
M b 1,5, a a. k –b 2, a a. G 3 a M. d 1 Z s. aMr4
11
117 Herpetological Conservation and Biology 16(1):117–127. Submitted: 10 December 2020; Accepted: 1 March 2021; Published: 30 April 2021. Copyright © 2021. Mohammad A. Abu Baker All Rights Reserved. DIET AND FOOD NICHE RELATIONSHIPS OF LIZARD ASSEMBLAGES IN JORDAN MOHAMMAD A. ABU BAKER 1,5 , AHMAD A. KATBEH–BADER 2 , ANSAM A. GHLELAT 3 , AHMAD M. DISI 1 , AND ZUHAIR S. AMR 4 1 Department of Biological Sciences, The University of Jordan, Queen Rania Street, Amman 11942, Jordan 2 Department of Plant Protection, Faculty of Agriculture, The University of Jordan, Queen Rania Street, Amman 11942, Jordan 3 Conservation and Monitoring Center, The Royal Society for the Conservation of Nature, Baker Al Baw Street, Amman 11941, Jordan 4 Department of Biology, Jordan University of Science & Technology, P.O. Box 3030, Irbid 22110, Jordan 5 Corresponding author, email: [email protected] Abstract.—We studied diet and food niche relationships among sympatric lizards in two areas in Jordan using stomach contents. We found that 77 stomachs from six species in the eastern deserts and 45 stomachs from three species in the western highlands resulted in 2,478 and 1,935 prey items, respectively. Stomachs from the eastern desert contained 1–216 prey items with an average of 31.5 items (median = 16) and 1–329 prey items with an average of 43.0 (median = 13) from the western highlands. The lizards were primarily insectivorous, with ants and beetles making up the highest percentages. Additionally, we identified a snake (Eirenis rothii), a freshwater fish, and four smaller lizards in the prey remains. The results showed similar niche breadths and high food-niche overlap values, especially among the agamid lizards, suggesting that some opportunistic feeding habits are likely influenced by prey availability and similar foraging strategies. Based on similar niche breadths and high food- niche overlap values, we believe that interspecific competition is an important factor in organizing these lizard assemblages. Therefore, habitat segregation and different hunting strategies may likely occur among these species and allow for species coexistence in these areas. Key Words.—deserts; diet partitioning; niche breadth; niche overlap; squamates; stomach contents INTRODUCTION The organization of natural communities is a central question in field ecology (Strong et al. 1984). Food, space, and time are the principal factors that determine the trophic niche of a species and organize assemblages of sympatric species (Pianka 1969; Schoener 1974). Morphological characteristics mediate the pattern of resource partitioning and determine the role of microhabitat selection (Ortega et al. 1982). Diversity and resource partitioning in lizard assemblages has been studied in several deserts of the world (Pianka 1973, 1986). Many studies have dealt with the ecological structure of lizard communities (e.g., Pianka 1975; Pianka and Huey 1978; Pianka et al. 1979; Toft 1985; Shenbrot et al. 1991); however, the data for Palaearctic deserts are scarce, except for some localized studies from the eastern Mediterranean, the Arabian Peninsula, and North Africa (e.g., Arnold 1984; Shenbrot and Krasnov 1997; Attum et al. 2004; Rifai et al. 2005; Rouag et al. 2007). Lizards are one of the most diversified and successful groups of vertebrates inhabiting these hot deserts and contribute highly to the local and regional biodiversity (Pianka 1986; Roll et al. 2017). Dietary studies of reptiles are crucial for understanding life-history traits, niche relationships, competitive processes, predator–prey and trophic interactions, and even energy fluxes within ecosystems (Pianka 1986; Luiselli and Amori 2016). The diets of lizard species may reflect their foraging modes and preferred microhabitats (Schoener 1971; Pianka 1973). Most species are mainly sit-and-wait foragers that rely on visual scanning (MacArthur and Pianka 1966; Schoener 1971; Pianka 1973; Arnold 1984). This mode of passive foraging is common in members of the family Agamidae, many of which feed mostly on ground-dwelling insects (Herrel et al. 1998; Ibrahim and El-Naggar 2013). Agamids usually remain motionless, elevate their bodies, and raise their heads while scanning their surroundings until a prey is spotted. The open, flat deserts or rocky terrains they occupy provide an ideal setting for this foraging mode. Other lizards, such as lacertids (the fringe-toed lizards, Acanthodactylus spp., and desert lizards, Mesalina spp.) and skinks (e.g., the Orange-tailed Skink, Eumeces schneideri) are active hunters feeding on larger percentages of beetles, orthopterans, and other flying and plant-dwelling insects (Arnold 1984; Perry et al.
Transcript of M b 1,5, a a. k –b 2, a a. G 3 a M. d 1 Z s. aMr4

117
Herpetological Conservation and Biology 16(1):117–127.Submitted: 10 December 2020; Accepted: 1 March 2021; Published: 30 April 2021.
Copyright © 2021. Mohammad A. Abu BakerAll Rights Reserved.
Diet anD FooD niche Relationships oF
lizaRD assemblages in JoRDan
MohaMMad a. abu baker1,5, ahMad a. katbeh–bader2, ansaM a. Ghlelat3, ahMad M. disi1, and Zuhair s. aMr4
1Department of Biological Sciences, The University of Jordan, Queen Rania Street, Amman 11942, Jordan2Department of Plant Protection, Faculty of Agriculture, The University of Jordan, Queen Rania Street,
Amman 11942, Jordan3Conservation and Monitoring Center, The Royal Society for the Conservation of Nature, Baker Al Baw Street,
Amman 11941, Jordan 4Department of Biology, Jordan University of Science & Technology, P.O. Box 3030, Irbid 22110, Jordan
5Corresponding author, email: [email protected]
Abstract.—We studied diet and food niche relationships among sympatric lizards in two areas in Jordan using stomach contents. We found that 77 stomachs from six species in the eastern deserts and 45 stomachs from three species in the western highlands resulted in 2,478 and 1,935 prey items, respectively. Stomachs from the eastern desert contained 1–216 prey items with an average of 31.5 items (median = 16) and 1–329 prey items with an average of 43.0 (median = 13) from the western highlands. The lizards were primarily insectivorous, with ants and beetles making up the highest percentages. Additionally, we identified a snake (Eirenis rothii), a freshwater fish, and four smaller lizards in the prey remains. The results showed similar niche breadths and high food-niche overlap values, especially among the agamid lizards, suggesting that some opportunistic feeding habits are likely influenced by prey availability and similar foraging strategies. Based on similar niche breadths and high food-niche overlap values, we believe that interspecific competition is an important factor in organizing these lizard assemblages. Therefore, habitat segregation and different hunting strategies may likely occur among these species and allow for species coexistence in these areas.
Key Words.—deserts; diet partitioning; niche breadth; niche overlap; squamates; stomach contents
intRoDuction
The organization of natural communities is a central question in field ecology (Strong et al. 1984). Food, space, and time are the principal factors that determine the trophic niche of a species and organize assemblages of sympatric species (Pianka 1969; Schoener 1974). Morphological characteristics mediate the pattern of resource partitioning and determine the role of microhabitat selection (Ortega et al. 1982). Diversity and resource partitioning in lizard assemblages has been studied in several deserts of the world (Pianka 1973, 1986). Many studies have dealt with the ecological structure of lizard communities (e.g., Pianka 1975; Pianka and Huey 1978; Pianka et al. 1979; Toft 1985; Shenbrot et al. 1991); however, the data for Palaearctic deserts are scarce, except for some localized studies from the eastern Mediterranean, the Arabian Peninsula, and North Africa (e.g., Arnold 1984; Shenbrot and Krasnov 1997; Attum et al. 2004; Rifai et al. 2005; Rouag et al. 2007). Lizards are one of the most diversified and successful groups of vertebrates inhabiting these hot deserts and contribute highly to the local and regional biodiversity (Pianka 1986; Roll
et al. 2017). Dietary studies of reptiles are crucial for understanding life-history traits, niche relationships, competitive processes, predator–prey and trophic interactions, and even energy fluxes within ecosystems (Pianka 1986; Luiselli and Amori 2016). The diets of lizard species may reflect their foraging modes and preferred microhabitats (Schoener 1971; Pianka 1973). Most species are mainly sit-and-wait foragers that rely on visual scanning (MacArthur and Pianka 1966; Schoener 1971; Pianka 1973; Arnold 1984). This mode of passive foraging is common in members of the family Agamidae, many of which feed mostly on ground-dwelling insects (Herrel et al. 1998; Ibrahim and El-Naggar 2013). Agamids usually remain motionless, elevate their bodies, and raise their heads while scanning their surroundings until a prey is spotted. The open, flat deserts or rocky terrains they occupy provide an ideal setting for this foraging mode. Other lizards, such as lacertids (the fringe-toed lizards, Acanthodactylus spp., and desert lizards, Mesalina spp.) and skinks (e.g., the Orange-tailed Skink, Eumeces schneideri) are active hunters feeding on larger percentages of beetles, orthopterans, and other flying and plant-dwelling insects (Arnold 1984; Perry et al.

118
Abu Baker et al.—Diet of lizard assemblages in Jordan.
1990; Cooper 1999; Cooper et al. 2000). Large species of Acanthodactylus walk jerkily over the substrate, touching the ground regularly with their snout detecting scents with their tongue to find prey (Arnold 1984). The habitats that they occupy include extensive vegetation cover and provide considerable prey diversity (Arnold 1984). The lizard fauna of Jordan consists of 56 species and subspecies belonging to eight families. The families Lacertidae (represented by 17 species) and Agamidae (represented by 12 species) are the most numerous in terms of species diversity (Disi et al. 2001; Disi 2011). Members of these families occur in a variety of habitats, including sand dunes, rocky terrains, and flat deserts (Disi et al. 2001). The lizard fauna of Jordan is diverse and forms an important component of the biodiversity in arid zones; however, ecological aspects of lizard communities in the region remain little known. We examined the dietary and niche relationships in two lizard assemblages through a comparative analysis
of species diets. We aimed to determine the diet composition for each lizard and the importance of food partitioning in organizing these lizard assemblages.
mateRials anD methoDs
We collected diet data from 122 lizard specimens. We obtained specimens from the Herpetological Collections of the Biological Sciences Department, University of Jordan in Amman. The lizards were captured during systematic, country-wide surveys (Disi et al. 1999; 2001; Disi 2011; Modry et al. 2004). Six lizard species (Fig. 1) comprised two assemblages from the eastern desert and the western highlands of Jordan (Fig. 2). The eastern desert of Jordan lies within the Saharo-Arabian biogeographical region and covers over 70% of the total area of the country with elevations ranging between 100–800 m. The area is characterized by arid and semiarid open areas and wadi beds dominated by Hammada (Hammada salicornicum), Lavender Cotton
FiguRe 1. Six of the lizards included in the study in their natural habitats in Jordan. (A) Bosk’s Fringe-toed Lizard (Acanthodactylus boskianus). (B) Orange-tailed Skink (Eumeces schneideri). (C) Starred Agama (Stellagama stellio). (D) Pale Agama (Trapelus agnetae). (E) Persian Agama (Trapelus persicus). (F) Sinai Agama (Pseudotrapelus sinaitus). (Photographed by Mohammad Abu Baker).

119
Herpetological Conservation and Biology
(Achillea fragrantissima), White Wormwood (Artemisia sieberi) and milk vetch (Astragalus spp.). The substrate consists of pebble, gravel, hamada (flat, gravely desert), saline, and sandy soils. The western highlands lie within the Mediterranean and Irano-Turanian biogeographical regions. These mountain ranges consist of forested vegetation at the highest elevations (700–1,500 m) surrounded by a narrow strip of dry to semi-dry habitats (400–700 m). Rocky, forested vegetation (Phoenician Juniper, Juniperus phoenicea, Atlantic Pistachio, Pistacia atlantica, Aleppo Pine, Pinus halepensis, and oak, Quercus spp.) occurs at the core with terra rosa (red Mediterranean soil), sandy, and sandy-loamy soil types. The surrounding vegetation is dominated by the Jointed Anabasis (Anabasis articulata), Artemisia sieberi, Spiny Milk-Vetch (Astragulus spinosum), and White Broom (Retama raetam) with a thin layer of surface soil. For the eastern desert assemblage, we checked stomach contents from 16 Acanthodactylus spp., three Eumeces schneideri, 10 Sinai Agama (Pseudotrapelus sinaitus), 12 Starred Agama (Stellagama stellio), 28 Pale Agama (Trapelus agnetae), and eight Persian Agama (Trapelus persicus). For the western highlands assemblage, we checked seven Eumeces schneideri, 10 Pseudotrapelus sinaitus, and 28 Stellagama stellio. We included three species of Acanthodactylus (12 Bosk’s Fringe-toed Lizards, A. boskianus, two Giant Fringe-toed Lizards, A. grandis, and two Fringe-toed Sand Lizards, A. schmidti), all very similar in morphology and ecology, in a single group due to the low number of specimens (see Arnold 1984; Al-Johany and Spellerberg 1989; Perry et al. 1990; Disi et al. 2001). We analyzed 77 stomachs from the eastern desert and 45 stomachs from the western highlands assemblages.
We analyzed diet composition in accordance with standard techniques for reptiles (see Luiselli and Amori 2016). We removed, sorted, and examined stomach contents using a stereoscopic microscope. We identified prey items to the family level (see Appendix 1, 2) but we analyzed them in terms of order except for Formicidae (Table 1, Appendix 3). Most items were fragmented, so it was difficult to obtain volumetric measures. Thus, we represented diet composition for each species as abundance (n) and frequency of occurrence (%). Abundance is represented by the number of items in each prey category and the corresponding percentage (number of items in each prey category/total number of items × 100). We estimated frequency of occurrence by dividing the percentage of stomachs containing a prey category / total number of stomachs analyzed × 100. We used the percentage of stomachs containing a prey category as an estimate of prey selection and to control for variation in number of lizards examined. We calculated Trophic Niche Breadth using the Simpson Diversity Index (Simpson 1949), Bs:
We also used the standardized form of Levins (Levins 1968), BL:
In both equations, Pi is the relative representation of the ith prey category in the sample.To investigate the presence of nonrandom patterns in trophic niche overlap, we used the Niche Overlap Module of EcoSim (www.garyentsminger.com/ecosim/index.htm). The data consisted of a matrix in which each species is a row and each prey category is a column. Entries in the matrix represent the abundance of prey categories in each species and the percentage of stomachs containing a prey category in a second run. The matrix is reshuffled to produce random patterns that would be expected in the absence of competitive interactions.
We used the Niche Overlap Index, Ojk (Pianka 1973), at 1,000 iterations to calculate a separate index for each species pair within each lizard assemblage:
where 0jk is niche overlap between species j and species k, Pij is the proportion of prey item i used by species j, and Pik is the proportion of prey item i used by species k. The niche overlap module varies from 0 (no overlap)
FiguRe 2. Map of Jordan showing the two areas from which the lizards were collected.
120
Abu Baker et al.—Diet of lizard assemblages in Jordan.
to 1 (complete overlap). To test for differences in diet composition between lizard species, we used the Kruskal-Wallis test using the number of prey items consumed from every prey category. We used the Mann-Whitney U test to compare diets of lizard species that occurred in the two study areas.
Results
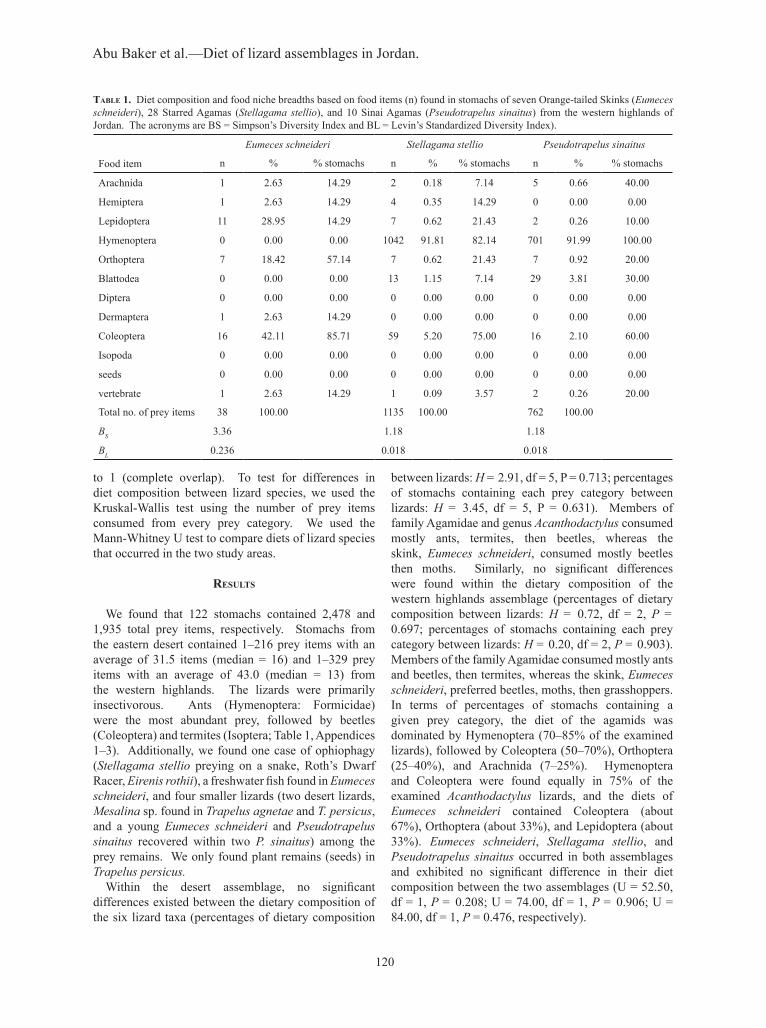
We found that 122 stomachs contained 2,478 and 1,935 total prey items, respectively. Stomachs from the eastern desert contained 1–216 prey items with an average of 31.5 items (median = 16) and 1–329 prey items with an average of 43.0 (median = 13) from the western highlands. The lizards were primarily insectivorous. Ants (Hymenoptera: Formicidae) were the most abundant prey, followed by beetles (Coleoptera) and termites (Isoptera; Table 1, Appendices 1–3). Additionally, we found one case of ophiophagy (Stellagama stellio preying on a snake, Roth’s Dwarf Racer, Eirenis rothii), a freshwater fish found in Eumeces schneideri, and four smaller lizards (two desert lizards, Mesalina sp. found in Trapelus agnetae and T. persicus, and a young Eumeces schneideri and Pseudotrapelus sinaitus recovered within two P. sinaitus) among the prey remains. We only found plant remains (seeds) in Trapelus persicus. Within the desert assemblage, no significant differences existed between the dietary composition of the six lizard taxa (percentages of dietary composition
between lizards: H = 2.91, df = 5, P = 0.713; percentages of stomachs containing each prey category between lizards: H = 3.45, df = 5, P = 0.631). Members of family Agamidae and genus Acanthodactylus consumed mostly ants, termites, then beetles, whereas the skink, Eumeces schneideri, consumed mostly beetles then moths. Similarly, no significant differences were found within the dietary composition of the western highlands assemblage (percentages of dietary composition between lizards: H = 0.72, df = 2, P = 0.697; percentages of stomachs containing each prey category between lizards: H = 0.20, df = 2, P = 0.903). Members of the family Agamidae consumed mostly ants and beetles, then termites, whereas the skink, Eumeces schneideri, preferred beetles, moths, then grasshoppers. In terms of percentages of stomachs containing a given prey category, the diet of the agamids was dominated by Hymenoptera (70–85% of the examined lizards), followed by Coleoptera (50–70%), Orthoptera (25–40%), and Arachnida (7–25%). Hymenoptera and Coleoptera were found equally in 75% of the examined Acanthodactylus lizards, and the diets of Eumeces schneideri contained Coleoptera (about 67%), Orthoptera (about 33%), and Lepidoptera (about 33%). Eumeces schneideri, Stellagama stellio, and Pseudotrapelus sinaitus occurred in both assemblages and exhibited no significant difference in their diet composition between the two assemblages (U = 52.50, df = 1, P = 0.208; U = 74.00, df = 1, P = 0.906; U = 84.00, df = 1, P = 0.476, respectively).
Eumeces schneideri Stellagama stellio Pseudotrapelus sinaitus
Food item n % % stomachs n % % stomachs n % % stomachs
Arachnida 1 2.63 14.29 2 0.18 7.14 5 0.66 40.00
Hemiptera 1 2.63 14.29 4 0.35 14.29 0 0.00 0.00
Lepidoptera 11 28.95 14.29 7 0.62 21.43 2 0.26 10.00
Hymenoptera 0 0.00 0.00 1042 91.81 82.14 701 91.99 100.00
Orthoptera 7 18.42 57.14 7 0.62 21.43 7 0.92 20.00
Blattodea 0 0.00 0.00 13 1.15 7.14 29 3.81 30.00
Diptera 0 0.00 0.00 0 0.00 0.00 0 0.00 0.00
Dermaptera 1 2.63 14.29 0 0.00 0.00 0 0.00 0.00
Coleoptera 16 42.11 85.71 59 5.20 75.00 16 2.10 60.00
Isopoda 0 0.00 0.00 0 0.00 0.00 0 0.00 0.00
seeds 0 0.00 0.00 0 0.00 0.00 0 0.00 0.00
vertebrate 1 2.63 14.29 1 0.09 3.57 2 0.26 20.00
Total no. of prey items 38 100.00 1135 100.00 762 100.00
BS 3.36 1.18 1.18
BL 0.236 0.018 0.018
table 1. Diet composition and food niche breadths based on food items (n) found in stomachs of seven Orange-tailed Skinks (Eumeces schneideri), 28 Starred Agamas (Stellagama stellio), and 10 Sinai Agamas (Pseudotrapelus sinaitus) from the western highlands of Jordan. The acronyms are BS = Simpson’s Diversity Index and BL = Levin’s Standardized Diversity Index).

121
Herpetological Conservation and Biology
The values of niche breadth were rather low for all species (Table 1, Appendix 3). The Bs values for the eastern desert assemblage ranged between 1.33 and 2.28 with species ranking from lowest to highest Bs as follows: Trapelus agnetae, Trapelus persicus, Eumeces schneideri, Stellagama stellio, Pseudotrapelus sinaitus, and Acanthodactylus sp., whereas in the western highlands assemblage, the values ranged between 1.18 and 3.36, and the species ranked as follows: Stellagama stellio, Pseudotrapelus sinaitus, and Eumeces schneideri. In this assemblage, Eumeces schneideri exhibited the most diverse diet. The estimated values of food niche overlap among lizard species within both assemblages were rather high, except for E. schneideri (Table 2). Acanthodactylus sp. also exhibited high niche overlap with the three agamids (Table 2). Among the agamids, niche overlap ranged between 0.88 and 0.981 in the eastern desert and was 0.999 between S. stellio and P. sinaitus in the western highlands. Conversely, the diet of E. schneideri was different from its coexisting species in both habitats, niche overlap values ranged between 0.003 and 0.102 in the eastern desert assemblage and 0.023 and 0.050 in the western highlands assemblage.
Discussion
The diet of the lizards included in our study were largely or entirely comprised of arthropods, with smaller insects (ants and termites) making up the highest percentages, especially for members of the family Agamidae. These prey items are small and provide little energy, yet they are common and easily obtained, which can reduce the energy costs of pursuit (Schoener 1971). Although ants were well represented in terms of abundance compared to other prey groups, ants were relatively less well represented in the stomachs of the examined lizards. Larger prey such as Coleoptera and Orthoptera were represented in up to 75% in the examined stomachs of agamids (Pseudotrapelus sinaitus, Stellagama stellio, Trapelus agnetae, T. persicus) and Acanthodactylus. Similar results were previously reported in Agama sp., Laudakia (Stellagama) sp., Acanthodactylus sp.
(Arnold 1984; Robson and Lambert 1980; Pérez-Mellado 1992; Ibrahim and El-Naggar 2013). The diet of A. boskianus has been described in desert populations, where the volumetric importance of Orthoptera exceeded that of Formicidae (Robson and Lambert 1980). Eumeces schneideri on the other hand relied on larger prey and had the most diverse diet, with Coleoptera, Lepidoptera, and Orthoptera on the menu. Stellagama stellio and Pseudotrapelus sinaitus are sympatric species of agamid lizards that share the same rocky habitats; however, the two species differ in the use of microhabitats. Pseudotrapelus sinaitus spends more time exposed on the rocks for thermoregulation and social communication (Norfolk et al. 2010). Stellagama stellio is a largest of the agamids in our study and is an omnivorous lizard, feeding on insects and other arthropods, gastropods, flowers, buds, and leaves. It also has been reported to enhance its diet with occasional vertebrate prey like mice, lizards, and there even is a record of ophiophagy on a small snake, the European Blind Snake (Xerotyphlops vermicularis; Karameta et al. 2015), as well as the European Bee-eater (Merops apiaster) eggs and House Sparrow (Passer domesticus) chicks (Kopan and Yom-Tov 1982; Lo Cascio et al. 2001).
In our study, we recovered remains of vertebrate prey items on six occasions, of which Stellagama stellio preying on the snake Eirenis rothii is noteworthy as a second record of ophiophagy (E. rothii is a common small colubrid that reaches up to 300 mm in length and weighs 2.4 g on average; Shai Meiri, pers. comm.). All three other agamids preyed on smaller lizards, including a case of cannibalism by Pseudotrapelus sinaitus. A freshwater fish was also found in an Eumeces schneideri stomach that came from Wadi Shuaeib, a wadi system with a stream, a dam, and agricultural farms. Energy-rich prey items, such as insects (primarily Coleoptera, Lepidoptera, and Orthoptera), gastropods, and arachnids, in addition to earthworms, leeches, crustaceans, and fish were similarly recovered from the Chinese Skink (Eumeces chinensis; Chen and Jiang 2006). Desert habitats are characterized by low and unpredictable productivity (Pianka 1986). The main insectivorous resources for lizards are ants and termites, with both prey groups found in clumped distributions in nests or along trails. It is common for Formicidae, Isoptera, and Coleoptera to be the main prey in desert ecosystems (Pianka 1986). Low values for diversity indices indicate limited variety of the taxonomic composition of the diet. This is likely due to the low resource availability in desert habitats. Other prey items (Orthoptera, Lepidoptera, Arachnida) may be taken by active foraging lizards. Lizards of the genus Acanthodactylus exhibit a foraging behavior of rapid runs from one bush to another, taking prey near bushes,
E. schneideri
S. stellio
P. sinaitus
T. agnetae
T. persicus
Acanthodactylus 0.211 0.839 0.987 0.962 0.931
E. schneideri 0.069 0.102 0.031 0.038
S. stellio 0.831 0.889 0.88
P. sinaitus 0.947 0.906
T. agnetae 0.981
table 2. Food niche overlap values among fringe-toed lizards (Acanthodactylus spp.), Orange-tailed Skinks (Eumeces schneideri), Starred Agamas (Stellagama stellio), Pale Agamas (Trapelus agnetae), Sinai Agamas (Pseudotrapelus sinaitus), and Persian Agamas (Trapelus persicus) from the lizard assemblage of the eastern desert of Jordan.

122
Abu Baker et al.—Diet of lizard assemblages in Jordan.
and then running again through open areas (Arnold 1984). The energetic costs of this foraging behavior would be high for these lizards; hence, more profitable prey are required for survival. The estimated values of food niche overlap among lizard species within both assemblages were rather high, except for E. schneideri. This suggests high overlap in the taxonomic composition of the diets in coexisting members of the family Agamidae in both habitats. The similar niche breadths and high food-niche overlap values (especially among the agamid lizards) suggest some opportunistic feeding habits by most species. These are likely influenced by prey availability and the foraging strategies for each species. We believe that interspecific competition is an important factor in organizing these lizard assemblages. We think that coexistence among species is maintained to a large degree by microhabitat segregation and the different foraging strategies employed by these species. Sympatric lizards are known to partition spatial resources in a wide variety of habitats and climatic conditions (Pianka 1966; Western 1974; Huey and Pianka 1977; Barbault and Maury 1981; Shenbrot and Krasnov 1997). We did not determine relative prey volume when calculating niche breadths because it tends to over-emphasize small prey, such as ants compared larger prey items such as beetles and orthopterans. This omission may have resulted in deficiencies in details when comparing the results between species, though. Unfortunately, this was unavoidable considering the ethical issue of sacrificing many individuals of reptile species that are in decline (Modry et al. 2004; Cox et al. 2012; Böhm et al. 2013). Future investigations that would be useful to determine spatial and temporal niche axes of lizard assemblages would be to study microhabitat partitioning, as well as comparisons of diel and seasonal activity patterns, prey availability, prey volumes, and home ranges among the lizard species.
Acknowledgments.—All material examined in this study were museum specimens that accumulated during systematic, country-wide surveys. A primary debt is owed for those who participated in the field surveys of the lizards of Jordan. We are also grateful to the staff at the Entomology Museum, Department of Plant Protection, Faculty of Agriculture, University of Jordan. Shai Meiri provided comments that improved the manuscript.
liteRatuRe citeD
Albergoni, A., I. Bride, C.T. Scialfa, M. Jocque, and S. Green. 2016. How useful are volunteers for visual biodiversity surveys? An evaluation of skill level and group size during a conservation expedition.
Biodiversity and Conservation 25:133–149.AI-Johany A.M.H., and I.F. Spellerberg. 1989. Home
range and vagility of the lizards Acanthodactylus schmidti and Acanthodactylus boskianus in Saudi Arabia. Journal of Arid Environments 16:79–86.
Arnold, E.N. 1984. Ecology of lowland lizards in the eastern United Arab Emirates. Journal of Zoology 204:329–354.
Attum, O., C. Covell, and P. Eason. 2004. The comparative diet of three Saharan sand dune skinksAfrican Journal of Herpetology 53:91–94.
Barbault, R., and M.E. Maury. 1981. Ecological organization of a Chihuahuan Desert lizard community. Oecologia 51:335–342.
Böhm, M., B. Collen, J.E.M. Baillie, P. Bowles, J. Chanson, N. Cox, G. Hammerson, M. Hoffmann, S.R. Livingstone, M. Ram, et al. 2013. The conservation status of the world’s reptiles. Biological Conservation 157:372–85.
Chen, X., and Y. Jiang. 2006. Diet of Chinese skink, Eumeces chinensis: is prey size important? Integrative Zoology 2:59–66.
Cooper, W.E., Jr. 1999. Supplementation of phylogenetically correct data by two-species comparison: support for correlated evolution of foraging mode and prey chemical discrimination in lizards extended by first intrageneric evidence. Oikos 87:97–104.
Cooper, W.E., Jr., A.M. Al-Johany, and L.J. Vitt. 2000. Responses to chemical cues from animal and plant foods by actively foraging insectivorous and omnivorous scincine lizards. Journal of Experimental Zoology 287:327–339.
Cox, N.A., D. Mallon, P. Bowles, J. Els, and M.F. Tognelli. 2012. The Conservation Status and Distribution of Reptiles of the Arabian Peninsula. International Union for Conservation of Nature, Gland, Switzerland, and Sharjah Environment and Protected Areas Authority, UAE.
Disi, A.M. 2011. Review of the lizard fauna of Jordan. Zoology in the Middle East 54:89–102.
Disi, A.M., D. Modry, F. Bunian, R.M. Al-Oran, and Z.S. Amr. 1999. Amphibians and reptiles of the Badia region of Jordan. Herpetozoa 12:135–146.
Disi, A.M., D. Modry, P. Necas, and L. Rifai. 2001. Amphibians and Reptiles of the Hashemite Kingdom of Jordan. An Atlas and Field Guide. Chimaira, Frankfurt am Main, Germany.
Herrel, A., J.P. Timmermans, and F. De Vree. 1998. Tongue flicking in agamid lizards: morphology, kinematics, and muscle activity patterns. Anatomical Record 252:102–116.
Huey, R.B., and E.R. Pianka. 1977. Patterns of niche overlap among broadly sympatric versus narrowly sympatric Kalahari lizards (Scincidae: Mabuya).

123
Herpetological Conservation and Biology
Ecology 58:119–128.Ibrahim, A.A., and M.H. El-Naggar. 2013. Diet and
reproductive biology of the Starred Agama, Laudakia stellio (Linnaeus, 1758) (Squamata: Agamidae), in the northern Sinai, Egypt. Zoology in the Middle East 59:136–143.
Karameta, E., V.V. Papadopoulos, and P. Pafilis. 2015. First record of ophiophagy in the Roughtail Rock Agama (Stellagama stellio): one of the most rare feeding behaviors among European lizards. Herpetology Notes 8:111–113.
Kopan, R., and Y. Yom–Tov. 1982. Agama stellio (Reptilia: Sauria) feeding on avian eggs and chicks. Israel Journal of Zoology 31:159–160.
Levins, R. 1968. Evolution in Changing Environments. Princeton University Press, Princeton, New Jersey, USA.
Lo Cascio, P., C. Corti, and L. Luiselli. 2001. Seasonal variations of the diet of Laudakia stellio (Agamidae) from Nisyros Island, Dodecanese (Greece). Herpetological Journal 11:33–35.
Luiselli, L., and G. Amori. 2016. Diet. Pp. 97–109 In Reptile Ecology and Conservation: A Handbook of Techniques. Dodd, C.K., Jr. (Ed.). Oxford University Press, Oxford, UK.
MacArthur, R.H., and E.R. Pianka. 1966. On optimal use of a patchy environment. American Naturalist 100:603–609.
Modry, D., L. Rifai, M. Abu Baker, Z. Amr. 2004. Amphibians and reptiles of the Hashemite Kingdom of Jordan. Denisia 14:407–420.
Norfolk, O., A. Melotte, F. Gilbert, S. Zalat, and T. Reader. 2010. A comparative study of two agamid lizards, Laudakia stellio and Pseudotrapelus sinaitus, in southern Sinai. Egyptian Journal of Biology 12:27–43.
Ortega, A., M.E. Maury, and R. Barbault. 1982. Spatial organization and habitat partitioning in a mountain lizard community of Mexico. Acta Oecologia 3:323–330.
Pérez-Mellado, V. 1992. Ecology of lacertid lizards in a desert area of eastern Morocco Journal of Zoology. 226:369–386.
Perry, G., I. Lampl, A. Lerner, D. Rotherstein, E. Shani, N. Sivan, and Y.L. Werner. 1990. Foraging mode in lacertid lizards: variation and correlates. Amphibia-Reptilia 11:373–384.
Pianka, E.R. 1966. Convexity, desert lizards and spatial heterogeneity. Ecology 47:1055–1059.
Pianka, E.R. 1969. Sympatry of desert lizards (Ctenotus) in western Australia. Ecology 50:1012–1030.
Pianka, E.R. 1973. The structure of lizard communities. Annual Reviews of Ecology and Systematics 4:53–74.
Pianka, E.R. 1975. Niche relations of desert lizards. Pp.
292–314 In Ecology and Evolution of Communities. Cody, M.L., and J.M. Diamond (Eds). Harvard University Press, Cambridge, Massachusetts, USA.
Pianka, E.R. 1986. The Ecology and Natural History of Desert Lizards. Princeton University Press, Princeton, New Jersey, USA.
Pianka, E.R., and R.B. Huey. 1978. Comparative ecology, resource utilization and niche segregation among gekkonid lizard in the southern Kalahari. Copeia 1978:691–701.
Pianka, E.R., R.B. Huey, and L.R. Lawlor. 1979. Niche segregation in desert lizards. Pp. 67–115 In Analysis of Ecological Systems. Horn, D.J., R. Mitchell, G.R. Stairs (Eds.). Ohio State University Press, Columbus, Ohio, USA.
Rifai, L., M. Abu Baker, D. Al Shafei, A. Disi, A. Mahasneh, and Z. Amr. 2005. Pseudopus apodus (PALLAS, 1775) from Jordan, with notes on its ecology. Herpetozoa 18:133–140.
Robson, G.M., and M.R.K. Lambert. 1980. Observations on the insect food of some semi-desert lizards in southern Morocco. Journal of Arid Environments 3:141–151.
Roll, U., A. Feldman, M. Novosolov, A. Allison, A.M. Bauer, R. Bernard, M. Böhm, F. Castro-Herrera, L. Chirio, B. Collen, et al. 2017. The global distribution of tetrapods reveal a need for targeted reptile conservation. Nature Ecology & Evolution 1:1677–1682.
Rouag, R., H. Djilali, H., Gueraiche, and L. Luiselli. 2007. Resource partitioning patterns between two sympatric lizard species from Algeria. Journal of Arid Environments 69:158–168.
Schoener, T.W. 1971. Theory of feeding strategies. Annual Review of Ecology and Systematics 2:369–404.
Schoener, T.W. 1974. Resource partitioning in ecological communities. Science 185:27–39
Schoener, T.W. 1975. Presence and absence of habitat shift in some widespread lizard species. Ecological Monographs 45:232–258.
Shenbrot, G., and B. Krasnov. 1997. Habitat relationships of the lizard fauna in the Ramon erosion cirque, Negev Highlands (Israel). Journal of Zoology 241:429–440.
Shenbrot, G.I., K.A. Rogovin, and A.V. Surov. 1991. Comparative analysis of spatial organization of desert lizard communities in Middle Asia and Mexico. Oikos 61:157–168.
Simpson, E.H. 1949. Measurement of diversity. Nature 163:688.
Strong, D.R., D. Simberloff, L.G. Abele, and A.B. Thistle (Eds.) 1984. Ecological Communities: Conceptual Issues and the Evidence. Princeton University Press, Princeton, New Jersey, USA.

124
Abu Baker et al.—Diet of lizard assemblages in Jordan.
Toft, C.A., 1985. Resource partitioning in amphibians and reptiles. Copeia 1985:1–21.
Western, D. 1974. The distribution, density and biomass
density of lizards in a semi-arid environment of northern Kenya. East African Wildlife Journal 12:49–62.
mohammaD a. abu bakeR is an Assistant Professor of Vertebrate Biology in the Department of Biological Sciences, University of Jordan, Amman. Mohammad earned a Ph.D. in Biology from the University of Illinois, Chicago, USA, where he studied habitat selection and coexistence of African small mammals. Thus far, Mohammad has over 70 publications on the ecology, behavior, and natural history of mammals and reptiles in Jordan, Qatar, South Africa, Egypt, and the USA. (Photographed by Bruce Patterson).
Ahmad A. Katbeh-Bader is a Professor of Entomology at the Department of Plant Protection, Fac-ulty of Agriculture, University of Jordan, Amman. He earned a Ph.D. in Entomology from Colorado University, Boulder, USA. He has published over 70 papers in entomology in Jordan. Ahmad has col-laborated on several occasions with Zuhair Amr on diet analysis of owls and on other vertebrate studies. (Photographed by Wafa Nasir).
ansam a. ghlelat is a Master’s student in Biological Sciences at the University of Jordan, Amman. Her research investigates the biology, ecology, and conservation of an endemic species of a freshwater fish in Jordan. She earned a B.Sc. in Biological Sciences from the University of Jordan. She is currently volunteering as a Conservation Ecologist at the Royal Society for the Conservation of Nature in Amman, Jordan. Ansam has much experience in classification and curation of museum specimens. (Photographed by Mo'men Ghlelat).
ahmaD m. Disi is an Emeritus Professor of Zoology in the Department of Biological Sciences, Uni-versity of Jordan, Amman. He earned a Ph.D. in Zoology from the University of Wisconsin, Madison, USA, and has published over 70 papers on various aspects of ecology and systematics of reptiles in Jordan. His book Amphibians and Reptiles of the Hashemite Kingdom of Jordan is considered by far as the most comprehensive reference for the reptiles of Jordan and the Middle East. (Photographed by Zuhair Amr).
zuhaiR s. amR is a Professor of Zoology and Animal Ecology in the Department of Biology, Jordan University of Science and Technology, Irbid. Zuhair earned a Ph.D. in Zoology from the University of Rhode Island, Kingston, USA, and has published over 200 papers and 10 books on various aspects of the ecology and systematics of mammals and reptiles in Jordan, Lebanon, Syria, the Palestinian Territories, and Saudi Arabia. Zuhair serves as the scientific authority for CITES in Jordan. (Photographed by Nashat Hamidan).

125
Herpetological Conservation and Biology
Acanthodactylus sp.
Eumeces schneideri
Stellagama stellio
Pseudotrapelus sinaitus
Trapelus agnetae
Trapelus persicus
Arachnida
Solifuga (Camel spiders) 1 0 0 1 1 0
Araneae (Spiders) 3 0 2 1 1 0
Scorpiones 0 0 0 0 0 2
Hemiptera
Cicadellidae 0 0 0 0 2 0
Pentatomidae 0 0 7 0 32 0
Unidentified Hemiptera 2 0 0 3 0 0
Lepidoptera
Noctuidae (Moths) 0 0 0 0 6 0
Lepidoptera larvae 5 2 0 0 1 0
Unidentified Lepidoptera 0 0 1 1 0 0
Hymenoptera
Ichneumonidae 0 0 5 0 0 0
Pompilidae 0 0 0 0 1 1
Sphecidae 0 0 0 0 1 0
Chrysididae 0 0 0 1 0 0
Unidentified wasp or bee 2 0 11 6 12 4
Formicidae 128 0 373 112 1093 125
Orthoptera
Acrididea (Grasshoppers) 0 1 2 4 4 1
Tettigoniidae (Bush crickets) 0 0 1 0 0 0
Orthoptera egg or larvae 0 0 196 0 0 0
Blattodea
Blattidae (Cockroaches) 0 0 1 0 0 0
Isoptera (Termites) 39 0 0 51 94 1
Diptera 1 0 4 0 1 0
Dermaptera 0 0 0 0 0 0
Coleoptera
Buprestidae 0 2 1 0 0 0
Tenebrionidae 20 3 7 9 24 3
Carabidae 1 0 0 0 1 0
Curculionidae 0 0 1 1 1 0
Scarabaeidae 0 5 0 1 0 0
glaphyridae 0 0 0 0 0 0
Meloidae 0 0 1 0 0 0
Unidentified Coleoptera 0 0 1 2 5 2
Unidentified Coleopteran larvae 8 0 0 0 2 0
Isopoda
Sowbugs 0 0 0 1 0 0
Seeds 0 0 0 0 0 23
Small vertebrates 0 0 0 0 1 1
Total no. of prey items 210 13 614 194 1283 163
appenDix 1. Prey items found in the stomachs of fringe-toed lizards (Acanthodactylus spp.), Orange-tailed Skinks (Eumeces schneideri), Starred Agamas (Stellagama stellio), Sinai Agamas (Pseudotrapelus sinaitus), Pale Agamas (Trapelus agnetae), and Persian Agamas (Trapelus persicus) from the eastern deserts of Jordan.
126
Abu Baker et al.—Diet of lizard assemblages in Jordan.
Eumeces schneideri Stellagama stellio Pseudotrapelus sinaitus
Arachnida
Solifuga (Camel spiders) 0 1 1
Acarina (Ticks) 0 0 2
Araneae (Spiders) 1 1 0
Scorpiones 0 0 2
Hemiptera
Cicadellidae 0 1 0
Rhopalidae 0 1 0
Pentatomidae 0 2 0
Unidentified Hemiptera 1 0 0
Lepidoptera
Noctuidae (Moths) 9 1 0
Lepidoptera larvae 2 5 2
Unidentified Lepidoptera 0 1 0
Hymenoptera
Unidentified wasp or bee 0 37 20
Unidentified Formicidae 0 1005 681
Orthoptera
Acrididea (Grasshoppers) 3 6 1
Gryllidae (Crickets) 1 0 0
Tettigoniidae (Bush crickets) 0 1 0
Orthoptera egg or larvae 3 0 6
Blattodea
Blattidae (Cockroaches) 0 1 0
Isoptera (Termites) 0 12 29
Dermaptera 1 0 0
Coleoptera
Buprestidae 0 1 1
Tenebrionidae 1 22 11
Carabidae 1 0 0
Coccinellidae 0 4 1
Chrysomelidae 0 5 0
Curculionidae 0 2 0
Scarabaeidae 0 13 0
Glaphyridae 1 7 0
Unidentified Coleoptera 12 2 2
Coleoptera larvae 1 3 1
Small vertebrates 1 1 2
Total no. of prey items 38 1135 762
appenDix 2. Prey items found in the stomachs of Orange-tailed Skinks (Eumeces schneideri), Starred Agamas (Stellagama stellio), and Sinai Agamas (Pseudotrapelus sinaitus) from the western highlands of Jordan.

127
Herpetological Conservation and Biology
app
en
Dix
3.
Die
t com
posi
tion
and
food
nic
he b
read
ths
base
d on
food
item
s fo
und
in s
tom
achs
of 1
7 fr
inge
-toed
liza
rds
(Aca
ntho
dact
ylus
spp
.), th
ree
Ora
nge-
taile
d Sk
inks
(E
umec
es s
chne
ider
i), 1
2 St
arre
d A
gam
as (
Stel
laga
ma
stel
lio),
28 P
ale
Aga
mas
(Tr
apel
us a
gnet
ae),
10 S
inai
Aga
mas
(Ps
eudo
trap
elus
sin
aitu
s), a
nd e
ight
Per
sian
Aga
mas
(T
rape
lus p
ersi
cus)
from
the
east
ern
dese
rt of
Jord
an.
The
acro
nym
s are
BS
= Si
mps
on’s
Div
ersi
ty In
dex
and
BL
= Le
vin’
s Sta
ndar
dize
d D
iver
sity
Inde
x.Ac
anth
odac
tylu
s sp.
Eum
eces
schn
eide
riSt
ella
gam
a st
ellio
Pseu
dotr
apel
us si
naitu
sTr
apel
us a
gnet
aeTr
apel
us p
ersi
cus
Gro
upn
%%
st
omac
hsn
%%
st
omac
hsn
%%
st
omac
hsn
%%
st
omac
hsn
%%
st
omac
hsn
%%
st
omac
hs
Ara
chni
da4
1.90
25.0
00
0.00
0.00
20.
3316
.67
21.
0320
.00
20.
167.
142
1.23
25.0
0
Hem
ipte
ra2
0.95
12.5
00.
000.
007
1.14
25.0
03
1.55
20.0
034
2.65
14.2
90
0.00
0.00
Lepi
dopt
era
52.
3831
.25
215
.38
33.3
31
0.16
8.33
10.
5210
.00
70.
5510
.71
00.
000.
00
Hym
enop
tera
130
61.9
75.0
00.
000.
0038
963
.36
75.0
011
961
.34
70.0
011
0786
.28
85.7
113
079
.75
75.0
0
Orth
opte
ra0
0.00
0.00
17.
6933
.33
199
32.4
125
.00
42.
0640
.00
40.
3114
.29
10.
6112
.50
Bla
ttode
a39
18.5
725
.00
0.00
0.00
10.
168.
3351
26.2
920
.00
947.
3325
.00
10.
6112
.50
Dip
tera
10.
486.
250
0.00
0.00
40.
658.
330
0.00
0.00
10.
083.
570
0.00
0.00
Der
map
tera
00.
000.
000
0.00
0.00
00.
000.
000
0.00
0.00
00.
000.
000
0.00
0.00
Col
eopt
era
2913
.81
75.0
1076
.92
66.6
711
1.79
50.0
013
6.70
70.0
033
2.57
53.5
75
3.07
62.5
0
Isop
oda
00.
000.
000
0.00
0.00
00.
000.
001
0.52
10.0
00
0.00
0.00
00.
000.
00
seed
s0
0.00
0.00
00.
000.
000
0.00
0.00
00.
000.
000
0.00
0.00
2314
.11
37.5
0
verte
brat
e0
0.00
0.00
00.
000.
000
0.00
0.00
00.
000.
001
0.08
3.57
10.
6112
.50
Tota
l no.
of p
rey
item
s21
010
0.00
1310
0.00
614
100.
0019
410
0.00
1283
100.
0016
310
0.00
B S2.
281.
611.
972.
221.
331.
52
B L0.
128
0.06
10.
097
0.12
20.
033
0.05
2












![Efficient, microwave-mediated synthesis of benzothiazole ... · building blocks for the synthesis of novel pyrazolo[1,5-a]pyrimidine and [1,2,4]-triazolo[1,5-a]pyramidine derivatives](https://static.fdocuments.in/doc/165x107/605da49ff8de38601e337786/efficient-microwave-mediated-synthesis-of-benzothiazole-building-blocks-for.jpg)





