MicroRNA-29 expression in various cancer types · Page 1 of 36 MicroRNA-29 expression in various...
36
Page 1 of 36 MicroRNA-29 expression in various cancer types 5.semester, efterår 2011 Studieretning: Molekylærbiologi og Medicinalbiologi Af Mette Neiegaard Witt & Stine Herold Madsen Vejleder: Annika Bagge
Transcript of MicroRNA-29 expression in various cancer types · Page 1 of 36 MicroRNA-29 expression in various...

Page 1 of 36
MicroRNA-29 expression in
various cancer types
5.semester, efterår 2011
Studieretning: Molekylærbiologi og Medicinalbiologi
Af Mette Neiegaard Witt & Stine Herold Madsen
Vejleder: Annika Bagge

Page 2 of 36
Index Abstract ...................................................................................................................................... 4
Resume ....................................................................................................................................... 4
Introduction ................................................................................................................................ 5
Problem statement .................................................................................................................. 5
MicroRNAs ............................................................................................................................ 6
MicroRNA biogenesis ........................................................................................................ 6
Insulin-signaling pathway and type 2 diabetes mellitus......................................................... 8
The role of miR-29 in type 2 diabetes mellitus .................................................................... 10
The correlation of glucose concentrations and miR-29 .................................................... 12
miR-29 and the insulin-signaling pathway ....................................................................... 14
Cancer and the cell cycle ...................................................................................................... 14
Cyclins and cyclin dependent kinases .............................................................................. 15
Analysis.................................................................................................................................... 16
miR-29 and cancer in insulin-responsive tissues ................................................................. 16
Muscle, rhabdomyosarcoma ............................................................................................. 19
Pancreas, endocrine tumors .............................................................................................. 20
miR-29 and cancer in non-insulin-responsive tissues .......................................................... 20
Lymphocytes, B-cell chronic lymphocytic leukemia ....................................................... 20
Lymphocytes, AML.......................................................................................................... 21
Bile duct, cholangiocarcinoma ......................................................................................... 23
Is miR-29 a tumor suppressor, an oncomiR or both? ........................................................... 24
Summative discussion and conclusion..................................................................................... 27
The connection between miR-29a, 29b1, 29b2 and 29c. ..................................................... 27
Using miRNA in cancer treatment ....................................................................................... 27
Models vs. the human body.................................................................................................. 28
The role of miR-29 in cancer development .......................................................................... 29

Page 3 of 36
Perspective ............................................................................................................................... 29
Refferences .............................................................................................................................. 30
Appendix 1 ............................................................................................................................... 36

Page 4 of 36
Abstract
The recent discovery of microRNA’s has presented a new pool of regulators of diverse
pathways of the human body. It is suggested that miR-29a has negative effects on some of the
steps in the insulin-signaling pathway. This indicates that clinical regulation of miR-29a
could be part of type 2 diabetes mellitus treatment. With every clinical regulation of
pathways of the body risks of undesirable consequences may follow. These consequences
could be numerous but cancer is a frequent result of pathways being deregulated, and
therefore we conducted a literature study of the miR-29 family and its expression levels in
various cancer types. Several experiments showed that all three miR-29 paralogs were
expressed in increased or decreased levels in different cancer types compared to the
corresponding normal tissue. The studies showed that miR-29 was down-regulated in brain,
lung, liver, cervix, muscle and stomach cancers, and up-regulated in cholangiocytic, prostate,
pancreatic and lymphocytic cancers with possible exceptions. Thus no overall tendency of an
up- or a down-regulation of miR-29 in cancer was found. But it seems that that miR-29 do
play a role in cancer development though the role differs between cancer types. miR-29 could
thereby possibly be a part of cancer treatment if it can be regulated tissue specifically.
Resume
Den nye opdagelse af microRNA’er har frembragt en ny pulje af regulatorer i
menneskekroppens mange signalveje. Det er blevet foreslået, at miR-29a har en negativ
effekt på nogle af trinene i insulinsignalvejen. Dette indikerer, at en klinisk regulering af
miR-29a kunne blive en del af type 2 diabetes mellitus behandling. Med enhver klinisk
regulering af signalveje i kroppen følger risici for uønskede konsekvenser. Disse
konsekvenser kan være mange, men kræft er et typisk resultat af dysregulerede signalveje, og
derfor laver vi her et litteraturstudie over miR-29 familien og dens ekspression i forskellige
kræfttyper. Flere eksperimenter viste at alle tre miR-29 paraloger blev udryk i øgede eller
mindskede niveauer i forskellige kræfttyper sammenlignet med det korresponderende
normale væv. Studierne viste, at miR-29 var nedreguleret i hjerne-, lunge-, lever-,
livmoderhals-, muskel- og mavekræft og opreguleret i cholangiocyt-, prostata-, bugspytkirtel-
og lymfocyt-kræft med mulige undtagelser. Der er altså ingen overordnet op- eller
nedreguleringstendens af miR-29 i kræft. Men det ser ud til, at miR-20 alligevel spiller en
rolle i kræftudvikling selvom rollen varierer fra kræfttype til kræfttype. miR-29 kunne
dermed være en del af kræftbehandling, hvis den kan reguleres vævsspecifikt.

Page 5 of 36
Introduction
MicroRNAs (miRNAs) are a recently discovered pool of regulators in vertebrates,
invertebrates and plants (Laegos-Quintana et al., 2001; Lee, 1993; Chen, 2005). It has been
brought to our attention that the miRNA miR-29 might have a negative regulatory effect on
the insulin signalling pathway. This indicates a possibility of utilizing down-regulation of
miR-29 in diabetes mellitus type 2 (T2DM) treatments. Therefore we started out by
investigating literature regarding miR-29, the insulin signalling pathway and the connection
between miR-29 and T2DM. Even though a down-regulation of miR-29 might help people
suffering from T2DM, clinical regulation of pathways in the body could have undesired
effects. It is therefore important to consider the possible consequences of interfering with the
pathways before doing so. The consequences could have great diversity but we have chosen
to focus on the coherence of cancer and the expression of miR-29 because this field of
investigation is gaining more and more ground.
Problem statement
• Do expression levels of miR-29 play a role in cancer development?
Investigation of the expression levels of miR-29 in various cancers and the possibility of
using regulation of miR-29 in cancer treatment

Page 6 of 36
MicroRNAs
MicroRNAs (miRNAs) are single stranded non-coding RNA’s that do not translate into
proteins like regular RNAs do. They function either as a silencing molecule by suppressing
the translation of a target-mRNA, or as a direct degrader of a target-mRNA (Kolfschoten,
2009). The most important sequence for miRNA target binding is called the seed region and
is located between nucleotides 2-8 in the
5’end of the miRNA (figure 1).
Complementarities between the seed
sequence and the target sequence is required
for the miRNA to bind to the mRNA
(Lynam-Lennon, 2008). miRNAs are
divided into families according to identical
seed sequences (Ambros, 2003). The miR-
29 family is comprised by three paralogs; miR-29a, 29b and 29c. miR-29a and miR-29b1 are
located on chromosome 7q32 whereas miR-29b2 and miR-29c are located on chromosome
1q23 (Garzon, 2009). miR-29b1 and miR-29b2 sequences are identical but they are
distinguished as b1 and b2 due to the difference in locus. Some miRNAs are tissue specific
(Lagos-Quintiana, 2002) but miR-29 is present in many different tissues (Xiong et al., 2010;
Roldo et al., 2006; Zhao, 2010; Mott et al., 2007).
MicroRNA biogenesis
MicroRNA biogenesis describes the process of miRNA maturation. The first step in the
biogenesis is transcription of the gene encoding the miRNA. miRNAs are encoded in the
genome and are transcribed by RNA Polymerase II and the primary transcript is called pri-
miRNAs (Lynam-Lennon, 2009). Pri-miRNAs have a characteristic shape, with one or
several stem-loop structures (figure 2). They have a 5’-cap and a polyadenylated 3’ end like
mRNAs and they consist of a couple of thousands nucleotides (Kolfschoten et al., 2009). The
pri-miRNA is processed by a microprocessor complex consisting of two molecules; the
RNAse Drosha and the protein DGCR8. The Drosha-DGCR8-complex reduces the pri-
miRNA to a pre-miRNA of around 80 nucleotides, by excising the hairpins from the long pri-
miRNA. The pre-miRNA is translocated out of the nucleus and into the cytoplasm by the
transport protein exportin-5. A complex consisting of three units: an RNAse named Dicer, a
Figure 1: The sequences of the microRNA-29 family Families of miRNAs have very similar base sequences and identical seed sequences. The seed sequence is located between nucleotide 2 and 8 (Modified from He et al., 2007).

Page 7 of 36
RNA-binding protein and an endonuclease named Argonaut-2 (Ago-2) cleaves the loop from
the stem yielding a double stranded miRNA in the cytoplasm (Lynam-Lennon, 2009). The
duplex is now unwound by RNA helicase leaving only the mature miRNA consisting of 21-
23 nucleotides and Ago-2. The unwinding yields two strands, and either of them could be the
future mature miRNA, but the strand with the most stable 5’ end is degraded (Winter, 2011).
The mature miRNA is incorporated into the RNA-induced silencing complex (RISC) and the
miRNA guides the RISC complex to the miRNA target-sequence located in the 3’UTR of
their mRNA target (Lynam-Lennon, 2009; Winter, 2011). Binding of the miRNA-RISC
complex to the mRNA target leads to either degradation or translational repression of the
mRNA (Winter, 2011).
Figure 2: microRNA biogenesis The microRNA primary transcript (pri-miRNA) is processed by Drosha/DGCR8 yielding precursor miRNA (pre-miRNA) with a characteristic stem-loop structure. Exportin-5 translocates pre-miRNA from the nucleus into the cytoplasm. Dicer complex cleaves of the loop yielding to complementary strands. RNA helicase unwinds the double strand. The unwinding yields two strands, and either of them could be the future mature miRNA, but the strand with the most stable 5’ end is degraded. The single-stranded mature miRNA is incorporated into the RISC complex, followed by binding of the miRNA to the target mRNA. (Modified from http://www.bioscience.org/2009/v14/af/3412/fulltext.asp?bframe=figures.htm&doi=yes).

Page 8 of 36
Because the miRNA most often is only partially complementary to their targets, the number
of potential targets is abundant (Shreenivasaiah, 2010). The field of miRNA-research is rather
new (Ambros, 2003), but several studies indicate that miRNAs are involved in regulating
many different pathways (Pekarsky, 2006; Garret, 2001; Wang, 2008; Garzon, 2009; Park et
al., 2009). Focus in this report is primarily on miR-29 and the correlation between miR-29
expression and cancer. Secondary focus is on the insulin signalling pathway and the
correlation of T2DM and miR-29, from where our motivation came.
Insulin-signaling pathway and type 2 diabetes mellitus
The body and in particular the brain and nervous system needs a constant supply of glucose
for optimal functioning. To maintain a constant glucose level in the blood, β-cells in the islets
of Langerhans in the pancreas produce and secrete the hormone insulin in response to
elevated glucose levels that follow food intake (Seeley, 2011; Hancock, 2010; Madsbad,
2007; Dirice, 2011). Insulin increases the uptake of glucose from the blood primarily into
muscle and adipose tissue by binding to the α-subunit of insulin receptors (IRs) on the surface
of the tissue cells. Binding to the α-subunit induces intracellular phosphorylation of the
transmembrane β-subunit, causing autophosphorylation of IR. These events lead to
translocation of intracellular vesicles containing the transmembrane glucose-transport protein
GLUT4 (figure 3). GLUT4 travels to the surface of the cell where it fuses with the plasma
membrane (Hancock, 2010; Skelly, 2006; Le Roith et al., 2003; Seeley, 2011).
Figure 3: The effect of insulin secretion on GLUT4 Insulin binds to the insulin receptor (IR) on the cell surface of muscle- and adipose tissue and induces a signaling pathway ultimately leading to translocation of glucose-transport protein GLUT4 from vesicles inside the cell to the cell membrane with which it fuses (modified from Hancock, 2010)

Page 9 of 36
The insulin-signaling pathway consists of many steps in between binding of insulin to IR and
translocation of GLUT4. miR-29 seems to regulate some of the steps and they are therefore
described in more details here and presented in figure 4. The binding of insulin to IR leads to
activation of insulin receptor substrates (IRS) which activates PI3-K. PI3-K has a regulatory
subunit called p85 and a catalytic subunit called p110 (Karlsson, 2007). When IRS binds to
p85 it leads to activation of p110 which catalyses phosphorylation of phosphatidylinositol
(3,4)-biphosphate (PIP2) yielding phosphatidylinositol (3,4,5)-triphosphate (PIP3) (Karlsson,
2007). The serine/threonine protein kinase Akt binds to PIP3 in the membrane and is then
phosphorylated and activated by either phosphoinositide dependent kinase 1 (PDPK1) at
Thr308 or rapamycin complex 2 (mTORC2) at Ser473 (Whitehead et al., 2000; Burén, 2003; Le
Roith et al., 2003). When Akt is activated it stimulates the translocation of GLUT 4 in the cell
(Karlsson, 2007; Whitehead et al., 2000; Lund, 2007).
Figure 4: Translocation of GLUT4 by the insulin signaling pathway The drawing shows some of the steps of the insulin-signalling pathway where binding of insulin ultimately leads to translocation of GLUT4. Insulin binds to insulin receptor (IR) thereby activating insulin receptor substrates (IRS) which binds to the p85 subunit of PI3-K. p85 then activates another subunit of PI3-K; p110, which leads to phosphorylation of phosphatidylinositol (3,4)-biphosphate (PIP2) to phosphatidylinositol (3,4,5)-triphosphate (PIP3). Akt then binds to PIP3 and is thereby phosphorylated by phosphoinositide dependent kinase 1 (PDPK1) or rapamycin complex 2 (mTORC2) and this stimulates the translocation of GLUT4 to the cell surface (own drawing, 2011).

Page 10 of 36
Insulin plays another and just as important role besides increasing glucose uptake. Insulin
inhibits glyconeogenesis and also induces glycogenesis in the liver thereby storing more
glucose as glycogen. Glucose enters the liver through glucose transport protein 2 (GLUT2)
which is not insulin dependent but has a low affinity for glucose. The activity of GLUT2 is
therefore proportional to the concentration of glucose in the blood (Lund, 2007).
T2DM is characterized by β-cell dysfunction and decreased insulin response. Although there
are still disagreements on the field it seems as if the β-cell deterioration happens in early
stages of the disease before the reduction in insulin response (Kahn, 2003). This deterioration
reduces the performance of the β-cells thereby decreased insulin secretion which leads to
diminished ability of muscle and adipose tissue to take up glucose. Although not well
understood it seems as if a constant high blood glucose level ultimately leads to reduced
insulin response and also to negative alterations in insulin secretion (Kahn, 2003). Decreased
insulin secretion together with decreased uptake of glucose causes the glucose concentration
to rise, which again has a negative effect on glucose-uptake and insulin secretion, and this
cascade results in a vicious circle (figure 5).
Figure 5: The effects of β-cell dysfunction Constantly elevated glucose levels down-regulate glucose uptake in muscle and adipose tissue, and decrease insulin secretion, and both events lead to further elevation of glucose (Own drawing, 2011).
The role of miR-29 in type 2 diabetes mellitus
The first scientists to discover a difference in miR-29 expression in T2DM liver, muscle and
adipose tissue compared to non-diabetic tissue of the same type was He et al. (2007). By
Northern blot they found an up-regulation of miR-29a, 29b and 29c in skeletal muscle tissue
from the hyperglycemic, diabetic Goto-Kakizaki (GK) rats compared to the normoglycemic,
non-diabetic Wistar Kyoto (WKY) rats. An up-regulation of miR-29a, 29b and 29c was also
observed in adipose tissue and liver, but the difference in miR-29 expression was not as
profound as for skeletal muscle tissue (figure 6).

Page 11 of 36
Figure 6: Northern blot analysis of miR-29a, 29b and 29c expression in muscle, adipose tissue and liver from GK and WKY rats MiR-29a, 29b and 29c are up-regulated in muscle-, fat- and liver tissue samples from GK (G1, G2, G3, G4 and G5) compared to WKY (N1 and N2). U6 is used control, and here it validates the elevated miR-29 expression levels in the Nothern blot (He et al., 2007).
The science group of Herrera et al. (2009) compared miR-29 expression in liver and adipose
tissue from GK rats and Brown Norway (BN) rats. BN rats are normoglycemic just as the
WKY rats, but they have lower plasma glucose concentrations. miR-29a in the GK rats was
increased by a 1,51 fold in adipose tissue and miR-29c was increased by a 1,5b fold in liver.
These results came from microarrays but were not validated. Microarray is a way of
investigating the expression levels of many miRNAs at the same time. It is an indicator of
differential miRNA expression of different miRNAs. Different oligonucleotide probes are
spotted onto a chip and a miRNA sample is spread onto the chip. The microRNAs are labeled
with a fluorescence gene and the more miRNA that binds to a probes, the more it will light
up. Microarray must be validated because the oligonucleotide probes not only bind to the
mature miRNA but also pri-miRNA and pre-miRNA, and only the mature miRNA are of
interest. Hence, validation by either Northern blotting where the pri-, pre- and mature miRNA
are separated by size or by real-time RT-Q-PCR with primers for the mature miRNA are
necessary. real-time RT-Q-PCR is short for real time Reverse Transcriptation Quantitative
Poly Chain Reaction. real-time RT-Q-PCR is a method used for amplification and
quantification of small RNAs.
A study (Herrera, 2010) combined GK, BN and WKY rat models and found a 1,18 fold
increase of miR-29a expression in adipose tissue from GK compared to BN, which supports
the findings of Herrera et al. (2010). miR-29a expression was reduced by a 1,68 in BN
compared to WKY, and unexpectedly reduced by a 1,42 fold in GK compared to WKY

Page 12 of 36
(table 1) (Herrera, 2010). This contradicts the results from He et al. (2007) who found
elevated expression of miR-29 in GK compared to WKY.
GK/BN BN/WKY GK/WKY
Tissue miRNA Fold change p value Fold
change p value Fold change p value
Adipose miR-29a 1,8 2,8*10-1 -1,68 3,3*10-4 -1,42 4,0*10-2
Table 1: Comparison of miR-29a expression in GK, WKY and BN rats Expression of miR-29a is increased by a 1,8 fold in GK compared to BN and is decreased 1,42 fold compared to WKY. Expression of miR-29a is decreased by a 1,68 fold in BN compared to WKY. The results were validated by real-time RT-Q-PCR (Extract from Herrera, 2010).
Yet another group of scientists found that miR-29a levels were increased in diabetic liver
tissue, this time by using db/db mice liver compared to the control (Pandey et al., 2011). The
results of the experiment were validated by Northern blot and real-time RT-Q-PCR (Pandey
et al., 2011). Muscle and adipose tissue were not included in this trial. db/db mice are obese
and not lean as the GK rats (Srinivasan, 2007). Obesity induces stress in cells and can
therefore possibly alter expression levels of different transcripts. One could therefore argue
that GK rats constitute better models than db/db mice, but in this case the experiments
showed an up-regulated level of miR-29a in liver from both db/db nice and GK rats.
The correlation of glucose concentrations and miR-29
Expression of miR-29a and 29b is up-regulated in 3T3-L1 adipocytes under hyperglycemia
(25mM) and hyperinsulinemia (100nM) compared to physiologically normal conditions
involving a homeostatic glucose concentration of 5mM and no insulin (He et al., 2007). The
up-regulation was validated by Northern blot. Another group of scientist also showed
elevated levels of miR-29a in 3T3-L1 adipocytes when glucose concentrations rose from
5mM to 25mM (insulin was kept constant at 100nM) (Herrera, 2010). An interesting
observation was that miR-29 expression was elevated when glucose concentrations rose from
the 5mmol/L to 15mmol/L, but a further elevation of glucose concentration to 25mmol/L had
no significant effect on miR-29a expression (Herrera, 2010). real-time RT-Q-PCR was used
to validate this finding. It was also suggested that 3T3-L1 adipocytes transfected with miR-29
reduced glucose uptake by approximately 50%, although this result was not validated (He et
al., 2007).

Page 13 of 36
Unpublished data from A. Bagge et al. (2011) have shown that elevated levels of glucose in
human islets of Langerhans and in the β-cell cell line INS-1E increased the expression of
miR-29a (p<0,05 and p<0,001 respectively). Over-expression of miR-29a in INS-1E
decreased Glucose-Stimulated Insulin Secretion (GSIS) in both high glucose and low glucose
concentrations. RT-Q-PCR was used as validation method.
If elevated miR-29a expression decreases glucose uptake in 3T3-L1 adipocytes and reduces
GSIS in INS-1E, and elevated glucose concentrations increases miR-29a expression in 3T3-
L1 adipocytes and INS-1E, and elevated miR-29a expression in β-cells reduces insulin
secretion it might indicate a role of miR-29a in the vicious circle mentioned earlier (figure 5).
It is possible to imagine miR-29a also having an effect on glucose uptake in muscle tissue
although experiments on the connection between miR-29a and glucose uptake in muscle
tissue have not been conducted. We have chosen to include muscle tissue in figure 7 below
that shows a possible connection between elevated glucose concentrations, the de-regulation
it leads to and miR-29a.
Figure 7: The possible effect of miR-29a in the vicious circle of elevated glucose levels in β-cells, muscle and adipose tissue Elevated glucose levels down-regulate glucose uptake in muscle and adipose tissue, and decrease insulin secretion, and both events lead to further elevation of glucose. An increase in glucose concentration in β-cells, 3T3-L1 adipocytes (and possibly in muscle tissue as well) elevates miR-29a expression in these tissues which leads to a decrease in glucose uptake (not validated) and insulin secretion. Again both of these events lead to an increase in glucose levels and thereby also in miR-29a levels. (Own drawing, 2011)

Page 14 of 36
miR-29 and the insulin-signaling pathway
miR-29a targets p85α, the regulatory subunit of PI3K in a human hepatocellular cell line
called HepG2 (Pandey et al., 2011). When miR-29a targets p85α, p85αt cannot activate p110
and this leads to inhibited phosphorylation of PIP2 to PIP3. To activate translocation of
GLUT4 to the cell surface Akt must bind to PIP3. Since the amount of PIP3 is reduced
glucose uptake will be diminished due to the lacking translocation of GLUT4.
Comparison of the results from He et al (2007), Herrera et al. (2009) , Herrera (2010),
Pandey et al. (2011) and Bagge et al. (unpublished data, 2011) indicate that down-regulation
of miR-29 could be a possible treatment of T2DM. But regulation of the complicated systems
of the body might induce undesirable results, and it is therefore important to investigate
possible consequences of such interferences. We have chosen to take a closer look at the
coherence between miR-29 and cancer, as this a field attracting more and more attention. To
understand a possible role of miR-29 in cancer development it is convenient to know
something about the cell cycle of cancer for which reason it is described below.
Cancer and the cell cycle
The majority of human cells are in a resting or non-proliferating state; G0. Due to the fact that
the cell is not dividing in this state it is not a phase of the cell division cycle. The cell division
cycle is divided into four phases: G1, Synthesis (S), G2 and Mitosis (M). Cells stimulated by
growth factors enters G1 thereby beginning the cell cycle (figure 8). During S the
chromosomes are replicated and during M the cell divides into two identical daughter cells
(Campbell, 2008). Five checkpoints around the cycle regulate the transition from one phase
to the following. The checkpoints only allow the cycle to proceed if the previous phases have
progressed correctly. The checkpoints monitor the phases by detecting signals such as DNA
damage (Garret, 2001; Campbell, 2008; Vermeulen, 2003). One checkpoint; the restriction
point is often referred to as the “point of no return” because the cell is committed to enter the
cell cycle and proceed throughout the cycle if it passes this checkpoint (Murakami, 2001;
Garret, 2001). The cells need growth factor stimulation to pass the restriction point.
Afterwards growth factors are no longer required (Murakami, 2001; Garret, 2001). Only the
restriction point is of interest to this report and the four other checkpoints are therefore not
further described.

Page 15 of 36
Figure 8: The cell cycle and its checkpoints The cell cycle starts from G1 proceeding towards
the final phase of mitosis (M). The cell cycle is
divided into four phases: G1, S, G2 and M. Another
fifth phase, not part of the cell cycle is G0 which is
a non-proliferation phase. Besides the transition
states, different checkpoints around the cycle are
monitoring whether the phases have progressed
correctly. The restriction point (R), the G1/S DNA
damage checkpoint, the S Phase DNA checkpoint,
the G2 DNA damage checkpoint and the mitotic
checkpoint (Modified from Lynam-Lennon, 2008).
Normal cells that follow the general aging pattern of the cell cycle proliferate into a
monolayer and require attachment to a matrix to grow. Cancer cells do not have the same
requirements. Cancer cells proliferate without growth factors, do not require attachment to a
matrix and can proliferate to become more than a single layer such as a clot (Campbell,
2008). Cancer appears when one or more checkpoints of the cell cycle are deregulated
(Garret, 2001).
Cyclins and cyclin dependent kinases
In addition to the checkpoints the cell cycle is also controlled by cyclins and cyclin dependent
kinases (CDK’s) (Campbell, 2008). There are different types of cylins; A, B, D and E. The
different types of cyclins are present during different stages of the cycle and in varying levels.
The level of a specific cyclin tops just before a transition state (Vermeulen, 2003).
The enzymatic activity of the CDK’s is regulated through phosphorylation, binding to a
partner cyclin (Garret, 2001) and through binding to two CDK-inhibitors; Cip1/Kip1 and
INK4 (Murakami, 2001). Inactive CDK’s are located in the cytoplasm and the levels are
constant throughout the cycle (Collins, 1996). A connection between certain CDK’s and miR-
29 has been found, and will be described in section “Lymphocytes, AML”.

Page 16 of 36
Analysis
If down-regulation of miR-29 is a possible treatment to T2DM patients it may only be
necessary to down-regulate it in insulin-responsive tissues. Therefore the first part of the
analysis will concern these tissues except for adipose tissue as to our knowledge no
experiments have been made concerning this insulin-responsive tissue. Connections between
miR-29 and cancer in non-insulin responsive tissues have also been detected and this is
interesting in case miR-29 cannot be down-regulated tissue-specifically. Therefore articles
regarding non-insulin responsive tissues will be analyzed in the section “miR-29 and cancer
in non-insulin-responsive tissues” below.
The scientists of the many articles describe a few targets of mir-29 in their articles, and they
are in a directory of targets of miR-29 in appendix I. Some of them will be described in the
analysis where relevant, and the rest will only be presented in the directory.
miR-29 and cancer in insulin-responsive tissues
Liver, hepatocellular cancer
A study conducted by Xiong et al. (2010) has shown reduced levels of miR-29 in
HepatoCellular Carcinoma (HCC) cells compared to normal liver cells. The differential
expression of miR-29 was investigated by Northern blot
in four human hepatocellular carcinoma cell lines:
HepG2, QGY-7703, MHCC97H and SMMC-7721.
When looking at the Northern blot in figure 9 it could
seem as if miR-29 was not particularly down-regulated
in the HCC cell lines compared to normal liver cells, but
the U6 control line at the bottom shows weaker
expression for normal liver cells than the other samples,
indicating that the down-regulation postulate is right.
Xiong et al., (2010) validated the down-regulation by RT-Q-PCR. The control line should
always be equally expression in every tissue, and a smaller expression in one lane could
indicate that a smaller amount of RNA (including the control) was administered here.
Figure 9: miR-29 in HCC Northern blotting showed a reduction in miR-29 levels in normal liver tissue compared to HCC cell lines. (Modified from Xiong et al., 2010).

Page 17 of 36
A detection of a down-regulation of miR-29 in HCC cell lines says nothing about which
came first; the decreased mir-29 expression or the cancer? A group of scientists; Braconi et
al. (2011) showed that the decrease in miR-29a might be first on the scene. They found that
miR-29a up-regulates the gene MEG3 and down-regulates the DNA methyltransferases
DNMT-1 and DNMT-3B in HCC. A decrease of MEG3 was found in HCC cell lines and
down-regulation of DNMT in human primary HCC increases MEG3 which suggests that
miR-29a might target MEG3 through DNMT (figure 10). Braconi et al. (2011) validated all
the experiments by real-time RT-Q-PCR. All together these results indicate an increase in
miR-29a being a source in the development of HCC.
Figure 10: The connection between miR-29a and the possible development of HCC miR-29a down-regulation increases DNMT expression and reduces the expression of MEG3 which has been shown to correlate with the development of HHC (Own drawing 2011).
miR-29 also targets Bcl-2 and Mcl-1 in HCC, this time shown in HepG2 and HEK293T cells
(Xiong et al., 2010). Bcl-2 and Mcl-1 are two apoptosis regulating proteins. Bcl-2 is encoded
by the gene BCL-2 and Mcl-1 is encoded by the gene MCL-1. The targets were predicted in
silico and validated by Luciferase (Luc.) assay. Wild type MCL-1-3’UTR, BCL-2-3’UTR,
mutated MCL-1-3’UTR or mutated BCL-2-3’UTR was cloned into one plasmid each
downstream of a parental Luc. gene. HepG2- and HEK293T cells were transfected with one
of the plasmids and then with mature miR-29 (figure 11).
Figure 11: Luciferase assay on Mcl-1 genes: MCL-1 Wild type MCL-1-3’UTR or mutated MCL-1-3’UTR is cloned into a plasmid downstream of a parental Luciferase gene. Cancer cells are transfected with plasmids cloned with either wild type or mutant and then with mature miR-29 (Own drawing, 2011).

Page 18 of 36
If miR-29 targets Bcl-2 or Mcl-1 there will be a decrease in Luc. activity in cells transfected
with wild-type compared to the mutant. The mutated MCL-1-3’UTR and BCL-2-3’UTR is
formed by point mutation, differing in only a few nucleotides from the wild type. The less
mutated the sequence is, the less nucleotides differ between the wild type 3’UTR and the
mutated 3’UTR, and the more precise the examination of the respective target is. Xiong et al.
(2010) do not state the number of mutated nucleotides.
As shown in figure 12A1 from Xiong et al. (2010) the Luc. activity is significantly (p<0,001)
decrease in cells transfected with wt-BCL-2-3’UTR when compared to the Negative Control
(NC). Figure 12A2 shows only a small decrease in Luc. activity in cells transfected with
mutated wt-BCL-2-3’UTR compared to NC. The cells transfected with wt-BCL-2-3’UTR
shows a reduced Luc. activity compared to the mutated BCL-2-3’UTR. This shows that miR-
29 binds to the wild type and not to the mutated type, hence it has been shown that miR-29
targets Bcl-2. Figure 12B1 and B2 shows the same tendency for Mcl-1. The cells transfected
with wt-BCL-1-3’UTR creates a significantly (p<0,01) reduced Luc. activity compared to
mutated MCL-1-3’UTR, showing that miR-29 targets Mcl-1. Used as control in A1, A2, B1
and B2 Luc. assays were HepG2 and HEK293T cells transfected with a non-humane
nucleotide sequence (Xiong et al,. 2010).
Figure 12: miR-29 targets Bcl-2 and Mcl-1 (A1+A2) Luciferase (Luc.) assay was conducted on HepG2- and HEK293T cells (cancerous cells). Negative control (NC) was none-human nucleotides. Cells transfected with wild type BCL-2-3’UTR showed a diminished Luc. activity compared to the NC. Cells transfected with mutated BCL-2-3’UTR showed no significant (***= p<0,001) decrease in Luc. activity indicating that miR-29 targets Bcl-2 (Modified from Xiong et al., 2010). (B1+B2) Cell transfected with wild type MCL-1-3’UTR and mutated MCL-1-3’UTR. Wild type transfected cells show a significant decrease (**= p<0,01) in Luc. activity compared to mutated transfected cells, showing that miR-29 targets Mcl-1 (Modified from Xiong et al., 2010).

Page 19 of 36
Control cells must always undergo the same treatment as the cells of investigation to make
sure that it is not very event of transfection that causes the differences in miRNA expression.
Non-human nucleotide sequences are often used because it is assumed that they cannot target
anything in a human cell.
Mcl-1 exists in three different isoforms. Mcl-1 isoform 1 inhibits apoptosis and is thus MCL-
1 isoform 1 is an oncogene. Mcl-1 isoform 2 and 3 both induces apoptosis and are thus tumor
suppressor proteins (http://www.ncbi.nlm.nih.gov/sites/entrez?Db=gene&Cmd=Show
DetailView&TermToSearch=4170). Xiong et al., (2010) showed a reduced level of miR-29
in HCC cells. A reduced level of miR-29 results in an increase in Mcl-1 and due to the cancer
development this Mcl-1 must be isoform 1. When miR-29 is down-regulated Mcl-1 isoform 1
is up-regulated resulting in a reduction of apoptosis repression. Repression of apoptosis
causes a diminished cell cycle arrest of the cells leading to uninhibited cell proliferation.
Muscle, rhabdomyosarcoma
Wang et al. (2008) showed a down-regulation of miR-29b2 and 29c in Rhabdomyosarcoma
(RMS) tumors versus normal human skeletal muscle, and also a down-regulation in RMS cell
lines (RH28, RH30, RH3, CW9019, SMS-CTR, RD2 and RH36) versus normal human
skeletal muscle cells. The down-regulations were validated by real-time RT-Q-PCR in both
trials. By Luc. assay they found that miR-29b2 targets Yin Yang 1 (YY1) in skeletal muscle
tissue and also that YY1 regulates mir-29b2 in a positive feedback mechanism. YY1 is a
regulatory protein of the NF-κB-pathway and it acts by inhibiting skeletal muscle cell
myogenesis (Wang et al., 2007). Activation of NF-κB is often amplified in RMS resulting in
too much reduction of miR-29, which causes an insufficient regulation of YY1 by miR-29.
This leads to a deregulation of YY1 resulting in RMS. Due to the targeting of YY1 by miR-
29, one might think that a down-regulation of miR-29 would result in an up-regulation of
YY1 which would repress cell proliferation and therefore unlikely result in cancer. But the
fact that a deregulation of YY1 causes RMS anyway indicates that any deregulation in a
pathway can induce undesirable consequences such as for instance cancer.

Page 20 of 36
Pancreas, endocrine tumors
miR-29 expression was also investigated pancreatic tissue and was shown to be up-regulated
in pancreatic endocrine tumors compared normal pancreas (Roldo et al., 2006). The increase
was shown by microarray and quantified using a computer program, but the result was not
validated. Consistent with this there is an elevated expression of miR-29a, 29b and 29c in
pancreatic cancer tissue specimens with respect to normal pancreatic tissue and in MiaPaCa-
2-, PANC-1- and BxPC-3-cell lines versus HPDE cells (Zhang, 2008). MiaPaCa-2, PANC-1
and BxPC-3 are epithelial-like human pancreatic carcinoma cell lines. HPDE cells are
immortalized human pancreatic duct epithelial cells. miR-29a and miR-29b is also elevated in
MiaPaCa-2 and PANC-1 cell lines compared to Mock controls (Lee et al., 2009). The
increase of miR-29 in pancreatic cell lines was found by microarray, but was not validated by
neither Zang (2008) nor Lee et al. (2009). Together these findings indicate a tendency of
elevated miR-29 expression in cancerous pancreatic tissue which is interesting for the
prospect of treating T2DM by down-regulating miR-29 expression in β-cells without
increasing the risk of developing pancreatic cancer.
miR-29 and cancer in non-insulin-responsive tissues
We have chosen to present the up- or down-regulation of miR-29 expression in three cancer
types in writing, and then present rest of the data found on the coherence between miR-29
expression and cancer in an accumulating figure in the end of the section (figure 15).
Lymphocytes, B-cell chronic lymphocytic leukemia
miR-29a and miR-29b is up-regulated in indolent and aggressive samples of B-cell Chronic
Lymphocytic Leukemia (CLL) compared to normal B-cells (Santanam et al., 2010). The up-
regulation was shown by real-time RT-Q-PCR. Over-expression of miR-29 in B-cells in
transfected mice lead to increased spleen-levels of the antigen CD5+ in 85% of the mice
(measured by real-time RT-PCR) and to an enlarged spleen over time (Santanam et al.,
2010). It has earlier been suggested that increased levels of the antigen CD5+ induces CLL
development (Bichi et al., 2002), and this supports the findings from Santanam et al. (2010)
of elevated miR-29a and miR-29b in CLL compared to normal B-cells. Together these events
indicate that an increase in miR-29 expression could induce CLL development. This is

Page 21 of 36
slightly supported by results from Santanam et al. (2010) showing that 4 of 20 miR-29
transfected mice developed frank leukemia and died.
However, contradicting findings have also been found. By Luc. assay it has been shown that
miR-29b targets tcl1 (Pekarsky, 2006), and that tcl1 over-expression might play an important
role in inducing the development of aggressive CLL (Pekarsky et al., 2008). The Luc. assay
did not involve a mutated target-3’UTR, and therefore it can be discussed whether the target-
result is valid. But if miR-29 targets tcl1, and tcl1 over-expression might play an important
role in inducing the development of aggressive CLL, a down-regulation of miR-29 could
induce CLL. But maybe the role of miR-29 in CLL is not that easy to categorize. Santanam et
al. (2010) surprisingly found that miR-29a expression was higher for indolent than for
aggressive CLL. All this suggest that any deregulation of miR-29 could induce CLL but that
a high up-regulation induces aggressive CLL and a smaller up-regulation, or maybe even a
down-regulation of miR-29 could induce indolent CLL.
A Western blot showed a decrease of CDK6 and DNMT-3A expression in B-cells from miR-
29 tranfected mice (Santanam et al., 2010). This correlates with the findings from Braconi et
al. ( 2011) where a miR-29a up-regulation in a HHC cell line causes a decrease in DNMT-3A
(and DNMT-1). The possible consequences of a decrease in CDK6 expression will be
discussed later in this section.
Lymphocytes, AML
miR-29a expression is elevated in Leukemia Stem Cells (LSC) collected from human primary
AML patients compared to normal Human Hematopoietic Stem cells (HSC) (Han et al.,
2010). Han et al. (2010) used different HSC’s: Lin-, CD34+, CD90+ and CD45Ra-, and
compared them to different LSC’s: Lin-, CD34+ and CD38+. The elevation was validated by
real-time PCR.
Another group of scientists have shown that miR-29b is down-regulated in primary Acute
Myolid Leukemia (AML) patient specimens compared to CD34+ cells from healthy donors
(Garzon, 2008). The down-regulation was shown by microarray and validated by real-time
RT-Q-PCR. The results contradict the findings from Han et al. (2010). Garzon (2008) did not
specify the names of the cells that were used for the miRNA expression profiling (Garzon,

Page 22 of 36
2008). The cells used by Garzon (2008) and Han et al. (2010) could differ from each other,
and this could be the reason for the different results.
Garzon (2009) showed that miR-29 targets cyclin dependent kinase 6 (CDK6) in ALM cells.
The target was validated by Luc. assay on K562 cells. K562 are human cancer cells from
AML and contains low levels of miR-29. The fact that miR-29 targets CDK6 is consistent
with the findings of Santanam et al. (2010) only they did validate their findings. Furthermore
Santanam et al. (2010) performed a Western blot using normal B-cells from miR-29-
transfected mice compared to wild type mice without miR-29. To understand how miR-29’s
targeting of CDK6 might induce cancer it is relevant to know how CDK6 functions in a
healthy cell.
CDK4 and CDK6 are active during G1 and their activity relies on cyclin D1. Cyclin D1 levels
top just before transition from G1 to S, and this results in an increase in cyclin D1/CDK4
complexes (cyclin D1/CDK4) and cyclin D1/CDK6 complexes (cyclin D1/CDK6) at this
transition state (Vermeulen, 2003). The transition from G1 to S requires activation of the
tumor suppressor protein Retinoblastoma (Rb). Rb is in a hypophosphorylated state bound to
the transcriptionfactor E2F in the beginning of G1. E2F is required for expression of genes
required for entry of S-phase (figure 13) (Giorgia, 2011).
Figure 13: Phosphorylation of Retinoblastoma Complexes of cyclin D1/CDK4, cyclin D1/CKD6 and cyclin E/CDK2 phosphorylate the tumor suppressor protein Retinoblastoma (Rb) thereby releasing the transcription factor E2F. E2F promotes cell cycle progression, gene transcription and the cell proceeds to S-phase. Two cyclin dependent inhibitors: INK4 and CIP/KIP, regulate the cycle by inhibiting the CDKs and indirectly regulate the phosphorylation of Rb (Modified from Giorgia, 2011).

Page 23 of 36
Cyclin D1/CDK4(6) phosphorylates Rb yielding a free E2F, and allowing cyclin E
expression. To be completely phosphorylated Rb requires enzymatic activity from cyclin
E/CDK2 complex. Expression of cyclin E results in formation of cyclin E/CDK2, and so the
Rb is fully phosphorylated (Sherr, 1996; Giorgia, 2011).
Mutations in the gene encoding cyclin D1 can cause undesirably elevated levels of cyclin D
which leads to increased levels of cyclin D1/CDK4(6) complexes resulting in an elevation of
phosphorylated Rb and thereby a deregulation of the checkpoint. Mutations in the gene
encoding cyclin D1accours in Mantle Cell Lymphoma (MCL) (Zhao, 2010). miR-29 levels in
MCL is decreased in comparison to non-cancerous CD19+ lymphocyte control cells (Zhao,
2010). This was shown by microarray and validated by real-time RT-Q-PCR where CD19+
peripheral blood lymphocytes and lymph nodes were used as controls. miR-29 is also
negatively correlated with CDK6 in MCL (Zhao, 2010). Due to the high expression of cyclin
D1 in MCL and the high expression of CDK6 transcripts caused by low miR-29 expression
there is a lot of cyclin D1/CDK6 complexes in MCL. These events result in uninhibited
phosphorylation of Rb and thereby uninhibited cell proliferation that could lead to MCL
developing.
Bile duct, cholangiocarcinoma
miR-29b is diminished in malignant cholangiocarcinoma cells (KMCH) with respect to non-
malignant H69 cholangiocytes (Mott et al., 2007). H69 cholangiocytes are epithelial cells of
the bile ducts. The finding was validated by real-time RT-Q-PCR (figure 14A). Consistent
with Xiong et al. (2010), Mott et al. ( 2007) found Mcl-1 to be a miR-29 target, this time by
Luc. assay on HeLa cells and KMCH cells (figure 14B) (Mott et al., 2007). HeLa is a human
cervical cancer cell line. A parental Luc. and a wt-MCL-1-‘3UTR was cloned into a plasmid.
The controls were cells transfected with either parental Luc. and single-nucleotide-mutated-
Mcl-1-‘3UTR or parental Luc. alone. miR-29b is down-regulated in KMCH and HeLa cells
and targets Mcl-1 in the same cells. This means that Mcl-1 is most likely increased in KMCH
and this reduces apoptosis and therefore induces risk of cancer development.

Page 24 of 36
Figure 14: miR-29 expression in malignant cholangiocarcinoma cells (KMCH) and Mcl-1 targeting (A) real-time RT-Q-PCR showed a decrease in miR-29 expression in KMCH cells compared to H69 cells. (B) Luciferase (Luc.) assay showed that miR-29b targets Mcl-1. Parental Luc. gene (pLuc.) was transfected into HeLa cells and KMCH cells. The control was HeLa/KMCH cells cotransfected with pLuc. mutated 3’UTR Mcl-1. Cells transfected with pLuc-Mcl-1 3’UTR and miR-29 (p29b) was down-regulated versus pLuc. and pLuc-Mutant 3’UTR (Modified from Mott et al., 2007).
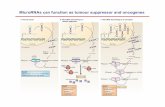
Is miR-29 a tumor suppressor, an oncomiR or both?
Targets such as CDK6 and Mcl-1 have been described in several articles regarding miR-29
and cancer. CDK6 is a key-player of cell cycle regulation and is a promoting factor of the
cycle, and is thus a proto-oncogene. In the context of targeting CDK6, miR-29 is therefore a
tumor suppressor miRNA. miR-29 has been shown to target Mcl-1 in HHC and AML (only
miR-29b), and miR-29/b is down-regulated in both cancer types. We therefore conclude that
the target of miR-29 in AML and HHC must be Mcl-1 isoform 1, and not isoform 2 or 3. One
could imagine that miR-29 could also target Mcl-1 isoform 2 and 3, and thereby be an
oncomiR instead of a tumor suppressor miR.
In general miR-29 can be an oncomiR or a tumor suppressor miR dependent on the target.
Four cascades can be proposed:
Target Regulation of miR-29 Induces cancer development Oncogene Up No Tumor suppressor Up Yes Oncogene Down Yes Tumor suppressor Down No

Page 25 of 36
Besides CDK6 and Mcl-1 we also encountered an article concerning miR-29 and another
tumor suppressor protein; p53. Park et al. (2009) found that miR-29 is positively correlated
with p53, and that miR-29 induces apoptosis in p53-positive HeLa cells compared to HeLa
cells with p53 knocked down suggesting that the induction of apoptosis by miR-29 happens
through p53. p53 regulates the G1/S-checkpoint (figure 8) (Vermeulen, 2003) and is activated
when the checkpoint receives a signal of DNA damage (Garret, 2001). Activation followed
by up-regulating of p53 increases expression of genes that induces cell arrest and apoptosis
(Garret, 2001). When not activated p53 levels are low in the cells due to MDM2-protein
binding leading to ubiquitin-degradation of p53. Ubiquitin is a cellular trash-machine that
guides proteins to degradation when they are no longer needed (Garret, 2001; Murakami,
2001). If p53 is not properly activated when DNA damage is detected it will not induce cell
arrest and apoptosis, and an uninhibited proliferation can occur.
By Luc. assay Park et al. (2009) found that miR-29 directly targets p85α and Cell Division
Cycle Protein 42 (CDC42) in HeLa cells. Through p85α miR-29 might reduce the
phosphorylation of Akt which normally inactivates MDM2 (Park et al., 2009). When MDM2
levels are not properly inactivated the levels become too high causing excessive binding to
p53 and thereby a degradation of p53.

Page 26 of 36
Figure 15: Overview of up- or down-regulation of miR-29 in cancers of various tissues
* = Validated

Page 27 of 36
Summative discussion and conclusion
Whether miR-29 is up- or down-regulated in cancer tissues compared to normal tissue is
highly dependent on the tissue type. miR-29 seems to be down-regulated in brain, lung, liver,
cervix, muscle and stomach cancers, and up-regulated in cholangiocytes, prostate, pancreas
and some lymphocytes with possible exceptions.
Up-regulation of miR-29a, 29b and 29c in pancreatic cancers are especially interesting in
relation to the possibility of using a repression of miR-29a to treat patients suffering from
T2DM. But it is important to notice that the findings were not validated! If these findings
become validated and miR-29a repression could be done tissue-specifically, then it might
serve as part of T2DM-treatment. Some miRNAs are tissue-specific, but miR-29 is clearly
not. In case a miR-29a repression occurs in all tissues and not only in pancreas one would
possibly expose the T2DM patients to an increased risk of developing cancer if miR-29a
expression was clinically repressed.
The connection between miR-29a, 29b1, 29b2 and 29c.
miR-29a and 29b1 is expressed on the same gene. The same accounts for miR-29b2 and 29c.
Li et al. (2011) found decreased levels of miR-29a and 29c in cervical cancer. The result was
not validated, and one would expect to see reduced levels of miR-29b1 and 29b2 too, if miR-
29a and 29c are both decreased. As for experiments conducted with reference to brain
(Silber, 2008) and cholangiocytes (Mott et al., 2007) differences in expression was detected
only for miR-29b, whereas experiments conducted with reference to lung (Yanaihara, 2006)
detected miR-29b2 expression differentiation (not validated). miR-29a or 29c should in this
case follow the increase or decrease that miR-29b exhibits in brain and cholangiocytes,
whereas only miR-29c should follow the increase or decrease miR-29b2 exhibits.
Using miRNA in cancer treatment
The detection of an either elevated or reduced level of a certain miRNA in a certain type of
cancer can be utilized in patient-treatment in at least three ways; prognosis assessment,
regulation of the particular miRNA in that cancer tissue type and for classification of the

Page 28 of 36
cancer. There have been examples of miRNA-levels rightfully placing tumors in the
appropriate category (Subramanian et al., 2007). The right categorization plays an important
role in choosing the optimal treatment.
A down- or up-regulation of the particular elevated or reduced miRNA could also be part of
cancer treatment. But as discussed earlier it is of utmost importance to make sure that the
clinical regulation of the miRNA occurs only in the tissue-type chosen as target of treatment.
Regulation of miRNAs could be done either by inducing a “fake target” of the miRNA to
which it will bind or by repressing the miRNA. TGF-β, YY1 and c-Myc are different genes
found to target and repress miR-29 (Winbanks, 2011; Wang, 2008; Mott, 2010). Two
miRNAs; miR-29a and 29b have been shown to induce apoptosis in cholangiocarcinoma and
AML cells (Mott et al. 2007; Garzon, 2009). Furthermore it has been shown that miR-29b
has a tumor suppressive effect in vivo on nude AML mice (Garzon, 2009; Xiong et al.,
2010). This indicates that induction of miR-29 is a possible treatment cholangiocarcinoma
and AML.
The detection of either down- or up-regulation of a certain miRNA can also be utilized in the
prognosis of cancer patients. The expression profiling of miRNA in a certain cancer type in
different stages of the cancer development compared to corresponding normal tissue can be a
guide in choosing the optimal treatment. Some scientists have investigated indolent versions
of a cancer type compared to the aggressive version (Pekarsky, 2006; Santanam et al., 2010;
Muniyappa 2009; Calin, 2007) instead of comparing cancer and non-cancer. The focus is
thereby the miRNA-induced or -repressed pathogenesis effect of the cancer. If regulation of
one or more miRNAs could shift the cancer from being aggressive to being indolent it gives
hope that the cancer might be beaten by additional cancer-treatments that alone would not
have killed the cancer.
Models vs. the human body
It is important to keep in mind that it is seldom possible to conduct experiments under
physiologically relevant or clinically relevant conditions. Cell lines are often utilized for
experiments, though they are not a perfect analogue to the human body because they are not
part of the overall unit that the body constitutes. But they have the advantage of being easy to
achieve, easy to grow in a petri dish and easy to transfect. A cell line used for experiments is

Page 29 of 36
chosen from many cell cultures, and it is chosen because it is judged to be the best analogue
to the tissue it resembles. Some scientists utilize specimens from human patients for their
experiments, which makes the experiment more physiological relevant than if cell lines were
used.
All the experiments described in this report are conducted on cell lines, specimens or mice,
which means that the results are only indications of what might occur in a real live human
being. And additionally it has been suggested that gender and starvation periods could alter
miRNA expression in rat liver (Cheung et al., 2009). It is possible that these parameters and
maybe other additional ones could also alter miRNA levels in humans. The experiments
might only give results that concern the particular model the experiment was conducted on,
and not general results.
The role of miR-29 in cancer development
There is no overall tendency of miR-29 being up- or down-regulated in cancer. miR-29
expression levels are highly dependent on the cancer tissue type. But it seems probable that
miR-29 do play a role in the development of cancer, although no absolute conclusions can be
made before clinical trials on humans are conducted.
Perspective
It would be interesting to investigate the role of other miRNA’s and their targets in cancer
development. Also there has been conducted several trials concerning miR-29 and other
diseases such as for instance sclerosis. This could also be interesting to further investigate.

Page 30 of 36
Refferences
Ambros, V., Lee, R.C., Lavanway, A., Williams, P.T. & Jewel, D. 2003, "MicroRNAs and Other Tiny Endogenous RNAs in C. elegans", Current Biologi, .
Bichi, R., , S., S. A., Martin, E.S., Koval, A., Calin, G.A., Cesari, R., Russo, G., Hardy, R.R. & Croce, C.M. 2002, "Human chronic lymphocytic leukemia modeled in mouse by targeted TCL1 expression", PNAS, vol. 99, no. 10, pp. 6955-6960.
Braconi, C., Kogure, T., Valeri, N., Huang, N., Nuovo, G., Costinean, S., Negrini, M., Miotto, E., Croce, C. M., Patel, J. 2011, "microRNA-29 can regulate expression of the long non-coding RNA gene MEG3 in hepatocellular cancer", Oncogene, vol. 30, pp. 4750-4756.
Burén, J., Liu, H., Lauritz, J. & Eriksson, J.W. 2003, "High glucose and insulin in combination cause insulin receptor substrate-1 and -2 depletion and protein kinase B desensitisation in primary cultured rat adipocytes: possible implications for insulin resistance in type 2 diabetes.", European Journal of Endocrinology, vol. 148, pp. 157-167.
Calin, G.A., Pekarsky, Y. & Croce, C.M. 2007, "The role of microRNA and other non-coding RNA in the pathogenesis of chronic lymphocytic leukemia ", Best Practice & Research Clinical Haematology, vol. 20, no. 3, pp. 425-437.
Campbell, N.A., Reece, J.B., Urry, L.A., Cain, M.L., Wasserman, S.T., Minorsky, P.V. & Jackson, R.B. 2008, "The Cell Cycle" in Biology, ed. B. Wilbur, 8th edn, Pearson Education, Inc., San Francisco, pp. 228-245.
Chen, X. 2005, "microRNA biogenesis and function in plants", FEBS Letters, vol. 579, pp. 5923-5931.
Cheung, L., Gustavsson, C., Nordsted, G. & Tollet-Egnell, P. 2009, "Sex-different and growth hormone-regulated expression of microRNA in rat liver", BMC Molecular Biology, vol. 10, no. 13.
Collins, K., Jacks, T. & Pavletich, N.P. 1997, "The cell cycle and cancer", The National Academy of Sciences of the USA, vol. 94, pp. 2776-2778.
Cui, Y., Su, W., Xing, J., Wang, Y., Wang, P., Chen, X., Shen, Z., Cao, H., Lu, Y. & Fang, J. 2011, "Mir-29a Inhibits Cell Proliferation and Induces Cell Cycle Arrest through down-regulation of p42.3 in Human Gastric Cancer", Plos One, vol. 6, no. 10.
Dirice, E. & Kulkarni, N. 2011, "Pathways Underlying beta-Cell Regeneration in Type 1, Type 2 and Gestational Diabetes" in Islet Cell Growth Factors, ed. R.N. Kulkarni, Landes Bioscience, USA, pp. 1-22.
Fabbri, M., Garzon, R., Cimmino, A., Liu, Z., Zanesi, N., Callegari, E., Liu, S., Alder, H., Costinean, S., Fernandez-Cymering, C., Volinia, S., Guler, G., Morrison, C. D., Chan, K. K., Marcucci, G., Calin, G. A., Huebner, K., Croce, C. M. 2007, "MicroRNA-29 family

Page 31 of 36
reverts aberrant methylation in lung cancer by targeting DNA methyltransferases 3A and 3B", PNAS, vol. 104, no. 40, pp. 15805-15810.
Garret, M.D. 2001, "Cell cycle control and cancer", Current Science, vol. 81, no. 5, pp. 515-522.
Garzon, R., Voliana, S. , Liu, C., Fernandez-Cymering, C., Polumbo, T., Pichiori, F., Fabri, M., Coombes, K., Alder, H., Nakamura, T., Flomenberg, N., Marcucci, G. , Calin, G. A. , Kornblau, S. M., Kantarjian, H., Bloofield, C. D., Andreeff, M., Croce, C. M. 2008, "MicroRNA signatures associated with cytogenetics and prognosis in acute myeloid leukemia", Blood, vol. 111, pp. 3183-3189.
Garzon, R., Heaphy, C.E.A., Havelange, V., Fabbri, M., Volinia, S., Tsao, T., Zanesi, N., Kornblau, S.M., Marcucci, G., Calin, G.A., Andreeff, M. & Croce, C.M. 2009, "MicroRNA 29b functions in acute myeloid leukemia", Blood, vol. 114, pp. 5331-5341.
Gebeshuber, C.A., Zatloukal, K. & Martinez, J. 2009, "miR-29a supresses tristetraprolin, which is a regulator of epithelial polarity and metastasis", European Molecular Biology Organization, vol. 10, no. 4, pp. 400-405.
Giorgia, S. & Bhushan, A. 2011, "Cell Cycle Regulation and Beta Cells" in Islet Cell Growth Factors, ed. R.N. Kulkarni, Landes Bioscience, USA, pp. 31-38.
Han, Y., Park, C.Y., Bhagat, G., Zhang, J., Wang, Y., Fan, J., Liu, M., Zou, Y., Weissman, I.L. & Gu, H. 2010, "microRNA-29a induces aberrant self-renewal capacity in hematopoietic progenitors, biased myeloid development and acute myeloid leukemia", Journal of Experimental Medicin, vol. 207, no. 3, pp. 475-489.
Hancock, J.T. 2010, "Insulin and the signal transduction cascades it invokes" in Cell Signaling, 3rd edn, Oxford University Press, Oxford, pp. 278.
He, A., Zhu, L., Gupta, N., Chang, Y., Fang, F. 2007, "Overexpression of Micro Ribonucleic Acid 29, Highly Up-Regulated in Diabetic Rats, Leads to insulin Resistance in 3T3-L1 Adipocytes", Molecular Endocrinology, vol. 21, no. 11, pp. 2785-2794.
Herrera, B. M., Lockstone, H. E., Taylor, J. M., Wills, Q. F., Kaisaki, P. J., Barrett, A., Camps, C., Fernandez, C., Ragoussis, J., Gauguier, D., McCarthy, M. I., Lindgren, C. M. 2009, "MicroRNA-125a is over-expressed in insulin target tissue in a spontaneous rat model of Type 2 Diabetes", BMC Medical Genomics, vol. 2, no. 54.
Herrera, B.M., Lockstone, H.E., Taylor, J.M., Ria, M., Barrett, A., Collins, S., Kaisaki, P., Argoud, K., Fernandez, C., Travers, M.E., Grew, J.P., Randall, J.C., Gloyn, A.L., Gauguier, D., McCarthy, M.I. & Lindgren, C.M. 2010, "Global microRNA expression profiles in insulin target tissues in a spontaneous rat model of type 2 diabetes", Diabetologia, vol. 53, pp. 1099-1109.
Kahn, S.E. 2003, "The relative contributions of insulin resistance and beta-cell dysfunction to the pathophysiology of Type 2 diabetes", Diabetologia, vol. 46, pp. 3-19.

Page 32 of 36
Karlsson, H.K.R. & Zierat, J.R. 2007, "Insulin signaling and glucose transport in insulin resistant humanskeletal muscle", Cell Biochem Biophys, vol. 48, pp. 103-113.
Kolfschoten, I. G. M., Roggli, E., Nesca, V. ,Regazzi, R.. 2009, "Role and therapeutic potential of microRNAs in diabetes", Diabetis, Obesity and metabolism, vol. 11, pp. 118-129.
Laegos-Quintana, M., Rauhut, R., Lendeckel, W. & Tuschl, T. 2001, "Identification of novel genes coding for small expressed RNAs", Science, vol. 294, no. 5543.
Lang, N., Liu, M., Tang, W., Chen, X., Liu, Z. & Bi, F. 2010, "Effects of microRNA-29 family members on proliferation and invasion of gastric cancer cell lines", Chinese Journal of Cancer, vol. 29, no. 6, pp. 603-610.
Le Roith, D., Quon, M. J., Zick, Y. 2003, "Molecular and cellular aspects of insulin resistance: implications for diabetes" in Signal Transduction and Human Disease, eds. T. Finkel & J.S. Gutkind, EDITION edn, John Wiley & Sons, Inc., New Jersey, pp. 171-191.
Lee, K., Lotterman, C., Karikari, C., Omura, N., Feldmann, G., Habbe, N., Goggins, M. G., Mendell, J. T., Maitra, A. 2009, "Epigenetic Silesing of MicroRNA miR-107 Regulates Cyclin-Dependent Kinase 6 Expression in Pancreatic Cancer", Pancreatology, vol. 9, pp. 293-301.
Lee, R.C., Feinbaum, R.L. & Ambros, V. 1993, "The C. elegans Heterochronic Gene lin-4 Encodes Small RNAs with Antisense Complementarity to &II-14", Cell, vol. 75, pp. 843-854.
Li, Y. , Wang, F., Xu, J., Ye, F., Shen, Y., Zhou, J., Lu, W., Wan, X., Ma, D., Xie, X. 2011, "Progressive miRNA expression profiles in cervical carcinogenesis and identificatiopn of HPV-related target genes for miR-29", Journal of Pathology, vol. 224, pp. 484-495.
Lund, S. & Jessen, N. 2007, "Insulins metaboliske og cellulære effekter" in Diabetes, eds. J. Hilsted, K. Borch-Johnsen & J.S. Christiansen, 1st edn, Munksgaard, København, pp. 21-29.
Lynam-Lennon, N., Maher, S.G. & Reynolds, J.V. 2009, "The roles of microRNA in cancer and apoptosis", Biological Reviews, vol. 84, pp. 55-71.
Madsbad, S. & Schmitz, O. 2007, "Ætiologi og patogenese type 2 (insulinsekretion)" in Diabetes, eds. J. Hilsted, K. Borch-Johnsen & J.S. Christiansen, 1st edn, Munksgaard, København, pp. 74-84.
Martinez et al. 2011, "miR-29 and miR-30 regulate B-Myb expression during cellular senescence", PNAS, vol. 108, no. 2, pp. 522-527.
Mott, J.L., Kobayashi, S., Bronk, S.F. & Gores, G.J. 2007, "mir-29 Regulates MCl-1 Protein Expression and Apoptosis", Oncogene, vol. 26, no. 42, pp. 6133-6140.

Page 33 of 36
Mott, J.L., Kurita, S., Cazanave, S.C., Bronk, S.F., Werneburg, N.W. & Fernandez-Zapico, M.E. 2010, "Transcriptional Suppression of mir-29b1/mir-29a Promoter by c-Myc, Hedgehog, and NF-kappaB", Journal of Cellular Biochemistry, vol. 110, pp. 1155-1164.
Muniyappa, M.K., Dowling, P., Henry, M., Meleady, P., Doolan, P., Gammell, P., Clynes, M. & Barron, N. 2009, "MiRNA-29a regulates the expresion of numerous proteins and reduces the invasiveness and proliferation of human carcinoma cell lines", European Journal of Cancer, vol. 45, pp. 3104-3118.
Murakami, M.S. & Woude, G.F.V. 2001, "Regulation of the cell cycle " in Molecular basis of cancer, eds. J. Mendelsohn, P.M. Howley, M.A. Israel & L.A. Liotta, 2nd edn, Saunders, Philadelphia, pp. 10-17.
Ozen, M., Creighton, C.J., Ozdemir, M. & Ittmann, M. 2008, "Widespread deregulation of microRNA expression in human prostate cancer", Oncogene, vol. 27, pp. 1788-1793.
Pandey, A. M., Verma, G., Vig, S., Srivastava, S., Srivastava, A. K., Datta, M. 2010, "miR-29a levels are elevated in the db/db mice liver and its overexpression leads to attenuation of insulin action on PEPCK gene expression in HepG2 cells", Molecular and Cellular Endocrinology, vol. 332, pp. 125-133.
Park, S., Lee, J.H., Ha, M., Nam, J. & Kim, N. 2009, "miR-29 miRNAs activate p53 by targeting p85alpha and CDC42", Nature structural & molecular biology, vol. 16, no. 1, pp. 23-29.
Pekarsky, Y., Palamarchyk, A., Maximov, V., Efaninov, A., Nanzaryan, N., Santanam, U., Rassenti, L., Kipps, T., Croce, C. M. 2008, "Tcl1 functions as a transcriptional regulator and is directly involved in the pathogenesis of CLL", PNAS, vol. 105, no. 50, pp. 19643-19648.
Pekarsky, Y., Santanam, U., Cimmino, A. & et al. 2006, "Tcl1 Expression in Chronic Lymphocytic Leukemia Is Regulated by miR-29 and miR-181", Cancer research, vol. 66, pp. 11590-11593.
Roldo, C., Missiaglia, E., Hagan, J. P., Falconi, M., Capelli, P., Bersani, S., Calin, G. A., Voliana, S., Liu, C., Scarpa, A., Croce, C. M. 2006, "MicroRNA Expression Abnormalities in Pancreatic Endocrine and Acinar Features and Clinical Behavior", Journal of Clinical Oncology, vol. 24, no. 29, pp. 4677-4684.
Santanam, U., Zanesi, N., Efanov, A., Costinean, S., Palarchuck, A., Hagan, J.P., Volinia, S., Alder, H., Rassenti, L., Kipps, T., Croce, C.M. & Pekarsky, Y. 2010, "Chonic lymphocytic leukemia modeled in mouse by targeted miR-29 expression", PNAS, vol. 107, pp. 12210-12215.
Seeley, R., VanPutte, C., Regan, J. & Russo, A. 2011, Seelsy's Anatomy & Physiology, 9th edn, McGraw-Hill, New York.
Sengupta, S., Boon, J.A., Chen, I.H., Newton, M.A., Stanhope, S.A., Cheng, Y.J., Chen, C.J., Hildesheim, A., Sugden, B. & Ahlquist, P. 2008, "MicroRNA 29c is down-regulated in

Page 34 of 36
nasopharyngeal carcinomas, up-regulating mRNAs encoding extracellular matrix proteins", PNAS, vol. 15.
Sherr, C.J. 1996, "Cancer cell cycles", Science, New Series, vol. 274, no. 5293, pp. 1672-1677.
Shreenivasaiah, P.K., Kim, D.H. & Wang, E. 2010, "microRNA Regulation of Networks of Normal and Cancer Cells" in Cancer systems biology, ed. E. Wang, pp. 107-123.
Silber, J., Lim, D.A., Petritsch, C., Parsson, A.I., Maunakea, A.K., Yu, M., Vandenberg, S.R., Ginzinger, D.G., James, D., Costello, J.F., Berges, G., Weiss, W.A., Alvarez-Buylla, A. & Hogdson, J.G. 2008, "miR-124 and miR-137 inhibit proliferation of glioblastoma multiforme cells and induce differentiation of bran tumor stem cells", MBC Medicine, vol. 6, no. 14.
Skelly, A.H. 2006, "Type 2 Diabetes Mellitus", Nursing clinics of north america, vol. 41, pp. 531-547.
Srinivasan, K. & Ramarao, P. 2007, "Animal models in type 2 diabetes research: An overview", The Indian Journal of Medical Research, vol. 125, pp. 451-472.
Subramanian, S., Lui, W., Lee, C. H., Espinosa, I., Nielsen, T. O., Heinrinch, N. C., Corless, C.L., Fire, A. Z., van de Rijn, M.2008, "MicroRNA expression signature of human sarcomas", Oncogene, vol. 27, pp. 2015-2026.
Ueda, T., Volinia, S., Okumura, H., Shimizu, M., Taccioli, C., Rossi, S., Alder, H., Liu, C., oue N., Yasui, W., Yoshida, K., Sasaki, H., Nomura, S., Seto, Y., Kaminishi, M., Calin, G.A. & Croce, C.M. 2010, "Relation between microRNA expression and progression and prognosis of gastric cancer: a microRNA expression analysis", Lancet, vol. 11, pp. 136-146.
Vermeulen, K., Bockstacle, D.R.V. & Berneman, Z.N. 2003, "The cell cycle: a review of regulation, deregulation and therapeutic targets in cancer", Cell prolifferation, vol. 36, pp. 131-149.
Wang, H., Garzon, R., Sun, H., Ladner, K.J., Singh, r., Dahlman, J., Cheng, A., Hall, B.M., Qualman, S.J., Chandler, D.S., Crose, C.M. & Cuttridge, D.C. 2008, "NF-KB-YY1-miR-29 Regulatory Circuitry in Skeletal Myogenesis and Rhabdomyosarcoma", Cancer cell, vol. 4, pp. 369-381.
Wang, H., Hertlein, E., Bakkar, N., Sun N., Acharyya, S., Wang, J., Carathers, M., Davuluri, R. & Guttridge, D.C. 2007, "NF-kB Regulation of YY1 Inhibits Skeletal Myogenesis through Transcriptional Silencing of Myofibrillar Genes", Molecular and Cellular Biology, vol. 27, no. 12, pp. 4374-4387.
Whitehead, J. P, Clark, S. F., Ursø, B., James, D. E. 2000, "Signalling through the insulin receptor", Current Opinion in Cell Biology, vol. 12, pp. 222-228.

Page 35 of 36
Winbanks, C.E., Wang, B., Beyer, C., Koh, P., White, L. & Kantharidis, P. 2011, "TGF-beta Regulates miR-206 and miR-29 to Control Myogenic Differentiation through Regulation of HDAC4", Journal of Biolological Chemistry, vol. 286, no. 16, pp. 13805-13814.
Winter, J. & Diederichs, S. 2011, "MicroRNA Biogenesis and Cancer" in MicroRNA and Cancer: Methods and Protocols, Methods in Molecular Biology, ed. W. Wu, 676th edn, Humana Press, , pp. 3-22.
Xiong, Y., Fang, J., Yun, J., Yang, J., Zhang, Y., Jia, W. & Zhuang, S. 2010, "Effects of MicroRNA-29 on Apoptosis, Tumorigenicity, and Prognosis of Hepatocellular Carcinoma", Hepatology, vol. 51, no. 3, pp. 836-845.
Xu, H., Cheung, I.Y., Guo, H. & et al. 2009, "MicroRNA miR-29 Modulates Expression of Immunoinhibitory Molecule B7-H3: Potential Implkications for Immune Based Therapy of Human Solid Tumors", Cancer Research, vol. 69, pp. 6275-6281.
Yanaihara, N., Caplen, N., Bowman, E., Seike, M., Kumamoto, K., yi, M., Stephens, R.M., Okamoto, A., Yokota, J., Tanaka, T., Calin, G.A., Liu, C., Croce, C.M. & Harris, C.C. 2006, "Unique microRNA molecular profiles in lung cancer diagnosis and prognosis", Cancer Cell, vol. 9, pp. 189-198.
Zanette, D. L., Rivadavia, F., Molfetta, G. A., Barbuzano, F. G., Proto-Siqueira, R., Falcão, R. P., Zago, M. A., Silva-Jr., W. A. 2007, "miRNA expression profiles in chronic lymphocytic and acute lymphocytic leukemia", Brazilian Journal of Medical and Biological Research, vol. 40, pp. 1435-1440.
Zhang, Y., Li, M., Wang, H., Fisher, W.E., Lin, P.H., Yao, Q. & Chen, C. 2009, "Profiling of 95 MicroRNAs in pancreatic Cancer Cell Lines and Surgical Specimens By Real-Time PCR Analysis", World Journal of Surgery, vol. 33, pp. 698-709.
Zhao, J., Lin, J., Lwin, T., Yang, H., Guo, J., Kong, W., Desseureaul, S., Moscinski, L.C., Rezania, D., Dalton, W.S., Sotomayor, E., Tao, J. & Cheng, J.Q. 2010, "microRNA expression profile and identification of miR-29 as a prognostic marker and pathogenic factor by targeting CDK6 in mantle", Blood, vol. 115, no. 13, pp. 2630-2639.

Page 36 of 36
Appendix 1
Directory of targets of miR-29
Abb
revi
atio
ns:
Luc.
ass
ay =
Luc
ifera
se
assa
y
N.V
.M. =
No
Val
idat
ion
Met
hod
was
use
d


















