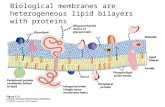
Lipid Bilayers and Membrane Dynamics: Insight into Thickness
Lipid Bilayers and Membrane Dynamics:
Insight into Thickness Fluctuation
Andrea C. Woodka,1 Paul D. Butler,1 Lionel Porcar,2 Bela Farago,2 and
Michihiro Nagao,1,2,3
1 NIST Center for Neutron Research, National Institute of Standards and Technology, Gaithersburg, MD 20899-6102, USA, 2 Institut Laue Langevin, 6 rue Jules Horowitz, BP 156-38042, Grenoble Cedex 9, France,
3 Center for Exploration of Energy and Matter, Indiana University, Bloomington, IN 47408, USA
Motivation
Lipid membranes are self-assembled highly flexible structures that have the ability to undergo an array of conformational and dynamic transitions which are essential for many biological functions.
Spectroscopic length scale:
Intermediate length scale : Membrane thickness fluctuations have been proposed as a mechanism for pore formation.(3)
Microscopic length scale:
Membrane stiffness and fluidity have been shown to have a large impact on cellular uptake and release.(2)
Cell signal transduction is affected by molecular lateral diffusion within the lipid membrane.(1)
(1) D Marguet, et al. EMBO J, 25, 3446 (2006)
(2) P. Weber, et. al. Adv Med Eng, 114, 377(2006)
(3) L. Movilenu, et al. Bull Math Biol, 58, 1231 (2006)
Membrane Dynamics
observation length scale
molecular motion collective motion
incoherent-quasi-elastic scattering
nuclear magnetic resonance etc.
coherent-quasi-elastic scattering
photon correlation spectroscopy
Molecular movements (incoherent movements of molecules)
Lateral diffusion Vertical vibration
Coherent movements of molecular assemblies
Rotation
Stretching Compression
Bending Stretching
Compression
Protrusion Thickness fluctuations
The Dynamics in Lipid Bilayers
Techniques to Measure Dynamics
Neutron Spin Echo
Elastic Scattering
http://www.ill.eu/instruments-support/instruments-groups/groups/tof/home/spin-echo-video/
Farago et al, Physica B 213&214, 712 (1995).; Farago, Physica B 226, 51 (1996).
• NSE measured Q
dependence of Deff • showing an increase at
the membrane thickness
length scale
Nagao, Phys. Rev. E 80, 031606 (2009).; Nagao et al., Soft Matter 7, 6598 (2011).; Nagao, J. Chem. Phys. 135, 074704 (2011).
bending
thickness
Surfactant Membranes
Breathing model of a lipid bilayer by Miller Miller, Top. Bioelectrochem. Bioenerg. 4, 161 (1981).; Bach and Miller, Biophys. J. 29, 183 (1980).; Miller, Biophys. J. 45, 643 (1984).
Amplitude of the fluctuations reaches ≈ 15 Å or more from
the geometrical constraints (volume conservation)
Thickness fluctuations by Hladky and Gruen
Thickness fluctuations occur, but the amplitude is small.
Long wavelength fluctuation amplitude is negligible
Short wavelength fluctuations (< 30 Å) are severely limited
Intermediate wavelength fluctuation amplitude < 10 Å
Hladky and Gruen, Biophys. J. 38, 251 (1982).
Deformation free energy of bilayer membranes by Huang Huang, Biophys. J. 50, 1061 (1986).
Theoretically, thickness fluctuations exist, their amplitude is very small
Thickness fluctuations in lipid
bilayers (theoretical studies)
Bilayer Preparation
(D54)DMPC = C14
(D62)DPPC = C16
(D70)DSPC = C18
Phospholipid Melting Temperature
1.112
1.110
1.108
1.106
1.104S
pecific
Gra
vity
70605040302010
Temperature (ºC)
DMPC DPPC DSPC
20.5ºC
37.5ºC
50.5ºC
Gel phase, below Tm
Above Tm lipid tails highly fluid, disordered and in constant motion
Fluid phase above Tm
Below Tm transition to a gel state, tails are fully extended with highly ordered packing
0.1
1
10
100
I(q)
0.012 3 4 5 6
0.12 3 4 5
q (A-1
)
DMPC_35C_ABS_i DPPC_50C_ABS_i DSPC_65C_ABS_i
Small Angle Neutron Scattering
Peak shifts to higher Q with decreasing tail
length
q ~ 2π / length scale
*Scattering intensity is offset to highlight peak shift
SANS and NSE are complimentary techniques
q
SANS static “snapshot” elastic scattering
NSE dynamic “snapshot”
quasielastic scattering
dqSqI ),()(
dtqStqI )cos(),(),(
1.0
0.8
0.6
0.4
I(q
,t)
/ I(
q,0
)
14121086420t (ns)
q (Å-1
)
0.056
0.070
0.075
0.113
0.132
0.164
0.183
0.201
fits
NSE : I(q,t)
])(exp[)0,(
),( 32 /tqI
tqI
1.0
0.8
0.6
0.4
0.2
14121086420
IQT_0p0456 IQT_0p0556 IQT_0p0651 IQT_0p0697 IQT_0p0750 IQT_0p0792 IQT_0p0845 IQT_0p0889 IQT_0p0935 IQT_0p0988 IQT_0p1031 IQT_0p1083 IQT_0p1126 IQT_0p1221 IQT_0p1228 IQT_0p1312 IQT_0p1323 IQT_0p1418 IQT_0p1514 IQT_0p1606 IQT_0p1635 IQT_0p1730
IQT_0p1826 IQT_0p1921 IQT_0p2013
1.0
0.9
0.8
0.7
0.6
0.5
14121086420
IQT_0p0615 IQT_0p0714 IQT_0p0815 IQT_0p0898 IQT_0p0909 IQT_0p0994 IQT_0p1004 IQT_0p1089 IQT_0p1184 IQT_0p1240 IQT_0p1275 IQT_0p1336 IQT_0p1431 IQT_0p1527 IQT_0p1618
DPPC 50ºC
DMPC 25ºC
DSPC 55ºC
Helfrich bending energy
Assuming the membrane is thin enough sheet, which is undulating
Zilman-Granek theory
Dynamics of a planar non-interacting Helfrich sheet
h(r)
k: bending modulus
h: solvent viscosity
I(q,t)
I(q,0)= exp - Gt( )
2 / 3[ ]
G = 0.025gkBT
k
æ
è ç
ö
ø ÷
1/ 2kBT
hq3
Watson-Brown theory
Extension of ZG theory including slipping of each monolayer
Inter-monolayer friction plays a role, where lateral
compressibility km of membrane
appears in dynamical equation
Zilman and Granek, Phys. Rev. Lett. 77, 4788 (1996).
Zilman and Granek, Chem. Phys. 284, 195 (2002).
Helfrich, Z. Natureforsch. 28, 693 (1973).
Watson and Brown, Biophys, J. 98, L09 (2010).
Evans and Yeung, Chem. Phys. Lipids. 73, 39 (1994).; Seifert and Langer, Europhys. Lett. 23, 71 (1993).
Surfactant Membrane Dynamic (bending)
Considering slipping friction
: decay rate, b=2/3
Zilman and Granek, Phys. Rev. Lett. 77, 4788 (1996).; Zilman and Granke,
Chem. Phys. 184, 195 (2002).
k: effective bending modulus,
h: solvent viscosity, a ≈ 1
~
Watson and Brown, Biophys. J. 98, L9 (2010).
Rawicz et al., Biophys. J. 79, 328 (2000).
Lee et al., Phys. Rev. Lett. 105, 038101 (2010).
Bending motion is explained as a single membrane dynamics model
Fitting I(q,t)
NSE : Γ vs. q DPPC @ 50ºC
32/1
23
025.0 qTkTk
OD
BBBEND
hk
4
6
0.001
2
4
6
0.01
2
4
6
0.1
(n
s-1)
3 4 5 6 7 8 9
0.12 3
q (A-1
)
6
80.1
2
4
6
81
2
4
6
810
I(q) (cm
-1)
q3
I(q)Γ deviates from the
pure q3 dependence expected from bending motions
This peak occurs at the same q as the SANS dip position
Bending
Thickness
22
0
333 )(1
1
qqqqqTFBEND
M. Nagao Zilman-Granek
NSE : Bending Modulus
Z. Yi, et. al, J. Phys. : Cond. Mater, 21, 155104 (2009).
Literature
Experiment
10
2
4
6
8100
2
4
6
81000
2
k (
kBT
)
100-10
T-Tm (ºC)
DMPC
DPPC
DSPC
● ■ ▲
q dependence of the decay rate:
Below vs. Above Tm
Below Tm DMPC
T = 16 ˚C
Bending
no enhancement =
suppressed thickness fluctuations
Bending
Enhancement =
thickness fluctuations Above Tm
DMPC
T = 35 ˚C
q dependence of the decay rate:
Lipid Tail Length
22
0
333 )(1
1
qqqqqTFBEND
NSE : Thickness fluctuations (Γ / q3)
accounts for bending motions damping frequency of thickness fluctuations
Proportional to the amplitude of thickness fluctuations
SANS dip position (Lorentzian peak position)
ΓBEND / q3 :
ΓTF :
ξ-1 :
q0 :
M. Nagao
40
30
20
10
0
/q
3 (
Å3/n
s)
0.200.150.100.05
q (Å-1
)
40
30
20
10
0
/q
3 (
Å3/n
s)
40
30
20
10
0
/q
3 (
Å3/n
s)
DMPC T=15ºC (NG5) T=16ºC (IN15) T=25ºC (NG5) T=35ºC (NG5) T=35ºC (IN15)
DPPC T=30ºC T=40ºC T=50ºC
DSPC T=45ºC T=55ºC T=65ºC
(b)
(c)
(d)
0.1
1
10
100
1000
I(q)
2 4
0.012 4
0.12 4
q (A-1
)
DPPC 30ºC 40ºC 50ºC
6
80.1
2
4I(
q)
0.250.200.150.100.05q (A
-1)
0.01 0.1
40
30
20
10
0
/q
3 (
Å3/n
s)
0.200.150.100.05
q (Å-1
)
40
30
20
10
0
/q
3 (
Å3/n
s)
40
30
20
10
0
/q
3 (
Å3/n
s)
DMPC T=15ºC (NG5) T=16ºC (IN15) T=25ºC (NG5) T=35ºC (NG5) T=35ºC (IN15)
DPPC T=30ºC T=40ºC T=50ºC
DSPC T=45ºC T=55ºC T=65ºC
(b)
(c)
(d)
0.1 0.15 0.2
40
30
20
10
0
/q
3 (
Å3/n
s)
0.200.150.100.05
q (Å-1
)
40
30
20
10
0
/q
3 (
Å3/n
s)
40
30
20
10
0
/q
3 (
Å3/n
s)
DMPC T=15ºC (NG5) T=16ºC (IN15) T=25ºC (NG5) T=35ºC (NG5) T=35ºC (IN15)
DPPC T=30ºC T=40ºC T=50ºC
DSPC T=45ºC T=55ºC T=65ºC
(b)
(c)
(d)
30
20
10
0
1000
100
10
1
0.1 0.1
1
10
100
I(q
)
0.012 3 4 5 6
0.12 3 4 5
q (A-1
)
DMPC_35C_ABS_i DPPC_50C_ABS_i DSPC_65C_ABS_i
Above Tm: independent of either
temperature or tail length
Membrane Thickness Fluctuations
Time scale
22
0
333 )(1
1
qqqqqTFBEND
An order of magnitude
slower (than surfactant
membranes)
≈ 100 ns
Width of Lorentzian peak relates to the fluctuation amplitude(1)
(1) Nagao et al., Soft Matter 7, 6598 (2011). (2) Huang, Biophys. J. 50, 1061 (1986). (3) Lindahl and Edholm, Biophys. J. 79, 426 (2000).
Mean amplitude = 3.7 Å ± 0.7 Å
Huang’s mean amplitude ≈ 4.5 Å(2)
Lindahl & Edholm’s amplitude ≈ 5 Å(3)
Experiment:
Theory:
Simulation:
≈ 8 % of the membrane thickness;
close to the value seen in surfactant
membranes (≈ 12 %)
Suggests amplitude is defined by physical constraints, like volume conservation
Membrane Thickness Fluctuations
Amplitude
Thickness Fluctuation Theory
21
121
2
0
1
21
2 tan2
tan2
CCCCCaK
TkD B
11
2aKC
2/1
1
22
Ka
BC
mdK
k1
adm 2
+ from NSE measurements ٭from SANS measurements # from literature
a0
Although membrane thickness fluctuations have not been previously measured Huang(7) has proposed a theory for thickness fluctuations in a lipid bilayer under the consideration of deformation free energy:
D ≈ 4.5Å
D B#
dm* k #
= thickness fluctuation amplitude = membrane compressibility = wavelength cut off = membrane thickness = bending modulus = surface tension
Conclusions
NSE was used to successfully measure lipid membrane thickness fluctuations
From SANS it is clear that these fluctuations appear at the length scale of the membrane thickness.
The relaxation time ≈100 ns and is independent of temperature and tail length. An order slower than that observed in surfactant membranes
Amplitude is ≈ 8 % of the thickness, consistent with surfactant membranes (12 %). Volume conservation may define the fluctuation amplitude.
Below Tm, thickness fluctuations are not observed, suggesting total suppression of the mode or much slower relaxation times which are not accessible by the current setup.
The experimental amplitude agrees well with both theory and simulation
FUTURE DIRECTION: What kind of effects do membrane associated molecules have on membrane dynamics such as thickness fluctuations?
Preliminary Data w/ Protein
Gramicidin
Alamethicin
http://rsif.royalsocietypublishing.org/content/early/2009/12/09/rsif.2009.0443/F4.large.jpghttp://rsif.royalsocietypublishing.org/content/early/2009/12/09/rsif.2009.0443/F4.large.jpg
Preliminary Data w/ Protein
45
40
35
30
25
20
15
k (
kBT
)
2.52.01.51.00.50.0
[Protein] mol%
DMPC(ILL)Ala/DMPCGram/DPPCGram/DMPC
12
10
8
6
4
2
0
T
F(x
10
-3n
s-1
)
2.01.00.0
[Protein] mol%
DMPC(ILL) Ala/DMPC Gram/DPPC Gram/DMPC
20
15
10
5
0
-1
(x10
-3Å
-1)
2.01.00.0
[Protein] mol%
DMPC(ILL) Ala/DMPC Gram/DPPC Gram/DMPC