al., 2001; Dapretto et al., 2006;
Hennenlotter et al., 2006; Kesler-West et al., 2001; Seitz et al., 2008), but whether the
region is critical for the facial expression recognition abilities of healthy adults
remained to be clarified. Using a four-forced-choice expression recognition task, the
findings indicated that cTBS to rSI impaired the recognition of emotional facial
expressions (happy and sad) relative to neutral expressions. This is consistent with
previous findings documenting a central role for somatosensory cortices in facial
expression discrimination (Adolphs et al., 2000; Pitcher et al., 2008; Pourtois et al.,
2004) and compliments the findings of chapter 7 by indicating that rSI activity is
involved in discriminating emotional expressions across modalities. No cTBS effect
was observed following stimulation to rIFG or right V5 / MT (visual control site).
This is interesting given that rIFG is considered to be a part of the human mirror

Chapter 9
173
system (Rizzolatti and Craighero, 2004) and has been reported in a number of fMRI
studies on facial expression evaluation (Carr et al., 2001; Dapretto et al., 2006;
Hennenlotter et al., 2006; Kesler-West et al., 2001; Seitz et al., 2008). The lack of
impairment following cTBS to rIFG would suggest that although this region may be
involved in facial expression recognition (Carr et al., 2001; Dapretto et al., 2006;
Hennenlotter et al., 2006; Kesler-West et al., 2001; Seitz et al., 2008), it may not be
critical to the process. Other components of motor simulation may play a more
critical in facial expression discrimination (e.g. premotor cortex as studied in chapter
7) and future studies should address this.
Further possibilities for future research include combining TMS with other
methodologies to consider the role of cortico-cortical interactions play in
discriminating another’s expressions. Moreover, while the effects of online TMS are
spatially discrete, the effects of offline stimulation will spread to other cortical areas
along the greatest lines of conductivity from the stimulated area. By combining cTBS
with fMRI paradigms one should be able to assess any secondary effects of cTBS on
other regions involved in expression recognition.
9.7 General Summary
In summary, this thesis has investigated the neurocognitive and perceptual
profiles of mirror-touch synaesthesia (Chapters 2-5). I have provided a
neurocognitive model of the condition (which provides testable predictions for future
studies) and used mirror-touch synaesthesia as a tool to inform us about the
neurocognitive mechanisms of synaesthesia more generally. The studies presented
have also used mirror-touch synaesthesia as a model to inform us about the impact
that heightened sensorimotor activity has on social cognition (Chapters 4 and 5), and
the findings from these studies are compatible with research presented in chapters 7

Chapter 9
174
and 8 which investigated the impact of suppression of sensorimotor activity on
expression recognition. This has resulted in a number of interesting possibilities for
further studies, some of which are currently in progress and others open to be
conducted.

References
175
REFERENCES
Adolphs, R. (2002). Neural systems for recognizing emotion. Current Opinion in
Neurobiology, 12, 169-77.
Adolphs, R. (2003). Cognitive neuroscience of human social behaviour. Nature
Reviews Neuroscience, 4, 165-178.
Adolphs, R., Baron-Cohen, S., and Tranel, D. (2002). Impaired recognition of social
emotions following amygdala damage. Journal of Cognitive Neuroscience,
14, 1264-1274.
Adolphs, R., Damasio, H., Tranel, D., Cooper, G., and Damasio, A. R. (2000). A role
for somatosensory cortices in the visual recognition of emotion as revealed by
three-dimensional lesion mapping. Journal of Neuroscience, 20, 2683-2690.
Aghajanian, G. K., and Marek, G. J. (1999). Serotonin and hallucinogens.
Neuropsychopharmacology, 21, 16S-23S.
Aleman, A., Rutten, G. J., Sitskoorn, M. M., Dautzenberg, G., and Ramsey, N. F.
(2001). Activation of striate cortex in the absence of visual stimulation: An
fMRI study of synesthesia. Neuroreport, 12, 2827-2830.
Aluja, A., Garcia, O., and Garcia, L. F. (2002). A comparative study of Zuckerman’s
three structural models for personality through the NEO-PI-R, ZKPQ-III-R,
EPQ-RS and Goldberg’s 50-bipolar adjectives. Personality and Individual
Differences, 33, 713-725.
Amassian, V.E., Cracco, R.Q., Maccabee, P.J., Cracco, J.B., Rudell, A.P., and Eberle,
L. (1993). Unmasking human visual perception with the magnetic coil and its
relationship to hemispheric asymmetry. Brain Res, 605, 312-316.

References
176
Amassian, V.E., Eberle, L., Maccabee, P., and Cracco, R.Q. (1992). Modeling
magnetic coil excitation of human cerebral cortex with a peripheral nerve
immersed in a brain-shaped volume conductor: The significance of fiber
bending in excitation. Electroencephalography and Clinical Neurophysiology,
85, 291–301.
Anderson, A. K., Spencer, D. D., Fullbright, R. K., and Phelps, E. A. (2000).
Contribution of the anteromedial temporal lobes to the evaluation of facial
emotion. Neuropsychology, 14, 526-536.
Armel, K. C., and Ramachandran, V. S. (2001). Acquired synethesia in retinitis
pigmentosa. Neurocase, 5, 293-296.
Arzy, S., Thut, G., Mohr, C., Michel, C. M., and Blanke, O. (2006). Neural basis of
embodiment: Distinct contributions of the temporoparietal junction and the
extrastriate body area. Journal of Neuroscience, 26, 8074-8081.
Ashbridge,E., Walsh,V., and Cowey, A. (1997). Temporal aspects of visual search
studied by transcranial magnetic stimulation. Neuropsychologia, 35, 1121-
1131.
Asher, J. E., Lamb, J. A., Brocklebank, D., Cazier, J. B., Maestrini, E., Addis, L., et
al. (2009). A whole-genome scan and fine-mapping linkage study of auditory-
visual synesthesia reveals evidence of linkage to chromosomes 2q24, 5p33,
6p12, and 12p12. Am J Hum Genet, 84, 279-285.
Ashley, V., Vuilleumier, P., and Swick, D. (2004). Time course and specificity of
event-related potentials to emotional expressions. NeuroReport, 15, 211-216.
Augstine, J. R. (1996). Circuitry and functional aspects of the insular lobe in primates
including humans. Brain Res Brain Res Rev., 22, 229-244.

References
177
Avenanti, A., Bueti, D., Galati, G., and Aglioti, S. M. (2005). Transcranial magnetic
stimulation highlights the sensorimotor side of empathy for pain. Nature
Neuroscience, 8, 955-960.
Avenanti, A., Minio-Paluello, I., Bufalari, I., and Aglioti, S. M. (2009). The pain of a
model in the personality of an onlooker: Influence of state-reactivity and
personality traits on embodied empathy for pain. Neuroimage, 44, 275-283.
Banissy, M. J., Cohen Kadosh, R., Maus, G. W., Walsh, V., and Ward, J. (2009).
Prevalence, characteristics, and a neurocognitive model of mirror-touch
synaesthesia. Experimental Brain Research, 198, 261-272.
Banissy, M. J., MacDonald, L., Ward, J., Walsh, V., Haggard, P., and Longo, M. R.
(in preparation). The rubber hand illusion in mirror-touch synaesthesia.
Banissy, M. J., Stewart, L., Ward, J., Walsh, V., and Kanai, R. (in prepartion).
Neuroanatomical correlates of tone-colour synaesthesia.
Banissy, M. J., and Ward, J. (2007). Mirror-touch synaesthesia is linked with
empathy. Nature Neuroscience, 10, 815-816.
Bargary, G., and Mitchell, K. J. (2008). Synaesthesia and cortical connectivity.
Trends in Neurosciences, 31, 335-342.
Barker, A.T., Freeston, I.L., Jalinous, R., Merton, P.A., and Morton, H.B. (1985).
Magnetic stimulation of the human brain. J. Physiol. (Lond.), 369, 3P.
Barnett, K. J., Feeney, J., Gormley, M., and Newell, F. N. (2009). An exploratory
study of linguistic-colour assocaitions across languages in multilingual
synaesthetes. Q J Exp Psychol, 62, 1343-1354.
Barnett, K. J., Finucane, C., Asher, J. E., Bargary, G., Corvin, A. P., Newell, F. N., et
al. (2008). Familial patterns and the origins of individual differences in
synaesthesia. Cognition, 106, 871-893.

References
178
Barnett, K. J., Fox, J. J., Molholm, S., Kelly, S. P., Shalgi, S., Mitchell, K. J., and
Newell, F. N. (2008). Differences in early sensory-perceptual processing in
synaesthesia: A visual evoked potential study. Neuroimage, 43, 605-613.
Barnett, K. J., and Newell, F. N. (2008). Synaesthesia is associated with enhanced,
self rated imagery. Consciousness and Cognition, 17, 1032-1039.
Baron-Cohen, S., Burt, L., Smith-Laittan, F., Harrison, J., and Bolton, P. (1996).
Synaesthesia: Prevalence and familiality. Perception, 25, 1073-1079.
Baron-Cohen, S., Harrison, J., Goldstein, L. H., and Wyke, M. (1993). Coloured
speech perception: Is synaesthesia what happens when modularity breaks
down. Perception, 22, 419-426.
Baron-Cohen, S., Richler, J., Biarya, D., Gurunathan, N., and Wheelwright, S. (2003).
The systemizing quotient: An investigation of adults with Aspergers syndrome
or high-functioning autism, and normal sex differences. Philos Trans R Soc
Lond B Biol Sci., 358, 361-374.
Baron-Cohen, S. and Wheelwright, S. (2004). The empathy quotient: An
investigation of adults with asperger syndrome or high functioning autism, and
normal sex differences. J. Aut. and Dev. Disorders, 34, 163-175.
Baron-Cohen, S., Wyke, M. A., and Binnie, C. (1987). Hearing words and seeing
colours: An experimental investigation of synaesthesia. Perception, 16, 761-
767.
Barrick, M. R., and Mount, R. K. (1991). The Big Five personality dimensions and
job performance: A meta-analysis. Personnel Psychology, 44, 1-26.
Basser, P. J. (1995). Inferring microstructural features and the physiological state of
tissues from diffusion-weighted images. NMR Biomed., 8, 333-344.

References
179
Batson, C. D. (1991). The altruism question: Towards a social-psychology answer.
Erlbaum, Hillside, NJ.
Beck, D., Muggleton, N., Walsh, V., and Lavie, N. (2006). The right parietal cortex
plays a critical role in change blindness. Cereb. Cortex, 16, 712-717.
Beeli, G., Esslen, M., and Jäncke, L. (2005). Synaesthesia: When coloured sounds
taste sweet. Nature, 434, 38.
Beeli, G., Esslen, M., and Jäncke, L. (2008). Time course of neural activity correlated
with colored-hearing synesthesia. Cereb Cortex, 18, 379-385.
Berlucchi, G., and Aglioti, S. (1997). The body in the brain: Neural basis of corporeal
awareness. Trends Neurosci., 20, 560-564.
Bestmann, S., Baudewig, J., Siebner, H.R., Rothwell, J.C., and Frahm, J. (2004).
Functional MRI of the immediate impact of transcranial magnetic stimulation
on cortical and subcortical motor circuits. European Journal of Neuroscience,
19, 1950-1962.
Bjoertomt, O., Cowey, A., and Walsh, V. (2002). Spatial neglect in near and far space
investigated by repetitive transcranial magnetic stimulation. Brain, 125, 2012-
2022.
Blake, R, Palmeri, T.J., Marois, R., and Kim, C. Y. (2004). On the perceptual reality
of synesthetic color. In: L. Robertson and N. Sagiv, Editors, Synesthesia:
Perspectives from Cognitive Neuroscience, Oxford University Press, Oxford
(2004), pp. 47–73.
Blakemore, S. J., Bristow, D., Bird, G., Frith, C., and Ward, J. (2005). Somatosensory
activations during the observation of touch and a case of vision-touch
synaesthesia. Brain, 128, 1571-1583.

References
180
Blanke, O., Landis, T., Spinelli, L., and Seeck, M. (2004). Out-of-body experiences
and autoscopy of neurological origin. Brain, 127, 243-258.
Blanke, O., and Mohr, C. (2005). Out-of-body experience, heautoscopy, and
autoscopic hallucinations of neurological origin: Implications for
neurocognitive mechanisms of corporeal awareness and self-consciousness.
Brain Res Brain Res Rev., 50, 184-199.
Blanke, O., Ortigue, S., Landis, T., and Seeck, M. (2002). Stimulating illusory own
body perceptions. Nature, 419, 269-270.
Bohning, D.E., Shastri, A., McConnell, K.A., Nahas, Z., Lorberbaum, J., and Roberts,
D. (1999). A combined TMS/fMRIstudy of intensity-dependent TMS over
motor cortex. Biol Psychiatry, 45, 385–394.
Botvinick, M., and Cohen, J. (1998). Rubber hands feel touch that I see. Nature, 391,
756.
Bradshaw, J. L., and Mattingley, J. B. (2001). Allodynia: A sensory analogue of
motor mirror neurons in a hyperaesthetic patient reporting instantaneous
discomfort to another’s perceived sudden minor injury? J Neurol Neurosurg
Psychiatry, 70, 135-136.
Bremmer, F., Schlak, A., Shah, N. J., Zafiris, O., Kubishik, M., Hoffmann, K., et al.
(2001). Polymodal motion processing in posterior parietal and premotor
cortex: A human fMRI study strongly implies equivalences between humans
and monkeys. Neuron, 29, 287-296.
Bruce, V., and Young, A. W. (1986). Understanding face recognition. Br. J. Psychol.,
77, 305-327.
Brugger, P. (2002). Reflective mirrors: Persepective-taking in autoscopic phenomena.
Cognitive Neuropsychiatry, 7, 179-194.

References
181
Bryant, G., and Barrett, H. C. (2007). Recognizing intentions in infant-directed
speech: Evidence for universals. Psychological Science, 18, 746-751.
Buccino, G., Binkofski, F., Fink, G. R., Fadiga, L., Fogassi, L., Gallese, V., et al.
(2001). Action observation activates premotor and parietal areas in a
somatotopic manner. European Journal of Neuroscience, 13, 400-404.
Bufalari, I., Aprile, T., Avenanti, A., Di Russo, F., and Aglioti, S. M. (2007).
Empathy for pain and touch in the human somatosensory cortex. Cereb
Cortex, 17, 2553-2661.
Bünning, S., and Blanke, O. (2005). The out-of body experience: Precipitating factors
and neural correlates. Prog Brain Res., 150, 331-350.
Burrack, A., Knoch, D., and Brugger, P. (2006). Mitempfindung in synaesthetes: Co-
incidence or meaningful association? Cortex, 42, 151-154.
Bushara, K. O., Grafman, J., and Hallett, M. (2001). Neural correlates of auditory-
visual stimulus onset asynchrony detection. J Neurosci, 21, 300-304.
Calder, A. J., Keane, J., Lawrence, A. D., and Manes, F. (2004). Impaired recognition
of anger following damage to the ventral striatum. Brain, 127, 1958-1969.
Calder, A. J., Keane, J., Manes, F., Antoun, N., and Young, A. W. (2000). Impaired
recognition and experience of disgust following brain injury. Nature
Neuroscience, 3, 1077-1078.
Calder, A. J., Lawrence, A. D., and Young, A. W. (2001). The neuropsychology of
fear and loathing. Nature Reviews Neuroscience, 2, 352-363.
Calder, A. J., and Young, A. W. (2005). Understanding the recognition of facial
identity and facial expression. Nature Reviews Neuroscience, 6, 641-651.
Campana, G., Cowey, A., and Walsh, V. (2002). Priming of motion direction and
area V5/MT: A test of perceptual memory. Cereb Cortex, 12, 663-669.

References
182
Carr, L., Iacoboni, M., Dubeau, M. C., Mazziotta, J. C., and Lenzi, G. L. (2003).
Neural mechanisms of empathy in humans: A relay from neural systems for
imitation to limbic areas. Proc. Natl. Acad. Sci., 100, 5497-5502.
Cohen, L., and Dehaene, S. (2004). Specialization within the ventral stream: The case
for the visual word form area. Neuroimage, 22, 466-476.
Cohen Kadosh, R., Cohen Kadosh, K., and Henik, A. (2007). The neural correlate of
bidirectional synaesthesia: A combined ERP and fMRI study. Journal of
Cognitive Neuroscience, 19, 2050-2059.
Cohen Kadosh, R., and Henik, A. (2006). When a line is a number: Colour yields
magnitude information in a digit-colour synaesthete. Neuroscience, 137, 3-5.
Cohen Kadosh, R., and Henik, A. (2007). Can synaesthesia research inform cognitive
science? Trends in Cognitive Sciences, 11, 177-184.
Cohen Kadosh, R., Henik, A., and Walsh, V. (2007). Small is bright and big is dark
in synaesthesia. Current Biology, 17, R834-R835.
Cohen Kadosh, R., Henik, A., Catena, A., and Walsh, V. (2009). Induced Cross-
Modal Synesthetic Experience without Abnormal Neuronal Connections.
Psychological Science, 20, 258-265.
Cohen Kadosh, R., Tzelgov, J., and Henik, A. (2008). A synethetic walk on the
mental number line: The size effect. Cognition, 106, 548-557.
Cohen Kadosh, R., and Walsh, V. (2006). Cognitive neuroscience: Rewired or
crosswired brains. Current Biology, 16, 962-963.
Cohen Kadosh, R., and Walsh, V. (2008). Synaesthesia and Cortical Connections:
Cause or Correlation? Trends in Neurosciences, 31, 549-550.
Colby, C. L. (1998). Action-orientated spatial reference frames in cortex. Neuron,
20, 15-24.

References
183
Coslett, B. (1998). Evidence for a disturbance of the body schema in neglect. Brain
and Cognition, 37, 527-544.
Costa, P. T., and McCrae, R. R. (1985). The NEO Personality Inventory manual.
Odessa, FL: Psychological Assessment Resources.
Cowey, A., and Gross, C. G. (1970). Effects of foveal prestriate and inferotemporal
lesions on visual discrimination by rhesus monkey. Experimental Brain
Research, 11, 128-144.
Craig, A. D. (2002). How do you feel? Interoception: The sense of the physiological
condition of the body. Nature Reviews Neuroscience, 3, 655-666.
Crawford, J. R., and Garthwaite, P. H. (2002). Investigation of the single case in
neuropsychology: Confidence limits on the abnormality of test scores and test
scores differences. Neuropsyhcologia, 40, 1196-1208.
Cytowic, R. E., and Wood, F. B. (1982). Synaesthesia II: Psychophysical relations in
the synaesthesia of geometrically shaped taste and colored hearing. Brain and
Cognition, 1, 36-49.
Damasio, A. R. (1990). Time-locked multiregional retroactivation: A systems-level
proposal for the neural substrates of recall and recognition. In Neurobiology
of Cognition, Edited by Eimas, P. D., and Galaburda, A. M. MIT Press,
Cambridge, 25-62.
Dapretto, M., Davies, M. S., Pfeifer, J. H., Scott, A. A., Sigman, M., Bookheimer, S.
Y., et al. (2006). Understanding emotions in others: Mirror neuron dysfunction
in children with autism spectrum disorders. Nature Neuroscience, 9, 28-30.
Davidoff, J. (2001). Language and perceptual categorisation. Trends in Cognitive
Sciences, 5, 382-387.

References
184
Davis, M. H. (1980). A multidimensional approach to individual differences in
empathy. Catal. Sel. Doc. Psychol., 10, 85.
Davis, M. H. (1994). Empathy: A social psychology approach. Colorado: Westview
Press.
Day, S. (2005). In: L. Robertson and N. Sagiv, Editors, Synesthesia: Perspectives
from Cognitive Neuroscience, Oxford University Press, Oxford (2004),
Decety, J., and Jackson, P. L. (2004). The functional architecture of human empathy.
Behav Cogn Neurosci Rev., 3, 71-100.
del Barrio, V., Aluja, A., and Garcia, L. F. (2004). Relationship between empathy and
the Big Five personality traits in a sample of Spanish adolescents. Social
Behavior and Personality, 32, 677-681.
De Waal, F. B. (2007). Putting the altruism back into altruism: The evolution of
empathy. Ann Rev Psychol, 59, 1-22.
Di Lazzaro, V., Pilato, F., Saturno, A., Oliviero, M., Dileone, M., Mazzone, P., et al.
(2005). Theta-burst repetitive transcranial magnetic stimulation suppresses
specific excitatory circuits in the human motor cortex. J. Physiology, 565 945-
950.
Dietrich, S., Szameitat, D., Ackermann, H., and Alter, K. (2006). How to disentangle
lexical and prosodic information? Psychoacoustic studies on the processing of
vocal interjections. Prog Brain Res. 156, 295-302.
Dimberg, U., Thunberg, M., Elmehed, K. (2000). Unconcious facial reactions to
emotional facial expressions. Psychological Science, 11, 86-89.
Dixon, M. J., Smilek, D., and Merikle, P. M. (2004). Not all synaesthetes are created
equal: Projector versus associator synaesthetes. Cogn Affect Behav Neurosci,
4, 335-343.

References
185
Donner, T. H., Kettermann, A., Diesch, E., Ostendorf, F., Villringer, A., and Brandt,
S. A. (2002). Visual feature and conjunction searches of equal difficulty
engage only partially overlapping frontoparietal networks. Neuroimage, 15,
16-25.
Downing, P.E., Jiang, Y., Shuman, M., and Kanwisher, N. A. (2001). Cortical area
selective for visual processing of the human body. Science, 293, 2470-2473.
Duchaine, B., and Nakayama, K. (2006). The Cambridge Face Memory Test: Results
from neurologically intact individuals and an investigation of its validity using
inverted performance and prosopagnosic subjects. Neuropsychologia, 16,
166-173.
Duchaine, B., Germine, L., and Nakayama, K. (2007). Family resemblance: Ten
family members with prosopagnosia and within-class object agnosia.
Cognitive Neuropsychology, 24, 419-430.
Duhamel, J. R., Colby, C. L., and Goldberg, M. E. (1998). Ventral intraparietal area
of the macaque: Congruent visual and somatic response properties. Journal
of Neurophysiology, 79. 126-136.
Dumoulin, S.O., Bittar, R. G., Kabani, N. J., Baker, C. L. Jr, Le Goualher, G., Bruce
Pike, G., et al. (2000). A new anatomical landmark for reliable identification
of human area V5 / MT: A quantative analysis of sulcal patterning. Cereb
Cortex, 10, 454-463.
Eagleman, D. M., Kagan, A. D., Nelson, S. S., Sagaram, D., and Sarma, A. K. (2007).
A standardized test battery for the study of synaesthesia. Journal of
Neurosciences Methods, 159, 139-145.

References
186
Ebisch, S. J. H., Perucci, M. G., Ferretti, A., Del Gratta, C., Luca Romani, G., and
Gallese, V. (2008). The sense of touch: Embodied simulation in a visuo-
tactile mirroring mechanism for observed animate or inanimate touch. Journal
of Cognitive Neuroscience, 20, 1611-1623.
Eimer, M., and Holmes, A. (2002). An ERP study on the time course of emotional
face processing. NeuroReport, 13, 427-431.
Eimer, M., and Holmes, A. (2007). Event-related brain potential correlates of
emotional face processing. Neuropsychologia, 45, 15-31.
Eimer, M., Holmes, A., and McGlone, F. (2003). The role of spatial attention in the
processing of facial expression: An ERP study of rapid brain responses to six
basic emotions. Cognitive, Affective and Behavioural Neuroscience, 3, 97-110.
Eisenberg, N. (2000). Emotion, regulation, and moral development. Annu Rev
Psychol., 51, 665-697.
Ekman, P., and Friesen, W. V. (1976). Pictures of facial affect. Palo Alto, CA:
Consulting Psychologists.
Elias, L. J., Saucier, D. M., Hardie, C., and Sart, C. E. (2003). Dissocaiting semantic
and perceptual components of synaesthesia: Behavioural and functional
neuroanatomical investigations. Cognitive Brain Research, 16, 232-237.
Esterman, M., Verstynen, T., Ivry, R. B., and Robertson, L. C. (2006). Coming
unbound: Disrupting automatic integration of synesthetic color and graphemes
by transcranial magnetic stimulation of the right parietal lobe. J Cogn
Neuroci, 18, 1570-1576.
Fadiga, L., Fogassi, L., Pavesi, G., and Rizzolatti, G. (1995). Motor facilitation during
action observation: A magnetic stimulation study. J. Neurophsyiology, 73,
2608-2611.

References
187
Fink, G. R., Markowitsch, H. J., Reinkemeier, M., Bruckbauer, T., Kessler, J., and
Heiss, W. D. (1996). Cerebral representations of one’s own past: Neural
networks involved in autobiographical memory. Journal of Neuroscience, 16,
4275-4282.
Flynn, F. G., Benson, D. F., and Ardila, A. (1999). Anatomy of the insula –
Functional and clinical correlates. Aphasiology, 13, 55-78.
Franz, E. A., Ford, S., and Werner, S. (2007). Brain and cognitive processes of
imitation in bimanual situations: Making inference about mirror neuron
systems. Brain research, 1145, 138-149.
Friedman-Hill, S. R., Robertson, L. C., and Treisman, A. (1995). Parietal
contributions to visual feature binding: Evidence from a patient with bilateral
lesions. Science, 269, 853-855.
Gallagher, S. (1995). Body schema and intentionality. In J. L. Bermudez, A. Marcel,
and N. Eilan (Eds.), The body and the self, Cambridge, MA: MIT Press.
Gallese, V. (2006). Intentional atonement: A neurophysiological perspective on
social cognition and its disruption in autism. Brain Res Cog Brain Res., 1079,
15-24.
Gallese, V., Fadiga, L., Fogassi, L., and Rizzolatti, G. (1996). Action recognition in
the premotor cortex. Brain, 119, 593-609.
Gallese, V., and Goldman, A. (1998). Mirror neurons and simulation theory of mind
reading. Trends in Cognitive Sciences, 12, 493-501.
Gallese, V., Keysers, C., and Rizzolatti, G. (2004). A unifying view of the basis of
social cognition. Trends in Cognitive Sciences, 8, 396-403.
Galton, F. (1880). Visualised numerals. Nature (1880), 252-256.

References
188
Garrido, L., Furl, N., Draganski, B., Weiskopf, N., Stevens, J., Tan, G. C-Y., et al. (in
press) VBM reveals reduced grey matter volume in the temporal cortex of
developmental prosopagnosics. Brain.
Gazzola, V., Aziz-Zadeh, L., and Keysers, C. (2006). Empathy and the somatotopic
auditory mirror system in humans. Current Biology, 16, 1824-1829.
Giummarra, M. J., Gibson, S. J., Georgiou-Karistianis, N., and Bradshaw, J. L.
(2008). Mechanisms underlying embodiment, disembodiment and loss of
embodiment. Neuroscience and Biobehavioural Reviews, 32, 143-160.
Goldberg, L. R. (1992). The development of markers for the Big-Five factor
structure. Psychological Assessment, 4, 26-42.
Goldman, A., and Sripada, C. S. (2005). Simulationist models of face-based emotion
recognition. Cognition, 94, 193-213.
Goldreich, D., and Kanics, I. M. (2003). Tactile acuity is enhanced in blindness.
Journal of Neuroscience, 23, 3439-3445.
Goller, A., Otten, L., and Ward, J. (2009). Seeing sounds and hearing colours: An
event-related potential study of audio-visual synaesthesia. Journal of
Cognitive Neuroscience, 21, 1869-1881.
Gray, J. A., Chopping, S., Nunn, J., Parslow, D., Gregory, L., Williams, S., et al.
(2002). Implications of synaesthesia for functionalism. Journal of
Consciousness Studies, 9, 5-31.
Gray, J. A., Parslow, D. M., Brammer, M. J., Chopping, S., Vythelingum, G. N., and
ffytche, D. H. (2006). Evidence against functionalism from neuroimaging of
the alien colour effect in synaesthesia. Cortex, 42, 309-318.

References
189
Grossenbacher, P.G., and Lovelace, C.T. (2001). Mechanisms of synaesthesia:
Cognitive and physiological constraints. Trends in Cognitive Sciences, 5, 36-
41.
Hadjikhani, N., and Roland, P. E. (1998). Cross-modal transfer of information
between the tactile and the visual representations in the human brain: A
positron emission tomographic study. Journal of Neuroscience, 18, 1072-
1084.
Haggard, P. (2006). Just seeing you makes me feel better: Interpersonal enhancements
of touch. Social Neuroscience, 1, 104-110.
Halligan, P. W., Hunt, M., Marshall, J. C., and Wade, D. T. (1996). When seeing is
feeling: Acquired synaesthesia or phantom touch? Neurocase, 2, 21-29.
Harris, J. A., Harris, I. M., Diamond, M. E., (2001). The topography of tactile
working memory. J Neurosci, 21, 8262-8269.
Haxby, J. V., Hoffman, E. A., and Gobbini, M. I. (2000). The distributed human
neural system for face processing. Trends in Cognitive Sciences, 4, 223-233.
Head, H., and Holmes, G. (1911-1912). Sensory disturbances from cerebral lesions.
Brain, 34, 102-254.
Hennenlotter, A., Schroeder, U, Erhard, P., Castrop, F., Haslinger, B., Stoecker, D.,
et al. (2005). A common neural basis for receptive and expressive
communication of pleasant facial affect. Neuroimage, 26, 581-591.
Hoekert, M., Bais, L., Kahn, R. S., and Aleman, A. (2008). Time course of the
involvement of the right anterior superior temporal gyrus and the right fronto-
parietal operculum in emotional prosody perception. PlOS One 3, doi:
10.1371/journal.pone.0002244.

References
190
Holmes, A., Vuilleumier, P., and Eimer, M. (2003). The processing of emotional
facial expressions is gated by spatial attention: Evidence from event-related
brain potentials. Cognitive Brain Research, 16, 174-184.
Hong, S. W. and Blake, R. (2008). Early visual mechanisms do not contribute to
synaesthetic colour experience. Vision Research, 48, 1018-1026.
Howell, D. C. (2002). Statistical Methods for Psychology. Thomson Learning: USA.
Huang, Y. Z., Edwards, M. J., Rounis, E., Bhatia, K. P., and Rothwell, J. C. (2005).
Theta burst stimulation of the human motor cortex. Neuron, 45, 201-206.
Huang, Y. Z., Rothwell, J. C., Lu, C. S., Wang, J., Weng, Y. H., Lai, S. C., et al.
(2009). The effect of continuous theta burst stimulation over premotor cortex
on circuits in primary motor cortex and spinal cord. Clin Neurophysiol., 120,
796-801.
Hubbard, E. M. (2007). A real red-letter day. Nat Neurosci, 10, 671-672.
Hubbard, E. M., Arman, A. C., Ramachandran V. S., and Boynton, G. M. (2005).
Individual differences among grapheme-color synesthetes: brain-behavior
correlations. Neuron, 45, 975–985.
Hubbard, E. M., Manohar, S., and Ramachandran, V. S. (2006). Contrast affects the
strength of synesthetic colors. Cortex, 42, 184-194.
Hubbard, E.M., and Ramachandran, V. S. (2003). Refining the experimental lever: A
reply to Shannnon and Pribram. J Consciousness Stud, 9, 77-84.
Hubbard, E.M., and Ramachandran, V. S. (2005). Neurocognitive mechanisms of
synaesthesia. Neuron, 48, 509-520.
Iacoboni, M., and Dapretto, M. (2006). The mirror neuron system and the
consequences of its dysfunction. Nat Rev Neurosci, 7, 942-951.

References
191
Iriki, A., Tanaka, M., and Iwamura, Y. (1996). Coding of modified body schema
during tool use by macaque postcentral neurones. Neuroreport, 7, 2325-2330.
Jabbi, M., Swart, M., and Keysers, C. (2007). Empathy for positive and negative
emotions in the gustatory cortex. Neuroimage, 34, 1744-1753.
Jäncke, L., Beeli, G., Eulig, C., and Hänggi, J. (2009). The neuroanatomy of
grapheme-color synethesia. Eur J Neurosci, 29, 1287-1293.
Jewanski, J., Day, S. A., and Ward, J. (2009). A colourful albino: The first
documented case of synaesthesia, by George Tobias Ludwig Sachs in 1812.
Journal of the History of Neurosciences, 18, 1-11.
John, O. P., Donahue, E. M., and Kentle, R. L. (1991). The Big Five Inventory –
Versions 4a and 54. Berkeley: University of California, Berkeley, Institute of
Personality and Social Research.
John, O. P. and Srivastava, S. (1999). The Big Five trait taxonomy: History,
measurement and theoretical perspectives. In Handbook of personality:
Theory and research (2nd ed.), Edited by Pervin, L., and John, O. P. New
York: Guilford.
Johnson, R., Burton, P., and Ro, T. (2006). Visually induced feelings of touch. Brain
Research, 1073-1074, 398-406.
Johnsen, K. O., and Philips, J. R. (1981). Tactile spatial resolution. I. Two-point
discrimination, gap detection, grating resolution, and letter recognition.
Journal of Neurophysiology, 46, 1177-1191.
Juan, C. H., and Walsh, V. (2003). Feedback to V1: A reverse hierarchy in vision. Exp
Brain Res., 150, 259-263.

References
192
Kalla, R., Muggleton, N., Cowey, G., and Walsh, V. (2009). Human dorsolateral
prefrontal cortex is involved in visual search for conjunctions but not features:
A theta TMS study. Cortex 45, 1085-1090.
Kammer. T. (1999). Phosphenes and transient scotomas induced by magnetic
stimulation of the occipital pole: their topographic relationship.
Neuropsychologia, 37, 191–198.
Kaplan, E. (1991). The receptive field of retinal ganglion cells in cat and monkey. In:
Leventhal, A. G. (Ed.), Vision and Visual Dysfunction. CRC Press: Boston. pp
10-40.
Kauffman, T., Théoret, H., Pascual-Leone, A. (2002). Braille character discrimination
in blindfolded human subjects. Neuroreport, 13, 571-574.
Keane, J., Calder, A. J., Hodges, J. R., and Young, A. W. (2002). Face and emotion
processing in frontal variant frontotemporal dementia. Neuropsychologia, 40,
655-665.
Kesler-West, M. L., Andersen, A. H., Smith, C. D., Avison, M. J., Davis, C. E.,
Kryscio, R. J., et al. (2001). Neural substrates of facial emotion processing
using fMRI. Brain Res. Cogn. Brain Res., 11, 213-226.
Keysers, C., and Gazzola, V. (2006). Towards a unifying theory of social cognition.
Progress in Brain Research, 156, 379-401.
Keyers, C., Wicker, B., Gazzola, V., Anton, J. L., Fogassi, L., and Gallese, V. (2004).
A touching sight: SII/PV activation during the observation and experience of
touch. Neuron, 42, 335-346.
Kim, C. Y., Blake, R., and Palmeri, T. J. (2006). Perceptual interaction between real
and synesthetic colors. Cortex, 42, 195–203.

References
193
Kipps, C. M., Duggins, A. J., McCusker, E. A., and Calder, A. J. (2007). Disgust and
happiness recognition correlate with anteroventral insula and amygdala
volume respectively in preclinical Huntigton’s disease. Journal of Cognitive
Neuroscience, 19, 1206-1217.
Kircher, T. T., Senior, C., Phillips, M. L., Rabe-Hesketh, S., Benson, P. J., Bullmore,
E. T., et al. (2001). Recognizing one’s own face. Cognition, 78, B1-B15.
Konen, C. S., and Kaster, S. (2008). Two hierarchically organized neural systems for
object information in human visual cortex. Nature Neuroscience, 11, 224-231.
Krolak-Salmon, P., Henaff, M. A., Isnard, J., Tallon-Baudry, C., Guenot, M.,
Vighetto, A., et al. (2003). A specific response to disgust moudlated by
attention in human ventral anterior insula. Annals of Neurology, 53, 446-453.
Lamm, C., Batson, C. D., and Decety, J. (2007). The neural substrate for human
empathy: Effects of perspective-taking and cognitive appraisal. Journal of
Cognitive Neuroscience, 19, 42-58.
Lavidor, M., and Walsh, V. (2003). A magnetic stimulation examination of
orthographic neighborhood effects in visual word recognition. Journal of
Cognitive Neuroscience, 15, 354-363.
Lawrence, A. D., Calder, A. J., McGowan, S. W., and Grasby, P. M. (2002). Selective
disruption of the recognition of facial expressions of anger. Neuroreport, 13,
881-884.
Lawrence, E. J., Shaw, P., Baker, D., Baron-Cohen, S., and David, A. S. (2004).
Measuring empathy: Reliability and validity of the Empathy Quotient.
Psychological Medicine, 34, 911-924.

References
194
Leslie, K. R., Johnson-Frey, S. H., and Grafton, S. T. (2004). Functional imaging of
face and hand imitation: Towards a motor theory of empathy. Neuroimage,
21, 601-607.
Lewis, S., Thoma, R. J., Lanoue, M. D., Miller, G. A., Heller, W., Edgar, C., et al.
(2003). Visual processing of facial affect. Neuroreport, 14, 1841-1845.
Lomber, S. (1999). The advantages and limitations of permanent or reversible
deactivation techniques in the assessment of neural function. Journal of
Neuroscience Methods, 86, 109-118.
Longo, M. R., Schüür, F., Kammers, M. P., Tsakiris, M., and Haggard, P. (in press).
Self awareness and the body image. Acta Psychologica.
Lundqvist, D., Flykt, A., and İhman, A. (1998). The Karolinska Directed Emotional
Faces (KDEF). Stockholm: Department of Neurosciences Karolinska
Hospital.
Macaluso, E., and Driver, J. (2003). Multimodal spatial representations in the human
parietal cortex: Evidence from functional neuroimaging. Adv Neurology, 93,
219-233.
MacDonald, D., Kabani, N., Avis, D., and Evans, A. C. (2000). Automated 3-D
extraction of inner and outer surfaces of cerebral cortex from MRI.
Neuroimage, 12, 340-356.
MacLeod, C. M., and Dunbar, K. (1988). Training and stroop-like interference:
Evidence for a continuum of automaticity. J Exp Pscyhol Learn Mem Cogn,
14, 126-135.
Maravita, A., and Iriki, A. (2002). Tools for the body (schema). Trends in Cognitive
Sciences, 8, 79-86.

References
195
Marks, L. E. (1975). On coloured hearing synaesthesia: Cross-modal translations of #
sensory dimensions. Psychological Bulletin, 82, 303-331.
Marzi, C. A., Miniussi, C., Maravita, A., Bertolasi, L., Zanette, G., Rothwell, J. C.,
and Sanes, J. N. (1998). Transcranial magnetic stimulation selectively impairs
interhemispheric transfer of visuo-motor information in humans. Exp. Brain.
Res., 118, 435-438.
McCabe, C., Rolls, E. T., Bilderbeck, A., and McGlone, F. (2008). Cognitive
influences on the affective representations of touch and the sight of touch in
the human brain. Social Cognitive and Affective Neuroscience, 3, 97-108.
Mesulam, M. M., and Mufson, E. (1982). Insula of the old world monkey, III:
Efferent cortical output and comments on function. Journal of Comparative
Neurology, 212, 38-52.
Meyer, M., Zysset, S., von Cramon, D. Y., and Alter, K. (2005). Distinct fMRI
responses to laughter, speech, and sounds along the human perisylvian cortex.
Cog Brain Res. 24, 291-306.
Milner, B. (1966). Amnesia following operation on the temporal lobes. In Amnesia,
C.W.M. Whitty and O.L. Zangwill (eds.). London: Botterworths, pp 109-133.
Mills, C. B., Boteler, E. H., and Oliver, G. K. (1999). Digit synaesthesia: A case
study using a stroop-type test. Cognitive Neuropsychology, 16, 181-191.
Mitchell, R. L., and Crow, T. J. (2005). Right hemisphere language functions and
schizophrenia: The forgotten hemisphere? Brain, 128, 963-978.
Montgomery, K. J., and Haxby, J. (2008). Mirror neuron system differentially
activated by facial expressions and social hand gestures: A functional
magnetic resonance imaging study. Journal of Cognitive Neuroscience, 20,
1866-1877.

References
196
Morrison, I., Lloyd, D., di Pellegrino, G., and Roberts, N. (2004). Vicarious responses
to pain anterior cingulated cortex: Is empathy a multisensory issue? Cogn
Affect Behav Neurosci, 4, 270-278.
Mottaghey, F. M., Gangitano, M., Sparing, R., Krause, B. J., Pascual-Leone, A.
(2002). Segregation of areas related to visual working memory in the
prefrontal cortex revealed by rTMS. Cereb Cortex, 12, 369-375.
Mufson, E., and Mesulam, M. M. (1982). Insula of the old world monkey, II:
Afferent cortical input and comments on the claustrum. Journal of
Comparative Neurology, 212, 23-37.
Muggleton, N., Tsakanikos, E., Walsh, V., and Ward, J. (2007). Disruption of
synaesthesia following TMS of the right posterior parietal cortex.
Neuropsychologia, 45, 1582-1585.
Muncer, S., and Ling, J. (2006). Psychometric analysis of the empathy quotient (EQ)
scale. Personality and Individual Differences, 40, 1111-1119.
Nagarajan, S.S., Durand, D., and Warman, E.N. (1993). Effects of induced electric
fields on finite neuronal structures: a simulation study. IEEE Trans. in
Biomedical Engineering, 40, 1175-1188.
Nikolić, D., Lichti, P., and Singer, W. (2007). Color opponency in synaesthetic
experiences. Psychological Science, 18, 481-486.
Nummenmaa, L., Hirvonen, J., Parkkola, R., and Hietanen, J. K. (2008). Is emotional
contagion special? An fMRI study on neural systems for affective and
cognitive empathy. Neuroimage, 43, 571-580.
Nunn. J. A., Gregory, L. J., Brammer, M., Williams, S. C., Parslow, D. M., Morgan,
M .J., et al. (2002). Functional magnetic resonance imaging of synesthesia:
Activation of V4 / V8 by spoken words. Nature Neuroscience, 5, 371-375.

References
197
Oberman, L. M., Hubbard, E. M., McCleery, J. P., Altschuler, E. L., Ramachandran,
V. S., and Pineda, J.A. (2005). EEG evidence of mirror neuron dysfunction in
autism spectrum disorders. Brain Res Cogn Brain Res., 24, 190-198.
Oberman, L. M., and Ramachandran, V. S. (2007). The simulating social mind: The
role of the mirror neuron system and simulation in the social and
communicative deficits of autism spectrum disorders. Psychol Bull., 133, 310-
327.
Oberman, L.M. , Winkielman, P., and Ramachandran, V. S. (2007). Face to face:
Blocking facial mimicry can selectively impair recognition of emotional
expressions. Social Neuroscience, 2, 167-178.
Olausson, H., Lamarre, Y., Backlund, H., Morin, C., Wallin, B. G., Starck, G.,
Ekholm, S., Strigo, I., Worsley, K., Valbo, Ǻ. B., and Bushnell, M. C. (2002).
Unmyelinated tactile afferents signal touch and project to insular cortex.
Nature Neuroscience, 5, 900-904.
Oliveri, M., Turriziani, P., Carlesimo, G. A., Koch, G., Tomaiuolo, F., Panella, M., et
al. (2001). Parieto-frontal interactions in visual-object and visual-spatial
working memory. Cereb Cortex, 11, 606-618.
Olson, I. R., Gatenby, J. C., and Gore, J. C. (2002). A comparison of bound and
unbound audio-visual information processing in the human cerebral cortex.
Cognitive Brain Research, 14, 129-138.
O'Shea, J., Muggleton, N. G., Cowey, A., and Walsh, V. (2004). Timing of target
discrimination in human frontal eye fields. Journal of Cognitive Neuroscience,
16, 1060-1067.

References
198
Parise, C. V., and Spence, C. (2009). ‘When birds of a feather flock together’:
Synesthetic correspondances modulate audiovisual integration in non-
synesthetes. PloS One, 4, e5664.
Pascual-Leone, A., Amedi, A., Fregni, F., and Merabet, L. B. (2005). The plastic
human brain cortex. Ann. Rev. Neuroscience, 28, 377-401.
Paulesu, E., Harrison, J., Baron-Cohen, S., Watson, J. D., Goldstein, L., Heather, J., et
al. (1995). The physiology of coloured hearing. A PET activation study of
colour-word synaesthesia. Brain, 118, 661-676.
Peelen, M. V., and Downing, P. E. (2005). Selectivity for the human body in
fusiform gyrus. Journal of Neurophysiology, 93, 603-608.
Petrides, M., Cadoret, G., and Mackey, S. (2005). Orofacial somatomotor responses in
the macaque monkey homolouge of Broca’s area. Nature, 435, 1235-1238.
Philips, M. L., Young, A. W., Scott, S. K., Calder, A. J., Andrew, C., Giampietro, V.,
et al. (1998). Neural responses to facial and vocal expressions of fear and
disgust. Proc. Biol. Sci., 265, 1809-1817.
Philips, M. L., Young, A. W., Senior, C., Brammer, M., Andrew, C., Calder, A. J., et
al. (1997). A specific substrate for perceiving facial expressions of disgust.
Nature, 389, 495-498.
Pitcher, D., Charles, L., Devlin, J. T., Walsh, V., and Duchaine, B. C. (2009). Triple
dissociation of faces, bodies, and objects in extrastraite cortex. Current
Biology, 19, 319-324.
Pitcher, D., Garrido, L., Walsh, V., and Duchaine, B. C. (2008). Transcranial
magnetic stimulation disrupts the perception and embodiment of facial
expressions. Journal of Neuroscience, 28, 8929-8933.

References
199
Pitcher, D., Walsh, V., Yovel, G., and Duchaine, B. C. (2008). TMS evidence for the
involvement of the right occipital face area in early face processing. Current
Biology, 17, 1568-1573.
Pourtois, G., Sander, D., Andres, M., Grandjean, D., Reveret, L., Olivier, E., et al.
(2004). Dissociable roles of the human somatosensory and superior temporal
cortices for processing social face signals. Eur. J. Neurosci., 20, 3507-3515.
Preston, S. D., and de Waal, F. B. (2002). Empathy: It’s ultimate and proximal basis.
Behav Brain Sci., 25, 1-20.
Ramachandran, V. S., and Hubbard, E. (2001). Synaesthesia – A window into
perception, thought and language. Journal of Consciousness Studies, 8, 3-34.
Rao, A., Nobre, A. C., Alexander, I., and Cowey, A. (2007). Auditory evoked visual
awareness following sudden ocular blindness: An EEG and TMS
investigation. Exp Brain Res, 176, 288-298.
Rich, A. N., and Mattingley, J. B. (2002). Anomalous perception in synaesthesia: A
cognitive neuroscience. Nature Reviews Neuroscience, 3, 43-52.
Rich, A. N., Bradshaw, J. L., and Mattingley, J. B. (2005). A systematic large-scale
study of synaesthesia: Implications for the role of early experience in lexical-
colour assocaitions. Cognition, 98, 53-84.
Rich, A. N., Williams, M. A., Pruce, A., Syngeniotis, A., Howard, M. A., McGlone,
F., et al. (2006). Neural correlates of imagined and synaesthetic colours.
Neuropsychologia, 44, 2918-2925.
Rizzo, M., and Eslinger, P. J. (1989). Colored hearing synesthesia: An investigation
of neural factors. Neurology, 39, 781-784.
Rizzolatti, G., and Craighero, L. (2004). The mirror-neuron system. Annual Review
of Neuroscience, 27, 169-192.

References
200
Ro, T., Farnè, A., Johnson, R. M., Wedeen, V., Chu, Z., Wang, Z. J., et al. (2007).
Feeling sounds after a thalamic lesion. Ann Neurol., 62, 433-441.
Robertson, L. C. (2003). Binding, spatial attention, and perceptual awarness. Nat Rev
Neurosci., 4, 93-102.
Robertson, D., Davies, I., and Davidoff, J. (2000). Colour categories are not
universal: Replications and new evidence from a stone-age culture. Journal of
Experimental Psychology, 129, 369-398.
Robertson, I. H., and Murre, J. M. J. (1999). Rehabilitation of brain damage: Brain
plasticity and principles of guided recovery. Psychological Bulletin, 125, 544-
575.
Rorden, C., Heutink, J., Greenfield, E., and Robertson, I. H. (1999). When a rubber
hand ‘feels’ what the real hand cannot. NeuroReport, 10, 135-138.
Rouw, R., and Scholte, H. S. (2007). Increased structural connectivity in grapheme-
colour synesthesia. Nat Neurosci., 10, 792-797.
Ruby, P., and Decety, J. (2001). Effect of subjective perspective taking during
simulation of action: A PET investigation of agency. Nature Neuroscience, 4,
546-550.
Ruff, C.C., Blankenburg, F., Bjoertomt, O., Bestmann, S., Freeman, E., Haynes, J-D.,
Rees, G., Josephs, O., Deichmann, R., and Driver, J. (2006). Concurrent TMS-
fMRI and psychophysics reveal frontal influences on human retinotopic visual
cortex. Current Biology, 16, 1479-1488
Rushworth, M., Ellison, A., and Walsh, V. (2001). Complementary localization and
lateralization of orienting and motor attention. Nature Neuroscience, 4, 656-
661.

References
201
Russell, R., Duchaine, B., and Nakayama, K. (2009). Super-recognizers: People with
extraordinary face recognition ability. Psychonomic Bulletin and Review, 16,
252-257.
Saarela, M. V., Hlushchuk, Y., Williams, A. C., Schürmann, M., Kalso, E., and Hari,
R. (2007). The compassionate brain: Humans detect intensity of pain from
another’s face. Cereb Cortex, 17, 230-237.
Sachs, G. T. L. (1812). Historiae naturalis duorum leucaetiopum: Auctoris ipsius et
sororis eius. Erlangen.
Saenz, M., and Koch, C. (2008). The sound of change: Visually-induced auditory
synesthesia. Current Biology, 18, 650-651.
Sagiv, N. (2004). In: L. Robertson and N. Sagiv, Editors, Synesthesia: Perspectives
from Cognitive Neuroscience, Oxford University Press, Oxford (2004),
Sagiv, N., Simner, J., Collins, J., Butterworth, B., and Ward, J. (2006). What is the
relationship between synaesthesia and visuo-spatial number forms?
Cognition, 101, 114-128.
Sagiv, N., and Ward, J. (2006). Cross-modal interactions: Lessons from Synaesthesia.
Progress in Brain Research, 155, 263-275.
Sauter, D. A., and Eimer, M. (in press). Rapid detection of emotion from human
vocalizations. Journal of Cognitive Neuroscience.
Sauter, D. A., and Scott, S. K. (2007). More than one kind of happiness: Can we
recognise vocal expressions of different positive states? Motivation and
Emotion, 31, 192-199.
Saxe, R., Jamal, N., and Powell, L. (2006). My body or yours? The effect of visual
perspective on cortical body representations. Cerebral Cortex, 16, 178-182.

References
202
Schiltz, K., Trocha, K., Wieringa, B. M., Emirch, H. M., Johannes, S., and Münte, T.
F. (1999). Neurophysiological aspecrs of synesthetic experience. J
Neuropsychiatry Clin Neurosci., 11, 58-65.
Schofield, W. N. (1976). Do children find movements which cross the body midline
difficult? Q. J. Exp. Psychol. 28, 571-582.
Schröder, M. (2003). Experimental study of affect bursts. Speech communication, 40,
99-116.
Schulte-Rüther, M., Markowitsch, H. J., Fink, G. R., and Piefke, M. (2008). Miror
neuron and theory of mind mechanisms involed in face-to-face interactions: A
functional magnetic resonance imaging approach to empathy. Journal of
Cognitive Neuroscience, 19, 1354-1372.
Scott, S. K., Sauter, D. A., and McGettigan, C. (In press). Brain mechanisms for
processing perceived emotional vocalizations in humans. To appear in
Handbook of Mammalian Vocalizations: A Neuroscience Approach, S
Brudzynski ed. (Elsevier).
Scott, S. K., Young, A. W., Calder, A. J., Hellawell, D. J., Aggleton, J. P., and
Johnsen, M. (1997). Impaired auditory recognition of fear and anger following
bilateral amygdala lesions. Nature, 385, 254-257.
Seitz, R. J., Schäfer, R., Scherfield, D., Friederichs, S., Popp, K., Wittsack, H. J., et al.
(2008). Valuating other people’s emotional face expression: A combined
functional magnetic resonance imaging and electroencephalography study.
Neuroscience, 152, 713-722.
Serino, A., Pizzoferrato, F., and Làdavas, E. (2008). Viewing a face (especially one’s
own face) being touched enhances tactile perception on the face.
Psychological Science, 19, 434-438.

References
203
Seron, X., Pesenti, M., Nöel, M. P., Deloche, G., and Cornet, J. A. (1992). Images of
numbers, or ‘When 98 is upper left and 6 is sky blue”. Cognition, 44, 159-
196.
Shallice, T. (1988). The neuropsychology of mental structure. Cambridge: Cambridge
University Press.
Shamay-Tsoory, S. G., Aharon-Peretz, J., and Perry, D. (2009). Two systems for
empathy: A double dissociation between emotional and cognitive empathy in
inferior frontal gyrus versus ventromedial prefrontal lesions. Brain, 132, 617-
627.
Shaw, P., Greenstein, D., Lerch, J., Clasen, L., Lenroot, R., Gogtay, N., et al. (2006).
Intellectual ability and cortical development in children and adolescents.
Nature, 440, 676-679.
Siebner HR, Willoch F, Peller M, Auer C, Boecker H, Conrad B, and Bartenstein P.
1998). Imaging brain activation induced by long trains of repetitive
transcranial magnetic stimulation. NeuroReport, 9, 943-948.
Simner, J. (2007). Beyond perception: Synaesthesia as a psycholingustic
phenomenon. Trends in Cognitive Sciences, 11, 23-29.
Simner, J., and Holenstein, E. (2007). Ordinal linguistic personification as a variant
of synaesthesia. Journal of Cognitive Neuroscience, 19, 694-703.
Simner, J., Mulvenna, C., Sagiv, N., Tsakanikos, E., Witherby, S., Fraser, C., Scott,
K., and Ward, J. (2006). Synaesthesia: The prevalence of atypical cross-modal
experiences. Perception, 35, 1024-1033.
Simner, J., Ward, J., Lanz, M., Jansari, A., Noonan, K., Glover, L., and Oakley, D.
(2005). Non-random associations of graphemes and colour in the synaesthetic
and normal populations. Cognitive Neuropsychology, 22, 1069-1085.

References
204
Singer, T., Seymour, B., O’Doherty, J., Kaube, H., Dolan, R. J., and Frith, C. D.
(2004). Empathy for pain involves the affective but not sensory components of
pain. Science, 303, 1157-1162.
Singh, K.D., Hamdy, S., Aziz, Q., and Thompson, D.G. (1997). Topographic mapping
of trans-cranial magnetic stimulation data on surface rendered MR images of
the brain. EEG Clin Neurophysiol, 105, 345-351.
Sirigu, A., Grafman, J., Bressler, K., and Sunderland, T. (1991). Multiple
representations contribute to body knowledge processing. Brain, 114, 629-
642.
Smilek, D., Dixon, M. J., Cudahy, C., and Merikle, P. M. (2001). Synaesthetic
photisms influence visual perception. Journal of Cognitive Neuroscience, 13,
930-936.
Smilek, D., Malcolmson, K. A., Carriere, J., Eller, M., Kwan, D., and Reynolds, M.
(2007). When “3” is a jerk and “E” is king: Personifying inanimate objects in
synaesthesia. Journal of Cognitive Neuroscience, 19, 981-992.
Smilek, D., Moffatt, B. A., Pasternak, J., White, B. N., Dixon, M. J., and Merikle, P.
M. (2002). Synaesthesia: A case study of discordant monozygotic twins.
Neurocase, 8, 338-342.
Smith, H. L. (1905). Synesthesia. John Hopkins Hospital Bulletin, 16, 258-263.
Southgate, V., and Hamilton, A. F. (2008). Unbroken mirrors: Challenging a theory
of autism. Trends in Cognitive Sciences, 12, 225-229.
Sperling. J. M., Prvulovic, D., Linden, D. E., Singer, W., and Stirn, A. (2006).
Neuronal correlates of colour-graphemic synaesthesia: A fMRI study. Cortex,
42, 295-303.

References
205
Spiller, M. J., and Jansari, A. S. (2008). Mental imagery and synaesthesia: Is
synaesthesia from internally-generated stimuli possible. Cognition, 109, 143-
151.
Sprengelmeyer, R., Raush, M., Eysel, U. T., and Przuntek, H. (1998). Neural
substrates associated with recognition of facial expressions of basic emotions.
Proc. Biol. Sci., 265, 1927-1931.
Sprengelmeyer, R., Young, A. W., Calder, A. J., Karnat, A., Lange, H., Hömberg, V.,
et al. (1996). Loss of disgust: Perception of faces and emotions in
Huntington’s disease. Brain, 119, 1647-1665.
Sprengelmeyer, R., Young, A. W., Mahn, K., Schroeder, U., Woitalla, D., Buttner, T.,
et al. (2003). Facial expression recognition in people with medicated and
unmedicated Parkinson’s disease. Neuropsychologia, 41, 1047-1057.
Stefan, K., Gentner, R., Zeller, D., Dang, S., and Classen, J. (2008). Theta-burst
stimulation: Remote physiological and local behavioural after-effects.
Neuroimage, 40, 265-274.
Steven, M. S., and Blakemore, C. (2004). Visual synaesthesia in the blind.
Perception, 33, 855-868.
Steven, M. S., Hansen, P. C., and Blakemore, C. (2006). Activation of color-selective
areas of the visual cortex in a blind synesthete. Cortex, 42, 304-308.
Stewart, L., Ellison, A., Walsh, V., and Cowey, A. (2001). The role of transcranial
magnetic stimulation in studies of vision, attention and language. Acta
Psychologica, 107, 275-291.
Stewart, L., Mulvenna, C., Griffiths, T., and Ward, J. (in prep). Neural correlates of
audio-visual synaesthesia induced from tone sequences.

References
206
Stewart, L., Walsh, V., Frith, U., and Rothwell, J. C. (2001). TMS produces two
dissociable types of speech arrest. Neuroimage, 13, 472-478.
Sugiura, M., Watanabe, J., Maeda, Y., Matsue, Y., Fukuda, H., and Kawashima, R.
(2005). Cortical mechanisms of visual self-recognition. Neuroimage, 24,
143-149.
Tang, J., Ward, J., and Butterworth, B. (2008). Number forms in the brain. Journal
of Cognitive Neuroscience, 20, 1547-1556.
Taylor, J. C., Wiggett, A. J., and Downing, P. E. (2007). Functional MRI analysis of
body and body part representations in the extrastriate and fusiform body areas.
Journal of Neurophysiology, 98, 1626-1633.
Taylor-Clarke, M., Kennett, S., and Haggard, P. (2004). Persistence of visual-tactile
enhancement in humans. Neuroscience Letters, 354, 22-25.
Terao, Y., Fukuda, H., Ugawa, Y., Hikosaka, O., Furubayashi, T., Hanajima, R.,
Sakai, K., and Kanazawa, I. (1998). Visualization of the information through
human oculomotor cortical regions by transcranial magnetic stimulation. J
Neurophysiol, 80, 936-946.
Terao, Y., Ugawa, Y., Suzuki, M., Sakai, K., Hanajima, R., Gemba-Shimizu, K., and
Kanazawa, I. (1997). Shortening of simple reaction time by peripheral
electrical and submotor-threshold magnetic cortical stimulation. Experimental
Brain Research, 115, 541-545.
Terhune, D. B. (2009). The incidence and determinants of visual phenomenology
during out-of-body experiences. Cortex, 45, 236-242.
Thilo, K. V., Santoro, L., Walsh, V., and Blakemore, C. (2004). The site of saccadic
suppression. Nature Neuroscience, 7, 13-14.

References
207
Treisman, A. (2004). In: L. Robertson and N. Sagiv, Editors, Synesthesia:
Perspectives from Cognitive Neuroscience, Oxford University Press, Oxford
(2004).
Tsakiris, M., and Haggard, P. (2005). The rubber hand illusion revisited: Visuotactile
integration and self-attribution. J Exp Psychol Hum Percept Perform., 31, 80-
91.
Tyrell, R. A., and Owens, D. A. (1988). A rapid technique to assess the resting states
of eyes and other threshold phenomena: The modified binary search (MOBS).
Behav. Res. Methods Instrum. Comput., 20, 137-141.
Uddin, L. Q., Iacoboni, M., Lange, C., and Keenan, J. P. (2007). The self and social
cognition: The role of cortical midline structures and mirror neurons. Trends
in Cognitive Sciences, 11, 153-157.
Uddin, L. Q., Kaplan, J. T., Molnar-Szakacs, I., Zaidel, E., and Iacoboni, M. (2005).
Self-face recognition activates a frontoparietal “mirror” network in the right
hemisphere: An event –related fMRI study. Neuroimage, 25, 926-935.
Uddin, L. Q., Molnar-Szakacs, I., Zaidel, E., and Iacoboni, M. (2006). rTMS to the
right inferior parietal lobule disrupts self-other recognition. Soc. Cogn. Affect.
Neurosci., 1, 65-71.
Ueno, T., Tashiro, T., and Harada, K. (1988). Localized stimulation of neural tissues
in the brain by means of a paired configuration of time-varying magnetic
fields. J. Appl. Phys, 64, 5862–5864.
Vallesi, A., Shallice, T., and Walsh, V. (2007). Role of the prefrontal cortex in the
foreperiod effect: TMS evidence for dual mechanisms in temporal preparation.
Cerebral Cortex, 17, 466-474.

References
208
Van Boven, R. W., and Johnson, K. (1994). A psychophysical study of the
mechanisms of sensory recovery following nerve injury in humans. Brain,
117, 149-167.
Van Boven, R. W., Hamilton, R. H., Kauffman, T., Keenan, P., and Pascual-Leone, A.
(2000). Tactile spatial resolution in blind Braille readers. Neurology, 54, 2230-
2236.
van der Gaag, C., Minderaa, R. B., and Keysers, C. (2007). Facial expressions: What
the mirror neuron system can and cannot tell us. Social Neuroscience, 2, 179-
222.
Van Lacker, D., and Fromkin, V. A. (1973). Hemispheric specialization for pitch and
tone: Evidence from Thai. Journal of Phonetics, 1, 101-109.
van Rijn, S., Aleman, A., van Diessen, E., Berckmoes, C., Vingerhoets, G., Kahn, R.
S. (2005). What is said or how it is said makes a difference: Role of the right
fronto-parietal operculum in emotional prosody as revealed by repetitive TMS.
Eur J. Neurosci. 21, 3195-3200.
Vega-Bermudez, F., and Johnsen, K. O. (1999). SA1 and RA receptive fields,
response variability, and population response mapped with a probe array.
Journal of Neurophysiology, 81, 2701-2710.
Wade, A. R., Brewer, A. A., Rieger, A. W., and Wandell, B. A. (2002). Functional
measurements of human ventral occipital cortex: Retinotopy and colour.
Philos. Trans. R. Soc. Lond. B. Biol. Sci., 357, 963-973.
Walsh, V., and Butler, S. R. (1996). Different ways of looking at seeing. Behavioural
Brain Research, 76, 1-3.
Walsh, V., and Cowey, A. (2000). Transcranial magnetic stimulation and cognitive
neuroscience. Nature Reviews Neuroscience, 1, 73-79.

References
209
Walsh, V., and Pascual-Leone, A. (2003). Transcranial Magnetic Stimulation: A
neurochromometrics of mind, (MIT Press).
Walsh, V., and Rushworth, M. (1999). A primer of magnetic stimulation as a tool for
neuropsychology. Neuropsychologia, 37, 125-135.
Ward, J. (2004). Emotionally mediated synaesthesia. Cognitive Neuropsychology,
71, 761-772.
Ward, J., Banissy, M. J., and Jonas, C. (2008). Haptic perception in synaesthesia. In
Human Haptic Perception: Basics and Applications, Edited by
M.Grunwald. Birkhäuser, Basel.
Ward, J., Huckstep, B., Tsakanikos, E. (2006). Sound-colour synaesthesia: To what
extent does it use cross-modal mechanisms common to us all. Cortex, 42,
264-280.
Ward, J., Li, R., Salih, S., and Sagiv, N. (2007). Varieties of grapheme-colour
synaesthesia: A new theory of phenomenological and behavioural differences.
Conscious Cogn., 16, 913-931.
Ward, J., and Mattingley, J. B. (2006). Synaesthesia: An overview of contemporary
findings and controversies. Cortex, 42, 129-136.
Ward, J., and Simner, J. (2003). Lexical-gustatory synaesthesia: Linguistic and
conceptual factors. Cognition, 89, 237-261.
Ward, J., and Simner, J. (2005). Is synaesthesia an X-linked trait lethality in males?
Perception, 34, 611-623.
Ward, J., Simner, J., and Auyeung, V. (2005). A comparison of lexical-gustatory and
grapheme-colour synaesthesia. Cognitive Neuropsychology, 22, 28-41.

References
210
Warren, J. E., Sauter, D. A., Eisner, F., Wiland, J., Dresner, M. A., Wise, R. J. S., et
al. (2006). Positive emotions preferentially engage an auditory-motor “mirror”
system. Journal of Neuroscience, 26, 13067-13075.
Wassermann, E. M. (1998). Risk and safety of repetitive transcranial magnetic
stimulation: Report and suggested guidelines from the International Workshop
on the Safety of Repetitive Transcranial Magnetic Stimulation.
Electroencephalogr Clin Neurophysiol., 108, 1-16.
Wassermann, E.M., McShane, L.M., Hallett, M., and Cohen, L.G. (1992).
Noninvasive mapping of muscle representations in human motor cortex.
Electroencephalogr Clin Neurophysiol, 85, 1-8.
Weiss, P. H., Shah, N. J., Toni, I., Zilles, K., and Fink, G. R. (2001). Associating
colours with people: A case of chromatic-lexical synaesthesia. Cortex, 37,
750-753.
Weiss, P. H., Zilles, K., and Fink, G. R. (2005). When visual perception causes
feelings: Enhanced cross-modal processing in grapheme-color synesthesia.
Neuroimage, 28, 859-868.
Wheeler, R. H., and Cutsforth, T. D. (1921). The role of synaesthesia in learning. J
Exp Psychology, 4, 448-468.
Wicker, B., Keysers, C., Plailly, J., Royet, J. P., Gallese, V., and Rizzolatti, G. (2003).
Both of us disgusted in my insula: The common neural basis of seeing and
feeling disgust. Neuron, 40, 655-664.
Wig, G., Grafton, S., Demos, K., and Kelley, W. (2005). Specificity of priming: a
cognitive neuroscience perspective. Nat. Neurosci. 8, 1228 – 1234.

References
211
Wildgruber, D., Riecker, A., Hertrich, I., Erb, M., Grodd, W., Ethofer, T., et al.
(2005). Identification of emotional intonation evaluated by fMRI.
Neuroimage, 24, 1233-1241.
Winston, J., O’Doherty, J., and Dolan, R. (2003). Common and distinct neural during
direct and incidental processing of multiple facial emotions. Neuroimage, 20,
84-97.
Wittenberg, G. F., Werhahn, K. J., Wassermann, E. M., Herscovitch, P., and Cohen,
L. G. (2004). Functional connectivity between somatosensory and visual
cortex in early blind humans. European Journal of Neuroscience, 20, 1923-
1927.
Witthoft, N., and Winawer, J. (2006). Synesthetic colors determined by having
refrigerator magents in childhood. Cortex, 42, 175-183.
Yaro, C., and Ward, J. (2007). Searching for Shereshevski: What is superior about
the memory of synaesthetes? Quarterly Journal of Experimental Psychology,
60, 681-695.

Appendix
212
Appendix 1: Questionnaire used to recruit potential synaesthetes in Experiment 2.1
(Chapter 2).
PLEASE READ THIS FIRST People with synaesthesia experience certain things (e.g. colours, tastes) when engaged in activities (e.g. reading) that would not elicit such a response in non-synaesthetic people. For instance, colours may be experienced in response to music or words, or shapes may be experienced in response to tastes. Synaesthesia is quite rare, but the questionnaire below asks whether you regularly have these types of experiences.
Everything you write will be treated in confidence, in accordance with the data protection act. You do not have to answer any questions if you feel uncomfortable about them. We may wish to contact a small number of people (by phone/e-mail/letter) to invite them to take part in a further study of memory and perception. None of the tasks are harmful or stressful. It would be helpful then, if you included contact details below, in case you are one of the people we would like to contact. You are in no way obliged to part in any further experiments. Your personal details (name, email, etc.) will not be passed on to anybody else.
Name: _________________________________________________________ Age: ______________ Profession/Degree Course: ______________________________________ Year: ________________ Telephone number: __________________________ E-mail: ___________________________
(1) Do you think about the letters of the alphabet (and/or words and numbers) as having specific colours (i.e. the letter A is experienced as red)? Strongly disagree Disagree Neither agree nor disagree Agree Strongly agree If SO, Which ones? Letters Words Numbers
Other? ______________
(2) Do you think about the letters of the alphabet (and/or days of the week/months of the
year/numbers) as being arranged in a specific pattern in space? Strongly disagree Disagree Neither agree nor disagree Agree Strongly agree If SO, Which ones? Letters Days Months Numbers
Other? ______________
(3) Do you experience taste sensations when you observe another person eating or
drinking something (i.e. observing someone eating strawberries and experiencing a sweet taste in your own mouth)?
Strongly disagree Disagree Neither agree nor disagree Agree Strongly agree

Appendix
213
(4) Do you experience touch sensations on your own body when you see them on another person’s body?
Strongly disagree Disagree Neither agree nor disagree Agree Strongly agree
(5) When experiencing touch to your own body do you experience visual sensations (i.e.
colour)?
Strongly disagree Disagree Neither agree nor disagree Agree Strongly agree (6) Do these experiences have specific locations (e.g. on your body, on words or objects
in the environment, in front of your eyes) or not (e.g. they feel as if they are in ‘your minds eye’)? Please describe. _____________________________________________________________________ _______________________________________________________________________________________________________________________________________________________________________________________________________________
(7) To the best of your knowledge have you always had these sensations?
YES NO DON’T KNOW If YES – at what age did you realise that other people did not have the same sensations as you? _____________________________________________________________________
If NO – at what age did they arise and was there a triggering incident?
_____________________________________________________________________
(8) Do the sensations that you have to particular things change over time or are they fixed
(e.g. if the word ‘book’ is green then is it always green and always has been)?
FIXED VARIABLE DON’T KNOW
(9) On the next page please match the triggers on the left with synaesthetic experiences on the right. For instance, if you experience colours in response to numbers then draw a line in between ‘numbers’ (left) and ‘colours’ (right).
IMPORTANT: Please do not connect the same things (e.g. colours–colours) as this is assumed true of everyone. We also assume (without you having to indicate) that letters/ words etc. as
experienced as shapes on a written page, and musical instruments/ voices/ spoken words as noise.
Moreover, if you have no synaesthetic experiences then there is no need to connect any triggers
with experiences.

Appendix
214
TRIGGERS EXPERIENCES Letters of alphabet Colours English words Shapes/Patterns Foreign words Tastes People’s names Smells Addresses/places Pains/touches Numbers Noises Days of week Flashes Months of year Music Pains/touches Movements Music (instrumental) Noises Smells Tastes Colours Shapes/Patterns
Thanks for your time.


















