
Localization of CGRP, CGRP receptor, PACAP and … glutamate in trigeminal ganglion. Relation to the...
17
www.elsevier.com/locate/brainres Available online at www.sciencedirect.com Research Report Localization of CGRP, CGRP receptor, PACAP and glutamate in trigeminal ganglion. Relation to the blood–brain barrier Sajedeh Eftekhari a,n , Christopher A. Salvatore b , Sara Johansson c , Tsing-bau Chen d , Zhizhen Zeng d , Lars Edvinsson a,c a Department of Clinical Sciences, Division of Experimental Vascular Research, Lund University, BMC A13, Sölvegatan 17, SE-22184 Lund, Sweden b Department of Pain and Migraine Research, Merck Research Laboratories, West Point, PA, USA c Department of Clinical Experimental Research, Glostrup Research Institute, Glostrup Hospital, Glostrup, Denmark d Department of Imaging, Merck Research Laboratories, West Point, PA, USA article info Article history: Accepted 13 November 2014 Available online 20 November 2014 Keywords: CGRP receptor antagonists CLR RAMP1 CGRP Evans blue Trigeminal ganglion abstract Calcitonin gene-related peptide (CGRP) receptor antagonists have demonstrated anti- migraine efficacy. One remaining question is where do these blockers act? We hypothe- sized that the trigeminal ganglion could be one possible site. We examined the binding sites of a CGRP receptor antagonist (MK-3207) and related this to the expression of CGRP and its receptor in rhesus trigeminal ganglion. Pituitary adenylate cyclase-activating polypeptide (PACAP) and glutamate were examined and related to the CGRP system. Furthermore, we examined if the trigeminal ganglion is protected by the blood–brain barrier (BBB). Autoradiography was performed with [ 3 H]MK-3207 to demonstrate receptor binding sites in rhesus trigeminal ganglion (TG). Immunofluorescence was used to correlate binding and the presence of CGRP and its receptor components, calcitonin receptor-like receptor (CLR) and receptor activity-modifying protein 1 (RAMP1), and the distribution of PACAP and glutamate in rhesus and rat TG. Evans blue was used to examine large molecule penetration into the rat TG. High receptor binding densities were found in rhesus TG. Immunofluorescence revealed expression of CGRP, CLR and RAMP1 in trigeminal cells. CGRP positive neurons expressed PACAP but not glutamate. Some neurons expressing CLR and RAMP1 co-localized with glutamate. Evans blue revealed that the TG is not protected by BBB. This study demonstrates CGRP receptor binding sites and expression of the CGRP receptor in rhesus and rat TG. The expression pattern of PACAP and glutamate http://dx.doi.org/10.1016/j.brainres.2014.11.031 0006-8993/& 2015 Elsevier B.V. All rights reserved. Abbreviations: BBB, blood–brain barrier; CGRP, calcitonin gene-related peptide; CLR, calcitonin receptor-like receptor; RAMP1, receptor activity-modifying protein 1; PACAP, pituitary adenylate cyclase-activating polypeptide; PBS, phosphate buffered-saline; BSA, bovine serum albumin; PFA, paraformaldehyde; Htx–eosin, hematoxylin–eosin; PBST, phosphate buffered- saline (PBS) containing 0.25% Triton X-100 n Corresponding author. E-mail address: [email protected] (S. Eftekhari). brain research 1600 (2015) 93–109
Transcript of Localization of CGRP, CGRP receptor, PACAP and … glutamate in trigeminal ganglion. Relation to the...

Available online at www.sciencedirect.com
www.elsevier.com/locate/brainres
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 9
http://dx.doi.org/100006-8993/& 2015 El
Abbreviations: BB
RAMP1, receptor a
buffered-saline; BSA
saline (PBS) containnCorresponding aE-mail address: S
Research Report
Localization of CGRP, CGRP receptor, PACAPand glutamate in trigeminal ganglion. Relationto the blood–brain barrier
Sajedeh Eftekharia,n, Christopher A. Salvatoreb, Sara Johanssonc,Tsing-bau Chend, Zhizhen Zengd, Lars Edvinssona,c
aDepartment of Clinical Sciences, Division of Experimental Vascular Research, Lund University, BMC A13, Sölvegatan 17,SE-22184 Lund, SwedenbDepartment of Pain and Migraine Research, Merck Research Laboratories, West Point, PA, USAcDepartment of Clinical Experimental Research, Glostrup Research Institute, Glostrup Hospital, Glostrup, DenmarkdDepartment of Imaging, Merck Research Laboratories, West Point, PA, USA
a r t i c l e i n f o
Article history:
Accepted 13 November 2014
Calcitonin gene-related peptide (CGRP) receptor antagonists have demonstrated anti-
migraine efficacy. One remaining question is where do these blockers act? We hypothe-
Available online 20 November 2014
Keywords:
CGRP receptor antagonists
CLR
RAMP1
CGRP
Evans blue
Trigeminal ganglion
.1016/j.brainres.2014.11.03sevier B.V. All rights res
B, blood–brain barrier
ctivity-modifying protein
, bovine serum albumi
ing 0.25% Triton [email protected]
a b s t r a c t
sized that the trigeminal ganglion could be one possible site. We examined the binding
sites of a CGRP receptor antagonist (MK-3207) and related this to the expression of CGRP
and its receptor in rhesus trigeminal ganglion. Pituitary adenylate cyclase-activating
polypeptide (PACAP) and glutamate were examined and related to the CGRP system.
Furthermore, we examined if the trigeminal ganglion is protected by the blood–brain
barrier (BBB). Autoradiography was performed with [3H]MK-3207 to demonstrate receptor
binding sites in rhesus trigeminal ganglion (TG). Immunofluorescence was used to
correlate binding and the presence of CGRP and its receptor components, calcitonin
receptor-like receptor (CLR) and receptor activity-modifying protein 1 (RAMP1), and the
distribution of PACAP and glutamate in rhesus and rat TG. Evans blue was used to examine
large molecule penetration into the rat TG. High receptor binding densities were found in
rhesus TG. Immunofluorescence revealed expression of CGRP, CLR and RAMP1 in
trigeminal cells. CGRP positive neurons expressed PACAP but not glutamate. Some neurons
expressing CLR and RAMP1 co-localized with glutamate. Evans blue revealed that the TG is
not protected by BBB. This study demonstrates CGRP receptor binding sites and expression
of the CGRP receptor in rhesus and rat TG. The expression pattern of PACAP and glutamate
1erved.
; CGRP, calcitonin gene-related peptide; CLR, calcitonin receptor-like receptor;
1; PACAP, pituitary adenylate cyclase-activating polypeptide; PBS, phosphate
n; PFA, paraformaldehyde; Htx–eosin, hematoxylin–eosin; PBST, phosphate buffered-
.se (S. Eftekhari).

Fig. 1 – In vitro autoradiography with [3H]to the left show [3H]MK-3207 high bindinalso its root (middle panel, red signal). Bstained with Htx–eosin. Since scanning owith different resolution, some discrepanusing unlabeled MK-3207 (non-displacea
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 994
suggests a possible interaction between the glutamatergic and CGRP system. In rat the TG
is outside the BBB, suggesting that molecules do not need to be CNS-penetrant to block
these receptors.
& 2015 Elsevier B.V. All rights reserved.
1. Introduction
Migraine is today recognized as a neurovascular disorder whichoriginates in the brain, involving the hypothalamus and thala-mus, as well as certain brainstem regions (Goadsby, 2012). Theacute migraine attack is often preceded by prodromal symptoms,which suggest the central nervous system (CNS) as a likely pointof origin (Charles, 2013). The pain during a migraine attack isassociated with the release of the peptide calcitonin gene-relatedpeptide (CGRP) which has a key role in migraine pathophysiology(Goadsby et al., 1988; Edvinsson and Goadsby, 1990; Ho et al.,2010). Clinical studies have demonstrated increased levels ofCGRP putatively from the trigeminal system that can be foundin serum, cerebrospinal fluid, and saliva (Goadsby et al., 1990;Goadsby and Edvinsson, 1993; Bellamy et al., 2006; Cernuda-Morollon et al., 2013). In addition, systemic infusion of CGRP cantrigger a “migraine-like headache” in patients (Hansen et al.,2010). It is hypothesized that CGRP acts at second order neuronsin the trigeminal nucleus caudalis (TNC) and at the C1–2 level ofthe spinal cord, to transmit pain signals to thalamus and highercortical pain regions (Goadsby, 2007; Levy et al. 2005). Thetrigeminovascular system is involved in the regulation of thecranial vasculature and is a key element in the transmission of
MK-3207, a CGRP recepg density within the trinding is also found inf the autoradiographycies may occur. In theble binding) is demons
pain. The trigeminal ganglion contains neurons that peripherally
innervate the intracranial vasculature and dura mater. The
trigeminal ganglion also centrally projects to the brainstem, in
related extensions down to the spinal cord and to parts of the
CNSwhere nociceptive information is processed to higher cortical
regions (Liu et al., 2009; Goadsby, 2012). Stimulation of the
trigeminal ganglion resulted in release of CGRP and elevation of
CGRP in the external jugular vein (Goadsby et al., 1988; Limmroth
et al., 2001). Neural activity in the trigeminal nociceptive system
in migraine patients has been demonstrated using imaging
techniques (Borsook et al., 2006). The wide distribution of CGRP
receptors in the trigeminovascular system is consistent with a
role in migraine pathophysiology (Lennerz et al., 2008; Eftekhari
and Edvinsson, 2010; Eftekhari et al., 2010; Bhatt et al., 2014).The most important evidence for the role of CGRP in
migraine pain came from the development of CGRP receptorantagonists (Olesen et al., 2004; Ho et al., 2008a, 2008b) whichact by blocking the action of CGRP on the CGRP-receptorcomplex. The CGRP receptor is a G protein-coupled receptor ofthe B-type consisting of calcitonin receptor-like receptor (CLR)and receptor activity-modifying protein 1 (RAMP1), both neces-sary to yield a functional CGRP receptor (Walker and Hay, 2013).It has been suggested that elevated neuronal RAMP1 could
tor antagonist, in the rhesus monkey. The two upper imagesigeminal ganglion (total binding, yellow and red signal), andareas with collagen. To the right, the same sections wereand the HTX-staining were performed in different scannerslower panel, self-block on the adjacent slide with no bindingtrated.

b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 9 95
potentially sensitize the trigeminal ganglia of individuals toCGRP actions (Zhang et al., 2007). However, little is currentlyknown about the regulation of RAMP1 levels in migraine.
Recent findings have identified the blockade of the CGRPreceptor as a mechanism to reduce migraine pain (Salvatoreand Kane, 2011; Moore and Salvatore, 2012). Clinical studiesusing CGRP receptor antagonists have demonstrated clinicalefficacy comparable to that of triptans in the treatment ofacute migraine attacks (Edvinsson and Linde, 2010; Ho et al.,2010; Salvatore and Kane, 2011). Therefore, it is of great
Fig. 2 – Immunohistochemistry of CGRP, CLR and RAMP1 in rhesuin the neurons (arrows). Thin fibers expressing CGRP is found (ar(arrows) and satellite glial cells (arrowheads). Negative cells forstaining nuclei, is used (blue). (C) Double immunolabelling of CG(small arrows) that lack expression of CLR or RAMP1 (large arroneurons (arrow heads). Auto-fluorescence from lipofuscin is detused in the merged pictures (blue).
interest to define where the CGRP receptor is expressed andon which possible sites drugs blocking CGRP signaling mayhave their therapeutic effect.
Two other neuronal messenger molecules that have beensuggested to have an important role in migraine pathophysiol-ogy are pituitary adenylate cyclase-activating polypeptide(PACAP) and glutamate. Recent studies suggest that PACAPmay have similar actions as CGRP and has been suggested tobe involved in migraine pathogenesis (Kaiser and Russo, 2013;Tuka et al., 2013; Edvinsson, 2014). Also glutamate is implicated
s monkey trigeminal ganglion. (A) CGRP is widely expressedrow heads). (B) CLR and RAMP1 immunoreactivity in neuronsCLR or RAMP1 are pointed out with small arrows. DAPI,RP and CLR or RAMP1. CGRP is mostly expressed in neuronsws). Co-localization is rarely found, and then in the smallerected in some neurons (asterisk). DAPI, staining nuclei, is
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 996
in migraine pathophysiology involving trigeminovascular activa-tion, central sensitization and cortical spreading depression.Therefore, the use of glutamate receptor antagonists has beensuggested for migraine treatment (Andreou and Goadsby, 2009;Marin and Goadsby, 2010). In relation to the trigeminal systemboth PACAP and glutamate are found in neurons in the trigem-inal ganglion and in the trigeminocervical complex (Kai-Kai andHowe, 1991; Tajti et al., 1999; Uddman et al., 2002). However, theirrelation to the CGRP system (mainly the CGRP receptor) in thetrigeminal ganglion has not been fully evaluated.
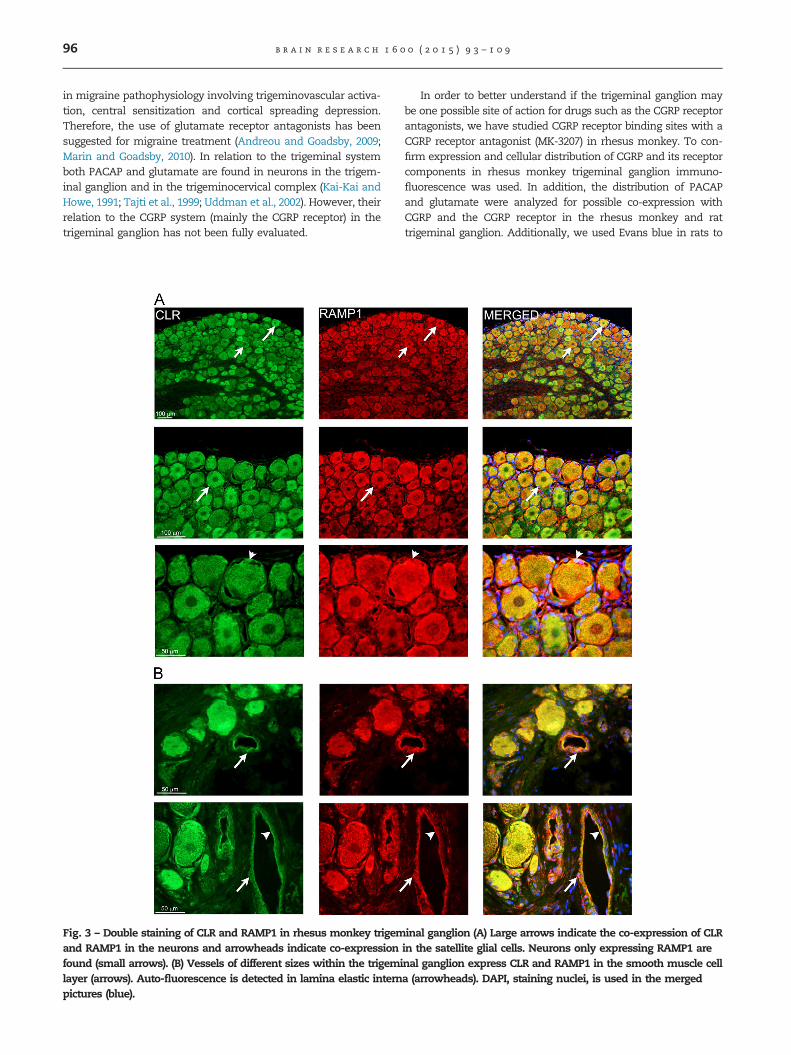
Fig. 3 – Double staining of CLR and RAMP1 in rhesus monkey trigemand RAMP1 in the neurons and arrowheads indicate co-expressionfound (small arrows). (B) Vessels of different sizes within the trigemilayer (arrows). Auto-fluorescence is detected in lamina elastic internpictures (blue).
In order to better understand if the trigeminal ganglion maybe one possible site of action for drugs such as the CGRP receptorantagonists, we have studied CGRP receptor binding sites with aCGRP receptor antagonist (MK-3207) in rhesus monkey. To con-firm expression and cellular distribution of CGRP and its receptorcomponents in rhesus monkey trigeminal ganglion immuno-fluorescence was used. In addition, the distribution of PACAPand glutamate were analyzed for possible co-expression withCGRP and the CGRP receptor in the rhesus monkey and rattrigeminal ganglion. Additionally, we used Evans blue in rats to
inal ganglion (A) Large arrows indicate the co-expression of CLRin the satellite glial cells. Neurons only expressing RAMP1 arenal ganglion express CLR and RAMP1 in the smooth muscle cella (arrowheads). DAPI, staining nuclei, is used in the merged

b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 9 97
study the relation of trigeminal ganglion to the blood brain-barrier (BBB).
2. Results
2.1. Binding of [3H]MK-3207 in the rhesus monkeytrigeminal ganglion
MK-3207 was labeled with tritium to high specific activity,and used for in vitro autoradiographic studies for binding sitelocalization in rhesus monkey trigeminal ganglia slices. Highbinding densities of [3H]MK-3207 were observed in the tri-geminal ganglion of rhesus monkey (Fig. 1). High binding wasfound in the central parts of the trigeminal ganglion wherethe ganglion is located and in the trigeminal root, verified byHtx–eosin staining on the same slide (Fig. 1). Areas with lowor no binding were also found. Minimal non-displaceablebinding was seen in the presence of 1 mM unlabeled MK-3207on an adjacent slice. The exact orientation of the ganglia wasmissing and regional variations (V1–V3) in the binding sitescould not be determined. Also, the exact cellular binding (inneurons or glial cells) could not be clearly delineated byautoradiography due to low resolution of the technique andtherefore the immunofluorescence method was used todemonstrate expression and cellular localization.
2.2. Expression of CGRP, CLR and RAMP1 in rhesusmonkey trigeminal ganglion
To correlate binding and the presence of CGRP and its receptorcomponents in rhesus monkey trigeminal ganglion, immuno-fluorescence was used. Immunoreactivity for CGRP, CLR andRAMP1 were observed in the trigeminal neurons (Fig. 2). TheCGRP immunoreactivity was mainly found in the neurons andin thin fibers but not in glial cells (Fig. 2A). The receptorcomponents, CLR and RAMP1, were expressed in neurons andin the satellite glial cells, which surround the neurons (Fig. 2B).Double-staining of CGRP with CLR or RAMP1 showed that the
Fig. 4 – Expression of PACAP and CGRP in rhesus monkey and rthe trigeminal neurons (large arrows). Not all PACAP positive neuor neurons lacking PACAP expression are found (asterisks). Arr
neuropeptide rarely co-expresses with the receptor components(Fig. 2C). The detailed distribution and cellular quantificationhave been previously described for rat and human trigeminalganglion (Eftekhari et al., 2010). Double-staining of CLR andRAMP1 revealed co-expression between the receptor compo-nents in some of the neurons and in the satellite glial cells(Fig. 3A), supporting expression of a functional CGRP receptor.In addition, RAMP1 and CLR were co-localized in the walls ofthe vessels within the trigeminal ganglion, suggestive of vas-cular receptor sites. This was apparent in both large and smallvessels in the trigeminal ganglion of rhesus monkey (Fig. 3B).
2.3. Distribution of PACAP, glutamate and their relationto the CGRP system in rhesus monkey and rat trigeminalganglion
Due to the difficulty in accessing rhesus tissues, rat trigeminalganglia were used as a surrogate to confirm the primate results.All the double-stainings were confirmed and similar stainingpatterns were found in primate and rat trigeminal ganglion.Double-staining of PACAP and CGRP revealed co-expression inthe trigeminal neurons of rhesus and rat (Fig. 4); however, not allPACAP positive neurons expressed CGRP (Fig. 4). Cell counting inrat trigeminal ganglia showed that immunoreactivity for CGRPwas found in 49% of the positive neurons (positive cells/total ofnumber of cells; 1184/2478), PACAP in 29% (713/2478) and co-expression of CGRP and PACAP was found in 23% of the cells(581/2478). Double immunolabeling of PACAP and the CGRPreceptor components showed that PACAP did not overallco-localize with RAMP1 or CLR either in rhesus monkey or rattrigeminal ganglion (Fig. 5). Very few neurons showedco-localization of PACAP and RAMP1 (Fig. 5).
There was generally no co-localization between glutamate andCGRP in the trigeminal neurons (Fig. 6) as confirmed in rattrigeminal ganglion (Fig. 6). Double-staining of glutamate andRAMP1 or CLR showed co-localization in the trigeminal neurons ofrhesus and rat (Figs. 7 and 8). Neurons expressing only the CGRPreceptor components or only glutamate were also found (Figs. 7A
at trigeminal ganglion Co-expression of PACAP and CGRP inrons express CGRP (small arrows). Thin CGRP positive fibers
owheads point at neurons negative for both markers.

Fig. 5 – Localization of PACAP with CLR or RAMP1 in rhesus monkey and rat trigeminal ganglion Double-staining of PACAPwith the CGRP receptor components shows that co-localization occurs rarely in the trigeminal neurons. CLR or RAMP1 positiveneurons are most often negative for PACAP (large arrows). Arrowheads point at neurons expressing only PACAP. Very fewneurons express RAMP1 and PACAP (small arrows). Thick fibers expressing CLR or RAMP1 lack PACAP expression (asterisks).DAPI, staining nuclei, is used in the merged pictures (blue).
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 998
and 8). Expression of RAMP1 in satellite glial cells surrounding aglutamate positive neuron was also observed (Fig. 7B).
2.4. Evans blue in rats
In order to evaluate if the trigeminal ganglion is located withinor outside the BBB, experiments with Evans blue were per-formed on rats. Evans blue has high affinity for serum albuminand forms a large complex when given systemically. This
complex cannot cross the barrier. Virtually all Evans blue isbound to albumin, and consequently the neural tissue typicallyremains unstained. Evans blue becomes fluorescent whenlinked to proteins and can be examined using the fluorescencemicroscope. The brains from the rats injected with Evans bluedisplayed as expected no dye uptake (Fig. 9A). However, dyeuptake was observed in the pituitary gland (Fig. 9A). Sectionsfrom cortex and cerebellum showed no fluorescent signal(Fig. 9B), confirming that Evans blue uptake was not present

Fig. 6 – CGRP and glutamate double staining in rhesus monkey and rat trigeminal ganglion Most CGRP positive neurons (red,large arrows) do not co-express glutamate. Few CGRP positive neurons co-express glutamate (small arrows). Glutamate (green)is found in separate neurons than those positive for CGRP (arrowheads). Asterisks point at thin CGRP positive fibers lackingglutamate expression. DAPI, staining nuclei, is used in the merged pictures (blue).
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 9 99
in these parts of the brain. Sections of pituitary gland andchoroid plexus, regions that are without BBB, displayed high redfluorescence signal (Fig. 9C). These results confirm that theexperiments with Evans blue were adequately performed.Dissected trigeminal ganglia showed uptake of Evans blue(Fig. 9A). Fluorescence signal was observed in trigeminal gang-lion sections, where high signal was found in the ganglionwhere there were numerous cell bodies (Fig. 10). The nervebundles displayed no or low signals, while some signal wasobserved around the nerve (Fig. 10A). Evans blue fluorescenceappeared proximal to the neurons and/or around the neurons(Fig. 10B). Blood vessels of various diameters within the gang-lion, often observed in close proximity to the neurons, dis-played red fluorescence signal throughout the vessel wall(Fig. 10C). Sections from trigeminal ganglia without Evans blueinjection showed no fluorescent signal (Fig.10D).
3. Discussion
This is the first study to examine CGRP binding sites, expressionof CGRP and its receptor in rhesus monkey trigeminal ganglion.In addition, we related the findings to expression of PACAP andglutamate in rhesus monkey trigeminal ganglion and confirmed
the staining patterns in rat trigeminal ganglion. We found highbinding densities of [3H]MK-3207, a selective CGRP receptorantagonist, and confirmed the presence of CGRP receptor com-ponents by immunofluorescence in the rhesus monkey trigem-inal ganglion. These results support the presence of functionalCGRP receptors in the ganglion that can putatively be occupiedby CGRP receptor antagonists. The clinically effective CGRPreceptor antagonist, telcagepant, has recently been shown tohave low central receptor occupancy (Hostetler et al., 2013).Therefore, we further evaluated if the trigeminal ganglion isprotected by the BBB using in vivo administration of Evans bluein rats. These experiments revealed that the trigeminal ganglionis located outside the BBB and can therefore be reached bysystemic CGRP receptor antagonists.
The autoradiographic binding experiments revealed highbinding densities of the CGRP receptor antagonist MK-3207 inthe trigeminal ganglion of rhesus monkey, suggesting thepresence of functional CGRP receptors in this region. Thepharmacological characterization of this antagonist has beenpreviously described (Bell et al., 2010; Salvatore et al., 2010).Although MK-3207 is selective for the CGRP receptor versus therelated adrenomedullin, calcitonin and amylin 3 (AMY3) recep-tor, it is less selective versus the amylin 1 (AMY1) receptor. Inthese studies the concentration of [3H]MK-3207 (0.045 nM) was

Fig. 7 – Double staining of glutamate, CLR and RAMP1 in rhesus monkey trigeminal ganglion (A) Large arrows indicate CLR orRAMP1 and glutamate positive neurons and their co-localization. Small arrows indicate neurons only expressing CLR orRAMP1. (B) Higher magnification image shows expression of RAMP1 (red) in the satellite glial cells (arrowheads) surrounding aglutamate positive neuron (green). Glutamate positive neurons lacking RAMP1 expression is found (arrows). DAPI, stainingnuclei, is used in the merged pictures (blue).
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 9100
carefully selected to be approximately 2-fold greater than itsaffinity value on the CGRP receptor (Ki¼0.024 nM) to limit thepotential for binding on the AMY1 receptor where MK-3207 hasa Ki¼0.75 nM (Salvatore et al., 2010). Although it cannot becompletely ruled out that some of the signal in rhesustrigeminal ganglion is a result of AMY1 receptor binding, it isunlikely to be a major contributor to the binding signal.
To further confirm the presence of CGRP and its receptor,immunofluorescence experiments were performed in rhesusmonkey trigeminal ganglion. CGRP immunoreactivity wasexpressed in the neurons of the trigeminal ganglion and inthin fibers distributed among the neurons. This is in agree-ment with previous studies on rat and human trigeminalganglion where CGRP has been shown to be predominantlyexpressed in small diameter ganglion cells (Tsai et al., 1988;Nozaki et al., 1990; Alvarez et al., 1991; Eftekhari et al., 2010)
and very thin unmyelinated nerve fibers (Liu-Chen et al.,1986; Eftekhari et al., 2013c). CLR and RAMP1 wereco-expressed mainly in the larger neurons and in satelliteglial cells, supporting the existence of functional CGRPreceptors in the trigeminal ganglion. The present work is inagreement with studies on human and rat trigeminal gang-lion (Lennerz et al., 2008; Eftekhari et al., 2010). Co-expressionof CLR and RAMP1 was also found in blood vessels walls ofthe trigeminal ganglion, opening up the possibility for effectson the microvasculature within the trigeminal ganglion.Since CGRP is a strong vasodilator of cranial vessels(Edvinsson et al., 2010), it is likely that CGRP can act as alocal vasodilator on these vessels.
The trigeminal ganglion cells consist of pseudounipolarneurons and two types of glial cells. As observed in humanand rat trigeminal ganglion (Lennerz et al., 2008; Li et al.,

Fig. 8 – Double-staining of glutamate, CLR and RAMP1 in rat trigeminal ganglion Co-expression of CLR or RAMP1 andglutamate in the trigeminal neurons (large arrows). Small arrows indicate neurons only expressing RAMP1. Glutamatepositive neurons lacking RAMP1 expression is found (arrowsheads). DAPI, staining nuclei, is used in the merged pictures(blue).
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 9 101
2008; Eftekhari et al., 2010; Seiler et al., 2013), there was co-expression of the receptor components in the rhesus satelliteglial cells. This cell type has been suggested to have animportant role in inflammation and pain (Li et al., 2008;Raddant and Russo, 2011). In primary cultures of rat trigem-inal ganglia CGRP treatment increased glial cell iNOS expres-sion and release of NO, which was inhibited by a CGRPreceptor antagonist (Li et al., 2008), suggesting that CGRPcan activate satellite glial cells. In addition, CGRP may triggeradditional inflammatory gene expression changes in gliain vitro (Vause and Durham, 2010). It has been demonstratedthat the anti-migraine drug sumatriptan, a 5-HT1B/1D receptoragonist, may inhibit the evoked CGRP release from naïvetrigeminal neuron cultures, as well as from cultured trigem-inal ganglia (de Corato et al., 2011). The present studysuggests that CGRP receptor antagonists may act in thetrigeminal ganglion on the satellite glial cells and secondarilyinfluence the neuronal responses. Further functional studiesare needed to address this hypothesis.
Several other signaling molecules have been found in thetrigeminal ganglion, these include substance P, neurokinin A,dynorphine, nitric oxide and PACAP (Tajti et al., 1999; Houet al., 2001; Dieterle et al., 2011). Yet clearly there may bemany other contributing neuropeptides that could act inconcert with CGRP. One interesting candidate is PACAP,which shares some of the actions as CGRP (Kaiser and
Russo, 2013). Human studies indicate that PACAP is a potentheadache trigger and altered plasma PACAP levels have beenshown in migraineurs (Schytz et al., 2009; Tuka et al., 2013).PACAP belongs to the vasoactive intestinal polypeptide (VIP)secretin superfamily (Vaudry et al., 2009) and has beenidentified in human and rodent sensory and parasympatheticganglia, as well as in second-order neurons of the TNC (Tajtiet al., 2001; Uddman et al., 2002). Three PACAP receptors havebeen identified, termed PAC1, VPAC1 and VPAC2 (Vaudry et al.,2009). Like CGRP, PACAP dilates cranial blood vessels inhumans in association with headache (Amin et al., 2012)and it has some shared actions of CGRP that may contributeto migraine pathophysiology. We observed co-expression ofPACAP and CGRP in the trigeminal neurons of rhesus monkeyand rat, which is in support of earlier studies showing co-expression of CGRP and PACAP in the dorsal horn of thespinal cord and in the trigeminal ganglion (Moller et al., 1993).In the rat trigeminal ganglion, we found CGRP expression inabout 50% of the cells and PACAP in around 30% of theneurons. Co-expression of PACAP and CGRP was found inaround 20% of the immunoreactive neurons. Recently wedemonstrated nerve fibers in dura mater co-expressingPACAP and CGRP (Eftekhari et al., 2013c). Altogether, thissuggests that PACAP may have 2 sites of origin, a minorpopulation co-expressed with sensory CGRP and a largerparasympathetic distribution (Edvinsson et al., 2001).

Fig. 9 – Evans blue experiments in rodents (A) Evans blue experiments show no uptake of the dye in the brain while thepituitary gland that is lacking BBB protection is blue from the dye uptake. In addition, the trigeminal ganglion shows uptake ofEvans blue. (B) No fluorescent signal from Evans blue was detected in cortex or cerebellum, however (C) red fluorescent signalwas detected in tissues located outside BBB, such as the pituitary gland and choroid plexus.
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 9102
Our results demonstrate that around 20% of the PACAPneurons co-express with sensory CGRP in the trigeminalganglion.
The CGRP receptor components are rarely co-expressed withCGRP, suggesting a low presence of putative auto-receptorswithin the ganglion (Eftekhari et al., 2010). In the present studywe did not find co-expression of the CGRP receptor componentsand PACAP in the trigeminal neurons. Therefore it is unlikelythat CGRP receptor antagonists would affect interganglionicPACAP levels. However this needs further evaluation.
Another important neurotransmitter that may contribute tomigraine pathogenesis is glutamate. Findings from both animaland human studies suggest a link between glutamate andmigraine (Vikelis and Mitsikostas, 2007). Potassium-stimulatedcultured trigeminal neurons release both glutamate and CGRP(Xiao et al., 2008). Elevated glutamine levels have been observed inthe cerebrospinal fluid of migraine patients (Rothrock et al., 1995).Therefore, it has been suggested that pharmacological manage-ment of glutamate receptors may provide further insight intopotential migraine treatment (Vikelis and Mitsikostas, 2007;Sprenger and Goadsby, 2009; Marin and Goadsby, 2010). Weinvestigated the localization of glutamate in the rhesus and rattrigeminal ganglion with a focus on its relation to CGRP and theCGRP receptor. Our results demonstrate that glutamate and CGRPare mostly expressed in separate neurons. This suggests that
trigeminal neurons that express CGRP do not contain glutamateas a co-transmitter. In support of this observation, most CGRPcontaining axons in autonomic ganglia and neurons in dorsal rootganglia lack expression of the glutamate transporters VGluT1 andVGluT2, suggesting thatmost peptide-containing sensory neuronsdo not use glutamate as transmitter (Morris et al., 2005).
Instead, some RAMP1 and/or CLR positive neurons wereshown for the first time to co-express glutamate. Neuronsexpressing CGRP receptor components only in their satellite glialcells and glutamate in their cell body were also observed. Thisopens up the possibility of interaction between the glutamatesystem and CGRP receptors. It remains to be evaluated if CGRPreceptor antagonists may have an effect on glutamate release.
The CGRP receptor antagonists represent a potential new groupof anti-migraine drugs that may offer a non-vasoconstrictiveapproach in the treatment of migraine (Chan et al., 2010). SeveralCGRP receptor antagonists (olcegepant, telcagepant, MK-3207, BI44370TA and BMS927711) have displayed efficacy in the treatmentof migraine (Durham and Vause, 2010; Negro et al., 2012; Dolgin,2013). The CGRP receptor antagonists have potentially opened up anew option in migraine treatment; however, there remains activediscussion as to where these molecules act to reduce migrainepain. It has been shown that the currently used anti-migrainedrugs, the triptans, cannot penetrate the BBB under normalcircumstances and a central effect in the trigeminal nucleus can

Fig. 10 – Fluorescence micrograph from trigeminal ganglion following injection of Evans blue (A) Demonstration of afluorescence microscopic figure showing the trigeminal ganglion with red fluorescent signal from the dye Evans blue,indicating the lack of the blood brain barrier. High signal is detected within the ganglion where the cells are located while noor low signal is detected within the nerve. Arrows demonstrate a higher magnification of the selected areas, from left to right:nerve, ganglion and vessel. (B) The Evans blue signal is detected around the neurons. (C) Vessels within ganglion (arrows),close to the neurons, also shows signal for Evans blue uptake. DAPI, staining nuclei, is used in the merged pictures (blue). (D)Trigeminal ganglion tissue without Evans blue showing no fluorescence signal.
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 9 103

b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 9104
only be seen after BBB disruption (Kaube et al., 1993). Someexperimental data have suggested that there might be a minorbreakdown or leakage of the BBB during migraine with activationof matrix metalloproteinases (MMPs) following repeated corticalspreading depression (Gursoy-Ozdemir et al., 2004). If the BBB iscompromised it would enable the passage of drugs which typicallydo not enter the CNS. However, these data are indirect and there isno clear proof of breakdown or leakage of the BBB in patientsduring migraine attacks (Edvinsson and Tfelt-Hansen, 2008).
Clinical studies have shown that CGRP receptor antago-nists need to achieve high plasma concentrations in relation-ship to their intrinsic potency in order to elicit therapeuticeffects (Ho et al., 2008a). However, the first PET study inhealthy individuals and migraine patients demonstrated thatthe CGRP receptor antagonist telcagepant achieved low CGRPreceptor occupancy (10%) in the CNS at the lowest clinicallyefficacious dose, whereas a supra-therapeutic dose of telca-gepant (1120 mg) resulted in only moderate receptor occu-pancy in healthy volunteers. These data suggest that CGRPreceptor antagonists do not have to act centrally for clinicalefficacy (Vermeersch et al., 2012; Hostetler et al., 2013).
If the current CGRP receptor antagonists cannot pass theBBB (or only to a low degree), it is important to determinewhich of the sites that can be reached by CGRP receptorantagonists contribute to the clinical efficacy. In the peripherythe CGRP receptor is found in the smooth muscle cell layer ofcerebral arteries, meningeal vessels, nerve fibers and rodentmast cells of the dura mater (Eftekhari et al., 2013c). Periph-erally the CGRP receptor antagonists may act on cranialvessels to inhibit CGRP-induced vasodilatation and on duralmast cells to block degranulation and the subsequent releaseof inflammatory mediators. However, the role of cerebrovas-cular dilatation is unclear. Recently, it was shown that acutemigraine pain was not accompanied by extracranial arterialdilatation and only by minor intracranial dilatation duringacute migraine attacks (Amin et al., 2013). In the same study, itwas demonstrated that effective treatment with sumatriptancaused no intracranial (cerebral artery) vasoconstriction. It isunclear if mast cell degranulation contributes to neurogenicinflammation in humans, since it has been shown that CGRPreceptors are not present in human mast cells (Eftekhari et al.,2013c). This agrees with the early finding that CGRP failed toinduce release of histamine from human dura mater mastcells (Ottosson and Edvinsson, 1997).
In the present study we demonstrated that the trigeminalganglion is located outside the BBB and thus accessible tosystemic drugs, which to our knowledge has not previouslybeen clearly determined. There is only limited previous workaddressing this issue. It has been suggested that capillaries insensory and sympathetic ganglia are fully permeable tomacromolecules (Kiernan, 1996). A study from 1973 showedEvans blue uptake in the trigeminal nerve of rabbits and mice(Arvidsson et al., 1973). In another study, injection of Fluoro-Gold into the tongue showed dense accumulation of the dyein cells of the trigeminal ganglion (Merchenthaler, 1991) andinjection of horseradish peroxidase (HRP) showed uptake ofHRP in the trigeminal ganglion (Ten Tusscher et al., 1989).
In the present study injection of the dye Evans blue intothe circulation of rats showed uptake of the dye in areasknown to be outside the BBB such as pituitary gland and
choroid plexus, whereas the brain showed no dye uptake. Dyeuptake was clearly found in the trigeminal ganglion and thefluorescence signal was found mostly around the cells andvessels in proximity to the cells. Within the trigeminal nervethere was low to no signal observed.
This suggests that the trigeminal ganglion is not protectedby the BBB and can be reached by CGRP receptor antagonistsregardless of the molecules ability to cross the BBB. Thetrigeminal ganglion may also be a site of action for the recentlydeveloped monoclonal anti-CGRP and anti-CGRP receptor anti-bodies, which in all likelihood are too large to pass the BBB(Dolgin, 2013). Support for this has been obtained in functionaltests in rodents with a CGRP antibody (Edvinsson et al., 2007).CGRP receptor antagonists, anti-CGRP and anti-CGRP receptorantibodies are currently under development (Bigal et al., 2013),and if these modalities are not able to penetrate the BBB, thetrigeminal ganglion may be one site of action.
Since the trigeminal ganglion is connected to the peripheryand also projects centrally, CGRP signaling may be blocked tothese parts by blunting the activation/signaling in the trigem-inal ganglion. However, it cannot be ruled out that additionalefficacy may be achieved with better access to the CNS sincethere is wide spread expression of CGRP and its receptorthroughout the CNS and the putative role of brain areas suchas the brainstem and hypothalamus in the pathophysiology ofmigraine. Lipophilic triptans with ability to pass the BBB didnot show improved efficacy (Pascual and Munoz, 2005).Recently, a series of CNS-penetrant CGRP receptor antagonistshave been characterized (Joshi et al., 2014). It will be of greatinterest to assess if CNS-penetrant CGRP receptor antagonistsdemonstrate better clinical efficacy. Interestingly, we haveshown binding of a CGRP receptor antagonist via in vitroautoradiography in the pineal gland and area postrema ofrhesus monkey (Eftekhari et al., 2013a). These CNS areas arenot protected by the BBB suggesting that these areas can alsobe reached by drugs such as CGRP receptor antagonists.
4. Conclusion
The present study demonstrates for the first time in primatebinding sites of a CGRP receptor antagonist and expression ofCGRP and its receptor within the rhesus trigeminal ganglion.The results suggest and are in support of the presence offunctional CGRP receptors in this area. The receptor compo-nents were co-expressed in neurons and satellite glial cells.The study also shows co-localization between PACAP andCGRP, while glutamate co-localizes with the CGRP receptorcomponents in the trigeminal ganglion. The study alsoreveals that trigeminal ganglion is located outside the BBBand therefore CGRP receptor antagonists do not need to beCNS-penetrant to block receptors in the trigeminal ganglion.
5. Experimental procedure
5.1. Rhesus monkey tissue samples
Rhesus trigeminal ganglion (Macaca mulatta, n¼3, age 13–15years old, females) was harvested in accordance with a Merck

Table 1 – Secondary antibodies used for immunohistochemistry.
Conjugate and host Against Dilution Supplier
DyLight 549 (goat) Anti-rabbit 1:200 Jackson Immuoresearch, West Grove, PADyLight 549 (donkey) Anti-goat 1:200 Jackson Immuoresearch, West Grove, PADyLight 549 (donkey) Anti-sheep 1:200 Jackson Immuoresearch, West Grove, PADyLight 488 (donkey) Anti-goat 1:200 Jackson Immuoresearch, West Grove, PADyLight 488 (donkey) Anti-rabbit 1:200 Jackson Immuoresearch, West Grove, PADyLight 488 (donkey) Anti-sheep 1:200 Jackson Immuoresearch, West Grove, PAAlexa 594 (goat) Anti-mouse 1:400 Invitrogen, La Jolla, CAAlexa 594 (donkey) Anti-rabbit 1:400 Jackson Immuoresearch, West Grove, PAAlexa 488 (goat) Anti-guinea pig 1:200 Invitrogen, La Jolla, CAFITC (goat) Anti-rabbit 1:100 Cayman Chemical, Ann Arbor, MITexas-Red (rabbit) Anti-sheep 1:200 Jackson Immuoresearch, West Grove, PA
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 9 105
Research Laboratories Institutional Animal Care and UseCommittee approved protocol. Tissues to be used for auto-radiography studies were quickly removed and frozen on dryice. The samples were cryosectioned at 20 mm (cryostat modelCM3050: Leica Microsystems, Inc., Deerfield, IL) and collectedon cold Superfrosts Plus slides, and stored at �80 1C. Anadditional rhesus monkey (9 years old, female) trigeminalganglion was harvested for immunohistochemistry. Thesamples were immersed overnight in fixative consisting of4% paraformaldehyde (PFA) and in 0.1 mol/l phosphate buf-fer, pH 7.2. After fixation, specimens were paraffin embedded.The tissues were sent to the laboratory in Sweden forimmunohistochemistry experiments. The rhesus sampleswere sectioned (5 mm) and stored at room-temperature.CITES, import permits, for the Swedish part was approvedand given the permit number Dnr34-10088/10 nr 51016-10.CITES permit for export from USA was approved and giventhe permit number 10US11621A/9. All animal experimentscarried out in accordance with EC Directive 86/609/EEC foranimal experiments.
5.2. Rat tissue samples
Trigeminal ganglia were removed from 10 male Sprague-Dawley rats weighing 300–350 g (Scanbur, Stockholm, Sweden).The experiments were approved by the University AnimalEthics Committee (M 126-12), Lund University, Sweden. Theganglia were placed in 4% PFA and fixed for 2–3 h. Afterfixation the ganglia were rinsed in rising concentrations ofsucrose in Sörensen’s phosphate buffer, embedded in gelatinmedium (30% egg albumin, 3% gelatin in distilled water) andcryosectioned at 12 mm. The sections were stored at �20 1Cuntil use.
5.3. Autoradiography studies
Autoradiography studies were conducted as previouslydescribed (Salvatore et al., 2010). Slides were preincubatedfor 15 min in binding buffer (0.9% NaCl, 50 mM Tris–HCl, pH7.5, 2 mM KCl, 1 mM MgCl2 and 1 mM CaCl2) followed by90 min incubation with [3H]MK-3207 (0.045 nM; specific activ-ity 76 Ci/mmol) at room temperature (two slides from eachanimal). Non-displaceable binding was defined by blockingwith 1 mM unlabeled MK-3207 using adjacent slides (twoslides from each animal). Slides were washed 3�1 min in
ice-cold buffer (0.9% NaCl and 50 mM Tris–HCl, pH 7.5)followed by rinse in ice-cold water. The slides were air-dried and exposed to phosphorimaging plates (TR2025; Fuji-film Medical Systems USA, Inc., Stamford, CT) for 3 weeks andscanned with a BAS 5000 scanner (Fuji, Tokyo, Japan). Imageanalysis was conducted with MCID software (MCIC, Linton,Cambridge,UK). After the autoradiography studies, the slideswere stained with hematoxylin–eosin following a standardprotocol (Htx 3 min, water rinse, eosin 1 min) for orientationand examination of the tissue condition.
5.4. Immunofluorescence studies
Immunofluorescence staining was performed to demonstratethe localization of CGRP, CLR, RAMP1, PACAP and glutamate inthe rhesus monkey trigeminal ganglia. Paraffin sections weredeparaffinized in xylene followed by serials of alcohol. Tounmask the antibody epitopes, heat-induced epitope retrievalwas performed, where the sections were heated in a microwavefor 10min in citric acid solution pH 6.0. Rat trigeminal gangliawere used for comparison and confirmation of the stainingpatterns. The sections were then washed for 10min in PBS pH7.2 containing 0.25% Triton X-100 (PBST). The sections wereblocked for 1 h in blocking solution of PBS and 5% normaldonkey or goat serum (depending on species origin of thesecondary antibody). After blocking, the sections were incu-bated overnight at 4 1C for single or double immunolabellingwith primary antibodies against CGRP (mouse, ab81887, Abcam,UK), CLR (rabbit, 3152, sheep, 132, Merck & Co.), RAMP1 (goat,844, Merck & Co.), PACAP-38 (rabbit, T-4473, Bachem, USA) andglutamate (rabbit, AB5018, Millipore, USA). For all doubleimmunostainings, the antibodies were applied separately andnot mixed together. The primary antibodies were diluted inPBST containing 1% BSA and 3% normal serum. After incuba-tion with primary antibodies, sections were equilibrated toroom temperature, rinsed in PBST for 3�15min and exposedto secondary antibodies diluted in PBST and 1% BSA for 1 h atroom temperature. For detailed description of the secondaryantibodies, see Table 1. The sections were subsequently washedwith PBST for 3�15min and mounted with an anti-fadingmounting medium containing DAPI (Vectashield, Vector Labora-tories). The development and specificity of CLR and RAMP1antibodies have been demonstrated in our previous study, wherethe specificity of the antibodies was confirmed in HEK293 cellsstably expressing the human CGRP receptor and was confirmed

b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 9106
by Western blotting (Eftekhari et al., 2010). In our recent study,the antibodies were used on rhesus monkey tissue (Eftekhariet al., 2013b). All stainings were repeated to confirm the resultsand all slides were analyzed. Omission of the primary antibodyserved as negative controls for all antibodies.
5.5. Cell-counting
Cell counting was performed to quantify the expression ofCGRP, PACAP and their co-expression in rat trigeminal gang-lion (n¼5). Three slides (with a minimum of five sectionsbetween each were used for measurements to avoid double-counting of cells) from each animal were stained with therespective antibody. Four to six images (depending on thesize of the ganglia) were taken at magnification 10� in eachsection to cover the entire trigeminal ganglion. The NIS-elements BR image analysis program (Nikon) was used tocalculate the number of positive cells. Only cells with visiblenuclei were counted. CGRP, PACAP positive and double-stained cells were counted in each image. The total numberof cells positive for CGRP, PACAP, double-stained and totalcounted cells was calculated for each animal.
5.6. Evans blue experiment
Sprague-Dawley rats weighing 300–350 g (n¼4) were used forthe Evans blue experiments. Rat experiments were approvedby the Animal Ethics Committee of Copenhagen University,Denmark and carried and the experiments conform to the“European Convention for the protection of vertebrate ani-mals used for experimental and other scientific purpose(council of Europe No 123, Strasbourg 1985). The animalswere deeply anaesthetized subcutaneously (dose of 2.5 ml/kg)with a mixture of hypnorm–midazolam (1:1:2) in sterile water(containing 0.079 mg/ml fentanyl, 2.50 mg/ml fluanison and1.25 mg/ml midazolam). 2% Evans blue in saline (Sigma-Aldrich, MO, USA) was injected via the tail vein (4 ml/kg).After 1 h, the animals were perfused transcardially with aprewash of phosphate-buffered saline (PBS, pH 7.2) for 2 minfollowed by 4% paraformaldehyde (PFA) in PBS for 5 min. Thebrains and trigeminal ganglia were dissected and the speci-mens were further fixed in PFA for 2–4 h followed by rinses inrising concentrations of sucrose in Sörensen’s phosphatebuffer. The specimens were embedded in a gelatin medium(30% egg albumin, 3% gelatin in distilled water) and cryosec-tioned (12 μm). The sections were stored at �20 1C until use.
5.7. Microscopic analysis
Immunostained sections were examined and images wereobtained using a light- and epifluorescence microscope (Nikon80i, Tokyo, Japan) coupled to a Nikon DS-2MV camera. AdobePhotoshop CS3 (v.8.0, Adobe Systems, Mountain View, CA) wasused to visualise co-labeling by superimposing the digitalimages and processed for brightness and contrast. Evans bluesections were examined by a filter with settings for excitation510–550 nm and emission at 590 nm.
Conflict of interest statement
Sajedeh Eftekhari conducted research at Merck ResearchLaboratories as a visiting scholar. Christopher Salvatore,Tsing-bau Chen, and Zhizhen Zeng are employees of MerckSharp & Dohme Corp. and potentially own stock and/or holdstock options in the Company.
Authors’ contributions
SE performed/participated in all the experiments, studydesign, analyzed all data and drafted the manuscript. CASparticipated in designing the study, analyzing the data andhelped to draft the manuscript. SJ performed the Evans blueexperiments. T-BC performed the autoradiography experi-ments. ZZ participated in designing the study and analyzingthe autoradiography data. LE participated in designing thestudy and helped to draft the manuscript. All authors haveread and approved of the final version of the manuscript.
Acknowledgments
Thanks are due to the following from Merck ResearchLaboratories: Kenneth Lodge and Anjali Patel for collectingand paraffin embedding the rhesus tissue, Brett Connolly forH&E staining, and Ian Bell and Carolee Lavey for synthesis ofthe radiotracer. Swedish Research council (grant no 5958) andthe Swedish Migraine Society (2013) for funding.
r e f e r e n c e s
Alvarez, F.J., Morris, H.R., Priestley, J.V., 1991. Sub-populations ofsmaller diameter trigeminal primary afferent neurons definedby expression of calcitonin gene-related peptide and the cellsurface oligosaccharide recognized by monoclonal antibodyLA4. J. Neurocytol. 20, 716–731.
Amin, F.M., Asghar, M.S., Guo, S., Hougaard, A., Hansen, A.E.,Schytz, H.W., van der Geest, R.J., de Koning, P.J., Larsson, H.B.,Olesen, J., Ashina, M., 2012. Headache and prolongeddilatation of the middle meningeal artery by PACAP38 inhealthy volunteers. Cephalalgia 32, 140–149.
Amin, F.M., Asghar, M.S., Hougaard, A., Hansen, A.E., Larsen, V.A., deKoning, P.J., Larsson, H.B., Olesen, J., Ashina, M., 2013. Magneticresonance angiography of intracranial and extracranial arteriesin patients with spontaneous migraine without aura: a cross-sectional study. Lancet Neurol. 12, 454–461.
Andreou, A.P., Goadsby, P.J., 2009. Therapeutic potential of novelglutamate receptor antagonists in migraine. Expert Opin.Investig. Drugs 18, 789–803.
Arvidsson, B., Kristensson, K., Olsson, Y., 1973. Vascularpermeability to fluorescent protein tracer in trigeminal nerveand gasserian ganglion. Acta Neuropathol. 26, 199–205.
Bell, I.M., Gallicchio, S.N., Wood, M.R., Quigley, A.G., Stump, C.A.,Zartman, C.B., Fay, J.F., Li, C.C., Lynch, J.J., Moore, E.L., Mosser,S.D., Prueksaritanont, T., Regan, C.P., Roller, S., Salvatore, C.A.,Kane, S.A., Vacca, J.P., Selnick, H.G., 2010. Discovery of MK-3207: a highly potent, orally bioavailable CGRP receptorantagonist. ACS Med. Chem. Lett. 1, 24–29.

b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 9 107
Bellamy, J.L., Cady, R.K., Durham, P.L., 2006. Salivary levels of CGRP and
VIP in rhinosinusitis and migraine patients. Headache 46, 24–33.Bhatt, D.K., Gupta, S., Ploug, K.B., Jansen-Olesen, I., Olesen, J.,
2014. mRNA distribution of CGRP and its receptor components
in the trigeminovascular system and other pain related
structures in rat brain, and effect of intracerebroventricular
administration of CGRP on Fos expression in the TNC.
Neurosci. Lett. 559, 99–104.Bigal, M.E., Walter, S., Rapoport, A.M., 2013. Calcitonin gene-
related peptide (CGRP) and migraine current understanding
and state of development. Headache 53, 1230–1244.Borsook, D., Burstein, R., Moulton, E., Becerra, L., 2006. Functional
imaging of the trigeminal system: applications to migraine
pathophysiology. Headache 46 (Suppl. 1), S32–S38.Cernuda-Morollon, E., Larrosa, D., Ramon, C., Vega, J., Martinez-
Camblor, P., Pascual, J., 2013. Interictal increase of CGRP levels in
peripheral blood as a biomarker for chronic migraine. Neurology
81 14:1191-1196.Chan, K., Edvinsson, L., Eftekhari, S., Kimblad, P.O., Kane, S., Lynch,
J., Hargreaves, R., de Vries, R., Garrelds, I., van den Bogaerdt, A.,
Danser, J., Maassenvandenbrink, A., 2010. Characterization of
the CGRP receptor antagonist telcagepant (MK-0974) in human
isolated coronary arteries. J. Pharmacol. Exp. Ther. 334:746-752.Charles, A., 2013. Migraine: a brain state. Curr. Opin. Neurol. 26,
235–239.de Corato, A., Capuano, A., Curro, D., Tringali, G., Navarra, P.,
Dello Russo, C., 2011. Trigeminal satellite cells modulate
neuronal responses to triptans: relevance for migraine
therapy. Neuron Glia Biol. 7, 109–116.Dieterle, A., Fischer, M.J., Link, A.S., Neuhuber, W.L., Messlinger,
K., 2011. Increase in CGRP- and nNOS-immunoreactive
neurons in the rat trigeminal ganglion after infusion of an NO
donor. Cephalalgia 31, 31–42.Dolgin, E., 2013. Antibody drugs set to revive flagging migraine
target. Nat. Rev. Drug Discov. 12, 249–250.Durham, P.L., Vause, C.V., 2010. Calcitonin gene-related peptide
(CGRP) receptor antagonists in the treatment of migraine. CNS
Drugs 24, 539–548.Edvinsson, L., 2014. PACAP and its receptors in migraine
pathophysiology. Br. J. Pharmacol 10.1111/bph.12772.Edvinsson, L., Chan, K.Y., Eftekhari, S., Nilsson, E., de Vries, R.,
Saveland, H., Dirven, C.M., Danser, A.H., MaassenVanDenBrink,
A., 2010. Effect of the calcitonin gene-related peptide (CGRP)
receptor antagonist telcagepant in human cranial arteries.
Cephalalgia 30, 1233–1240.Edvinsson, L., Elsas, T., Suzuki, N., Shimizu, T., Lee, T.J., 2001.
Origin and Co-localization of nitric oxide synthase, CGRP,
PACAP, and VIP in the cerebral circulation of the rat. Microsc.
Res. Tech. 53, 221–228.Edvinsson, L., Goadsby, P.J., 1990. Extracerebral manifestations in
migraine. A peptidergic involvement? J. Intern. Med. 228, 299–304.Edvinsson, L., Linde, M., 2010. New drugs in migraine treatment and
prophylaxis: telcagepant and topiramate. Lancet 376, 645–655.Edvinsson, L., Nilsson, E., Jansen-Olesen, I., 2007. Inhibitory effect of
BIBN4096BS, CGRP(8-37), a CGRP antibody and an RNA-Spiegelmer
on CGRP induced vasodilatation in the perfused and non-perfused
rat middle cerebral artery. Br. J. Pharmacol. 150, 633–640.Edvinsson, L.1, Tfelt-Hansen, P., 2008. The blood–brain barrier in
migraine treatment. Cephalalgia 12, 1245–1258.Eftekhari, S., Edvinsson, L., 2010. Possible sites of action of the
new calcitonin gene-related peptide receptor antagonists.
Ther. Adv. Neurol. Disord. 3, 369–378.Eftekhari, S., Gaspar, R., Roberts, R., Villarreal, S., Chen, T., Zeng,
Z., Edvinsson, L., Salvatore, C., 2013a. Localization of CGRP
receptor components, CGRP and receptor binding sites in
rhesus brainstem: a detailed study using in situ hybridization
and autoradiography. Cephalalgia 2013, 33 (Supplement).
Eftekhari, S., Salvatore, C.A., Calamari, A., Kane, S.A., Tajti, J.,
Edvinsson, L., 2010. Differential distribution of calcitonin
gene-related peptide and its receptor components in the
human trigeminal ganglion. Neuroscience 169, 683–696.Eftekhari, S., Salvatore, C.A., Gaspar, R.C., Roberts, R., O’Malley, S.,
Zeng, Z., Edvinsson, L., 2013b. Localization of CGRP receptor
components, CGRP, and receptor binding sites in human and
rhesus cerebellar cortex. Cerebellum 12, 937–949.Eftekhari, S., Warfvinge, K., Blixt, F.W., Edvinsson, L., 2013c.
Differentiation of nerve fibers storing CGRP and CGRP receptors in
the peripheral trigeminovascular system. J. Pain 14, 1289–1303.Goadsby, P.J., 2007. Recent advances in understanding migraine
mechanisms, molecules and therapeutics. Trends Mol. Med.
13, 39–44.Goadsby, P.J., 2012. Pathophysiology of migraine. Ann. Indian
Acad. Neurol. 15, S15–S22.Goadsby, P.J., Edvinsson, L., 1993. The trigeminovascular system
and migraine: studies characterizing cerebrovascular and
neuropeptide changes seen in humans and cats. Ann. Neurol.
33, 48–56.Goadsby, P.J., Edvinsson, L., Ekman, R., 1988. Release of vasoactive
peptides in the extracerebral circulation of humans and the
cat during activation of the trigeminovascular system. Ann.
Neurol. 23, 193–196.Goadsby, P.J., Edvinsson, L., Ekman, R., 1990. Vasoactive peptide
release in the extracerebral circulation of humans during
migraine headache. Ann. Neurol. 28, 183–187.Gursoy-Ozdemir, Y., Qiu, J., Matsuoka, N., Bolay, H., Bermpohl, D.,
Jin, H.e.t., et al., 2004. Cortical spreading depression activates
and upregulates MMP-9. J. Clin. Invest. 113, 1447–1455.Hansen, J.M., Hauge, A.W., Olesen, J., Ashina, M., 2010. Calcitonin
gene-related peptide triggers migraine-like attacks in patients
with migraine with aura. Cephalalgia 30, 1179–1186.Ho, T.W., Edvinsson, L., Goadsby, P.J., 2010. CGRP and its receptors
provide new insights into migraine pathophysiology. Nat. Rev.
Neurol. 6, 573–582.Ho, T.W., Ferrari, M.D., Dodick, D.W., Galet, V., Kost, J., Fan, X.,
Leibensperger, H., Froman, S., Assaid, C., Lines, C., Koppen, H.,
Winner, P.K., 2008a. Efficacy and tolerability of MK-0974
(telcagepant), a new oral antagonist of calcitonin gene-related
peptide receptor, compared with zolmitriptan for acute
migraine: a randomised, placebo-controlled, parallel-
treatment trial. Lancet 372, 2115–2123.Ho, T.W., Mannix, L.K., Fan, X., Assaid, C., Furtek, C., Jones, C.J.,
Lines, C.R., Rapoport, A.M., 2008b. Randomized controlled trial
of an oral CGRP receptor antagonist, MK-0974, in acute
treatment of migraine. Neurology 70, 1304–1312.Hostetler, E.D., Joshi, A.D., Sanabria-Bohorquez, S., Fan, H., Zeng,
Z., Purcell, M., Gantert, L., Riffel, K., Williams, M., O’Malley, S.,
Miller, P., Selnick, H.G., Gallicchio, S.N., Bell, I.M., Salvatore, C.
A., Kane, S.A., Li, C.C., Hargreaves, R.J., de Groot, T., Bormans,
G., Van Hecken, A., Derdelinckx, I., de Hoon, J., Reynders, T.,
Declercq, R., De Lepeleire, I., Kennedy, W.P., Blanchard, R.,
Marcantonio, E.E., Sur, C., Cook, J.J., Van Laere, K., Evelhoch, J.
L., 2013. In vivo quantification of calcitonin gene-related
peptide receptor occupancy by telcagepant in rhesus monkey
and human brain using the positron emission tomography
tracer [11C]MK-4232. J. Pharmacol. Exp. Ther. 347, 478–486.Hou, M., Kanje, M., Longmore, J., Tajti, J., Uddman, R., Edvinsson, L.,
2001. 5-HT(1B) and 5-HT(1D) receptors in the human trigeminal
ganglion: co-localization with calcitonin gene-related peptide,
substance P and nitric oxide synthase. Brain Res. 909, 112–120.Joshi, P., Anderson, C., Binch, H., Hadida, S., Yoo, S., Bergeron, D.,
Decker, C., Terhaar, E., Moore, J., Garcia-Guzman, M., Termin,
A., 2014. Identification of potent CNS-penetrant
thiazolidinones as novel CGRP receptor antagonists. Bioorg.
Med. Chem. Lett. 24, 845–849.

b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 9108
Kai-Kai, M.A., Howe, R., 1991. Glutamate-immunoreactivity in the
trigeminal and dorsal root ganglia, and intraspinal neurons and
fibres in the dorsal horn of the rat. Histochem. J. 23, 171–179.Kaiser, E.A., Russo, A.F., 2013. CGRP and migraine: could PACAP
play a role too?. Neuropeptides 47, 451–461.Kaube, H., Hoskin, K.L., Goadsby, P.J., 1993. Inhibition by
sumatriptan of central trigeminal neurones only after blood–
brain barrier disruption. Br. J. Pharmacol. 109, 788–792.Kiernan, J.A., 1996. Vascular permeability in the peripheral
autonomic and somatic nervous systems: controversial
aspects and comparisons with the blood–brain barrier.
Microsc. Res. Tech. 35, 122–136.Lennerz, J.K., Ruhle, V., Ceppa, E.P., Neuhuber, W.L., Bunnett, N.W.,
Grady, E.F., Messlinger, K., 2008. Calcitonin receptor-like receptor
(CLR), receptor activity-modifying protein 1 (RAMP1), and
calcitonin gene-related peptide (CGRP) immunoreactivity in the rat
trigeminovascular system: differences between peripheral and
central CGRP receptor distribution. J. Comp. Neurol. 507, 1277–1299.Levy, D., Burstein, R., Strassman, A.M., 2005. Calcitonin gene-
related peptide does not excite or sensitize meningeal
nociceptors: implications for the pathophysiology of migraine.
Ann. Neurol. 58 (5), 698–705.Li, J., Vause, C.V., Durham, P.L., 2008. Calcitonin gene-related
peptide stimulation of nitric oxide synthesis and release from
trigeminal ganglion glial cells. Brain Res. 1196, 22–32.Liu Y, Broman J, Zhang M, Edvinsson L 2009. Brainstem and
thalamic projections from a craniovascular sensory nervous
centre in the rostral cervical spinal dorsal horn of rats.
Cephalalgia 29:935-948.Liu-Chen, L.Y., Liszczak, T.M., King, J.C., Moskowitz, M.A., 1986.
Immunoelectron microscopic study of substance P-containing
fibers in feline cerebral arteries. Brain Res. 369, 12–20.Limmroth, V., Katsarava, Z., Liedert, B., Guehring, H., Schmitz, K.,
Diener, H.C., Michel, M.C., 2001. An in vivo rat model to study
calcitonin gene related peptide release following activation of
the trigeminal vascular system. Pain 92, 101–106.Marin, J.C., Goadsby, P.J., 2010. Glutamatergic fine tuning with
ADX-10059: a novel therapeutic approach for migraine?.
Expert Opin. Investig. Drugs 19, 555–561.Merchenthaler, I., 1991. Neurons with access to the general
circulation in the central nervous system of the rat: a retrograde
tracing study with fluoro-gold. Neuroscience 44, 655–662.Moller, K., Zhang, Y.Z., Hakanson, R., Luts, A., Sjolund, B.,
Uddman, R., Sundler, F., 1993. Pituitary adenylate cyclase
activating peptide is a sensory neuropeptide:
immunocytochemical and immunochemical evidence.
Neuroscience 57, 725–732.Moore, E.L., Salvatore, C.A., 2012. Targeting a family B GPCR/
RAMP receptor complex: CGRP receptor antagonists and
migraine. Br. J. Pharmacol. 166, 66–78.Morris, J.L., Konig, P., Shimizu, T., Jobling, P., Gibbins, I.L., 2005.
Most peptide-containing sensory neurons lack proteins for
exocytotic release and vesicular transport of glutamate.
J. Comp. Neurol. 483, 1–16.Negro, A., Lionetto, L., Simmaco, M., Martelletti, P., 2012. CGRP
receptor antagonists: an expanding drug class for acute
migraine?. Expert Opin. Investig. Drugs 21, 807–818.Nozaki, K., Uemura, Y., Okamoto, S., Kikuchi, H., Mizuno, N., 1990.
Origins and distribution of cerebrovascular nerve fibers
showing calcitonin gene-related peptide-like
immunoreactivity in the major cerebral artery of the dog.
J. Comp. Neurol. 297, 219–226.Olesen, J., Diener, H.C., Husstedt, I.W., Goadsby, P.J., Hall, D.,
Meier, U., Pollentier, S., Lesko, L.M., 2004. Calcitonin gene-
related peptide receptor antagonist BIBN 4096 BS for the acute
treatment of migraine. N. Engl. J. Med. 350, 1104–1110.
Ottosson, A., Edvinsson, L., 1997. Release of histamine from duralmast cells by substance P and calcitonin gene-related peptide.Cephalalgia 17, 166–174.
Pascual, J., Munoz, P., 2005. Correlation between lipophilicity andtriptan outcomes. Headache 45, 3–6.
Raddant, A.C., Russo, A.F., 2011. Calcitonin gene-related peptidein migraine: intersection of peripheral inflammation andcentral modulation. Expert Rev. Mol. Med. 13, e36.
Rothrock, J.F., Mar, K.R., Yaksh, T.L., Golbeck, A., Moore, A.C., 1995.Cerebrospinal fluid analyses in migraine patients andcontrols. Cephalalgia 15, 489–493.
Salvatore, C.A., Kane, S.A., 2011. CGRP receptor antagonists:toward a novel migraine therapy. Curr. Pharm. Biotechnol. 12,1671–1680.
Salvatore, C.A., Moore, E.L., Calamari, A., Cook, J.J., Michener, M.S., O’Malley, S., Miller, P.J., Sur, C., Williams Jr., D.L., Zeng, Z.,Danziger, A., Lynch, J.J., Regan, C.P., Fay, J.F., Tang, Y.S., Li, C.C.,Pudvah, N.T., White, R.B., Bell, I.M., Gallicchio, S.N., Graham, S.L., Selnick, H.G., Vacca, J.P., Kane, S.A., 2010. Pharmacologicalproperties of MK-3207, a potent and orally active calcitoningene-related peptide receptor antagonist. J. Pharmacol. Exp.Ther. 333, 152–160.
Schytz, H.W., Birk, S., Wienecke, T., Kruuse, C., Olesen, J., Ashina,M., 2009. PACAP38 induces migraine-like attacks in patientswith migraine without aura. Brain 132, 16–25.
Seiler, K., Nusser, J.I., Lennerz, J.K., Neuhuber, W.L., Messlinger, K.,2013. Changes in calcitonin gene-related peptide (CGRP)receptor component and nitric oxide receptor (sGC)immunoreactivity in rat trigeminal ganglion followingglyceroltrinitrate pretreatment. J. Headache Pain 14, 74.
Sprenger, T., Goadsby, P.J., 2009. Migraine pathogenesis and stateof pharmacological treatment options. BMC Med. 7, 71.
Tajti, J., Uddman, R., Edvinsson, L., 2001. Neuropeptidelocalization in the “migraine generator” region of the humanbrainstem. Cephalalgia 21, 96–101.
Tajti, J., Uddman, R., Moller, S., Sundler, F., Edvinsson, L., 1999.Messenger molecules and receptor mRNA in the humantrigeminal ganglion. J. Auton. Nerv. Syst. 76, 176–183.
Ten Tusscher, M.P., Klooster, J., Vrensen, G.F., 1989. Satellite cellsas blood-ganglion cell barrier in autonomic ganglia. Brain Res.490, 95–102.
Tsai, S.H., Tew, J.M., McLean, J.H., Shipley, M.T., 1988. Cerebral arterialinnervation by nerve fibers containing calcitonin gene-relatedpeptide (CGRP): I. Distribution and origin of CGRP perivascularinnervation in the rat. J. Comp. Neurol. 271, 435–444.
Tuka, B., Helyes, Z., Markovics, A., Bagoly, T., Szolcsanyi, J., Szabo, N.,Toth, E., Kincses, Z.T., Vecsei, L., Tajti, J., 2013. Alterations inPACAP-38-like immunoreactivity in the plasma during ictal andinterictal periods of migraine patients. Cephalalgia 33, 1085–1095.
Uddman, R., Tajti, J., Hou, M., Sundler, F., Edvinsson, L., 2002.Neuropeptide expression in the human trigeminal nucleuscaudalis and in the cervical spinal cord C1 and C2. Cephalalgia22, 112–116.
Walker, C.S., Hay, D.L., 2013. CGRP in the trigeminovascularsystem: a role for CGRP, adrenomedullin and amylinreceptors? Br. J. Pharmacol, Dec 2013; 170(7): 1293–1307.
Vaudry, D., Falluel-Morel, A., Bourgault, S., Basille, M., Burel, D.,Wurtz, O., Fournier, A., Chow, B.K., Hashimoto, H., Galas, L.,Vaudry, H., 2009. Pituitary adenylate cyclase-activatingpolypeptide and its receptors: 20 years after the discovery.Pharmacol. Rev. 61, 283–357.
Vause, C.V., Durham, P.L., 2010. Calcitonin gene-related peptidedifferentially regulates gene and protein expression intrigeminal glia cells: findings from array analysis. Neurosci.Lett. 473, 163–167.
Vermeersch S.D., De Saint-Huber B., Derdelinckx I.P.K., Stekers M.S.K., Bormans G., Reynders T., Declercq R., De Lepeleire I.,Kennedy W.D., Blancard R., Marcantonio E., Hargreaves R.Li C.-

b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 9 3 – 1 0 9 109
C., Sanabria S., Hosteoer E., Joshi A.Evelhoch J.Van Laere K.,(2012). PET imaging in healty subjects and migraineurssuggests CGRP receptor antagonists do not have to actcentrally to achieve clinical efficacy. In: European Headacheand Migraine Trust International Congress, London, 20–23September 2012, Poster number LB08 2012.
Vikelis, M., Mitsikostas, D.D., 2007. The role of glutamate and itsreceptors inmigraine. CNS Neurol. Disord. Drug Targets 6, 251–257.
Xiao, Y., Richter, J.A., Hurley, J.H., 2008. Release of glutamate and
CGRP from trigeminal ganglion neurons: role of calcium
channels and 5-HT1 receptor signaling. Mol. Pain. 4, 12.Zhang, Z., Winborn, C.S., Marquez de Prado, B., Russo, A.F., 2007.
Sensitization of calcitonin gene-related peptide receptors by
receptor activity-modifying protein-1 in the trigeminal
ganglion. J. Neurosci. 27, 2693–2703.


















