
Mechanismy koncentrování CO C4 a CAM fotosyntézakfrserver.natur.cuni.cz/lide/lukasf/emr/6.pdf ·...
22
Mechanismy koncentrování CO 2 C4 a CAM fotosyntéza Mechanismy koncentrování CO 2 (carbon-concentrating mechanisms) Sun, Y., Casella, S., Fang, Y., Huang, F., Faulkner, M., Barrett, S. and Liu, L. (2016). Light modulates the biosynthesis and organization of cyanobacterial carbon fixation machinery through photosynthetic electron flow. Plant Physiol. 171: 530-541 . Engel, B.D., Schaffer, M., Kuhn Cuellar, L., Villa, E., Plitzko, J.M. and Baumeister, W. (2015). Native architecture of the Chlamydomonas chloroplast revealed by in situ cryo-electron tomography. eLife. 4: e04889 . Sinice: Karboxyzómy flagella chloroplast nucleus py pyrenoid Řasy: Pyrenoid Rostliny: C4 a CAM metabolismus HCO 2 - CO 2 C 4 acids CO 2 CB cycle PEPC Rubisco CA ME n. PEPCK Cíl: snížení oxygenázové akt. Rubisco (fotorespirace)
Transcript of Mechanismy koncentrování CO C4 a CAM fotosyntézakfrserver.natur.cuni.cz/lide/lukasf/emr/6.pdf ·...

Mechanismy koncentrování CO2
C4 a CAM fotosyntéza
Mechanismy koncentrování CO2(carbon-concentrating mechanisms)
Sun, Y., Casella, S., Fang, Y., Huang, F., Faulkner , M., Barrett, S. and Liu, L. (2016). Light modulat es the biosynthesis and organization of cyanobacteri al carbon fixation machinery through photosynthetic electron flow. Pla nt Physiol. 171: 530-541. Engel, B.D., Schaffer, M., Kuhn Cuellar, L., Villa, E., Plitzko, J.M. and Baumeister, W. (2015). Native architecture of the Chlamydomonas chloroplast revealed by in situ cryo-electron tomography. eLife. 4: e04889.
Sinice:Karboxyzómy
flagella
chloroplast
nucleus
py
pyrenoid
Řasy:Pyrenoid
Rostliny:C4 a CAM
metabolismus
HCO2- CO2
C4 acids
CO2
CB cycle
PEPC
Rubisco
CA
ME n. PEPCK
Cíl: snížení oxygenázové akt. Rubisco (fotorespirace)

Karboxyzóm sinic koncentruje Rubisco a CO2
Adapted from Price, G.D., Badger, M.R. and von Caemmerer, S. (2011). The prospect of using cyanobacterial bicarbonate transporters to improve leaf photosynthesis in C3 crop plants. Plant Physiol. 155: 20-26; Badger, M.R., and Price, G.D. (2003). CO2 concentrating mechanisms in cyanobacteria: molecular components, their diversity and evolution. J. Exp. Bot. 54: 609 –622. Toyeates
HCO3-
CO2 HCO3-
HCO3-
CO2CA
Rubisco
Carbonic anhydrase (CA) anhydráza kys. uhli čité:
CO2 + H2O HCO3- + H+
obalové proteiny: - tvo ří dvacetist ěn- umožňují vstup HCO 3
-
- brání úniku CO 2- váží Rubisco- vstup a výstup metabolit ů
- asi všechny sinice- koncentrování až 1000 x- nutné transportéry - nutná CA v karboxyzómu
chloroplast
nucleus
py
Engel, B.D., Schaffer, M., Kuhn Cuellar, L., Villa, E., Plitzko, J.M. and Baumeister, W. (2015). Native architecture of the Chlamydomonaschloroplast revealed by in situ cryo-electron tomography. eLife. 4: e04889.
Tylakoidy
škrob
Rubisco Rubisco
Pyrenoidy řas a růžkatců koncentrují Rubisco a CO2
- škrobovými zrny obklopený kompartment- aglomeráty Rubisco v proteinové matrix (EPYC1 + tubuly tylakoid ů)- tvorba často indukována malou [CO 2]
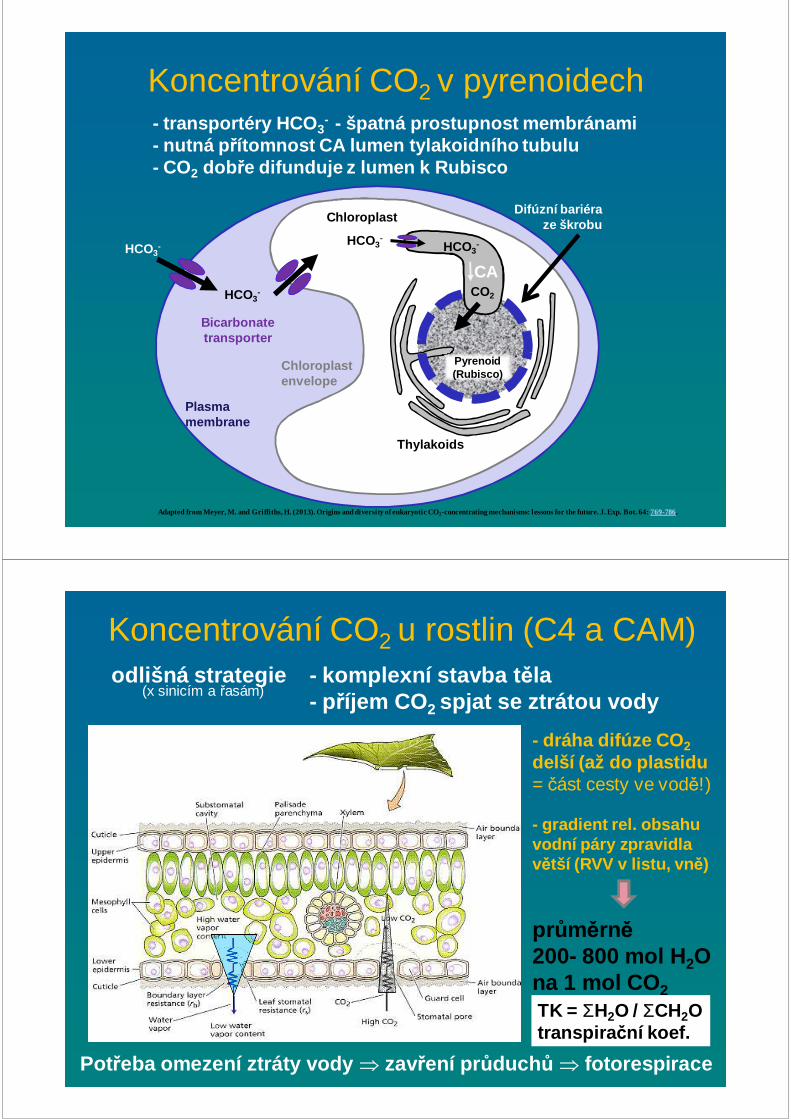
Koncentrování CO2 v pyrenoidech
Adapted from Meyer, M. and Griffiths, H. (2013). Origins and diversity of eukaryotic CO2-concentrating mechanisms: lessons for the future. J. Exp. Bot. 64: 769-786.
Plasma membrane
Chloroplast envelope
Chloroplast
Pyrenoid (Rubisco)
Difúzní bariéra ze škrobu
Thylakoids
HCO3-
HCO3-
HCO3-
Bicarbonate transporter
HCO3-
CO2
CA
- transportéry HCO 3- - špatná prostupnost membránami
- nutná p řítomnost CA lumen tylakoidního tubulu- CO2 dob ře difunduje z lumen k Rubisco
Koncentrování CO2 u rostlin (C4 a CAM)odlišná strategie - komplexní stavba t ěla
- příjem CO 2 spjat se ztrátou vody
- dráha difúze CO 2 delší (až do plastidu = část cesty ve vodě!)
- gradient rel. obsahu vodní páry zpravidlavětší (RVV v listu, vn ě)
průměrně 200- 800 mol H 2Ona 1 mol CO 2
(x sinicím a řasám)
Potřeba omezení ztráty vody ⇒ zavření průduch ů ⇒ fotorespirace
TK = ΣH2O / ΣCH2Otranspira ční koef.

vacuole
PEP
Koncentrování CO2 u rostlin (C4 a CAM)Cíl: omezit ztráty vody bez současné indukce fotorespirace
Koncentrování CO2 u rostlin - princip1) vazba CO 2 (i v nízké koncentraci - karboxylace C3 kys.)
PEPc (bez rizika oxygenázové aktivity): PEP + CO2 → OAA (fosfoenolpyruvátkarboxyláza) C3 + CO2 → C4 kyselina (OAA)
PEPc: Km = 0,7- 0,05 µM (CO2) Rubisco: Km = 15 µM (CO2)
2) místn ě či časově specifické uvoln ění CO2 (dekarboxylace C4 kys.)
ME (NADP či NAD dependentní) – jablečný enzym:(dekarboxylující malátdehydrogenáza)
nebo
PEPCK (fosfoenolpyruvátkarboxykináza):
3) fixace v Calvinov ě cyklu ( Rubisco )- omezená oxygenázová aktivita díky vysoké koncentraci CO2
( )
( )

2) B. pochev cévních sv. (BSC)
dekarboxylace C4 molekuly (Mal, OAA)
vlastní (sekundární) fixace CO 2(Rubisco v Calvinov ě cyklu)
→ PGA (C3)
C4 fotosyntéza
Dvojí fixace CO 2
1) Mezofylové bu ňky (MC)
primární fixace PEPc → OAA (C4)
Reprinted from Denton, A. K., Simon, R., and Weber, A.P.M. (2013). C4 photosynthesis: from evolutionary analyses to strategies for synthetic reconstruction of the trait. Curr. Opin. Plant Biol. 16: 315–321with permission from Elsevier.
C4 fotosyntézamístně specifické uvolnění CO2 z C4 kys. v BSC
(v buňkách pochev cévních svazků)
C3
C4
Vascular bundle
Vascular bundle
Vascular bundle
Vascular bundle
BSC
Mesophyll cells
Mesophyll cells
C3: Rubisco ve všech chloroplastechC4: Rubisco jen v chlp BSC
R R
R
RR R
R
R
RR
R
R

Předpoklady pro funkční C4 fotosyntézu
- buněčně specifická exprese genů- PEPc v mesofylu - Rubisco v BSC- častá absence PSII v BSC
PROČ?
- masivní symplastická spojení mezi zdroji a sinky pro cyklující C3 a C4 molekuly
- difusní bariéra pro CO2 na povrchu pochev (suberinová vrstva)
hranice pochvy cévního svazku
BSC
MC
Plasmodesmy
Evoluce C4 fotosyntézy- paraleln ě vznikla min. 62x (19 čeledí), 3 % druh ů- 6-22 mil. let - Poaceae, Cyperaceae, Amaranthaceae- u některých C3 rostlin (tabák) je C4 fotosyntéza ve sto ncích a v řapících
Sage et al. 2011, JXB
Vývojová centra dvoud ěložných C4
Cíl: omezit ztráty vody bez současné indukce fotorespirace
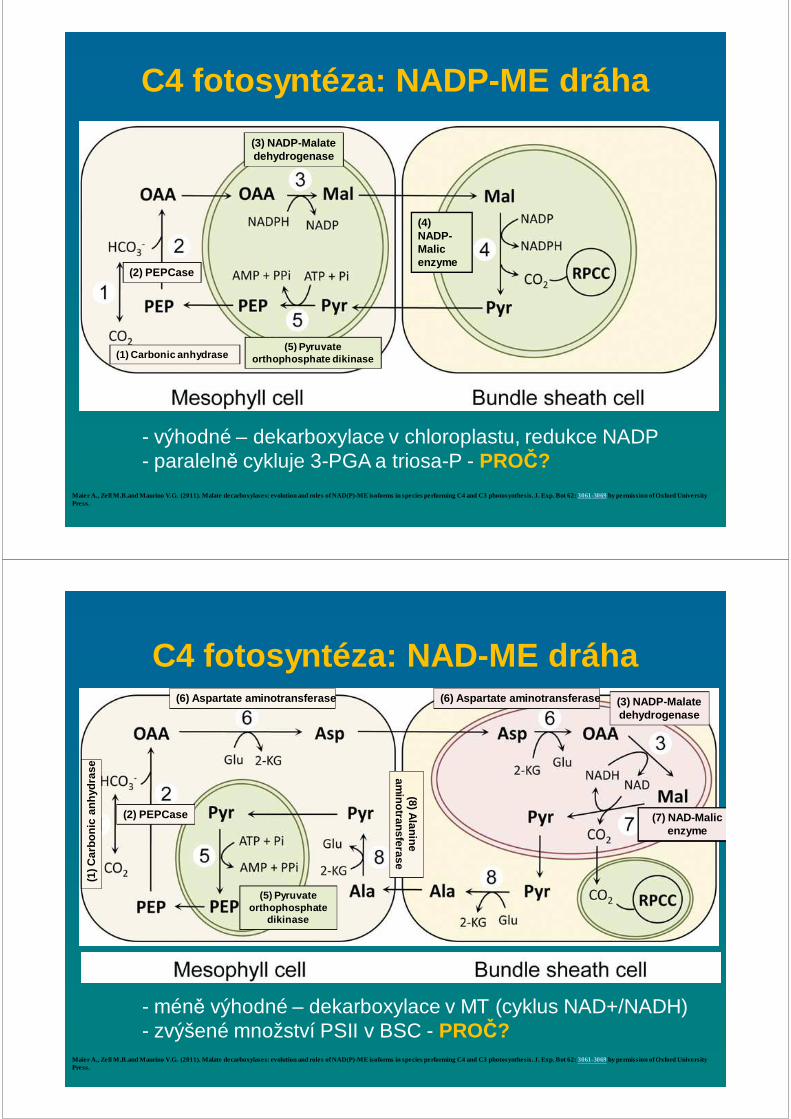
C4 fotosyntéza: NADP -ME dráha
Maier A., Zell M.B.and Maurino V.G. (2011). Malate decarboxylases: evolution and roles of NAD(P)-ME isoforms in species performing C4 and C3 photosynthesis. J. Exp. Bot 62: 3061-3069by permission of Oxford University Press.
(1) Carbonic anhydrase
(2) PEPCase
(3) NADP-Malate dehydrogenase
(4) NADP-Malic enzyme
(5) Pyruvateorthophosphate dikinase
- výhodné – dekarboxylace v chloroplastu, redukce NADP- paralelně cykluje 3-PGA a triosa-P - PROČ?
(1) C
arbo
nic
anhy
dras
e
(2) PEPCase
(5) Pyruvateorthophosphate
dikinase
(3) NADP-Malate dehydrogenase
(6) Aspartate aminotransferase (6) Aspartate aminotransferase
(8) Alanine
aminotransferase
(7) NAD-Malic enzyme
Maier A., Zell M.B.and Maurino V.G. (2011). Malate decarboxylases: evolution and roles of NAD(P)-ME isoforms in species performing C4 and C3 photosynthesis. J. Exp. Bot 62: 3061-3069by permission of Oxford University Press.
C4 fotosyntéza: NAD -ME dráha
- méně výhodné – dekarboxylace v MT (cyklus NAD+/NADH)- zvýšené množství PSII v BSC - PROČ?
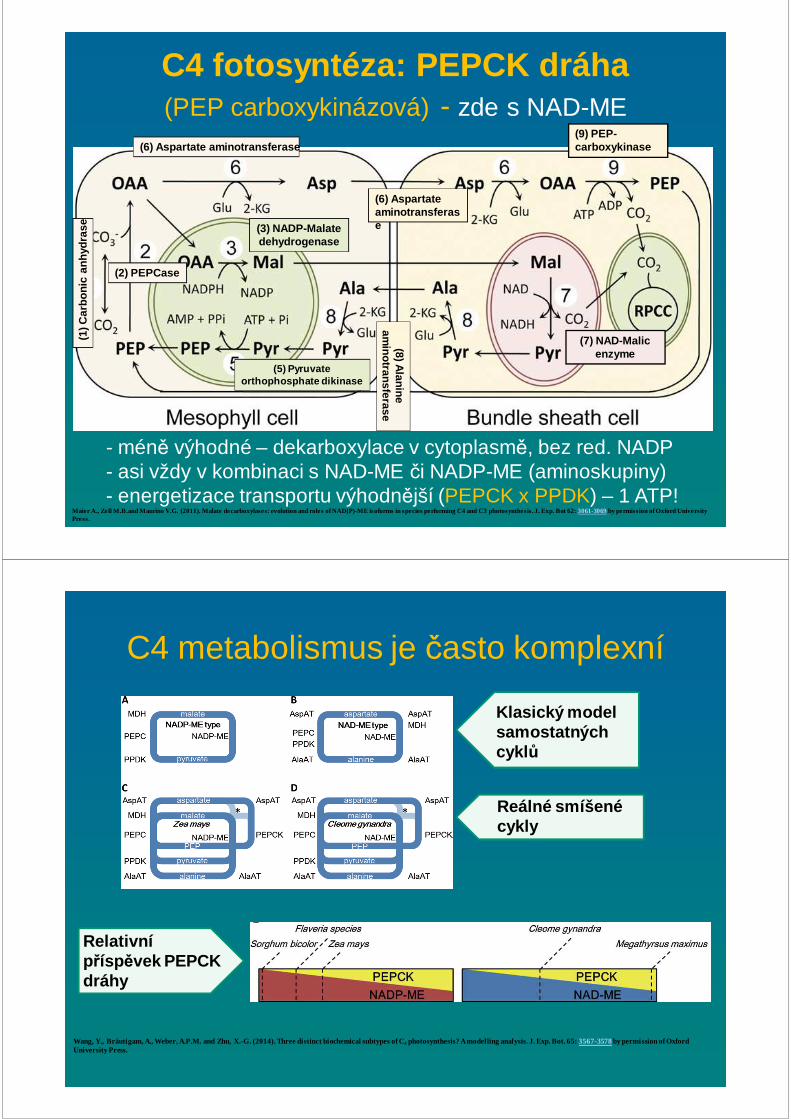
(1) C
arbo
nic
anhy
dras
e
(2) PEPCase
(3) NADP-Malate dehydrogenase
(5) Pyruvateorthophosphate dikinase
(6) Aspartate aminotransferase
(6) Aspartate aminotransferase
(9) PEP-carboxykinase
(7) NAD-Malic enzyme
(8) Alanine
aminotransferase
Maier A., Zell M.B.and Maurino V.G. (2011). Malate decarboxylases: evolution and roles of NAD(P)-ME isoforms in species performing C4 and C3 photosynthesis. J. Exp. Bot 62: 3061-3069by permission of Oxford University Press.
C4 fotosyntéza: PEPCK dráha(PEP carboxykinázová) - zde s NAD-ME
- méně výhodné – dekarboxylace v cytoplasmě, bez red. NADP- asi vždy v kombinaci s NAD-ME či NADP-ME (aminoskupiny)- energetizace transportu výhodnější (PEPCK x PPDK) – 1 ATP!
C4 metabolismus je často komplexní
Wang, Y., Bräutigam, A., Weber, A.P.M. and Zhu, X.-G. (2014). Three distinct biochemical subtypes of C4 photosynthesis? A modelling analysis. J. Exp. Bot. 65: 3567-3578by permission of Oxford University Press.
Klasický modelsamostatných cykl ů
Reálné smíšené cykly
Relativní příspěvek PEPCK dráhy

Fylogeneze typů C4 u lipnicovitých
(Rao and Dixon, FiPS 2016)
- všechny enzymy přítomné i u C3 – umožnilo nezávislou evoluci
Rod Flaveriaškála druhů od C3 po C4 metabolismus
Flaveria trinervia
Mallmann, J., Heckmann, D., Bräutigam, A., Lercher, M.J., Weber, A.P.M., Westhoff, P. and Gowik, U. (2014). The role of photorespiration during the evolution of C4 photosynthesis in the genus Flaveria. eLife. 3: e02478; Forest & Kim Starr .Whitney et al. PNAS 2011.
- výborný modelstudia evoluce C4 metabolismu
C2 metabolismus
Substituce Met-309-Ile ve velké podjednotce Rubisco - switch mezi pomalým s vyšší afinitou a rychlým s nižší afinitou

C2 fotosyntéza koncentruje CO2uvolněný fotorespirací v BSC
Sage R.F., Khoshravesh R., and Sage T.L. (2014). From proto-Kranzto C4 Kranz: building the bridge to C4 photosynthesis. J. Exp. Bot. 65: 3341-3356by permission of Oxford University Press.
C3 C4C2
- C2 pumpa (glycinová pumpa) – dekarboxylace jen v BSC
Opakovaná evoluce C4 fotosyntézy
Edwards E.J. (2014). The inevitability of C4 photosynthesis. eLife 3: e03702
Model evoluce – klíčové kroky zvyšující fitness:1) genové duplikace2) zvýšení poměru BSC:MC3) glycinová pumpa

Brown et al. 2011, Science
C4 metabolismus – dávná strukturn ě regula ční predispozice?
v kódující sekvenci NAD-ME je úsek navozující expres i v pochvách cévních svazk ů (i u ortolog ů z C3 !)
Independent and Parallel Recruitment of Preexisting Mechanisms Underlying C4 Photosynthesis
C4 fotosyntéza v jedné buňce- dva typy chloroplastů (periferní bez Rubisco, central compartment: s Rubisco)
Lung, S.-C., Yanagisawa, M. and Chuong, S.D. (2012). Isolation of dimorphic chloroplasts from the single-cell C4 species Bienertia sinuspersici. Plant Methods. 8: 1-13; Rosnow, J., Yerramsetty, P., Berry, J.O., Okita, T.W. and Edwards, G.E. (2014). Exploring mechanisms linked to differentiation and function of dimorphic chloroplasts in the single cell C4 species Bienertia sinuspersici. BMC Plant Biology. 14: 1-18.
Bienertia sinuspersici

C4 metabolismus v jedné bu ňcePEPc perifern ě, NAD-ME centráln ě (Bc ) či u cévních sv.v mitochondriích obklopených plastidy s Rubisco ( Ba)
Borszczowia aralocaspica Bienertia cycloptera
C4 fotosyntéza
- 3 % druh ů, 5 % rostlinné biomasy- až 20 % celkové rostlinné fixace CO 2! (travní porosty)Hospodá řsky významné C4 rostliny :
- plodiny: kukuřice, cukr. třtina, proso, merlík- plevely: lebeda, merlík
C4 se vyplatí v sušších a teplejších podmínkách:
- šet ří vodu (!!!) – přivřené průduchy (stačí méně CO2) - ale energeticky náročnější: PPDK/PEPCK + 1-2 ATP/CO2
- málo účinná při nižší teplotě (PPDK, Rubisco)
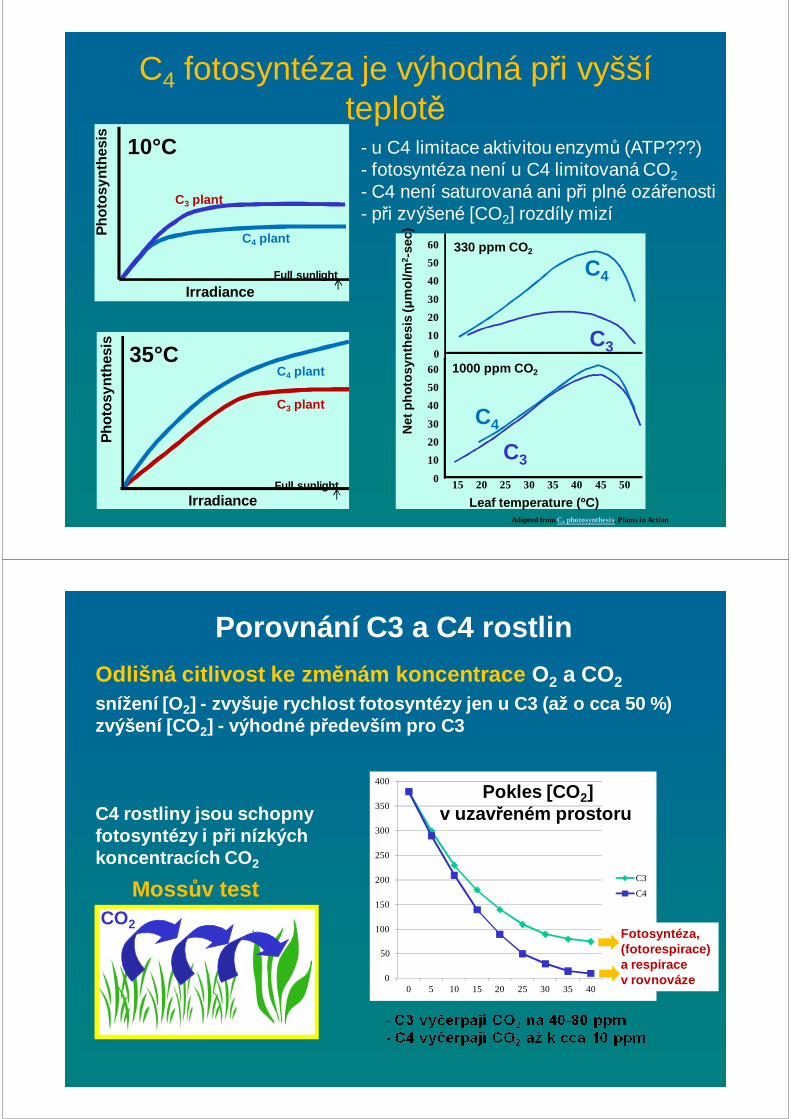
C4 fotosyntéza je výhodná při vyšší teplotě
C3 plant
C4 plant35°C
IrradianceFull sunlight
Pho
tosy
nthe
sis
10°C
C3 plant
C4 plant
IrradianceFull sunlight
Pho
tosy
nthe
sis
Adapted from C4 photosynthesis, Plants in Action
- u C4 limitace aktivitou enzymů (ATP???) - fotosyntéza není u C4 limitovaná CO2
- C4 není saturovaná ani při plné ozářenosti- při zvýšené [CO2] rozdíly mizí
50
Net
pho
tosy
nthe
sis
(µ
mol
/m2 -
sec)
0
10
20
40
30
50
60
0
10
20
40
30
50
60
Leaf temperature (ºC)20 30 4015 3525 45
330 ppm CO 2
1000 ppm CO 2
C4
C3
C4
C3
snížení [O 2] - zvyšuje rychlost fotosyntézy jen u C3 (až o cca 50 %) zvýšení [CO 2] - výhodné p ředevším pro C3
C4 rostliny jsou schopny fotosyntézy i p ři nízkých koncentracích CO 2
0
50
100
150
200
250
300
350
400
0 5 10 15 20 25 30 35 40
C3
C4
Pokles [CO 2]v uzav řeném prostoru
Porovnání C3 a C4 rostlin
Odlišná citlivost ke zm ěnám koncentrace O2 a CO2
CO2Fotosyntéza, (fotorespirace) a respirace v rovnováze
Mossův test

Rozdíly mezi rostlinami typu C3 a C4
Parametry fotosyntézy rubisco PEPc
CO2:ATP:NADPH (teoreticky – netěsnosti!) 1 : 3 :2 1 :5 :2
teplotní optimum °C (rubisco, PPDK) 15 - 25 30 - 45
kompenzační koncentrace CO2 (ppm) 30 - 50 0 - 10
důsledky poklesu koncentrace O2 PN se zvýší PN se nemění
max. rychlost čisté fotos. PN(mol dm-2 h-1) 15 - 35 40 - 60
produkce sušiny (t ha-1 rok-1) 22 38
transpirační koeficient Tk (g H2O/g CH2O) 450 - 1100 250 - 350
účinnost využití N (produkce sušiny na množství vázaného dusíku) Nízká Vysoká
isotopová diskriminace δ13CO2 (promile) -22 až -34 -11 až -19
C3 C4Primární karboxylace
Účinnost využití vody a dusíku u C3 a C4 rostlin
Rýže (C3) Kuku řice (C4)
Vody (g DM / kg vody) 1,4 2,6 (1,9x)Dusíku (µmol CO2 s-1 / mmol N) 0,26 0,74 (2,8x)Záření (g DM / MJ pohlcené FAR) 2,2 3,3 (1,5x)
Účinnost využití dusíku = rychlost asimilace CO2 vztažená na obsah dusíku v listu
- většina dusíku je vázaná v proteinech, - u C3 rostlin tvoří až polovinu všech proteinů enzym rubisco - u C4 je méně rubisco, tedy dusík je efektivněji využit
Účinnost využití vody (produkce biomasy na jednotku vytranspirované vody)– převrácená hodnota TK (transpiračního koeficientu)- menší otevřenost průduchů (C4 fotosyntéze stačí nižší [CO2])
(orientační hodnoty: velký vliv stanoviště, klimatu, genotypu apod.!)

Diskriminace 13CO2 ve fotosyntéze je vyšší u C3
- Rubisco p řednostn ě fixuje 12CO2, pokud má na výb ěr(u C4 má rubisco v uzavřeném prostoru méně na výběr)
δ13C = ((13C/12C)vzorek - (13C/12C)atm. )) / (13C/12C)atm.
C3C4
⇒ diskriminuje rubisco, PEPc nerozlišuje 13C a 12C
Isotopová diskriminace nejen Rubisco - projevuje se i v mnoha dalších
procesech a reakcích
Von Caemmerer, S., Ghannoum, O., Pengelly, J.J.L. and Cousins, A.B. (2014). Carbon isotope discrimination as a tool to explore C4 photosynthesis. J. Exp. Bot. 65: 3459-3470by permission of Oxford University Press.
Respiration (mesophyll)
Isotope discrimination during diffusion into mesophyll cell
Isotope discrimination by carbonic anhydrase and PEPC
Isotope discrimination
by Rubisco
Respiration (bundle sheath)

CAM fotosyntéza = „sukulentní“(Crassulacean acid metabolism)
- speciální typ C4 fotosyntézy (= C4 meziprodukt)- adaptace na kritický nedostatek vody- první výskyt už před 200 mil let (Proteaceae), - recentní sukulenty především v třetihorách (40 mil let, hlavní radiace 15 mil let)- 6-7 % druhů (33 čeledí)
Problém : - E pro fixaci CO2 lze získat jen za dne- otevření průduchů = obrovské ztráty vody
HYPOTETICKÁ ŘEŠENÍ?
Frerea indica- CAM ve stoncích, C3 v opadavých listech
CAM fotosyntéza = C4 odd ělená časově !Noc- otevřené průduchy (↓ ztráta vody)- primární fixace CO2 (PEPc)- štěpení škrobu (zdroj PEP, NADH)- skladování MA ve vakuole
Den- zavřené průduchy- fotochemické reakce- dekarboxylace malátu (ME)- fixace CO2 Calvinovým cyklem
Plastická regulace: je-li dost vody, pak fixace i přes den (sezónní změny), je-li extrémní sucho, průduchy zavřené i v noci (CAM idling)
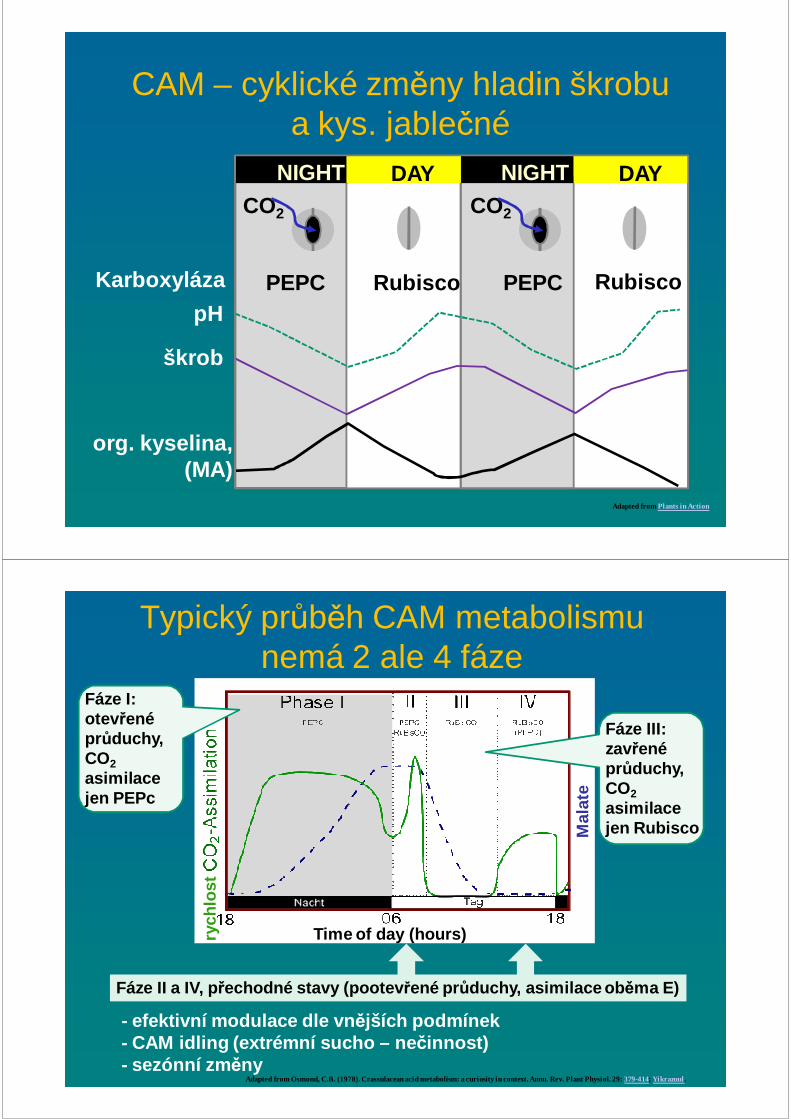
CAM – cyklické změny hladin škrobu a kys. jablečné
Adapted from Plants in Action
NIGHT NIGHTDAY DAYCO2
org. kyselina,(MA)
CO2
Karboxyláza PEPC PEPC RubiscoRubiscopH
škrob
Mal
ate
Time of day (hours)
Typický průběh CAM metabolismu nemá 2 ale 4 fáze
Adapted from Osmond, C.B. (1978). Crassulacean acid metabolism: a curiosity in context. Annu. Rev. Plant Physiol. 29: 379-414; Yikrazuul .
Fáze I: otev řené průduchy,CO2asimilacejen PEPc
Fáze III: zavřené průduchy, CO2asimilacejen Rubisco
Fáze II a IV, přechodné stavy (pootev řené průduchy, asimilace ob ěma E)
rych
lost
- efektivní modulace dle vn ějších podmínek- CAM idling (extrémní sucho – ne činnost)- sezónní zm ěny

CAM cyklování (∼ C3) a nečinnost
Adapted from Kerbauy, G.B., Takahashi, C.A., Lopez, A.M., Matsumura, A.T., Hamachi, L., Félix, L.M., a Pereira, P.N., Freschi, L., and Mercier, H. (2012) Crassulacean acid metabolism in epiphytic orchids: Current knowledge, future perspectives. In Applied Photosynthesis. Pp. 81-104.
Stomatal features
Atmospheric CO 2 uptake
Organic acid accumulation
Carbohydrate fluctuation
CAM cycling: - kombinace C3 a CAM - fixace b ěhem dne Rubisco- v noci PEPc
CAM idling:- extrémní sucho- reasimilace vydýchaného CO 2
Rozšíření CAM je spojené s poklesem CO2 a desertifikací
Reprinted with permission from Arakaki, M., Christi n, P.-A., Nyffeler, R., Lendel, A., Eggli, U., Ogburn, R.M., Spriggs, E., Moore, M.J. and Edwards, E.J. (2011). Contemporaneous and recent radiations of the world's major succulent plant lineages. Proc. Natl. Acad. Sci. USA. 108: 8379-8384.
[CO2]
[CO2]
Relativní globální teplota
CAM

Výskyt CAM druhůKDE?
Reprinted from Borland, A.M., Hartwell J., Weston D.J., Schlauch K.A., Tschaplinski T.J., Tuskan G.A., Yang X., and Cushman J.C. (2014). Engineering crassulacean acid metabolism to improve water-use efficiency. Trends Plant Sci. 19: 327-338with permission from Elsevier; Adapted from Lüttge U. 2010. Ability of crassulacean acid metabolism plants to overcome interacting stresses in tropical environments. AoBPLANTS 2010: plq005.
~300
~9500
~1250
~400
~1500
Počet druh ů- především xerofyty a epifyty
Fakultativní CAM při stresu suchem
Winter K., and Holtum, J.A.M. (2014). Facultative crassulacean acid metabolism (CAM) plants: powerful tools for unravelling the functional elements of CAM photosynthesis. J. Exp.Bot. 65: 3425-3441by permission of Oxford University Press; Brilhaus, D., Bräutigam, A., Mettler-Altmann, T., Winter, K. and Weber, A.P.M. (2016). Reversible burst of transcriptional changes during induction of crassulacean acid metabolism in Talinum triangulare. Plant Physiol. 170: 102-122.
Daytime gas exchange Daytime gas exchangeNighttime gas exchange
Water withheld Water provided
Suchem indukovaná no ční akumulace org. kyselin
Talinum triangulare
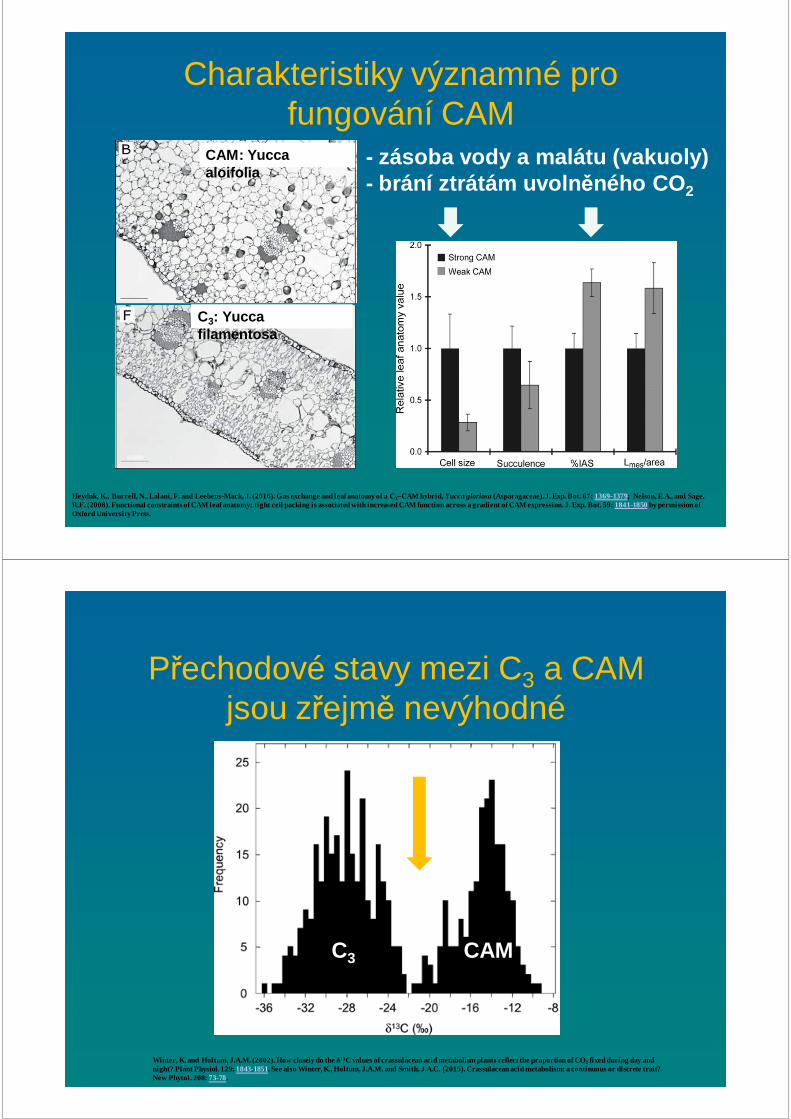
Charakteristiky významné pro fungování CAM
Heyduk, K., Burrell, N., Lalani, F. and Leebens-Mack, J. (2016). Gas exchange and leaf anatomy of a C3–CAM hybrid, Yucca gloriosa (Asparagaceae). J. Exp. Bot. 67: 1369-1379; Nelson, E.A., and Sage, R.F. (2008). Functional constraints of CAM leaf anatomy: tight cell packing is associated with increased CAM function across a gradient of CAM expression. J. Exp. Bot. 59: 1841-1850by permission of Oxford University Press.
CAM: Yucca aloifolia
C3: Yucca filamentosa
- zásoba vody a malátu (vakuoly)- brání ztrátám uvoln ěného CO 2
Přechodové stavy mezi C3 a CAM jsou zřejmě nevýhodné
Winter, K. and Holtum, J.A.M. (2002). How closely do the δ13C values of crassulacean acid metabolism plants reflect the proportion of CO2 fixed during day and night? Plant Physiol. 129: 1843-1851. See also Winter, K., Holtum, J.A.M. and Smith, J.A.C. (2015). Crassulacean acid metabolism: a continuous or discrete trait? New Phytol. 208: 73-78.
C3 CAM
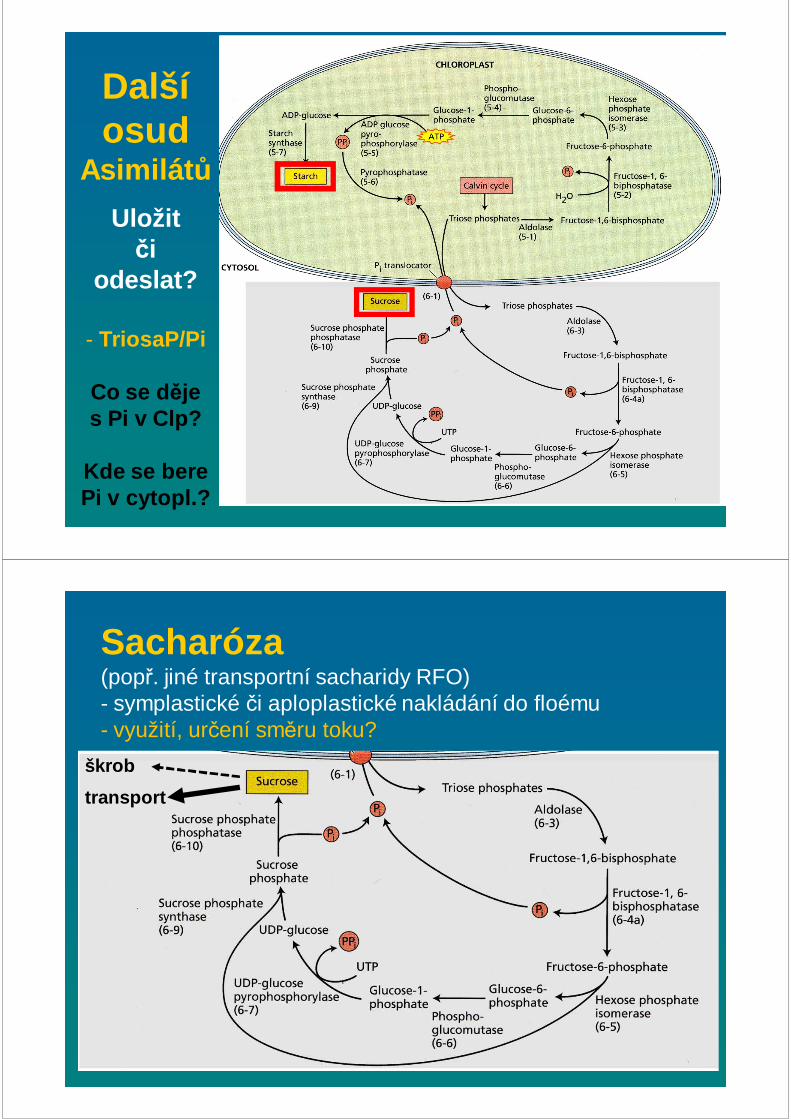
Další osud
Asimilát ů
Uložit či
odeslat?
- TriosaP/Pi
Co se d ěje s Pi v Clp?
Kde se berePi v cytopl.?
Sacharóza (popř. jiné transportní sacharidy RFO)- symplastické či aploplastické nakládání do floému- využití, určení směru toku?
škrob
transport
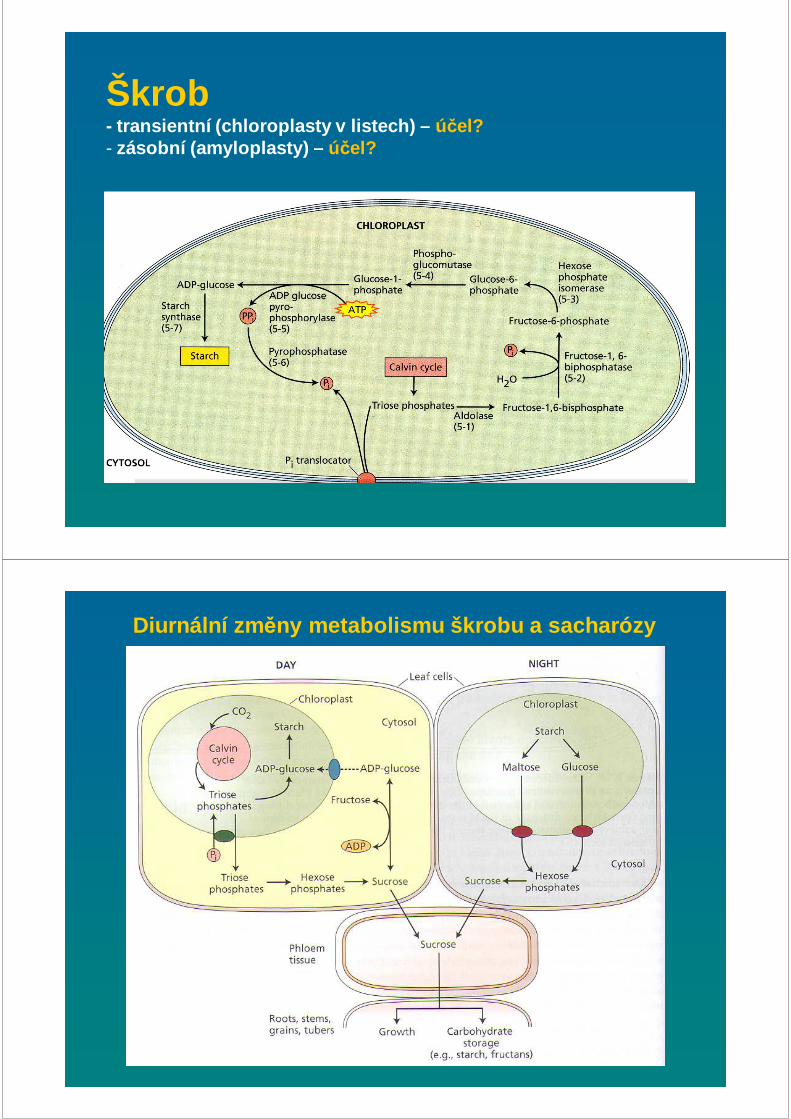
Škrob - transientní (chloroplasty v listech) – účel?- zásobní (amyloplasty) – účel?
Diurnální zm ěny metabolismu škrobu a sacharózy


















