
MENSAJE BIOQUÍMICO - Taller de Actualización …tab.facmed.unam.mx/files/5-GARFIAS-2017.pdfen las...
12
Memoria del 44º Taller de Actualización Bioquímica, Facultad de Medicina; UNAM La nucleolina como potencial blanco terapéutico en la neovascularización aberrante Nucleolin as a potential therapeutic target for aberrant neovascularization Alvarado Hernández, Beatriz 1, 2 ; Vivanco Rojas, Óscar 1,2 ; Quiroz Mercado, Joaquín 3 y Garfias Becerra, Yonathan Omar 1,2 * 1. Coordinación de Inmunología, Departamento de Bioquímica, Facultad de Medicina; UNAM. 2. Instituto de Oftalmología, Fundación Conde de Valenciana IAP. Unidad periférica de la Facultad de Medicina. 3. Hospital Veterinario de Especialidades del Departamento de Medicina, Cirugía y Zootecnia para pequeñas especies, Facultad de Medicina Veterinaria y Zootecnia, UNAM. *Correspondencia. Unidad de Investigación, Instituto de Oftalmología, Fundación Conde de Valenciana IAP, Chimalpopoca 14, Col. Obrera, Cuauhtémoc, Ciudad de México, CP 06800. Tel: +52(55)54421700 Ext.3134, [email protected] Versión electrónica en http://tab.facmed.unam.mx MENSAJE BIOQUÍMICO Mens. Bioquim. 41 (2017) 37-48 Resumen La nucleolina es una proteína altamente conservada en las células eucariotas y es una de las proteínas más abundantes del nucléolo. Adicionalmente, esta proteína se encuentra distribuida en el núcleo (donde representa el 10% de todas las proteínas presentes en este organelo), citoplasma y superficie celular. Aunque su localización es mayoritariamente en el núcleo y por ello su principal función se desarrolla en la biogénesis del ribosoma, en los últimos años se ha estudiado con gran interés a la nucleolina que se expresa en la superficie celular, ya que ésta se expresa de manera diferencial, es decir, se expresa solo en aquellas células que se encuentran en proliferación, como sucede tanto en la neovascularización o angiogénesis como en procesos tumorales; se ha demostrado que hay una expresión diferencial de la nucleolina en la superficie de aquellas células endoteliales que forman neovasos aberrantes, además se ha demostrado que la localización de la nucleolina en la superficie, está mediada por el factor de crecimiento vascular endotelial y proteínas de citoesqueleto. Por otro lado también se ha descrito que la neovascularización puede revertirse cuando se bloquea a la nucleolina de superficie con algunos de sus ligandos sintéticos como el psudopéptido HB-19 o el aptámero AS1411. Finalmente, en este capítulo también se describen algunos hallazgos sobre la expresión de la nucleolina Abstract Nucleolin is a protein highly conserved among eukaryotic cells and it is one of the more abundant proteins in the nucleolus, which accounts for approximately 10% of the protein content within the nucleolus; also, it has been reported that nucleolin is located in the nucleus, cytoplasm and cell surface, as well. Due to its nuclear localization nucleolin’s main function is in the ribosome biogenesis. It has recently been studied with great interest the cell surface nucleolin, because, it is only expressed in proliferating cells, like neovascularization or angiogenesis processes as well as tumorigenesis. It has been shown that there is a differential expression of surface nucleolin in those endothelial cells in aberrant new vessels, also it has been shown that its cell surface localization is dependent on vascular endothelial grown factor and cytoskeleton proteins. On the other hand, it is shown that new vessels can be inhibited when nucleolin is blocked with some artificial ligands, such as HB-19 pseudopeptide and AS1411 aptamers. At the end of this chapter, some evidences of nucleolin cell surface expression on corneal angiogenesis are discussed and we propose the cell surface nucleolin as a novel target for the treatment of neoangiogenesis diseases.
Transcript of MENSAJE BIOQUÍMICO - Taller de Actualización …tab.facmed.unam.mx/files/5-GARFIAS-2017.pdfen las...

Memoria del 44º Taller de Actualización Bioquímica, Facultad de Medicina; UNAM
La nucleolina como potencial blanco terapéutico en la
neovascularización aberrante
Nucleolin as a potential therapeutic target for aberrant neovascularization
Alvarado Hernández, Beatriz1, 2; Vivanco Rojas, Óscar1,2; Quiroz Mercado, Joaquín3 y Garfias Becerra, Yonathan Omar 1,2 *
1. Coordinación de Inmunología, Departamento de Bioquímica, Facultad de Medicina; UNAM. 2. Instituto de Oftalmología, Fundación Conde de Valenciana IAP. Unidad periférica de la Facultad de Medicina.
3. Hospital Veterinario de Especialidades del Departamento de Medicina, Cirugía y Zootecnia para pequeñas especies, Facultad de
Medicina Veterinaria y Zootecnia, UNAM. *Correspondencia. Unidad de Investigación, Instituto de Oftalmología, Fundación Conde de Valenciana IAP, Chimalpopoca 14, Col.
Obrera, Cuauhtémoc, Ciudad de México, CP 06800. Tel: +52(55)54421700 Ext.3134, [email protected]
Versión electrónica en http://tab.facmed.unam.mx
MENSAJE BIOQUÍMICO Mens. Bioquim. 41 (2017) 37-48
Resumen La nucleolina es una proteína altamente conservada en las células eucariotas y es una de las proteínas más abundantes del nucléolo. Adicionalmente, esta proteína se encuentra distribuida en el núcleo (donde representa el 10% de todas las proteínas presentes en este organelo), citoplasma y superficie celular. Aunque su localización es mayoritariamente en el núcleo y por ello su principal función se desarrolla en la biogénesis del ribosoma, en los últimos años se ha estudiado con gran interés a la nucleolina que se expresa en la superficie celular, ya que ésta se expresa de manera diferencial, es decir, se expresa solo en aquellas células que se encuentran en proliferación, como sucede tanto en la neovascularización o angiogénesis como en procesos tumorales; se ha demostrado que hay una expresión diferencial de la nucleolina en la superficie de aquellas células endoteliales que forman neovasos aberrantes, además se ha demostrado que la localización de la nucleolina en la superficie, está mediada por el factor de crecimiento vascular endotelial y proteínas de citoesqueleto. Por otro lado también se ha descrito que la neovascularización puede revertirse cuando se bloquea a la nucleolina de superficie con algunos de sus ligandos sintéticos como el psudopéptido HB-19 o el aptámero AS1411. Finalmente, en este capítulo también se describen algunos hallazgos sobre la expresión de la nucleolina
Abstract Nucleolin is a protein highly conserved among eukaryotic cells and it is one of the more abundant proteins in the nucleolus, which accounts for approximately 10% of the protein content within the nucleolus; also, it has been reported that nucleolin is located in the nucleus, cytoplasm and cell surface, as well. Due to its nuclear localization nucleolin’s main function is in the ribosome biogenesis. It has recently been studied with great interest the cell surface nucleolin, because, it is only expressed in proliferating cells, like neovascularization or angiogenesis processes as well as tumorigenesis. It has been shown that there is a differential expression of surface nucleolin in those endothelial cells in aberrant new vessels, also it has been shown that its cell surface localization is dependent on vascular endothelial grown factor and cytoskeleton proteins. On the other hand, it is shown that new vessels can be inhibited when nucleolin is blocked with some artificial ligands, such as HB-19 pseudopeptide and AS1411 aptamers. At the end of this chapter, some evidences of nucleolin cell surface expression on corneal angiogenesis are discussed and we propose the cell surface nucleolin as a novel target for the treatment of neoangiogenesis diseases.

Alvarado-Hernández et al. Mens. Bioquim. 41 (2017): 37 - 48
© 2017 Mensaje Bioquímico. Todos los derechos reservados. ISSN-0188-137X Comité Editorial: Cárdenas Monroy, C.A.; González Andrade, M.; Lara Lemus, R.; Martínez González, J.J.; Molina Jijón, E.; Torres
Durán, P.V. Publicado por el Departamento de Bioquímica de la Facultad de Medicina; UNAM.
38
Genética y biología molecular de la nucleolina
La nucleolina (NCL) es una de las proteínas más abundantes del núcleo, que se expresa de manera constitutiva en altos niveles y es esencial en la biogénesis del ribosoma. Fue descrita por primera vez en 1973 en el hígado de rata [1], es una proteína de células eucariotas muy conservada durante la evolución, ya que podemos encontrarla desde vegetales [2,3] hasta el humano [4]. La NCL de humano se encuentra codificada en el cromosoma 2, la secuencia nucleotídica es de aproximadamente 11 kb y está organizada en 14 exones y 13 intrones [5].
La región promotora del gen de la NCL de
humano consta de 1069 pb, el sitio de inicio de la transcripción se encuentra en la posición 1,070, mientras que el sitio de término se encuentra en la posición 10,499. Aunque el sitio de inicio de la transcripción de los genes eucariontes inicia frecuentemente en un nucleótido adenina (A), en el gen de la NCL humana inicia en citosina (C). La secuencia promotora consenso CCATT y la caja TATA, no se encuentran en las posiciones usuales -75 y -25 respectivamente, ya que así como con otros genes involucrados en el crecimiento celular o genes housekeeping, la NCL carece de la secuencia consenso tradicional, sin embargo se ha descrito a la secuencia GTTACTG en el sitio -49 (Figura 1A), como su sitio de regulación transcripcional, esta característica única además se encuentra conservada en la nucleolina de humano, ratón y otros roedores [5].
En la NCL de humano se encuentran diversos
elementos regulatorios putativos reconocidos por los factores CP1/CP2 y por CP1/CP2-RC (complemento reverso), los cuales flanquean el extremo 5’ y el primer intrón. También la secuencia consenso NNGCGGGGCN en las regiones ricas en GC en las posiciones -112, -155, -174, -213, -266, -308, -441, -782, 33, 91, 100, 181, 433 y 476 son elementos reguladores negativos para la transcripción del gen de la NCL y se encuentran en el extremo 5’ y en el primer intrón. En la posición -126 se encuentra la secuencia consenso TTGGCTNNNAGCCAA, la
cual es reconocida por el factor nuclear I (NF-1/CTPEl); por otro lado el elemento control UCE (CGGCC) es reconocido por la proteína de unión UBF1 y está localizado en las posiciones -112, -174, -257, -558, 35, 134, 434, 477, 496, 539 y 643 en la misma área. Hay siete secuencias en las posiciones -1006, -672, 224, 613, 807, 987, 1013 y 1066 de las cuales coinciden al menos 5 de cada 6 bases con el elemento de respuesta a glucocorticoides TGTTCT. La secuencia TGACTC en la posición -84 y 394 es idéntica al elemento reconocido por AP-1, GCN4 y CREB. La secuencia consenso CCCCAGGC es reconocida por AP-2 y factores de unión al ácido retinóico y están presentes en las posiciones -919, -767, -219, -194, -160 y 39. Aunque se han identificado todas estas secuencias consenso y putativas, hoy en día no se han realizado estudios experimentales para demostrar que esto ocurre en condiciones fisiológicas [5]. Estos datos abren la posibilidad de estudios futuros que demuestren los factores de transcripción que regulan positiva o negativamente la transcripción del gen de la NCL. Hoy en día existen únicamente dos reportes sobre la regulación transcripcional del gen de la NCL, uno de ellos demuestra que la actividad promotora del gen de la NCL es regulada por repetidos dinucleótidos polimórficos, los cuales forman estructuras Z-DNA, que podrían ser uno de los factores contribuyentes a la variación genética [6]. Y por otro lado se ha demostrado que el gen de la NCL es regulado por la proteína Myc, la cual se une a la secuencia consenso CACGTG (E-box), que se encuentra en los primeros intrones del gen de la NCL [7].
La longitud de los 14 exones de la NCL es variada, varían de 87 hasta 478 pb, mientras que los intrones varían de 104 a 1164 pb (Figura 1B). Cada splice donador inicia con GT y cada sitio aceptor termina con AG precedido por un tracto de polipirimidinas. Todos los intrones también contienen sitios aceptores potenciales lariat río arriba de las uniones de splice 3’ [5].
La información que se tiene hoy en día sobre
la NCL ha sido en gran parte por los estudios que se han hecho en el gen de la NCL de humano y de
en un modelo de neovascularización corneal y su repercusión como blanco terapéutico en procesos de angiogénesis aberrante. Palabras clave: angiogénesis, factor de crecimiento vascular endotelial (VEGF), AS1411.
Keywords: angiogenesis, vascular endothelial grown factor (VEGF), AS1411.

Alvarado-Hernández et al. Mens. Bioquim. 41 (2017): 37 - 48
© 2017 Mensaje Bioquímico. Todos los derechos reservados. ISSN-0188-137X Comité Editorial: Cárdenas Monroy, C.A.; González Andrade, M.; Lara Lemus, R.; Martínez González, J.J.; Molina Jijón, E.; Torres
Durán, P.V. Publicado por el Departamento de Bioquímica de la Facultad de Medicina; UNAM.
39
ratón, por ello se sabe que comparten muchas características, entre ellas que ambos contienen 14 exones y 13 intrones y que son interrumpidos por secuencias codificantes en posiciones idénticas. Sin embargo, los tamaños y las secuencias de los intrones tienen poca o ninguna similitud, excepto
por 300 pb en el primer intrón. La localización de las uniones de splice de las secuencias consenso RNP1 están conservadas en el extremo 3’ de cada exón (6,8,10 y 12)y en el extremo 5’ de los exones impares (7, 9, 11 y 13) [5].
Figura 1. Características del gen, RNAm y proteína de la NCL. A) Características del gen de la NCL, donde se marca el sitio +1 o sitio de inicio transcripcional (GTTAGCTG, secuencia de regulación transcripcional). B) Tamaño de los exones o regiones codificantes del gen de la NCL. C) RNAm de la NCL, sitio de inicio de la traducción y tamaño de las regiones UTR. D) Dominios de la proteína NCL, dominio N-terminal y sus regiones ácidas y básicas; dominio central, RBD 1,2,3,4; dominio C-terminal, señal de localización nuclear bipartita. Modificaciones postraduccionales reportadas, fosforilaciones y glicosilaciones. Modificado de Rong Cong et. al 2011 [12].

Alvarado-Hernández et al. Mens. Bioquim. 41 (2017): 37 - 48
© 2017 Mensaje Bioquímico. Todos los derechos reservados. ISSN-0188-137X Comité Editorial: Cárdenas Monroy, C.A.; González Andrade, M.; Lara Lemus, R.; Martínez González, J.J.; Molina Jijón, E.; Torres
Durán, P.V. Publicado por el Departamento de Bioquímica de la Facultad de Medicina; UNAM.
40
La señal de localización nuclear (Pro-Gly,Lys-Arg-Lys-Lys), la cual se encarga del transporte de la proteína al núcleo, está codificada en el exón 5. Las cuatro regiones ácidas del dominio terminal están codificadas entre los exones 2 a 4. La proteína NCL posee cuatro dominios de unión a RNA y están codificados por 2 exones consecutivos e independientes que además se ubican en posiciones idénticas; cuya organización genómica es muy conservada en humano, ratón y otros roedores. Finalmente, en el extremo 3’ el exón 13 contiene la información de dos tercios de la región rica en glicina y el exón 14 codifica para la porción restante [5]. Otra característica importante del gen de la NCL, es que los intrones 11 y 5 codifican para los snoRNAs (ARN pequeños nucleolares) U20 y U82 respectivamente [8,9].
El gen de la NCL codifica a un RNAm de
2406 nt con regiones 5’ y 3’ UTR (por su siglas en inglés, UnTranslated Regions), de 114 y 332 nt respectivamente, además de una cola poly-A de alrededor de 200 nt (Figura 1C) [10]. El codón de inicio ATG se encuentra en la posición 1069 y tiene una secuencia Kozak conservada. La región 3’ UTR incluye una señal de poliadenilación TATAAA en la posición 2424, la cual es ligeramente distinta en ratón. Finalmente, el codón de paro se encuentra en la posición 10214 [5]. Al igual que a nivel transcripcional, se sabe muy poco sobre la regulación a nivel post-transcripcional. Se ha reportado que la proteína HuR es un regulador positivo de la expresión de la NCL en algunos cánceres de mama y células no malignas, mientras que los microRNAs miR-194 y miR-206 son reclutados a la región 3’ UTR del RNAm de la NCL para regularla negativamente [11].
El marco de lectura abierta de la NCL de
humano, codifica para una proteína con una longitud de 707 aminoácidos (Figura 1D), cuya región codificante es 84% y 81% idéntica a la de hámster y ratón respectivamente, la cual corresponde a un peso molecular in sílico de 77 KDa y un peso molecular experimental de 100-110 KDa, que ha sido atribuido a las regiones ácidas del dominio N-terminal ya que son altamente fosforiladas [5]. Bioquímica de la nucleolina y sus modificaciones postraduccionales
La proteína NCL que se expresa en el citoplasma y superficie celular posee un punto
isoeléctrico (pI) de alrededor de 4.5, mientras que la que se encuentra en el núcleo varía de 4 a 6; las diferencias mostradas en los pI están mediados por las características particulares de sus dominios y modificaciones postraduccionales [13]. El dominio N-terminal posee repeticiones ácidas de glutamato (Glu) y aspartato (Asp) separadas por regiones básicas, este dominio se ha asociado a interacciones proteína-proteína, como a la histona H1 y a ribonucleoproteínas, además en este dominio se encuentra la señal de localización nuclear bipartita que le permite ser reclutada al núcleo [14]; el dominio central que posee cuatro dominios de unión a RNA o RBD (por sus siglas en inglés, RNA binding domain) los cuales están más conservados entre especie que entre ellos mismos, y que le permite a la NCL participar en el procesamiento de los pre-RNAr; y finalmente el dominio C-terminal, que posee regiones ricas en glicinas y argininas, denominado dominio GAR (por sus siglas en inglés Glycine Arginine Rich) que le permite unirse a proteínas y a RNAs (Fig. 1D) [15]. Aún no se cuenta con la cristalografía completa de la NCL, pero sí con la cristalografía de los dominios RBD1 y 2. El modelo teórico de las regiones RBD1 y 2 han permitido determinar que se encuentran organizadas espacialmente cercanas y que el cambio en la estructura terciaria de la NCL al interaccionar con el RNA, logra modificar los residuos básicos del dominio central, donde el RBD1 y 2 genera un cambio en su estructura completamente, provocando un torcimiento respecto a la estructura inicial (Figura 2). Estos datos nos ayudan a proponer que posiblemente este cambio conformacional le permite a la NCL interaccionar con otras moléculas, lo que le confiere la capacidad de cambiar su localización y/o función.
Las múltiples funciones de la NCL se deben a las diversas características que posee cada uno de sus dominios, los cuales además sufren modificaciones postraduccionales que permiten que la NCL pueda adoptar actividades y localizaciones distintas en la célula, dependiendo de la señal extracelular que reciba (Figura 1D). La localización de la NCL en el núcleo depende de su señal de localización nucleolar (NLS) y de otros factores con los que interacciona (Fig. 1D), ya que se ha propuesto que la localización nucleolar es también consecuencia de la afinidad de sus dominios, por factores nucleolares como el RNAr [17] u otros componentes nucleolares, además de las fosforilaciones que sufre, ya que cuando se desfosforila se mejora la eficiencia de su

Alvarado-Hernández et al. Mens. Bioquim. 41 (2017): 37 - 48
© 2017 Mensaje Bioquímico. Todos los derechos reservados. ISSN-0188-137X Comité Editorial: Cárdenas Monroy, C.A.; González Andrade, M.; Lara Lemus, R.; Martínez González, J.J.; Molina Jijón, E.; Torres
Durán, P.V. Publicado por el Departamento de Bioquímica de la Facultad de Medicina; UNAM.
41
translocación nuclear, mientras que cuando se fosforila su localización es citoplasmática [18].
Figura 2. Modelaje de la NCL. A) NCL (cian) estructura teórica modelada por Phyre2 (16). B) Regiones de unión a RNA (RBD1 y 2) (amarillo) y RNA (azul/magenta) (2KKR). C) Interacción de la NCL con el RNA y el cambio conformacional de RBD1 y 2. D) E) F) Secuencia de movimiento de las regiones centrales, RBD 1 y 2 en la estructura inicial de la NCL, el cambio en la conformación genera un torcimiento casi total del RBD2 y desfase de un asa (flecha).
Aunque no existen estudios que describan a detalle todas las posibles modificaciones postraduccionales de la NCL, varios reportes demuestran que la NCL puede estar fuertemente fosforilada [19]. La fosforilación se lleva a cabo en residuos de serina (Ser) y treonina (Thr) por diferentes cinasas [20]. Los residuos de Ser predominantemente en regiones ácidas del dominio N-terminal (Figura. 1D), son fosforilados por la cinasa de caseína-II (CK2), mientras que la cinasa dependiente de ciclina 1 (Cdk1) lo hace en dominios básicos con repeticiones TPXKK; ambas fosforilaciones están asociadas al proceso de división celular. Se ha propuesto que la fosforilación modula la interacción de la NCL con ácidos nucleicos y con otras proteínas durante el ciclo celular, además durante un daño genotóxico la unión de la NCL con el RNA incrementa la actividad de la proteína cinasa activada por mitógeno-38 (p38) [21], la cual está asociada a procesos de apoptosis. En células malignas, la NCL fosforilada (NCL-p) probablemente por la cinasa de fosfoinositol 3-fosfato (PI3K) y/o PKC, puede competir por la unión al promotor del gen REST (factor silenciador restrictivo), cuya interacción parece ser un mecanismo clave para la activación de genes por PKC y también para la regulación de la proliferación celular y apoptosis [22]. Se ha descrito que en células troncales
embrionarias, la unión de la NCL-p con factores de transcripción (Oct4) tiene efectos en la diferenciación y desarrollo espontáneo de mamíferos [23]. Existen reportes de que la NCL podría interactuar con la tirosina fosfatasa de la regeneración de hígado-3 (PRL-3), y que la actividad de PRL-3 es necesaria para la supresión de la NCL-p en el citoplasma y su acumulación en el nucléolo [24] lo que sugiere que la PRL-3 podría defosforilar a la NCL-p en el citoplasma, regulando así su distribución nucleolar.
Aunque la NCL se expresa mayoritariamente
en el núcleo y el citoplasma, se ha demostrado que en procesos de proliferación se expresa en la superficie celular de distintos tipos celulares [13,25,26]. Se ha demostrado que la expresión y translocación de la NCL en la superficie celular no es convencional, ya que carece de un péptido señal que la trasloque o una secuencia transmembranal que le permita anclarse a la superficie celular y aunque no se ha definido como se lleva a cabo su traslocación a la superficie, se sabe que es independiente del tráfico vesicular de retículo endoplásmico y complejo de Golgi [13].
Se ha demostrado que una pequeña fracción de
la NCL que se encuentra en la superficie de diferentes tipos celulares, es blanco de sitios

Alvarado-Hernández et al. Mens. Bioquim. 41 (2017): 37 - 48
© 2017 Mensaje Bioquímico. Todos los derechos reservados. ISSN-0188-137X Comité Editorial: Cárdenas Monroy, C.A.; González Andrade, M.; Lara Lemus, R.; Martínez González, J.J.; Molina Jijón, E.; Torres
Durán, P.V. Publicado por el Departamento de Bioquímica de la Facultad de Medicina; UNAM.
42
potenciales de glicosilación [27]; ya que la inhibición de la N-glicosilación mediante el tratamiento de células con tunicamicina impide la expresión de NCL en la superficie celular [28], mostrando que esta modificación postraduccional es absolutamente necesaria para su localización en la superficie. Los dos sitios posibles de N-glicosilación, N317 y N492, se encuentran localizados en los RBD 1 y 3 respectivamente (Figura 1D) [29]. Se ha determinado que la N-glicosilación podría presentar carbohidratos como N-acetil glucosamina (GlcNAc), mientras que para la O-glicosilación podrían ser la Galactosa (Gal) y el ácido N-acetil neuramínico (NeuAc α2-6).
La expresión de la NCL en la superficie
celular determina su función como receptor de diversas moléculas, el efecto que desencadena al expresarse depende de sus ligandos extracelulares, algunos implicados en la proliferación,
diferenciación, adhesión, mitogénesis y angiogénesis como en tumores [30,31] en donde la fosforilación y la glicosilación de la NCL son necesarias para estas interacciones. Por otro lado se ha reportado que el factor de crecimiento vascular endotelial (VEGF, por sus siglas en inglés Vascular Endothelial Growth Factor), promueve la expresión de la NCL en la superficie celular [32] y que la activación de la cascada de señalización vía PI3K es la que promueve esta expresión (Figura 3). Varios procesos que determinan su expresión en la superficie celular no están completamente dilucidados, aunque existen experimentos que proponen que la fosforilación por CK2 o por PKC-ξ ocasiona que se forme un complejo con la proteína de choque térmico Hsc70, cuya interacción le permite unirse con la miosina no muscular de cadena pesada (MyH9), que se encuentra asociada a proteínas del citoesqueleto, lo que le permitiría traslocarse a la superficie (Figura 3).
Figura 3. Propuesta de la translocación de la NCL a la superficie celular. La activación de vías de señalización por factores de crecimiento (VGEF) y su receptor (VGEF-R) activan la vía de PI3K u otras cinasas como PKC-ξ, CdK-1, CK2 que fosforilan a la NCL, permitiéndole asociarse a moléculas como Hsc70 y MyH9, esta última asociada a proteínas del citoesqueleto lo que le permitiría expresarse en la superficie celular.
La expresión de la NCL en la superficie celular tiene consecuencias más allá de las propias funciones de la NCL, ya que se ha demostrado en linfomas de células de linfocitos B, que pueden mostrar resistencia a la apoptosis por la interacción de la NCL (en las regiones RBD-4 y GAR) con el receptor de muerte Fas o CD95 [33] y que aún después de su interacción con su ligando FasL (CD95L) logra bloquear la
apoptosis. Otro tipo de interacción se demostró en células de carcinoma hepatocelular, donde la presencia de la NCL con el receptor de quimiocina 6 (CCR6) y su ligando CCL20 en la superficie, incrementa la señalización de las vías pAKT y pERK, ocasionando la expresión de metaloproteasas (MMP2 y 9) y moléculas de adhesión (ICAM-1), lo que le confiere capacidad de adhesión e invasión a la célula, proponiendo la presencia de la NCL y CCR6 en la superficie

Alvarado-Hernández et al. Mens. Bioquim. 41 (2017): 37 - 48
© 2017 Mensaje Bioquímico. Todos los derechos reservados. ISSN-0188-137X Comité Editorial: Cárdenas Monroy, C.A.; González Andrade, M.; Lara Lemus, R.; Martínez González, J.J.; Molina Jijón, E.; Torres
Durán, P.V. Publicado por el Departamento de Bioquímica de la Facultad de Medicina; UNAM.
43
como un posible factor de pronóstico en el carcinoma hepatocelular [34]. También se ha determinado en el carcinoma papilar de tiroides, que la presencia de la NCL y el receptor de quimiocina 4 (CXCR4) incrementa el crecimiento, migración e invasividad, cuya asociación está restringida sólo a la interacción del dominio 212 del C-terminal de la NCL [35]. De la misma forma se ha descrito que la NCL interacciona con moléculas de adhesión como la P-selectina o CD62L, permitiendo formar un complejo con PI3K y p38, incrementando la adhesión e invasión en células de carcinoma de colon, por lo que no solo participa de manera directa en el reconocimiento de ligandos, si no afectando complejos de vías de señalización [36]. Localización celular y funciones de la nucleolina
Como ya se ha mencionado, la NCL es una de las proteínas más abundantes del nucléolo y representa alrededor del 10% del contenido de proteínas dentro de este compartimento (Figura 4) [14].
Figura 4. Inmunofluorescencia de la NCL en cultivo celular Hela. NCL en verde, cuyo marcaje se observa en los nucleolos. Núcleos marcados con DAPI (azul).
Se localiza principalmente en las regiones fibrilar densa y granular del nucléolo [37], sin embargo durante la interfase y la mitosis, la NCL se identifica en la región periférica cerca del cinetocoro de los cromosomas, mientras que en el período de prometafase a anafase se relaciona con los polos del huso [14]. La localización de la NCL dentro del nucléolo está influenciada por el
dominio central y extremo carboxilo de la proteína o dominio GAR, ya que estos dominios le permiten unirse a componentes como el pre-RNAr y RNAr (38). Además se han identificado varias proteínas asociadas con la NCL en extractos nucleares, por ejemplo, B23, fibrilarina (Figura 5) y topoisomerasa 1 entre otras proteínas de tipo no ribosomal y sobre todo de tipo ribosomal, por ello su participación en eventos esenciales de la biogénesis de los ribosomas [13,14,39,40,41]. En el núcleo celular, la NCL puede tener actividad de helicasa, también interactúa con varios factores de la trascripción y componentes nucleares como la polimerasa tipo I y tipo II, cumpliendo una función de regulación de la replicación del DNA y del crecimiento celular [14] [42]. Por otro lado se ha relacionado a la NCL con una inhibición de la replicación del DNA inducida por choque de calor, lo que parece estar mediado por su asociación con el factor de replicación A [43].
Figura 5. Inmunofluorescencia de la NCL en células de cultivo celular epitelial. Se observa el marcaje de la NCL (verde) en el nucléolo celular, colocalizando (flecha) con la proteína nucleolar fibrilarina (rojo). Núcleo marcado con DAPI (azul).
Aunque carece de un dominio transmembranal o una secuencia señal, la NCL se ha encontrado en la superficie de varios tipos de células [13,26], sin embargo aún no se conoce bien como se transporta a la superficie celular [30], aunque se ha reportado una asociación estrecha entre la NCL de membrana y la actina del citoesqueleto [13]. Sobre la superficie celular sirve como una proteína de unión para varios ligandos, desde factores de crecimiento hasta microorganismos y partículas virales [30,31,46].
Se considera que la NCL de citoplasma y de superficie celular es proteína recién sintetizada en la célula [37]. En el citoplasma la NCL transporta proteínas hacia el núcleo y regula negativa o positivamente la traducción de diferentes RNA mensajeros como el de la proteína pro-oncogénica de linfoma de células B (Bcl-2) , la proteína precursora amiloide (APP), interlucina 2 (IL)-2, Akt1, ciclina I entre otras [42]; así como la

Alvarado-Hernández et al. Mens. Bioquim. 41 (2017): 37 - 48
© 2017 Mensaje Bioquímico. Todos los derechos reservados. ISSN-0188-137X Comité Editorial: Cárdenas Monroy, C.A.; González Andrade, M.; Lara Lemus, R.; Martínez González, J.J.; Molina Jijón, E.; Torres
Durán, P.V. Publicado por el Departamento de Bioquímica de la Facultad de Medicina; UNAM.
44
traducción del RNA de algunos virus como el del rinovirus, poliovirus y calicivirus felino [44,45] (Figura 6).
Figura 6. Inmunofluorescencia de la NCL en retina de murino. Reconstrucción tridimensional de cortes en XYZ realizados con microscopio confocal. Marcaje de la NCL (verde) en la monocpa nuclear interna de la retina de murino; cuyo marcaje se observa dentro del núcleo, aunque en algunas células se observa el marcaje mayoritariamente en el citoplasma. Núcleo marcado con DAPI (azul).
Existen reportes que relacionan ligandos de la NCL de superficie con la diferenciación y proliferación celular como en el desarrollo de
tumores y en la angiogénesis. Entre estas moléculas se encuentran el factor de crecimiento de hepatocitos, el péptido regulador afín a la heparina, midkina, el receptor del factor de crecimiento epitelial ErbB y endostatina [47,48]. Por otro lado existen otras moléculas de unión a NCL de superficie involucradas en otro procesos como la adhesión celular, el tráfico de leucocitos y la inflamación, como la laminina-1, la glicoproteína catiónica conocida como factor J y las selectinas L y P [47].
Se ha identificado la presencia de la NCL en la superficie de células tumorales así como en células endoteliales de vasos sanguíneos durante el desarrollo de neoplasias [31], por ello se ha reportado a la NCL como un marcador específico de células endoteliales en estado de angiogénesis (Figura 7) [30,49]. Esta proteína en la superficie es importante para la migración y formación de túbulos de células del endotelio y su incremento en la membrana se atribuye a un transporte desde el núcleo.
Figura 7. Presencia de NCL en la superficie celular durante la angiogénesis. Microfotografía de tejido corneal donde se observa una colocalización de nucleolina e isolectina B4 en la superficie celular de células del estroma de una córnea humana con desarrollo de vasos sanguíneos. Marcaje de NCL en células del estroma (a). Marcaje con isolectina B4 para identificar la superficie celular (b). Sobreposición del marcaje de NCL e isolectina B4 para identificar la colocalización (c). Identificación de los núcleos de las células que conforman la estructura vascular para evidenciar el marcaje de NCL e isolectina B4 en el espacio extranuclear (d).

Alvarado-Hernández et al. Mens. Bioquim. 41 (2017): 37 - 48
© 2017 Mensaje Bioquímico. Todos los derechos reservados. ISSN-0188-137X Comité Editorial: Cárdenas Monroy, C.A.; González Andrade, M.; Lara Lemus, R.; Martínez González, J.J.; Molina Jijón, E.; Torres
Durán, P.V. Publicado por el Departamento de Bioquímica de la Facultad de Medicina; UNAM.
45
El VEGF puede estimular la translocación de
la NCL de manera sinérgica cuando las células endoteliales se adhieren a la matriz extracelular, también se ha relacionado con eventos de señalización para la entrada de calcio y en procesos de adhesión celular y migración al promover la expresión de moléculas como TNF-α e ILI-β [42]. La nucleolina de membrana como blanco terapéutico
La búsqueda de la NCL de superficie celular mediante sus ligandos específicos sintéticos, como el pseudopéptido HB-19 han hecho de esta molécula un blanco terapéutico para la inhibición de neovasos expresados en tumores. El pseudopéptido HB-19 reconoce de manera específica al carboxilo terminal de la NCL de superficie formando un complejo estable, el cual es internalizado sin afectar la síntesis de la NCL que se encuentra en el núcleo. Se demostró en un modelo de cáncer de mama que el pseudopéptido HB-19 no sólo es capaz de identificar neovasos aberrantes sino que además genera una reducción significativa del tumor debido a la disminución en el aporte sanguíneo, concluyendo que la inhibición de esta molécula conlleva a la
inhibición de la proliferación celular de las células que conforman a los neovasos aberrantes, de la misma forma se demostró que la NCL de superficie es responsable de la citocinesis de las células que conforman los neovasos, así que la inhibición funcional de esta molécula se ha convertido en blanco para la terapia en tumores sólidos [31]. Además del pseudopéptido HB-19, también se ha descrito un aptámero específico para la región extracelular de la NCL. Este aptámero conocido como AS1411 induce apoptosis selectiva de diversas líneas celulares derivadas de tumores sólidos. Aunque no se conocen del todo los mecanismos por los cuáles estos aptámeros ejercen su función antiproliferativa, se han propuesto como reguladores positivos de la macropinocitosis que produce un desbalance en el tráfico intracelular que provoca la muerte de las células con una tasa de proliferación elevada [50]; asimismo se ha demostrado que otro mecanismo de inducción de apoptosis es mediante el arresto celular en fase S inhibiendo la replicación celular e inhibiendo las helicasas endógenas de las células neoplásicas [51].
Figura 8. Translocación de la NCL del núcleo a la región extranuclear en células del epitelio anterior de la córnea luego de someterse a un daño tisular por colocación de puntos de sutura. (a) Marcaje anti-nucleolina localizado en la región del núcleo de células de la capa basal del epitelio anterior de una córnea sana (C. control). (b) Microfotografía de inmunofluorescencia de corte histológico de córnea obtenido una semana después de la colocación de puntos de sutura, donde se observa un marcaje en la región extranuclear de células de varias capas del epitelio anterior de la córnea bajo probable estado de proliferación (C. sut.). En azul se identifican los núcleos celulares marcados con DAPI.
Alvarado-Hernández et al. Mens. Bioquim. 41 (2017): 37 - 48
© 2017 Mensaje Bioquímico. Todos los derechos reservados. ISSN-0188-137X Comité Editorial: Cárdenas Monroy, C.A.; González Andrade, M.; Lara Lemus, R.; Martínez González, J.J.; Molina Jijón, E.; Torres
Durán, P.V. Publicado por el Departamento de Bioquímica de la Facultad de Medicina; UNAM.
46
De manera interesante, el uso de estos aptámeros ya se encuentra en fases clínicas para el tratamiento de cáncer renal en etapas avanzadas, teniendo resultados positivos en cuanto a la reducción del tamaño en el tumor renal, inhibiendo la proliferación de las células del tumor, así como las células endoteliales, apoyando la función de esta proteína como reguladora en la angiogénesis [52]. Recientemente se ha descrito que la NCL de superficie, además se encuentra en procesos angiogénicos en el sistema visual. En un estudio con roedores donde se indujo neoangiogénesis coroidea mediante láser se demostró que la inyección intravítrea de aptámeros AS1411 redujo significativamente el proceso de formación de vasos coroideos aberrantes, así como la disminución del infiltrado leucocitario en la retina, además se demostró que la nucleolina se localizaba en los sitios de neovascularización; estos hallazgos sugieren que la NCL tiene una función importante en el proceso de degeneración macular relacionada con la edad [53].
En nuestro laboratorio hemos encontrado que
la NCL de superficie se expresa específicamente en neovasos corneales aberrantes y que la inhibición de la angiogénesis corneal mediante el uso de un anticuerpo humanizado contra el VEGF (bevacizumab) disminuye concomitantemente la
presencia de la NCL [54]. Para determinar que esta proteína se expresa de manera preferencial en los neovasos aberrates, es decir aquellos que crecieron anormalmente en una córnea con daño, se realizaron estudios de inmunodetección de NCL en córneas humanas con neoangiogénesis e interesantemente se demostró su presencia de manera similar que en nuestro modelo murino (Figura 8). De igual manera, se realizaron estudios comparando la presencia de la NCL en los vasos aberrantes de córneas murinas y de los vasos retinianos murinos maduros sin alteraciones, encontrándose una presencia exclusiva de la NCL en los neovasos de córnea, mientras que en los vasos maduros normales de retina murina no se identificó su presencia. Estos datos sugieren que la NCL de superficie se presenta de manera preferencial en estructuras vasculares aberrantes.
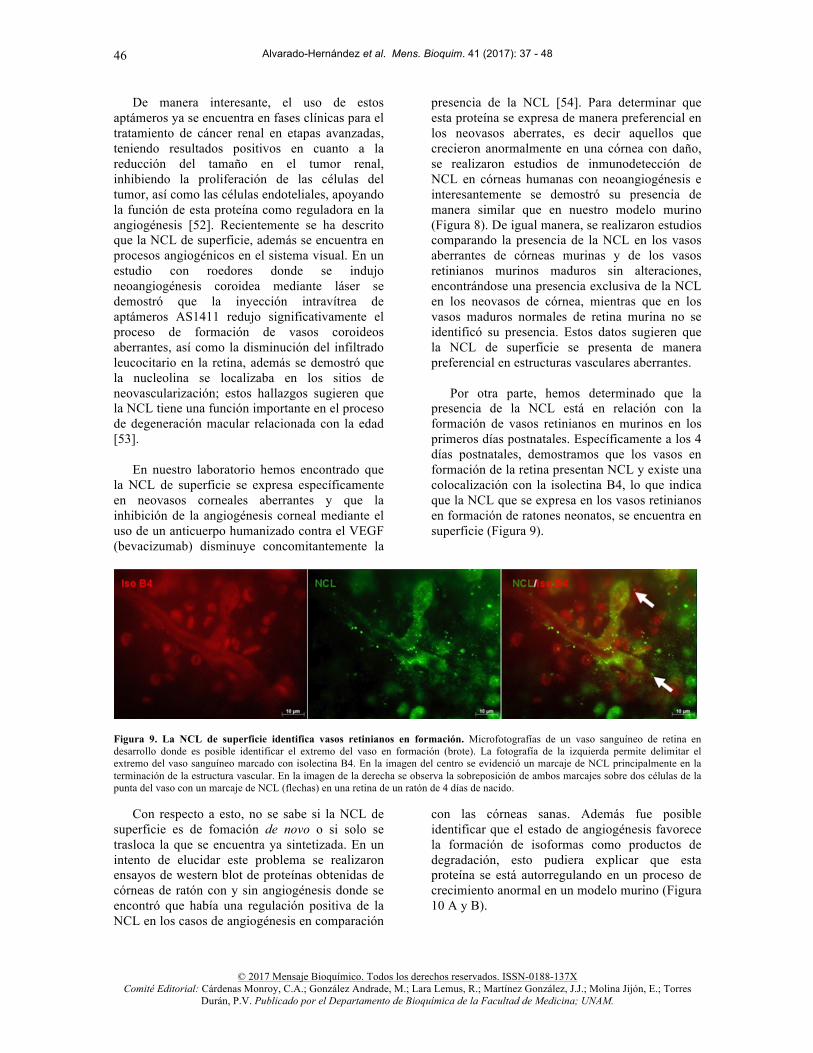
Por otra parte, hemos determinado que la
presencia de la NCL está en relación con la formación de vasos retinianos en murinos en los primeros días postnatales. Específicamente a los 4 días postnatales, demostramos que los vasos en formación de la retina presentan NCL y existe una colocalización con la isolectina B4, lo que indica que la NCL que se expresa en los vasos retinianos en formación de ratones neonatos, se encuentra en superficie (Figura 9).
Figura 9. La NCL de superficie identifica vasos retinianos en formación. Microfotografías de un vaso sanguíneo de retina en desarrollo donde es posible identificar el extremo del vaso en formación (brote). La fotografía de la izquierda permite delimitar el extremo del vaso sanguíneo marcado con isolectina B4. En la imagen del centro se evidenció un marcaje de NCL principalmente en la terminación de la estructura vascular. En la imagen de la derecha se observa la sobreposición de ambos marcajes sobre dos células de la punta del vaso con un marcaje de NCL (flechas) en una retina de un ratón de 4 días de nacido.
Con respecto a esto, no se sabe si la NCL de superficie es de fomación de novo o si solo se trasloca la que se encuentra ya sintetizada. En un intento de elucidar este problema se realizaron ensayos de western blot de proteínas obtenidas de córneas de ratón con y sin angiogénesis donde se encontró que había una regulación positiva de la NCL en los casos de angiogénesis en comparación
con las córneas sanas. Además fue posible identificar que el estado de angiogénesis favorece la formación de isoformas como productos de degradación, esto pudiera explicar que esta proteína se está autorregulando en un proceso de crecimiento anormal en un modelo murino (Figura 10 A y B).

Alvarado-Hernández et al. Mens. Bioquim. 41 (2017): 37 - 48
© 2017 Mensaje Bioquímico. Todos los derechos reservados. ISSN-0188-137X Comité Editorial: Cárdenas Monroy, C.A.; González Andrade, M.; Lara Lemus, R.; Martínez González, J.J.; Molina Jijón, E.; Torres
Durán, P.V. Publicado por el Departamento de Bioquímica de la Facultad de Medicina; UNAM.
47
Figura 10. Incremento de la expresión de NCL en córneas con inducción de angiogénesis y formación de posibles isoformas de la nucleolina durante el proceso de angiogénesis. A) Western Blot realizado en córneas con angiogénesis y corneas sin angiogénesis (control). En el carril de la izquierda se aprecia la expresión incrementada de NCL en el tejido corneal con angiogénesis inducida por colocación de sutura, los carriles de la derecha muestran la expresión de NCL en córneas control (a). En la imagen de la izquierda se observa un marcaje anti-NCL correspondiente a la banda de 100KDa siendo más marcado en el carril correspondiente al tejido corneal con angiogénesis (sut) en comparación con los siguientes tres carriles que corresponden con tejido control (sin sut). B) En la imagen de la izquierda se observan además de la banda de 100KDa, bandas de menor peso molecular que podrían relacionarse con fragmentos de NCL. La imagen de la derecha corresponde a la membrana tratada sin anticuerpo primario (control negativo). M: marcadores de peso molecular; Sut: córneas con angiogénesis; Sin Sut: córneas sin angiogénesis.
Con todos estos hallazgos que demuestran una selectiva expresión de la NCL en la superficie de las células endoteliales de neovasos aberrantes, como sucede en diferentes neoplasias, es posible proponer a esta proteína ubicua y pleiotrópica como un blanco para la detección y terapia de enfermedades que involucran el crecimiento aberrante de neovasos en el sistema visual.
Agradecimientos Este trabajo se realizó bajo el apoyo de los siguientes fondos: CONACYT-Ciencia Básica 167438; CONACYT-Problemas Nacionales 2015-311; PAPIIT-DGAPA-UNAM: IN215617; CVU No. 545374/Posgrado en Ciencias Biológicas, UNAM y CVU No. 332602/Programa de Doctorado en Ciencias Biomédicas y de la Salud, UNAM/Coordinación de la Investigación Científica/Facultda de Medicina. Los autores agradecen la participación de la Dra. Beatriz Buentello Volante y de la Biol. Exp. Mariana Angélica García Mejía.
Referencias 1. Orrick, L. R., Olson, M. O., and Busch, H. (1973) Proc
Natl Acad Sci U S A 70, 1316-1320. 2. Tong, C. G., Reichler, S., Blumenthal, S., Balk, J., Hsieh,
H. L., and Roux, S. J. (1997) Plant Physiol 114, 643-652. 3. de Carcer, G., Cerdido, A., and Medina, F. J. (1997)
Planta 201, 487-495 4. Srivastava, M., Fleming, P. J., Pollard, H. B., and Burns,
A. L. (1989) FEBS Lett 250, 99-105. 5. Srivastava, M., McBride, O. W., Fleming, P. J., Pollard,
H. B., and Burns, A. L. (1990) J Biol Chem 265, 14922-14931.
6. Rothenburg, S., Koch-Nolte, F., Rich, A., and Haag, F. (2001) Proc Natl Acad Sci U S A 98, 8985-8990.
7. Greasley, P. J., Bonnard, C., and Amati, B. (2000) Nucleic Acids Res 28, 446-453.
8. Rebane, A., and Metspalu, A. (1999) Biochim Biophys Acta 1446, 426-430.
9. Nicoloso, M., Caizergues-Ferrer, M., Michot, B., Azum, M. C., and Bachellerie, J. P. (1994) Mol Cell Biol 14, 5766-5776.
10. Bourbon, H. M., Lapeyre, B., and Amalric, F. (1988) J Mol Biol 200, 627-638.
11. Bose, S., Tholanikunnel, T. E., Reuben, A., Tholanikunnel, B. G., and Spicer, E. K. (2016) Mol Cell Biochem 417, 141-153.
12. Rong Cong, S. D., and Philippe Bouvet. (2011) The Multiple Properties and Functions of Nucleolin, Jackson,, Mississippi.
13. Hovanessian, A. G., Puvion-Dutilleul, F., Nisole, S., Svab, J., Perret, E., Deng, J. S., and Krust, B. (2000) Exp Cell Res 261, 312-328.
14. Tajrishi, M. M., Tuteja, R., and Tuteja, N. (2011) Commun Integr Biol 4, 267-275.
15. Ginisty, H., Sicard, H., Roger, B., and Bouvet, P. (1999) Journal of Cell Science 112, 761-772.
16. Kelley, L. A., Mezulis, S., Yates, C. M., Wass, M. N., and Sternberg, M. J. E. (2015) Nat. Protocols 10, 845-858.
17. Olson MO, T. B. (1983) Biochemistry 21, 3187-3193. 18. Schwab MS1, D. C. (1997) Eur J Cell Biol 73, 287-297. 19. Lischwe, M. A., Richards, R. L., Busch, R. K., and Busch,
H. (1981) Experimental Cell Research 136, 101-109. 20. Caizergues-Ferrer, M., Belenguer, P., Lapeyre, B.,
Amalric, F., Wallace, M. O., and Olson, M. O. J. (1987) Biochemistry 26, 7876-7883.
21. Yang, C., Maiguel, D. A., and Carrier, F. (2002) Nucleic Acids Research 30, 2251-2260.

Alvarado-Hernández et al. Mens. Bioquim. 41 (2017): 37 - 48
© 2017 Mensaje Bioquímico. Todos los derechos reservados. ISSN-0188-137X Comité Editorial: Cárdenas Monroy, C.A.; González Andrade, M.; Lara Lemus, R.; Martínez González, J.J.; Molina Jijón, E.; Torres
Durán, P.V. Publicado por el Departamento de Bioquímica de la Facultad de Medicina; UNAM.
48
22. Tediose, T., Kolev, M., Sivasankar, B., Brennan, P., Morgan, B. P., and Donev, R. (2010) Nucleic Acids Research 38, 2799-2812.
23. Johansson, H., Svensson, F., Runnberg, R., Simonsson, T., and Simonsson, S. (2010) PLOS ONE 5, e13678.
24. Semba, S., Mizuuchi, E., and Yokozaki, H. (2010) Cancer Science 101, 2254-2261.
25. Derenzini, M. (2000) Micron 31, 117-120. 26. Chen, X., Kube, D. M., Cooper, M. J., and Davis, P. B.
(2008) Molecular therapy : the journal of the American Society of Gene Therapy 16, 333-342.
27. Lapeyre, B., Bourbon, H., and Amalric, F. (1987) Proceedings of the National Academy of Sciences 84, 1472-1476.
28. Losfeld, M.-E., Leroy, A., Coddeville, B., Carpentier, M., Mazurier, J., and Legrand, D. (2011) FEBS Journal 278, 2552-2564.
29. Carpentier, M., Morelle, W., Coddeville, B., Pons, A., Masson, M., Mazurier, J., and Legrand, D. (2005) Biochemistry 44, 5804-5815.
30. Fogal, V., Sugahara, K. N., Ruoslahti, E., and Christian, S. (2009) Angiogenesis 12, 91-100.
31. Destouches, D., El Khoury, D., Hamma-Kourbali, Y., Krust, B., Albanese, P., Katsoris, P., Guichard, G., Briand, J. P., Courty, J., and Hovanessian, A. G. (2008) PLoS One 3, e2518.
32. Huang, Y., Shi, H., Zhou, H., Song, X., Yuan, S., and Luo, Y. (2006) Blood 107.
33. Wise, J. F., Berkova, Z., Mathur, R., Zhu, H., Braun, F. K., Tao, R.-H., Sabichi, A. L., Ao, X., Maeng, H., and Samaniego, F. (2013) Blood 121, 4729-4739.
34. Qiu, W., Wang, G., Sun, X., Ye, J., Wei, F., Shi, X., and Lv, G. (2015) Medical Oncology 32, 75.
35. Niu, H., Yang, X., Xu, Z., Du, T., and Wang, R. (2015) Tumor Biology 36, 1099-1104.
36. Reyes-Reyes, E. M., and Akiyama, S. K. Experimental Cell Research 314, 2212-2223.
37. Koutsioumpa, M., and Papadimitriou, E. (2014) Recent Pat Anticancer Drug Discov 9, 137-152.
38. Srivastava, M., and Pollard, H. B. (1999) Faseb J 13, 1911-1922.
39. Pinol-Roma, S. (1999) Mol Biol Cell 10, 77-90.
40. Ugrinova, I., Monier, K., Ivaldi, C., Thiry, M., Storck, S., Mongelard, F., and Bouvet, P. (2007) BMC Mol Biol 8, 66.
41. Wang, W., Luo, J., Xiang, F., Liu, X., Jiang, M., Liao, L., and Hu, J. (2014) PLoS One 9, e110101.
42. Abdelmohsen, K., and Gorospe, M. (2012) RNA Biol 9, 799-808.
43. Velichko, A. K., Petrova, N. V., Kantidze, O. L., and Razin, S. V. (2012) Molecular biology of the cell 23, 3450-3460.
44. Izumi, R. E., Valdez, B., Banerjee, R., Srivastava, M., and Dasgupta, A. (2001) Virus Res 76, 17-29.
45. Hernandez, B. A., Sandoval-Jaime, C., Sosnovtsev, S. V., Green, K. Y., and Gutierrez-Escolano, A. L. (2016) Virology 489, 51-62.
46. Krust, B., El Khoury, D., Nondier, I., Soundaramourty, C., and Hovanessian, A. G. (2011) BMC cancer 11, 333.
47. Birmpas, C., Briand, J. P., Courty, J., and Katsoris, P. (2012) BMC cell biology 13, 32.
48. Ding, Y., Song, N., Liu, C., He, T., Zhuo, W., He, X., Chen, Y., Song, X., Fu, Y., and Luo, Y. (2012) Arterioscler Thromb Vasc Biol 32, e126-134.
49. Christian, S., Pilch, J., Akerman, M. E., Porkka, K., Laakkonen, P., and Ruoslahti, E. (2003) The Journal of cell biology 163, 871-878.
50. Bates, P. J., Reyes-Reyes, E. M., Malik, M. T., Murphy, E. M., O'Toole, M. G., and Trent, J. O. (2016) Biochim Biophys Acta .
51. Xu, X., Hamhouyia, F., Thomas, S. D., Burke, T. J., Girvan, A. C., McGregor, W. G., Trent, J. O., Miller, D. M., and Bates, P. J. (2001) J Biol Chem 276, 43221-43230.
52. Rosenberg, J. E., Bambury, R. M., Van Allen, E. M., Drabkin, H. A., Lara, P. N., Jr., Harzstark, A. L., Wagle, N., Figlin, R. A., Smith, G. W., Garraway, L. A., Choueiri, T., Erlandsson, F., and Laber, D. A. (2014) Invest New Drugs 32, 178-187.
53. Leaderer, D., Cashman, S. M., and Kumar-Singh, R. (2015) Exp Eye Res 140, 171-178.
54. Quiroz-Mercado, J., Ramirez-Velazquez, N., Partido, G., Zenteno, E., Chavez, R., Agundis-Mata, C., Jimenez-Martinez, M. C., and Garfias, Y. (2016) Graefes Arch Clin Exp Ophthalmol 254, 1753-1763.
DR. YONATHAN OMAR GARFIAS BECERRA
Médico Cirujano egresado de la Facultad de Medicina de la UNAM. Doctorado en Ciencias Biomédicas. Profesor Titular B de Tiempo Completo Definitivo en la Coordinación de Inmunología del Departamento de Bioquímica, Unidad de Investigación Fundación Conde de Valenciana-UNAM, de la Facultad de Medicina. Tutor de diversos programas de posgrados. Publicación de más de 40 artículos nacionales e internacionales con más de 250 citas. Miembro del Sistema Nacional de Investigadores desde 2003, actualmente en el nivel 1. Formación de más de 30 recursos humanos desde licenciatura hasta postdoctorado. Estancia de investigación en la Universidad de Tufts, Boston MA, EUA e Investigador invitado en el Institut de la Vision-Institut National de la Santé et la Recherche Médicale, París, Francia.


















