
Mating systems, sociality and mate choice in animals: contributions of molecular ecology.
64
Mating systems, sociality and mate choice in animals: contributions of molecular ecology
-
Upload
julie-carol-mathews -
Category
Documents
-
view
217 -
download
1
Transcript of Mating systems, sociality and mate choice in animals: contributions of molecular ecology.

Mating systems, sociality and mate choice in animals:
contributions of molecular ecology

Mating Systems and Sexual Selection
• Diversity of mating systems– Polyandry, Polygyny, Polygynandry, and
Monogamy
• Consequences: male-male competition, female choice– Honest signals, good genes– Runaway selection, exaggerated characters,
linkage of genes for female preference with genes for male traits

Mating Systems and Sexual Selection• Polyandry
– One female, several males– Male jacanas (Microparra spp.) do most nest building– After the female has laid a clutch of four eggs, the
male takes over the parenting responsibilities– Jacana nests are built on mostly submerged plants. If
the nest starts to sink, the male carries to a new site– Meanwhile, the female has left the male to find more
males to breed with– She does not participate in raising chicks– If the eggs or chicks are lost, she will return to breed
and produce a replacement clutch with the first male

Mating Systems and Sexual Selection
• Polygyny– One male, several females– Much male-male competition. Males
compete for females by defending important resources (like foraging grounds in blackbirds) or by attracting females to display grounds (like grouse leks)
– In birds, some males defend female harems• Montezuma oropendola• Males defend colonies of females, alpha male
rules (like elephant seal) and gets most of copulations (and pass on most genes)
• Females do all the nestling care– Clue to occurrence is sexual dimorphism
(larger males) and grouping behavior of females

• In a Costa Rica population of this bird, high-ranking males defended groups of females at nesting colonies
• Alpha vs. beta vs. low-ranking males• DNA fingerprinting assessed paternity

Apparent alpha (RRR) and beta (OMO) male mating success (based upon behavior observation)

Analysis of DNA fingerprint data
• Jeffreys 33.15 probe, and M13 probe• Multilocus band patterns on gels• Band sharing S=2n/T
– N=# bands shared– T=total # bands in both lanes
• Within group relatedness r=(Sw-Sb)/(1-Sb)
– Sw = band sharing within groups
– Sb =band sharing between groups
– Group = colony presumably governed by one male
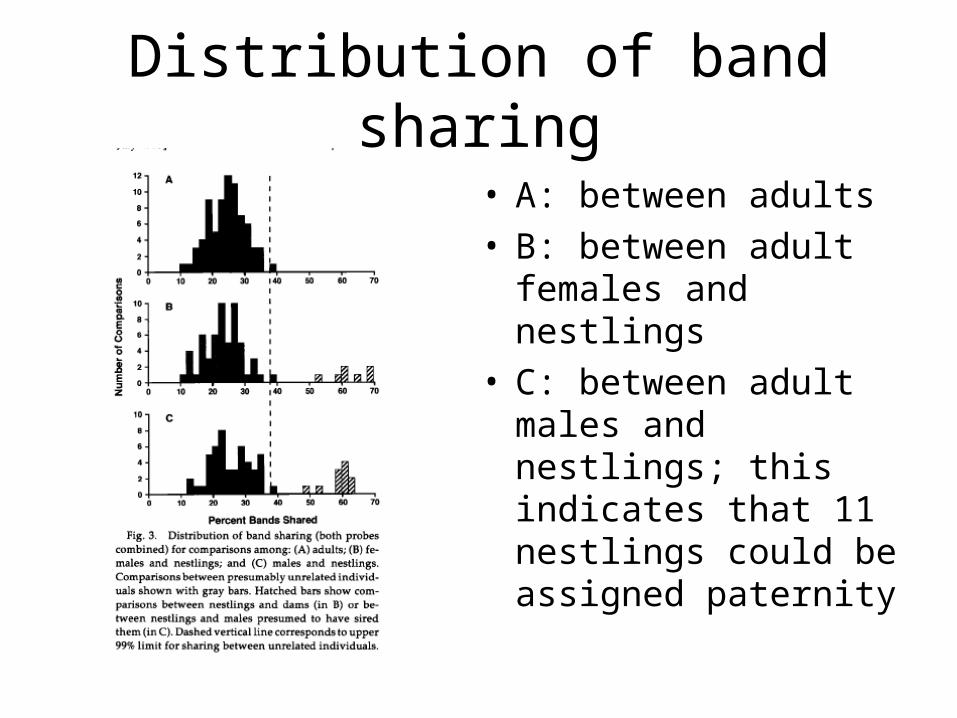
Distribution of band sharing
• A: between adults• B: between adult
females and nestlings• C: between adult
males and nestlings; this indicates that 11 nestlings could be assigned paternity

Revelations due to fingerprinting
• Paternity was examined for 21 nestlings from four colony sites.
• Seven nestlings matched with the alpha male at their colony, 4 matched with the beta male, and 10 did not match any sampled male.
• Fertilization success of alpha males was significantly lower than expected from the observed copulations
• Paternity assignment and levels of band sharing among nestlings indicate that most nestlings not attributable to the alpha were sired by several low-ranking males copulating away from the colony.

Within group relatedness
r=(Sw-Sb)/(1-Sb)


Mating system of sea lions
• Pinnipeds often show ‘‘unconventional’’ and ‘‘sneaky’’ mating tactics to increase their reproductive success, and these can be less easy to observe.
• Males tend to control a “harem” (Harem holders)
• They sometimes control more than one harem at the same time, or switch between being the holder of one harem and the peripheral of another harem.
• Use DNA to discover the true pattern of mating success!

% of harem progeny sired by harem holder brackets are number of progeny sired by harem holder


Issues in this study
• Compares reproductive patterns with size of population
• But power of paternity inference decreases with population size


Mating Systems and Sexual Selection
• Polygynandry – multiple males and females– Smith’s Longspur
• Females pair and copulate with more than one male for a single clutch of eggs
• Males pair and copulate with two or more females• Intense sperm competition due to promiscuous
females• Several males routinely feed a single brood• All males in a neighborhood sing SAME song
– Allows males to easily recognize intruders and females to mate only with local males who will help her raise brood
Briskie 1999

• Paternity analysis: – digest DNA with MboI or Alu1– Run on gel, probe with one of several probes (per, Jeffreys,
33.15, 33.6 and lambda)





Monogamy
• Common Crossbill (Kleven et al. 2008)
– Many passerines seem to have mates for life
– However extra-pair paternity may be common in socially monogamous passerines


Monogamy
• Common Crossbill (Kleven et al. 2008)– no evidence of extrapair paternity among 96 offspring
in 34 examined broods. – Common Crossbills thus seem to represent an
exception to the rule of extrapair mating among socially monogamous passerine bird species.
– A potentially important selective pressure preventing promiscuity
– in Common Crossbills is the harsh environmental conditions experienced during breeding at wintertime, which may increase the importance of paternal care and limit the time available for seeking extrapair copulations.

Tradeoffs
• Migration may limit resources (energy and time) available for investment in parental care and territory defense
• Sexual selection for parental care may reduce time and energy available for migration– Breeding systems may
determine or respond to migration behavior
Shorebirds: Garcia-Pena et al. 2009)

Costs of breeding systems
More male:male competition leads to increased bias toward male mortality
Female-female competition does not
Parental care also is costly
Liker and Szekely 2005

Conservation implications• Polygyny can be costly
• In sage-grouse modelling study Ne was only 19% of N because of variation in reproductive success between years and the skew in breeding sex ratio
• Resulted in small effective population (42) which may suffer inbreeding effects (low hatchability was observed
• Need to manage for larger than one might expect population sizes
(Stiver et al. 2008)

Conservation implications• Flexible mating systems may
counteract synchronized environmental fluctuations
• When sex ratio becomes skewed, if polygamy can occur, then chance of extinction is reduced
• Lesser Spotted Woodpecker in Germany– Models suggest that when male
skew in sex ratio occurs, then polyandry would reduce probability of extinction
(Rossmanith et al. 2006)

Optimal Group Size
• Sociality can reduce individual workload, fear, predation, finding unpredictable foods, etc
• Sociality can increase disease, competition, conspicuousness, cuckoldry
• Tradeoffs can give optimal size
• Kin selection theory may explain evolution of groups


Raven Information Centers• (1) Roosts comprised both knowledgeable
and naive foragers. • (2) Departures from roosts were highly
synchronized, with most members departing in one direction.
• (3) Direction of departure often changed from day to day.
• (4) Birds made naive of food sources (by being withheld from the wild and then allowed to join roosts) followed roost-mates to new feeding sites, whereas control birds held and released outside of roosts rarely found the local food bonanzas.
• (5) Birds made knowledgeable of food sources (by being released at new carcasses) joined roosts and led roost-mates to the food on three of 20 occasions.
• (6) The same individuals switched leader and follower roles depending upon their knowledge of feeding opportunities.
(Marzluff et al. 1996)

Why do groups form?
Not Kin Selection
Parker et al. 1994

Animal mating systems:viewpoint of a quantitative
geneticist (Stevan J. Arnold)

Qualitative classification of mating systems
• Monogamy, polygamy, polyandry (Darwin 1871)
• Monogamy, resource defense polygyny, harem defense polygyny, explosive mating assemblage, leks, female access polyandry …(since Darwin)

Limitations of qualitative classifications
• Progeny can be produced by matings that are difficult to observe.
• Difficult to specify how the categories grade into one another.
• Essential differences may masquerade under the same name.
• For all these reasons, we need quantitative characterizations.

Determination vs characterization of mating systems
Spatial distribution of
resources
System of mating
“Intensity of sexual selection”
Temporal availability of
the limiting sexOperational
Sex ratio
Emlen & Oring 1977
Variation in reproductive success

Fundamental information about the mating system is captured in the
parental table
POLYGYNY + POLYANDRYFemales
Males 1 2 3 4 5 6 7 8 no. mates no. offspring1 5 6 2 3 132 3 8 2 3 133 2 6 2 3 104 2 3 6 6 4 175 1 1 16 2 1 27 2 1 28 0 0
no. mates 2 1 3 1 4 2 2 1no. offspring 6 6 7 8 8 9 8 6
Arnold & Duvall 1994

Selection theory measures
• Quantify Bateman’s three principles (variance in mating success, variance in offspring number, relationship between offspring number and mating success)
• Standardized variances, regression slopes
• Direct connection to theory for selection on quantitative traits
• Is, Is; I, I; βss, βss
Bateman 1948, Crow 1958, Wade 1979, Wade & Arnold 1980, Arnold & Duvall 1994, Shuster & Wade 2003

The relationship between βss, Is, and I
3 2 1
12
Male Bateman Gradient
0
2
4
6
8
10
12
14
0 1 2 3 4 5 6 7
Number of mates
Nu
mb
er o
f o
ffsp
rin
g
βss=slope= 1.46 offspring/mate
Is=0.21
I=0.18

Properties of a selection opportunity, I
• Equals variance in relative fitness
• Sets upper limit on the magnitude of directional, stabilizing (disruptive), and correlational selection
• When this variance is zero, there can be no sexual selection

Properties of a Bateman gradient
• Equals the slope of the regression that relates reproductive success (offspring) to mating success (mates that bear progeny)
• Part of the selection that acts on every sexually-selected trait
• The final common path between sexually-selected traits and fitness
• When this gradient is zero, there can be no sexual selection
Arnold & Duvall 1994

The relationship between βss, Is, and I
3 2 1
12
Male Bateman Gradient
0
2
4
6
8
10
12
14
0 1 2 3 4 5 6 7
Number of mates
Nu
mb
er o
f o
ffsp
rin
g
βss=slope= 1.46 offspring/mate
Is=0.21
I=0.18

A parental table and Bateman plots derived from it
POLYGYNY + POLYANDRYFemales
Males 1 2 3 4 5 6 7 8 no. mates no. offspring1 5 6 2 3 132 3 8 2 3 133 2 6 2 3 104 2 3 6 6 4 175 1 1 16 2 1 27 2 1 28 0 0
no. mates 2 1 3 1 4 2 2 1no. offspring 6 6 7 8 8 9 8 6
Male Bateman Gradient
-5
0
5
10
15
20
0 1 2 3 4 5
Number of mates
Nu
mb
er
of
off
sp
rin
g
Female Bateman Gradient
0
2
4
6
8
10
0 1 2 3 4 5
Number of mates
Nu
mb
er
of
off
sp
rin
g

The Bateman gradient as a part of selection on a trait
Arnold & Duvall 1994

Same thing with plant mating systems, but with inbreeding

Parental table and Bateman plots for a population with partial selfing
Parental Table with number of offspring as entries = selfed progenyPARTIAL SELFING =outcross progeny
FemalesMales 1 2 3 4 5 6 7 8 no. mates no. offspring
1 1 1 2 22 4 2 1 1 2 5 103 4 1 1 3 64 5 1 3 2 4 115 1 2 1 2 4 1 6 116 6 1 3 3 107 2 4 1 4 4 118 5 1 5
no. mates 3 5 5 4 3 4 4 0no. offspring 6 14 10 7 7 10 12 0
Male Bateman GradientPollen parentage
0
2
4
6
8
10
12
14
0 2 4 6 8
Number of mates
Nu
mb
er o
f o
ffsp
rin
g
Female Bateman GradientOvule parentage
-2
0
2
4
6
8
10
12
14
16
0 1 2 3 4 5 6
Number of mates
Nu
mb
er o
f o
ffsp
rin
g

Theoretical perspective: connections to evolutionary theory
Inbreeding depression
Inbreeding coefficient
Selection on selfing rate Inheritance
Evolution of selfing rate
Selfing rate
Lande & Schemske 1985

Summary of insights from the empirical perspective
DATA EVOLUTIONARY PARAMETERS THAT CAN BE ESTIMATED
Mating success Opportunity for sexual selection
Reproductive success Opportunity for fecundity selection, Bateman gradients
Traits in males and females
Sexual and fecundity selection gradients
Traits in offspring Heritabilities (G-matrix), response to selection
Fitness of offspring Heritability of mating and reproductive success, parental selection
Inbreeding coefficients or pedigree
Inbreeding depression, coefficients of inbreeding

Summary
• Characterization of mating systems using selection and inbreeding theory measures has advantages over other characterizations.
• The parental table offers a useful empirical perspective on mating systems, and can be inferred via paternity analysis.
• In some mating systems and for some purposes, the parental table needs to be supplemented with additional information (e.g., parental traits, offspring fitness).

MHC and Mate ChoiceMHC evolution and how we select a mate

Among the constellation of genes that control the immune system are those known as the major histocompatibility complex (MHC), which influence tissue rejection.
If the smell of MHC isn't a deal maker or breaker, the taste is. Saliva also contains the compound, a fact that Haselton believes may partly explain the custom of kissing, particularly those protracted sessions that stop short of intercourse. "Kissing," she says simply, "might be a taste test." (Kluger, 2008)
Conceive a child with a person whose MHC is too similar to your own, and the risk increases that the womb will expel the fetus.

Why Sex?
• Considered “the deepest mystery in all of biology.” (Trivers, 1985)

“Red Queen” hypothesis about parasite resistance
• New combinations of genes for resistance are required in every generation to cope with the currently dominating parasites.
“It takes all the running you can do, to keep in the same place.” (Carroll, 1872)

Major histocompatibility complex(MHC)
• 1940: Discovered in mice in connection with skin grafting
• Name refers to the region of several linked genes that controls skin grafting
• Connection to immune system not determined until 1960s/70s
Abbas and Lichtman, 2005

Abbas and Lichtman, 2005

MHC Genes
• Human MHC molecules are called Human Leukocyte Antigens (HLA)
• Most polymorphic genes in genome• Codominantly expressed• Categorized as Class I or Class II
Abbas and Lichtman, 2005

Three requirements for MHC to be a basis of mate choice
• Females must…– Know there is an enormous variation in resistance
genes in the population– Know her immune genes– Recognize the immune genes of potential mates and
choose

The infamous T-Shirt test• Males and Females
were typed for HLA- A, -B, and -DR
• Males wore shirts for two consecutive nights
• Women rated smell of shirts on pleasantness, intensity, and memory association
• Two sample populations:– Oral contraceptive users– Non-oral contraceptive
users


Results

“Red Queen” is still a hypothesis
• Need evidence showing selection for rare MHC alleles, in response to the ever-changing parasite load
• This would the mechanism that maintains MHC polymorphism

Mate choice motives to choose different MHC genotype
• Reduce inbreeding?
• Specific MHC selection?
• Increase MHC heterozygosity?
• Maximize diversity across several loci?


















