
Mass mortality of Diadema...
11
Marine Biology 104, 67-77 (1990) Marine ................ Biology © Springer-Verlag 1990 Mass mortality of Diadema antillarum I. Long-term effects on sea urchin population-dynamics and coral reef algal communities R.C. Carpenter Department of Biology, California State University, Northridge, California 91330, USA Abstract The mass mortality of the echinoid Diadema antillarum Philippi in 1983/1984 resulted in dramatic changes in the benthic algal community. This study reports data on the population densities of sea urchins prior to and following the mass mortality and data on the biomass, species compo- sition, and rates of primary productivity of algal communi- ties at several study sites in St. Croix, U.S. Virgin Islands. The mass mortality reduced D. antillarum population densi- ties by 95 to 99%. Population densities were reduced further by a second, less severe mortality event in October 1985. Over a period of 2 yr, recruitment of D. antillarum larvae to the study sites was low and population densities remained at 2 to 30% of their premortality levels. The effects of the mass mortality on the algal community were significant. Algal biomass increased by 22 to 439% across reef zones 16 mo after the die-off. Rates of primary productivity per unit biomass (chlorophyll a) remained at approximately 60% of premortality levels in shallow-reef habitats for 25 mo fol- lowing the die-off. The magnitude of the response of the algal community was positively correlated with the previous population densities of D. antillarum. The species composi- tion of the algal community also shifted dramatically. Prior to the mass mortality, algal communities were dominated by algal turfs and crustose algae, and macroalgae were either rare or absent. Twenty-five months after the mass mortality, algal turfs covered 40% of the area and macroalgae covered 47%. Many of the macroalgal species are not consumed by herbivores and much of the algae is dislodged by storm waves and is exported from the reef. This represents a tran- sition from a community dominated by a grazing-based food web to one where the majority of primary production • may be exported to adjacent communities. Introduction Regular echinoids are important consumers of plant biomass in many marine communities (Lawrence 1975). The effect of grazing by sea urchins often depends on their pop- ulation density. Grazing by "fronts" of sea urchins in tem- perate and boreal kelp beds results in reduced algal biomass and species diversity (Leighton 1971, Pearse and Hines 1979, Scheibling 1986, Johnson and Mann 1988). Grazing by some species of sea urchins can also increase algal communi- ty diversity in some coral reef algal communities (Sammarco 1980, Carpenter 1981) and rocky intertidal habitats (Paine and Vadas 1969). The role of predators in controlling the population sizes of sea urchins, and thereby influencing the plant communi- ty, has received much attention, particularly the relation- ships between sea otters, sea urchins, and kelp beds on the west coast of North America (Estes and Palmisano 1974, Estes et al. 1978, Duggins 1980) and between lobsters, sea urchins, and kelp beds on the east coast of North America (Breen and Mann 1976, Wharton and Mann 1981, Vadas et al. 1986). Population sizes of sea urchins can also be affected significantly by disease which results in mass mor- tality. Mass mortalities of red sea urchins (Strongylocentro- tus franciscanus) in California resulted in the expansion of kelp forests (Pearse and Hines 1979, Dean et al. 1984) and annual die-offs of green sea urchins (S. droebachiensis) off the coast of Nova Scotia allowed the reestablishment of kelp beds in crustose coralline "barren grounds" that had been previously overgrazed (Chapman 1981, Scheibling 1986). In 1983-1984, the echinoid Diadema antillarum suffered mass mortalities throughout its range in the western Atlantic Ocean, including all areas of the Caribbean (Lessios et al. 1984). This is thought to be one of the largest documented mass mortalities to affect any marine organism (Lessios 1988a). Although not uniformly abundant in all regions prior to the mass mortality (Bauer 1980), this species was a major source of grazing disturbance and bioerosion in many coral reef and seagrass environments (Ogden et al. 1973, Sammarco et al. 1974, Lawrence 1975, Ogden 1977, Sam- marco 1980, 1982, Carpenter 1981, 1986). The mass mortal- ity provided the opportunity to use the near extinction of this species as a "natural experiment" to examine the effects
Transcript of Mass mortality of Diadema...

Marine Biology 104, 67-77 (1990) Marine . . . . . . . . . . . . . . . . Biology
© Springer-Verlag 1990
Mass mortality of Diadema antillarum
I. Long-term effects on sea urchin population-dynamics and coral reef algal communities
R . C . Carpenter
Department of Biology, California State University, Northridge, California 91330, USA
Abstract
The mass mortality of the echinoid Diadema antillarum Philippi in 1983/1984 resulted in dramatic changes in the benthic algal community. This study reports data on the population densities of sea urchins prior to and following the mass mortality and data on the biomass, species compo- sition, and rates of primary productivity of algal communi- ties at several study sites in St. Croix, U.S. Virgin Islands. The mass mortality reduced D. antillarum population densi- ties by 95 to 99%. Population densities were reduced further by a second, less severe mortality event in October 1985. Over a period of 2 yr, recruitment of D. antillarum larvae to the study sites was low and population densities remained at 2 to 30% of their premortality levels. The effects of the mass mortality on the algal community were significant. Algal biomass increased by 22 to 439% across reef zones 16 mo after the die-off. Rates of primary productivity per unit biomass (chlorophyll a) remained at approximately 60% of premortality levels in shallow-reef habitats for 25 mo fol- lowing the die-off. The magnitude of the response of the algal community was positively correlated with the previous population densities of D. antillarum. The species composi- tion of the algal community also shifted dramatically. Prior to the mass mortality, algal communities were dominated by algal turfs and crustose algae, and macroalgae were either rare or absent. Twenty-five months after the mass mortality, algal turfs covered 40% of the area and macroalgae covered 47%. Many of the macroalgal species are not consumed by herbivores and much of the algae is dislodged by storm waves and is exported from the reef. This represents a tran- sition from a community dominated by a grazing-based food web to one where the majority of primary production • may be exported to adjacent communities.
Introduction
Regular echinoids are important consumers of plant biomass in many marine communities (Lawrence 1975). The
effect of grazing by sea urchins often depends on their pop- ulation density. Grazing by "fronts" of sea urchins in tem- perate and boreal kelp beds results in reduced algal biomass and species diversity (Leighton 1971, Pearse and Hines 1979, Scheibling 1986, Johnson and Mann 1988). Grazing by some species of sea urchins can also increase algal communi- ty diversity in some coral reef algal communities (Sammarco 1980, Carpenter 1981) and rocky intertidal habitats (Paine and Vadas 1969).
The role of predators in controlling the population sizes of sea urchins, and thereby influencing the plant communi- ty, has received much attention, particularly the relation- ships between sea otters, sea urchins, and kelp beds on the west coast of North America (Estes and Palmisano 1974, Estes et al. 1978, Duggins 1980) and between lobsters, sea urchins, and kelp beds on the east coast of North America (Breen and Mann 1976, Wharton and Mann 1981, Vadas et al. 1986). Population sizes of sea urchins can also be affected significantly by disease which results in mass mor- tality. Mass mortalities of red sea urchins (Strongylocentro- tus franciscanus) in California resulted in the expansion of kelp forests (Pearse and Hines 1979, Dean et al. 1984) and annual die-offs of green sea urchins (S. droebachiensis) off the coast of Nova Scotia allowed the reestablishment of kelp beds in crustose coralline "barren grounds" that had been previously overgrazed (Chapman 1981, Scheibling 1986).
In 1983-1984, the echinoid Diadema antillarum suffered mass mortalities throughout its range in the western Atlantic Ocean, including all areas of the Caribbean (Lessios et al. 1984). This is thought to be one of the largest documented mass mortalities to affect any marine organism (Lessios 1988a). Although not uniformly abundant in all regions prior to the mass mortality (Bauer 1980), this species was a major source of grazing disturbance and bioerosion in many coral reef and seagrass environments (Ogden et al. 1973, Sammarco et al. 1974, Lawrence 1975, Ogden 1977, Sam- marco 1980, 1982, Carpenter 1981, 1986). The mass mortal- ity provided the opportunity to use the near extinction of this species as a "natural experiment" to examine the effects

68 R.C. Carpenter: Diadema mass-mortality effects/population dynamics and algae
of the removal of sea urchins on coral reef communities and allow comparisons with the results of previous experimental manipulations (Carpenter 1985 a). This study presents data on the population dynamics of D. antillarum and on reef algal-community parameters and compares them to data collected for two years at the same study locations following the mass mortality.
Study site
This study was conducted at several sites on Tague Bay Reef (see Carpenter 1986) and one site at Grass Point on the northeast coast and south shore, respectively, of St. Croix, U.S. Virgin Islands (17°45'N; 64°42'W). Tague Bay reef is a bank barrier reef (sensu Adey 1978) comprised largely of the scleractinian corals Aeropora palmata, Montastrea annu- laris, Porites porites, Diploria strigosa, and Agaricia agari- cites, and the hydrocoral Millepora complanata. Much of the Acropora palmata has been killed within the past 10 yr by white-band disease (Gladfelter 1982) and is covered by crus- rose and filamentous algae. The study area at Grass Point is on the leeward side of the point and consists of a coral cobble reef that extends from shore westward for 50 m where it grades into a fringing-type reef comprised primarily of A. palmata and Millepora complanta.
Two sites, approximately 10 m apart, were established in each of four reef zones (i.e., depths) on Tague Bay Reef in July 1983 for placement of the experimental algal communi- ties (see "Materials and methods" - Algal community dy- namics). These zones were the backreef/reefcrest (2 m depth), shallow forereef (2 m depth), mid-forereef (5 m depth), and deep forereef (10 m depth).
Materials and methods
Population densities and sizes of Diadema antillarum
The number of individuals and the sizes (maximum test diameter, MTD) of all Diadema antillarum Philippi occur- ring in 20 to 50, 1 m 2 quadrats were recorded during ten sampling times between December 1983 and March 1986 at the Tague Bay Reef sites. The quadrats were located con- tiguously along a transect between the two established sites within each reef zone. The areas censused spanned the range of microhabitats within the study areas. Sizes of the sea urchins were measured to the nearest millimeter using long- jawed calipers. The only data collected at the Grass Point study site were for population densities and individual sizes of D. antillarum. These data were collected in December 1983, September 1984, December 1985, and March 1986.
Algal community dynamics
The response of reef algal communities to the mass mortal- ity of Diadema antillarum was quantified by measuring
changes in algal communities on natural reef substrata for 25 mo following the mass mortality. The substrata were cross-sections of the coral Acropora palmata (64 cm 2) that were first submerged in December 1981 at the backreef/ reefcrest site (Carpenter 1986). Six months after the plates had been submerged, the algal communities on the plates were similar to those on the surrounding natural substra- tum. Plate assemblies were haphazardly allocated to one of the two sites within each reef zone (depth) in July 1983.
Data on the rates of algal primary productivity, algal biomass, and algal species composition were collected on the same sampling dates given above. For each sampling date, three plates were chosen randomly from each replicate plate assembly in each reef zone. Rates of primary productivity were estimated by incubating the plates in in situ acrylic chambers at saturating photon-flux densities and measuring changes in dissolved oxygen. Respiration rates were estimat- ed by covering the chambers with black acrylic boxes, allow- ing the algae to acclimate, and again measuring changes in dissolved oxygen. Details on the methodology to estimate rates of primary productivity are given in Carpenter (1985b).
Algal biomass was estimated as decalcified dry mass/ area and as the mass of chlorophyll a/area. Subsamples (three 1 c m 2 samples, located randomly using an overlying coordinate grid) for dry algal mass were taken from the plates after they were returned to the laboratory. Algae were scraped from the plates to a depth of I mm into the substra- tum to sample endolithic forms and placed in weak acid (5% HC1) and 1% formalin solution to decalcify, filtered onto a preweighed filter, rinsed with deionized water, and dried to constant mass at 60 °C. Subsamples for chlorophyll a deter- mination were scraped from the substratum as described above, ground in a mortar and pestle with I ml of a saturat- ed MgCO 3 solution and 90% acetone, and kept in the dark for 20 to 24 h at 4°C. Chlorophyll a was determined spec- trophotometrically using the equations of Jeffrey and Humphrey (1975).
Algal species composition was estimated by several tech- niques to provide adequate sampling of algal communities with widely varying community structure. Before the mass mortality of Diadema antillarum, reef algal communities were dominated by algal turfs (Carpenter 1981, 1985a, b, 1986) that were best sampled by scraping subsamples (4 four l-cm 2 randomly located samples) from the plates as de- scribed above, decalcifying the algae, and mounting them on microscope slides. All subsamples for a plate were pooled and the presence of algal taxa noted as the entire slide was scanned at 100 x magnification. The relative abundances of algal taxa were estimated by counting the number of inter- sections of a 10 x 10 ocular grid that overlaid each taxon in 50 to 84 different viewing fields. The percent relative abun- dances were calculated by dividing the total number of inter- sections for each taxon by the total number of intersections for all taxa and multiplying by 100.
As algal community structure changed following the sea urchin mass-mortality, it became necessary to sample the plates additionally for larger macroalgae. This was done by

R.C. Carpenter: Diadema mass-mortality effects/population dynamics and algae 69
counting the number of intersections of an 8 cm x 8 cm grid (64 total points) that overlaid each macroalgal taxon on the plates. Addi t ional ly , the abundances of algal turfs, crustose algae (primari ly corallines), and macroalgae were est imated in 25 marked photoquadra t s (388 cm 2 in area) that were established at the backreef/reefcrest site. Photographs were taken with a Nikonos camera with a 28 mm lens and close- up framer in December 1983, Feb rua ry and Apri l 1984, and in March 1986. The resulting 35 mm color slides were pro- jected onto a sheet of paper with 130 dots spaced at 2 cm intervals. The number of dots falling on each algal compo- nent (algal turf, crustose algae, and macroalgae) was summed and expressed as a percent of the total algal cover (Littler and Lit t ler 1985).
To provide a par t ia l control for this uncontrol led natura l experiment (the mass mortal i ty) , experimental algal commu- nities were placed inside inclusion cages at the backreef/ reefcrest site with densities of Diadema antillarum that were typical at this site pr ior to the mass mor ta l i ty (6 individuals m - Z ; Carpenter 1986). This control t reatment was estab- lished in Apri l 1984, and da ta on algal pr imary product ivi ty and algal biomass were obtained in June and September 1984, December 1985, and March 1986.
Results
Popula t ion densities and size-frequency distr ibutions of Diaderna antillarum
Popula t ion densities of Diadema antillarum were highest in shallow forereef zones pr ior to the mass morta l i ty with a mean density of 13.4 individuals m -2 (Table 1). Popula t ion densities decreased with increasing water depth to less than 6 individuals m - 2 at 10 m, approximate ly the same popula- t ion density that was present in backreef/reefcrest habitats.
Signs of the mass morta l i ty of Diadema antillarum first appeared in St. Croix in late January 1984 (Lessios et al. 1984, Ogden personal communicat ion) on the west end of the island. The mor ta l i ty spread to the east, reached the established study sites in early February , and 5 to 7 d later resulted in the el imination of 95 to 99% of the D. antillarum popula t ion (Carpenter 1985a, 1988). Popula t ion densities were reduced over all reef zones (Table 1) as well as in other habi tats a round the island, such as seagrass beds and man- groves. Censuses of sea urchins conducted 5 and 12 d after signs of the mor ta l i ty had disappeared, suggested that a por t ion of the surviving popula t ion was within the reef framework. More D. antillarum individuals were found at each site after 12 d than had been present immediately fol- lowing the mortal i ty.
In October 1985, Diadema antillarum again showed signs of disease that paral leled those observed in 1984. Mor ta l i ty rates were lower than in 1984 and averaged approximate ly 50% (Table 1), a l though this is based on small sample sizes at each site. The second wave of mass morta l i ty affected D. antillarum popula t ions at all the study sites. As with the pr ior mor ta l i ty event, no other echinoid species was directly affected.
Table 1. Diadema antillarum. Mean (+_ SD) number of individuals per m 2 at all study sites over time before (December 1983) and after mass mortality, n: no. of 1 m 2 quadrats, nd: no data
Date Site
Backreef/ Forereef Forereef Forereef Grass reefcrest (2 m) (5 m) (10 m) Point
1983 Dec. 6.4 (3.6) 13.4 (4.1) 9 (5.9) 5.8 (6.5) 7.4 (1.5)
n = 20 n = 20 n = 20 n = 20 n = 34
1984 Feb. 0.1 (0.2) 1.1 (1.3) 0.3 (0.5) 0.3 (0.6) nd
n=20 n=20 n=25 n=33 Apr. 0.4 (0.8) 1.2 (2.1) 0.6 (1) 0.2 (0.5) nd
n=20 n=20 n=20 n=20 June 0.2 (0.4) 1.1 (1.5) 0.7 (1.1) 0.2 (0.4) nd
n=20 n=19 n=20 n=20 Sep. 0.t (0.3) 1.3 (1.2) 0.3 (0.4) 0 (-) 1.4 (1.2)
n=17 n=20 n=20 n=20 n=30 Dec. 0.1 (0.2) 1.6 (1.6) 0.3 (0.6) 0 ( ) nd
n=20 n=20 n=20 n=20
1985 June
Sep.
Dec.
1986 Mar.
0.1 (0.5) 0.5 (1.1) 0.2 (0.4) 0.2 (0.5) 5.6 (2.1) n=20 n=20 n=20 n=20 n=30
0.1 (0.4) 1 (1.3) 0.3 (0.8) 0.1 (0.3) nd n = 30 n = 40 n = 40 n = 20
0 (-) 0.4 (0.8) 0.3 (0.7) 0 (-) 3.1 (1.9) n=20 n=20 n=20 n=20 n=40
0 (-) 0.5 (0.9) 0.3 (0.8) 0.1 (0.2) 2.1 (2.4) n = 20 n = 20 n = 20 n = 20 n = 40
Mean individual sizes of Diadema antillarum ranged f rom 25 to 82 mm M T D before the morta l i ty and 32 to 66 mm M T D 5 d after the morta l i ty had run its course (Fig. 1). However, differences in the size-frequency distribu- tions before and immediately after the mass morta l i ty were not statistically significant (Kolmogorov-Smirnov test, p > 0.05).
Size-frequency distr ibutions of Diadema antillarum over all reef zones showed little evidence of recruitment since the mass mortal i ty. Where sea urchins were most abundant fol- lowing the mor ta l i ty event (forereef 2 m and forereef 5 m sites) and sample sizes were greatest, size-frequency distribu- tions shifted gradual ly into larger size classes. Only during Apri l 1984 at the forereef 2 m site was there a peak in the smaller size classes ( < 30 m m MTD), indicating pr ior re- crui tment of larvae. This pat tern of extremely low rates of recruitment has generally been mainta ined since March 1986, as the abundance o fD . antillarum larvae in the plank- ton and rates of settlement remain very low (up to June 1989, Carpenter unpublished data).
Algal communi ty dynamics
Algal biomass
The increase in algal biomass immediately following the mass morta l i ty (Carpenter 1988) was sustained over time.

70 R.C. Carpenter: Diadema mass-mortality effects/population dynamics and algae
Backreef/ Reefcrest (2 m) Forereef (2 m)
6 0 ~ 80t Dec" 1983 l
% 4O 20
0
o 6°1 II ' /"4°1 II I
. . . . . . . . . .
0
~;4 June 1984
% 4 0 ~ 2o
Forereef (5 m) Forereef (10 m)
April 1984
! II .... June 1984
% 40 No Diaderna Found
20 0 . . . . . . . . .
o ¢ ~ o o o 0 1 ~ o o o o o o o o o o o ¢ ~ o o o o o o o o o
MT° (mm) MTD (mm) MTD (mm) MTD (mm)
Backreef/ Reefcrest (2 m)
80 J Dec. 1984
0 ] . . . . . . . .
Forereef (2 m) Forereef (5 m) Forereef (10 m)
Dec. 1984
No Diaderna Found
6080 I June 198il % 40
20 0 . . . . . . . . .
80 "1 Sept. 1985
% 4 0
2 0
0
86~ t Dec. 1985
% 40 1 N°DiademaF°und I
. . . . . . . . I
80 .~ March 1986 6O
°/o 40 No D~adema Found 2O
0 . . . . . . . .
June 1985
Sept. 1985
0ec19851 Oec 198,
No Oiadema Found
o o o o o o o o o o o o o o o o o o o o o o o o o o o o ,~ ~, ,~ ~ ~ ~. = ~ 0 , . , ~ ~ ~- ~o o . . . . . . . ~ ~ ~. ~= . ~. ~. =. o ~ & - . ~ . - - o o . . . . . . . o ~ ' ~ ' = ~
MTD (ram) MTD (mm) MTD (ram) MTD ( r a m )
Fig. 1. Diadema antillarum. Mean test diameter (MTD) in four reef zones on Tague Bay Reef in St. Croix, U.S. Virgin Islands, before (December 1983) and after mass mortality. Number of individuals measured ranged from 20 to 100 within each reef zone
20 ~ a)'Backreef/Reeferest 20 T b) Forereef (2 m)
0 . . . . . . . . . . . 0 . . . . . . . . . m
~ 2 0 ]1 C) Forereef (5 rn) 20 ]1 d) Forereef (10 m)
10 10
0 0
•
03
Fig. 2. Mean ( 4- SD) algal biomass (mg decalcified dry mass cm- 2) at study sites in four reef zones before (for backreef/reefcrest site) and after (for all reef zones) mass mortality. ND: no data. Arrow marks approximate time of mass mortality
Decalcified algal biomass ranged from 1.5 to 3.5 mg cm -2 pr ior to the die-off of Diadema antillarum. Algal biomass increased in all reef zones over a per iod of two months following the morta l i ty and cont inued to increase thereafter at the backreef/reefcrest, forereef 2 m, and forereef 5 m sites (Fig. 2). Sixteen months after the die-off, algal biomass ranged from 4.5 to 15.1 mg cm -2 and was 319, 439, 305, and 22% higher at the backreef/reefcrest, forereef 2 m, forereef 5 m, and forereef 10 m sites, respectively, than when D. antillarum was abundan t (Steneck 1983). Algal biomass in some backreef and reefcrest habi tats increased to > 50 mg c m - z.
Algal species composition
Algal species composi t ion changed rapidly following the mass mortal i ty. The pat terns described here are for the backreef/reefcrest site; however, similar pat terns were noted in all of the reef zones. The relative abundance of algal components (algal turfs, crustose algae, and macroalgae) began to change within two mouths following the die-off (Fig. 3). Pr ior to the mass mortal i ty, algal turfs and crustose algae covered 90 and 10% of the area respectively. They covered only 50 and 1%, respectively, 2 mo following the mass mortal i ty. Algal turfs accounted for < 40% of the algal cover and crustose algae were reduced to < 1%, 25 mo after the die-off. Macroalgae, which were previously absent or extremely rare in these algal communities, increased in abundance from 12 to 38% cover 25 mo after the mass morta l i ty (see Fig. 4).
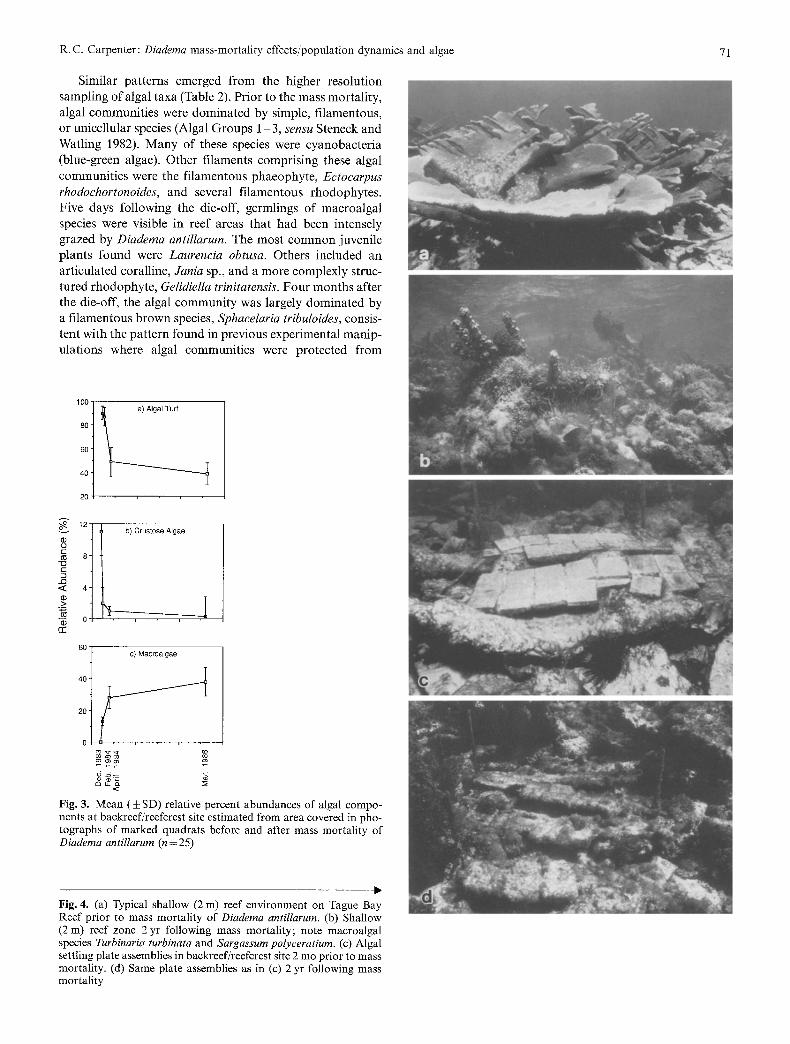
R.C. Carpenter: Diaderna mass-mortality effects/population dynamics and algae 71
Similar patterns emerged from the higher resolution sampling of algal taxa (Table 2). Prior to the mass mortality, algal communities were dominated by simple, filamentous, or unicellular species (Algal Groups 1 -3 , sensu Steneck and Watling 1982). Many of these species were cyanobacteria (blue-green algae). Other filaments comprising these algal communities were the filamentous phaeophyte, Ectocarpus rhodochortonoides, and several filamentous rhodophytes. Five days following the die-off, germlings of macroalgal species were visible in reef areas that had been intensely grazed by Diaderna antillarum. The most common juvenile plants found were Laurencia obtusa. Others included an articulated coralline, Jania sp., and a more complexly struc- tured rhodophyte, Gelidiella trinitatensis. Four months after the die-off, the algal community was largely dominated by a filamentous brown species, Sphacelaria tribuloides, consis- tent with the pattern found in previous experimental manip- ulations where algal communities were protected from
1 O0 "
8 0
6 0
40"
20
o~ 12 v
~9 t - 8
A21 < 4 CO >
- ~ 0 . . . . .
6O
4 0
2 0
0
c o c o
c) Macroa/gae
Fig. 3. Mean (4- SD) relative percent abundances of algal compo- nents at backreef/reefcrest site estimated from area covered in pho- tographs of marked quadrats before and after mass mortality of Diadema antillarum (n = 25)
Y
Fig. 4. (a) Typical shallow (2 m) reef environment on Tague Bay Reef prior to mass mortality of Diadema antillarum. (b) Shallow (2m) reef zone 2 yr following mass mortality; note macroalgal species Turbinaria turbinata and Sargassurn polyceratium. (c) Algal settling plate assemblies in backreef/reefcrest site 2 mo prior to mass mortality. (d) Same plate assemblies as in (c) 2 yr following mass mortality

72 R.C. Carpenter: Diadema mass-mortality effects/population dynamics and algae
Table 2. Mean relative percent abundances (-+ SD) of algal taxa occurring on settling plates at backreef/reefcrest site before sea urchin mass-mortality (December 1983), and 1 wk (February 1984), 8 wk (April 1984), and 108 wk (March 1986) following mass mortality (six settling plates sampled at each date). Asterisk denotes macroalgal taxa. Taxa identified using descriptions by Taylor (1960), Norris and Bucher (1982), and Kapraun et al. (1983)
Algal taxa Dec. Feb. April March 1983 1984 1984 1986
Cyanophyta Anabaina inequalis (K/itzing) Trevison 0 Brachytrichia quoyi (C. Agardh) < 1
Bornet and Flahault Calothrix crustacea Schousboe and Thuret 13± 3 Chrooeoceus minutus (Kfitzing) Nageli < 1 Hyella caespitosa Bornet and Flahault 2 ± 3 Lyngbya sordida (Zanardini) Gomont 2 ± 1 Lyngbya sp. 8 ± 7 Mastigocoleus testarum Lagerheim 3-4- 4 Schizothrix laeustris A. Braun 13_ 8 Scytonemafuliginosum Tilden 1 ± 1 Spirulina major K/itzing 0 Unidentified coccoid blue-greens 23 ± 6
Chlorophyta Acetabularia pusilIa (Howe) Collins < 1 Cladophora sp. < 1 Cladophoropsis membranaeea Taylor 0 Derbesia sp. < 1 Enteromorpha sp. < 1 Ostreobium quekettii Bomet and Flahault 2 ± 1 Unidentified encrusting greens 1 ± 1
Chrysophyta Pennate diatoms < 1
Phaeophyta Dictyota sp. * 0 Dictyotaceae sp. * 0 Eetoearpus rhodoehortonoides Borgensen 18 ± 17 Giffordia rallsiae (Vickers) Taylor 2_+ 1 Sphacelaria tribuloides Meneghini 1 ± 1
Rhodophyta Amphiroa fragilissima (Linnaeus) Lamouroux * 0 Amphiroa sp.* 0 Asparagopsis taxiformis (Delile) Collins < 1
and Hervey (Falkenbergia stage) Asteroeytis ramosa (Twaites) Gob± 0 AudouineIla sp. 0 Bangia atropurpurea (Roth) C. Agardh Montagne < 1 Centroeeras clavulatum (C. Agardh) 0 Ceramium flaecidum (Harvey ex Kiitzing) 0
Ardisonne Ceramium sp. 0 Champia sp. 0 Encrusting Corallines 8 ± 3 Gelidiaceae sp.* 0 Gelid±ella trinitatensis Taylor * 0 Herposiphonia seeunda (C. Agardh) Ambronn 2 ± 2 Jania sp.* 0 Laurencia sp.* 0 Polysiphonia atlantica Kapraun and Norris < 1 Polysiphonia sp. 1 + 1 Spermothamnion sp. 0 Taenioma nanum (Kfitzing) Papenfuss < 1 Wurdemania miniata (Draparnaud)* 0
Feldmann and Hamel
0 0 < 1 < 1 0 0
6_+3 2 ± 1 0 < 1 0 0 < 1 0 0 < 1 0 0
2 ± 3 < 1 < l 6 ± 8 < 1 < l 3 ± 3 < 1 < 1
< 1 < 1 < 1 < 1 0 0
7 ± 8 8 ± 5 2 5 ± 7
0 0 3_+4 0 0 12±17 0 0 < 1
11_+ 9 0 < 1 4 ± 5 49±25 11±13
0 0 4 ± 4 0 0 < 1 2 ± 2 0 0
< 1 0 0 < 1 0 0 < 1 0 0
4 ± 9 0 5±11 3 ± 3 0 < 1
0 0 < 1 0 0 < 1 3 ± 2 1 2 ± 7 3 ± 4 0 < 1 20±18
< 1 2 ± 3 2 ± 4 1 0 ± 8 2 ± 3 0
< 1 < 1 2 ± 5 < 1 5 ± 1 5 ± 8
0 0 < 1 1 ± 1 < 1 < 1
< 1 0 0 6 ± 5 < 1 < 1 0 0 1 ± 2
< 1 0 < 1
2 ± 4 < 1 0 < 1 < 1 1 ± 2 < 1 0 0
0 0 0 0 0 < 1
< 1 1 ± 1 < 1 26±16 19±27 < 1

R.C. Carpenter: Diadema mass-mortality effects/population dynamics and algae 73
1.4
1.2 ̧
1.0
0.8
0.6'
< 0.4
=.,~- 0.2
~ , 0.0'. = E 4 -~ 1.4]
.~_ 1.0
Z
; . ; . ; . , . , . ; . ; . ; . , . ,
1 . 4 1 1.2t
1.01
o.81
o.61
0.4t
0.2t
0.01, . , . . . . . . . . . . . . . , . ,
e) Forereef(Srn)
0.6
0.4
Oo t . , ~ , - , - , , , , , . , , , , , , ,
c l l a _
1 . 4 4 d) Forereef (10 m)
1.0
0.8
0.6
0.4
0.2 " 4
0 . 0 i , . , . , . , . , . , . , . , . , . ,
N a u. _~
Fig. 5. Mean (_+SD) net primary productivity per unit area of algal communities over time in each of the four reef zones before and after mass mortality. Arrow marks approximate time of mass mor- tality in St. Croix. Filled data points represent data from caged treatment where grazing by Diadema antillarurn was reestablished
>
:,.2=
o_~
30 t a) Backreef/Reefcrest }
~-=" 30 t
m
Z
c) Forereef (5 m )
o u_ ~ N o ~ o
30
20
10
0
30
20
10
, . , . , . . . , . , . . , , . , . ,
d) Forereef (10 m)
, . , . , . , . , . , . , . , . , . ,
D u _
Fig. 6. Mean (+ SD) net biomass-specific primary productivity of algal communities in each of the four reef zones before and after mass mortality. Symbols as in Fig. 5
D. antillarum grazing (Carpenter 1986). Twenty-five months following the mass mortality, the algal community was dom- inated by macroalgal species (relative abundance of all macroalgal species =47%). Macroalgal taxa included spe- cies in the phaeophyte order Dictyotales, the articulated corallines Amphiroa spp. and Jania sp., and other larger rh0dophytes such as Laurencia sp. and species in the Geli- diaceae. Both chlorophyte and rhodophyte crustose algae decreased between 4 and 25 mo after the die-off.
Algal community primary-productivity
Rates of algal community primary productivity were affect- ed significantly by the mass mortality of Diadema antillarum (Figs. 5 and 6). Rates of primary productivity per unit area in the backreef/reefcrest treatment were 37% lower (p<0.01, one-way ANOVA, Scheff6's multiple compari- sons) than pre-mortality levels 5 d after the die-off (Carpen- ter 1988). Primary productivity rates per unit area increased following the mass mortality as algal biomass increased sig- nificantly (Fig. 2 a). Rates of primary producitivity per unit area in the treatment with reestablished grazing by D. antil- larum were lower than in the non-grazed treatments until March 1986, when the rates were comparable (Fig. 5a). However, the amount of algal biomass in the treatment grazed by D. antillarum was only 30% of that in the un- grazed treatments.
Primary productivity per unit area also decreased dra- matically at the forereef 2 m sites (p<0.001, one-way ANOVA, Scheff6's multiple comparisons). Although pro-
ductivity rates increased within 2 mo, unlike the backreef/ reefcrest sites, they did not return to premortality levels. There were no significant decreases in primary productivity per unit area at the deeper 5 and 10 m sites.
Rates of primary productivity per unit algal biomass (chlorophyll a) decreased significantly at all sites except the forereef 10 m sites (p < 0.001 for backreef/reefcrest, p < 0.001 for forereef 2 m, p < 0.01 for forereef 5 m; one-way ANOVA, Scheff6's multiple comparisons). Productivity rates per unit algal biomass did not increase significantly after the initial decrease in February 1984, except in the backreef/reefcrest caged treatment where grazing by Diadema antillarum oc- curred. The magnitude of the decrease in biomass-specific productivity rates that immediately followed the cessation of grazing by D. antillarum was positively and significantly correlated with the previous population densities of D. antil- larum (r s = 1.0, p < 0.01, Spearman's coefficient of rank cor- relation).
D i s c u s s i o n
Effects of mass mortality on population dynamics of Diadema antiIIarum
The effects of the mass mortality on the population dynam- ics ofDiadema antillarum in St. Croix have been significant. Decreases in population densities from 95 to 99% are simi- lar to those reported from Jamaica (Hughes et al. 1985), Curagao (Bak et al. 1984, Hunte et al. 1986), and Panamfi (Lessios et al. 1984), and elsewhere in the Caribbean (Les-

74 R.C. Carpenter: Diadema mass-mortality effects/population dynamics and algae
sios et al. 1984). Consistent with data from other sites, there is no evidence that the mass mortality affected size classes of sea urchins disproportionately, or that sea urchins in partic- ular habitats were more or less susceptible to the causative agent (Lessions 1988a). The result of this relatively indis- criminate mortality was to proportionately reduce the popu- lation density of D. antillarum of most sizes across all habi- tats.
Increases in the population densities of Diadema antil- larum following the mass mortality have been very limited. Immediately following the initial mortality event, the distri- bution of sea urchins shifted as individuals from deeper habitats migrated into the shallow forereef (R. Carpenter personal observations). Likewise, the appearance of addi- tional individuals within two days of the die-off suggests that some portion of the population was living within the interstices of the reef framework. Intraspecific competition for scarce algal resources under conditions of high popula- tion density may have resulted in part of the population being displaced to sub-optimal habitats within the reef struc- ture, where they probably grazed upright and encrusting invertebrates as well as crustose coralline algae (Jackson and Winston 1982, Jackson and Kaufman 1987). After much of the population was decimated, these individuals emerged to graze on the upper reef surfaces.
A second incidence of mortality has also been reported to have occurred in Panamfi in November 1985 (Lessios 1988 b). The mortality rate was much lower during this sec- ond event (ca. 50% in St. Croix and < 1% in Panam/t) than during the initial die-off. This may have resulted from a degree of immunity exhibited by survivors of the first mass mortality or from reduced virulence of the causative agent.
Recruitment of Diadema antillarum larvae immediately following the mass mortality was low in the habitats studied. In Curaqao and Panamfi there was a peak in juvenile recruit- ment shortly after the mortality event (Bak 1985, Lessios 1988 b). There is no evidence that such an influx of larvae settled on the study reef in St. Croix. The mass mortality occurred in February 1984 in St. Croix, nearly 14 mo after it appeared in Panam/t and 4 mo after it occurred in Cura- 9ao. The source of larvae for St. Croix populations of D. an- tillarum is not known, however, a reasonable explanation for the absence of a post-mortality peak in recruitment is the timing of the mortality relative to the time of spawning peaks of the parental population(s). If the parental popula- tion(s) died prior to the seasonal spawning peaks (April- May, August, October-November; Lewis 1966, Lessios 1988b), no larvae would be available for settlement.
For two years after the mass mortality, recruitment of Diadema antillarum larvae to the study sites has been low. This is correlated with a paucity of larvae in the plankton (Carpenter unpublished data) and suggests that inadequate larval supply rather than the absence or rarity of con- specifics to provide settlement cues is the reason for the poor recruitment at these reef sites (Lessios 1988b). The higher rate of recruitment of juveniles at the Grass Point site sug- gests that larvae may be concentrated and/or entrained in an eddy that forms behind the point. Grass Point is perpendic-
ular to the prevailing current and current-flow patterns indi- cate that planktonic organisms may be retained behind the point. Similar processes on a larger spatial scale have been proposed to explain the relatively high rates of recruitment of D. antillarum in Barbados (Hunte and Younglao 1988).
The general pattern of recruitment across the study sites is in agreement with patterns seen elsewhere in the Carib- bean, as reviewed by Lessios (1988 a). Although recruitment rates to reef habitats in Barbados are high (Hunte and Younglao 1988), recruitment rates in Jamaica, St. John, St. Croix, and Panamfi are low (Lessios 1988b, Levitan 1988, Hughes 1989). Rearing experiments with larvae of Diadema antillarum indicate that the feeding pluteus stage is viable for at least 52 d (Carpenter in preparation). Based on this larval longevity and considering current directions and velocities, as specific sites such as Barbados become repopu- lated, larvae from these reestablished populations should disperse quickly to habitats throughout the Caribbean. It is unclear whether there is a threshold parental population size that will lead to the production of sufficient larvae for re- population (Levitan 1988), or if larval settlement is depen- dent on the density of conspecific adults.
Ogden et al. (1973) and Hay (t984) have speculated that abnormally high population densities of Diadema antillarum in some areas of the Caribbean were the result of overfishing and the removal of sea urchin predators and competitors. This hypothesis assumes that sea urchin populations were at equilibrium. The mass mortality of D. antillarum and the subsequent slow and patchy recovery of populations at most sites suggests an alternative hypothesis about the underlying factors affecting the population densities of this species throughout the Caribbean prior to the mortality event. Al- though overfishing is likely to be important to some degree on a local scale in determining sea urchin abundances, anec- dotal evidence of high population densities of D. antillarum in the Caribbean in the early 1900's (Clark 1933) and the lack of correlation between sea urchin population-sizes and human population densities throughout the Caribbean evi- dent in the data published by Bauer (1980), suggest that other, more general processes, are responsible for the overall distribution and abundances of this species prior to the mass mortality. Although there is no evidence that a mass mortal- ity of D. antillarum has occurred previously, this natural experiment indicates that substantial variation in recruit- ment exists between sites on both local and regional scales (Bak 1985, Hunte and Younglao 1988). Similar spatial and temporal variation in the recruitment of D. antillarum was also demonstrated by Bak (1985) prior to the mass mortali- ty. Differential recruitment of larvae into sea urchin popula- tions that are not at equilibrium may be an equally plausible hypothesis to explain the variation in population densities of D. antillarum prior to the mass mortality.
Effects of sea-urchin mass mortality on algal biomass and community structure
The removal of Diadema antillarum from reef habitats in St. Croix resulted in significant increases in algal biomass in all

R.C. Carpenter: Diaderna mass-mortality effects/population dynamics and algae 75
reef zones despite increased grazing rates (Carpenter 1988) and increased population sizes of herbivorous fishes (Car- penter 1990). The magnitude of the increases is correlated with the previous population densities of sea urchins within each zone; as a result, algal biomass has increased much more in shallow backreef and forereef habitats than in deep- er forereef areas. Although most previous studies have not reported algal biomass values both before and after the sea urchin mass mortality, several studies have reported increas- es in the percent cover of algae following the mortality event. Increases in algal percent cover of 42% (Liddell and Ohlhorst 1986) and 93% (Hughes et al. 1987) are reported from Jamaica, and an increase of 30% in algal cover was reported in Curagao (de Ruyter van Steveninck and Bak 1986) following the mass mortality. These data support the hypothesis that grazing by D. antillarum is the primary de- terminant of algal distribution and biomass on many shal- low reefs throughout the Caribbean (Sammarco et al. 1974, Carpenter 1986, Foster 1987). Unfortunately, there are no published data on the effects of the sea urchin die-off on algal communities from sites where the abundances of her- bivorous fishes are reported to be high, such as Belize (Hay 1984, Lewis and Wainwright 1985) or the San Blas Islands in Panamfi (Lessios 1988a). However, data from one sea urchin-removal experiment conducted in the San Blas Is- lands prior to the mass mortality suggest that changes in algal biomass similar to those reported here would be ex- pected (Foster 1987).
Accompanying the increases in the biomass of the algal community were significant shifts in algal community struc- ture that are qualitatively similar to changes documented previously when sea urchins were removed experimentally (Sammarco et al. 1974, Sammarco 1982). Algal communi- ties grazed by Diaderna antillarum prior to the mass mortal- ity were dominated by unicells and simple filamentous algal species (Adey and Steneck 1985, Carpenter 1985a, b, 1986, Hackney et al. 1989). Although some seasonal variation in species composition normally occurs, species shifts are with- in the algal turf functional group (sensu Littler and Littler 1980 and Steneck and Watling 1982). The dramatic decreas- es in abundance of algal turfs and crustose algae and the increase in percent cover of macroalgal species support the hypothesis that grazing by D. antillarum controls the abun- dance of macroalgal species, most probably by grazing them indiscriminately when the plants are small (Carpenter 1986). Macroalgae only become established where they have a spa- tial or temporal refuge from intense herbivory (Lubchenco and Gaines 1981). The demonstrated control of algal com- munity structure, particularly the relative abundance of macroalgae, by sea urchin grazing contrasts with some of the recent literature that suggests that the relative abun- dances of reef algae are determined by the avoidance or preference of algal species by mainly herbivorous fishes based on algal defensive (usually chemical) characteristics (Littler et al. 1983, Paul and Hay 1986, Targett et al. 1986, Hay et al. 1987, 1988, Morrison 1988). The data presented here and obtained from experiments conducted at several reefs in the Caribbean and Pacific suggest instead that in-
tense herbivory limits the types of algal species that can persist in shallow-reef habitats to those species either pos- sessing fast growth rates (i.e., turf species) or those that exhibit resistance (i.e., crusts) to herbivory (Scott and Russ 1987, Foster 1987, Hackney et al. 1989). I f herbivory is re- duced, macroalgal species can become established and in- crease in abundance by outcompeting turf and crust species. Macroalgae have also overgrown corals and other sessile benthic invertebrates following the mass mortality (Hughes et al. 1987, Hughes 1989, Lessios 1988 a). As a result of the die-off of D. antillarum, reef-community structure has shift- ed from a community dominated by low-biomass algal turfs and corals to one dominated primarily by macroalgae, de- spite increased grazing by herbivorous fishes and increases in their local population size (Carpenter 1990). This is prob- ably the result of differences in the intensity and frequency of herbivore-induced disturbance created by most herbivo- rous fishes compared to D. antillarum (Carpenter 1986).
Effects of sea-urchin mass mortality on algal community primary-productivity
The immediate decreases in rates of biomass-specific prima- ry productivity following the mass mortality support con- clusions drawn from previous studies on the effects of graz- ing by Diadema antillarum on the metabolism of algal turf species (Carpenter 1981, 1986, 1988). The decrease in the rate of primary productivity was greatest in shallow back- reef and forereef habitats and more moderate in mid- and deep-forereef sites, correlated both with the previous popu- lation densities of D. antillarurn and the amount of in situ light. The recent demonstration that algal turfs are nitrogen- limited (Williams and Carpenter 1988) indicates that ammo- nium excretions from sea urchins may be a source of recy- cled nitrogen for algal turf species, allowing them to main- tain elevated rates of photosynthesis. Such an effect should be most evident in shallow habitats where algal photosyn- thesis is light-saturated for much of the day (Adey and Steneck 1985, Carpenter 1985b, Klumpp et al. 1987).
Overall, the mass mortality has resulted in an increase in reef algal primary-productivity per unit area in the shallow- est reef zones in St. Croix as algal biomass has increased by 300 to 440%. However, much of this biomass is comprised of species such as Turbinaria turbinata, Sargassum polycera- tiurn, Dictyopteris delicatula, and Laurencia obtusa; all spe- cies that are not readily consumed by most herbivores (Litt- ler et al. 1983). The result is that the reef algal community is more productive per unit area, but that much of the organic matter produced is not utilized by reef herbivores, leading to increased algal detritus in backreef and lagoonal areas as macroalgae die or are dislodged by storm waves and are exported off the reef (Carpenter unpublished data). This is a fundamental change in the functioning of the reef com- munity, representing a transition from a community previ- ously dominated by a grazing-based food web with 60 to 97% of the algal biomass removed by herbivores (Hatcher 1981, Carpenter 1986), to a community that exports a signif- icant amount of organic matter to adjacent communities.

76 R.C. Carpenter: Diadema mass-mortality effects/population dynamics and algae
Acknowledgement. I would like to thank E. Telemaque and L. Robinson who assisted in the field, Dr. J. Ogden and the staff of the West Indies Laboratory for facilitating the research, and especially H. Carpenter who assisted with every phase of the project. Helpful criticism on an earlier draft was kindly provided by Dr. S. L. Williams and two anonymous reviewers. This research was funded by the University of Puerto Rico Sea Grant Program (UPR PD-21, UPR R/ES-40-2). This is Contribution No. 186 of the West Indies Laboratory.
Literature cited
Adey, W. H. (1978). Coral reef morphogenesis: a multidimensional model. Science, N.Y. 202:831-837
Adey, W. H., Steneck, R. S. (1985). Highly productive eastern Caribbean reefs: synergistic effects of biological, physical and geological factors. In: Reaka M. L. (ed.) The ecology of coral reefs. National Oceanic and Atmospheric Administration, Rockville Maryland p. 163-187
Bak, R. P. M. (1985). Recruitment patterns and mass mortalities in the sea urchin Diadema antillarum. Proc. 5th int. coral Reef Congress, Tahiti 5 :267-272 [Gabri6, C., et al. (eds.) Antenne Museum - EPHE, Moorea, French Polynesia]
Bak, R. P. M., Carpay, M. J. E., de Ruyter van Steveninck, E. D. (1984). Densities of the sea urchin Diadema antillarum before and after mass mortalities on the coral reefs of Curacao. Mar. Ecol. Prog. Set. 17:105-108
Bauer, J. C. (1980). Observations on geographical variations in population density of the echinoid Diadema antillarum within the western North Atlantic. Bull. mar. Sci. 30:509-515
Breen, P. A., Mann, K. H. (1976). Changing lobster abundance and the destruction of kelp beds by sea urchins. Mar. Biol. 34:137 142
Carpenter, R. C. (1981). Grazing by Diadema antillarum (Philippi) [sic] and its effects on the benthic algal community. J. mar. Res. 39:749-765
Carpenter, R. C. (1985 a). Sea urchin mass mortality: effects on reef algal abundance, species composition, and metabolism and oth- er reef herbivores. Proc. 5th int. coral Reef Congr. 4 : 5 3 - 6 0 [Gabri6, C., et al. (eds.) Antenne Museum - EPHE, Moorea, French Polynesia]
Carpenter, R. C. (1985b). Relationships between primary produc- tion and irradiance in coral reef algal communities. Limnol. Oceanogr. 30:784-793
Carpenter, R. C. (1986). Partitioning herbivory and its effects on coral reef algal communities. Ecol. Monogr. 56:345-363
Carpenter, R. C. (1988). Mass mortality of a Caribbean echinoid: immediate effects on community metabolism and other herbi- vores. Proc. natn. Acad. Sci. U.S.A. 85:511-514
Carpenter, R. C. (1990). Mass mortality of Diadema antillarum. II. Effects on population densities and grazing intensity of parrot- fishes and surgeonfishes. Mar. Biol. 104:79-86
Chapman, A. R. O. (1981). Stability of sea urchin dominated barren grounds following destructive grazing of kelp in St. Margaret's Bay, Eastern Canada. Mar. Biol. 62:307-311
Clark, H. L. (1933). A handbook of the littoral echinoderms of Porto Rico and other West Indian islands. Scientific Survey of Porto Rico and the Virgin Islands. Vol. XVI. New York Acade- my of Sciences, New York
Dean, T. A., Schroeter, S. C., Dixon, J. D. (1984). Effects of grazing by two species of sea urchins (Strongylocentrotus franciseanus and Lytechinus anamesus) on recruitment and survival of two species of kelp (Macrocystis pyrifera and Pterygophora californi- ca). Mar. Biol. 78:301-313
Duggins, D. O. (1980). Kelp beds and sea otters: an experimental approach. Ecology 61:447-453
Estes, J. A., Palmisano, J. F. (1974). Sea otters: their role in struc- turing nearshore communities. Science, N.Y. 18:1058-1060
Estes, J. A., Smith, N. S., Palmisano, J. F. (1978). Sea otter preda- tion and community organization in the western Aleutian Is- lands, Alaska. Ecology 59:822-833
Foster, S. A. (1987). The relative impacts of grazing by Caribbean coral reef fishes and Diadema: effects of habitat and surge. J. exp. mar. Biol. Ecol. 105:1-20
Gladfelter, W B. (1982). White-band disease in Acropora palmata: implications for the structure and growth of shallow reefs. Bull. mar. Sci. 32:639-643
Hackney, J. M., Carpenter, R. C., Adey, W. H. (1989). Character- istic adaptations to grazing among algal turfs on a Caribbean coral reef. Phycologia 28:109-119
Hatcher, B. G. (1981). The interaction between grazing organisms and the epilithic algal community of a coral reef: a quantitative assessment. Proc. 4th int. coral Reef Syrup. 2 :515- 524 [Gomez, E. D., et al. (eds.) Marine Sciences Center, University of the Philippines, Quezon City]
Hay, M. E. (1984). Patterns of fish and urchin grazing on Caribbean coral reefs: are previous results typical? Ecology 65:446-454
Hay, M. E., Fenical, W., Gustafson, K. (1987). Chemical defense against diverse coral-reef herbivores. Ecology 68:1581-1591
Hay, M. E., Paul, V. J., Lewis, S. M., Gustafson, K., Tucker, J., Trindell, R. (1988). Can tropical seaweeds reduce herbivory by growing at night? Diel patterns of growth, nitrogen content, herbivory, and chemical versus morphological defenses. Oecolo- gia 75:233-245
Hughes, T. P. (1989). Community structure and diversity of coral reefs: the role of history. Ecology 70:275-279
Hughes, T. P., Keller, B. D., Jackson, J. B. C., Boyle, M. J. (1985). Mass mortality of the echinoid Diadema antillarum Philippi in Jamaica. Bull. mar. Sci. 36:377-384
Hughes, T. P., Reed, D. C., Boyle, M. J. (1987). Herbivory on coral reefs: community structure following mass mortalities of sea urchins. J. exp. mar. Biol. Ecol. 113:39-59
Hunte, W., Cote, I., Tomascik, T. (1986). On the dynamics of the mass mortality of Diadema antillarum in Barbados. Coral Reefs 4:135-139
Hunte, W., Younglao0 D. (1988). Recruitment and population re- covery in the black sea urchin Diadema a~tillarum (Echinoder- mata: Echinoidea) in Barbados. Mar. Ecol. Prog. Ser. 45: 109- 119
Jackson, J. B. C., Kaufman, K. W. (1987). Diadema antillarum was not a keystone predator in cryptic reef environments. Science, N.Y. 35:687-689
Jackson, J. B. C., Winston, J. E. (1982). Ecology of cryptic reef communities. I. Distribution and abundance of major groups of encrusting organisms. J. exp. mar. Biol. Ecol. 57:135-147
Jeffrey, S. W., Humphrey, G. F. (1975). New spectrophotometric equations for determining chlorophylls a, b, cl, e2 in higher plants, algae, and natural phytoplankton. Biochem. Physiol. Pfl. 167:191 194
Johnson, C. R., Mann, K. H. (1988). Diversity, patterns of adapta- tion, and stability of Nova Scotia kelp beds. Ecol. Monogr. 58: 129-154
Kapraun, D. F., Lemus, A. J., Bula-Meyer, G. (1983). Genus Polysiphonia (Rhodophyta, Ceramiales) in the tropical Western Atlantic. I. Colombia and Venezuela. Bull. mar. Sci. 33:881-898
Klumpp, D. W., McKinnon, D., Daniel, P. (1987). Damselfish territories: zones of high productivity on coral reefs. Mar. Ecol. Prog. Ser. 40:41-51
Lawrence, J. M. (1975). On the relationships between marine plants and sea urchins. Oceanogr. mar. Biol. A. Rev. 13:213-286
Leighton, D. L. (1971). Grazing activities of benthic invertebrates in southern California kelp beds. Nova Hedwigia 32: 421- 453
Lessios, H. A. (1988 a). Mass mortality ofDiadema antillarum in the Caribbean: what have we learned? A. Rev. Ecol. Syst. 19: 371- 393
Lessios, H. A. (1988 b). Population dynamics of Diadema antillarum (Echinodermata: Echinoidea) following mass mortality in Pana- mh. Mar. Biol. 99:515 526

R.C. Carpenter: Diadema mass-mortality effects/population dynamics and algae 77
Lessios, H. A., Robertson, D. R., Cubit, J. D. (1984). Spread of Diadema mass mortality through the Caribbean. Science, N.Y. 226:335 337
Levitan, D. R. (1988). Asynchronous spawning and aggregative behavior in the sea urchin Diadema antillarum (Philippi) [sic]. In: Burke, R. D., Mladenov, P. V., Lambert, P., Parsley R. L. (eds.) Echinoderm biology. Proceedings of the 6th International Echinoderm Conference. A. A. Balkema, Rotterdam, p. 181- 186
Lewis, J. B. (1966). Growth and breeding in the tropical echinoid Diadema antillarum Philippi. Bull. mar. Sci. 16:151-158
Lewis, S. M., Wainwright, P. C. (1985). Herbivore abundances and grazing intensity on a Caribbean coral reef. J. exp. mar. Biol. Ecol. 87:215-228
Liddell, W. D., Ohlhorst, S. L. (1986). Changes in benthic commu- nity composition following the mass mortality of Diadema at Jamaica. J. exp. mar. Biol. Ecol. 95:271-278
Littler, M. M., Littler, D. S. (1980). The evolution of thallus form and survival strategies in benthic marine macroalgae: field and laboratory tests of a functional form hypothesis. Am. Nat. 116: 25 -44
Littler, M. M., Littler, D. S. (1985). Nondestructive sampling. In: Littler, M. M., Littler, D. S. (eds.) Handbook of phycological methods: macroalgae. Cambridge University Press, Cambridge, p. 161-175
Littler, M. M., Taylor, P. R., Littler, D. S. (1983). Algal resistance to herbivory on a Caribbean coral reef. Coral Reefs 2:111 118
Lubchenco, J. L., Gaines, S. D. (1981). A unified approach to marine plant-herbivore interactions. I. Populations and commu- nities. A. Rev. Ecol. Syst. 12:405-437
Morrison, D. (1988). Comparing fish and urchin grazing in shallow and deeper coral reef algal communities. Ecology 69:1367-1382
Norris, J. N., Bucher, K. E. (1982). Marine algae and seagrasses from Carrie Bow Cay, Belize. In: Rutzler, K., Macintyre, I. G. (eds.) The Atlantic Barrier Reef Ecosystem at Carrie Bow Cay, Belize. Smithsonian Institution Press, Washington, D.C., p. 167 238
Ogden, J. O. (1977). Carbonate-sediment production by parrotfish and sea urchins on Caribbean reefs. Stud. Geol. Am. Ass. Pet- rol. Geol., Tulsa 4:281-288
Ogden, J. O., Brown, R. A., Salesky, N. (1973). Grazing by the echinoid Diadema antillarum Philippi: formation of halos around West Indian patch reefs. Science, N.Y. 182:715-717
Paine, R. T., Vadas, R. L. (1969). The effects of grazing by sea urchins Strongylocentrotus spp. on benthic algal populations. Limnol. Oceanogr. 14:710 719
Paul, V. J., Hay, M. E. (1986). Seaweed susceptibility to herbivory: chemical and morphological correlates. Mar. Ecol. Prog. Set. 33:255 264
Pearse, J. S., Hines, A. H. (1979). Expansion of a Central California kelp forest following the mass mortality of sea urchins. Mar. Biol. 51:83-91
Ruyter van Steveninck, E. D. de, Bak, R. P. M. (1986). Changes in abundance of coral-reef bottom components related to mass mortality of the sea urchin Diadema antillarum. Mar. Ecol. Prog. Set. 34:87-94
Sammarco, P. W. (1980). Diadema and its relationship to coral spat mortality: grazing, competition and biological disturbance. J. exp. mar. Biol. Ecol. 45:245-272
Sammarco, P. W. (1982). Effects of grazing by Diadema antillarum Philippi (Echinodermata: Echinoidea) on algal diversity and community structure. J. exp. mar. Biol. Ecol. 65:83-105
Sammarco, P. W., Levinton, J. S., Ogden, J. O. (1974). Grazing and control of community structure by Diadema antillarum Philippi (Echinodermata: Echinoidea): a preliminary study. J. mar. Res. 32:47-53
Scheibling, R. (1986). Increased macroalgat abundance following mass mortalities of sea urchins (Strongylocentrotus droebachien- sis) along the Atlantic coast of Nova Scotia. Oecologia 68: 186- 198
Scott, F. J., Russ, G. R. (1987). Effects of grazing on species com- position of the epilithic algal community on coral reefs of the Great Barrier Reef. Mar. Ecol. Prog. Ser. 39:293-304
Steneck, R. S. (1983). Quantifying herbivory on coral reefs: just scratching the surface and still biting off more than we can chew. In: Reaka, M. L. (ed.) The ecology of deep and shallow coral reefs. National Oceanic and Atmospheric Administration, Rockville, Maryland, p. 103-111
Steneck, R. S., Watling, L. (1982). Feeding capabilities and limita- tion of herbivorous molluscs: a functional group approach. Mar. Biol. 68:299 319
Targett, N. M., Targett, T. E., Vrolijk, N. H., Ogden, J. C. (1986). Effect of macrophyte secondary metabolites on feeding prefer- ences of the herbivorous parrotfish Sparisoma radians. Mar. Biol. 92:141-148
Taylor, W. R. (1960). Marine algae of the eastern tropical and subtropical coasts of the Americas. University of Michigan Press, Ann Arbor
Vadas, R. L., Elner, R. W., Garwood, P. E., Babb, I. G. (1986). Experimental evaluation of aggregation behavior in the sea urchin Strongylocentrotus droebachiensis. A reinterpretation. Mar. Biol. 90:433-448
Wharton, W. G., Mann, K. H. (1981). Relationship between de- structive grazing by the sea urchin, Strongylocentrotus droe- bachiensis, and the abundance of American lobster, Homarus americanus, on the Atlantic coast of Nova Scotia. Can. J. Fish. aquat. Sciences 38: 1339-1349
Williams, S. L., Carpenter, R. C. (1988). Nutrient-limited primary productivity of coral reef algal turfs: potential contribution of ammonium excreted by Diadema antillarum. Mar. Ecol. Prog. Set. 47:145-152
Date of final manuscript acceptance: September 28, 1989. Communicated by M. G. Hadfield, Honolulu


















