
MESTRADO EM MICROBIOLOGIA APLICADA -...
53
UNIVERSIDADE DE LISBOA FACULDADE DE CIÊNCIAS DEPARTAMENTO DE BIOLOGIA VEGETAL EMERGÊNCIA DE CARBAPENEMASES EM KLEBSIELLA PNEUMONIAE: O DESAFIO DE BACTÉRIAS MULTIRRESISTENTES E VIRULENTAS Filipa Alexandra Gomes Calisto MESTRADO EM MICROBIOLOGIA APLICADA 2011
Transcript of MESTRADO EM MICROBIOLOGIA APLICADA -...

UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA VEGETAL
EMERGÊNCIA DE CARBAPENEMASES EM KLEBSIELLA
PNEUMONIAE: O DESAFIO DE BACTÉRIAS
MULTIRRESISTENTES E VIRULENTAS
Filipa Alexandra Gomes Calisto
MESTRADO EM MICROBIOLOGIA APLICADA
2011

UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA VEGETAL
EMERGÊNCIA DE CARBAPENEMASES EM KLEBSIELLA
PNEUMONIAE: O DESAFIO DE BACTÉRIAS
MULTIRRESISTENTES E VIRULENTAS
Dissertação orientada pelas Professoras Doutoras
Aida Duarte (Faculdade de Farmácia da Universidade de Lisboa)
Ana Reis (Faculdade de Ciências da Universidade de Lisboa)
Filipa Alexandra Gomes Calisto
MESTRADO EM MICROBIOLOGIA APLICADA
2011

EMERGÊNCIA DE CARBAPENEMASES EM KLEBSIELLA
PNEUMONIAE: O DESAFIO DE BACTÉRIAS
MULTIRRESISTENTES E VIRULENTAS
Filipa Alexandra Gomes Calisto
DISSERTAÇÃO DE MESTRADO
2011
Esta dissertação foi realizada no Laboratório de Microbiologia do Instituto de
Investigação de Ciências Médicas e Farmacêuticas (iMed.UL) sob a orientação
directa da Prof.ª Doutora Aida Duarte no âmbito do Mestrado em Microbiologia
Aplicada da Faculdade de Ciências da Universidade de Lisboa.
A Prof.ª Doutora Ana Reis foi a orientadora interna designada no âmbito do
Mestrado em Microbiologia Aplicada da Faculdade de Ciências da
Universidade de Lisboa.

IV
AGRADECIMENTOS
A dificuldade da realização duma tese depende das adversidades, mas também dos apoios
que vão surgindo, ao longo do trabalho. Neste sentido, gostaria de expressar a minha gratidão
a todos aqueles que de alguma forma contribuíram para a realização do presente trabalho.
À Prof.ª Dr.ª Aida Duarte, pela amizade e orientação. Por ter confiado no meu trabalho e por
permitir o meu crescimento como microbiologista. Fica aqui a minha admiração, consideração e
respeito.
À Prof.ª Dr.ª Ana Reis pelo apoio, orientação, disponibilidade e interesse demonstrados.
Ao Instituto de Investigação de Ciências Médicas e Farmacêuticas da Faculdade de Farmácia
da Universidade de Lisboa (iMed.UL) pela disponibilização dos meios necessários para a
realização deste trabalho.
À Ana Narciso e Ana Eusébio pela ajuda e momentos de descontracção.
À Alessa e ao Flávio pelos momentos cinematográficos.
Agradeço aos meus amigos que me apoiam constantemente, pelo companheirismo e amizade
inestimáveis.
À minha família um agradecimento muito especial. Por todo o seu apoio sem limites, por todo o
seu amor e carinho, por tudo aquilo que representam.
Ao Filipe, por ser o meu companheiro de todos os momentos, pela partilha, porque sem ele eu
seria diferente.

V
TRABALHOS APRESENTADOS COM BASE NESTA DISSERTAÇÃO
Publicações em actas de encontros científicos:
F. Calisto, A. Narciso, A. Duarte. (2011) "Como identificar uma carbapenemase?". Revista Portuguesa
de Farmácia. vol.LII (nº5) Suplemento, P-D-09
F. Calisto, A. Narciso, A. Duarte. (2011) "Disseminação da resistência ao imipenemo em
enterobactérias". Revista Portuguesa de Farmácia. vol.LII (nº5) Suplemento, P-D-010
F. Calisto, L. Lito, J. Melo-Cristino, A. Duarte. (2011) "IncF plasmids responsible by dissemination of
blaKPC gene among Enterobacteriaceae". BMC Proceedings vol.5, suppl.6, P132
C. Caneiras, F. Calisto, A. Narciso, S. Arriaga, J. Melo-Cristino, A. Duarte. (2011) "Resistance and
virulence characterization of CTX-M15 and KPC-3 Klebsiella pneumoniae clinical isolates". Clin.
Microbiol. Infect. vol.17, suppl.s4, S270 (Impact Factor: 4.784)
Comunicações orais por convite:
"Plasmídeos como veículos de disseminação de genes de resistência e virulência". F. Calisto, A.
Narciso, C. Caneiras, A. Duarte. 4º Congresso Pandemias na Era da Globalização, Coimbra, Portugal.
16 - 18 Junho 2011.
Comunicações em painel "poster":
"Disseminação da resistência ao imipenemo em enterobactérias". F. Calisto, A. Narciso, A. Duarte.
COIFFA, Lisboa, Portugal. 21 - 23 Junho 2011.
"Como identificar uma carbapenemase?". F. Calisto, A. Narciso, A. Duarte. COIFFA, Lisboa, Portugal.
21 - 23 Junho 2011. F. Calisto, A. Narciso, A. Duarte. COIFFA, Lisboa, Portugal. 21 - 23 Junho 2011.
"Plasmídeos como veículos de disseminação de genes de resistência e virulência". F. Calisto, A.
Narciso, C. Caneiras, A. Duarte. 4º Congresso Pandemias na Era da Globalização, Coimbra, Portugal.
16 - 18 Junho 2011.
"KPC-3 em Portugal e no Mundo". F. Calisto, A. Narciso, J. Delgado, A. Duarte. 4º Congresso
Pandemias na Era da Globalização, Coimbra, Portugal. 16 - 18 Junho 2011.
"IncF plasmids responsible by dissemination of blaKPC gene among Enterobacteriaceae". F. Calisto, L.
Lito, J. Melo-Cristino, A. Duarte. 1st ICPIC - International Conference on Prevention & Infection Control,
Geneva, Switzerland, 29 June – 2 July 2011
"Resistance and virulence characterisation of CTX-M15 and KPC-3 Klebsiella pneumoniae clinical
isolates". C. Caneiras, F. Calisto, A. Narciso, S. Arriaga, J. Melo Cristino, A. Duarte. 21st ECCMID -
European Congress of Clinical Microbiology and Infectious Diseases, Milan, Italy, 7 - 10 May 2011,
P1043.

VI
RESUMO
A emergência e disseminação global de Enterobacteriaceae resistentes aos carbapenemos é
uma ameaça à saúde pública, pois estão associadas a altas taxas de morbilidade e
mortalidade. Estirpes produtoras de carbapenemases são caracterizadas por resistência a
todos os antibióticos β-lactâmicos, fluoroquinolonas e aminoglicosídeos. Este estudo tem como
objectivo identificar e caracterizar mecanismos de resistência aos carbapenemos.
Vinte e seis isolados clínicos de Klebsiella pneumoniae, K. oxytoca, Enterobacter aerogenes,
Escherichia coli e Citrobacter freundii, com resistência ou reduzida susceptibilidade aos
carbapenemos, identificados em 2010 de dezoito doentes do Hospital de Santa Maria, foram
estudados em termos de perfil genético, genes de resistência e do ambiente genético em que
se inserem, presença/ausência de porinas na membrana externa, plasmídeos transportados e
factores de virulência.
Entre os vinte e seis isolados clínicos em estudo, dezoito foram positivos para a presença da
carbapenemase KPC-3. Associada a esta carbapenemase foram detectados as β-lactamases
TEM-1 e SHV-1, as ESBLs SHV-12 e SHV-35 e a cefalosporinase DHA-1. A ausência das
porinas OmpK35 e OmpK36 da membrana externa aumentam a resistência dos isolados para
os carbapenemos e cefalosporinas.
A resistência às quinolonas em alguns isolados deve-se há presença dos genes aac(6’)-Ib-cr e
qnr, à perda da porina OmpK36 e/ou a mutações na DNA topoisomerase.
O gene blaKPC-3 destes isolados está associado ao transposão Tn4401-like com capacidade de
transposição replicativa que foi detectado no plasmídeo do grupo de incompatibilidade IncF.
Os isolados de K. pneumoniae estudados são predominantemente do serótipo K2 e possuem
os genes fimH e/ou mrkD, conferindo a capacidade de aderir às células do hospedeiro.

VII
ABSTRACT
The emergence and global spread of Enterobacteriaceae carbapenem-resistant is a threat to
public health because they are associated with high morbidity and mortality. Strains producing
carbapenemases are characterized by resistance to all β-lactams, aminoglycosides and
fluoroquinolones. This study aims is to identify and characterize mechanisms of resistance to
carbapenems.
Twenty-six clinical isolates of Klebsiella pneumoniae, K. oxytoca, Enterobacter aerogenes,
Escherichia coli and Citrobacter freundii, with reduced susceptibility or resistance to
carbapenems, identified in 2010 in eighteen patients of the Hospital of Santa Maria, were
studied in terms of genetic profile, resistance genes and genetic environment in which they
operate, presence/absence of porins in outer membrane, carried plasmids by each on and
virulence factors.
Among the twenty-six clinical isolates under study, with resistance or reduced susceptibility to
carbapenems, eighteen were positive for the presence of KPC-3 carbapenemase. Associated
with this carbapenemases were detected the TEM-1 and SHV-1 β-lactamases, the SHV-12 and
SHV-35 ESBLs and DHA-1 cephalosporinase. The absence of porins OmpK35 OmpK36 in
outer membrane, increase the resistance of the isolates to cephalosporins and carbapenems.
The quinolone resistance in some isolates is given by the genes aac (6 ')-Ib-cr and qnr, loss of
porin OmpK36 and/or mutations in DNA topoisomerase.
blaKPC-3 gene of these isolates is associated with the transposon Tn4401-like with transposition
replicative capacity and this transposon was detected in the plasmid incompatibility group IncF.
The isolates of K. pneumoniae studied are predominantly K2 serotype and have the fimH and/or
mrkD genes, which giving the ability to adhere to host cells.

VIII
ÍNDICE
AGRADECIMENTOS ........................................................................................................................ IV
RESUMO ........................................................................................................................................ VI
ABSTRACT .................................................................................................................................... VII
1. INTRODUÇÃO .......................................................................................................................... 1
1.1 Resistência aos β-lactâmicos ........................................................................................ 1
1.1.1 Diminuição da acumulação intracelular do antibiótico .......................................... 2
1.1.2 Modificação do local alvo do antibiótico ................................................................ 3
1.1.3 Produção de β-lactamases .................................................................................... 3
1.2 Disseminação de genes de resistência ......................................................................... 5
1.2.1 Klebsiella pneumoniae carbapenemase ................................................................... 6
1.3 Klebsiella spp. ............................................................................................................... 7
2. OBJECTIVOS ........................................................................................................................... 9
3. MATERIAL E MÉTODOS ......................................................................................................... 10
3.1 Caracterização fenotípica dos isolados ...................................................................... 10
Determinação do perfil de susceptibilidade aos antibióticos .............................................. 10
3.2 Caracterização genotípica dos isolados...................................................................... 11
Extracção de DNA ............................................................................................................... 11
Tipificação dos isolados bacterianos................................................................................... 11
Multilocus Sequence Typing (MLST) .................................................................................. 11
Identificação dos determinantes genéticos de resistência .................................................. 12
Determinação do ambiente genético dos genes de resistência ......................................... 13
Identificação dos plasmídeos transportadores de β-lactamases ........................................ 14
Identificação dos determinantes genéticos de virulência .................................................... 14
Sequenciação e análise das sequências nucleotídicas ...................................................... 15
4. RESULTADOS E DISCUSSÃO ................................................................................................. 16
4.1 Susceptibilidade aos antibióticos ................................................................................ 17
4.2 Tipificação dos isolados bacterianos........................................................................... 19
4.4 Determinantes genéticos de resistência ..................................................................... 21
4.5 Disseminação de genes de resistência ....................................................................... 26
4.6 Determinantes de virulência em Klebsiella spp. ......................................................... 30
5. CONCLUSÃO ......................................................................................................................... 32
6. REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................................ 33

1
1. INTRODUÇÃO
As infecções associadas aos cuidados de saúde são adquiridas após a entrada do paciente
num hospital em consequência dos cuidados e procedimentos de saúde prestados e que
podem, também, afectar os profissionais de saúde durante o exercício da sua actividade. Estas
infecções dificultam o tratamento adequado de doentes em todo o mundo e são uma causa
importante de morbilidade e mortalidade, bem como do consumo acrescido de recursos quer
hospitalares, quer da comunidade.1 As principais bactérias de Gram-negativo, responsáveis por
infecções hospitalares, pertencem à família Enterobacteriaceae, nomeadamente Escherichia
coli, Klebsiella spp. e Enterobacter spp.. O tratamento destas infecções bacterianas é
extremamente complicado devido à capacidade das bactérias para desenvolverem resistência
a antibióticos.1; 2
Os antibióticos têm a capacidade de impedir a multiplicação de bactérias, interferindo na
síntese proteica, ou de as destruir, sem ter efeitos tóxicos para as células humanas.
Conhecem-se diversos mecanismos de acção antibacteriana: os β-lactâmicos (penicilinas,
cefalosporinas, monobactâmicos e carbapenemos) inibem a síntese de peptidoglicano da
parede celular; os aminoglicosídeos, tetraciclinas e macrólidos ligam-se às subunidades
ribossomais inibindo a síntese proteica; as quinolonas impedem a replicação do DNA por
interacção com a DNA girase ou a topoisomerase IV; e as sulfonamidas inibem a síntese de
folato, essencial na formação de purinas e alguns aminoácidos.3
1.1 Resistência aos β-lactâmicos
Os antibióticos β-lactâmicos são o grupo de antibióticos mais importantes, dada a sua eficácia
terapêutica e a sua baixa toxicidade para os animais, incluindo o Homem.3 Nos últimos anos a
emergência de bactérias patogénicas resistentes aos β-lactâmicos em ambientes hospitalares
tem-se tornado um problema mundial.4
As bactérias de Gram-negativo possuem diversos mecanismos de resistência que afectam as
diferentes classes de antibióticos (Figura 1). Estas bactérias podem ter: (i) diminuição da
expressão de porinas da membrana externa (OMPs) e bombas de efluxo que diminuem a
acumulação intracelular do antibiótico; (ii) mutações que alteram o sítio activo de ligação do
antibiótico (PBP – Penicillin Binding Protein), diminuindo a afinidade com o antibiótico; (iii)
genes que codificam enzimas, como as β-lactamases, que destroem o antibiótico.5, 6
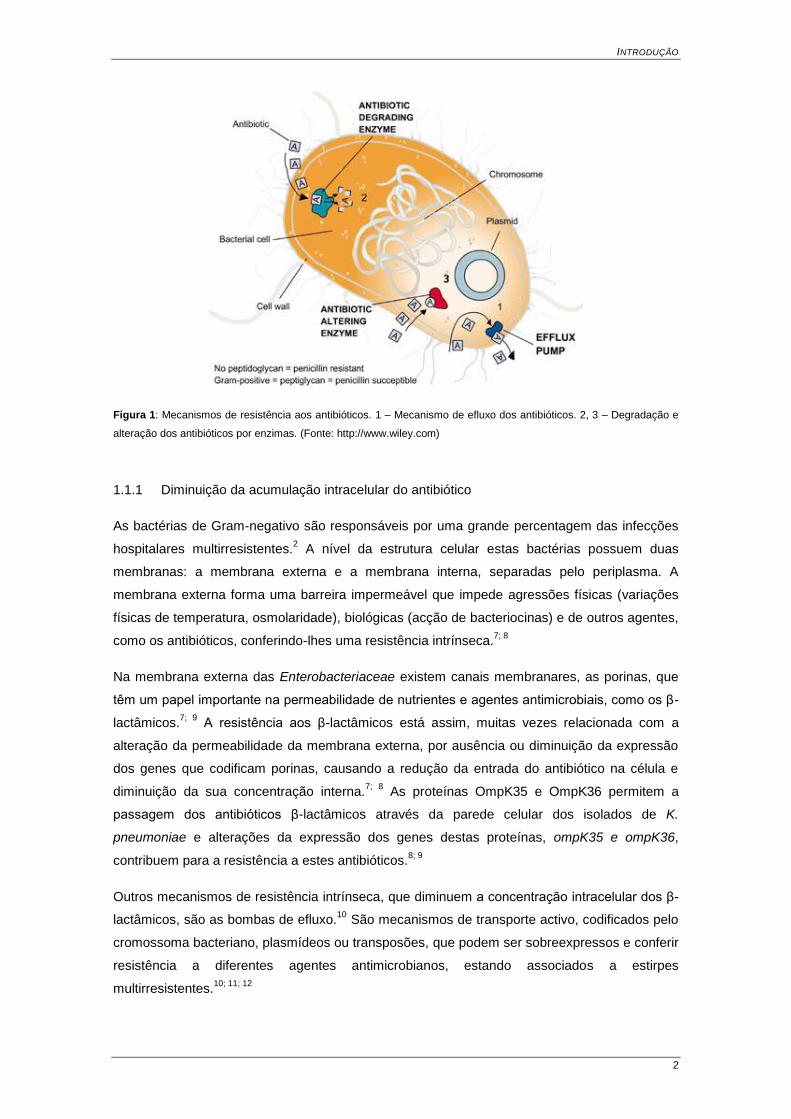
INTRODUÇÃO
2
Figura 1: Mecanismos de resistência aos antibióticos. 1 – Mecanismo de efluxo dos antibióticos. 2, 3 – Degradação e
alteração dos antibióticos por enzimas. (Fonte: http://www.wiley.com)
1.1.1 Diminuição da acumulação intracelular do antibiótico
As bactérias de Gram-negativo são responsáveis por uma grande percentagem das infecções
hospitalares multirresistentes.2 A nível da estrutura celular estas bactérias possuem duas
membranas: a membrana externa e a membrana interna, separadas pelo periplasma. A
membrana externa forma uma barreira impermeável que impede agressões físicas (variações
físicas de temperatura, osmolaridade), biológicas (acção de bacteriocinas) e de outros agentes,
como os antibióticos, conferindo-lhes uma resistência intrínseca.7; 8
Na membrana externa das Enterobacteriaceae existem canais membranares, as porinas, que
têm um papel importante na permeabilidade de nutrientes e agentes antimicrobiais, como os β-
lactâmicos.7; 9
A resistência aos β-lactâmicos está assim, muitas vezes relacionada com a
alteração da permeabilidade da membrana externa, por ausência ou diminuição da expressão
dos genes que codificam porinas, causando a redução da entrada do antibiótico na célula e
diminuição da sua concentração interna.7; 8
As proteínas OmpK35 e OmpK36 permitem a
passagem dos antibióticos β-lactâmicos através da parede celular dos isolados de K.
pneumoniae e alterações da expressão dos genes destas proteínas, ompK35 e ompK36,
contribuem para a resistência a estes antibióticos.8; 9
Outros mecanismos de resistência intrínseca, que diminuem a concentração intracelular dos β-
lactâmicos, são as bombas de efluxo.10
São mecanismos de transporte activo, codificados pelo
cromossoma bacteriano, plasmídeos ou transposões, que podem ser sobreexpressos e conferir
resistência a diferentes agentes antimicrobianos, estando associados a estirpes
multirresistentes.10; 11; 12

INTRODUÇÃO
3
1.1.2 Modificação do local alvo do antibiótico
Relativamente à parede celular da célula bacteriana, os β-lactâmicos ligam-se
irreversivelmente às D-D-carboxitranspeptidases (PBPs), impedindo o estabelecimento de
pontes interpeptídicas entre cadeias peptídicas vizinhas do peptidoglicano em crescimento.
Assim, mutações nos genes produtores de PBPs, recombinações homólogas entre genes de
PBPs, que não alterem a função das proteínas, ou a síntese de novos PBPs sem grande
afinidade para os β-lactâmicos conferem às estirpes resistência a estes antibióticos.3; 12
1.1.3 Produção de β-lactamases
As β-lactamases, expressas por genes plasmídicos ou cromossomais, hidrolisam o anel β-
lactâmico, sendo a principal causa de resistência bacteriana a estes antibióticos.13
Podem ser
divididas em quatro classes de acordo com a sua sequência de aminoácidos: as classes A (β-
lactamases de espectro alargado), C (cefalosporinas) e D (oxacilinases) compreendem as
serino-β-lactamases (serina no centro activo) e a classe B (metalo-β-lactamases) compreende
as β-lactamases com iões de zinco no centro activo.14
A primeira β-lactamase, uma cefalosporinase cromossómica AmpC, inibida por cloxacilina e
ácido borónico,15; 16
foi identificada em Escherichia coli, em 1940, e conferia resistência à
penicilina.17
Em 1963, foi descrita a primeira β-lactamase plasmídica, TEM-1, identificada numa
estirpe de E. coli. 18,19
A presença de plasmídeos e transposões facilitaram a disseminação da
β-lactamase TEM-1 para outras espécies bacterianas e em poucos anos espalhou-se por todo
o mundo, podendo actualmente ser identificada, principalmente, em diferentes espécies de
Enterobacteriaceae.20
A β-lactamase TEM-2, derivada da TEM-1 por alteração de um
aminoácido, surgiu pouco tempo depois.21
O gene blaSHV-1, que codifica para uma β-lactamase com actividade contra penicilinas, foi
descrito pela primeira vez no cromossoma de K. pneumoniae.22
Em E. coli O gene blaSHV-1 é
normalmente encontrado em plasmídeos.20
A introdução na terapêutica das cefalosporinas de 3ª geração, como a cefotaxima e
ceftazidima, no início dos anos 80, em resposta à elevada prevalência de β-lactamases em
determinados microrganismos, como a TEM-1 e SHV-1 em E. coli e K. pneumoniae, levou a
surgirem genes blaSHV e blaTEM mutados que codificavam β-lactamases de espectro alargado
(Extended-Spectrum β-Lactamases – ESBLs).20
A 1ª destas enzimas, SHV-2, foi isolada de K.
ozaenae na Alemanha.23
A maioria das ESBLs pode ser dividida em três grupos: TEM, SHV e
CTX-M.24
A 1ª β-lactamase da família CTX-M, CTX-M-1, isolada de K. pneumoniae, hidrolisava
preferencialmente a cefotaxima.25
São as ESBLs geneticamente mais heterogéneas e

INTRODUÇÃO
4
prevalentes no globo,26
sendo a β-lactamase CTX-M-15, descrita em 2000 na Índia, a que tem
maior sucesso de disseminação desta família.27
Outras enzimas da família ESBL são as β-lactamases OXA, que conferem resistência à
ampicilina e cefalotina. Estão descritas como sendo as β-lactamases plasmídicas mais
prevalentes no fim dos anos 70 e início dos anos 80.28
A utilização de cefalosporinas de 3ª geração para tratamento de infecções causadas por
Citrobacter spp. e Enterobacter spp. levou à mobilização das β-lactamases AmpC para
plasmídeos.29
Assim como as ESBLs, as β-lactamases AmpC plasmídicas actuam sobre um
amplo número de antibióticos, como a cefotaxima e a ceftazidima.30
A primeira cefalosporinase
AmpC plasmídica, CMY-1, surgiu na Coreia em 1988.31
Posteriormente foi identificada em K.
pneumoniae, em 1990, a β-lactamase CMY-2, sendo actualmente, a β-lactamase AmpC mais
prevalente e globalmente distribuída.32
Em 1992, é identificada a β-lactamase DHA-1 em
Salmonella enteritidis.33
O aparecimento de Enterobacteriaceae multirresistentes produtoras de ESBLs e β-lactamases
AmpC plasmídicas levou à introdução de outros antibióticos, como o imipenemo e
meropenemo, na quimioterapia bacteriana. A produção de carbapenemases, β-lactamases com
capacidade de hidrolisar os carbapenemos e outros β-lactâmicos, faz emergir
Enterobacteriaceae resistentes aos carbapenemos.34
As carbapenemases pertencem às
classes moleculares: A, que inclui as famílias das β-lactamases KPC, IMI, SME, NMC e GES,
inibidas por ácido borónico;35
classe B, metalo-β-lactamases (MBL), que incluem as famílias
das β-lactamases VIM, IMP, SPM e NDM e que são inibidas por EDTA;16
e classe D, que inclui
as carbapenemases OXA, encontradas maioritariamente em Acinetobacter baumannii.16; 36
A primeira MBL cromossómica foi descrita em 1966, num isolado de Bacillus cereus,37
no
entanto só no final dos anos 80 é que estas enzimas ganham importância clinica com a
detecção no Japão da primeira MBL plasmídica, a IMP-1.38
Na Europa, em 1997, é identificada
a VIM-139
com uma elevada afinidade para os carbapenemos. Estas enzimas são
predominantemente encontradas em Klebsiella spp. e Enterobacter spp.40
A MBL SPM-1 foi
identificada posteriormente numa estirpe de Pseudomonas aeruginosa, no fim do século XXI,41
e encontra-se actualmente confinada ao Brasil.40
Recentemente foi identificada em K.
pneumoniae a enzima NDM-1,42
com um enorme potencial de disseminação.43
Na Índia
encontra-se amplamente disseminada pelas Enterobacteriaceae.42
A disseminação de genes
que codificam para MBL entre bactérias patogénicas de Gram-negativo é mediada por
elementos móveis de DNA associados a transposões e plasmídeos.44
As carbapenemases cromossómicas de classe A das famílias IMI, SME e NMC apenas são
encontradas num baixo número de isolados de Serratia e Enterobacter. Conferem resistência
aos carbapenemos mas são sensíveis às cefalosporinas de 3ª geração.45
As β-lactamases da
família GES, localizadas em plasmídeos, foram inicialmente classificadas como ESBL,46
mas

INTRODUÇÃO
5
em 2001 com o aparecimento da GES-2 e a hidrólise do imipenemo foram classificadas como
carbapenemases.47
Klebsiella pneumoniae carbapenemase (KPC), identificada nos Estados Unidos da América,
em 1996,48
é a β-lactamase de classe A mais frequente em K. pneumoniae.34
Após a
identificação da KPC-1 surgiram várias publicações com uma nova variante, a KPC-2.49; 50; 51
Uma única alteração num aminoácido da β-lactamase KPC-2 gerou a β-lactamase KPC-3.52
A primeira OXA β-lactamase com actividade hidrolítica para os carbapenemos, ARI-1, foi
descrita em Acinetobacter baumannii, em 1993.53
Estas carbapenemases são mais frequentes
em A. baumannii, tendo sido identificada em 2004 num isolado de K. pneumoniae a OXA-48.54
Recentemente identificou-se em K. pneumoniae as variantes OXA-163 e OXA-181.55; 56
1.2 Disseminação de genes de resistência
A mobilidade dos elementos genéticos bacterianos torna possível a disseminação dos genes
de resistência aos diferentes antibióticos e a sua acumulação em alguns clones
multirresistentes. São determinantes para este fenómeno a existência de plasmídeos,
transposões, integrões e cassetes genéticas de resistência nas células bacterianas.3
Os plasmídeos, pequenas moléculas circulares de DNA extracromossómico que se replicam
autonomamente, não possuem genes essenciais para o crescimento e multiplicação bacteriana
mas têm uma variedade de genes que permite a iniciação e controlo da replicação e que
fornecem à célula vantagem adicional em determinadas condições ambientais, como a
resistência aos antibióticos.3; 57
Frequentemente as bactérias patogénicas possuem plasmídeos
conjugativos que transportam genes que conferem simultaneamente resistência a diferentes
antibióticos e que são transferidos vertical e horizontalmente entre bactérias.58
Vários genes de resistência também estão localizados em transposões e estes por sua vez em
plasmídeos. Os transposões são sistemas genéticos que se podem mover dentro do
cromossoma, do plasmídeo para o cromossoma e vice-versa e entre plasmídeos. Estão
flanqueados por sequências de inserção (IS) que são essenciais para a transposição deste
elemento. Associados aos transposões podem estar integrões e cassete genéticas. 3; 57; 58
Os integrões são compostos por duas regiões conservadas e uma região variável. A região
conservada 5’ (5’-CS) contém o gene intI, que codifica uma integrase, um local de
recombinação (attI) e uma região promotora P1-P2 que controla a transcrição de genes de
resistência que possam estar integrados na região variável; a região conservada 3’ (3’-CS)
codifica os genes de resistência a sulfonamidas (suII), a compostos quaternários de amónio e
brometo de etídio (qacEΔ1) e a ORF5 (Figura 2).59; 60

INTRODUÇÃO
6
As cassetes genéticas são pequenos elementos circulares de DNA com uma região
codificante, onde estão os genes de resistência, que podem inserir-se por recombinação num
local attI de um integrão.3; 57; 58
Figura 2: Representação esquemática da estrutura de um integrão. Os genes e a sua orientação de transcrição são
representados por setas horizontais. Os quadrados vermelhos representam promotores. O rectângulo branco delineado
por azul representa a zona de recombinação. O rectângulo branco delineado por preto representa uma cassete
genética. (Fonte: Adaptado de Levesque et al., 1995)
1.2.1 Klebsiella pneumoniae carbapenemase
A carbapenemase KPC, que confere resistência aos carbapenemos, cefalosporinas e
aztreonam,48
é responsável por infecções nosocomiais severas,61
e foi identifica nos Estados
Unidos da América.33; 47
Registou-se uma rápida disseminação de isolados produtores desta
carbapenemase por todo o mundo, tendo sido identificada em Portugal em 2009.62
A mobilidade do gene blaKPC está associada a plasmídeos com diferentes tamanhos e grupos
de incompatibilidade63; 64; 65
e ao transposão Tn4401, um transposão do tipo Tn3 com
aproximadamente 10 kb (Figura 3).66
O transposão Tn4401 possui os genes que codificam
para a transposase (tnpA) e para a resolvase (tnpR) e duas sequências de inserção, ISKpn6 e
ISKpn7, e está na origem da disseminação desta β-lactamase, assim como outros transposões
com estrutura similar (Tn4401-like).66; 67; 68
Este transposão tem uma elevada taxa de
transposição replicativa.69
Figura 3: Representação esquemática do transposão Tn4401. Os genes e a sua orientação de transcrição são
representados por setas horizontais. O transposão é delimitado por duas sequências invertidas representadas por
triângulos cinzentos. Os triângulos brancos representam as sequências de inserção do ISKpn7 e ISKpn6. A linha
vertical representa uma zona de deleções. (Fonte: Adaptado de Naas et al., 2008)
P1 P2 P P
P
intI
qacEΔ1 sulI orf5
attI
tnpR tnpA IstA IstB KPC tnpA
ISKpn7 ISKpn6

INTRODUÇÃO
7
1.3 Klebsiella spp.
As bactérias do género Klebsiella são uma causa importante de infecções hospitalares e
podem ser também adquiridas na comunidade, sendo as mais comuns as infecções urinárias,
as infecções respiratórias e as bacterémias causadas por K. pneumoniae.70 A emergência e
disseminação de estirpes hospitalares de K. pneumoniae, resistentes aos principais antibióticos
usados como opção terapêutica, realçam a importância de estudar os mecanismos pelos quais
esta bactéria interage com o hospedeiro.71
K. pneumoniae desenvolve, geralmente, cápsulas proeminentes compostas por polissacáridos.
As subunidades capsulares podem ser classificadas em 77 serótipos.70
As estirpes com
antigénios capsulares K1, K2, K4 e K5 são consideradas as mais virulentas, sendo menos
virulentas as estirpes com antigénios K6 e K superior a 6.71
Os serótipos K1 e K2 são os
predominantes entre os isolados clínicos humanos.70
O gene cromossomal magA, um gene de virulência associado a estirpes de K. pneumoniae
invasivas, codifica para a proteína MagA, específica do serótipo capsular K1, responsável pelo
fenótipo de hipermucoviscosidade e que está associada à resistência ao soro e à fagocitose.72
Alguns isolados de K. pneumoniae encapsulados possuem colónias mucóides de consistência
viscosa. É a proteína RmpA, codificada pelo gene plasmídico rmpA, que controla positivamente
a biossíntese do polissacárido capsular, e assim, o fenótipo mucóide.73
A capacidade da bactéria para aderir às superfícies dos tecidos do hospedeiro é importante no
desenvolvimento da infecção. As propriedades adesivas nas Enterobacteriaceae são mediadas
por diferentes tipos de fímbrias. As fímbrias, projecções filamentosas não flagelares na
superfície bacteriana, são constituídas por subunidades de pilina. Nos isolados de K.
pneumoniae predominam dois tipos de fímbrias: as do tipo 1 e as do tipo 3. As adesinas são
proteínas que permitem a aderência da bactéria às células epiteliais assim como a superfícies
abióticas, podendo ou não estar localizadas nas fímbrias.70
As fímbrias do tipo 1, codificadas
no operão fim, medeiam a adesão a receptores com a manose através de adesina FimH, como
mucosas ou células epiteliais do trato urinário, respiratório e intestinal. As fímbrias do tipo 3,
codificadas no operão mrk, são manose resistente e promovem a ligação às células
endoteliais, do epitélio do trato respiratório e células uropiteliais através da adesina MrkD. As
fímbrias do tipo 3 desempenham um papel crucial na formação de biofilmes.74
O crescimento das bactérias no tecido hospedeiro é limitado por mecanismos de defesa do
hospedeiro e pela quantidade de ferro disponível. No hospedeiro o ferro livre disponível para a
bactéria, factor essencial para o crescimento bacteriano, é extremamente baixo, pois encontra-
se ligado a proteínas intracelulares como a hemoglobina, ferritina e hemosiderina e a proteínas
extracelulares como a lactoferrina e transferrina.75
Muitas bactérias tentam garantir o seu
suprimento de ferro no hospedeiro secretando substâncias quelantes, os sideróforos, que
solubilizam e ajudam na importação do ferro requerido.70
O género Klebsiella sintetiza dois

INTRODUÇÃO
8
grupos químicos de sideróforos, os do tipo fenolato, como a enterobactina, e do tipo
hidroxaqmato, como a aerobactina. A enterobactina tem maior afinidade para o ferro que a
aerobactina, no entanto a aerobactina é mais virulenta devido à sua maior estabilidade e
solubilidade.76; 77; 78
Em 1985 um estudo de Albesa et al.,79
descreveu isolados de K. pneumoniae com actividade
hemolítica em Rabbit Bolld Agar mas sem actividade hemolítica para os eritrócitos humanos.
Posteriormente, vários estudos mostraram que esta hemolisina também tem um efeito
hemolítico em células de ovelhas, ratos e humanos.80; 81
No entanto, a hemolisina, codificada
pelo gene khe, e o seu papel como factor de virulência em K. pneumoniae não se encontram
plenamente estudados.82; 83

9
2. OBJECTIVOS
Este estudo teve como objectivo identificar e caracterizar genes de resistência aos
carbapenemos, o seu ambiente genético e outras resistências associadas, em bactérias
resistentes e com reduzida susceptibilidade aos carbapenemos, isoladas no Hospital Santa
Maria, em Lisboa.
De modo a atingir este objectivo:
• Avaliou-se a susceptibilidade dos isolados bacterianos a diferentes grupos de
antibióticos pelo método de difusão em agar Kirby-Bauer;
• Pesquisou-se genes de resistência e elementos móveis associados à resistência aos
antibióticos, pela técnica de PCR, com primers específicos ou desenhados no
laboratório. Efectuou-se a confirmação, por sequenciação, dos produtos de
amplificação com tamanho esperado;
• Pesquisou-se factores de virulência, nomeadamente adesinas fimbriais; hemolisinas,
sideróforos entre outros, específicos das espécies bacterianas identificadas, pela
técnica de PCR com primers específicos ou desenhados no laboratório. Efectuou-se a
confirmação, por sequenciação, dos produtos de amplificação com tamanho esperado;

10
3. MATERIAL E MÉTODOS
Este estudo incidiu sobre isolados de Klebsiella pneumoniae (n=16), Klebsiella oxytoca (n=5),
Enterobacter aerogenes (n=1), Escherichia coli (n=2) e Citrobacter freundii (n=2) identificados
em 2010 de vários produtos biológicos no Hospital de Santa Maria, em Lisboa. Recaiu
principalmente sobre K. pneumoniae por esta espécie ser um dos principais agentes de
infecção nocomial. Foram identificados pelo sistema automatizado do Laboratório de
Bacteriologia do hospital e seleccionados por apresentarem resistência ou reduzida
susceptibilidade aos carbapenemos.
3.1 Caracterização fenotípica dos isolados
Determinação do perfil de susceptibilidade aos antibióticos
Testou-se a susceptibilidade de cada isolado a antimicrobianos segundo o método de difusão
em disco Kirby-Bauer, em meio Müller-Hinton agar (MHA), com discos de amoxicilina/ácido
clavulânico (20/10 μg), cefoxitina (30 μg), cefotaxima (30 μg), ceftazidima (30 μg), cefepima (30
μg), aztreonamo (30 μg), imipenemo (10 μg), meropenemo (10 μg), ciprofloxacina (5 μg),
levofloxacina (5 μg), gentamicina (10 μg), tigeciclina (15 μg), nitrofurantoína (300 μg) e
fosfomicina (200 μg). Os halos de inibição foram interpretados de acordo com os critérios do
Clinical and Laboratory Standards Institute (CLSI) (2010),84
excepto para a tigeciclina. Os halos
para a tigeciclina foram interpretados de acordo com o European Committee on Antimicrobial
Susceptibility Testing.85
Com o objectivo de identificar mecanismos específicos de resistência, realizou-se um ensaio
com inibidores. Adicionou-se 10 µL de um agente inibidor a cada disco de antibiótico utilizado
no antibiograma. Um aumento de 5 mm, ou mais, no diâmetro da zona de inibição na presença
destes compostos, comparado com uma caixa padrão sem a adição destes agentes inibidores,
é considerado um teste positivo. A pesquisa de metalo-β-lactamases e carbapenemases do
grupo A foi realizada com a adição de EDTA (ácido etilenodiamino tetra-acético)86
e ácido
borónico (20 mg/mL),87
respectivamente. A pesquisa de cefalosporinases AmpC foi realizada
com a utilização de cloxacilina (10 mg/mL).88
A combinação de amoxicilina e ácido clavulânico
permite identificar a produção de β-lactamases de espectro alargado, pois a sua actividade é
inibida pelo ácido clavulânico.

MATERIAL E MÉTODOS
11
3.2 Caracterização genotípica dos isolados
Extracção de DNA
Extraiu-se o DNA total de cada isolado pelo método de boiling-centrifugation, que consiste na
lise de uma suspensão bacteriana densa, por fervura (94˚C, 10 minutos), seguida da remoção
dos resíduos celulares por centrifugação (12000 rpm, 2 minutos), recolhendo-se posteriormente
o sobrenadante.
Tipificação dos isolados bacterianos
De modo a proceder à caracterização molecular e avaliar a relação clonal entre os isolados
clínicos da mesma espécie recorreu-se ao método genotípico M13 e ERIC-PCR fingerprinting.
• M13-PCR fingerprinting
O perfil genético dos isolados foi obtido por M13-PCR fingerprinting, usando Ready-To-Go
RAPD Analysis Beads (GE Healthcare) para a reacção de PCR. Os primers e as condições de
PCR usados foram descritos por Grundmann et al., 1997.89
A análise dos perfis de
fingerprinting foi feita em gel de agarose a 2%, em TBE (1x) (Tris 45 mM, ácido borónico 45
mM, EDTA 1 mM), corado com brometo de etídio.
• ERIC-PCR fingerprinting
O perfil genético dos isolados foi obtido por ERIC-PCR fingerprinting, usando a Phusion Flash
Master Mix (Finnzymes) para a reacção de PCR. Os primers e as condições de PCR usados
foram previamente descritos por Adamus-Bialek et al., 2009.90
A análise dos perfis de
fingerprinting foi feita em gel de agarose a 2%, em TAE (0,5x) (Tris 40 mM, ácido acético 20
mM, EDTA 1 mM), corado com brometo de etídio.
Multilocus Sequence Typing (MLST)
A técnica de MLST, baseada na análise de fragmentos internos de genes housekeeping, foi
aplicada em três isolados de K. pneumoniae, correspondentes aos perfis de M13 e ERIC-PCR
fingerprinting mais representativos. Foram usadas Pure Taq Ready-To-Go PCR Beads (GE
Healthcare) para amplificar os sete genes housekeeping: gapA (gliceraldeído 3-fosfato
desidrogenase), infB (factor 2 de iniciação da tradução), mdh (malato desidrogenase), pgi
(fosfoglucose isomerase), phoE (fosfoporina E), rpoB (subunidade β da RNA polimerase B) e
tonB (transdutor energético periplasmático). Os primers e condições de PCR foram descritos
por Diancourt et al., 2005. Após a corrida em gel de agarose a 1%, em TAE (1x) e coloração
com brometo de etídio, os produtos foram purificados e sequenciados. As sequências
nucleotídicas obtidas permitiram definir os alelos de cada locus e, de acordo com a

MATERIAL E MÉTODOS
12
combinação de alelos de cada isolado, identificar os Sequence Types (STs) na base de dados
do Instituto Pasteur (www.pasteur.fr/mlst).
Identificação dos determinantes genéticos de resistência
Todas as reacções de PCR foram efectuadas num volume de 20 μL, usando a DyNAzymeTM
II
PCR Master MIX (Finnzymes), de acordo com o protocolo dos fabricantes. Posteriormente os
produtos de PCR foram resolvidos em gel de agarose a 1%, em TAE (1x), e corados com
GelRed (Biotium).
• Carbapenemases
A presença de carbapenemases, sugeridas pela resistência ou baixa susceptibilidade aos
carbapenemos, foi pesquisada através de reacções de PCR utilizando primers direccionados
para os genes blaKPC,48
blaNDM, blaIMP,91
blaOXA54
e blaVIM.92
Os primers para o gene blaNDM foram
desenhados no laboratório de acordo com as sequências disponíveis no Genbank (AB604954)
(Tabela 1).
• β-lactamases
Para além dos genes que codificam para carbapenemases, foram pesquisados os genes de
resistência blaTEM, blaSHV,93
blaCTX-M,94
blaDHA,95
e blaCMY.96
Os primers para os genes blaTEM e
blaCTX-M-2 foram desenhados no laboratório de acordo com as sequências disponíveis no
Genbank (HM749966 e DQ125241). Os primers desenhados e as condições de PCR utilizadas
estão descritos na Tabela 1.
Tabela 1: Primers desenhados no laboratório e condições das reacções de PCR para amplificação das β-lactamases
Reacção de PCR
Gene Primers
5' - 3'
Desnaturação
inicial
Nº de
ciclos Ciclos
Extensão
final
Tamanho
fragmento
blaNDM
F: TATCGCCGTCTAGTTCTGCTG 94º C
10 min 30
94º C, 45 seg
59º C, 45 seg
72º C, 1min
72º C
10 min 871 pb
R: ACTGCCCGTTGACGCCCAAT
blaTEM
F: GAAAGGGCCTCGTGATAC 94º C
3 min 35
94º C, 1 min
56º C, 1 min
72º C, 1 min
72º C
9 min 1058 pb
R: TTACCAATGCTTAATCAGTGA
blaCTX-M-2
F: ATGACTCAGAGCATTCGCC 94º C
3 min 30
94º C, 1 min
61º C, 1 min
72º C, 2 min
72º C
10 min 879 pb
R: GCATCAGAAACCGTGGGT
F, primer forward; R, primer reverse

MATERIAL E MÉTODOS
13
• Porinas
Para os isolados de Klebsiella spp. pesquisou-se, também, a presença dos genes ompK35 e
ompK36 que codificam para as porinas OmpK35 e OmpK36 da membrana externa,
respectivamente. Os primers para os genes ompK35 e ompK36 foram desenhados no
laboratório, de acordo com as sequências disponíveis no Genbank (AJ303057 e GU461279), e
estão descritos na Tabela 2, assim como as condições de PCR utilizadas.
• Resistência às quinolonas
As quinolonas e fluoroquinolonas entram nas células das bactérias de Gram-negativo através
de porinas ou atravessam directamente a membrana lipídica e citoplasmática inibindo
principalmente a acção da DNA girase e topoisomerase IV. A resistência a estes antibióticos
resulta: (i) da acumulação de mutações nos genes gyrA e parC que codificam as enzimas DNA
girase e topoisomerase IV, respectivamente;97
(ii) da acção de determinantes plasmídicos,
qnrA, qnrB ou qnrS, que protegem a DNA girase da actividade inibitória das quinolonas; (iii) da
actividade da enzima aminoglicosido-acetiltransferase, codificada pela variante aac(6’)-Ib-cr do
gene aac(6’)-Ib, que tem capacidade de acetilar algumas quinolonas; (iv) e/ou da diminuição da
permeabilidade membranar pelo aumento da expressão de bombas de efluxo ou por alteração
da natureza e quantidade das porinas.98; 99
Para os isolados resistentes às quinolonas pesquisou-se a presença dos genes qnrA,100
qnrB,101
qnrS102
e do alelo aac(6’)-Ib-cr103
com primers e condições de PCR previamente
descritas.
Tabela 2: Primers e condições das reacções de PCR para amplificação dos genes das porinas OmpK35 e Ompk36
Reacção de PCR
Gene Primers
5' - 3'
Desnaturação
inicial
Nº de
ciclos Ciclos
Extensão
final
Tamanho
fragmento
ompK35
F: ATATTCTGGCAGTGGTGATCC 94º C
10 min 35
94º C, 30 seg
60º C, 30 seg
72º C, 1min
72º C
10 min 1012 pb
R: GCTTTGGTGTAATCGTTGTC
ompK36
F: TAGCAGGCGCAGCAAATGC 94º C
10 min 35
94º C, 30 seg
60º C, 30 seg
72º C, 1min
72º C
10 min 1031 pb
R: TGCAACCACGTCGTCGGTA
F, primer forward; R, primer reverse
Determinação do ambiente genético dos genes de resistência
Muitos genes de resistência aos antibióticos estão localizados em transposões e integrões.
Investigou-se, assim, a presença do transposão Tn4401 frequentemente associado ao gene
blaKPC66; 68
nos isolados com o gene blaKPC-3 identificado. Para esse efeito pesquisou-se a
presença dos genes tnpA e ISKpn7 a montante e ISKpn6 a jusante do gene. Utilizaram-se
primers descritos por Naas et al. (2008)66
para amplificar os genes tnpA e ISKpn6, enquanto

MATERIAL E MÉTODOS
14
para o gene ISKpn7 os primers foram desenhados no laboratório de acordo com as sequências
disponíveis no Genbank (EU176012.1).
Pesquisou-se em todos os isolados a presença de integrões de classe I e III com primers e
condições de PCR descritos previamente por Lévesque et al. (1995).60
Para esse efeito
amplificou-se a zona da cassete genética e as regiões conservadas 5’ e 3’ a montante e
jusante, respectivamente, da cassete genética. Os produtos de PCR dos elementos móveis
foram resolvidos em gel de agarose a 1%, em TAE (1x) e corados com GelRed (Botium).
Identificação dos plasmídeos transportadores de β-lactamases
A identificação dos grupos de incompatibilidade dos plasmídeos, dos vinte e sete isolados, foi
efectuada pela técnica de Replicon Typing, desenvolvida por Carattoli et al. (2005).104
Esta
técnica consiste na realização de cinco reacções de PCR multiplex e três reacções de PCR
simplex, com primers específicos para origens de replicação de plasmídeos pertencentes a
diferentes grupos de incompatibilidade (IncHI1, IncHI2, IncI1-Iγ, IncX, IncL/M, IncN, IncFIA,
IncFIB, IncW, IncY, IncP, IncFIC, IncA/C, IncT, IncFIIAs, IncK, IncB/O, IncF). Para as reacções
de PCR foi usado o Supreme NZYTaq Green Master Mix (NZYTech) Os produtos de PCR
foram analisados em gel de agarose a 1%, em TBE (1x), e corados com brometo de etídio.
Posteriormente foram purificados e sequenciados.
Posteriormente, para confirmar a associação das carbapenemases aos plasmídeos,
seleccionaram-se dois isolados de K. pneumoniae e um isolado de E. coli, representativos de
cada estirpe, para se proceder à conjugação. Utilizou-se como estirpe receptora a E. coli K12
J53 azida resistente. Inoculou-se 5 mL de meio Brain Heart Infusion (BHI) com 100 μL de
estirpe dadora e 200 μL da estirpe receptora, crescidas previamente over-night, e incubou-se
sem agitação a 25 e 37ºC. Posteriormente plaqueou-se em MH suplementado com azida e
cefotaxima. Fez-se o antibiograma das estirpes transformantes e extraiu-se o DNA, como já foi
descrito. Realizou-se ainda um screening por PCR para β-lactamases e para os grupos de
incompatibilidade dos plasmídeos.
Identificação dos determinantes genéticos de virulência
Para os isolados de K. pneumoniae pesquisou-se a presença dos genes de virulência através
de amplificação por PCR e análise em gel de agarose a 1%, em TBE, e corado com GelRed
(Biotium). Posteriormente purificou-se e enviou-se para sequenciação. Os genes pesquisados
foram magA (mucoviscosidade), rmpA (regulador do fenótipo mucóide),105
k2A (antigénio
capsular),106
khe (hemolisina), fimH (proteína das fímbrias tipo 1), mrkDV1, mrkV2, mrkV3 e
mrkV4 (adesinas das fímbrias tipo 3) e iucC (sideróforo). Os primers desenhados no
laboratório, de acordo com as sequências disponíveis no Genbank (magA: NC_012731.1; khe:
NC_012731.1; fimH: NC_012731.1; mrkDV1: EU682505.2; mrkDV2: AY225463.1; mrkDV3:

MATERIAL E MÉTODOS
15
AY225464.1; mrkDV4: AY225465.1; iucC: NC_005249.1), e as condições de PCR usadas
estão descritos na Tabela 3.
Sequenciação e análise das sequências nucleotídicas
Os produtos de PCR para sequenciação foram purificados através do kit JETQUICK Spin
Column Technique (GENOMED), segundo as instruções do fabricante, e ambas as cadeias
foram sequenciadas pelo serviço da Macrogen Korea.
As sequências nucleotídicas foram analisadas e comparadas com as existentes na base de
dados Genbank, recorrendo aos programas informáticos BLASTN e ClustalW, disponíveis
online.
Tabela 3: Primers desenhados e condições das reacções de PCR para amplificação dos genes de virulência
Reacção de PCR
Gene Primers
5' - 3'
Desnaturação
inicial
Nº de
ciclos Ciclos
Extensão
final
Tamanho
fragmento
magA
F: TCTGTCATGGCTTAGACCGAT 94º C
10 min 30
94º C, 30 seg
58º C, 30 seg
72º C, 1min
72º C
10 min 1137 pb
R: GCAATCGAAGTGAAGAGTGC
khe
F: TGATTGCATTCGCCACTGG 94º C
10 min 30
94º C, 30 seg
60º C, 30 seg
72º C, 1min
72º C
10 min 428 pb
R: GGTCAACCCAACGATCCTGG
fimH
F: TGTTCACCACCCTGCTGCTG 94º C
10 min 30
94º C, 30 seg
61º C, 30 seg
72º C, 1min
72º C
10 min 512 pb
R: CACCACGTCGTTRTTGGCGT
mrkDV1
F: CGGTGATGCTGGACATGGT 94º C
10 min 30
94º C, 30 seg
60º C, 30 seg
72º C, 1min
72º C
10 min 300 pb
R: CYTCSAGSGAATAGTTGGTG
mrkDV2
mrkDV3
mrkDV4
F: CTTRATGGCGMTGGGSACCA 94º C
10 min 30
94º C, 30 seg
61º C, 30 seg
72º C, 1min
72º C
10 min 950 pb
R: TCATRTGCGAYTCCACCTCR
iucC
F: GTGCTGTCGATGAGCGATGC 94º C
10 min 30
94º C, 30 seg
62º C, 30 seg
72º C, 1min
72º C
10 min 944 pb
R: GTGAGCCAGGTTTCAGCGTC
F, primer forward; R, primer reverse
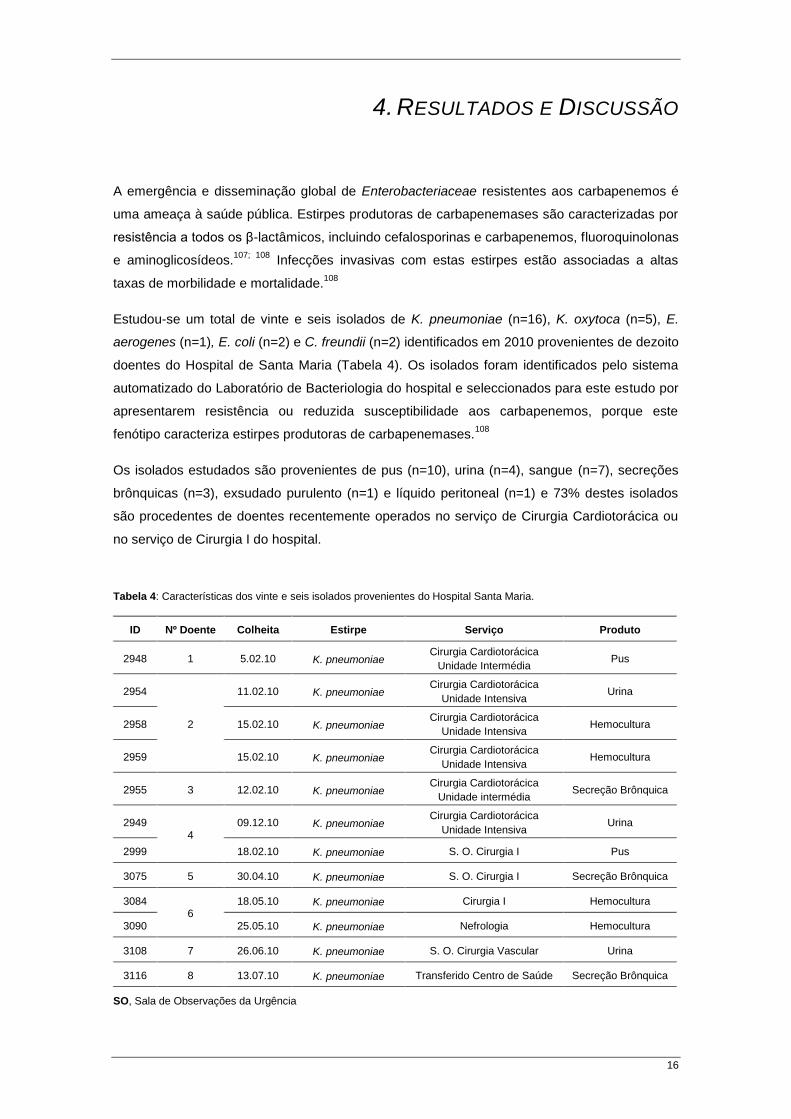
16
4. RESULTADOS E DISCUSSÃO
A emergência e disseminação global de Enterobacteriaceae resistentes aos carbapenemos é
uma ameaça à saúde pública. Estirpes produtoras de carbapenemases são caracterizadas por
resistência a todos os β-lactâmicos, incluindo cefalosporinas e carbapenemos, fluoroquinolonas
e aminoglicosídeos.107; 108
Infecções invasivas com estas estirpes estão associadas a altas
taxas de morbilidade e mortalidade.108
Estudou-se um total de vinte e seis isolados de K. pneumoniae (n=16), K. oxytoca (n=5), E.
aerogenes (n=1), E. coli (n=2) e C. freundii (n=2) identificados em 2010 provenientes de dezoito
doentes do Hospital de Santa Maria (Tabela 4). Os isolados foram identificados pelo sistema
automatizado do Laboratório de Bacteriologia do hospital e seleccionados para este estudo por
apresentarem resistência ou reduzida susceptibilidade aos carbapenemos, porque este
fenótipo caracteriza estirpes produtoras de carbapenemases.108
Os isolados estudados são provenientes de pus (n=10), urina (n=4), sangue (n=7), secreções
brônquicas (n=3), exsudado purulento (n=1) e líquido peritoneal (n=1) e 73% destes isolados
são procedentes de doentes recentemente operados no serviço de Cirurgia Cardiotorácica ou
no serviço de Cirurgia I do hospital.
Tabela 4: Características dos vinte e seis isolados provenientes do Hospital Santa Maria.
ID Nº Doente Colheita Estirpe Serviço Produto
2948 1 5.02.10 K. pneumoniae Cirurgia Cardiotorácica
Unidade Intermédia Pus
2954
2
11.02.10 K. pneumoniae Cirurgia Cardiotorácica
Unidade Intensiva Urina
2958 15.02.10 K. pneumoniae Cirurgia Cardiotorácica
Unidade Intensiva Hemocultura
2959 15.02.10 K. pneumoniae Cirurgia Cardiotorácica
Unidade Intensiva Hemocultura
2955 3 12.02.10 K. pneumoniae Cirurgia Cardiotorácica
Unidade intermédia Secreção Brônquica
2949 4
09.12.10 K. pneumoniae Cirurgia Cardiotorácica
Unidade Intensiva Urina
2999 18.02.10 K. pneumoniae S. O. Cirurgia I Pus
3075 5 30.04.10 K. pneumoniae S. O. Cirurgia I Secreção Brônquica
3084 6
18.05.10 K. pneumoniae Cirurgia I Hemocultura
3090 25.05.10 K. pneumoniae Nefrologia Hemocultura
3108 7 26.06.10 K. pneumoniae S. O. Cirurgia Vascular Urina
3116 8 13.07.10 K. pneumoniae Transferido Centro de Saúde Secreção Brônquica
SO, Sala de Observações da Urgência

RESULTADOS E DISCUSSÃO
17
Tabela 4 (continuação): Características dos vinte e seis isolados provenientes do Hospital Santa Maria.
ID Nº Doente Colheita Estirpe Serviço Produto
94182 9
1.08.10 K. pneumoniae Cirurgia I Pus
3191 23.09.10 K. pneumoniae Cirurgia I Pus
3030 10 14.03.10 K. pneumoniae Transferido Centro de Saúde Hemocultura
3046 11 26.03.10 K. pneumoniae S. O. Cirurgia I Hemocultura
3034
12
16.03.10 K. oxytoca S. O. Cirurgia I Exsudado Purulento
3042 18.03.10 K. oxytoca S. O. Cirurgia I Líquido Peritoneal
3051 29.03.10 K. oxytoca Medicina Intensiva Hemocultura
3066 13 16.04.10 K. oxytoca Transferido Centro de Saúde Pus
3074 14 04.05.10 K. oxytoca Transferido Centro de Saúde Urina
3016 15 5.03.10 E. aerogenes Cirurgia Cardiotorácica Pus
3023 16 10.03.10 E. coli Cirurgia I Pus
3052 17 30.03.10 E. coli Transferido Centro de Saúde Pus
3192 18
2.10.10 C. freundii Cirurgia I Pus
3193 2.10.10 C. freundii Cirurgia I Pus
SO, Sala de Observações da Urgência
4.1 Susceptibilidade aos antibióticos
Foram realizados testes de susceptibilidade aos vinte e seis isolados para compreender o
comportamento destes perante um conjunto de antibióticos (amoxicilina/ácido clavulânico,
carbapenemos, cefalosporinas, fluoroquinolonas, aminoglicosídeo, tetraciclina, nitrofurantoína,
fosfomicina). Os antibióticos seleccionados pretendem representar os grupos antimicrobianos
frequentemente utilizados na prática clínica.
Na Tabela AI, em anexo, estão representados os perfis de susceptibilidade dos isolados. Pela
análise desta tabela verifica-se que as principais características exibidas por todos os isolados
é um perfil de multirresistência e resistência ou baixa susceptibilidade aos carbapenemos.
A actividade antimicrobiana menos eficaz foi apresentada pelos agentes antimicrobianos β-
lactâmicos (amoxicilina/ácido clavulânico, cefotaxima, cefoxitina, ceftazidima, cefepima,
imipenemo, meropenemo e aztreonamo) (Gráfico 1). Dentro do grupo dos β-lactâmicos, os
antibióticos menos eficazes foram a ceftazidima, com uma resistência bacteriana de 100%, e a
amoxicilina/ácido clavulânico, cefotaxima e aztreonamo com uma resistência de 96%. Para a
cefoxitina e a cefepima observou-se uma resistência de 69%. Os carbapenemos, imipenemo e
meropenemo, dentro do grupo dos β-lactâmicos, foram os antibióticos com menor percentagem
de resistência, 34% e 65% respectivamente, no entanto os isolados classificados como
sensíveis segundo as normas do CLSI (2010)84
apresentaram, para estes antibióticos, baixa
susceptibilidade quando comparados com estirpes de referência. Um estudo de Endimiani et al.

RESULTADOS E DISCUSSÃO
18
(2009) revela que 60% dos isolados produtores da carbapenemase KPC são classificados
como sensíveis para o imipenemo e meropenemo.
O antibiótico com actividade mais eficaz foi a fosfomicina, para a qual não foi observada
nenhuma resistência. As fluoroquinolonas ciprofloxacina e levofloxacina também tiveram uma
baixa percentagem de resistência, 30% e 23%, respectivamente. A nitrofurantoína teve 27% de
resistência. Para a gentamicina observou-se uma resistência de 62% e 27% dos isolados foram
classificados com um fenótipo intermédio. A tigeciclina teve uma resistência de 54%.
Gráfico 1: Susceptibilidade aos antibióticos dos vinte e seis isolados de Enterobacteriaceae em estudo.
amc, amoxicilina/ácido clavulânico; fox, cefoxitina; ctx, cefotaxima; caz, ceftazidima; fep, cefepima; atm, aztreonamo;
imp, imipenemo; mem, meropenemo; cip, ciprofloxacina; lvx, levofloxacina; gm, gentamicina; tgc, tigeciclina; ft,
nitrofurantoína; fos, fosfomicina.
Na detecção fenotípica dos mecanismos de resistência, os isolados 2948, 2954, 2958, 2959,
2955, 2949, 2999, 3075, 3084, 3108, 3116, 94182, 3046, 3034, 3042, 3051, 3016 e 3023
apresentaram um aumento do diâmetro da zona de inibição para a amoxicilina/ácido
clavulânico, cefoxitina, cefotaxima, ceftazidima e imipenemo na presença de ácido borónico
(Figura 4), o que indica que estes isolados codificam para uma carbapenemase de classe A
que é responsável pela hidrólise destes antibióticos. Os isolados 3034, 3042 e 3051 de K.
oxytoca, provenientes do mesmo doente, na presença de cloxacilina, tiveram maior
susceptibilidade às cefalosporinas, sugerindo a presença simultânea de uma cefalosporinase
AmpC.
Com o EDTA não se verificou nenhuma alteração do diâmetro da zona de inibição para os
antibióticos testados, sugerindo assim que nenhum dos isolados codifica para metalo-β-
lactamases.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
amc ctx fox caz fep imp mem atm cip lvx gm tgc ft fos
Resistente Intermédio Sensível

RESULTADOS E DISCUSSÃO
19
Figura 4: Antibiograma do isolado de K. pneumoniae produtora de carbapenemase A) sem ácido borónico e B) com
ácido borónico, usando discos dos antibióticos amoxicilina/ácido clavulânico (AMC), cefotaxima (CTX), ceftazidima
(CAZ), cefoxitina (FOX), ciprofloxacina (CIP), imipenemo (IMP) e gentamicina (GM).
O isolado 3030, na presença de cloxacilina, teve um aumento do diâmetro da zona de inibição
para a amoxicilina/ácido clavulânico, cefotaxima, cefoxitina e ceftazidima, indicando que uma
cefalosporinase AmpC está presente e é a responsável pela hidrólise destes β-lactâmicos. Os
isolados 3066, 3192 e 3193 apresentaram, simultaneamente, na presença de cloxacilina e
ácido borónico, um aumento do diâmetro da zona de inibição para os antibióticos
amoxicilina/ácido clavulânico, cefotaxima, cefoxitina e ceftazidima, o que também indica a
presença de uma cefalosporinase AmpC, pois estas também são inibidas por ácido borónico.109
A presença de uma carbapenemase é excluída porque não existe uma alteração significativa
do diâmetro da zona de inibição para o imipenemo.
Apenas o isolado 3074 de K. oxytoca foi classificado com um fenótipo intermédio, segundo as
normas do CLSI (2010),84
para a amoxicilina/ácido clavulânico, sugerindo que pode codificar
para uma β-lactamase de espectro alargado.
4.2 Tipificação dos isolados bacterianos
O controlo de uma infecção associada aos cuidados de saúde e o estudo epidemiológico
pressupõem o conhecimento do parentesco biológico e genético entre as bactérias
multirresistentes. A necessidade de determinar a relação entre os organismos surge, durante o
aparecimento de surtos com origem em organismos da mesma espécie e com o mesmo
fenótipo de resistência antimicrobiana. É necessário identificar se estamos perante uma
disseminação clonal ou plasmídica e qual a fonte de infecção. Diversos métodos moleculares
são aplicados para originarem um fingerprint (impressão digital) de cada microrganismo.110; 111
AMC
IMP
GM
CIP
CAZ
FOX
CTX
AMC
IMP
GM
CIP
CAZ
FOX
CTX
A B

RESULTADOS E DISCUSSÃO
20
As técnicas de M13 e ERIC-PCR fingerprintig tiveram uma boa correlação e permitiram
classificar entre os isolados de K. pneumoniae, nove perfis genotípicos (Figura 5). O principal
perfil genotípico é o A. Cinco isolados (2948, 2954, 2958, 2959 e 2955) provenientes de três
doentes do Serviço de Cirurgia Cardiotorácica, apresentaram este perfil. Estes resultados
sugerem que estamos na presença de um clone de K. pneumoniae produtor de uma
carbapenemase com capacidade de disseminação e de adaptação. Ainda neste serviço temos
o perfil B, que difere do perfil A em uma banda, sugerindo que mutações, ou inserções
alteraram o seu genoma, levando-o a divergir do perfil A. Os restantes isolados de K.
pneumoniae, maioritariamente provenientes do Serviço de Cirurgia I, têm perfis genómicos
distintos. Estes resultados mostram que existe uma grande variabilidade genética nos isolados
de K. pneumoniae estudados e mesmo com um perfil de susceptibilidade comum, estamos
perante diferentes clones.
Figura 5: Perfil de ERIC-PCR fingerprinting. O rectângulo laranja representa o perfil A, dominante no Serviço de
Cirurgia Cardiotorácica. O rectângulo verde representa os dois isolados de K. oxytoca com perfil igual (perfil M). O
rectângulo vermelho representa os dois isolados de C. freundii com perfil igual (perfil T).
Os isolados de K. oxytoca definiram-se em quatro perfis genómicos. Dois isolados tinham o
mesmo perfil. O isolado 3051, do doente 12, é caracterizado por um perfil fenótipo igual ao dos
outros isolados de K. oxytoca mas tem um perfil genómico diferente, no entanto bastante
similar, sugerindo que mutações, ou inserções alteraram o seu genoma.
Os dois isolados de E. coli apresentaram diferentes perfis genómicos, o que era esperado, pois
são provenientes de diferentes doentes e foram isolados em serviços distintos.
Os isolados de C. freundii provenientes do doente 18 tinham um perfil genotípico igual apesar
do perfil fenotípico ser diferente para os carbapenemos.
A B M T
K. pneumoniae K. oxytoca E. coli C. freundii

RESULTADOS E DISCUSSÃO
21
4.3 Multilocus Sequence Typing
O método MLST, baseado na análise de sequências de fragmentos internos de genes
housekeeping que fornecem um perfil alélico para cada isolado consoante as alterações
nucleotídicas observadas, é utilizado actualmente para caracterizar isolados bacterianos e
estabelecer facilmente relações entre eles. Os perfis obtidos por MLST são cruzados numa
base de dados internacional, facilitando a troca de informação e o estudo da disseminação das
estirpes à escala global.112; 113
Entre os isolados de K. pneumoniae, as sequências dos genes housekeeping permitiram definir
os seguintes alelos para cada locus:
Tabela 5: Perfil alélico obtido através da técnica de MLST
Klebsiella pneumoniae MLST
gapA infB mdh pgi phoE rpoB tonB
1 6 1 1 1 1 1
De acordo com a combinação de alelos, identificou-se uma única Sequence Type (ST) para
todos os isolados, a ST14. Para uma única ST foram detectados diferentes perfis genómicos.
Este resultado é consistente com as técnicas utilizadas, M13 e ERIC-PCR fingerprinting são
técnicas que permitem detectar rearranjos cromossomais causados por elementos móveis,
enquanto o MLST só analisa sete genes housekeeping, sendo assim mais apropriado para
estabelecer filogenias e epidemiologias a larga escala. Estirpes com um mesmo ST podem
então ter diferentes perfis genómicos com M13 e ERIC-PCR fingerprinting.114
K. pneumoniae ST14, descrita com um elevado potencial para causar outbreaks num estudo de
Oteo et al. (2009), está a ser referida como o “novo MRSA” devido à sua distribuição
internacional e aos marcadores de resistência aos antibióticos que lhe estão associados, como
a carbapenemase NDM-1.115; 116
4.4 Determinantes genéticos de resistência
A caracterização dos mecanismos moleculares e biológicos de resistência aos antimicrobianos
aumenta a capacidade de microbiologistas e epidemiologistas para monitorizar a propagação
da resistência aos antibióticos dentro e entre unidades de saúde e comunidades.117
Neste trabalho foram pesquisados nos isolados clínicos os genes que codificam para as
carbapenemases KPC, NDM, IMP, VIM e OXA, por estes apresentarem resistência ou baixa
susceptibilidade aos carbapenemos. Para além dos genes que codificam para
carbapenemases foram também pesquisados os genes das β-lactamases TEM, SHV, CTX-M,

RESULTADOS E DISCUSSÃO
22
DHA e CMY e em Klebsiella spp. os genes que codificam para as porinas OmpK35 e OmpK36,
pois a presença destas β-lactamases e/ou a ausência das porinas contribui para a resistência
aos β-lactâmicos. Os mecanismos de resistência de cada isolado estão representados na
Tabela AII em anexo.
Dos vinte e seis isolados, dezoito foram positivos para a amplificação do gene blaKPC (Figura 6),
os mesmos isolados que anteriormente mostraram um perfil de inibição com o ácido borónico,
o que revela que o ácido borónico tem uma elevada sensibilidade e especificidade para a
pesquisa fenotípica da carbapenemase KPC, concordante com o estudo de Tsakris et al.
(2011), no qual foi definido que a utilização deste inibidor com imipenemo é um método
específico para a detecção das carbapenemases KPC.118
Os dezoito produtos de PCR foram
posteriormente sequenciados e identificou-se a variante blaKPC-3, que hidrolisa o imipenemo,
meropenemo, ertapenemo, aztreonamo, ceftazidima, cefotaxima, cefepima e a cefoxitina.52
Este gene foi identificado pela primeira vez em Portugal e no Hospital Santa Maria em 2009,
numa K. pneumoniae ST11.62
Neste estudo todas as K. pneumoniae produtoras da
carbapenemase KPC-3 têm um ST14, não tendo ocorrido disseminação clonal da K.
pneumoniae ST11 mas a disseminação de um integrão e/ou plasmídeo contendo o gene
blaKPC-3.
Figura 6: Corrida de electroforese dos produtos de PCR da amplificação do gene blaKPC em gel de agarose a 1%.
Dos isolados produtores da carbapenemase KPC-3, dezassete isolados apresentaram
resistência para um dos carbapenemos testados ou para ambos. A presença da
carbapenemase KPC-3 não explica, por si só, o perfil de resistência aos carbapenemos. A
resistência ao imipenemo e meropenemo para estes isolados pode estar então associada a um
elevado número de cópias do gene blaKPC-3, a uma elevada expressão do mesmo gene ou à
alteração da permeabilidade da membrana externa.119
A impermeabilidade das porinas, por
mutação ou deleção dos genes ompK35 e/ou ompK36 em Klebsiella spp. contribui para uma
maior resistência ao meropenemo, ertapenemo, cefoxitina, cefotaxima, ceftazidima e cefepima,
uma cefalosporina de 4ª geração.9; 120; 121
Dos catorze isolados de Klebsiella spp. produtores da carbapenemase KPC-3, 86% não
apresentaram alteração dos genes ompK35 e ompK36, não havendo então alteração da

RESULTADOS E DISCUSSÃO
23
permeabilidade da membrana, estando a sua resistência aos carbapenemos associada a um
elevado número de cópias ou a uma elevada expressão do gene blaKPC-3. O isolado 2958, sem
alteração da permeabilidade, está classificado com um fenótipo sensível para o imipenemo e
meropenemo, segundo as normas do CLSI (2010),84
indicando que não tem um número
elevado de cópias nem uma elevada expressão do gene blaKPC-3.
O isolado 3046, para a amplificação do gene ompK35 teve um produto de PCR superior ao
esperado (2000 pb), o que aponta para a inserção de um elemento móvel neste gene que
impede a sua expressão. A inclusão de sequências de inserção nos genes ompK35 e ompK36,
também descrito nos estudos de Song et al. (2009) e Doumith et al. (2009), alteram a
permeabilidade da membrana externa, o que se observa com o aumento da resistência para a
cefotaxima, cefoxitina, cefepima, meropenemo e ligeiramente para o imipenemo.122; 123
Os isolados 2959, 3034, 3042 e 3051 não tiveram amplificação para o gene ompK35, indicando
a deleção deste fragmento de DNA ou a existência de diversas mutações e/ou deleções na
zona de emparelhamento do primer, logo não há expressão do gene, não havendo na
membrana externa destes isolados a porina OmpK35, o que diminui a permeabilidade aos
carbapenemos, cefotaxima, cefoxitina e cefepima. O isolado 3034 apresenta ainda um
mecanismo adicional de resistência aos carbapenemos, como um aumento da expressão ou
um elevado número de cópias do gene blaKPC-3. No estudo de Doumith et al. (2009) também foi
observada a ausência de amplificação dos genes ompK35 e ompK36.123
A associação da β-lactamase SHV-1 e/ou TEM-1 com a carbapenemase KPC-3 é observada
para 83% dos isolados produtores desta carbapenemase (Gráfico 2). As β-lactamases,
encontradas frequentemente em Enterobacteriaceae desde a introdução de cefalosporinas de
largo espectro na terapêutica, conferem resistência às penicilinas e às cefalosporinas de 1ª
geração.124; 125
As β-lactamases AmpC plasmídicas estão disseminadas pelos isolados clínicos de
Enterobacteriaceae. No entanto, as cefalosporinases do grupo DHA, com actividade hidrolítica
contra antibióticos β-lactâmicos, são pouco frequentes nas enterobactérias.126
A presença de
uma cefalosporinase DHA-1 foi confirmada por PCR para os isolados 3034 e 3042 de K.
oxytoca produtores da carbapenemase KPC-3, ambos provenientes do doente 12. Estes
isolados ainda codificam para a β-lactamase TEM-1. A associação da cefalosporinase DHA-1
com a carbapenemase KPC-3 aumenta a resistência dos isolados para a cefotaxima,
cefoxitina, ceftazidima e aztreonamo em relação a isolados que só codificam para a
carbapenemase.126; 127
Até à data não foi descrita a associação da DHA-1 com a
carbapenemase KPC-3, no entanto existem estudos que descrevem a associação da DHA-1
com a carbapenemase KPC-2.128; 129
A presença da cefalosporinase DHA-1 com a ausência da
porina OmpK35 na membrana externa destes isolados contribui para um aumento da
resistência aos carbapenemos.130

RESULTADOS E DISCUSSÃO
24
Gráfico 2: Combinação da carbapenemase KPC-3 com outras β-lactamases nos isolados clínicos em estudo.
As β-lactamases de espectro alargado estão presentes em 14% dos isolados de K.
pneumoniae multirresistentes.131
A detecção fenotípica de β-lactamases de espectro alargado
por inibição com o ácido clavulânico, pode ser mascarada pela expressão de carbapenemases
e/ou cefalosporinases que hidrolisam este inibidor. Após pesquisa molecular e sequenciação
dos produtos de PCR, identificou-se em dois isolados a presença de uma β-lactamase de
espectro alargado. O isolado 2999 produtor da carbapenemase KPC-3 codifica para a ESBL
SHV-35, não existindo nenhum estudo publicado sobre a actividade desta β-lactamase, no
entanto parece conferir uma maior resistência à cefepima, quando comparamos o isolado 2999
com o isolado 2949, proveniente do mesmo doente e com um fingerprinting genómico igual. No
isolado 3051, produtor da carbapenemase KPC-3, cefalosporinase DHA-1 e β-lactamase TEM-
1, identificou-se a ESBL SHV-12, anteriormente encontrada num plasmídeo com a
carbapenemase KPC-2 em França.132
A β-lactamases de espectro alargado SHV-12 difere da
SHV-1 por substituição de três aminoácidos e aumenta a resistência deste isolado à
ceftazidima e cefotaxima.133
A resistência aos carbapenemos em Enterobacteriaceae pode surgir por aquisição de genes
que codificam para carbapenemases (MBLs, KPC, GES ou OXA) ou por associação da
expressão de genes que codificam para cefalosporinases AmpC ou β-lactamases de espectro
alargado com a alteração da expressão de porinas da membrana externa.9; 120; 122; 130; 134
Oito isolados com reduzida susceptibilidade para os carbapenemos não codificam para
nenhuma carbapenemase. O isolado 3090 de K. pneumoniae, proveniente do doente 6 com
uma diferença de sete dias da data de isolamento do isolado 3084 produtor da carbapenemase
KPC-3, apenas codifica para as β-lactamases SHV-1 e TEM-1, no entanto é resistente para as
cefalosporinas e meropenemo e tem um perfil intermédio para o imipenemo. Este isolado 3090
não possui os genes que codificam para as porinas OmpK35 e OmpK36 da membrana externa
e a ausência destas conjuntamente com a actividade das β-lactamases SHV-1 e TEM-1
conferem a resistência ou baixa susceptibilidade para o imipenemo. Yang et al. (2010) e Tsai et
0
1
2
3
4
5
6
7
nº
de isola
dos
KPC-3 KPC-3 TEM-1
KPC-3 SHV-type
KPC-3 TEM-1
SHV-type
KPC-3 TEM-1 DHA-1
KPC-3 TEM-1 SHV-12 DHA-1

RESULTADOS E DISCUSSÃO
25
al. (2011) também verificaram que mesmo na ausência de carbapenemases, ESBLs e
cefalosporinases AmpC, a inexistência das duas porinas na membrana externa conferiam
resistência aos carbapenemos, sendo esta resistência mais elevada para o ertapenemo.9; 129
Procedentes do doente 9, temos o isolado 94182, que expressa a carbapenemase KPC-3, e o
isolado 3191 que não codifica para nenhuma das β-lactamases pesquizadas nem para as
porinas da membrana externa, o que justifica uma baixa susceptibilidade aos carbapenemos e
cefalosporinas mas a ausência de resistência.
Os isolados 3030 e 3066 possuem um perfil de resistência para as cefalosporinas e uma ligeira
diferença na susceptibilidade para os carbapenemos testados. O isolado 3030 não possui
alterações na expressão das porinas OmpK35 e OmpK36, no entanto tem baixa
susceptibilidade para os carbapenemos, alguns estudos indicam que além destas porinas,
existe outras, como a OmpK26, importantes na permeabilidade dos carbapenemos nas
espécies de Klebsiella e a sua ausência combinada com uma cefalosporinase contribui para a
resistência a estes antibióticos.135; 136; 137
Para o isolado 3066 não houve amplificação do gene ompK36 e a ausência da porina OmpK36
da membrana combinada com a cefalosporinase DHA-1 justificam a resistência aos
carbapenemos e cefalosporinas. A ausência da expressão da porina OmpK36 e a actividade da
cefalosporinase DHA-1 já foram descritas em vários estudos como factor de resistência para os
carbapenemos (imipenemo e meropenemo) e cefalosporinas (cefotaxima e cefoxitina).122; 129; 130
A β-lactamase CTX-M-15, identificada no Hospital de Santa Maria em 2005,94
é considerada
uma das β-lactamases de espectro alargado mais prevalentes no meio hospitalar.138
O isolado
3074 apresenta a ESBL CTX-M-15 associada com a β-lactamase SHV-11 e com a perda da
porina OmpK36 da membrana externa. Esta associação permite que este isolado hidrolise os
carbapenemos e possua assim um perfil de baixa susceptibilidade para estes antibióticos.129; 134
O isolado de E. coli 3052 produtor da β-lactamase TEM-1 possui uma baixa susceptibilidade
aos carbapenemos que parece ser conferida por alteração de permeabilidade por alteração ou
ausência da expressão de porinas.139
Os isolados 3192 e 3193 de C. freundii, provenientes do doente 18, têm perfis diferentes de
susceptibilidade para os carbapenemos. Ambos os isolados possuem as cefalosporinases
CMY-47 e DHA-1. A β-lactamase CMY-47, identificada em C. freundii em 2008, hidrolisa a
amoxicilina/ácido clavulânico, cefotaxima, cefoxitina, ceftazidima, aztreonamo e parcialmente o
a cefepima.140
A resistência para os carbapenemos no isolado 3191 pode ser explicada por
uma alteração da permeabilidade da membrana. A inibição da expressão de porinas pode
surgir frequentemente, durante terapias com carbapenemos. O que justifica que dois isolados
idênticos tenham uma alteração de susceptibilidade para os carbapenemos, apenas para um
deles surgiu mutações.141

RESULTADOS E DISCUSSÃO
26
Apesar da pesquisa fenotípica de metalo-β-lactamases, com a adição de EDTA, ter sido
negativa fez-se, para confirmação, a pesquisa molecular das metalo-β-lactamases blaNDM,
blaIMP e blaVIM. Não houve amplificação dos genes codificantes para metalo-β-lactamases para
nenhum dos isolados em estudo, resultado concordante com a pesquisa fenotípica. A pesquisa
molecular da carbapenemase OXA-48 também teve um resultado negativo para os isolados.
Relativamente aos genes e alelos que conferem resistência às quinolonas, verificou-se que
seis isolados apresentavam o alelo aac(6’)-Ib-cr, sete isolados o gene qnr e três isolados
apresentavam simultaneamente o alelo aac(6’)-Ib-cr e o gene qnr. O alelo aac(6’)-Ib-cr
aumenta a resistência dos isolados para a ciprofloxacina. Verificou-se que este alelo está
muitas vezes associado à carbapenemase KPC, à cefalosporinase DHA-1e principalmente à
ESBL CTX-M-15.142
O alelo do gene qnr prevalente nos isolados em estudo foi o qnrB4, que está frequentemente
associado a ESBL SHV-12 e à cefalosporinase DHA-1,98; 143
o que também foi verificado neste
estudo. As proteínas QnrB1, QnrB4 e QnrS1 conferem resistência ao ácido nalidixico e baixam
a susceptibilidade para a ciprofloxacina.143
Os isolados resistentes para as fluoroquinolonas
têm alterações de permeabilidade ou mutações na DNA girase e/ou DNA topoisomerase.98
A
perda da porina OmpK36 e mutações na DNA topoisomerase também causam resistência às
fluoroquinolonas,121
podendo estar na origem da resistência dos isolados 3066 e 3074.
4.5 Disseminação de genes de resistência
A disseminação de genes de resistência aos antibióticos entre estirpes bacterianas aumenta a
ocorrência de doenças infecciosas. Os genes de resistência podem estar localizados em
plasmídeos, transposões e integrões, permitindo assim a sua transferência entre diferentes
espécies bacterianas.60
A localização do gene blaKPC tem sido observada em plasmídeos de diversos tamanhos, que
podem apresentar mais de uma cópia do gene, demonstrando alta capacidade de
mobilização.68
Recentemente, um estudo de Naas et al. (2008), descreveu que o gene blaKPC-2
estava inserido no transposão Tn4401 e um estudo de Curiao et al. (2010) descreveu uma
variante do transposão Tn4401, com uma deleção de 100 pb a jusante do gene blaKPC-3.66; 67
Um outro estudo de Shen et al. (2009) descreve um transposão Tn4401 onde ocorre a
substituição do gene ISKpn7 a montante do gene blaKPC-2, pelo gene ISKpn8.144
No presente estudo, detectaram-se cinco ambientes genéticos diferentes para o gene blaKPC-3
(Figura 7). O ambiente genético dominante, presente em oito isolados de K. pneumoniae (n=5),
K. oxytoca (n=2) e E. coli (n=1), é idêntico à estrutura do transposão descrito por Naas et al.
(2008). 61
Este transposão foi identificado em isolados com diferentes origens geográficas e
diferentes ST-types, em Enterobacteriaceae.66; 145; 146
O Tn4401, localizado em diferentes loci e
inserido em plasmídeos com diferentes tamanhos e grupos de incompatibilidade, 66; 145
é capaz

RESULTADOS E DISCUSSÃO
27
de transposição replicativa. A transposição replicativa, que ocorre com elevada frequência para
este transposão, está na origem das inúmeras cópias presentes num mesmo genoma e/ou
plasmídeo.69
A
B
C
D
Figura 7: Representação esquemática das variantes do transposão Tn4401-like encontrados no presente estudo: a)
Tn4401a; b) Tn4401b; c) Tn4401c; d) Tn4401d. Os genes e a sua orientação de transcrição são representados por
setas horizontais. O transposão é delimitado por duas sequências invertidas representadas por triângulos cinzentos. Os
triângulos brancos representam as sequências invertidas do ISKpn7 e ISKpn6. A linha vertical representa uma zona de
deleções.
O segundo ambiente genético, mais comum entre os isolados, difere do transposão Tn4401 a
montante do gene ISKpn7 (Figura 7-A). Apesar de ter havido amplificação para o gene tnpA
este não se encontra a montante do gene ISKpn7. Entre os genes tnpA e ISKpn7 parece existir
uma zona de inserção que impede a amplificação, por PCR, da região média entre os genes. A
inserção não parece afectar a transposição replicativa deste transposão, pois este encontra-se
disseminado entre os isolados produtores de carbapenemase KPC-3 do Hospital Santa Maria.
Um outro ambiente genético (Figura 7-B), presente num isolado de K. pneumoniae e num
isolado de E. aerogenes, é caracterizado por uma região com inserção a montante do gene
blaKPC-3 que impede a amplificação por PCR, da região média entre os genes ISKpn7 e blaKPC-3.
tnpR tnpA IstA IstB blaKPC tnpA
ISKpn7 ISKpn6
Zona de inserção
tnpR tnpA IstA IstB blaKPC tnpA
ISKpn7 ISKpn6
Zona de inserção
tnpR tnpA blaKPC tnpA
ISKpn6
tnpR tnpA IstA IstB blaKPC tnpA IstA IstB bla tnpA
ISKpn7 ISKpn6

RESULTADOS E DISCUSSÃO
28
Os restantes ambientes genéticos encontrados são caracterizados por deleções. No ambiente
genético C da Figura 7, presente apenas no isolado 2958, houve a deleção da região ISKpn7.
No ambiente genético D da Figura 7, houve a eliminação da região ISKpn6. Ambos os
transposões perderam a sua capacidade de transposição, pois as sequências de inserção são
essenciais para a sua mobilidade.
Um nível excessivo de transposição replicativa e/ou a acumulação de um número considerável
de transposões, no mesmo plasmídeo ou genoma, pode estar na origem destes ambientes
genéticos mutados.147
Em quatro dos vinte e seis isolados, em que se pesquisaram integrões, os resultados foram
positivos. Os isolados 3034 e 3042 de K. oxytoca, provenientes do mesmo doente, têm
integrões com diferentes enzimas na zona variável de cada um, demonstrando uma elevada
capacidade para aquisição e perda de genes destes elementos móveis. O integrão presente no
isolado 3034 tem na zona variável os genes dhfrVII-aadA4, que conferem resistência às
sulfonamidas e estreptomicina, respectivamente. O isolado 3042 tem na zona variável do seu
integrão o gene dfr17 que confere resistência ao trimethoprim. Os isolados de C. freundii, com
um perfil genómico igual, possuem um integrão com os mesmos genes na zona variável
(dfrA12-orfF-aadA2a) que conferem resistência ao trimethoprim e aos aminoglicosídeos.
Foram detectados diferentes grupos de incompatibilidade de plasmídeos nos vinte e seis
isolados estudados. O grupo de incompatibilidade predominante entre os isolados foi o IncF
(65%) (Gráfico 3). Verificou-se também a existência dos grupos de incompatibilidade IncFIA
(27%), IncA/C (19%), IncFIC (23%), IncHI1 (23%), IncHI2 (12%), IncFIIA (4%).
Gráfico 3: Grupos de incompatibilidade dos plasmídeos detectados nos isolados em estudo.
0%
10%
20%
30%
40%
50%
60%
70%
IncF IncFIA IncA/C IncFIC IncHI1 IncHI2 IncFIIA

RESULTADOS E DISCUSSÃO
29
Para identificar o plasmídeo responsável pela disseminação do gene blaKPC-3 entre os isolados,
escolheram-se os isolados 2948, 2949 e 3023. Os isolados 2948 e 2949 têm plasmídeos com
os grupos de incompatibilidade IncFIA e IncF e o isolado 3023 de E. coli possui apenas um
plasmídeo com o grupo de incompatibilidade IncF. Por conjugação, os três isolados
transferiram o plasmídeo IncF para a estirpe de E. coli K12 J53 azida resistente. Por PCR
confirmou-se a presença do gene blaKPC-3 na estirpe de E. coli K12 J53 azida resistente e
verificou-se a aquisição de resistência à amoxicilina/ácido clavulânico, cefotaxima e
ceftazidima. A aquisição do gene blaKPC-3 também diminuiu a susceptibilidade da E. coli K12
J53 azida resistente para a cefoxitina e imipenemo, reduzindo o diâmetro da zona de inibição
em média em 13 mm e 9 mm respectivamente. Com este ensaio verifica-se que a transferência
do gene blaKPC-3 está associada ao plasmídeo do grupo IncF denotando-se a importância dos
plasmídeos na disseminação desta resistência independentemente do género bacteriano.
Para os isolados em que não foi observada a presença de plasmídeos, a presença do gene
blaKPC-3 pode estar no cromossoma bacteriano ou num plasmídeo de um grupo de
incompatibilidade que não está inserido neste conjunto de origem de replicação estudadas. Os
plasmídeos multirresistentes têm uma maior instabilidade, o que pode estar na origem da perda
destes plasmídeos. Se a bactéria perdeu o plasmídeo, o gene blaKPC-3 associado ao transposão
Tn4401-type pode ter sofrido translocação replicativa para o cromossoma antes deste
evento.148
Disseminação do gene blaKPC-3 no Hospital Santa Maria
No Serviço de Cirurgia Cardiotorácica foram caracterizados dois perfis genéticos entre os
isolados de K. pneumoniae. O isolado 2949, do doente 4 deste serviço, apresentou as mesmas
características fenotípicas que os restantes isolados de K. pneumoniae descritos neste serviço,
no entanto têm perfis genéticos diferentes, o que sugere que ocorreu transferência horizontal
dos genes de resistência entre eles.
O doente 4, inicialmente no Serviço de Cirurgia Cardiotorácica, foi transferido para o serviço de
Cirurgia I, desde então foram detectadas estirpes de Klebsiella e de outras espécies noutros
doentes com as mesmas características dos isolados do Serviço de Cirurgia Cardiotorácica, em
outros doentes presentes neste serviço.
Na origem desta disseminação entre os isolados das espécies Klebsiella spp., E. coli e E.
aerogenes está associado o plasmídeo do grupo de incompatibilidade IncF, o que foi verificado
por conjugação.

RESULTADOS E DISCUSSÃO
30
4.6 Determinantes de virulência em Klebsiella spp.
As bactérias do género Klebsiella estão frequentemente associadas a infecções do tracto
urinário, pneumonia e outras infecções nosocomiais com elevada morbidade e mortalidade.70
A maioria dos isolados clínicos produz uma cápsula polissacárida que cobre a superfície
bacteriana. As cápsulas, essenciais para a virulência de Klebsiella spp., podem apresentar
diferentes serótipos e intensidade de virulência. 70; 149
Neste estudo, os isolados do género
Klebsiella tinham um fenótipo mucóide e entre eles detectou-se o serótipo K2 em 57% dos
isolados (Tabela 8). Os restantes isolados não eram nem do serótipo K1 nem do serótipo K2. O
serótipo capsular K2 está associado às estirpes de Klebsiella spp. mais virulentas e
predominantes entre as infecções,70
pois este serótipo aumenta a resistência à fagocitose,
opsonização e ao efeitos bactericidas do soro.149
A adesão da bactéria às superfícies mucosas é um passo essencial para o desenvolvimento da
infecção. Nas bactérias de Gram-negativo a adesão é mediada por adesinas fimbriais do tipo 1
e/ou tipo 3.70
95% dos isolados clínicos em estudo codificam para adesinas do tipo 1 e/ou do
tipo 3 (Tabela 7). As adesinas do tipo 1, codificadas pelo gene fimH, medeiam a adesão a
estruturas das células do hospedeiro que contêm manose, sendo um importante factor de
virulência em infecções urinárias.150
As adesinas do tipo 3, codificadas pelo gene mrkD,
efectuam a sua adesão na ausência de manose e promovem a formação de biofilmes.150; 151
Para o gene mrkD existem quatro alelos diferentes (mrkDv1, mrkDv2, mrkDv3 e mrkDv4) com
diferentes forças adesivas, sendo o alelo mrkDv1 o que codifica para a adesina com menos
força adesiva.151
Entre os vinte e um isolados de Klebsiella spp., em 95% dos isolados foi
identificado, por PCR, o alelo mrkDv1 em simultâneo com um dos alelos mrkDv2, mrkDv3 ou
mrkDv4.
O crescimento da bactéria nos tecidos do hospedeiro é limitado pelos mecanismos de defesa e
a disponibilidade de ferro. Como a concentração de ferro livre é baixa, as bactérias secretam
sideróforos com elevada afinidade para o ferro, de modo a assegurarem o seu fornecimento.152
Apenas quatro isolados tiveram um PCR positivo para a presença do gene iucC, que codifica
para uma aerobactina. Normalmente este gene está associado ao gene rmpA,152
o qual não foi
detectado entre os isolados em estudo. No entanto existem outros sistemas de aquisição de
ferro que não foram pesquisados neste trabalho.
O gene khe, que codifica para uma hemolisina com actividade hemolítica para os glóbulos
vermelhos humanos, foi identificado apenas em dois isolados de pus.

RESULTADOS E DISCUSSÃO
31
Tabela 7: Factores de virulência nos isolados em estudo de Klebsiella spp..
ID Nº Doente k2A rmpA magA fimH mrkDv1 mrkDv2,v3,v4 khe iucC
2948 1 + - - + + + - -
2954
2
+ - - + + + - -
2958 + - - + + + - -
2959 + - - + + + - -
2955 3 + - - + + + - -
2949 4
+ - - + + + - -
2999 - - - + + + - -
3075 5 + - - + + + - -
3084 6
+ - - + + + - -
3090 + - - + + + - -
3108 7 - - - - + + - +
3116 8 - - - + + + - -
94182 9
+ - - + + - + -
3191 - - - - - - - -
3030 10 - - - + + + - -
3046 11 + - - + + + - -
3034
12
- - - + + + - +
3042 - - - + + + - +
3051 - - - + + + - +
3066 13 - - - + + + + -
3074 14 + - - + + + - -
+, amplificação do gene por PCR; -, não se observou amplificação do gene por PCR

32
5. CONCLUSÃO
Entre os vinte e seis isolados clínicos em estudo, com resistência ou reduzida susceptibilidade
aos carbapenemos, dezoito foram positivos para a presença da carbapenemase KPC-3. O
ácido borónico mostrou uma elevada sensibilidade e especificidade para a identificação da
carbapenemase KPC-3.
Na ausência da carbapenemase KPC-3, a resistência ou baixa susceptibilidade para os
carbapenemos é explicada pela presença de penicilinases CTX-M-15, cefalosporinases DHA-1
e/ou a ausência das porinas OmpK35 e OmpK36 da membrana externa. A associação da
cefalosporinase com a ausência das porinas confere um grau de resistência aos
carbapenemos mais elevado que com as penicilinases.
O gene blaKPC-3 está inserido num transposão composto do tipo Tn4401-like descrito por outros
autores. No entanto no nosso estudo encontramos quatro variantes não descritas e que se
caracterizaram pela deleção de regiões importantes na disseminação do gene blaKPC-3 ou
inserção de regiões no transposão. Os transposões têm capacidade de transposição replicativa
e a sua transferência entre géneros bacterianos diferentes é facilitada quando associados a
plasmídeos.
O gene blaKPC-3, associado a outros genes bla, encontrava-se em plasmídeos, com a mesma
origem de replicação, do grupo de incompatibilidade IncF, o que permitiu a disseminação entre
diferentes espécies bacterianas tais como: K. pneumoniae, K. oxytoca, E. aerogenes e E. coli.
O gene blaKPC-3 foi encontrado pela primeira vez em isolados com o sequence type ST14. Estas
estirpes multirresistentes de K. pneumoniae ST14 têm sido descritas com um elevado potencial
disseminativo e de virulência, aumentando o risco de transmissão destas bactérias,
frequentemente responsáveis por surtos hospitalares. Os isolados de K. pneumoniae eram
predominantemente do serótipo K2, um serótipo responsável por infecções invasivas
nomeadamente abcessos em órgãos como o fígado. Nestes isolados encontraram-se genes
fimH e/ou mrkD, os quais codificam para adesinas que conferem a habilidade de aderir às
células do hospedeiro, essencial para o desenvolvimento da infecção urinária.
A resistência a um largo espectro de antibióticos é um problema entre os isolados de
Enterobacteriaceae e os carbapenemos podem ser uma alternativa no tratamento de infecções
por estas bactérias. A emergência e disseminação de β-lactamases com actividade hidrolítica
para os carbapenemos contribuem para o aumento da multirresistência e para a falência
terapêutica de infecções graves constituindo uma ameaça à saúde pública

33
6. REFERÊNCIAS BIBLIOGRÁFICAS
1. World Health Organization (2002). Prevention of hospital-acquired infections: A practical guide
(WHO/CDS/CSR/EPH/2002.12, ed.).
2. Peleg, A. Y. & Hooper, D. C. (2010). Hospital-acquired infections due to gram-negative bacteria.
N Engl J Med 362, 1804-13.
3. Sousa, J. C. d. (2006). Manual de Antibióticos Antibacterianos (Pessoa, U. F., Ed.), Porto.
4. Pfeifer, Y., Cullik, A. & Witte, W. (2010). Resistance to cephalosporins and carbapenems in
Gram-negative bacterial pathogens. Int J Med Microbiol 300, 371-9.
5. Tenover, F. C. (2006). Mechanisms of antimicrobial resistance in bacteria. Am J Med 119, S3-10;
discussion S62-70.
6. Drawz, S. M. & Bonomo, R. A. (2010). Three decades of beta-lactamase inhibitors. Clin Microbiol
Rev 23, 160-201.
7. Pages, J. M. (2004). [Bacterial porin and antibiotic susceptibility]. Med Sci (Paris) 20, 346-51.
8. Nikaido, H. (2003). Molecular basis of bacterial outer membrane permeability revisited. Microbiol
Mol Biol Rev 67, 593-656.
9. Tsai, Y. K., Fung, C. P., Lin, J. C., Chen, J. H., Chang, F. Y., Chen, T. L. & Siu, L. K. (2011).
Klebsiella pneumoniae outer membrane porins OmpK35 and OmpK36 play roles in both
antimicrobial resistance and virulence. Antimicrob Agents Chemother 55, 1485-93.
10. Poole, K. (2004). Efflux-mediated multiresistance in Gram-negative bacteria. Clin Microbiol Infect
10, 12-26.
11. Cattoir, V. (2004). [Efflux-mediated antibiotics resistance in bacteria]. Pathol Biol (Paris) 52, 607-
16.
12. Strelkauskas, A., Strelkauskas, J. & Moszyk-Strelkauskas, D. (2010). Microbiology, a clinical
approach. 1st edit, Garland Science, Taylor & Francis Group, LLC, New York, USA.
13. Bush, K. & Jacoby, G. A. (2010). Updated Functional Classification of -Lacamases. Antimicrob
Agents Chemother 54, 969-976.
14. Ambler, R. P. (1980). The structure of beta-lactamases. Philos Trans R Soc Lond B Biol Sci 289,
321-31.
15. Bush, K., Jacoby, G. A. & Medeiros, A. A. (1995). A functional classification scheme for beta-
lactamases and its correlation with molecular structure. Antimicrob Agents Chemother 39, 1211-
33.
16. Thomson, K. S. (2010). Extended-spectrum-beta-lactamase, AmpC, and Carbapenemase issues.
J Clin Microbiol 48, 1019-25.
17. Abraham, E. P. & Chain, E. (1940). An enzyme from bacteria able to destroy penicillin. Nature
146, 837.
18. Datta, N. & Kontomichalou, P. (1965). Penicillinase synthesis controlled by infectious R factors in
Enterobacteriaceae. Nature 208, 239-41.
19. Medeiros, A. A. (1984). Beta-lactamases. Br Med Bull 40, 18-27.
20. Bradford, P. A. (2001). Extended-spectrum beta-lactamases in the 21st century: characterization,
epidemiology, and detection of this important resistance threat. Clin Microbiol Rev 14, 933-51,
table of contents.

REFERÊNCIAS BIBLIOGRÁFICAS
34
21. Barthelemy, M., Peduzzi, J. & Labia, R. (1985). [Distinction between the primary structures of
TEM-1 and TEM-2 beta-lactamases]. Ann Inst Pasteur Microbiol 136A, 311-21.
22. Heritage, J., M'Zali, F. H., Gascoyne-Binzi, D. & Hawkey, P. M. (1999). Evolution and spread of
SHV extended-spectrum beta-lactamases in gram-negative bacteria. J Antimicrob Chemother 44,
309-18.
23. Kliebe, C., Nies, B. A., Meyer, J. F., Tolxdorff-Neutzling, R. M. & Wiedemann, B. (1985). Evolution
of plasmid-coded resistance to broad-spectrum cephalosporins. Antimicrob Agents Chemother
28, 302-7.
24. Paterson, D. L. & Bonomo, R. A. (2005). Extended-spectrum beta-lactamases: a clinical update.
Clin Microbiol Rev 18, 657-86.
25. Sirot, D., Sirot, J., Labia, R., Morand, A., Courvalin, P., Darfeuille-Michaud, A., Perroux, R. &
Cluzel, R. (1987). Transferable resistance to third-generation cephalosporins in clinical isolates of
Klebsiella pneumoniae: identification of CTX-1, a novel beta-lactamase. J Antimicrob Chemother
20, 323-34.
26. Bonnet, R. (2004). Growing group of extended-spectrum beta-lactamases: the CTX-M enzymes.
Antimicrob Agents Chemother 48, 1-14.
27. Karim, A., Poirel, L., Nagarajan, S. & Nordmann, P. (2001). Plasmid-mediated extended-spectrum
beta-lactamase (CTX-M-3 like) from India and gene association with insertion sequence ISEcp1.
FEMS Microbiol Lett 201, 237-41.
28. Matthew, M. (1979). Plasmid-mediated beta-lactamases of Gram-negative bacteria: properties
and distribution. J Antimicrob Chemother 5, 349-58.
29. Hawkey, P. M. (2008). The growing burden of antimicrobial resistance. J Antimicrob Chemother
62 Suppl 1, i1-9.
30. Black, J. A., Moland, E. S. & Thomson, K. S. (2005). AmpC disk test for detection of plasmid-
mediated AmpC beta-lactamases in Enterobacteriaceae lacking chromosomal AmpC beta-
lactamases. J Clin Microbiol 43, 3110-3.
31. Bauernfeind, A., Chong, Y. & Schweighart, S. (1989). Extended broad spectrum beta-lactamase
in Klebsiella pneumoniae including resistance to cephamycins. Infection 17, 316-21.
32. Bauernfeind, A., Chong, Y. & Lee, K. (1998). Plasmid-encoded AmpC beta-lactamases: how far
have we gone 10 years after the discovery? Yonsei Med J 39, 520-5.
33. Barnaud, G., Arlet, G., Verdet, C., Gaillot, O., Lagrange, P. H. & Philippon, A. (1998). Salmonella
enteritidis: AmpC plasmid-mediated inducible beta-lactamase (DHA-1) with an ampR gene from
Morganella morganii. Antimicrob Agents Chemother 42, 2352-8.
34. Nordmann, P., Cuzon, G. & Naas, T. (2009). The real threat of Klebsiella pneumoniae
carbapenemase-producing bacteria. Lancet Infect Dis 9, 228-36.
35. Pournaras, S., Poulou, A. & Tsakris, A. (2010). Inhibitor-based methods for the detection of KPC
carbapenemase-producing Enterobacteriaceae in clinical practice by using boronic acid
compounds. J Antimicrob Chemother 65, 1319-21.
36. Poirel, L., Pitout, J. D. & Nordmann, P. (2007). Carbapenemases: molecular diversity and clinical
consequences. Future Microbiol 2, 501-12.
37. Sabath, L. D. & Abraham, E. P. (1966). Zinc as a cofactor for cephalosporinase from Bacillus
cereus 569. Biochem J 98, 11C-3C.

REFERÊNCIAS BIBLIOGRÁFICAS
35
38. Osano, E., Arakawa, Y., Wacharotayankun, R., Ohta, M., Horii, T., Ito, H., Yoshimura, F. & Kato,
N. (1994). Molecular characterization of an enterobacterial metallo beta-lactamase found in a
clinical isolate of Serratia marcescens that shows imipenem resistance. Antimicrob Agents
Chemother 38, 71-8.
39. Lauretti, L., Riccio, M. L., Mazzariol, A., Cornaglia, G., Amicosante, G., Fontana, R. & Rossolini,
G. M. (1999). Cloning and characterization of blaVIM, a new integron-borne metallo-beta-
lactamase gene from a Pseudomonas aeruginosa clinical isolate. Antimicrob Agents Chemother
43, 1584-90.
40. Cornaglia, G., Giamarellou, H. & Rossolini, G. M. (2011). Metallo-beta-lactamases: a last frontier
for beta-lactams? Lancet Infect Dis 11, 381-93.
41. Robin, F., Aggoune-Khinache, N., Delmas, J., Naim, M. & Bonnet, R. (2010). Novel VIM metallo-
beta-lactamase variant from clinical isolates of Enterobacteriaceae from Algeria. Antimicrob
Agents Chemother 54, 466-70.
42. Kumarasamy, K. K., Toleman, M. A., Walsh, T. R., Bagaria, J., Butt, F., Balakrishnan, R.,
Chaudhary, U., Doumith, M., Giske, C. G., Irfan, S., Krishnan, P., Kumar, A. V., Maharjan, S.,
Mushtaq, S., Noorie, T., Paterson, D. L., Pearson, A., Perry, C., Pike, R., Rao, B., Ray, U.,
Sarma, J. B., Sharma, M., Sheridan, E., Thirunarayan, M. A., Turton, J., Upadhyay, S., Warner,
M., Welfare, W., Livermore, D. M. & Woodford, N. (2010). Emergence of a new antibiotic
resistance mechanism in India, Pakistan, and the UK: a molecular, biological, and
epidemiological study. Lancet Infect Dis 10, 597-602.
43. Rolain, J. M., Parola, P. & Cornaglia, G. (2010). New Delhi metallo-beta-lactamase (NDM-1):
towards a new pandemia? Clin Microbiol Infect 16, 1699-701.
44. Walsh, T. R., Toleman, M. A., Poirel, L. & Nordmann, P. (2005). Metallo-beta-lactamases: the
quiet before the storm? Clin Microbiol Rev 18, 306-25.
45. Livermore, D. M. (2002). The impact of carbapenemases on antimicrobial development and
therapy. Curr Opin Investig Drugs 3, 218-24.
46. Giakkoupi, P., Tzouvelekis, L. S., Tsakris, A., Loukova, V., Sofianou, D. & Tzelepi, E. (2000). IBC-
1, a novel integron-associated class A beta-lactamase with extended-spectrum properties
produced by an Enterobacter cloacae clinical strain. Antimicrob Agents Chemother 44, 2247-53.
47. Poirel, L., Weldhagen, G. F., Naas, T., De Champs, C., Dove, M. G. & Nordmann, P. (2001).
GES-2, a class A beta-lactamase from Pseudomonas aeruginosa with increased hydrolysis of
imipenem. Antimicrob Agents Chemother 45, 2598-603.
48. Yigit, H., Queenan, A. M., Anderson, G. J., Domenech-Sanchez, A., Biddle, J. W., Steward, C. D.,
Alberti, S., Bush, K. & Tenover, F. C. (2001). Novel carbapenem-hydrolyzing beta-lactamase,
KPC-1, from a carbapenem-resistant strain of Klebsiella pneumoniae. Antimicrob Agents
Chemother 45, 1151-61.
49. Yigit, H., Queenan, A. M., Rasheed, J. K., Biddle, J. W., Domenech-Sanchez, A., Alberti, S.,
Bush, K. & Tenover, F. C. (2003). Carbapenem-resistant strain of Klebsiella oxytoca harboring
carbapenem-hydrolyzing beta-lactamase KPC-2. Antimicrob Agents Chemother 47, 3881-9.
50. Miriagou, V., Tzouvelekis, L. S., Rossiter, S., Tzelepi, E., Angulo, F. J. & Whichard, J. M. (2003).
Imipenem resistance in a Salmonella clinical strain due to plasmid-mediated class A
carbapenemase KPC-2. Antimicrob Agents Chemother 47, 1297-300.
51. Smith, M. E., Hanson, N. D., Herrera, V. L., Black, J. A., Lockhart, T. J., Hossain, A., Johnson, J.
A., Goering, R. V. & Thomson, K. S. (2003). Plasmid-mediated, carbapenem-hydrolysing beta-
lactamase, KPC-2, in Klebsiella pneumoniae isolates. J Antimicrob Chemother 51, 711-4.

REFERÊNCIAS BIBLIOGRÁFICAS
36
52. Woodford, N., Tierno, P. M., Jr., Young, K., Tysall, L., Palepou, M. F., Ward, E., Painter, R. E.,
Suber, D. F., Shungu, D., Silver, L. L., Inglima, K., Kornblum, J. & Livermore, D. M. (2004).
Outbreak of Klebsiella pneumoniae producing a new carbapenem-hydrolyzing class A beta-
lactamase, KPC-3, in a New York Medical Center. Antimicrob Agents Chemother 48, 4793-9.
53. Paton, R., Miles, R. S., Hood, J. & Amyes, S. G. (1993). ARI 1: beta-lactamase-mediated
imipenem resistance in Acinetobacter baumannii. Int J Antimicrob Agents 2, 81-7.
54. Poirel, L., Heritier, C., Tolun, V. & Nordmann, P. (2004). Emergence of oxacillinase-mediated
resistance to imipenem in Klebsiella pneumoniae. Antimicrob Agents Chemother 48, 15-22.
55. Castanheira, M., Deshpande, L. M., Mathai, D., Bell, J. M., Jones, R. N. & Mendes, R. E. (2011).
Early dissemination of NDM-1- and OXA-181-producing Enterobacteriaceae in Indian hospitals:
report from the SENTRY Antimicrobial Surveillance Program, 2006-2007. Antimicrob Agents
Chemother 55, 1274-8.
56. Poirel, L., Castanheira, M., Carrer, A., Rodriguez, C. P., Jones, R. N., Smayevsky, J. &
Nordmann, P. (2011). OXA-163, an OXA-48-Related Class D β-Lactamase with Extended Activity
Toward Expanded-Spectrum Cephalosporins. Antimicrob Agents Chemother 55, 2546-2551.
57. Bennett, P. M. (2008). Plasmid encoded antibiotic resistance: acquisition and transfer of antibiotic
resistance genes in bacteria. Br J Pharmacol 153 Suppl 1, S347-57.
58. Willey, J. M., Sherwood, L. M. & Woolverton, C. J. (2008). Microbiology. Seventh Edition edit,
McGraw-Hill, New York, USA.
59. Di Conza, J. A. & Gutkind, G. O. (2010). [Integrons: gene collectors]. Rev Argent Microbiol 42, 63-
78.
60. Levesque, C., Piche, L., Larose, C. & Roy, P. H. (1995). PCR mapping of integrons reveals
several novel combinations of resistance genes. Antimicrob Agents Chemother 39, 185-91.
61. Cuzon, G., Naas, T. & Nordmann, P. (2010). [KPC carbapenemases: what is at stake in clinical
microbiology?]. Pathol Biol (Paris) 58, 39-45.
62. Machado, P., Silva, A., Lito, L., Melo-Cristino, J. & Duarte, A. (2010). Emergence of Klebsiella
pneumoniae ST-11 producing KPC-3 carbaoenemase at a Lisbon hospital. Clin Microbiol Infect
16.
63. Endimiani, A., Carias, L. L., Hujer, A. M., Bethel, C. R., Hujer, K. M., Perez, F., Hutton, R. A., Fox,
W. R., Hall, G. S., Jacobs, M. R., Paterson, D. L., Rice, L. B., Jenkins, S. G., Tenover, F. C. &
Bonomo, R. A. (2008). Presence of plasmid-mediated quinolone resistance in Klebsiella
pneumoniae isolates possessing blaKPC in the United States. Antimicrob Agents Chemother 52,
2680-2.
64. Hossain, A., Ferraro, M. J., Pino, R. M., Dew, R. B., 3rd, Moland, E. S., Lockhart, T. J., Thomson,
K. S., Goering, R. V. & Hanson, N. D. (2004). Plasmid-mediated carbapenem-hydrolyzing
enzyme KPC-2 in an Enterobacter sp. Antimicrob Agents Chemother 48, 4438-40.
65. Rasheed, J. K., Biddle, J. W., Anderson, K. F., Washer, L., Chenoweth, C., Perrin, J., Newton, D.
W. & Patel, J. B. (2008). Detection of the Klebsiella pneumoniae carbapenemase type 2
Carbapenem-hydrolyzing enzyme in clinical isolates of Citrobacter freundii and K. oxytoca
carrying a common plasmid. J Clin Microbiol 46, 2066-9.
66. Naas, T., Cuzon, G., Villegas, M. V., Lartigue, M. F., Quinn, J. P. & Nordmann, P. (2008). Genetic
structures at the origin of acquisition of the beta-lactamase blaKPC gene. Antimicrob Agents
Chemother 52, 1257-63.

REFERÊNCIAS BIBLIOGRÁFICAS
37
67. Curiao, T., Morosini, M. I., Ruiz-Garbajosa, P., Robustillo, A., Baquero, F., Coque, T. M. &
Canton, R. (2010). Emergence of bla KPC-3-Tn4401a associated with a pKPN3/4-like plasmid
within ST384 and ST388 Klebsiella pneumoniae clones in Spain. J Antimicrob Chemother 65,
1608-14.
68. Gootz, T. D., Lescoe, M. K., Dib-Hajj, F., Dougherty, B. A., He, W., Della-Latta, P. & Huard, R. C.
(2009). Genetic organization of transposase regions surrounding blaKPC carbapenemase genes
on plasmids from Klebsiella strains isolated in a New York City hospital. Antimicrob Agents
Chemother 53, 1998-2004.
69. Cuzon, G., Naas, T. & Nordmann, P. (2011). Functional characterization of Tn4401, a Tn3-based
transposon involved in blaKPC gene mobilization. Antimicrob Agents Chemother.
70. Podschun, R. & Ullmann, U. (1998). Klebsiella spp. as nosocomial pathogens: epidemiology,
taxonomy, typing methods, and pathogenicity factors. Clin Microbiol Rev 11, 589-603.
71. Simoons-Smit, A. M., Verwey-van Vught, A. M., Kanis, I. Y. & MacLaren, D. M. (1984). Virulence
of Klebsiella strains in experimentally induced skin lesions in the mouse. J Med Microbiol 17, 67-
77.
72. Fang, C. T., Chuang, Y. P., Shun, C. T., Chang, S. C. & Wang, J. T. (2004). A novel virulence
gene in Klebsiella pneumoniae strains causing primary liver abscess and septic metastatic
complications. J Exp Med 199, 697-705.
73. Nassif, X., Honore, N., Vasselon, T., Cole, S. T. & Sansonetti, P. J. (1989). Positive control of
colanic acid synthesis in Escherichia coli by rmpA and rmpB, two virulence-plasmid genes of
Klebsiella pneumoniae. Mol Microbiol 3, 1349-59.
74. Schroll, C., Barken, K. B., Krogfelt, K. A. & Struve, C. (2010). Role of type 1 and type 3 fimbriae in
Klebsiella pneumoniae biofilm formation. BMC Microbiol 10, 179.
75. Bullen, J. J., Rogers, H. J. & Griffiths, E. (1978). Role of iron in bacterial infection. Curr Top
Microbiol Immunol 80, 1-35.
76. Wooldridge, K. G. & Williams, P. H. (1993). Iron uptake mechanisms of pathogenic bacteria.
FEMS Microbiol Rev 12, 325-48.
77. Miles, A. A. & Khimji, P. L. (1975). Enterobacterial chelators of iron: their occurrence, detection,
and relation to pathogenicity. J Med Microbiol 8, 477-90.
78. Nassif, X. & Sansonetti, P. J. (1986). Correlation of the virulence of Klebsiella pneumoniae K1
and K2 with the presence of a plasmid encoding aerobactin. Infect Immun 54, 603-8.
79. Albesa, I., Barberis, L. I., Pajaro, M. C., Farnochi, M. C. & Eraso, A. J. (1985). A thiol-activated
hemolysin in gram-negative bacteria. Can J Microbiol 31, 297-300.
80. Barberis, L. I., Eraso, A. J., Pajaro, M. C. & Albesa, I. (1986). Molecular weight determination and
partial characterization of Klebsiella pneumoniae hemolysins. Can J Microbiol 32, 884-8.
81. Albesa, I. (1989). Klebsiella pneumoniae haemolysin adsorption to red blood cells. J Appl
Bacteriol 67, 263-6.
82. Braun, V. & Focareta, T. (1991). Pore-forming bacterial protein hemolysins (cytolysins). Crit Rev
Microbiol 18, 115-58.
83. Yin-Ching, C., Jer-Horng, S., Ching-Nan, L. & Ming-Chung, C. (2002). Cloning of a gene
encoding a unique haemolysin from Klebsiella pneumoniae and its potential use as a species-
specific gene probe. Microb Pathog 33, 1-6.
84. Clinical and Laboratory Standards Institute (2010). Performance Standards for Antimicrobial
Susceptibility Testing; Twentieth Informational Supplement. M-100-S20 edit., Vol. M100-S20
vol.30. Clinical and Laboratory Standards Institute Wayne, PA.

REFERÊNCIAS BIBLIOGRÁFICAS
38
85. European Committee on Antimicrobial Susceptibility Testing (2011). Breakpoint tables for
interpretation of MICs and zone diameters. Version 1.3, January 5, 2011.
86. Manoharan, A., Chatterjee, S. & Mathai, D. (2010). Detection and characterization of metallo beta
lactamases producing Pseudomonas aeruginosa. Indian J Med Microbiol 28, 241-4.
87. Pasteran, F., Mendez, T., Guerriero, L., Rapoport, M. & Corso, A. (2009). Sensitive screening
tests for suspected class A carbapenemase production in species of Enterobacteriaceae. J Clin
Microbiol 47, 1631-9.
88. Madi, A., Lakhdari, O., Blottiere, H. M., Guyard-Nicodeme, M., Le Roux, K., Groboillot, A.,
Svinareff, P., Dore, J., Orange, N., Feuilloley, M. G. & Connil, N. (2010). The clinical
Pseudomonas fluorescens MFN1032 strain exerts a cytotoxic effect on epithelial intestinal cells
and induces Interleukin-8 via the AP-1 signaling pathway. BMC Microbiol 10, 215.
89. Grundmann, H. J., Towner, K. J., Dijkshoorn, L., Gerner-Smidt, P., Maher, M., Seifert, H. &
Vaneechoutte, M. (1997). Multicenter study using standardized protocols and reagents for
evaluation of reproducibility of PCR-based fingerprinting of Acinetobacter spp. J Clin Microbiol 35,
3071-7.
90. Adamus-Bialek, W., Wojtasik, A., Majchrzak, M., Sosnowski, M. & Parniewski, P. (2009). (CGG)4-
based PCR as a novel tool for discrimination of uropathogenic Escherichia coli strains:
comparison with enterobacterial repetitive intergenic consensus-PCR. J Clin Microbiol 47, 3937-
44.
91. Senda, K., Arakawa, Y., Ichiyama, S., Nakashima, K., Ito, H., Ohsuka, S., Shimokata, K., Kato, N.
& Ohta, M. (1996). PCR detection of metallo-beta-lactamase gene (blaIMP) in gram-negative rods
resistant to broad-spectrum beta-lactams. J Clin Microbiol 34, 2909-13.
92. Lee, K., Lim, J. B., Yum, J. H., Yong, D., Chong, Y., Kim, J. M. & Livermore, D. M. (2002).
bla(VIM-2) cassette-containing novel integrons in metallo-beta-lactamase-producing
Pseudomonas aeruginosa and Pseudomonas putida isolates disseminated in a Korean hospital.
Antimicrob Agents Chemother 46, 1053-8.
93. Pitout, J. D., Thomson, K. S., Hanson, N. D., Ehrhardt, A. F., Moland, E. S. & Sanders, C. C.
(1998). beta-Lactamases responsible for resistance to expanded-spectrum cephalosporins in
Klebsiella pneumoniae, Escherichia coli, and Proteus mirabilis isolates recovered in South Africa.
Antimicrob Agents Chemother 42, 1350-4.
94. Conceicao, T., Brizio, A., Duarte, A., Lito, L. M., Cristino, J. M. & Salgado, M. J. (2005). First
description of CTX-M-15-producing Klebsiella pneumoniae in Portugal. Antimicrob Agents
Chemother 49, 477-8.
95. Yan, J. J., Ko, W. C., Jung, Y. C., Chuang, C. L. & Wu, J. J. (2002). Emergence of Klebsiella
pneumoniae isolates producing inducible DHA-1 beta-lactamase in a university hospital in
Taiwan. J Clin Microbiol 40, 3121-6.
96. Navarro, F., Perez-Trallero, E., Marimon, J. M., Aliaga, R., Gomariz, M. & Mirelis, B. (2001).
CMY-2-producing Salmonella enterica, Klebsiella pneumoniae, Klebsiella oxytoca, Proteus
mirabilis and Escherichia coli strains isolated in Spain (October 1999-December 2000). J
Antimicrob Chemother 48, 383-9.
97. Hooper, D. C. (2000). Mechanisms of action and resistance of older and newer fluoroquinolones.
Clin Infect Dis 31 Suppl 2, S24-8.
98. Nordmann, P. & Poirel, L. (2005). Emergence of plasmid-mediated resistance to quinolones in
Enterobacteriaceae. J Antimicrob Chemother 56, 463-9.

REFERÊNCIAS BIBLIOGRÁFICAS
39
99. Padilla, E., Llobet, E., Domenech-Sanchez, A., Martinez-Martinez, L., Bengoechea, J. A. &
Alberti, S. (2010). Klebsiella pneumoniae AcrAB efflux pump contributes to antimicrobial
resistance and virulence. Antimicrob Agents Chemother 54, 177-83.
100. Cambau, E., Lascols, C., Sougakoff, W., Bebear, C., Bonnet, R., Cavallo, J. D., Gutmann, L.,
Ploy, M. C., Jarlier, V., Soussy, C. J. & Robert, J. (2006). Occurrence of qnrA-positive clinical
isolates in French teaching hospitals during 2002-2005. Clin Microbiol Infect 12, 1013-20.
101. Jacoby, G. A., Walsh, K. E., Mills, D. M., Walker, V. J., Oh, H., Robicsek, A. & Hooper, D. C.
(2006). qnrB, another plasmid-mediated gene for quinolone resistance. Antimicrob Agents
Chemother 50, 1178-82.
102. Chen, Y. T., Shu, H. Y., Li, L. H., Liao, T. L., Wu, K. M., Shiau, Y. R., Yan, J. J., Su, I. J., Tsai, S.
F. & Lauderdale, T. L. (2006). Complete nucleotide sequence of pK245, a 98-kilobase plasmid
conferring quinolone resistance and extended-spectrum-beta-lactamase activity in a clinical
Klebsiella pneumoniae isolate. Antimicrob Agents Chemother 50, 3861-6.
103. Ambrozic Avgustin, J., Keber, R., Zerjavic, K., Orazem, T. & Grabnar, M. (2007). Emergence of
the quinolone resistance-mediating gene aac(6')-Ib-cr in extended-spectrum-beta-lactamase-
producing Klebsiella isolates collected in Slovenia between 2000 and 2005. Antimicrob Agents
Chemother 51, 4171-3.
104. Carattoli, A., Bertini, A., Villa, L., Falbo, V., Hopkins, K. L. & Threlfall, E. J. (2005). Identification of
plasmids by PCR-based replicon typing. J Microbiol Methods 63, 219-28.
105. Yu, W. L., Ko, W. C., Cheng, K. C., Lee, C. C., Lai, C. C. & Chuang, Y. C. (2008). Comparison of
prevalence of virulence factors for Klebsiella pneumoniae liver abscesses between isolates with
capsular K1/K2 and non-K1/K2 serotypes. Diagn Microbiol Infect Dis 62, 1-6.
106. Turton, J. F., Perry, C., Elgohari, S. & Hampton, C. V. (2010). PCR characterization and typing of
Klebsiella pneumoniae using capsular type-specific, variable number tandem repeat and
virulence gene targets. J Med Microbiol 59, 541-7.
107. Bilavsky, E., Schwaber, M. J. & Carmeli, Y. (2010). How to stem the tide of carbapenemase-
producing enterobacteriaceae?: proactive versus reactive strategies. Curr Opin Infect Dis 23,
327-31.
108. Cohen Stuart, J. & Leverstein-Van Hall, M. A. (2010). Guideline for phenotypic screening and
confirmation of carbapenemases in Enterobacteriaceae. Int J Antimicrob Agents 36, 205-10.
109. Beesley, T., Gascoyne, N., Knott-Hunziker, V., Petursson, S., Waley, S. G., Jaurin, B. &
Grundstrom, T. (1983). The inhibition of class C beta-lactamases by boronic acids. Biochem J
209, 229-33.
110. Pfaller, M. A., Acar, J., Jones, R. N., Verhoef, J., Turnidge, J. & Sader, H. S. (2001). Integration of
molecular characterization of microorganisms in a global antimicrobial resistance surveillance
program. Clin Infect Dis 32 Suppl 2, S156-67.
111. Inês, A. (2007). Abordagem polifásica na caracterização e selecção de bactérias do ácido láctico
de vinhos da Região Demarcada do Douro, Universidade de Trás-os-Montes e Alto Douro.
112. Diancourt, L., Passet, V., Verhoef, J., Grimont, P. A. & Brisse, S. (2005). Multilocus sequence
typing of Klebsiella pneumoniae nosocomial isolates. J Clin Microbiol 43, 4178-82.
113. Maiden, M. C., Bygraves, J. A., Feil, E., Morelli, G., Russell, J. E., Urwin, R., Zhang, Q., Zhou, J.,
Zurth, K., Caugant, D. A., Feavers, I. M., Achtman, M. & Spratt, B. G. (1998). Multilocus
sequence typing: a portable approach to the identification of clones within populations of
pathogenic microorganisms. Proc Natl Acad Sci U S A 95, 3140-5.

REFERÊNCIAS BIBLIOGRÁFICAS
40
114. Vimont, S., Mnif, B., Fevre, C. & Brisse, S. (2008). Comparison of PFGE and multilocus sequence
typing for analysis of Klebsiella pneumoniae isolates. J Med Microbiol 57, 1308-10.
115. Poirel, L., Al Maskari, Z., Al Rashdi, F., Bernabeu, S. & Nordmann, P. (2011). NDM-1-producing
Klebsiella pneumoniae isolated in the Sultanate of Oman. J Antimicrob Chemother 66, 304-6.
116. Oteo, J., Cuevas, O., Lopez-Rodriguez, I., Banderas-Florido, A., Vindel, A., Perez-Vazquez, M.,
Bautista, V., Arroyo, M., Garcia-Caballero, J., Marin-Casanova, P., Gonzalez-Sanz, R., Fuentes-
Gomez, V., Ona-Compan, S., Garcia-Cobos, S. & Campos, J. (2009). Emergence of CTX-M-15-
producing Klebsiella pneumoniae of multilocus sequence types 1, 11, 14, 17, 20, 35 and 36 as
pathogens and colonizers in newborns and adults. J Antimicrob Chemother 64, 524-8.
117. Bergeron, M. G. & Ouellette, M. (1998). Preventing antibiotic resistance using rapid DNA-based
diagnostic tests. Infect Control Hosp Epidemiol 19, 560-4.
118. Tsakris, A., Themeli-Digalaki, K., Poulou, A., Vrioni, G., Voulgari, E., Koumaki, V., Agodi, A.,
Pournaras, S. & Sofianou, D. (2011). Comparative Evaluation of Combined-Disk Tests Using
Different Boronic Acid Compounds for Detection of Klebsiella pneumoniae Carbapenemase-
Producing Enterobacteriaceae Clinical Isolates. J Clin Microbiol 49, 2804-9.
119. Kitchel, B., Rasheed, J. K., Endimiani, A., Hujer, A. M., Anderson, K. F., Bonomo, R. A. & Patel,
J. B. (2010). Genetic factors associated with elevated carbapenem resistance in KPC-producing
Klebsiella pneumoniae. Antimicrob Agents Chemother 54, 4201-7.
120. Jacoby, G. A., Mills, D. M. & Chow, N. (2004). Role of beta-lactamases and porins in resistance
to ertapenem and other beta-lactams in Klebsiella pneumoniae. Antimicrob Agents Chemother
48, 3203-6.
121. Domenech-Sanchez, A., Martinez-Martinez, L., Hernandez-Alles, S., del Carmen Conejo, M.,
Pascual, A., Tomas, J. M., Alberti, S. & Benedi, V. J. (2003). Role of Klebsiella pneumoniae
OmpK35 porin in antimicrobial resistance. Antimicrob Agents Chemother 47, 3332-5.
122. Song, W., Suh, B., Choi, J. Y., Jeong, S. H., Jeon, E. H., Lee, Y. K., Hong, S. G. & Lee, K. (2009).
In vivo selection of carbapenem-resistant Klebsiella pneumoniae by OmpK36 loss during
meropenem treatment. Diagn Microbiol Infect Dis 65, 447-9.
123. Doumith, M., Ellington, M. J., Livermore, D. M. & Woodford, N. (2009). Molecular mechanisms
disrupting porin expression in ertapenem-resistant Klebsiella and Enterobacter spp. clinical
isolates from the UK. J Antimicrob Chemother 63, 659-67.
124. Simpson, I. N., Harper, P. B. & O'Callaghan, C. H. (1980). Principal beta-lactamases responsible
for resistance to beta-lactam antibiotics in urinary tract infections. Antimicrob Agents Chemother
17, 929-36.
125. Bush, K. (2010). Bench-to-bedside review: The role of beta-lactamases in antibiotic-resistant
Gram-negative infections. Crit Care 14, 224.
126. Giakkoupi, P., Tambic-Andrasevic, A., Vourli, S., Skrlin, J., Sestan-Crnek, S., Tzouvelekis, L. S. &
Vatopoulos, A. C. (2006). Transferable DHA-1 cephalosporinase in Escherichia coli. Int J
Antimicrob Agents 27, 77-80.
127. Mammeri, H., Guillon, H., Eb, F. & Nordmann, P. (2010). Phenotypic and biochemical comparison
of the carbapenem-hydrolyzing activities of five plasmid-borne AmpC beta-lactamases.
Antimicrob Agents Chemother 54, 4556-60.
128. Jiang, Y., Yu, D., Wei, Z., Shen, P., Zhou, Z. & Yu, Y. (2010). Complete nucleotide sequence of
Klebsiella pneumoniae multidrug resistance plasmid pKP048, carrying blaKPC-2, blaDHA-1,
qnrB4, and armA. Antimicrob Agents Chemother 54, 3967-9.

REFERÊNCIAS BIBLIOGRÁFICAS
41
129. Yang, Q., Wang, H., Sun, H., Chen, H., Xu, Y. & Chen, M. (2010). Phenotypic and genotypic
characterization of Enterobacteriaceae with decreased susceptibility to carbapenems: results
from large hospital-based surveillance studies in China. Antimicrob Agents Chemother 54, 573-7.
130. Chen, J. H., Siu, L. K., Fung, C. P., Lin, J. C., Yeh, K. M., Chen, T. L., Tsai, Y. K. & Chang, F. Y.
(2010). Contribution of outer membrane protein K36 to antimicrobial resistance and virulence in
Klebsiella pneumoniae. J Antimicrob Chemother 65, 986-90.
131. Paterson, D. L., Ko, W. C., Von Gottberg, A., Mohapatra, S., Casellas, J. M., Goossens, H.,
Mulazimoglu, L., Trenholme, G., Klugman, K. P., Bonomo, R. A., Rice, L. B., Wagener, M. M.,
McCormack, J. G. & Yu, V. L. (2004). International prospective study of Klebsiella pneumoniae
bacteremia: implications of extended-spectrum beta-lactamase production in nosocomial
Infections. Ann Intern Med 140, 26-32.
132. Kassis-Chikhani, N., Decre, D., Ichai, P., Sengelin, C., Geneste, D., Mihaila, L., Dussaix, E. &
Arlet, G. (2010). Outbreak of Klebsiella pneumoniae producing KPC-2 and SHV-12 in a French
hospital. J Antimicrob Chemother 65, 1539-40.
133. Nuesch-Inderbinen, M. T., Kayser, F. H. & Hachler, H. (1997). Survey and molecular genetics of
SHV beta-lactamases in Enterobacteriaceae in Switzerland: two novel enzymes, SHV-11 and
SHV-12. Antimicrob Agents Chemother 41, 943-9.
134. Martinez-Martinez, L., Pascual, A., Hernandez-Alles, S., Alvarez-Diaz, D., Suarez, A. I., Tran, J.,
Benedi, V. J. & Jacoby, G. A. (1999). Roles of beta-lactamases and porins in activities of
carbapenems and cephalosporins against Klebsiella pneumoniae. Antimicrob Agents Chemother
43, 1669-73.
135. Bradford, P. A., Urban, C., Mariano, N., Projan, S. J., Rahal, J. J. & Bush, K. (1997). Imipenem
resistance in Klebsiella pneumoniae is associated with the combination of ACT-1, a plasmid-
mediated AmpC beta-lactamase, and the foss of an outer membrane protein. Antimicrob Agents
Chemother 41, 563-9.
136. Cao, V. T., Arlet, G., Ericsson, B. M., Tammelin, A., Courvalin, P. & Lambert, T. (2000).
Emergence of imipenem resistance in Klebsiella pneumoniae owing to combination of plasmid-
mediated CMY-4 and permeability alteration. J Antimicrob Chemother 46, 895-900.
137. Garcia-Sureda, L., Domenech-Sanchez, A., Barbier, M., Juan, C., Gasco, J. & Alberti, S. (2011).
OmpK26, a novel porin associated to carbapenem resistance in Klebsiella pneumoniae.
Antimicrob Agents Chemother.
138. Marcade, G., Deschamps, C., Boyd, A., Gautier, V., Picard, B., Branger, C., Denamur, E. & Arlet,
G. (2009). Replicon typing of plasmids in Escherichia coli producing extended-spectrum beta-
lactamases. J Antimicrob Chemother 63, 67-71.
139. Chia, J. H., Siu, L. K., Su, L. H., Lin, H. S., Kuo, A. J., Lee, M. H. & Wu, T. L. (2009). Emergence
of carbapenem-resistant Escherichia coli in Taiwan: resistance due to combined CMY-2
production and porin deficiency. J Chemother 21, 621-6.
140. Ahmed, A. M. & Shimamoto, T. (2008). Emergence of a cefepime- and cefpirome-resistant
Citrobacter freundii clinical isolate harbouring a novel chromosomally encoded AmpC beta-
lactamase, CMY-37. Int J Antimicrob Agents 32, 256-61.
141. Mainardi, J. L., Mugnier, P., Coutrot, A., Buu-Hoi, A., Collatz, E. & Gutmann, L. (1997).
Carbapenem resistance in a clinical isolate of Citrobacter freundii. Antimicrob Agents Chemother
41, 2352-4.
142. Strahilevitz, J., Jacoby, G. A., Hooper, D. C. & Robicsek, A. (2009). Plasmid-mediated quinolone
resistance: a multifaceted threat. Clin Microbiol Rev 22, 664-89.

REFERÊNCIAS BIBLIOGRÁFICAS
42
143. Park, Y. J., Yu, J. K., Lee, S., Oh, E. J. & Woo, G. J. (2007). Prevalence and diversity of qnr
alleles in AmpC-producing Enterobacter cloacae, Enterobacter aerogenes, Citrobacter freundii
and Serratia marcescens: a multicentre study from Korea. J Antimicrob Chemother 60, 868-71.
144. Shen, P., Wei, Z., Jiang, Y., Du, X., Ji, S., Yu, Y. & Li, L. (2009). Novel genetic environment of the
carbapenem-hydrolyzing beta-lactamase KPC-2 among Enterobacteriaceae in China. Antimicrob
Agents Chemother 53, 4333-8.
145. Cuzon, G., Naas, T., Truong, H., Villegas, M. V., Wisell, K. T., Carmeli, Y., Gales, A. C., Venezia,
S. N., Quinn, J. P. & Nordmann, P. (2010). Worldwide diversity of Klebsiella pneumoniae that
produce beta-lactamase blaKPC-2 gene. Emerg Infect Dis 16, 1349-56.
146. Leavitt, A., Chmelnitsky, I., Ofek, I., Carmeli, Y. & Navon-Venezia, S. (2010). Plasmid pKpQIL
encoding KPC-3 and TEM-1 confers carbapenem resistance in an extremely drug-resistant
epidemic Klebsiella pneumoniae strain. J Antimicrob Chemother 65, 243-8.
147. Lewin, B. (2004). Genes VIII, Pearson Prentice Hall, USA.
148. Smith, M. A. & Bidochka, M. J. (1998). Bacterial fitness and plasmid loss: the importance of
culture conditions and plasmid size. Can J Microbiol 44, 351-5.
149. Kabha, K., Nissimov, L., Athamna, A., Keisari, Y., Parolis, H., Parolis, L. A., Grue, R. M.,
Schlepper-Schafer, J., Ezekowitz, A. R., Ohman, D. E. & et al. (1995). Relationships among
capsular structure, phagocytosis, and mouse virulence in Klebsiella pneumoniae. Infect Immun
63, 847-52.
150. Struve, C., Bojer, M. & Krogfelt, K. A. (2008). Characterization of Klebsiella pneumoniae type 1
fimbriae by detection of phase variation during colonization and infection and impact on virulence.
Infect Immun 76, 4055-65.
151. Struve, C., Bojer, M. & Krogfelt, K. A. (2009). Identification of a conserved chromosomal region
encoding Klebsiella pneumoniae type 1 and type 3 fimbriae and assessment of the role of
fimbriae in pathogenicity. Infect Immun 77, 5016-24.
152. Yu, V. L., Hansen, D. S., Ko, W. C., Sagnimeni, A., Klugman, K. P., von Gottberg, A., Goossens,
H., Wagener, M. M. & Benedi, V. J. (2007). Virulence characteristics of Klebsiella and clinical
manifestations of K. pneumoniae bloodstream infections. Emerg Infect Dis 13, 986-93.

43
7. ANEXOS
Tabela AI: Susceptibilidade aos antibióticos de vinte e seis isolados de Enterobacteriaceae.
ID Nº Doente Estirpe amc ctx fox caz fep imp mem atm cip lvx gm tgc ft fos
2948 1 K. pneumoniae 6 13 11 6 13 11 9 7 19 18 13 12 16 22
2954
2
K. pneumoniae 7 14 12 6 15 11 13 8 21 18 12 12 16 21
2958 K. pneumoniae 7 9 6 6 6 18 16 6 8 7 10 14 6 20
2959 K. pneumoniae 7 16 15 7 14 19 13 8 21 18 13 12 17 22
2955 2 K. pneumoniae 8 13 17 9 16 11 15 11 26 22 10 16 18 21
2949 4
K. pneumoniae 6 12 10 6 19 9 6 8 18 6 12 10 17 22
2999 K. pneumoniae 7 16 16 8 13 13 12 8 9 10 12 15 9 22
3075 5 K. pneumoniae 8 16 13 9 11 19 13 7 23 20 14 13 16 23
3084 6
K. pneumoniae 7 13 10 8 9 10 7 6 21 21 13 13 15 24
3090 K. pneumoniae 8 15 14 8 12 15 11 7 21 21 13 12 16 24
3108 7 K. pneumoniae 8 17 20 9 16 16 11 6 26 26 14 16 20 20
3116 8 K. pneumoniae 8 16 16 7 13 13 11 6 21 20 6 17 22 25
94182 9
K. pneumoniae 9 16 13 8 14 14 12 6 22 18 23 10 15 23
3191 K. pneumoniae 11 30 27 13 30 25 25 32 15 23 18 15 9 21
3030 10 K. pneumoniae 11 8 6 6 10 20 16 6 6 7 8 20 6 18
3046 11 K. pneumoniae 7 11 11 6 8 14 9 6 22 19 13 12 14 24
3034
12
K. oxytoca 6 7 6 6 6 6 6 6 21 22 6 16 21 21
3042 K. oxytoca 6 15 11 6 13 17 10 6 22 22 6 16 24 27
3051 K. oxytoca 7 11 9 6 14 16 11 6 22 22 6 18 25 26
3066 13 K. oxytoca 11 10 7 6 22 25 23 8 6 6 6 18 8 21
3074 14 K. oxytoca 17 7 20 9 12 24 19 8 6 6 6 13 6 20
3016 15 E. aerogenes 8 16 14 7 12 15 10 6 26 25 12 18 21 24
3023 16 E. coli 7 14 13 6 10 14 10 6 26 20 15 12 16 25
3052 17 E. coli 10 18 19 12 14 18 15 12 21 20 11 21 24 29
3192 18
C. freundii 7 16 6 11 20 23 23 11 8 18 11 17 24 30
3193 C. freundii 6 18 6 14 19 10 15 8 15 18 11 18 20 30
amc, amoxicilina/ácido clavulânico; fox, cefoxitina; ctx, cefotaxima; caz, ceftazidima; fep, cefepima; atm, aztreonamo;
imp, imipenemo; mem, meropenemo; cip, ciprofloxacina; lvx, levofloxacina; gm, gentamicina; tgc, tigeciclina; ft,
nitrofurantoína; fos, fosfomicina.

ANEXOS
44
Tabela AII: Características genéticas dos isolados em estudo
ID Nº Doente Estirpe Mecanismo de resistência Plasmídeo Transposão Integrão
2948 1 K. pneumoniae KPC-3, TEM-1, aac(6')-Ib-cr FIA, F Tn4401a -
2954
2
K. pneumoniae KPC-3, TEM-1, aac(6')-Ib-cr A/C, F Tn4401 -
2958 K. pneumoniae KPC-3, SHV-1 FIC, F Tn4401c -
2959 K. pneumoniae KPC-3, SHV-1, TEM-1, perda OmpK35 - Tn4401a -
2955 3 K. pneumoniae KPC-3, SHV-1, TEM-1 A/C, F Tn4401 -
2949 4
K. pneumoniae KPC-3, TEM-1 F Tn4401a -
2999 K. pneumoniae KPC-3, SHV-35, TEM-1, aac(6')-Ib-cr FIA, F Tn4401 -
3075 5 K. pneumoniae KPC-3, TEM-1 F Tn4401 -
3084 6
K. pneumoniae KPC-3, SHV-1, TEM-1 F Tn4401b -
3090 K. pneumoniae SHV-1, TEM-1, aac(6')-Ib-cr, perda OmpK35 A/C, F - -
3108 7 K. pneumoniae KPC-3 - Tn4401d -
3116 8 K. pneumoniae KPC-3, QnrB1 - Tn4401a -
94182 9
K. pneumoniae KPC-3, TEM-1, aac(6')-Ib-cr - Tn4401 -
3191 K. pneumoniae Perda OmpK35 e OmpK36 - - -
3030 10 K. pneumoniae TEM-1, DHA-1, QnrB4 FIA - -
3046 11 K. pneumoniae KPC-3, aac(6')-Ib-cr, perda OmpK35 F Tn4401a -
3034
12
K. oxytoca KPC-3, TEM-1, DHA-1, QnrB4, perda OmpK35 HI1, HI2, FIC, F Tn4401 dhfrVII-aadA4
3042 K. oxytoca KPC-3, TEM-1, DHA-1, QnrB4, perda OmpK35 HI1, HI2, FIC, F Tn4401 dfrA17
3051 K. oxytoca KPC-3, SHV-12, TEM-1, DHA-1, QnrB4, perda da OmpK35 HI1, HI2, FIC, F Tn4401a -
3066 13 K. oxytoca DHA-1, QnrB4, aac(6')-Ib-cr, perda OmpK36 FIA, A/C - -
3074 14 K. oxytoca SHV-11, CTX-M-15, QnrB1, aac(6')-Ib-cr, perda OmpK36 FIA, A/C - -
3016 15 E. aerogenes KPC-3, TEM-1, aac(6')-Ib-cr HI1, F Tn4401b -
3023 16 E. coli KPC-3, SHV-1, TEM-1 F Tn4401 -
3052 17 E. coli TEM-1, QnrS1 FIA, F - -
3192 18
C. freundii CMY-47, DHA-1, QnrB4, aac(6')-Ib-cr HI1, FIC, FIIA - dfrA12-orfF-aadA2a
3193 C. freundii CMY-47, DHA-1, QnrB4 HI1, FIC - dfrA12-orfF-aadA2a

ANEXOS
45


















