
Migration and aggregation of embryonic chicken neurons in vitro: possible functional implication of...
12
Developmental Brain Research, 57 (1990) 223-234 223 Elsevier BRESD 51188 Migration and aggregation of embryonic chicken neurons in vitro: possible functional implication of polysialogangliosides Christian Greis and Harald R6sner Institute of Zoology, University of Hohenheim-Stuttgart, Stuttgart (F.R. G.) (Accepted 28 August 1990) Key words: Primary neuronal culture; Cell migration; Cell aggregation; Fasciculation; Development; Polysialoganglioside; Modulatory cell surface molecule The presented study reports a primary culture system of embryonic chicken optic lobe neurons, which turned out to be a suitable model for cell migration and aggregation: Freshly dissociated neurons developed short processes, contacted one another and formed fasciculated bundles, on which neurons migrated as long-shaped cells, similar to migrating neurons in vivo. We used this system to study the possible involvement of c-pathway polysialogangliosidesfor neuronal migration and aggregation. These highly negative charged glycosphingolipids are the predominant gangliosides of migrating and outgrowing neurons in vivo. Addition of a purified ganglioside mixture (50/~M), extracted from brains of the corresponding embryonic stage, strongly enhanced neuronal migration and aggregation, while incubation of the cells with monoclonal antibody Q211, specifically binding c-polysialogangliosides, reduced aggregate formation in a dose-dependent manner. Cultures treated with 10 pg/ml Q211, instead, displayed a more divergent growth, leading to the formation of a fine network of single neuronal processes. These results suggest a functional implication of c-polysialogangliosides in neuronal fasciculation, migration and aggregation. INTRODUCTION The development of vertebrate nervous system in- volves spatial and temporal patterned differentiation processes, leading to appropriate, highly ordered neuro- nal connections. The underlying basic events are cell proliferation, cell migration, cell aggregation, cyto-dif- ferentiation and formation of specific neuronal connec- tions and pathways 4. The genetic regulation of these events is epigenetically modulated by specific interactions of membrane-bound surface molecules with receptor molecules of neighbour cells and extracellular matrix TM ~8,24, which are suggested to act in a mechanochemicai manner, directly mediating cellular processes 12. There is strong evidence that glycolipids in addition to or in combination with glycoproteins are involved in such modulatory events, e.g. during developmental differen- tiation or oncogenesis 6'18"28. Gangliosides are neuraminic acid-containing glycosphingolipids, predominantly local- ized at the surface of the plasma membrane and espe- cially concentrated in neuronal cells. Because of their location and structural diversity, they are well suited to function as cell surface receptors and as modulators of various membrane processes, playing a distinctive role in cellular recognition and interaction 17'~8, as well as in neuronal differentiation 25"42. In embryonic chicken brain, striking changes of the ganglioside pattern have been shown to parallel neuro- genesis and progress in neuronal growth and matu- ration 1°'23'34'36'37. Therefore, gangliosides have been des- ignated as cytochemical markers of neuronal develop- ment and maturation 34. However, the functional involvement of gangliosides remained an open question. To get more information about the function of ganglio- sides in neuronal development and differentiation, we established a primary cell culture from embryonic chicken retina and optic lobes, which turned out as a simple and highly reproducible model for neuronal migration, aggregation, sprouting and contact initiation. In this study, we investigated the migration and aggre- gation of embryonic chicken neurons in vitro, with respect to the possible implication of polysialoganglio- sides, which are the major gangliosides of these neurons during migration and aggregation in vivo 34"36 MATERIALS AND METHODS Abbreviations Abbreviations of gangliosides follow the nomenclature of Sven- nerholm 46. PBS, phosphate buffered saline; BSA, bovine serum albumine; DMEM, Dulbecco's modified Eagle's medium; HEPES, 4-(2-hydroxyethyl)-2-piperazine-ethanesulfonic acid; HPTLC, high- performance thin-layer chromatography; mAb, monoclonal antibody. Correspondence: H. ROsner, Institute of Zoology (220), University of Hohenheim-Stuttgart, Garbenstrasse 30, D-7000 Stuttgart-70, F.R.G. 0165-3806/90/$03.50 © 1990 Elsevier Science Publishers B.V. (Biomedical Division)
-
Upload
christian-greis -
Category
Documents
-
view
214 -
download
2
Transcript of Migration and aggregation of embryonic chicken neurons in vitro: possible functional implication of...

Developmental Brain Research, 57 (1990) 223-234 223 Elsevier
BRESD 51188
Migration and aggregation of embryonic chicken neurons in vitro: possible functional implication of polysialogangliosides
Christian Greis and Harald R6sner
Institute of Zoology, University of Hohenheim-Stuttgart, Stuttgart (F. R. G.)
(Accepted 28 August 1990)
Key words: Primary neuronal culture; Cell migration; Cell aggregation; Fasciculation; Development; Polysialoganglioside; Modulatory cell surface molecule
The presented study reports a primary culture system of embryonic chicken optic lobe neurons, which turned out to be a suitable model for cell migration and aggregation: Freshly dissociated neurons developed short processes, contacted one another and formed fasciculated bundles, on which neurons migrated as long-shaped cells, similar to migrating neurons in vivo. We used this system to study the possible involvement of c-pathway polysialogangliosides for neuronal migration and aggregation. These highly negative charged glycosphingolipids are the predominant gangliosides of migrating and outgrowing neurons in vivo. Addition of a purified ganglioside mixture (50/~M), extracted from brains of the corresponding embryonic stage, strongly enhanced neuronal migration and aggregation, while incubation of the cells with monoclonal antibody Q211, specifically binding c-polysialogangliosides, reduced aggregate formation in a dose-dependent manner. Cultures treated with 10 pg/ml Q211, instead, displayed a more divergent growth, leading to the formation of a fine network of single neuronal processes. These results suggest a functional implication of c-polysialogangliosides in neuronal fasciculation, migration and aggregation.
INTRODUCTION
The deve lopment of ver tebra te nervous system in-
volves spatial and tempora l pa t t e rned differentiat ion
processes, leading to appropr ia te , highly ordered neuro-
nal connections. The underlying basic events are cell
prol i fera t ion, cell migrat ion, cell aggregation, cyto-dif-
ferent ia t ion and format ion of specific neuronal connec-
tions and pathways 4. The genetic regulat ion of these
events is epigenet ical ly modula ted by specific interactions
of membrane -bound surface molecules with receptor
molecules of ne ighbour cells and extracel lular matrix TM
~8,24, which are suggested to act in a mechanochemicai
manner , directly media t ing cellular processes 12. There is
s t rong evidence that glycolipids in addi t ion to or in
combinat ion with glycoproteins are involved in such
modula tory events, e.g. during deve lopmenta l differen-
t iat ion or oncogenesis 6'18"28. Gangl iosides are neuraminic
acid-containing glycosphingolipids, p redominant ly local-
ized at the surface of the p lasma membrane and espe-
cially concent ra ted in neuronal cells. Because of their
locat ion and structural diversity, they are well suited to
function as cell surface receptors and as modula tors of
various membrane processes, playing a distinctive role in
cellular recognit ion and interact ion 17'~8, as well as in neuronal different ia t ion 25"42.
In embryonic chicken brain, str iking changes of the
ganglioside pa t te rn have been shown to paral le l neuro-
genesis and progress in neuronal growth and matu- ration 1°'23'34'36'37. Therefore , gangliosides have been des-
ignated as cytochemical markers of neuronal develop-
ment and matura t ion 34. However , the functional
involvement of gangliosides remained an open question.
To get more informat ion about the function of ganglio-
sides in neuronal deve lopment and different iat ion, we
establ ished a pr imary cell culture from embryonic
chicken ret ina and optic lobes, which turned out as a
simple and highly reproducib le model for neuronal
migrat ion, aggregat ion, sprout ing and contact init iation.
In this study, we invest igated the migrat ion and aggre-
gation of embryonic chicken neurons in vitro, with
respect to the possible implicat ion of polysialoganglio-
sides, which are the ma jo r gangliosides of these neurons during migrat ion and aggregat ion in vivo 34"36
MATERIALS AND METHODS
Abbreviations Abbreviations of gangliosides follow the nomenclature of Sven-
nerholm 46. PBS, phosphate buffered saline; BSA, bovine serum albumine; DMEM, Dulbecco's modified Eagle's medium; HEPES, 4-(2-hydroxyethyl)-2-piperazine-ethanesulfonic acid; HPTLC, high- performance thin-layer chromatography; mAb, monoclonal antibody.
Correspondence: H. ROsner, Institute of Zoology (220), University of Hohenheim-Stuttgart, Garbenstrasse 30, D-7000 Stuttgart-70, F.R.G.
0165-3806/90/$03.50 © 1990 Elsevier Science Publishers B.V. (Biomedical Division)

224
Media and buffers PBS (Dulbecco) contained (per liter): 8.0 g NaCI, 0.2 g KCI, 1.44 g
Na2HPO4.2H~O, 0.2 g KH2PO 4, and was adjusted to pH 7.4. Cell buffer consisted of PBS, supplemented with 1 g/l glucose. Preparation medium consisted of DMEM (Sigma), supplemented with 0.37 g/l NaHCO 3 and 2.4 g/l HEPES, and was adjusted to pH 7.4. Culture medium consisted of DMEM (Sigma), supplemented with 3.7 g/l NaHCO 3, and was adjusted to pH 7.4. All buffers and media used for isolation and culture of cells were steril filtered through a 0.2/tin filter (Sartorius).
Monoclonal antibodies Anti-neurofilament 200 antibody (IgG) was purchased as ascites
fluid from BioMakor (Israel). Anti-polysialoganglioside antibody Q211 (IgM) was prepared from Hybridoma cells 2° and used as ascites fluid (protein concentration of 18 mg/ml) or alternatively as ammonium sulphate precipitate. The specificity of mAb Q211, recognizing c- pathway polysialogangliosides with 3 and more sialic acid residues has been shown previously 16'37. Secondary antibodies, anti-mouse-IgG- fluorescein isothiocyanate (FITC), anti-mouse-IgM-FITC and anti- mouse-IgM-alkaline phosphatase, were affinity-isolated antibodies, purchased from Sigma (Munich).
Preparation of embryonic chicken neurons Fertilized eggs (White Leghorn) were kept at 37.8 °C in a humidified
incubator. Embryos from 5 to 15 days (C5-C15), corresponding to Hamburger and Hamilton ~9 stages 28-40, were used for the establish- ment of primary cultures. After decapitation, the heads were immedi- ately transferred in cold preparation medium. All following steps were done under sterile conditions. Brains and eyes were removed and collected in cold preparation medium (4 °C). Optic lobes and retinae were carefully dissected under microscopic control, freed of adhering tissue and washed twice in cold preparation medium. For dissociation, the tissue was transferred into tubes with 2 ml cell buffer, containing 0.5 mg trypsin and 0.05 mg DNAse I (Serva), carefully triturated 20 times with a 5 ml plastic pipette and incubated for 20 min at 37 °C in an agitating water bath. After 10 and 20 min, trituration was repeated as described above. Dissociation was stopped by adding 5 ml of cold preparation medium, and cells were collected by centrifugation (10 rain, 200 g) in a cooled centrifuge (4 °C). Cells were washed twice by
resuspension in cold preparation medium and ccntrifugation ~s described above. Afterwards. cells were suspended in 2 ml cold preparation medium and stored on ice. An aliquot was taken fol counting the cells in a Thoma counting chamber. Isolated cells usually possessed a viability /> 95%.
Cell culture Cells were cultured on 35 mm tissue culture dishes (Greiner). The
surfaces were precoated with polylysine, using the following protocol: each 35 mm dish was filled with 1.5 ml sterile solution of poly-D-lysine (Sigma) in aqua dest. (16 ~tg/ml), and incubated under repeated agitation for 60 min at ambient temperature. Then, the solution was removed by aspiration, and the dishes were washed 3 times with sterile aqua dest. to remove free polylysine. The dishes were air dried and sterilized for 30 min by UV radiation.
Cells were seeded at a density of 5-10 ~' cells/dish and cultivated in culture medium (see above) in a humidified incubator with 5% CO 2 and 95% air at 37 °C. After 30 min, the medium was supplemented by addition of 5% heat-inactivated horse serum, 2% fetal calf serum, 100 U/ml penicillin, 100 /~g/ml streptomycin and 10 k~g/ml tetracychn (Sigma). Living cultures were counted and photographed at different culture times with a Zeiss Axiovert microscope, using phase-contrast or interference-contrast optics, respectively.
lmmunostaining of cell cultures Cultures were washed 3 times with PBS, fixed for 2 h with 3.8%
freshly prepared paraformaldehyde in PBS, and washed again 3 times with PBS. For labelling of membrane-bound polysialogangliosides, cultures were overlayed with mAb Q211, diluted 1:400 in PBS containing 10 mg/ml BSA, and incubated 1 h at 20 °C. Then the cultures were washed 3 times with PBS and overlayed with anti- mouse-IgM-FITC, diluted 1:400 in PBS containing I0 mg/ml BSA (1 h, 20 °C). Afterwards, the cultures were washed again 3 times and covered with PBS.
For the labelling of intracellular neurofilaments, cultures were first treated for 15 min with 0.1% (v/v) Triton X-100 in PBS, to permeabilize the cell membranes. Then the cultures were washed 3 times in PBS and stained as described above, using anti-neurofila- ment 200 antibody (1:800) as primary, and ant i ,mouse-IgG'FITC (1:400) as secondary antibody.
Fig. 1. Migration and aggregation of C7 optic lobe cells in vitro at indicated time intervals after plating: at 5 min cells were evenly distributed and adhered to the ground; with increasing culture time, cells formed fasciculating processes, which were used as migration routes between increasing aggregates; insets demonstrate magnifications of cells stained by the neuron-specific mAb Q211; interference contrast, bar = 200 /*m (insets: 70/*m).
225
Immunostained cultures were examined and photographed with a Universal microscope (Zeiss) with epifluorescence and alternative phase contrast illumination, using BP 546, FT 580 and LP 590 filters.
Isolation and purification of embryonic chicken gangliosides Eggs (White Leghorn) were incubated for 11 days at 37.8 °C as
described above. Embryos were decapitated and the optic lobes were removed and freed of meninges and connective tissue. The lobes were pooled, immediately frozen at -80 °C, lyophylilized and stored at -80 °C. The samples were pre-extracted twice with ice-cold acetone 37, to remove lipophilic contaminants. Gangliosides were extracted according to the method of Svennerholm and Fredman 47, and dialyzed 3 days against running deionized water. The ganglio- side extracts were further purified by LH-20 gel chromatography (Pharmacia), to remove lipophilic protein contaminations 2.
Pure GM1, used for control experiments, was a gift of Fidia, Abano Terme, Italy.
Characterization of isolated gangliosides The purified gangliosides were dried, resolved in water and
lyophyllized. The gangliosides were quantified by measuring the ganglioside sialic acid, according the method of Svennerholm ~5, modified by Miettinen and Takki-Luukkainen 27. Qualitative anal- ysis of the gangliosides was done by HPTLC and by immuno-TLC with mAb Q211, as described earlier in detail 16'3s'37. Briefly, gangliosides were separated by one-dimensional TLC on precoated silicagel HPTLC plates (Merck), using a dual solvent system (1: C/M/12 mM MgCIz/15 N NH4OH = 60/36/6.5/4.5; and 2:C/M/12 mM MgCI 2 = 58/40/9, v/v). After separation, plates were cut into two halves, containing identical samples. One half was sprayed with resorcinol-HCl reagent 4s to visualize gangliosides, and the other half was immunostained with mAb Q211, using a alkaline phosphatase- labelled second antibody.
Quantitative estimation of isolated ganglioside fractions was done by scanning the resorcinol-HCl-stained TLC-plate at 580 nm with a KM3 remission-spectralphotometer (Zeiss).
In vitro assay with exogenous gangliosides and mAb Q211 Isolated and purified gangliosides from l 1-day-old chicken em-
bryos (see above) were dissolved in culture medium, sonicated for 30 rain, sterile filtered, and stored at -20 °C. The isolated mixture contained 2.3% GM1, 10.3% GD3, 17.3% GDla , 5.5% GDlb ,
c 5
~} -~
C 8
21.1% GTlb, 18.5% GQlb/c, 10.5% GPlc, 1.6% G 'H ' , and 12.9% minor fractions. For calculation, an average molecular weight of 2000 Da was assumed. For experiments, ganglioside medium was diluted with fresh DMEM to a final concentration of 50 pM.
Monoclonal antibody Q211 was precipitated from ascites fluid or cell supernatants by adding 40% saturated ammonium sulphate, dialyzed against PBS, sterile filtered and stored at -80 °C. For experiments, Q211 was dissolved in fresh DMEM in final concen- trations from 5 to 15 #g/ml protein. For in vitro assay, cells were seeded in normal culture medium and allowed to adhere to the ground. After 30 min, culture medium was changed against ganglioside- or antibody-containing culture medium, respectively. Thirty min later, the culture medium was supplemented with serum and antibiotics, as described above. After different times in culture, photographs were taken at low magnification, to obtain a repre- sentative image of the whole culture dish.
Quantitative evaluation of neuronal migration The diameters of formed aggregates in living cultures of chicken
optic lobes were measured and used as scale for neuronal migratiom Only cultures of high viability and without detached cells were used, Each value represents mean and standard deviation of 38 up to 621 measured aggregates, taken from 2 to 4 different preparations. The statistically significant difference of the corresponding values was proved by Student's t-test (P ~< 0.001).
RESULTS
D i s s o c i a t e d e m b r y o n i c c h i c k e n n e u r o n s can easi ly be
c u l t u r e d o n p o l y l y s i n e - c o a t e d p las t i c d i shes . W i t h i n a few
m i n u t e s , t h e y a t t a c h to t h e g r o u n d a n d d e v e l o p sho r t
f i lopod ia . A t low s e e d i n g d e n s i t i e s (~<1.106 ce l l s /d ish =
1.105 cel ls /cm2), n e u r o n s r e m a i n i so l a t ed a n d e s t a b l i s h
long, n e u r o n a l p r o c e s s e s 3'14'29. A t h i g h e r cell dens i t i e s
(5.106 cei ls /d ish = 5.105 cel ls /cm2) , h o w e v e r , t he ou t -
g rowing n e u r o n a l p r o c e s s e s c o n t a c t e a c h o t h e r , a n d t he
n e u r o n s b e g i n to m i g r a t e , l e a d i n g to t he f o r m a t i o n of cell
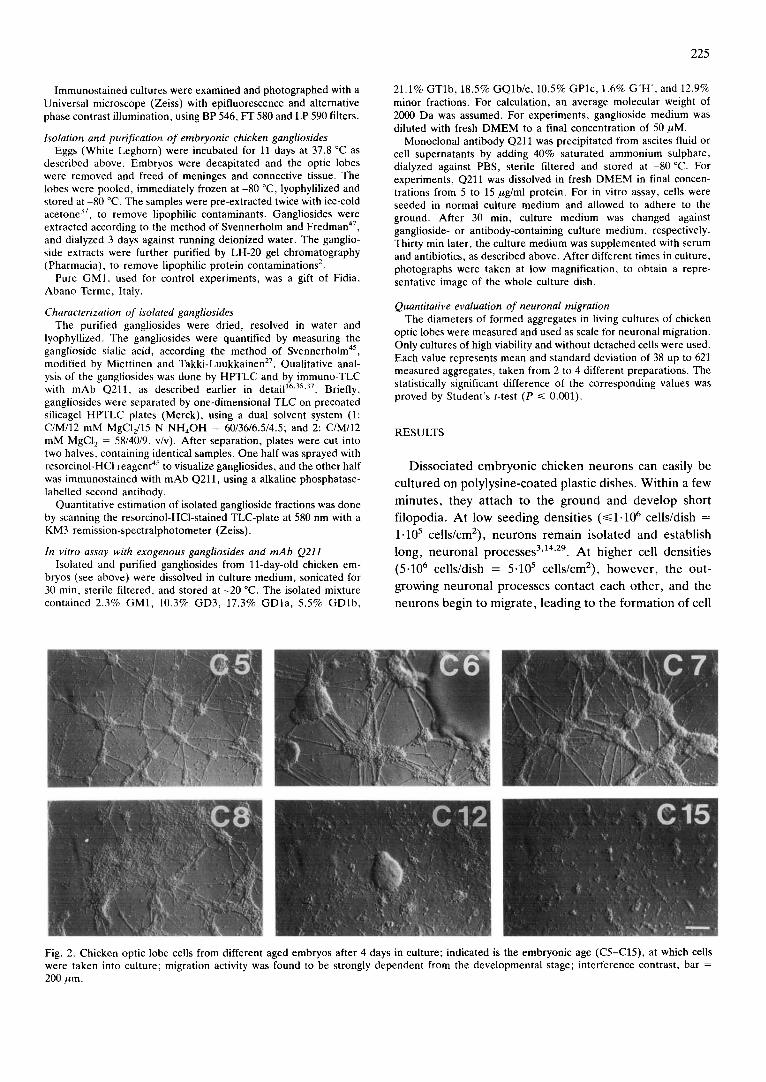
Fig. 2. Chicken optic lobe cells from different aged embryos after 4 days in culture; indicated is the embryonic age (C5-C15), at which cells were taken into culture; migration activity was found to be strongly dependent from the developmental stage; interference contrast, bar = 200 ~m.

226
i
i
l
Fig. 3 .Chicken retina cells from different aged embryos after 4 days in culture; indicated is the embryonic age (C6-C9) , at which cells were taken into culture; neurons formed carpets with 'crater-like' structures, but no 'migrat ion-bundles ' ; interference contrast, bar = 200 ,um.

227
Fig. 4. Morphology of migrating optic lobe neurons in vitro; cells possess a flat, long shaped perikaryon (large arrow) and two processses (small arrows), termed leading and trailing process; resting neurons showed a more rounded shape (open arrow); a: phase contrast; b: Q211-immunofluorescence, bar = 50 rum.
aggregates. Fig. 1 shows the typical development of a culture of chicken optic lobe cells, isolated from a
7-day-old embryo. After 5 rain, the dissociated cells were attached to the ground and homogenously distributed.
Twelve hours later, many of the cells showed an obviously neuronal morphology, with bipolar cell bodies
and short filopodial processes. Staining with neuron- specific mAb Q211 confirmed the neuronal character of the outgrowing cells (see insets). After 1 day in culture, the first accumulations of cells could be seen, being connected during the next 2 days by straight bundles, which were used as migration routes. On these routes, cells migrated in both directions, changing from one aggregation center to another, predominantly from the
smaller to the greater ones, as could be clearly seen by
means of time-lapse film recordings. This led within 4
days to the formation of great aggregates, which were connected by thick bundles with migrating neurons. This time course of migration and aggregation behaviour was highly reproducible, but greatly dependent on the seed-
ing density, the age of the embryo, and the neuronal organ from which the cells were taken.
Fig. 2 shows comparable cultures of optic lobe cells from 5- to 15-day-old chicken embryos (C5-C15), after 4 days in culture, demonstrating the dependency of neu- ronal migration and aggregation intensity from the embryonic age. Maximal migration activity was found with cells from 6- to 7-day-old embryos, whereas prep-

228
Fig. 5. Migration of single optic lobe neurons on a 'migration bundle'; two different neurons (open and closed arrows) migrated in opposite direction; neurons often run short distances with high speed (closed arrow in a, b), then rounded up and stopped for a while (closed arrow in b, c); pictures were taken at intervals of 1 h; phase contrast, bar = 50/~m.
arations from older embryos obviously possessed a greater number of non-migrating cells. This was plainly visible in the areas between the aggregates, which were completely free of cells in C6 cultures, but were covered with great areas of cell monolayers in C12 cultures, although few aggregates could still be found there. In C15 cultures, only some small, local aggregates arised, with- out any connecting migration routes.
Fig. 3 shows similar cultures from retinas of differently aged embryos (C6-C9), after 4 days in culture. Like optic
GM1 GD3
GDla GDla
GD2 GDlb GDlb G Tlb G Tlb GQIb
GQle
GPIo
G'I-I"
1 2 3
Fig. 6. Thin-layer chromatographic separation of a ganglioside mixture from Cll chicken optic lobes (lanes 2 and 3), in comparison with co-chromatographed standards from bovine brain (lane 1), stained with resorcinol-HCl reagent (lanes 1 and 2) or immuno- stained with mAb Q211, specific for polysialogangliosides (lane 3).
lobe cells, retina cells showed a well-developed migration
behaviour. However , they did not form any bundles, which connect the aggregation centers, serving as migra-
tion routes. Instead, the cells migrated directly on the
polylysine-coated bot tom, forming local monolayers. The aggregates, formed by retina cells, often show a 3- dimensional morphology with a 'crater-like' structure, which were never seen in cultures of optic lobe cells.
After some days, when migration was terminated, pro- nounced neuronal sprouting occurred, leading to a dense
neuronal network around the aggregates (Fig. 3c). As in
the case of optic lobe cells, migration and aggregation intensity was age-dependent.
In contrast to retina cells, which migrated directly on
the polylysine surface, optic lobe cells were migrating mainly in contact to the straight bundles between the
aggregates. Fig. 4 demonstrates the morphology of such a migrating cell (large arrow), which possessed a flat- tened, bipolar soma with an ovoid nucleus and 2 cellular processes (small arrows). Immunostaining with neuron- specific m A b Q21133"36'37, directed against membrane-
bound polysialogangliosides (Fig. 4b), confirms the bi- polar morphology of the neurons, migrating in close contact to the surface of the fibre bundle. The time course of neuronal migration was studied by time-lapse film and video recordings of living cultures. Fig. 5 demonstrates the migration of 2 neurons (arrows) along

I d
229
3d
c o n t r o l 50 laM GMi x
c o n t r o l 50 ~M GMi x
Fig. 7. Influence of exogenous gangliosides (GMix) on the migration of C7 optic lobe neurons; gangliosides were isolated from Cll chicken optic lobes and added to a final concentration of 50pM; a marked acceleration of neuronal migration and aggregation in comparison to control cultures can be seen after 1 and 3 days in culture; interference contrast, bar = 200 pm.
a common bundle. In this case, the 2 neurons migrated in opposite direction, indicating the absence of direction- specifying factors on the bundle. Migrating cells pos- sessed an elongated shaped cell body (open arrows in Fig. 5), which changed into a more rounded cell body, when cells became resting (closed arrow in Fig. 5c).
Effect of polysialogangliosides and mAb Q211 on neuro- nal migration and aggregation
Since outgrowing chicken neurons highly express c-pathway polysialogangliosides in vivo and in vitro (Figs. 4b and 6), we used aggregating C7 optic lobe cultures to elucidate a possible involvement of these surface mole- cules. For these experiments, gangliosides isolated from optic lobes of 11 days incubated embryonic chicken were used, possessing the same composition as the endogenous gangliosides in this developmental stage. Fig. 6 demon- strates a thin-layer chromatographic separation of the purified ganglioside mixture (lane 2), in comparison with a co-chromatographed standard (lane 1). The mixture
contained a high proportion of Q211-binding polysialo- gangliosides (>30%), as indicated by Q211-immuno- staining (lane 3). Fig. 7 shows typical cultures, grown with or without addition of the ganglioside mixture
(GMix, 50 pM), respectively, Ganglioside-supplemented cultures (right) showed a markedly increased migration compared to control cultures (left) at the same culture time. After 1 day in culture, in the presence of GMi x aggregates had formed, which were comparable to those of 3-day-old controls. After 3 days, almost all ganglio- side-treated cells were gathered in a few large aggregates. One day later, these large aggregates usually rounded off and detached from the ground. However, most of these neurons were still viable, because after renewed dissoci- ation and seeding they attached again to the ground and started to form neuronal processes (not shown here).
In contrast, addition of mAb Q211 (10 pg/ml) to the cultures reduced neuronal migration and aggregation intensity (Fig. 8). Even after 4 days in culture, aggregates appeared more diffuse and a much higher number of

230
lct
c o n t r o l 10 B g / m l Q211
c o n t r o l 10 j a g / m l Q21l
Fig. 8. Influence of monoclonal antibody Q211 on the migration of C7 optic lobe neurons; Q211 was added to a final concentration of 10 #g/ml; a reduction of neuronal migration and aggregation in comparison to control cultures can be seen, especially after 4 days in culture; note the fine distributed network of single neuronal processes in the Q211 treated culture (4d); interference contrast, bar = 2(~)~m.
non-migrating cells could be found in the spaces between the aggregates. The development of neuronal processes was not reduced, but the fasciculation of processes to bundles was markedly decreased, leading to a fine fibre network (Figs. 8 and 9). This aggregation- and fascicu- lation-inhibiting effect of the Q211 antibody appeared only in a critical concentration (Fig. 9). At 5/~g/ml Q211 had no effect (Fig. 9b), while at 10/~g/ml fasciculation and aggregation was clearly diminished (Fig. 9c). At 15 ~g/ml, there was only negligible neuronal sprouting and no fasciculation and migration of neurons occurred (Fig. 9d). However, the cells were still viable and adhered to the ground.
Quantitative estimation of the aggregate size (average diameter) of optic lobe cultures (Fig. 10) confirmed these observations and revealed a significant pronouncement (P ~< 0.001) of neuronal aggregation in ganglioside (50 /~M) treated (+92.7% after 3 days) and a significant reduction in Q211 (10/~g/ml) treated cultures (-33.0% after 3 days). Cultures treated with 50/~M pure GM1
showed also an enhanced neuronal aggregation, but the increase in the average aggregate size (+54.9%) was only half as much as in the case of embryonic chicken ganglioside mixture.
DISCUSSION
High-density primary cultures of embryonic chicken neurons turned out as a highly reproducible model, to study neuronal migration and aggregation. In accordance with previous findings z9, the polylysine-coated surface was found to permit sufficient adhesion and survival of the neurons, while the growth of dividing, non-neuronal cells was rather suppressed. While in sparse cultures most neurons remain dispersed, forming neuronal processes under nearly all conditions allowing appropriate adhesion to the ground, in our dense cultures, the neurons form fasciculated bundles, which are used as migration routes. Most of the neurons then lose their contact to the artificial substrate and aggregate to large cell clusters.

- ,~ ..... ~,,. ....
t ~ . . . . p
231
control 5 pg/ml Q211
• ~"~,,
:k ?
10 pg/ml Q211 15 ~g/ml Q211 Fig. 9. Dose-dependent reduction of neuronal migration and aggregation of C7 optic lobe cells by mAb Q211, after 3 days in culture; at a final concentration of 5 #g/ml no obvious effect is visible; at 10 ~g/ml aggregates were smaller, more cells remained between the aggregates and a fine network of single neuronal processes is formed; at 15 #g/ml aggregation is completely inhibited, although most of the cells were still alive and adhered to the ground; interference contrast, bar = 100 :tm.
In the case of optic lobe neurons particular long
migration routes were formed, connecting rounded ag- gregation centers (Fig. 1). In vivo, such migration routes are formed by neuronal pioneer fibres 2~ and/or by radial glial fibres 43, and the migration of neurons is restricted to
a distinct developmental period. A comparable age- dependent restriction of neuronal migration was also found in vitro (Fig. 2), suggesting that the ability to
migrate is transiently regulated by the neuronal genetic
machinery, as well as dependent on permissive environ- mental structures. A similar temporal restriction of cell migration had been found by Trenkner et al. 49 in cultures
of cerebellar granule cells, for which the migration period of a single cell lasted about 4 days.
In the case of retinal cultures, cells were also observed to migrate, but without forming any fibre bundles. They rather moved directly on the polylysine-coated dish,
forming large monolayer carpets and later some 3- dimensional , 'crater-like' structures (Fig. 3). This indi-
average size of aggregates in C7 optic lobe cultures
[~m} 120
110
100
90
80
70
60
50
40
30
20
10
0 1 2 3
GMix Control
Q211
J 4
da~s hi (-~[turc
Fig. 10. Quantitative evaluation of aggregate size (average diame- ter) of C7 optic lobe cultures grown with 50 /~M embryonic ganglioside mixture (GMix) or 10 /~g/ml mAb Q211 (Q211); each point represents average values and standard deviation of cultures from 2 to 4 separate experiments; in ganglioside supplemented cultures, after 4 days aggregates usually had already detached from the ground•

232
cates that the neuronal migration and aggregation be- havior is strongly dependent on cell types, present in the culture, and is not induced by the artificial culture conditions, which were the same in both cases. Migrating optic lobe cells in vitro showed a flat, long shaped perikaryon with a leading and a trailing process, which were in close contact with a fibre bundle (Fig. 4). This morphology looks exactly like that of migrating neurons in vivo, as found in chicken optic lobes 8'9'31 or monkey cerebellum 32.
The migration routes, formed by optic lobe neurons in vitro, consisted of fasciculated processes. An intense staining of the bundles with neuron-specific mAb Q21133. 36 suggests that most if not all of them were neuronal processes. In addition, all migrating cells and aggregates were stained by mAb Q211 (Fig. 4b). The neuronal character of the migrating cells and the majority of the processes contacted by them was further confirmed by immunostaining with another neuron-specific antibody, directed against intracellular neurofilaments 7 (not shown). From these data it can be concluded, that most neurons migrate directly on the surface of other neurons and thus belong to the neurophilic type 32. It is important to note, that on a given bundle neurons migrate in both directions (Fig. 5), and single neurons sometimes stop migration and move then to the opposite direction. Thus, along the bundles no direction-determining surface in- formation seems to exist. In vivo as in vitro, the expression of c-pathway polysialogangliosides by embry- onic chicken neurons is developmentally regulated and restricted to the period of neuronal migration and fibre outgrowth 15'34"36. The functional importance of these
highly negative charged glycosphingolipids for neuronal migration is supported by the presented data, demon- strating an acceleration in the presence of exogenous embryonic chicken gangliosides (Fig. 7). Exogenous added gangliosides have been shown to be in part properly incorporated into the plasma membrane and act like the endogenous ones 13'41'5°. The migration and
aggregation promoting effect of the exogenous ganglio- sides seems not to be a simple consequence of complex- ing free Ca 2+3°, because addition of equivalent amounts of EGTA did not produce similar effects, but rather fascilitated detachment of the cells (data not shown).
We used a ganglioside mixture, which had been isolated from embryonic chicken optic lobes of the same developmental stage as used for cultured neurons, with the same pattern as the endogenous gangliosides. Incor- poration of this ganglioside mixture therefore will only raise the ganglioside content in the membrane of the cells, without changing the ganglioside composition. The used mixture had a high content of c-pathway polysialo- gangliosides, which first expression in vivo has been
shown to coincide strictly with neurogenesis -~4. The special importance of these polysialogangliosides for neuronal migration is further emphasized by a dose- dependent reduction of migration and aggregation in cultures grown in the presence of the specific monoclonal antibody Q211 (Figs. 8 and 9), binding selectively to gangliotetraoses of the c-pathway polysialogangliosides ~6. Initial sprouting and growth per se of the neurons was not inhibited by Q211-treatment, but the single neuronal processes formed a fine distributed network and failed to fasciculate to well-ordered migration routes. The diverg- ing growth resembled more dendritic arborisation of neurons after reaching their target field
Both effects, acceleration of neuronal fasciculation, migration and aggregation of added embryonic chicken brain gangliosides and reduction of this growth behaviour by 'masking' c-polysialogangliosides with mAb Q211, could be quantitatively confirmed in several preparations and were in spite of high standard deviations between different series highly significant, as proved by the Student's t-test (P ~< 0.001).
The opposite effects of c-polysialoganglioside contain- ing chicken brain extract and of mAb Q211 on neuronal migration and aggregation in vitro together with the developmental restriction of polysialoganglioside expres- sion to the period of migration and fibre outgrowth in vivo suggest, that the observed aggregation-stimulating effect of the ganglioside extract is specific to the c-polysialogangliosides, although other gangliosides were present in this extract too. This idea is supported by the much lower stimulating effect of equivalent amounts of pure GM1, which can probably be explained by a conversion of inserted GM1 by cellular sialyltransferases, which are highly active in this developmental stage. Such a conversion could take place either in the cell membrane or after internalization during the normal turnover process in the Golgi apparatus.
Our results are in accordance with previous findings of Spoerri 44, who also found a stimulation of neuronal clustering in primary cultures of embryonic chicken cerebrum after addition of exogenous gangliosides. The inhibitory effect of the Q211 antibody is interestingly similar to the effect of antibodies against cell adhesion molecules (CAMs). Monoclonal antibodies to neural CAM (N-CAM) had been shown to inhibit fasciculation and aggregation of chicken embryonic neurons in vitro and in vivo, and led to formation of a fine neuronal network 3s'39'48. Other monoclonal and polyclonal anti-
bodies to L1 antigen (which corresponds to the neuron- glia CAM (Ng-CAM) of embryonic chicken) and to AMOG (adhesion molecule on glia) inhibit granule cell migration in slices of mouse cerebellar cortex x'26.
In the case of N-CAM, the adhesion property of the

m o l e c u l e can be di rec t ly co r re l a t ed to the n u m b e r of
sialic acid mo lecu l e s 4°, which dec rease dur ing deve lop -
m e n t 5. In te res t ing ly ' n e w - b o r n ' neu rons express also
highly s ia lyla ted polys ia logangl ios ides which are r ep laced
by less s ia lyla ted gangl ios ides dur ing synaptogenes i s , i .e.
a f ter the p e r i o d o f mig ra t ion and initial neur i togenes i s 33'
34. T h e da ta p r e s e n t e d in this p a p e r suggest that Q211
b ind ing po lys ia logangl ios ides are in addi t ion to cell
adhes ion p ro te ins i m p o r t a n t for ce l l - ce l l and /or c e l l -
REFERENCES
1 Antonicek, H., Person, E. and Schachner, M., Biochemical and functional characterization of a novel neuron-glia adhesion molecule that is involved in neuronal migration, J. Cell Biol., 104 (1987) 1587-1595.
2 Byrne, M.C., Sbaschnig-Agler, M., Aquino, D.A., Scalafani, J.R. and Ledeen ,R.W., Procedure for isolation of gangliosides in high yield and purity: simultaneous isolation of neutral glycosphingolipids, Anal. Biochem., 148 (1985) 163-173.
3 Collins, F., Axon initiation by ciliary neurons in culture, Dev. Biol., 65 (1978) 50-57.
4 Cowan, W.M., A synoptic view of the development of the vertebrate central nervous system. In J.G. Nicholls (Ed.), Repair and Regeneration of the Nervous System, Springer, Berlin, 1982, pp. 7-24.
5 McCoy, R.D., Vimr, E.R. and Troy, EA. , CMP-NeuNAc:Poly- a-2,8-sialosyl sialyltransferase and the biosynthesis of polysia- losyl units in neural cell adhesion molecules, J. Biol. Chem., 260 (1985) 12695-12699.
6 Curatolo, W., Glycolipid function, Biochim. Biophys Acta, 906 (1987) 137-160.
7 Doherty, P., Dickson, J.G., Flanigan, T.P. and Walsh, F.S., Quantitiative evaluation of neurite outgrowth in cultures of human foetal brain and dorsal root ganglion cells using an enzyme-linked immunoadsorbent assay for human neurofila- ment protein, J. Neurochem., 42 (1984) 1116-1122.
8 Domesick, V.B. and Morest, D.K., Migration and differentia- tion of ganglion cells in the optic tectum of the chick embryo, Neuroscience, 2 (1977) 459-475.
9 Domesick, V.B. and Morest, D.K., Migration and differentia- tion of shepherd's crook cells in the optic tectum of the chick embryo, Neuroscience, 2 (1977) 477-491.
10 Dreyfus, H., Urban, P.E, EdeI-Harth, S. and Mandel, P., Developmental patterns of gangliosides and of phospholipids in chick retina and brain, J. Neurochem., 25 (1975) 245-250.
11 Edelma'n, G.M., Modulation of cell adhesion during induction, histogenesis and perinatal development of the nervous system, Annu. Rev. Neurosci., 7 (1984) 339-377.
12 Edelman, G.M. and Thiery, J.P., General introduction. In G.M. Edelman and J.P. Thiery (Eds.), The Cell in Contact; Adhesions and Junctions as Morphogenetic Determinants, Wiley, New York, 1985, pp. 3-7.
13 Fishman, P.H., Recent advances in identifying the functions of gangliosides, Chem. Phys. Lipids, 42 (1986) 137-151.
14 Greis, Ch., R6sner, H., Rodemann, H.P., Henke-Fahle, S. and Hirabayashi, Y., Expression of ganglioside-antigens by fibro- blasts and primary neuron cultures. In N. Eisner and EG. Barth (Eds.), Sense Organs, Interfaces Between Environment and Behaviour, Thieme, Stuttgart, 1988, p. 311.
15 Greis, Ch. and R6sner, H., Possible involvement of polysialo- gangliosides in neuronal sprouting, migration and aggregation in vitro. In N. Eisner and W. Singer (Eds.), Dynamics and Plasticity in Neuronal System, Thieme, Stuttgart, 1989, p. 220.
16 Greis, Ch. and R6sner, H., C-pathway-polysialogangliosides in the nervous tissue of vertebrates, reacting with the monoclonal antibody Q211, Brain Res., 517 (1990) 105-110.
233
ex t race l lu la r mat r ix in te rac t ions , unde r l ay ing neu rona l
migra t ion and neur i t e fasc icula t ion. If these polysia lo-
gangl ios ides s imply act as r ecep to r s for e m b r y o n i c N-
C A M , as specu la ted by J0 rgensen ~2, o r as part of a
comp lex mach ine ry in addi t ion to o r in c o m b i n a t i o n with
m e m b r a n e - b o u n d pro te ins , has to be clarif ied.
Acknowledgements. We wish to thank the Fidia Research Labs, Abano Terme, Italy for supporting this project.
17 Hakomori, S.I., Ganglioside-mediated modulation of growth factor receptor function and cell adhesion. In H. Rahmann (Ed.), Gangliosides and Modulation of Neuronal Functions; Nato ASI Series H7, Springer, Berlin, 1987, pp. 465-479.
18 Hakomori, S.I., Glycosphingolipids in cellular interaction, dif- ferentiation, and oncogenesis, Annu. Rev. Biochem., 50 (1981) 733-764.
19 Hamburger, V. and Hamilton, H.L., A series of normal stages in the development of the chick embryo, J. Morphol., 88 (1951) 49-92.
20 Henke-Fahle, S., Monoclonal antibodies recognize gangliosides in the chick brain, Neurosci. Lett., 14 (1983) 160.
21 Hicks, S.P. and D'Amato, C.J., Cell migrations to the isocortex in the rat, Anat. Rec., 160 (1968) 619-634.
22 JOrgensen, O.S., The embryonic form of D2-protein (N-CAM) is linked to A2B5-binding polysialoganglioside-like carbohy- drates, J. Neurochem., 44 (1985) 33.
23 Landa, C.A. and Moscona, A.A., Changes in ganglioside profile in chick embryo retina: studies on tissue and cell cultures, Int. J. Dev. Neurosci., 3 (1985) 77-78.
24 Lander, A.D., Understanding the molecules of neural cell contacts: emerging patterns of structure and function, Trends Neurosci., 12 (1989) 189-195.
25 Ledeen, R.W., Biology of gangliosides: neuritogenic and neu- ronotrophic properties, J. Neurosci. Res., 12 (1984) 147-159.
26 Lindner, J., Rathjen, EG. and Schachner, M., L1 mono- and polyclonal antibodies modify cell migration in early postnatal mouse cerebellum, Nature, 305 (1983) 427-430.
27 Miettinen, T. and Takki-Luukkainen, I.T., Use of butylacetate in determination of sialic acid, Acta Chem. Scand., 13 (1959) 856-858.
28 Nagai, Y. and Tsuji, S., Cell biological significance of ganglio- sides in neural differentiation and development: critique and proposals. In R.W. Ledeen, E.L. Hogan, G. Tettamanti, A.J. Yates and R.K. Yu (Eds.), New Trends in Ganglioside Research, Springer, Berlin, 1988, pp. 329-350.
29 Pettman, B., Louis, J.C. and Sensenbrenner, M., Morphological and biochemical maturation of neurons cultured in the absence of glial cells, Nature, 281 (1979) 378-380.
30 Probst, W., R6sner, H., Wiegandt, H. and Rahmann, H., Das KomplexationsvermOgen yon Gangliosiden fiir Ca 2. . I. Einfluss mono- and divalenter Kationen sowie yon Acetylcholin, Hoppe- Seyler's Z. Physiol. Chem., 360 (1979) 979-986.
31 Puelles, L. and Bendala, M.C., Differentiation of neuroblasts in the chick optic rectum up to eight days of incubation: a Golgi study, Neuroscience, 3 (1978) 307-325.
32 Rakic, P., Contact regulation of neuronal migration. In G.M. Edelman and J.P. Thiery (Eds.), The Cell in Contact, Wiley, New York, 1985, pp. 67-91.
33 Rohrer, H., Henke-Fahle, S., El-Sharkawy, T., Lux, H.D. and Thoenen, H.C., Progenitor cells from embryonic chick dorsal root ganglia differentiate in vitro to neurons: biochemical and electrophysiological evidence, EMBO J., 4 (1985) 1709-1714.
34 R6sner, H., Ganglioside changes in the chicken optic lobes as biochemical indicators of brain development and maturation, Dev. Brain Res., 236 (1982) 49-61.
35 ROsner, H., A new thin-layer chromatographic approach for

234
separation of multisialogangliosides. Novel ganglioside fractions in the embryonic chicken brain, Anal. Biochem., 109 (1980) 437-442.
36 ROsner, H., AI-Aqtum, M. and Henke-Fahle, S., Developmen- tal expression of GD3 and polysialogangliosides in embryonic chicken nervous tissue reacting with monoclonal antiganglioside antibodies, Dev. Brain Res., 18 (1985) 85-95.
37 R6sner, H., Greis, Ch. and Henke-Fahle, S., Developmental expression in embryonic rat and chicken brain of a polysialo- ganglioside-antigen reacting with the monoclonal antibody Q211, Dev. Brain Res., 42 (1988) 161-171.
38 Rutishauser, U., Gall, W.E. and Edelman, G.M., Adhesion among neural cells of the chick embryo; IV. Role of the cell surface molecule CAM in the formation of neurite bundles in cultures of spinal ganglia, J. Cell Biol., 79 (1978) 382-393.
39 Rutishauser, U., Thiery, J.P., Brackenbury, R. and Edelman, G.M., Adhesion among neural cells of the chick embryo; III. Relationship of the surface molecule CAM to cell adhesion and the development of histiotypic patterns, J. Cell Biol., 79 (1978) 371-381.
40 Rutishauser, U., Watanabe, M., Silver, J., Troy, EA. and Vimr, E.R., Specific alteration of NCAM-mediated cell adhesion by an endoneuraminidase, J. Cell Biol., 101 (1985) 1842-1849.
41 Schwarzmann, G., Hoffmann-Beihauer, P., Schubert, J., Sand- hoff, K. and Marsh, D., Incorporation of gangliosides analogues into fibroblast cell membranes. A spin-label study, Biochemistry, 22 (1983) 5041-5048.
42 Seyfried, T.N., Ganglioside abnormalities associated with failed neural differentiation in a T-locus mutant mouse embryo, Dev.
Biol., 123 (1987) 286-291. 43 Sidman, R.L. and Rakic, P., Neuronal migration with special
reference to developing human brain: a review, Brain Res.. 62 (1973) 1-35.
44 Spoerri, P.E., Facilitated-establishment of contacts and synapses in neuronal cultures: ganglioside-mediated neurite sprouting and outgrowth. In G. Tettamanti, R.W. Ledeen, K. Sandhoff, Y. Nagai and G. Toffano (Eds.), Gangliosides and Neuronal Plasticity, Liviana Press, Padova, 1986, pp. 3[)9-325.
45 Svennerholm, L., Quantitative estimation of sialic acids. A colorimetric resorcinol-hydrochloric acid method, Biochim. Bio- phys. Acta, 24 (1957) 604-611.
46 Svennerholm, L., Chromatographic separation of human brain gangliosides, J. Neurochem., 10 (1963) 613-623.
47 Svennerholm, L. and Fredman, P., A procedure for the quantitative isolation of brain gangliosides, Bioehim. Biophys. Acta, 617 (1980) 97-109.
48 Thanos, S., Bonhoeffer, F. and Rutishauser, U., Fiber-fiber interaction and tectal cues influence the development of the chicken retinotectal projection, Proc. Natl. Acad. Sci. U.S.A., 81 (1984) 1906-1910.
49 Trenkner, E., Smith, D. and Segil, N., Is cerebellar granule cell migration regulated by an internal clock?, J. Neurosci., 4 (1984) 2850-2855.
50 Varon, S., Pettmann, B. and Manthorpe, M., Extrinsic regula- tions of neuronal maintenance and repair. In R.W. Ledeen, E.L. Hogan, G. Tettamanti, A.J. Yates and R.K. Yu (Eds.), New Trends in Ganglioside Research, Springer, Berlin, 1988, pp. 607-623.

















