
Learned Helplessness: Theory and Evidence · 30/06/1975 · Journal ol Experimental Psychology:...
44
\, Journal ol Experimental Psychology: General 1976, Vol. 105, No. 1, 3-46 Learned Helplessness: Theory and Evidence Steven F. Maier Martin E. P. Seligman University of Colorado University of Pennsylvania SUMMARY In 1967, Overmier and Seligman found that dogs exposed to inescapable and unavoidable electric shocks in one situation later failed to learn to escape shock in a different situation where escape was possible. Shortly thereafter Seligman and Maier (1967) demonstrated that this effect was caused by the uncontrollability of the original shocks. In this article we review the effects of exposing organisms to aversive events which they cannot control, and we review the explanations which have been offered. There seem to be motivational, cognitive, and emotional effects of uncontrol- lability. (a) Motivation. Dogs that have been exposed to inescapable shocks do not subsequently initiate escape response in the presence of shock. We review parallel phenomena in cats, fish, rats, and man. Of particular interest is the discussion of learned helplessness in rats and man. Rats are of interest because learned helplessness has been difficult to demonstrate in rats. However, we show that inescapably shocked rats do fail to learn to escape if the escape task is rea- sonably difficult. With regard to man, we review a variety of studies using in- escapable noise and unsolvable problems as agents which produce learned helpless- ness effects on both instrumental and cognitive tasks, (b) Cognition. We argue that exposure to uncontrollable events interferes with the organism's tendency to perceive contingent relationships between its behavior and outcomes. Here we review a variety of studies showing such a cognitive set. (c) Emotion. We re- view a variety of experiments which show that uncontrollable aversive events pro- duce greater emotional disruption than do controllable aversive events. We have proposed an explanation for these effects, which we call the learned helplessness hypothesis. It argues that when events are uncontrollable the organism learns that its behavior and outcomes are independent, and that this learning pro- duces the motivational, cognitive, and emotional effects of uncontrollability. We describe the learned helplessness hypothesis and research which supports it. Finally, we describe and discuss in detail alternative hypotheses which have been offered as accounts of the learned helplessness effect. One set of hypotheses argues that organisms learn motor responses during exposure to uncontrollable shock that compete with the response required in the test task. Another explanation holds that uncontrollable shock is a severe stressor and depletes a neurochemical necessary for the mediation of movement. We examine the logical structure of these explanations and present a variety of evidence which bears on them directly.
Transcript of Learned Helplessness: Theory and Evidence · 30/06/1975 · Journal ol Experimental Psychology:...

\,
Journal ol Experimental Psychology: General1976, Vol. 105, No. 1, 3-46
Learned Helplessness: Theory and Evidence
Steven F. Maier Martin E. P. SeligmanUniversity of Colorado University of Pennsylvania
SUMMARY
In 1967, Overmier and Seligman found that dogs exposed to inescapable andunavoidable electric shocks in one situation later failed to learn to escape shock ina different situation where escape was possible. Shortly thereafter Seligman andMaier (1967) demonstrated that this effect was caused by the uncontrollability ofthe original shocks. In this article we review the effects of exposing organismsto aversive events which they cannot control, and we review the explanationswhich have been offered.
There seem to be motivational, cognitive, and emotional effects of uncontrol-lability. (a) Motivation. Dogs that have been exposed to inescapable shocks donot subsequently initiate escape response in the presence of shock. We reviewparallel phenomena in cats, fish, rats, and man. Of particular interest is thediscussion of learned helplessness in rats and man. Rats are of interest becauselearned helplessness has been difficult to demonstrate in rats. However, we showthat inescapably shocked rats do fail to learn to escape if the escape task is rea-sonably difficult. With regard to man, we review a variety of studies using in-escapable noise and unsolvable problems as agents which produce learned helpless-ness effects on both instrumental and cognitive tasks, (b) Cognition. We arguethat exposure to uncontrollable events interferes with the organism's tendency toperceive contingent relationships between its behavior and outcomes. Here wereview a variety of studies showing such a cognitive set. (c) Emotion. We re-view a variety of experiments which show that uncontrollable aversive events pro-duce greater emotional disruption than do controllable aversive events.
We have proposed an explanation for these effects, which we call the learnedhelplessness hypothesis. It argues that when events are uncontrollable the organismlearns that its behavior and outcomes are independent, and that this learning pro-duces the motivational, cognitive, and emotional effects of uncontrollability. Wedescribe the learned helplessness hypothesis and research which supports it.
Finally, we describe and discuss in detail alternative hypotheses which have beenoffered as accounts of the learned helplessness effect. One set of hypotheses arguesthat organisms learn motor responses during exposure to uncontrollable shockthat compete with the response required in the test task. Another explanationholds that uncontrollable shock is a severe stressor and depletes a neurochemicalnecessary for the mediation of movement. We examine the logical structure ofthese explanations and present a variety of evidence which bears on them directly.

STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
What follows are three instances of thephenomenon to be explained:
1. When placed in a shuttle box an ex-perimentally naive dog, at the onset of thefirst electric shock, runs frantically about,until it accidentally scrambles over the bar-rier and escapes the shock. On the nexttrial, the dog, running frantically, crosses thebarrier more quickly than on the precedingtrial. Within a few trials the animal be-comes very efficient at escaping and soonlearns to avoid shock altogether. After about50 trials the dog becomes nonchalant andstands in front of the barrier. At the onsetof the signal for shock, the dog leaps grace-fully across and rarely gets shocked again.But dogs first given inescapable shock in aPavlovian hammock show a strikingly dif-ferent pattern. Such a dog's first reactionsto shock in the shuttle box are much thesame as those of a naive dog. He runsaround frantically for about 30 sec, but thenstops moving, lies down, and quietly whines.After 1 min. of this, shock terminates auto-matically. The dog fails to cross the barrierand escape from shock. On the next trial,the dog again fails to escape. At first hestruggles a bit and then, after a few seconds,seems to give up and passively accept theshock. On all succeeding trials, the dogcontinues to fail to escape.
2. A college student is confronted with aseries of 25 letter anagrams, each with thesame pattern, 34251. He has a little troublewith the first one, taking about 45 sec tosolve it. He solves each of the next threein about 30 sec, and now he sees the pattern.Each of the last 16 anagrams is solved im-mediately. In striking contrast is the collegestudent who has first faced a series of dis-crimination problems which are unsolvableor a series of loud tones which are inescap-able. He works hard on the first anagram,trying many rearrangements of letters, butfails to solve it in the 100 sec allowed. He
The preparation of this manuscript was supportedby Grant MH26827-01 to Steven F. Maier andGrant MH19604-01 to Martin E. P. Seligman.
Requests for reprints should be sent to StevenF. Maier, Department of Psychology, Universityof Colorado, Boulder, Colorado 80302.
fails to solve the second one also. The thirdanagram, a relatively easy one, he solves,after about 60 sec. He fails to solve thenext eight, appearing to give up after about60 sec with, each one. He then solves sixin a row, but very slowly, and finally sees thepattern. He now solves the final three im-mediately.
3. A naive rat is placed in a shuttle boxand trained to escape from shock. Shockterminates immediately upon his running tothe opposite side; he learns readily, andescapes efficiently. A second rat who hadreceived inescapable shock earlier in anotherapparatus learns just as well as the first ratto escape in the shuttle box. Now, how-ever, the contingency between crossing theshuttle box and shock termination is ob-fuscated ; shock does not terminate im-mediately upon crossing, but only after 3 secelapse. The first rat continues to escapereadily, learning to bridge a 3-sec delay ofreinforcement. The second rat, however,fails to respond; on other trials, he runsacross during shock, but overall he shows nolearning curve.
We believe these three phenomena are allinstances of "learned helplessness," instancesin which an organism has learned that out-comes are uncontrollable; by his responsesand is seriously debilitated by this knowl-edge. This article explores the evidence forthe phenomenon of learned helplessness, anddiscusses a variety of theoretical interpreta-tions. Since the phenomenon results fromexperience with uncontrollable outcomes, webegin by defining uncontrollability.
TJJNCONTROLLABILITY AND THE INSTRU-! MENTAL TRAINING SPACE
llearning theorists have usually viewedthe I relations between instrumental respond-ing! and outcomes to which organisms aresensitive in terms of the _ conditional prob-ability of an outcome or reinforcer followinga response />(RF/R), which can have valuesranging from 0 to 1.0. At 1.0, every re-sponse produces a reinforcer or outcome(continuous reinforcement). At- 0, a re-sponse never produces a reinforcer (extinc-tion). Intermediate values represent vari-ous degrees of partial reinforcement.
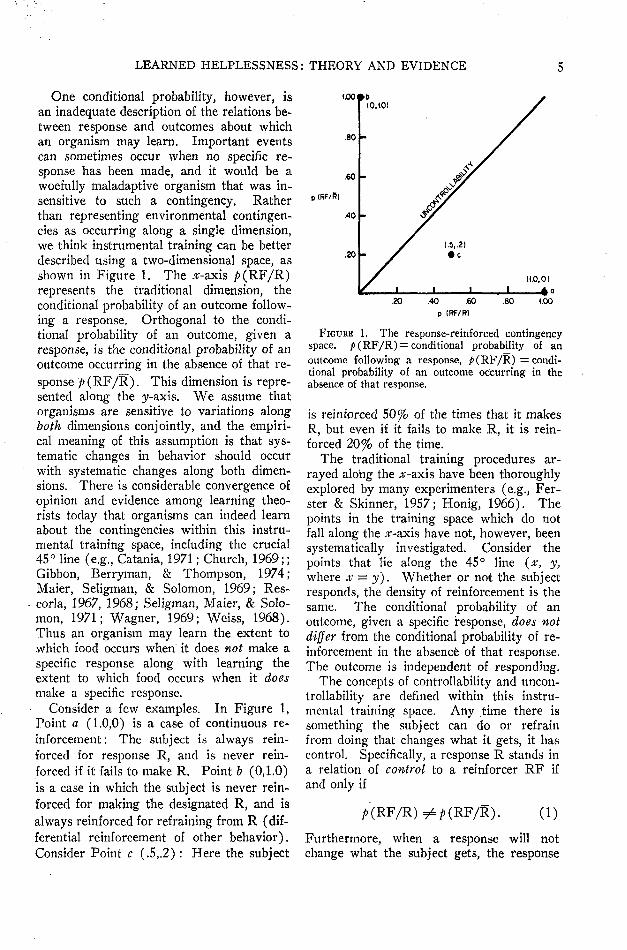
LEARNED HELPLESSNESS: THEORY AND EVIDENCE
One conditional probability, however, isan inadequate description of the relations be-tween response and outcomes about whichan organism may learn. Important eventscan sometimes occur when no specific re-sponse has been made, and it would be awoefully maladaptive organism that was in-sensitive to such a contingency. Ratherthan representing environmental contingen-cies as occurring along a single dimension,we think instrumental training can be betterdescribed using a two-dimensional space, asshown in Figure 1. The .ar-axis />(RF/R)represents the traditional dimension, theconditional probability of an outcome follow-ing a response. Orthogonal to the condi-tional probability of an outcome, given aresponse, is the conditional probability of anoutcome occurring in the absence of that re-sponse />(RF/R). This dimension is repre-sented along the y-axis. We assume thatorganisms are sensitive to variations alongboth dimensions conjointly, and the empiri-cal meaning of this assumption is that sys-tematic changes in behavior should occurwith systematic changes along both dimen-sions. There is considerable convergence ofopinion and evidence among learning theo-rists today that organisms can indeed learnabout the contingencies within this instru-mental training space, including the crucial45° line (e.g., Catania, 1971; Church, 1969;;Gibbon, Berryman, & Thompson, 1974;Maier, Seligman, & Solomon, 1969; Res-corla, 1967, 1968; Seligman, Maier, & Solo-mon, 1971; Wagner, 1969; Weiss, 1968).Thus an organism may learn the extent towhich food occurs when it does not make aspecific response along with learning theextent to which food occurs when it doesmake a specific response.
Consider a few examples. In Figure 1,Point a (1.0,0) is a case of continuous re-inforcement : The subject is always rein-forced for response R, and is never rein-forced if it fails to make R. Point b (0,1.0)is a case in which the subject is never rein-forced for making the designated R, and isalways reinforced for refraining from R (dif-ferential reinforcement of other behavior).Consider Point c (.5,.2): Here the subject
p (RF/R)
.40 -
.20 -
(.00
FIGURE 1. The response-reinforced contingencyspace. p( RF/R) = conditional probability of anoutcome following a response, />(RF/R) = condi-tional probability of an outcome occurring in theabsence of that response.
is reinforced 50% of the times that it makesR, but even if it fails to make R, it is rein-forced 20% of the time.
The traditional training procedures ar-rayed along the #-axis have been thoroughlyexplored by many experimenters (e.g., Fer-ster & Skinner, 1957; Honig, 1966). Thepoints in the training space which do notfall along the #-axis have not, however, beensystematically investigated. Consider thepoints that lie along the 45° line (x, y,where x = y). Whether or not the subjectresponds, the density of reinforcement is thesame. The conditional probability of anoutcome, given a specific response, does notdiffer from the conditional probability of re-inforcement in the absence of that response.The outcome is independent of responding.
The concepts of controllability and uncon-trollability are defined within this instru-mental training space. Any time there issomething the subject can do or refrainfrom doing that changes what it gets, it hascontrol. Specifically, a response R stands ina relation of control to a reinforcer RF ifand only if
/» (RF/R) ^# (RF/R). (1)
Furthermore, when a response will notchange what the subject gets, the response
STEVEN F. MAIER AND MARTIN E. P. SELIGMAN,
and reinforcer are independent. Specifically,a response R stands in relation of independ-ence to a reinforcer RF if and only if
/>(RF/R) = (2)
When this is true of all emitted responses(as in Pavlovian conditioning) the subjectcannot control the reinforcer, and the rein-forcer is defined as uncontrollable.
How can we tell that the phenomena wewill discuss result from experiencing uncon-trollable outcomes as opposed to merely ex-periencing the outcome itself? To put itanother way, how can we tell whether help-lessness is a psychological phenomenon asopposed to merely being the result of physi-cal stimulation?
There is a simple and elegant experimentaldesign which isolates the effects of con-trollability from the effects of the outcomebeing controlled. In this "triadic" design,three groups are used: One group receivesas its pretreatment an outcome that it cancontrol by some response. A second groupis "yoked"—it receives exactly the samephysical outcome as its counterpart in thefirst group, but there is no response theyoked subject can make which modifies theseoutcomes. A third group receives no pre-treatment. Later, all groups are tested on anew task.
Helplessness does not result from traumaper se: In the studies we discuss, deficitsdo not occur in the groups that controlshock, but only in the yoked group (Hiroto& Seligman, 1975; Maier, 1970; Maier,Anderson, & Lieberman, 1972; Seligman &Beagley, 1975; Seligman & Maier, 1967).1
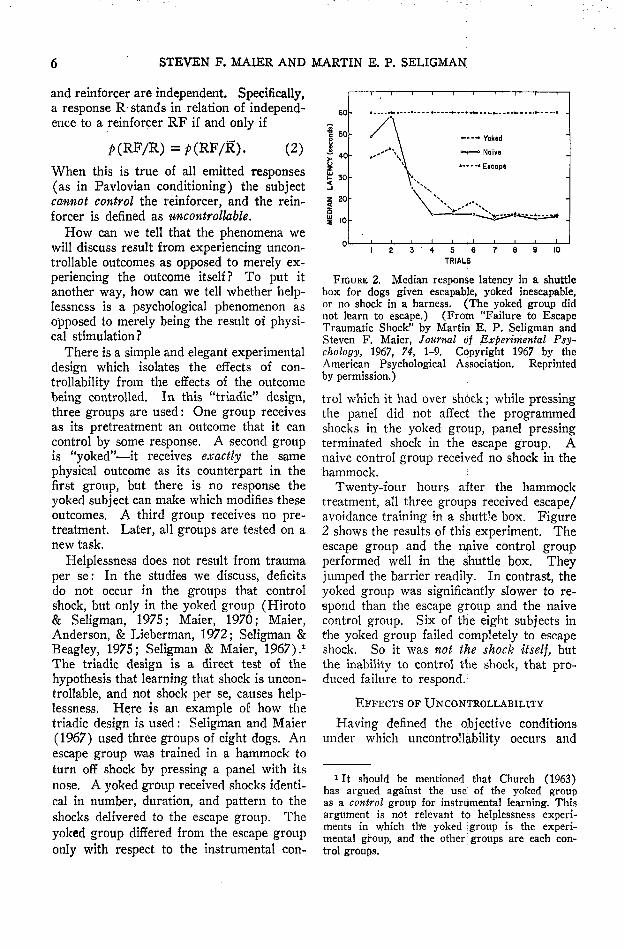
The triadic design is a direct test of thehypothesis that learning that shock is uncon-trollable, and not shock per se, causes help-lessness. Here is an example of how thetriadic design is used: Seligman and Maier(1967) used three groups of eight dogs. Anescape group was trained in a hammock toturn off shock by pressing a panel with itsnose. A yoked group received shocks identi-cal in number, duration, and pattern to theshocks delivered to the escape group. Theyoked group differed from the escape grouponly with respect to the instrumental con-
4 5 6TRIALS
FIGURE 2. Median response latency in a shuttlebox for dogs given escapable, yoked inescapable,or no shock in a harness. (The yoked group didnot learn to escape.) (From "Failure to EscapeTraumatic Shock" by Martin E. P. Seligman andSteven F. Maier, Journal of Experimental Psy-chology, 1967, 74, 1-9, Copyright 1967 by theAmerican Psychological Association. Reprintedby permission.)
trol which it had over shdck; while pressingthe panel did not affect the programmedshocks in the yoked group, panel pressingterminated shock in the escape group. Anaive control group received no shock in thehammock. :-
Twenty-four hours after the hammocktreatment, all three groups received escape/avoidance training in a shuttle box. Figure2 shows the results of this experiment. Theescape group and the naive control groupperformed well in the shuttle box. Theyjumped the barrier readily. In contrast, theyoked group was significantly slower to re-spond than the escape group and the naivecontrol group. Six of the eight subjects inthe yoked group failed completely to escapeshock. So it was not the shock itself, butthe inability to control the shock, that pro-duced failure to respond.1
EFFECTS OF UNCONTEOLLABILITY
Having defined the objective conditionsunder which uncontrollability occurs and
lit should be mentioned: that Church (1963)has argued against the use of the yoked groupas a control group for instrumental learning. Thisargument is not relevant to helplessness experi-ments in which the yoked ;group is the experi-mental group, and the other groups are each con-trol groups.

LEARNED HELPLESSNESS: THEORY AND EVIDENCE
delineated the kind of experimental designwhich isolates the effects of uncontrollabilityfrom the effects of stimulation per se, we nowreview the deficits produced by uncontrol-lable outcomes. In general, when an organ-ism experiences uncontrollable events, threedeficits often ensue: motivational, cognitive,and emotional.
a) The motivation to respond in the faceof later aversive events seems to wane, b)Moreover, even if the subject does respondand the response succeeds in producing re-lief, the subject often has difficulty learningthat the response worked, c) Finally, emo-tional balance may be distributed; depressionand anxiety, measured in a variety of ways,may predominate. The motivational deficitsproduced by helplessness are in many waysthe most striking, so we turn to them firstfor review and analysis.
Motivational Deficits
Dogs. These sets of experiments form thebase of the pyramid on which we constructa theory of learned helplessness, so thesestudies will be examined thoroughly. Thebehavior of dogs exposed to inescapableshock seems to typify what many species dowhen they are faced with uncontrollability.
Here is the typical procedure that pro-duces learned helplessness in the dog (Over-mier, 1968; Overmier & Seligman, 1967;Seligman & Groves, 1970; Seligman &Maier, 1967; Seligman, Maier, & Geer,1968). On the first day, the subject isstrapped into a hammock and given 64 in-escapable electric shocks, each 5.0 sec longand of 6.0 mA (moderately painful) in-tensity. The shocks are not predicted by anysignal and they occur randomly in time.Twenty-four hours later, the subject is given10 trials of signalized escape/avoidance train-ing in a two-way shuttle box. The dogmust jump over the barrier from one com-partment into the other to escape or avoidshock. Shocks can occur in either com-partment, so there is no place that is alwayssafe, but the response of shuttling or jump-ing always leads to shock termination.
The onset of a signal (light dimming) be-gins each trial, and the signal stays on untilthe trial ends. The interval between the start
of the signal and the shock is 10 sec. If thedog jumps the shoulder-high barrier duringthis interval, the signal terminates and shockis prevented. Failure to jump during thesignal-shock interval leads to a 4.5-mAshock which remains until .the dog jumps thebarrier. If the dog fails to jump the barrierwithin 60 sec after signal onset, the trialautomatically ends.
Between the years 1965 and 1969 the be-havior of about 150 dogs that received priorinescapable shock was studied. Of these,-two thirds (about 100) did not learn toescape and went through the striking se-quence of behaviors that we described. Theother one third seemed completely normal.Like naive dogs, they escaped efficiently.There was no intermediate outcome. In-terestingly enough, of the several hundrednaive dogs who had been given shuttle boxtraining, about 95% have been efficient re-sponders. The other 5% failed to learneven with no prior inescapable shock. Webelieve that the prior history of these dogsbefore they arrived at the laboratory maydetermine whether a naive dog will show alearned helplessness effect and whether adog given inescapable shock will be immuneto this effect. When we discuss the theoryand prevention of the learned helplessnesseffect below, we will be more explicit abouthow to immunize against failure to escape.
Since dogs exposed to inescapable shockseem to be physically capable of jumping thebarrier, the behavioral deficit must have apsychological base. Occasionally, they jumpthe barrier between trials. Further, if a doghas been sitting and taking shock after shockon the left side of the box, and the door onthe right side is opened at the end of thesession, he will often come bounding acrossto escape from the box altogether
The learned helplessness effect in the dogoccurs in a variety of situations and is easilyproduced. Within limits, it does not de-pend on the use of any particular shockparameters. We have varied the frequency,intensity, duration, and temporal pattern ofshocks, and still produced the effect. Further-more, it does not matter if the inescapableshock is or is not preceded by a signal.Finally, it does not matter what apparatus

STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
the inescapable shocks are given in or wherethe escape/avoidance training takes place.The shuttle box and hammock are inter-changeable. If the dog is given inescapableshock in the shuttle box and then requiredto press panels with its head to escape inthe hammock, he is still helpless. Further,after uncontrollable shock, dogs are not onlydebilitated at fleeing from the shock itself(escape), but they also seem to be unableto prevent or "avoid" shock. Overmier(1968) gave dogs inescapable shock in thehammock and then tested them in a shuttlebox. If the dog jumped after the signalwent on, but before the shock came on, hecould avoid the shock. But no escape wasallowed, for if the dog failed to jump in thesignal-shock interval, the barrier was closedand inescapable shock occurred. The help-less dogs failed to avoid, just as they hadfailed to escape. So, dogs previously ex-posed to inescapable shocks failed to copeadaptively with signals for shock as well aswith shock.
Debilitation of response initiation as a con-sequence of uncontrollable outcomes hasbeen reported in cats, rats, mice, birds, pri-mates, fish, and man, as well as in dogs.The learned helplessness effect seems rathergeneral among species that learn.
Cats. Thomas and Baiter (in press) re-ported an effect in cats which seems identicalto learned helplessness in dogs. (See alsoMasserman, 1943, 1971; Seward & Hum-phrey, 1967; Zielinski & Soltysik, 1964, forother reports of debilitation in cats caused byinescapable shock). They designed a ham-mock for cats and gave them inescapableshock. When later placed in a cat shuttlebox, those cats failed to escape. Like dogs,they sat and took the shock. Thomas andBaiter also reported that blocking activity ofthe septal area of the brain breaks up help-lessness, and directly stimulating the septumelectrically produces a learned helplessness-like effect. When we discuss the theory andtherapy for helplessness, we will examinethese physiological data in more detail.
Fish. Following inescapable shock, fishalso show poor escape and avoidance re-sponding. Padilla, Padilla, Ketterer, andGiacolone (1970) gave inescapable shock to
goldfish and then tested them in an aquaticshuttle box. These fish were slower toavoid than naive controls. (See Behrend& Bitterman, 1963; Bintz, 1971; Frumkin& Brookshire, 1969; Padilla, 1973, for re-lated goldfish data.)
Rats. Until recently, it has proven dif-ficult to produce a learned helplessness effectin rats. A substantial number of experi-ments were performed involving inescapableshock; by and large, however, these showedrather small, if any, effects on later responseinitiation. (Maier et al., 1969, and Selig-man et al., 1971, reviewed the complicatedliterature, and the interested reader is re-ferred there for details. See also Anderson,Cole, & McVaugh, 1968; de Toledo & Black,1967; Dinsmoor & Campbell, 1965a, 1965b;Looney & Cohen, 1972; Mullin & Mogen-son, 1963; Weiss, Krieckhaus, & Conte,1968, for representative experimentalstudies.) Unlike dogs, a rat given priorinescapable shock was typically only a bitslower to escape shock on the first few trials,or slower to acquire avoidance—it did notfail to learn.
After intensive experimentation, however,several investigators have :now independentlyproduced substantial learned helplessness ef-fects in rats. (Maier, Albin, & Testa, 1973;Maier & Testa, 1975; Seligman & Beagley,1975; Seligman, Rosellini, & Kozak, 1975.)«In doing so, one crucial factor emerged—theresponse used in the test for learned help-lessness must be difficult, and not somethingthe rat does very readily. So, for example,if rats are first exposed to inescapable shockand then tested on a simple escape responselike pressing a bar once (FR-1), or fleeingto the other side of a shuttle box, no deficitsare found. If, however, the response re-quirement is increased—so the bar must bepressed three times in order for shock toend (FR-3 lever pressing) or the rat has torun across the shuttle box and back (FR-2shuttling)—then the rat that has experiencedinescapable shock fails to escape. In con-
2 In passing it should be mentioned that micehave been reported to show response deficits fol-lowing inescapable shock (Braud, Wepman, &Russo, 1969).
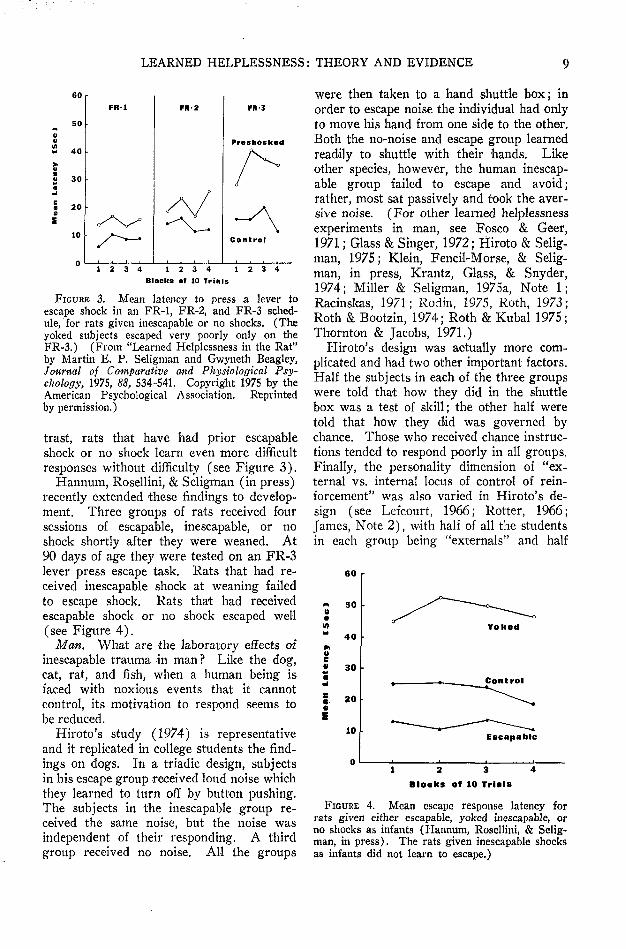
LEARNED HELPLESSNESS: THEORY AND EVIDENCE 9
60
50
40
30
20
10
FR-2 FR-3
Preshocked
1 2 3 4 1 2 3 4 1 2 3 4Blocks of 10 Trials
FIGURE 3. Mean latency to press a lever toescape shock in an FR-1, FR-2, and FR-3 sched-ule, for rats given inescapable or no shocks. (Theyoked subjects escaped very poorly only on theFR-3.) (From "Learned Helplessness in the Rat"by Martin E. P. Seligman and Gwyneth Beagley,Journal of Comparative and Physiological Psy-chology, 1975, 88, 534-541. Copyright 1975 by theAmerican Psychological Association. Reprintedby permission.)
trast, rats that have had prior escapableshock or no shock learn even more difficultresponses without difficulty (see Figure 3).
Hannum, Rosellini, & Seligman (in press)recently extended these findings to develop-ment. Three groups of rats received foursessions of escapable, inescapable, or noshock shortly after they were weaned. At90 days of age they were tested on an FR-3lever press escape task. Rats that had re-ceived inescapable shock at weaning failedto escape shock. Rats that had receivedescapable shock or no shock escaped well(see Figure 4).
Man. What are the laboratory effects ofinescapable trauma in man? Like the dog,cat, rat, and fish, when a human being isfaced with noxious events that it cannotcontrol, its motivation to respond seems tobe reduced.
Hiroto's study (1974) is representativeand it replicated in college students the find-ings on dogs. In a triadic design, subjectsin his escape group received loud noise whichthey learned to turn off by button pushing.The subjects in the inescapable group re-ceived the same noise, but the noise wasindependent of their responding. A thirdgroup received no noise. All the groups
were then taken to a hand shuttle box; inorder to escape noise the individual had onlyto move his hand from one side to the other.Both the no-noise and escape group learnedreadily to shuttle with their hands. Likeother species, however, the human inescap-able group failed to escape and avoid;rather, most sat passively and took the aver-sive noise. (For other learned helplessnessexperiments in man, see Fosco & Geer,1971; Glass & Singer, 1972; Hiroto & Selig-man, 1975; Klein, Fencil-Morse, & Selig-man, in press, Krantz, Glass, & Snyder,1974; Miller & Seligman, 1975a, Note 1;Racinskas, 1971; Rodin, 1975, Roth, 1973;Roth & Bootzin, 1974; Roth & Kubal 1975;Thornton & Jacobs, 1971.)
Hiroto's design was actually more com-plicated and had two other important factors.Half the subjects in each of the three groupswere told that how they did in the shuttlebox was a test of skill; the other half weretold that how they did was governed bychance. Those who received chance instruc-tions tended to respond poorly in all groups.Finally, the personality dimension of "ex-ternal vs. internal locus of control of rein-forcement" was also varied in Hiroto's de-sign (see Lefcourt, 1966; Rotter, 1966;James, Note 2), with half of all the studentsin each group being "externals" and half
60 r
- SOuv
40
30
20
10
Yoked
Control
Escapable
1 2 3 4Blocks of 10 Trials
FIGURE 4. Mean escape response latency forrats given either escapable, yoked inescapable, orno shocks as infants (Hannum, Rosellini, & Selig-man, in press). The rats given inescapable shocksas infants did not learn to escape.)
10 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
"internals." An external is a person whobelieves, as shown by his answers on a per-sonality inventory, that reinforcements occurin his life by chance or luck and are beyondhis control. An internal believes that hecontrols his own reinforcers and that skillwill out. Hiroto found that externals be-came helpless in his experiment more easilythan did internals (see also Dweck, 1975).So three independent factors, the laboratoryprocedure of uncontrollability, the cognitiveset induced by chance instructions, and ex-ternality, each produced a learned helpless-ness effect. Given this convergence, Hirotoconcluded that these three factors all erodemotivation to escape by contributing to theexpectation that responding and relief areindependent.
This concludes the survey of motivationaldeficits produced by exposure to uncontrol-lable aversive events in different species. Itseems to be generally true that uncontrol-lability produces deterioration in the readi-ness of dogs, cats, rats, fish, monkeys, andmen to respond actively to trauma.
Generality. Does uncontrollability pro-duce a habit limited to situations very likethe ones in which uncontrollability is experi-enced, or does it produce a more general ef-fect ? The question we are really asking hereis "Is helplessness just an isolated set ofhabits or does it involve a more basicchange?" We believe that what is learnedwhen the environment is uncontrollable canhave consequences for a wide range of be-havior.
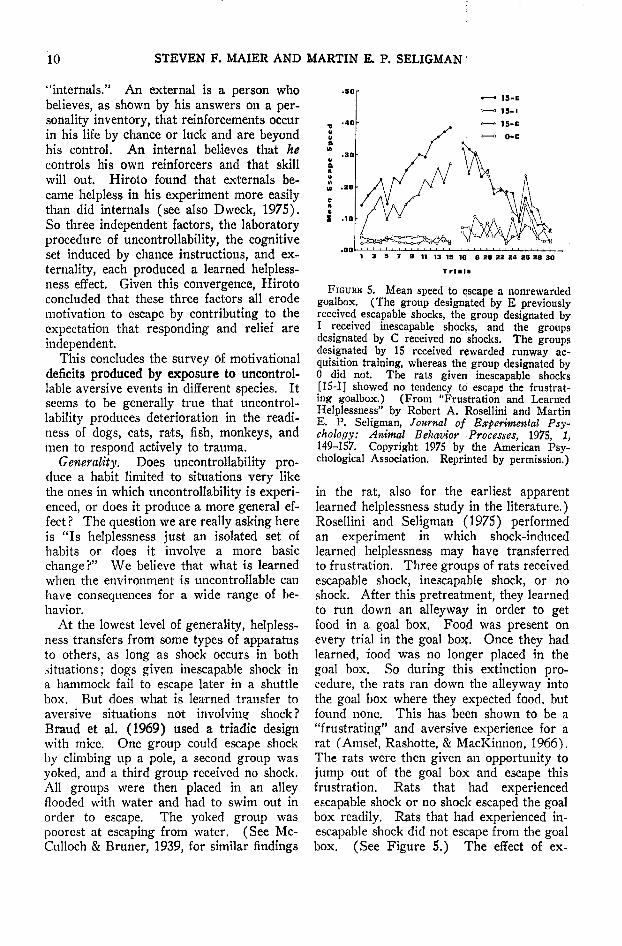
At the lowest level of generality, helpless-ness transfers from some types of apparatusto others, as long as shock occurs in both.situations; dogs given inescapable shock ina hammock fail to escape later in a shuttlebox. But does what is learned transfer toaversive situations not involving shock?Braud et al. (1969) used a triadic designwith mice. One group could escape shockby climbing up a pole, a second group wasyoked, and a third group received no shock.All groups were then placed in an alleyflooded with water and had to swim out inorder to escape. The yoked group waspoorest at escaping from water, (See Mc-Culloch & Bruner, 1939, for similar findings
1 3 5 7 13 IS 16 8 2O 22 2« 26 28 30
T r i a l s
FIGURE S. Mean speed to escape a nonrewardedgoalbox. (The group designated by E previouslyreceived escapable shocks, the group designated byI received inescapable shocks, and the groupsdesignated by C received no shocks. The groupsdesignated by IS received rewarded runway ac-quisition training, whereas the group designated by0 did not. The rats .given inescapable shocks[15-1] showed no tendency to escape the frustrat-ing goalbox.) (From "Frustration and LearnedHelplessness" by Robert A. Rosellini and MartinE. P. Seligman, Journal of Experimental Psy-chology: Animal Behavior Processes, 197S, 1,149-157. Copyright 1975 by the American Psy-chological Association. Reprinted by permission.)
in the rat, also for the earliest apparentlearned helplessness study in the literature.)Rosellini and Seligman (1975) performedan experiment in which shock-inducedlearned helplessness may have transferredto frustration. Three groups of rats receivedescapable shock, inescapable shock, or noshock. After this pretreatment, they learnedto run down an alleyway in order to getfood in a goal box. Food was present onevery trial in the goal box. Once they hadlearned, food was no longer placed in thegoal box. So during this extinction pro-cedure, the rats ran down the alleyway intothe goal box where they expected food, butfound none. This has been shown to be a"frustrating" and aversive experience for arat (Amsel, Rashotte, & MacKinnon, 1966).The rats were then given an opportunity tojump out of the goal box and escape thisfrustration. Rats that had experiencedescapable shock or no shock escaped the goalbox readily. Rats that had experienced in-escapable shock did not escape from the goalbox. (See Figure 5.) The effect of ex-
LEARNED HELPLESSNESS: THEORY AND EVIDENCE 11
40-1
XO
30H
u 20"CONFINED^-*ESCAPED =YOKED* •
1 2 3 4 5
SESSIONS
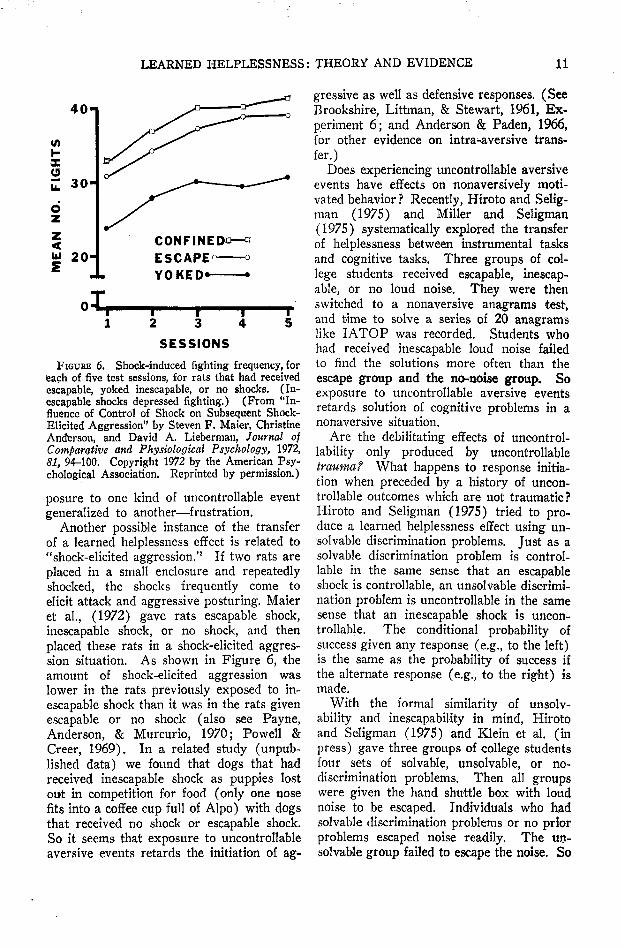
FIGURE 6. Shock-induced fighting frequency, foreach of five test sessions, for rats that had receivedescapable, yoked inescapable, or no shocks. (In-escapable shocks depressed fighting.) (From "In-fluence of Control of Shock on Subsequent Shock-Elicited Aggression" by Steven F. Maier, ChristineAnderson, and David A. Lieberman, Journal ofComparative and Physiological Psychology, 1972,SI, 94-100. Copyright 1972 by the American Psy-chological Association. Reprinted by permission.)
posure to one kind of uncontrollable eventgeneralized to another—frustration.
Another possible instance of the transferof a learned helplessness effect is related to"shock-elicited aggression.'' If two rats areplaced in a small enclosure and repeatedlyshocked, the shocks frequently come toelicit attack and aggressive posturing. Maieret al., (1972) gave rats escapable shock,inescapable shock, or no shock, and thenplaced these rats in a shock-elicited aggres-sion situation. As shown in Figure 6, theamount of shock-elicited aggression waslower in the rats previously exposed to in-escapable shock than it was in the rats givenescapable or no shock (also see Payne,Anderson, & Murcurio, 1970; Powell &Creer, 1969). In a related study (unpub-lished data) we found that dogs that hadreceived inescapable shock as puppies lostout in competition for food (only one nosefits into a coffee cup full of Alpo) with dogsthat received no shock or escapable shock.So it seems that exposure to uncontrollableaversive events retards the initiation of ag-
gressive as well as defensive responses. (SeeBrookshire, Littman, & Stewart, 1961, Ex-periment 6; and Anderson & Paden, 1966,for other evidence on intra-aversive trans-fer.)
Does experiencing uncontrollable aversiveevents have effects on nonaversively moti-vated behavior ? Recently, Hiroto and Selig-man (1975) and Miller and Seligman(1975) systematically explored the transferof helplessness between instrumental tasksand cognitive tasks. Three groups of col-lege students received escapable, inescap-able, or no loud noise. They were thenswitched to a nonaversive anagrams test,and time to solve a series of 20 anagramslike IATOP was recorded. Students whohad received inescapable loud noise failedto find the solutions more often than theescape group and the no-noise group. Soexposure to uncontrollable aversive eventsretards solution of cognitive problems in anonaversive situation.
Are the debilitating effects of uncontrol-lability only produced by uncontrollabletrauma? What happens to response initia-tion when preceded by a history of uncon-trollable outcomes which are not traumatic?Hiroto and Seligman (1975) tried to pro-duce a learned helplessness effect using un-solvable discrimination problems. Just as asolvable discrimination problem is control-lable in the same sense that an escapableshock is controllable, an unsolvable discrimi-nation problem is uncontrollable in the samesense that an inescapable shock is uncon-trollable. The conditional probability ofsuccess given any response (e.g., to the left)is the same as the probability of success ifthe alternate response (e.g., to the right) ismade.
With the formal similarity of unsolv-ability and inescapability in mind, Hirotoand Seligman (1975) and Klein et al. (inpress) gave three groups of college studentsfour sets of solvable, unsolvable, or no-discrimination problems. Then all groupswere given the hand shuttle box with loudnoise to be escaped. Individuals who hadsolvable discrimination problems or no priorproblems escaped noise readily. The un-solvable group failed to escape the noise. So

12 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
we believe that response initiation to con-trol noxious events may be impaired by ex-perience with uncontrollable reward.
Equally intriguing is the possibility thatdelivery of appetitive events independent ofbehavior may produce a failure to learn re-sponses to procure such events, an effectanalogous to learned helplessness. Engberg,Hansen, Welker, and Thomas (1973)trained one group of pigeons to press atreadle to obtain food. A second group ofpigeons received food delivered indepen-dently of behavior, and a third group re-ceived only a small amount of magazinetraining. Following this pretreatment, allpigeons were tested with a key peck auto-shaping procedure. In autoshaping (Brown& Jenkins, 1968) the pigeon key is periodi-cally illuminated for a brief period of time,and food is presented at the end of each keyillumination period whether or not a peckhas occurred. Pigeons normally acquirekey pecking under such an arrangement.The result was that the group given previoustreadle press training was fastest to acquirekey pecking and the group given noncon-tingent food was the slowest to acquire keypecking. Engberg et al. interpreted theirresults as constituting an appetitive analogueto learned helplessness; food was deliveredindependently of behavior, and this retardedthe acquisition of a response to procure food.
However, the Engberg et al. experimenthas been criticized on a number of grounds(Gamzu, Williams, & Schwartz, 1973).First, Gamzu et al. argued that autoshapingis not a reasonable task to use in attemptingto establish an appetitive analogue to learnedhelplessness because autoshaped key pecksare largely controlled by Pavlovian con-tingencies (Moore, 1973) rather than instru-mental contingencies; learned helplessness isa debilitation of instrumental responding.Second, Gamzu et al. noted that it is easyto account for the Engberg et al. experimentin terms of competing motor responses.Both the group given treadle training andthe group given noncontingent food wouldhave been expected to learn a motor re-sponse incompatible with key pecking(treadle responding and standing by thegrain hopper, respectively). However, Eng-
berg et al. removed the treadle during theautoshaping test, and so it could be arguedthat the stimulus controlling the competingbehavior was removed for the treadle group.The stimulus controlling any competing be-havior was not removed for the noncontin-gent food group, and this difference couldaccount for the results. In response tothese criticisms Welker (1974) conducteda further investigation of the phenomenon.One group of pigeons was trained to pecka key for food, a second was given non-contingent food, and a third was given onlya small amount of magazine training. Allpigeons were then given a test in whichtreadle pressing produced food on a con-tinuous reinforcement schedule. The keywas present during the treadle test, so itcould not be argued that the stimulus con-trolling the competing behavior was re-moved for the contingent food group. Theresults were the same as in Engberg et al.;noncontingent food presentation retarded ac-quisition. Taking a hint from the helpless-ness experiments in rats (see pp. 8-9), Wel-ker repeated his experiment with an FR-3treadle response as the test task. That is,the treadle had to be pressed three times inorder to produce food. This magnified thesize of the observed effect.
A potentially related finding was recentlyreported by Bainbridge (1973). At SO daysof age, rats were given a solvable black-white discrimination problem, an unsolvableproblem, or no problem. Twenty days laterthey were tested with the same tasks butdifferent stimuli, or in a Hebb-Williamsmaze based on spatial cues. The rats givenan initial unsolvable problem performedvery poorly in both the same and the differ-ent apparatus. Thus exposure to appetitiveevents delivered independently of behaviormay interfere with the acquisition of ap-petitively motivated responses, just as ex-posure to noncontingent shock interfereswith the acquisition of shock-motivated re-sponses.
So we believe that the psychological stateproduced by uncontrollability may under-mine response initiation quite generally.Dogs, rats, cats, fish, and people make fewerresponses to escape shock after receiving

LEARNED HELPLESSNESS: THEORY AND EVIDENCE 13
uncontrollable shock. Furthermore, thesemotivational deficits may not be limited toshock or even noxious events. Escape fromfrustration, aggressive behavior, and eventhe propensity to solve anagrams might beundermined by inescapable aversive events.Conversely, uncontrollable rewards disruptescape from loud noise, learning to procurefood, and competitiveness. However, itshould be stressed that many of the experi-ments cited here are subject to alternativeinterpretation, and much more research willbe required before the true extent of theseeffects will be known. Clearly, there mustbe a limit to the degree of generalization oflearned helplessness effects, and these limitsmust be explored and delineated. We can-not yet say what factors will prove import-ant. All that can be said now is that thereappears to be some degree of transfer.
Cognitive Deficits
We have argued that a major consequenceof experience with uncontrollable events ismotivational; uncontrollable events under-mine the motivation to initiate voluntary re-sponses to control future events. Experiencewith uncontrollable events may also have asecond consequence that is cognitive; ex-perience with uncontrollability may producea difficulty in learning that responses havesucceeded, even when responding is actuallysuccessful. Uncontrollability may retard theperception of control.
This phenomenon seems to appear inhelpless dogs, rats, and men. Occasionally,a naive dog sits and takes shock on the firstthree or four trials in the shuttle box, andthen on the next trial jumps the barrier andescapes shock successfully for the first time.Once a naive dog makes one response thatproduces shock termination, it learns rapidly.On all further trials, the subject respondsvigorously and goes on to learn to avoidshock altogether. But dogs that first re-ceived inescapable shock were different inthis respect also. About one third of themgo through a similar pattern—sitting throughshock on the first three or four trials andthen escaping successfully on the next. Thesedogs, however, then revert to taking theshock, and they fail to escape on future trials.
It appears as if one success is just notenough to make a dog that has experiencedinescapable shock learn that his respondingnow produces shock termination.
Miller and Seligman (1975) and Kleinet al. (in press) found that such a negativecognitive set results from uncontrollabilityin man. Three groups of students receivedescapable, inescapable, or no loud noise.Then they confronted two new tasks, a taskof skill and a task of chance. On each of 10trials in the skill task, they sorted 15 cardsinto 10 categories of shape, attempting tocomplete it within IS sec. Unknown tothem, the experimenter arranged to havethem succeed or fail on any given trial bysaying that time was up before they hadfinished or after. So they went through aprearranged run of successes and failures.At the end of each trial, the subjects rated.(on a 0-10 scale) what they thought theirchances of succeeding on the next trialwould be. Subjects who were previously ex-posed to inescapable noise showed very littlechange in their expectancy for successesafter each new success and failure. Theyhad difficulty perceiving that their responsewould affect succeeding or failing. Controlsubjects and subjects who had escaped noiseshowed large expectancy changes followingeach .success and failure. This showed thatthey believed outcomes to be dependent ontheir actions. The three groups did notdiffer in expectancy changes following suc-cess and failure in a "chance" task that theyperceived as a guessing game. So inescap-able loud noise produced a cognitive set inwhich people believed that success and fail-ure is independent of their own skilled;
actions within the experimental situation,and they therefore had difficulty perceivingthat skilled responses were effective.
Hiroto and Seligman (1975), Miller andSeligman (Note 1) and Klein et al. (inpress) also reported this negative cognitiveset in another form. After escapable, in-escapable, or no noise, as the reader willrecall, students had to solve anagrams (seep. 11). Two kinds of deficits emerged.Inescapable noise interfered with theirability to solve any given anagram. Inaddition, there was a pattern to the 20 ana-

14 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
TABLE 1MEAN NUMBER AND STANDARD DEVIATION or
CONSECUTIVE SUCCESSES BEFOREPATTERN SOLUTION
Noise
Subject Inescapable Escapable None
DepressedMSD
NondepressedMSD
5.92.53
6.93.36
4.21.36
4.51.58
7.11.22
3.7.86
Note. Adapted from Miller and Seligman (Note 1), In-escapable noise increased the number of successes required innondepressed subjects.
grams to be solved; each was arrangedwith its letters in this order: 53124, forexample, ISOEN, DERRO, OURPG, etc.As can be seen in Table 1, students who hadreceived inescapable noise required aboutseven consecutive successes before catchingon to the pattern; students who receivedescapable noise or no noise needed aboutthree consecutive successes. Unsolvable dis-crimination problem's, incidentally, producedthe same disruption of anagram solution asdid inescapable noise.
The existence of a negative cognitive setproduced by independence between respond-ing and outcomes bears on an importantissue in learning theory; When two eventsare presented independently of each other,for example, a tone and a shock presentedat random, does the subject learn anythingat all about the tone or does he merely cometo ignore it? A helplessness point of viewholds that men and animals actively learnthat responses and outcomes are independentof each other, and one way the learning ismanifested is by the difficulty they laterhave learning that the response produces theoutcome when it does. Merely substituteshock for outcome and tone for response.This suggests that organisms should alsoactively learn when a tone and shock areindependent and that organisms show thisby having trouble learning later that the toneis followed by the shock when it actually is.Rescorla (1967) held the contrary view.Independence between a tone and shock is aneutral condition in which nothing is learned.
Recent investigation has shown that activelearning does indeed occur when condi-tioned stimuli (CSs) and unconditionedstimuli (USs) are independently presented.Mellgren and Ost (1971) reported that agroup for which CSs were presented inde-pendently of food took longer to learn laterthat the CSs were associated with the foodthan did naive rats (or even rats for whomthe CSs predicted the opposite relationshipwith food). Kemler and Shepp (1971)showed that irrelevant stimuli were mostslowly learned about when they became therelevant stimuli for the solution of a discrimi-nation problem for children. Thomas, Free-man, Svinicki, Burr, and Lyons (1970)showed that pigeons that had two colorspresented independently of food tended notto discriminate later between two line tilts,one of which predicted the presence of foodand the other its absence. MacKintosh(1973) also reported retardation of condi-tioning by prior CS-US independence.
So independence between two stimulimay produce active learning, and this learn-ing seems to retard the ability of rats, pi-geons, and men to learn later that the stimulidepend on one another. This evidence co-heres with the effects of response-outcomeindependence on cognition and bolsters thehypothesis that such independence retardsthe ability of the organism to perceive thathis responding has contingent consequences(also see the discussion of experiments byMaier & Testa, 1975, pp. 24-25).
Emotional Deficits
Our first hint that uncontrollable aversiveevents had emotional as well as motivationaland cognitive consequences came when wefound that the motivational effects dissi-pated in time under some circumstances.Overmier and Seligman (1967) found thatafter one session of inescapable shock in thehammock, dogs were helpless in the shuttlebox 24 hours later. If, however, the dogswere not tested in 24 hours, but at 48, 72,or 168 hours after inescapable shock, theywere normal. This is the only 'circumstancewe know under which helplessness dissipatesin time. If multiple sessions of inescapable

LEARNED HELPLESSNESS: THEORY AND EVIDENCE 15
shock are given to the dog in the hammock,he will fail to escape a week later in theshuttle box (Selignian & Groves, 1970).Further, if the dog is reared from birth inlaboratory cages, having minimal experiencewith control over events, small amounts ofinescapable shock produce nontransient help-lessness (Seligman & Groves, 1970). Finally,no time course has been observed in rats;following one session of inescapable shock,rats fail to escape 5 minutes, 1 hour, 24hours, and 168 hours later. It must beborne in mind however, that under one cir-cumstance—a single session of inescapableshock in dogs of unknown past history—helplessness dissipates in time, and such atime course hints of a transient emotionaldisturbance.
Other evidence indicates the emotionalconsequences of uncontrollability. Onewidely quoted study (Brady, Porter, Con-rad, & Mason, 1958) is related to uncon-trollability and helplessness, but appears toshow less emotionality with uncontrollability.Two groups of four monkeys were givenshocks; half of them—the "executives"—had control over the shock and could avoidthem by bar pressing. The other four wereyoked, or helpless, since they could notmodify shock. The executives formed stom-ach ulcers and died, but the helpless mon-keys did not. Unfortunately, this resultmay be an artifact of the way the monkeyswere assigned to the two groups. All eightmonkeys were placed on the executive sched-ule originally, and the first four to startthe lever pressing became the executives.The last four became the yoked subjects.It is possible that the more emotional a mon-key is, the more quickly it begins to barpress when it is shocked (Sines, Cleeland, &Adkins, 1963). So the four most emotionalanimals may have become the executives,and the four least emotional may have be-come the yoked subjects.
Jay Weiss (1968, 1971a, 1971b, 1971c)has recently repeated the executive monkeydesign with rats. (See also Moot, Ceballa,& Crabtree, 1970.) Three groups of ratswere randomly assigned to the triadic de-sign. The executive animals showed fewerand less severe ulcers than the yoked ani-
mals. Moreover, the yoked rats lost moreweight, defecated more, and drank less thanthe executive. So rats given inescapableshock show more stress or emotional effectswhen measured by stomach ulcers than ratswho can control shock.
There is further evidence in rats that un-controllable shock produces more severeemotional reactions than does controllableshock. Mowrer and Viek (1948) shockedtwo groups of rats while the rats wereeating. One group could control the shockby jumping in the air, and the other groupreceived uncontrollable shock. The ratsgetting uncontrollable shock subsequentlyate less than those controlling shock (seeBrimer & Kamin, 1963; Desiderate & New-man, 1971; Lindner, 1968; and Payne, 1972,for an ongoing controversy surrounding thesedata). In an analogous human study byHokanson, DeGood, Forrest, and Brittain(1971), subjects performed a symbol match-ing task while being shocked. The scheduleswere individually arranged so that each sub-ject received an average of one shock every45 sec. Subjects in the controllability groupwere allowed to take as many time-outs asthey wished whenever they wanted. A yokedcontrol group received the same number oftime-outs at the same times. Measures ofblood pressure taken at 30-sec intervals in-dicated that yoked subjects showed con-sistently higher blood pressures. (See Averill& Rosenn, 1972; Bandler, Madaras, & Bern,1968; Corah & Boffa, 1970; Elliot, 1969;and Stotland & Blumenthal, 1964, for re-lated studies in humans using a variety ofother measures of emotional arousal. Thisis a complex and inconsistent literature andis reviewed by Averill, 1973.)
Hearst (1965) found that the presenta-tion of uncontrollable shocks resulted in abreakdown of a well-trained appetitive dis-crimination in rats. During uncontrollableshock his rats no longer discriminated be-tween two stimuli, one of which signaled thepresence and the other the absence of food.During controllable shock the appetitivediscrimination was maintained.
Such breakdown of appetitive discrimina-tions is reminiscent of the classic work on"experimental neurosis." The concept of

16 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
experimental neurosis is not a homogeneousone; neither is it well denned. Unlike theother studies reviewed, controllability hasnot been manipulated explicitly to producethe "neurosis." Yet from an operationalpoint of view, we can speculate that thelack or loss of control may be important intheir etiology. Typically, an animal is re-strained in some type of harness seriouslylimiting what it can do. Often the procedureis Pavlovian, and by definition the organismhas no control over the onset or the offsetof the stimuli presented. In Shenger-Krest-nikova's (in Pavlov, 1927) classic experi-ment, an appetitive discrimination deterio-rated and signs of 'distress were noted whenthe dog could no longer tell the differencebetween the rewarded and nonrewarded stim-ulus. In the work of Liddell, James, andAnderson (1934), sheep developed a rangeof maladaptive behaviors following uncon-trollable electric shock. Masserman (1943)trained monkeys to feed in response to asignal and made them "neurotic" by pre-senting a fear-arousing stimulus duringfeeding. Without "therapy" these monkeysremained disturbed almost indefinitely. Ac-cording to Masserman (1943) :
Markedly different, however, was the case of ani-mals that had been taught to manipulate variousdevices that actuated the signals and feeder be-cause in this way they could exert at least partialcontrol over their environment. This stood themin good stead even after they were made neuroticin as much as when their hunger increased they.gradually made hesitant, but spontaneous, attemptsto reexplore the operation of the switches, signalsand food boxes, and were bolder and more suc-cessful as food began to reappear, (p. 82)
In a dramatic primate study, Stroebel (1969)trained a group of rhesus monkeys to air-condition their overheated chamber by press-ing a lever, and also to control loud noise,annoying light, and mild shocks by pressingthe same lever. He then retracted the leverso that it could still be seen, but could nolonger be pressed. No further physical stres-sors were presented. Initial responding wasfrantic. Brain temperature became irregularand the circadian rhythm was disrupted.
As rhythm disturbance developed, members of this. . . group of subjects began to show lassitude andweakness; their fur became knotted, mottled and
poorly groomed; b'ehaviorally they performed un-predictably if at all on the right hand lever prob-lems, pausing often for naps and rest. The be-haviors exhibited by these animals were clearlynonadaptive in nature; for example, two subjectsspent hours in catching "imaginary" flying insects,one subject masturbated almost continuously,three subjects became almost compulsive hairpullers, and all tended to show movement stereo-typy alternating with an almost total lack of in-terest in their external environment. (Stroebel,1969, p. 97)
It is not clear whether there can be anyone theory which can account for "experi-mental neuroses," nor is it clear whether allthese phenomena are closely related. Butuncontrollability is prominently present, andemotional disruption is the frequent result.
In summary, three types of disruptionseem to be caused by uncontrollability ofaversive events in the laboratory: The mo-tivation to respond is reduced, the propensityto perceive success is undermined, and emo-tionality is modified. These effects holdacross a variety of situations and species. Weneed a theory to put all this together and wewill now propose one.
LEARNED HELPLESSNESS THEORY
The basic facts about learned helplessnesseffects in the laboratory have now been ar-rayed before the reader. - What must anadequate theory of helplessness accomplish?It must account for the three facets of theeffects of uncontrollable aversive events:motivational, cognitive, and emotional. Itmust be testable; there should be experi-ments that can be performed which wouldconfirm it if it is true, or disconfirm it if it isfalse. Finally, it must be; applicable outsidethe laboratory—it must be: useful in explain-ing any helplessness effects found in the realworld. This final requirement is beyond thescope of this article, but the interested readershould see Seligman (1975).
The theory to be presented is straight-forward, and the groundwork for it has beenprepared by the way we laid out the dataabove. It accounts directly for the motiva-tional and cognitive deficits and, with anadditional premise, can account for the emo-tional disturbance. The j theory has beentested in several ways, and as a bonus it

LEARNED HELPLESSNESS: THEORY AND EVIDENCE 17
predicts ways to eliminate helplessness effectsand ways to prevent them.
When an organism is faced with an out-come that is independent of his responses,he sometimes learns that the outcome is in-dependent of his responses.
This is the cornerstone of our view andprobably seems obvious to all but the mostsophisticated learning theorist. You will re-call our discussion of the response contin-gency space (Figure 1). Learning theoristswould much prefer that the kinds of con-tingencies that .can be learned about be assimple as possible. First they believed thatthe most that could be learned about was asimple pairing of a response and an outcome,and the pairing of the response with the ab-sence of the outcome. But this had to bebroadened to include partial reinforcement,with the subject integrating both kinds ofpairings. So what could be learned aboutwas broadened to the probability of an out-come given a response. Then, it was shownthat organisms could also learn about theprobability of an outcome given that it didnot make the indicated response. The addedstep which our view makes is that the organ-ism can learn about both these probabilitiesconjointly, that variation of experience cor-responding to the points in the responsecontingency space will produce systematicparticular, we have argued that exposingchanges in behavior and cognition. Inorganisms to the 45° line, in which the prob-ability of the outcome is the same whether ornot the response of interest occurs, produceslearning. Behaviorally, this learning shouldtend to produce lack of response initiation tocontrol the outcome; cognitively it shouldproduce a belief in the inefficacy of respond-ing and difficulty in learning that respondingsucceeds: and emotionally when the outcomeis traumatic it might produce emotionalchanges.
The basic triadic design employed in allthe helplessness studies reviewed above is, ofcourse, directly relevant to the premise thatmen and animals learn about, and form ex-pectations concerning, independence betweenoutcome and responses. So, for example,in the Seligman and Maier (1967) studyonly the yoked dogs later failed to escape
shock, while the dogs who could escape bypanel pressing and the dogs who were notshocked did not fail to respond later. Clearlysomething different happened to the dogsthat received shock independently of theirresponses. We believe they learned thatresponding was futile and therefore expectedfuture responding to shock to be futile. Wehave gone to pains to give an objectivedefinition of uncontrollabtiity and response-outcome independence. This is because thetheory is basically a three-step affair.
Information about contingency —>Cognitive representation of the contingency(learning, expectation, perception, belief)—*
Behavior
An organism must begin with the informa-tion about the contingency between responseand outcome. This information is a prop-erty of the world out there, a set of stimuli,and not a property of the receiver. We havecarefully defined the kind of information thatcan be called information that a response andan outcome are independent.
But the middle step in the chain is crucialand is easily overlooked. The informationabout the contingency must be processed andtransformed into a cognitive representationof the contingency.3 Such a representationhas been variously called "learning that out-come and response are independent," "per-ceiving that response and outcome are in-dependent," or "believing that response andoutcome are independent." We prefer tocall the representation "the expectation thatresponding and an outcome are independent."
A person or animal can be exposed to theenvironmental contingency in which an out-come response are independent, yet notform such an expectation. Immunization,as we shall see later in this article ';(refer top. 25), is an example. Conversely, a per-
3 For attempts to spell out in detail the relation-ship between the contingency information and itscognitive representation, the interested reader shouldconsult, for example, Kelly (1967, 1972), Weiner,Frieze, Kukla, Reed, Rest, and Rosenbaum (1971)from an attribution theory point of view; Irwin(1971) and Seligman and Johnston (1973) froma cognitive-learning theory point of view; as wellas Lazarus (1966) and Stotland (1969).

18 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
son can show a helplessness effect withoutbeing exposed to the contingency as such;he can merely be told that events are uncon-trollable.
Glass and Singer (1972) have done anextensive series of studies on the role ofcontrollability in reducing stress and havefound that merely telling a human subjectabout controllability duplicates the effects ofactual controllability. They attempted tomimic the stress of the urban environmentby having their subjects (college students)listen to a very loud melange of sound: twopeople speaking Spanish, one person speak-ing Armenian, a mimeograph machine, acalculator, and a typewriter. When sub-jects could actually turn off the noise bybutton pushing, they were more presistentin problem solving, they found the noiseless irritating, and they did better at proof-reading than yoked subjects.
Glass and Singer also presented anothergroup of subjects with the same noise, butthis time it was uncontrollable. However,this group had a panic button and was told:"You can terminate the noise by pressingthe button. But we'd prefer you not do it."None of the subjects in fact tried to turnoff the noise. All they had was the falseinformation that they could control the noiseif they had to. These subjects did just aswell as the subjects who actually controlledthe noise. Actual controllability and actualuncontrollability may often produce cor-responding expectations. But this type ofexperiment, in which the expectation is in-valid, highlights the fact that it is the ex-pectation and not the objective conditions ofcontrollability that is the cornerstone of ourtheory. (See also Geer, Davison, & Gatchel,1970; Geer & Maisel, 1972; Klein et al., inpress; Langer, 1974; Pervin, 1963; andStotland & Blumenthal, 1964, for relatedeffects of perceived control.)
So the first step of the theory is that theorganism acquires an expectation of re-sponse-outcome independence, when out-comes are uncontrollable. The second stepin the theory is the means by which the ex-pectation of response-outcome independenceproduces the effects associated with helpless-ness.
We assume that the incentive to initiatevoluntary responses in a traumatic situa-tion is partly produced by the expectationthat responding produces relief. In the ab-sence of this incentive, voluntary respondingwill decrease in likelihood. When a personor animal has learned that relief is indepen-dent of responding, the expectation that re-sponding produces relief is negated, andtherefore response initiation is reduced.Most generally put, the incentive to initiatevoluntary responses to control any outcome(e.g., food, sex, shock termination) comes,in part, from the expectation that respondingproduces that outcome. (See Bolles, 1972,for a discussion of expectational mechanismsand incentive motivation.)* In the absenceof this incentive, voluntary responding willdecrease in likelihood. When a person oranimal has learned the outcome is indepen-dent of responding, the expectation that re-sponding will produce the outcome, andtherefore response initiation, decreases.
The way in which this undermining ofmotivation works has been seen with crystalclarity in a human helplessness experiment(Thornton & Jacobs, 1971). Following in-escapable shock, college students sat andtook the shock. When asked why they didnot respond appropriately, 60% of the sub-jects reported they had no control overshocks, so why try. These subjective re-ports suggest that a belief in uncontrollabilityundermines the incentive to initiate re-sponses.
We also assume that learning that an out-come is independent of a response makes itmore difficult later to learn that responsesproduce that outcome.
Response^outcome independence is an ac-tive form of learning, and like any otheractive form of learning, it can proactivelyinterfere with contravening forms of learn-ing through an associative interference. At
*It should also be mentioned that innatelyelicited struggling is another source of respondingin a traumatic situation, but it is the waxing andwaning of voluntary (outcome-sensitive) responseswhich is our concern here. This does not denythat innate responses can be transformed intovoluntary responses (Schwartz & Williams, 1972).

LEARNED HELPLESSNESS: THEORY AND EVIDENCE 19
first the dog is not a passive recipient ofshock but emits many responses, but each ofthe responses is unrelated to shock termina-tion. The dog might, for example, turn hishead and shock might happen to go off onthe trial, but on other trials he might turnhis head and shock would not go off. Shockwould also terminate when he had not turnedhis head. Now when he is tested in theshuttle box and jumps the barrier, which inreality causes shock termination, the dog hastrouble learning this. This is because he hashad many experiences in which a pairing be-tween a response and shock terminationproved spurious and, as for head turning, hestill expects that shock will just as likely gooff if he fails to jump the barrier. Such adog may return to taking shock even afterhe makes one or two "successful" jumps.In contrast, a naive dog has no interferingexpectation that shock termination is inde-pendent of responding, so one experiencewith barrier jumping leading to shock term-ination is sufficient for him to learn. Forsome evidence regarding such associativeinterference see the discussion of Maier andTesta (1975), pp. 24-25.
This, then, is the theory of helplessness.The expectation that an outcome is inde-pendent of responding (a) reduces the mo-tivation to control that outcome and (b)interferes with learning that responding con-trols the outcome.
Seligman (1975) has recently extendedthis theory to changes in emotionality. Heargued that when a traumatic event firstoccurs it causes a heightened state of emo-tionality, which has been called "fear." Thefear continues until the subject learns thathe can or cannot control the trauma. If thesubject learns he can control the trauma,fear is reduced and may disappear altogether.If the subject learns that he cannot controlthe traumatic event, fear decreases and isreplaced with depression. For a more de-tailed discussion see Seligman (1975).
ALTERNATIVE EXPLANATIONS
Now that we have described the learnedhelplessness hypothesis and the manner inwhich it is able to account for the phe-nomena under consideration, we will describe
some theoretical positions which have beenadvanced as alternatives to the helplessnesshypothesis. It should be noted at the outsetthat these hypotheses have been offered asexplanations of only the fact that organismsexposed to inescapable shock later fail tolearn to escape and avoid shock in a shuttlebox, rather than as explanations of thebroad range of phenomena considered above.Thus we will describe these theoretical posi-tions in the context of the basic learnedhelplessness effect. These alternatiye the-ories can be divided into two groups, motiva-tional accounts and motor accounts.
Motivational Alternatives
Adaptation. The adaptation hypothesismaintains that a subject adapts to shockduring pretreatment with inescapable shocksand is therefore not sufficiently motivated toescape from shock in the shuttle box. Thehypothesis is inadequate:
1. Adaptation to repeated, intense shockhas never been demonstrated (Church Lo-Lordo, Overmier, Solomon, & Turner,1966).
2. It is unlikely that very much adapta-tion could persist for as long as 24 hoursand beyond.
3. In experiments with dogs, the dogs donot look as though they are adapted; theyhowl, defecate, and urinate to the firstshock presentation in the shuttle box. Onlater trials, the dogs are passive; but theywhimper and jerk with the shock.
4. We have disconfirmed the adaptationhypothesis experimentally. Raising the shocklevel in the shuttle box should increasemotivation to escape. However, Overmierand Seligman (1967) found that increasingthe shock level from 4.5 mA to 6.5 mA doesnot eliminate the interference effect.
5. A series of escapable shocks in thehammock does not produce failure to escapein the shuttle box (Seligman & Maier,1967), while the same shocks, if inescapable,do produce failure to escape. By this hy-pothesis, both conditions should lead toequal adaptation to shock and to similar be-havior in the test situation, but they do not.

20 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
6. Seligman and Maier (1967) gave onegroup of dogs 10 trjals of escape/avoidancetraining in a shuttle box before treatmentwith inescapable shock in the harness. Fol-lowing exposure to inescapable shocks in theharness, the dogs escaped and avoided shocknormally when returned to the shuttle box.Thus prior exposure to controllable shock"immunized" the dogs against the interfer-ing effects of exposure to inescapable shocks.This result follows from the helplessness hy-pothesis because prior experience with con-trollable shock should proactively interferewith the subject's learning that shock is un-controllable and should also allow the sub-ject to discriminate between the places whereshocks are controllable and uncontrollable.However, this result is not consistent withthe adaptation hypothesis. Prior exposureto escapable shock in the shuttle box shouldnot eliminate any adaptation produced byinescapable shocks in the harness.
7. If the subject is tested and fails toescape 24 hours after inescapable shocks,the learned helplessness effect will persist inchronic form; the subject will fail to escapeon later opportunities. The helplessnesshypothesis suggests a way to eliminate chro-nic failure to escape. By this hypothesis,the dog does not try to escape becausehe does not expect that any instru-mental response will produce shock termina-tion. By forcibly exposing the dog to theescape and avoidance contingencies, this ex-pectation might be altered. This type oftraining by "putting through" has been usedby others with mixed success (Loucks,1935; Miller & Konorski, 1928; Woodbury,1942). Seligman et al. (1968) reasonedthat forcibly dragging the dog from side toside in the shuttle box, in such a way thatthe dog's changing compartments terminatedshock, might effectively expose the dog tothe response-reinforcement contingency. Thiswas the case. The experimenter pulled threechronically helpless dogs back and forthacross the shuttle box with long leashes.This was done during CS and shock, whilethe barrier was absent. After being pulledacross the center of the shuttle box (and thusterminating shock and CS) 20, 35, and 50times, respectively, each dog began to re-
spond on his own. Then the barrier was re-placed, and the subject continued to escapeand avoid. The recovery from helplessnesswas complete and lasting. This type of ex-posure to the escape and avoidance contin-gencies should not affect any adaptation toshock that might have been present. Selig-man et al. (1975) replicated these findingsin rats.
Sensitization. Perhaps the inescapableshocks received in the harness sensitize thesubject to shock so that it is too motivatedto enable it to make organized responses inthe shuttle box. This hypothesis is inade-quate, (a) Sensitization explains the in-efficiency of responding but not the absenceof responding, (b) Lowering the shocklevel in the shuttle box should permit thesubject to make organized responses. How-ever, Overmier and Seligman (unpublisheddata) found that the interference effect isnot attenuated when shock in the shuttle boxis reduced to 3.0 mA. (3) Arguments 5,6, and 7 in the previous paragraph, whichinvalidate the adaptation hypothesis, also in-validate the sensitization argument.
Motor Activity Alternatives
Within the last 2 years three hypotheseshave been offered as explanations of thelearned helplessness effect which are moredifficult to dismiss than are sensitization andadaptation. These explanations have theadvantage that they are able to account forthe fact that the learned helplessness effectis determined by the controllability of theshock during the initial treatment. We haveclassified them as motor activity hypothesesbecause they all maintain that exposure toinescapable shock interferes with subsequentshuttle box acquisition because it changesmotor activity. Two of these hypothesesargue that exposure to inescapable shockestablishes a motor response that is incom-patible with shuttling and therefore competeswith shuttling. Thus these two hypothesesargue that the organism does not learn shut-tling because it is performing a. motor re-sponse that prevents the occurrence of shut-tling. The remaining hypothesis maintainsthat inescapable shock is a powerful stressor

LEARNED HELPLESSNESS: THEORY AND EVIDENCE 21
and depletes a neurochemical necessary forthe occurrence of movement. Thus this viewholds that inescapably shocked animals failto acquire shuttling because they cannotmove sufficiently. :
Incompatible motor response theories. Asalready noted, two different incompatiblemotor response explanations of the learnedhelplessness effect have been proposed. Thefirst to be described was offered by Brace-well and Black (1974) and was suggestedby three experiments. In their first experi-ment they examined the effects of restraintand inescapable shock upon simple FR-1shuttlebox escape/avoidance acquisition.They found that restraint in the absence ofany exposure to inescapable shock produceda decrement in FR-1 shuttlebox acquisition,but that exposure to inescapable shock in theabsence of restraint did not produce a reli-able decrement. In their second experimentBracewell and Black found that exposure to"high" intensity (1.0 mA) inescapable shockproduces a small decrement in FR-1 shuttlebox acquisition even when delivered in theunrestrained condition. In their final ex-periment Bracewell and Black explicitlypunished movement by arranging a positivecorrelation between movement and shockintensity. This procedure led to a decre-ment in subsequent FR-1 shuttle box ac-quisition.
Bracewell and Black (1974) proposed thefollowing interpretation of their data:
One explanation that accounts for these results isthe operant conditioning of responses incompatiblewith the shuttle box escape response that occurredin the third case [where movement was explicitlypunished], also occurred in the first two. Thatis, restrained rats may have been punished forstruggling and consequently, learned to hold still,and unrestrained rats may have learned to holdstill or to perform some other incompatible re-sponse to reduce the high intensity shock, (p. 67)
Thus Bracewell and Black (1974) arguedthat because explicit punishment of move-ment produced a retardation in shuttle boxacquisition, the other instances of retardedacquisition might also be due to punishmentof movement during the pretreatment phase.They went on to argue that the learnedhelplessness effect might be explainable inthe same terms. It should be noted that
this hypothesis can explain why escapableshock does not produce a subsequent decre-ment. If shock is escapable, the organismlearns a response that completely eliminatesshock (the escape response) and so doesnot have to learn a response that only miti-gates shock (the putative incompatible re-sponse). It need only be argued that theformer is not incompatible with shuttling butthe latter is.
We will present a large amount of datawhich we feel to be inconsistent with theBracewell and Black (1974) hypothesis.However, these experiments also bear on thesecond competing motor response theory andso they will not be presented until that theoryhas been described. Nevertheless, a fewcomments are appropriate here. First, thefact that explicit punishment of movementproduces a decrement in subsequent escape/avodiance acquisition does not imply thatother procedures which produced escape/avoidance decrements do so because theypunish movement. There are undoubtedlymany ways to produce poor escape acquisi-tion, and it is unlikely that all operatethrough a similar mechanism. Paralyzingthe animal with a drug would retard escapeacquisition, but this does not mean that thedebilitating effect of inescapable shocks oc-curs because inescapable shock induces pa-ralysis. Thus the retarding effect of move-ment punishment does not imply that in-escapable shock retards acquisition becauseit punishes movement. Moreover, the ex-plicit punishment of movement in the Brace-well and Black experiment led to a muchsmaller escape acquisition decrement thandid inescapable shock delivered to restrainedsubjects. The punishment procedure in-creased the subsequent mean escape latencyacross the 10 test trials from the controllevel of 2.4 sec to 9.1 sec. The inescapableshock procedure resulted in a 21.5-sec meanescape latency. So it can be argued that theexplicit training of an incompatible responsedid not duplicate the effects of exposure toinescapable shock in restrained subjects, andso the argument that the movement punish-ment effect accounts for the inescapable shockeffect seems questionable. It can, of course,be argued that a more effective or different

22 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
punishment procedure might yield a largerescape decrement, but that remains to bedemonstrated.
Further, it is not clear that Bracewell andBlack (1974) employed a test procedure thatis generally sensitive to learned helplessnesseffects* In order for an effect to be called alearned helplessness effect, it must be demon-strated that the effect is caused by the con-trollability of the events delivered duringpretreatment. There is no evidence thatdecrements in FR-1 shuttle box acquisitionproduced by prior exposure to shock arecaused by the inescapability of the shocks inrats. There is not a single study employinga triadic design in rats (see p. 6) that hasfound appropriate effects with FR-1 shuttlingas the dependent variable. In fact, therehave been both reported (Anderson, Schwen-diman & Payne, Note 3; Maier et al.,1973; Seligman & Beagley, 1975) and un-reported (Carder, Leaf, Note 4) failures tofind any effect of prior exposure to shock onFR-1 .shuttle box escape acquisition in therat. It is to be noted that both Andersonet al. and Maier et al. (1973) delievered in-escapable shock to restrained rats and stillfailed to observe any effects. There are anumber of differences between the variousFR-1 shuttle box experiments, and it is notknown which are responsible for the differ-ence in results. Nevertheless, the dependentvariable used by Bracewell and Black maynot be one that is sensitive to helplessnesseffects in rats and so the implications oftheir results for learned helplessness is dif-ficult to assess.
A further problem concerns the generalityof the effects found by Bracewell and Black(1974). It will be recalled that they foundrestraint in the absence of shock to producea subsequent escape deficit and inescapableshock in the absence of restraint to produceonly a small escape decrement. However,Maier et al. (1973) used an escape/avoid-ance task known to be sensitive to helpless-ness Affects in rats (see p. 8) and foundno effect of restraint alone. Cohen (1970)found no effect of restraint on shuttle boxescape/avoidance acquisition in dogs.Further, Looney and Cohen (1972), Selig-man and Beagley (1975), and Seligman
et al. (1975) reported large escape deficitsproduced by exposure tO; inescapable shockin unrestrained rats. Again there are manydifferences between studies, but it is clearthat restraint does not always produce asubsequent escape decrement nor does in-escapable shock have to be delivered to re-strained subjects for a large effect to occur.
A second incompatible motor responsetheory has been proposed by Anisman andWaller (1973). They argued that exposureto shock induces response repertoire changesin the organism, and if no, coping response isavailable, "freezing" becomes the organism'sdominant reaction to shock. Freezing would,of course, compete with ;the occurrence ofactive responding in a subsequent escape/avoidance task. As evidence for their hy-pothesis, Anisman and Waller cite a varietyof studies which show that manipulationsdesigned to increase freezing have a detri-mental effect on avoidance behavior, andmanipulations designed to reduce freezingfacilitate avoidance performance. So, forexample, strong shock, which causes morefreezing in rats, produced more interferencewith shuttle avoidance than weak shock(Anisman & Waller, 1972) and scopalamine,a drug that reduces freezing, makes ratsbetter avoiders (Anisman, 1973).
A number of comments are appropriate.First, the experiments discussed by Anismanand Waller are not directly relevant to thelearned helplessness effect. They are allstudies of effects on avoidance, not escape.There is no evidence that the manipulationsdescribed by Anisman and Waller affectescape acquisition. In fact, escape latenciesare not even reported in many of the experi-ments (e.g., Anisman, 1973). The learnedhelplessness effect is a failure to escape—toflee from shock itself—a more profounddebilitation than failure to prevent shock.Second, even if it could be shown that in-ducing freezing debilitates escape, this doesnot mean that inescapable shock interfereswith escape by inducing freezing. In addi-tion, dogs do not seem to freeze, people whoreceive unsolvable discrimination problemsor inescapable noise do not freeze, and organ-isms that receive noncontingent food do notfreeze, yet these conditions may all produce

LEARNED HELPLESSNESS: THEORY AND EVIDENCE 23
helplessness. Finally, why does inescapableshock, but not escapable shock, producefreezing (if it does) in rats? We have notbeen able to think of an answer that does notimply that the rat has learned that shock isinescapable, and this is the heart of thehelplessness view.
Experimental evidence. We feel that thereare six lines of evidence that bear on the in-compatible motor response theories, andeach will be described.
1. It has already been noted that rats ex-posed to inescapable shock often do not failto learn an FR-1 shuttle box response. Thishas been investigated by Maier et al. (1973),and in agreement with a number of previousinvestigators they found that rats exposedto inescapable shock under a wide range ofparameters subsequently responded asrapidly as did controls in a two-way shuttlebox escape/avoidance acquisition test. How-ever, they noted that the rats in their experi-ments seemed to acquire this shuttling escaperesponse in a manner different from thatexhibited by dogs. The rats responded veryrapidly, but the response was- as rapid onthe very first acquisition trial as it was after30 trials—the acquisition curves were flat.The mean response latency from shock onseton the very first shuttle box trial was 2.67sec. The escape responding of dogs in ashuttle box is quite different. Averagedacross experiments, the naive control dogs inthe Maier, Overmier, and Seligman experi-ments took slightly over 30.0 sec of shockbefore escaping on the first trial and didnot emit short latency escape responsesuntil the fourth trial. This suggested toMaier et al. (1973) that an interferenceeffect in rats might be found if a moreslowly acquired escape response were usedas the test task. The escape response theyused was one in which the rats had to crossthe shuttle box twice in order to terminateshock. However, these FR-2 trials were pre-ceded by five ordinary shuttle box trials (FR-1), since pilot investigation revealed thatnaive rats sometimes fail to acquire the FR-2shuttling response if such prior training wasnot provided. The result was that a largeinterference effect appeared—half of the rats
exposed to inescapable shocks did not learnthe FR-2 escape response even though theyresponded rapidly on the five FR-1 trials,while all control rats learned the FR-2escape response. It should be noted that wehave replicated this effect several times, ashave other laboratories (Seligman &Beagley, 1975; Weisman, Note 5)i
The fact that rats exposed to inescapableshock will learn FR-1 shuttling but will notlearn FR-2 shuttling might be explained ina variety of ways. The helplessness hy-pothesis asserts that prior exposure to in-escapable shock interferes with the subse-quent formation of associations betweenresponding and shock termination and, inaddition, reduces the organism's incentive toattempt to escape. From the foregoing dis-cussion it is clear that crossing the shuttlebox once is a high probability initial responseto shock in the rat, perhaps a species-specificdefense response (Bolles, 1970) and, atleast on early trials, may be elicited. Con-sistent with this view of shuttle box crossing,Maier et al. (1973) found that if conditionswere arranged such that there was no. con-tingency between crossing the shuttle boxand shock termination, naive rats would stillcross the shuttle box in response to shockwith short latencies for about 10 trials.
It should be noted that this strong tend-ency of the rat to cross the shuttle boxrapidly in response to shock guarantees thatthe preshocked rats will be exposed to theFR-1 contingency a relatively large numberof times even if such a rat is slow to learn.In addition, the contingency between crossingthe shuttle box and shock termination isrelatively simple under the FR-1 procedure,and so the preshocked rats might learn FR-1escape even though associative interference isactually present. If associative interferencewere present, some difference in FR-1 ac-quisition between inescapably shocked andcontrol rats, at least on early trials, mightbe expected. However, such a differencecould be masked by the elicited nature ofshuttling on early trials. Consistent withthis argument, Testa, Juraska, and Maier(1974) found that FR-1 shuttling extin-guished more rapidly in groups given previ-ous exposure to inescapable shock than in

24 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
controls, even though these groups did notdiffer in the acquisition of FR-1 shuttling.
The FR-2 shuttling response, on the otherhand, is not as probable an initial uncondi-tioned response to shock as is FR-1 shut-tling, and presents a more complicated con-tingency between crossing the shuttle boxand shock termination than does FR-1 shut-tling. Thus the associative interference pro-duced by learning that shock is inescapablemight be able to prevent the acquisition ofcan alsb explain why exposure to inescap-the FR-2 shuttle box response.
Both competing motor response hypothesesable shock has little or no effect on FR-1shuttling. The competing motor responsetheories could argue that the competingresponse is not strong enough to interferewith the highly elicited FR-1 but can com-pete with the initially weaker FR-2. Inaddition the FR-2 procedure exposes therat to more shock than does the FR-1 pro-cedure, and it could be argued that this in-tensifies the competing response.
Although these hypotheses are not mutu-ally exclusive, they point to different aspectsof the FR-2 procedure as the crucial featurein producing an interference effect. The help-lessness5 hypothesis emphasizes the difficultyof learriing the contingency between crossingthe shuttle box and shock termination in-herent in the FR-2 procedure, while thecompeting response explanations emphasizethe physical difficulty of the FR-2 responseand/or the large amount of shock to whichthe FR-2 procedure exposes an organism.There are two obvious ways to determinewhich of these aspects of the FR-2 pro-cedure is crucial. The first is to simplifythe contingency between crossing the shut-tle box and shock termination while at thesame time maintaining the physical difficultyand shock exposure characteristics of theFR-2. The second is to arrange a com-plex contingency between shuttling and shocktermination while at the same time reducingthe physical difficulty of the response and theextent of shock exposure. Maier and Testa(1975) have conducted experiments usingeach of these strategies.
The contingency between crossing theshuttle box and shock termination should be
relatively difficult to learn with the FR-2procedure, for the obvious reason that shockdoes not terminate after each crossing. Inthe Maier et al. (1973) FR-2 experiment,shock was simultaneously presented to thegrids of both chambers of the shuttle box,so that there was no consequence for the firstresponse of the FR-2, not even a momentaryinterruption of shock presentation. Thecontingency between the shuttle response andshock termination can be improved by ar-ranging a brief interruption of shock fol-lowing the first response in the FR-2. Itshould be noted that such a procedure willnot change the effortfulness of the FR-2nor will it have an appreciable effect on theduration of shock exposure produced bythe FR-2 procedure, if the interruption inshock is very brief.
Maier and Testa (1975) conducted pre-cisely the experiment suggested above. Itwas found that although inescapably shockedrats fail to learn ordinary FR-2 shuttling,they learn as well as nonshocked controlswhen there is a 1-sec interruption of shockafter the first crossing of the FR-2. Sincethe only obvious difference between the pro-cedures which did and did not yield a learnedhelplessness effect was the nature of thecontingency between responding and shocktermination, it seems likely that associativefactors are strongly involved.
The previous experiment suggests thatthe inescapably shocked rats learn the FR-1task but do not learn the FR-2 task becausethe FR-1 task contains an obvious con-tingency between shuttling and shock term-ination while the FR-2 task contains a de-graded contingency. If this is the case, thenit ought to be possible to produce a learnedhelplessness effect with a single crossing ofthe shuttle box if the contingency betweenthe single crossing and shock terminationwere degraded.
In a second experiment, Maier and Testa(1975) degraded the contingency betweena single crossing of the shuttle box andshock termination by interposing a time de-lay between crossing of the shuttle box andshock termination. Thus the requiredescape response was no more effortful thanthe ordinary FR-1 task which does not

LEARNED HELPLESSNESS: THEORY AND EVIDENCE 25
TABLE 2DESIGN OF THE MAIER AND RHOADES EXPERIMENT
Group
ES-ISIS-ISNS-ISNS-NS
i
Escapable wheel turnInescapable wheel turnNo-shock wheel turnNo-shock wheel turn
Phase
2
Inescapable tubeInescapable tubeInescapable tubeNo-shock tube
3
Escapable shuttle boxEscapable shuttle boxEscapable shuttle boxEscapable shuttle box
yield an interference effect, but containedan obscured contingency between shuttlingand shock termination, as does the FR-2task which does yield an interference effect.We found that a delay in shock terminationas short as 1 sec severely retarded the ac-quisition of FR-1 shuttle box escape in ratspreviously exposed to inescapable shocks,but had no effect on subjects not previouslyexposed to inescapable shocks. It shouldbe noted that a procedure designed to con-trol for the extra amount of shock producedby the delay revealed that this factor didnot account for the results. Thus priorexposure to inescapable shock will retardthe acquisition of even low-effort escape re-sponses if the contingency between theescape responses and shock termination ismade less obvious.
Taken together, the results of these twoexperiments indicate that the nature of thecontingency between the escape response andshock termination is more important than arethe effort and shock exposure characteristicsof the escape response in determiningwhether an interference effect will occur.Inescapably shocked rats fail to learn FR-2shuttle box escape because the contingencybetween crossing the shuttle box and shocktermination is here more complex, not be-cause the FR-2 response is effortful or ex-poses the subject to long shock durations.It is therefore difficult to account for the ratinterference effect with an explanation thatdoes not posit an associative deficit followinginescapable shock.
The incompatible motor response hy-potheses cannot handle these data. It isdifficult to conceive of a motor response thatwould interfere with FR-2 shuttling whenthere is no shock interruption, but not whenthere is. Similarly, what motor response
could be incompatible with FR-1 shuttlingif there is a short delay in shock terminationbut not incompatible when there is no delay ?
2. It will be recalled (p. 20) that Selig-man and Maier (1967) found that prior ex-posure to controllable shock immunizes theorganism against the deleterious effects ofexposure to uncontrollable aversive events.This finding might seem inconsistent with theincompatible motor response hypotheses sinceprior exposure to escapable shock should notprevent the establishment of the incompatibleshock-mitigating response or freezing dur-ing exposure to uncontrollable shock. How-ever, in the Seligman and Maier experimentthe subjects' initial experience with con-trollable shock was given with the same taskthat was used as the test task following ex-posure to inescapable shock. That is, thesubjects were first given escape training in ashuttle box, then given inescapable shock ina harness, and then tested in a shuttle box.Thus the immunization effect may not havebeen caused by the organisms' initial con-trol over shock but rather by the acquisitionof the specific response later used as a test.
The confounding of initial control overshock with the acquisition of the test taskresponse allows the incompatible motor re-sponse theories to account for the Seligmanand Maier immunization experiment. Theycould argue that the Seligman and Maierprocedure strengthened shuttling sufficientlyto overcome the putative competing re-sponse.
Clearly, what is needed is an experimentwhich separates the effects of initial controlover shock and the prior strengthening ofthe specific response later to be tested. Anunpublished experiment by Maier andRhoades meets this requirement. (See alsoSeligman et al., 1975). The design of tfie

26 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
s-uLUH-«t_J
<LUE
35-
30-
25-
20"
is-le-s'
IS-IS0
ES-IS*NS-IS° /--»o '̂0\NS-NS* / XX
D^ .̂D °/ J^
v!>f\- —o %S *^
a
i i i i i i1 2 3 4 5 6
BLOCKS OF 5 TRIALS
FIG. 7. Mean shuttle box FR-2 escape responselatency for rats previously given either escapableshock (ES), yoked inescapable shock (IS), or noshock (NS) in a wheel-turn apparatus, and thengiven either inescapable shock (IS) or no shock(NS) in a restraining tube (Maier & Rhoades, un-published data, 1975). (The rats given escapetraining in the wheel-turn box and then given in-escapable shocks in the restraining tube [ES-IS]were not helpless in the shuttle box.)
experiment can be seen in Table 2. Theexperiment involved three phases and fourgroups of rats. In the first phase one groupof rats was trained to escape shock by turn-ing a small wheel with their paws, a secondgroup received yoked inescapable shock in awheel-turn box, and two groups were placedin the wheel-turn box but not shocked. Inthe second phase both of the shocked groupsand one of the control groups received in-escapable shock in a restraining tube, andthe remaining group was only restrained. Inthe third phase all groups were given fivetrials of FR-1 followed by 25 trials of FR-2escape/avoidance training in a shuttle box,our standard helplessness test task.
The results found by Maier and Rhoadesare seen in Figure 7. Prior experience withescapable shock in the wheel-turn situationmitigated the interfering effects of inescap-able shock on shuttle box escape acquisition.Although prior experience with escapableshock in the wheel-turn situation did notcompletely eliminate the effects of exposure
to inescapable shock, the immunizing effectwas reliable. It is difficult for competingresponse theories to account for an immuniz-ing effect that is caused by the experienceof control per se rather than by the priorstrengthening of the test response. It isdifficult to argue that learning to turn a smallwheel with the paws in a wheel-turn boxdirectly strengthens running in a shuttlebox. This wheel-turn escape training shouldnot have an immunizing effect, but it does.
3. If inescapable shock interferes withsubsequent escape acquisition because it hasled to the development of a motor responseincompatible with the escape response, thenthe explicit reinforcement of such a responseshould duplicate the effects of exposure toinescapable shock. As already noted, Brace-well and Black (1974) found that the explicitpunishment of movement led to a smalldecrement in subsequent acquisition in ashuttle box. However, this experiment isdifficult to interpret for the reasons alreadynoted (see pp. 21-22). Another study usingthis strategy was reported by Maier (1970).One group of dogs was trained to escapeshock by holding still, a response directlyincompatible with jumping over a hurdle ina shuttle box. The dogs were restrained ina hammock and panels were placed | in.(.64 cm) above and | in. (.64 cm) to eachside of the dog's head. The dogs could notprevent the onset of shock, but they couldterminate shock by not touching any of thepanels for a specified period of time once theshock came on. Since shock elicits headmovements in restrained dogs, the dogswere required to actively inhibit movementselicited by shock. Because the panels werepositioned 1 in. (.64 cm) from the dog'shead, the dogs could refrain from touchingthe panels only by remaining almost per-fectly motionless, by freezing. A secondgroup of dogs received inescapable shockyoked to the first group and a third receivedno pretreatment. Following this treatmentall groups received 12 days of testing in ashuttle box.
If inescapable shock produces a learnedhelplessness effect because it induces freez-ing or leads to the punishment of movement,then the group taught to inhibit movement as
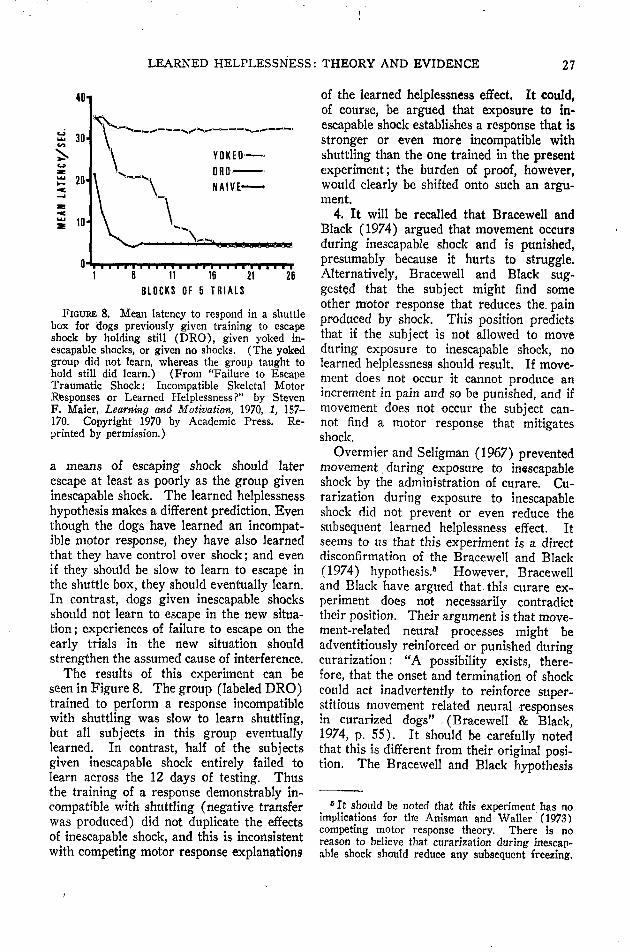
LEARNED HELPLESSNESS: THEORY AND EVIDENCE 27
40-1
S 30-CO
£E 20-
ae
£ 10-
0-
^ „-_. — -\ Y O K E D - •
\ """A NAIVE-—
V v11 16 21
B L O C K S OF 5 T R I A L S26
FIGURE 8. Mean latency to respond in a shuttlebox for dogs previously given training to escapeshock by holding still (DRO), given yoked in-escapable shocks, or given no shocks. (The yokedgroup did not learn, whereas the group taught tohold still did learn.) (From "Failure to EscapeTraumatic Shock: Incompatible Skeletal MotorResponses or Learned Helplessness?" by StevenF. Maier, Learning and Motivation, 1970, 1, 157-170. Copyright 1970 by Academic Press. Re-printed by permission.)
a means of escaping shock should laterescape at least as poorly as the group giveninescapable shock. The learned helplessnesshypothesis makes a different prediction. Eventhough the dogs have learned an incompat-ible motor response, they have also learnedthat they have control over shock; and evenif they should be slow to learn to escape inthe shuttle box, they should eventually learn.In contrast, dogs given inescapable shocksshould not learn to escape in the new situa-tion ; experiences of failure to escape on theearly trials in the new situation shouldstrengthen the assumed cause of interference.
The results of this experiment can beseen in Figure 8. The group (labeled DRO)trained to perform a response incompatiblewith shuttling was slow to learn shuttling,but all subjects in this group eventuallylearned. In contrast, half of the subjectsgiven inescapable shock entirely failed tolearn across the 12 days of testing. Thusthe training of a response demonstrably in-compatible with shuttling (negative transferwas produced) did not duplicate the effectsof inescapable shock, and this is inconsistentwith competing motor response explanations
of the learned helplessness effect. It could,of course, be argued that exposure to in-escapable shock establishes a response that isstronger or even more incompatible withshuttling than the one trained in the presentexperiment; the burden of proof, however,would clearly be shifted onto such an argu-ment.
4. It will be recalled that Bracewell andBlack (1974) argued that movement occursduring inescapable shock and is punished,presumably because it hurts to struggle.Alternatively, Bracewell and Black sug-gested that the subject might find someother motor response that reduces the. painproduced by shock. This position predictsthat if the subject is not allowed to moveduring exposure to inescapable shock, nolearned helplessness should result. If move-ment does not occur it cannot produce anincrement in pain and so be punished, and ifmovement does not occur the subject can-not find a motor response that mitigatesshock.
Overmier and Seligman (1967) preventedmovement during exposure to inescapableshock by the administration of curare. Cu-rarization during exposure to inescapableshock did not prevent or even reduce thesubsequent learned helplessness effect. Itseems to us that this experiment is a directdisconfjrmation of the Bracewell and Black(1974) hypothesis." However, Bracewelland Black have argued that this curare ex-periment does not necessarily contradicttheir position.. Their argument is that move-ment-related neural processes might beadventitiously reinforced or punished duringcurarization: "A possibility exists, there-fore, that the onset and termination of shockcould act inadvertently to reinforce super-stitious movement related neural responsesin curarized dogs" (Bracewell & Black,1974, p. 55). It should be carefully notedthat this is different from their original posi-tion. The Bracewell and Black hypothesis
slt should be noted that this experiment has noimplications for the Anisman and Waller (1973)competing motor response theory. There is noreason to believe that curarization during inescap-able shock should reduce any subsequent freezing.

28 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
argued that an incompatible response is ac-quired because it produces a positive out-come of reduced pain or discomfort. ThusBracewell and Black provided a mechanismwhereby the incompatible response is ac-quired, an explicit reinforcement mechanism.But in the case of the curarized subjectBracewell and Black do not argue that "themovement related neural processes" reducepain and are therefore explicitly reinforcedsince there is no obvious way that suchprocesses could reduce pain. Thus Brace-well and Black argue that in the case ofcurarized subjects movement-related neuralprocesses are adventitiously rather than ex-plicitly reinforced, and this seems gratuitous.Although there is evidence that movement-related neural processes can be explicitlypunished (Black, Young, & Batenchuk,1970), there is no evidence that they can beadventitiously or superstitiously reinforced.Further, even if such processes could beadventitiously reinforced or punished there isno reason to assume that the overt movementproduced by such a process would be incom-patible with shuttling. It could just as easilyfacilitate shuttling. To assume that it will beincompatible is, again, gratuitous.
5. We have already described an experi-ment by Rosellini and Seligman (1975) (seep. 10) in which rats were given escapableshock, inescapable shock, or no shock, andwere then trained to traverse a runway forfood. Following training, the rats wereextinguished and then allowed to escapefrom the presumably frustrating goal box.The result was that the rats previously ex-posed to inescapable shock were slow toescape the goal box but did not differ fromcontrols in either acquisition or extinction.If a motor response is established duringexposure to inescapable shock, it is difficultto see why it would interfere with escapefrom the goal box but not interfere withrunway acquisition and extinction.
6. The incompatible motor responsetheories have difficulty dealing with situa-tions that do not employ electric shocks orother noxious unconditioned stimuli. Theparticular competing response hypothesesoffered by Bracewell and Black (1974) andby Anisman and Waller (1973) cannot ac-
count for appetitive analogues to the learnedhelplessness effect (see p. 12) in animals orlearned helplessness effects in human beingsinduced by exposure to loud noises (see p.9). Moreover, it is difficult to see howany incompatible motor response view couldaccount for the noise-escape failure producedby unsolvable discrimination problems inhuman subjects (see p. 11). It should benoted that the incompatible motor responsehypotheses which we have discussed weredesigned to account only for the effects ofinescapable shock in animals, and it couldbe argued that these other effects are differ-ent phenomena and therefore need not all beexplained by the same theory. Such a moveseems ad hoc and our preference is for thattheory which is able to integrate the largestamount of data.
Motor Activation Deficit Hypothesis
A somewhat different motor response ex-planation of the learned helplessness phe-nomenon has been offered by Weiss et al.,(1975). Their explanation of the learnedhelplessness effect is as follows:
When animals were exposed to severe inescapableshock, a deficiency in central naradrenergic activityoccurred, deriving in part at least, from the de-pletion in the level of norepinephrine seen in thiscondition. As a consequence of this noradrenergicdeficiency the animals could mediate only a limitedamount of motor activity, an amount insufficientfor learning and performance of the correct re-sponse in the shuttle avoidance task on which theywere tested. On the other hand, animals exposedto the same shocks but able to control then! didnot develop this noradrenergic deficiency and there-fore could mediate sufficient motor behavior toperform adequately, (p. 7)
A number of factors led Weiss et al. (1975)to develop this explanation of the learnedhelplessness effect. First, Miller and Weiss(1969) argued that the dissipation of thelearned helplessness effect over a 48-hourinterval following one session of exposureto inescapable shock in dogs suggests media-tion by a time-dependent physiological changerather than by learning. They argued thatthings that are learned do not disappearacross an interval as short as 48 hours.Second, it is known that central catechol-amines have a time course of recovery fol-

LEARNED HELPLESSNESS: THEORY AND EVIDENCE 29
lowing depletion (e.g., Rech, Bovys, &Moore, 1966) and are involved in the media-tion of movement (e.g., Herman, 1970).Third, Weiss (1968, 1971a, 1971b, 1971c)has shown that rats given inescapable shockundergo greater stress than do rats givenequivalent escapable shocks, as measured bygastric lesions, loss of body weight, plasmasteroid levels, and fearfulness. Finally, thisled Weiss, Stone, and Harrell (1970) toassay whole brain norepinephrine followingexposure to escapable or yoked inescapableshock. They found norepinephrine levels tobe lower following exposure to inescapableshock. This finding has been replicated inseparate brain regions by Weiss, Pohorecky,Dorros, Williams, Emmel, Whittlesey, andCase (Note 6) and has been shown to oc-cur regardless of the amount of motor activ-ity required to escape and avoid shock(Weiss, Pohorecky, Emmel, & Miller, citedin Weiss et al., 1975). Following thesedemonstrations of effect on endogenousnorepinephrine levels, Weiss, Pohorecky,McMenima, Berkeley, and Jaffe (cited inWeiss et al., 1975) estimated norepinephrinereuptake and release in vivo by infusion ofradioactive (3H) norepinephrine. Theyfound that reuptake occurred at a higherrate in rats previously exposed to inescap-able than escapable shocks. This reuptakedifference could account for the differencesin the level of endogenous norepinephrine.
Weiss et al. (1975) present 12 experi-ments in support of the notion that thelearned helplessness effect occurs becauseinescapable shock is a severe stressor, pro-duces a temporary depletion of norepi-nephrine, and thus depresses the movementthe organism can emit below what is re-quired for shuttle box acquisition. We willdescribe each of these 12 experiments. Inthe first three experiments, Weiss et al.attempted to show that a procedure whichdeplete norepinephrine but does not involvethe factors stressed by the learned helpless-ness hypothesis duplicates the learned help-lessness phenomenon. It is known that aforced swim in 2 °C water rapidly depletesbrain norepinephrine levels and reduces itsrelease (Stone, 1970a, 1970b). Thus themotor activation deficit hypothesis would
expect a forced swim in cold water to pro-duce a learned helplessness effect. However,a forced swim in warm water (28 °C) doesnot produce a depletion of brain norepi-nephrine and so the motor activation deficithypothesis would not expect a warm swim toproduce a learned helplessness effect. Weisset al. argued that animals forced to swim ineither warm or cold water should experi-ence the same lack of control, and that thelearned helplessness hypothesis would thusexpect similar effects resulting from a warmor a cold swim. In Experiment 1, rats wereput in either warm (28 °C) or cold (2 °C)water for either 3.5 or 6.5 minutes. Thirtyminutes later the rats were all given an FR-1shuttle box shock escape/avoidance acquisi-tion test. The results were that the rats ex-posed to cold swim were severely retardedin shuttle box acquisition, while the warm-swim subjects showed normal acquisition.
Experiment 1 led Weiss et al. (1975) toconclude that cold swim duplicated the fail-ure-to-learn aspect of the learned helpless-ness phenomenon. Another feature of thelearned helplessness phenomenon is thatprior training to escape in the shuttle boxeliminates the interfering effects of sub-sequent exposure to inescapable shock. InExperiment 2 Weiss et al. gave rats 25trials of FR-1 shuttle box training 24 hoursbefore a 3.5 minute cold swim. The ratswere again tested in the shuttle box 30minutes after the cold swim. The pretrain-ing in the shuttle box reduced the deficitproduced by cold swim. Another feature ofthe learned helplessness phenomenon is thatit has a time course in dogs following asingle session of inescapable shock. In Ex-periment 3 Weiss et al. gave rats a coldswim and then tested them in a shuttle box30 minutes, 2 hours, or 48 hours later. Witha 48-hour delay, there was no longer anydetectable effect of the cold swim. Further,rats tested at the 30-minute point were re-tested 48 hours later. A deficit was stillpresent upon retest, as was the case withlearned helplessness. These three experi-ments led Weiss et al. to conclude that coldswim duplicates the effects of inescapableshocks.

30 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
The next six experiments employed a dif-ferent strategy. In these experiments, Weisset al. attempted to find situations in whichthe motor deficit and the learned helpless-ness hypotheses make different or opposedpredictions. The motor activation deficithypothesis implies that the magnitude and thelikelihood of finding a failure to learn toescape depends on the amount of motoractivity required to escape. Weiss et al.claimed that the learned helplessness hy-pothesis would not expect the amount ofeffort required to escape to be a potentvariable. Experiment 4 by Weiss et al.demonstrated that the detrimental effects ofcold swim are strengthened by increasing theheight of the barrier in the shuttle box, andExperiment 5 showed that cold swim did notproduce a deficit in learning to escape shockwhen the escape response only required re-strained rats to poke their nose through ahole in the front of the tube in which theywere confined. In Experiment 6 Weisset al. attempted to demonstrate that exposureto inescapable shock interferes with acquisi-tion of escape behavior in their FR-1 testtask. They mentioned that they have haddifficulty in producing such an effect, butshowed that a 50-min. exposure to 4.0 mA2.0-sec shocks occurring every 20 sec pro-duces an escape deficit when the rat is tested30 min later. Experiment 7 showed thatsuch shocks interfered with shuttle box butnot with nose-poke acquisition, as predictedby the motor deficit hypothesis.
The next three experiments by Weiss etal. are based on data (Zigmond & Harvey,1970) which indicate that the central nora-drenergic response to stress habituates withrepeated exposure to stress. That is, thedegree of norepinephrine depletion producedby a stressor is much reduced if the organ-ism has been repeatedly stressed previously.This leads the motor activation deficit hy-pothesis to predict that repeated exposuresto the stressful condition should reduce anysubsequent deficit in a test task. The learnedhelplessness hypothesis surely does not makesuch a prediction. In Experiments 8 and 9Weiss et al. found that 14 sessions of ex-posure to cold swim or inescapable shockeliminated the FR-1 shuttle box escape
deficit produced by cold swim or inescap-able shock. In addition, these experimentsconfirmed the finding that norepinephrine isnot depleted after repeated stress.
A variety of drug treatments were ex-plored by Weiss et al. in the final three ex-periments. Experiment 10 found that tetra-benazine, a relatively nonspecific depletor ofmonoamines, produced poor performance ina shuttle box 30 minutes after administra-tion. Experiment 11 showed that repeatedadministration of tetrabenazine did not af-fect shuttle box performance. Finally,Experiment 12 found that pargyline, a drugwhich inhibits monoamine oxidase and there-fore prevents intraneuronal degradation ofmonoamines, prevented inescapable shockfrom affecting FR-1 shuttle box perform-ance.
Discussion of the motor activation deficithypothesis. We have presented the entirelogical and evidential bases of the motordeficit hypothesis without comment or in-terruption because it presents, at least on thesurface, a convincing case and should beseen as such by the reader. However, wefeel that every step in the argument whichsuggested the motor deficit hypothesis andeach piece of data which seems to supportthe hypothesis is questionable and open toalternative interpretation, and we will dis-cuss each, point by point. Following thisdiscussion we will present a variety of linesof evidence which we feel are inconsistentwith the motor deficit hypothesis.
It will be recalled that a physiological de-pletion hypothesis was first suggested byMiller and Weiss (1969), who argued thatthe time course of the learned helplessnesseffect is not consistent with the notion thatsomething learned during exposure to in-escapable shock is the cause of the effect.They argued that if something is learned itshould still be present 48 hours later. Al-though this point is not crucial to the motordeficit hypothesis, it should be pointed outthat it is highly debatable. Proactive andretroactive interference produce memorylosses in animals (Maier, Allaway, & Gleit-man, 1967; Maier & Gleitman, 1967) aswell as man, and both proactive and retro-active interference effects generally increase

LEARNED HELPLESSNESS: THEORY AND EVIDENCE 31
with time since learning. It is possible thatprior experiences with learning to controlevents could interfere with the retention ofthe learning that shocks are uncontrollableafter only one sesison with inescapableshocks. In addition, there is ample evidencein animals for retention losses over shortintervals of time (see D'Amato, 1973; Spear,1973).
More importantly, Weiss et al. (1975)ignored aspects of the learned helplessnesstime course data which are not as congenialto their hypothesis as is the fact that theeffect does not occur after 48 hours follow-ing a single experience with inescapableshocks. Seligman and Groves (1970) haveshown that the time course of the learnedhelplessness effect is eliminated in dogs ifthe dogs are given four sessions rather thanone session of inescapable shock. Such dogsfailed to learn to escape even when tested7 days after their last exposure to inescap-able shock. Further, if dogs are raised inlaboratory cages and thus deprived of ex-tensive experience controlling events, onlytwo sessions of inescapable shock are re-quired to produce nontransient learned help-lessness effects (Seligman & Groves, 1970).It should be noted that these are preciselythe results that would be expected from amemory interpretation of the time course andare not explicable by the motor deficit hy-pothesis in any obvious way. Norepi-nephrine levels should certainly be recoveredby 7 days following exposure to inescapableshock. Finally, Seligman and Beagley(1975) failed to find a time course of learnedhelplessness with rats as subjects after onlyone session of inescapable shock. Ratsgiven inescapable shock failed to learn toescape when tested 7 days after exposure toa single session of inescapable shock. Thisfinding is particularly damaging to the motordeficit hypothesis because this hypothesisrests on a base of data obtained with rats,not dogs, as subjects. It would be interest-ing if rats raised in a natural environment,rather than in laboratory cages, showed atransient helplessness effect. In summary,the transient effect required by the motordeficit hypothesis occurs only under onecondition—a single session of inescapable
shock with mongrel dogs not raised in thelaboratory. The time course data taken asa whole does not invalidate a learning inter-pretation and may be more consistent withthe learned helplessness hypothesis than withthe motor activation deficit hypothesis.
Of greater importance in establishing theplausibility of the motor activation deficithypothesis are the several demonstrations byWeiss and his colleagues that exposure toinescapable shock is highly stressful and re-duces endogenous norepinephrine levelswhile exposure to escapable shock is lessstressful and does not lead to such a reduc-tion. It is this that allows the motor deficithypothesis to explain why the learned help-lessness effect occurs and is determined bythe controllability of the initial shock. Sev-eral comments are in order. Weiss et al.(1975) argue that the learned helplessnesseffects which we have found are caused bynorepinephrine depletion produced by theinescapable shock which we deliver. There-fore, it is required that Weiss and his col-leagues demonstrate that our inescapableshock conditions produce intense stress andnorepinephrine depletion. It is thus crucialto inquire whether the experiments by Weissand his colleagues employed conditionsroughly similar to ours. Table 3 summarizessome of the relevant aspects of our rat help-lessness experiments and the Weiss studies.As can be seen, the behavioral helplessnessstudies use 1.0-mA shocks with sessionlengths lasting from 1 to 1-J hours. Therat's behavior is tested 24 hours or longerafter the inescapable shock session. Theconditions used by Weiss and his colleaguesbear no resemblance at all to these condi-tions. All but one of the norepinephrinestudies exposed the subjects to a minimum of
\20 hours of inescapable shock, and shock in-tensity was a minimum of 3.0 mA. Further,norepinephrine was typically assayed im-mediately following the end of the inescap-able shock session. It is worth noting thatthe one norepinephrine study which deviatedfrom these parameters, even though stillvery different from our parameters (Weisset al., 1970, Experiment 1), did not finddepletion of norepinephrine in the groupgiven inescapable shocks. This experiment

32 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
TABLE 3SUMMARY OF THE PARAMETERS USED IN THE RAT LEARNED HELPLESSNESS EXPERIMENTS
ExperimentMaximum
shock levelSessionlength
Time between inescapableshock and test
Behavioral helplessness
Maier, Albin, & TestaGoeckner, Greenough, & MaierTesta, Juraska, & MaierMaier & TestaSeligman & BeagleySeligman, Rosellini, & KozakSeligman, Hannum, & Rosellini
1.0mA1.0mA1.0mA1.0mA1.0mA1,0mA1.0mA
IJhr.1 hr.
Hhr.l| hr.IJhr.lihr.IJhr ,
24 hr.24 hr.24 hr.24 hr.24 hr.
S min.; 1 hr. ; 24 hr.65 days
; 168 hr.
Ulcers, steroids
Weiss (1968), Experiment 2Weiss (1971a)Weiss (1971b)Weiss (1971c)
1.6mA4.0 mA4.0 mA4.0mA
21 hr.48 hr.48 hr.48 hr.
12 hr.nonenonenone
Norepinephrine
Weiss, Stone & Harrell, ExWeiss, Pohorecky, Dorros,
Whittlesey, & CaseWeiss, Pohorecky, Emmel,Weiss, Pohorecky, Emmel,
Berkeley, & Jaffe
periment 1Williams, Emmel,
& MillerMcMenima,
3.0mA
3.4mAnot known
not known
3hr.
20 hr.24 hr.
20 hr.
20 min. ; 40 min.
nonenone
2 min.
found a reliable overall difference in norepi-nephrine levels between inescapable, escap-able, and no-shock groups. However, theinescapable group did not differ reliably fromthe no-shock group, and the overall differ-ence was due to an elevation of norepi-nephrine in the escapable shock group.
It should be clear that, in order to ex-plain the learned helplessness effect, themotor activation deficit hypothesis requiresthat a 1-hour session employing 1.0-mAshocks (occurring about one per minute)produce a depletion in norepinephrine whichis still present 24 hours following the termi-nation of the inescapable shock session.There is not a shred of evidence for such aneffect. It is a curious argument that holdsa behavioral deficit which occurs 24 hoursafter a 1-hour session with 1.0-mA shocks isattributable to norepinephrine depletion be-cause norepinephrine levels are low whenmeasured immediately after massive 20-hoursessions with 3.4- or 4.0-mA shocks.
At this point it should be noted that Weisset al. (1975) have restricted their explana-
tion to only the dog helplessness experi-ments, and not the rat helplessness experi-ments. However, all of the data obtainedby Weiss and his colleagues came from rats,and so we feel that a comparison of theirparameters with those used in our rat studieshas been proper. How can one argue thatdata obtained from rats apply to data ob-tained from dogs but not to parallel data ob-tained from rats? Further, the parametersused in our dog studies also differ massivelyfrom those used by Weiss et al.—we havetypically used 64 shocks spread over about1^ hours and have tested the dogs 24 hourslater.
Moreover, Weiss et al, (1975) have re-ported that endogenous levels of norepi-nephrine are depleted in rats given inescap-able shock only if the rats were group housedprior to the experiment. That is, in all ofthe neurochemical studies referred to above,the rats were group housed before treatment.If the rats were individually housed beforetreatment inescapable shock did not depleteendogenous norepinephrine levels. This may

LEARNED HELPLESSNESS: THEORY AND EVIDENCE 33
be because individual housing in rats inducesincreased activity of tyrosine hydroxylase(Segal, Knapp, Kuczenski, & Mandell,1973), a rate-limiting enzyme in the syn-thesis of norepinephrine. It should be notedthat the rats in all of our experiments havebeen individually housed prior to treatment.In fact, Goeckner, Greenough, and Maier(1974) manipulated housing conditions fromweaning to adulthood and found that in-escapable shock produced a learned helpless-ness effect whether the rats were grouphoused or individually housed.
We conclude that the neurochemical datasaid to provide plausibility to the motoractivation deficit explanation of learned help-lessness, in fact, provide little if any support.
We now turn to a consideration of the 12experiments reported by Weiss et al. (1975)as supporting the motor deficit interpreta-tion of the learned helplessness effect. Re-call that the first three experiments weredesigned to show two things: (a) that atreatment which depletes norepinephrine(cold swim) but cannot be said to lead tothe learning that aversive events and re-sponding are independent produces all of thecharacteristics of the learned helplessnessphenomenon; (b) that a procedure whichinvolves the same degree of uncontrol-lability as the first but does not depletenorepinephrine (warm swim) does not pro-duce a learned helplessness effect. We donot feel that the experiments actually demon-strate that which is claimed.
Recall that Weiss et al. compared the ef-fects of a cold swim and a warm swim onFR-1 shuttle box acquisition and arguedthat the learned helplessness hypothesiswould expect equivalent effects from thesetwo treatments since they involve equal un-controllability. This is not correct. Thelearned helplessness hypothesis does notargue that failure to learn to escape shockresults from exposure to sheer uncontrol-lability, but by exposure to uncontrollableaversive events. For example, the learnedhelplessness hypothesis would not expectfailure to escape shock to result from ex-posure to a light presented independently ofbehavior. Cold swim and warm swim are
certainly not equally aversive, and warmswim may not be aversive at all. Further,although we do not wish to argue in thisdirection, it is not clear to us why cold swimcannot be described as a condition in whichescape from the highly aversive freezing wa-ter is independent of behavior. Cold swimproduces muscular debilitation while warmswim does not. Thus the trauma in a coldswim may indeed be more uncontrollablethan in a warm swim.
More important than whether the learnedhelplessness hypothesis predicts equivalenteffects of cold and warm swim, we do notbelieve that Weiss et al. (1975) have demon-strated that cold swim produces a behavioraldeficit that is the same as the learned help-lessness effect. Learned helplessness is de-fined as an effect resulting from the uncon-trollability of aversive events. As notedpreviously, FR-1 shuttle box acquisition maynot be sensitive to such effects in rats, and avariety of experimenters have failed to findany effect of inescapable shock on subse-quent FR-1 shuttle box acquisition. Thereare a few experiments which find an effectof inescapable shock on subsequent FR-1shuttle box acquisition (e.g., Weiss et al.,1975, Experiment 6) but these experimentsomit comparison of the effects of equivalentescapable and inescapable shock so it can-not be concluded that a learned helplessnesseffect with FR-1 shuttling has been demon-strated. Since Weiss et al. employed FR-1shuttling as their dependent variable, theymay have been using a test that is not sensi-tive to learned helplessness effects.
Further, Weiss et al. (1975) used a 30-minute interval between cold swim and thebehavioral task, whereas our learned help-lessness experiments use a 24-hour intervalbetween inescapable shock exposure and test.Thus Weiss et al. must demonstrate thatcold swim produces a deficit 24 hours later,not 30 minutes later. Experiment 3 ofWeiss et al. examined the time course of thecold-swim effect, but it does not help ushere because Weiss et al. did not have a 24-hour time point. Their experiment onlyinvolved testing at 30 minutes, 2 hours, or48 hours. An examination of the Weiss et al.

34 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
25
20
15
10
5
COLD 30'—»GOLD 24°"-"°SHOCK 30 —SHOCK 24-—
- .̂ ——•*..,
1 2 3 4 5 6BLOCKS OF 5 TRIALS
FIG. 9. Mean shuttle box FR-1 escape responselatency for rats given either cold swim or inescap-able shocks 3 minutes or 24 hours before the shut-tle box test (Jackson & Maier, unpublished data1975). (Only the rats given cold swim 30 minutesbefore the test failed to respond.)
data reveals that the cold-swim effect wasalmost absent at 2 hours. However, the24-hour point is still needed, and Maier hasnow supplied the missing point. Jacksonand Maier (unpublished data) gave rats 3.5minutes of cold swim in 2 °C water andtested them in a FR-1 shuttle box task 30minutes or 24 hours later. The results canbe seen in Figure 9. Jackson and Maierfound the same debilitation as found byWeiss et al. 30 minutes after cold swim, butthere was no deficit at all if 24 hours hadintervened. It should also be noted that thelearned helplessness effect in rats, the effectsof exposure to uncontrollable shock on sub-sequent escape acquisition in a task sensitiveto controllability, may not show any dissipa-tion with time.
Another aspect of Experiment 3 is worthyof comment. Weiss et al. (1975) note thatdogs that were tested 24 hours after ex-posure to inescapable shock and that failedto learn to escape, would fail to learn againif retested later. That is, the failure be-came permanent. Weiss et al. retested the
rats that failed to learn to escape 30 minutesfollowing cold swim after an additional 48hours. A deficit was still present and Weisset al. concluded that this duplicates the cor-responding characteristic of the learned help-lessness phenomenon in dogs. We do notthink so. In the dog experiments, there wasno improvement at all during retest; thedogs showed no tendency to learn. Thiswas not true of the rats in the Weiss et al.study. There were 25 trials on the retest,and by the last block of 5 trials the responselatency of the cold swim rats was down toabout 7 sec, as compared with about 4 secin a control group. In fact, in a later ex-periment (Experiment 7) the retest effectdisappeared completely, although Weiss etal. did not comment on this aspect of thedata.
The final experiment (Experiment 2 inWeiss et al., 1975) said to reveal the identityof the cold swim and learned helplessnesseffects is the immunization effect. It isworth noting that although Weiss et al.found prior training in the shuttle box toreduce the effects of cold swim on shuttlebox performance, the reduction was notcomplete; some debilitation still occurred.The immunizing effect on learned helpless-ness is complete in both dogs (Seligman &Maier, 1967) and rats (Seligman et al.,1975) when the immunization treatment isgiven in the shuttle box.
We conclude that the experiments pro-vided by Weiss et al. (1975) do not demon-strate that the behavioral deficit producedby cold swim is the same as the deficit pro-duced by the uncontrollable shocks used inthe learned helplessness experiments. Evenmore convincing to us than the discrepancybetween the Weiss et al. and the learned help-lessness experiments are our observations ofsubjects 30 minutes after cold swim, thepoint at which Weiss et al. tested such sub-jects. A severe motor debilitation does ap-pear to be present. The rats have great dif-ficulty moving and simply remain motionlesswhen placed on a table. Further, such ratshave some difficulty righting themselveswhen placed on their sides. When they areremoved from the 2 °C water they are com-

LEARNED HELPLESSNESS: THEORY AND EVIDENCE 35
pletely limp. Stone (1970b) has extensivelystudied the effects of cold swim. Observingthe rats after a 20-minute swim in 14.5 °Cwater, Stone notes:
Shortly after the stress, the swim stressed ratsappear exhausted; they lie in a prostrate position,are unable to move their rear limbs, and have dif-ficulty righting themselves when laid on one side.During the later stages of recovery, the animalsremain immobile in a crouched position and showptosis with continued and violent shivering, (p. 53)
We have observed similar behavior after3.5 minutes in 2 °C water.
The inescapable shocks used in our learnedhelplessness experiments do not produce any-thing like these obvious signs of motor debil-itation. As noted previously, the shockswhich we use have no effect on runway ac-quisition 24 hours later, nor do they affectFR-1 shuttle box performance. We doubtthat Weiss's subjects could even locomote tothe end of a runway 30 minutes after coldswim. Thus we conclude that Weiss et al.(1975) are indeed correct in arguing thatcold swim produces a motor deficit, perhapsvia norepinephrine depletion. However,there is no reason to believe that the levelsof inescapable shock used in the learnedhelplessness dog or rat experiments producea motor deficit 24 hours later.
The factors just discussed are also im-portant in interpreting the subsequent ex-periments by Weiss et al. in which the motoractivation deficit hypothesis and the learnedhelplessness hypothesis were said to makeopposed predictions. Experiments 4 and 5demonstrated that the amount of motoractivity required to perform the task re-sponse given 30 minutes after cold swimdetermined whether a deficit occurred, andthus supported the notion that cold swim re-sults in a temporary motor deficit. Theseexperiments do support this notion, butthere is nothing in the learned helplessnessposition which denies that a cold swim pro-duces a motor deficit. The learned helpless-ness position does not predict this, but thereis no reason why it should.
Experiments 6 and 7 of Weiss et al.(1975) use electric shocks during pretreat-ment, but 150 4.0-mA shocks were de-
livered whereas the learned helplessness ex-periments with rats used 64 to 80 1.0-mAshocks. The level of shock used by Weisset al. is almost of tetanizing intensity andmay indeed lead to a trauma-induced motordeficit 30 minutes later. Our dog experi-ments used 64 6-mA shocks, and testing oc-curred 24 hours later. It is difficult to com-pare shock intensities across species as dif-ferent as dogs and rats, but our observa-tions indicate 4 mA in rats to be much moresevere than 6 mA in dogs. Jackson andMaier (unpublished data) investigatedwhether exposure to shocks characteristic oflearned helplessness experiments would pro-duce the sort of deficit reported by Weisset al. Rats were exposed to 64 1.0-mAshocks and tested in an FR-1 shuttle boxtask either 30 minutes or 24 hours later. Ascan be seen in Figure 10, these shocks didnot produce a deficit even 30 minutes later.Thus Experiments 6 and 7 employed condi-tions very different from those employed inthe learned helplessness experiments withrats and may indeed be correctly interpretedin terms of a motor deficit. The fact that150 4.0-mA shocks produce a motor deficit30 minutes later does not imply that 64 1.0-mA shocks produce a motor deficit 24 hourslater.
The conceptual base for Experiments 8and 9 may indeed be adequate for a test be-tween the motor activation deficit andlearned helplessness explanations of thelearned helplessness phenomenon. Weiss etal. noted that the reduction in norepinephrinelevels produced by exposure to stressorshabituates with repeated exposure to thestressor. Thus if inescapable shock pro-duces a learned helplessness effect becauseit produces a motor deficit through norepi-nephrine depletion, repeated exposure to in-escapable shock should not produce a learnedhelplessness effect. The learned helpless-ness hypothesis certainly does not make thisprediction. It will be recalled that Weisset al. found that rats repeatedly exposed tocold swim or to 4.0-mA shocks did indeedperform well when tested on FR-1 shuttling.On the surface this result seems quite deci-
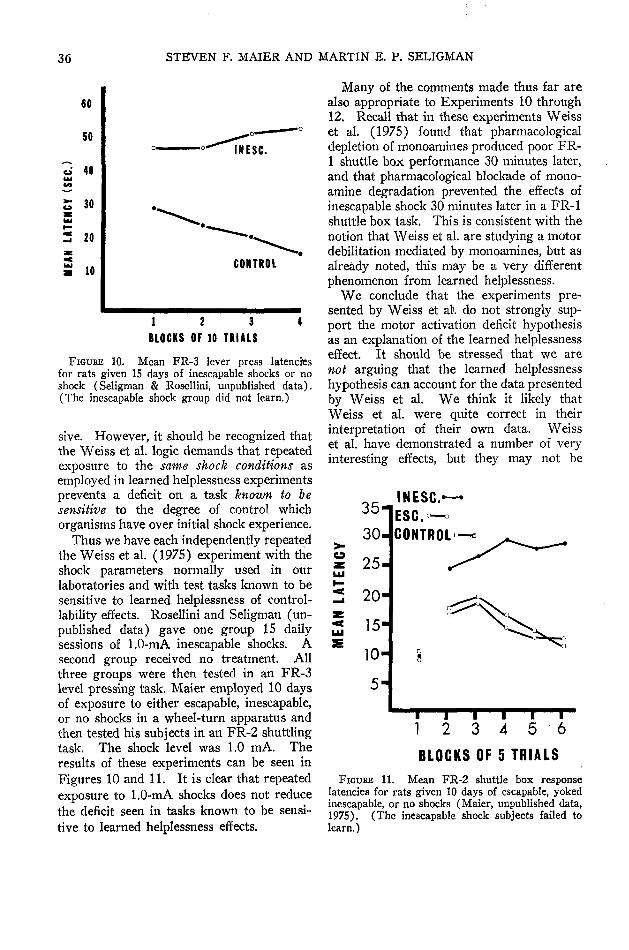
36 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
60
50
• 40
5 30Ul
2 20
i 10
INESC.
CONTROL
1 2 3 4BLOCKS OF 10 T R I A L S
FIGURE 10. Mean FR-3 lever press latenciesfor rats given 15 days of inescapable shocks or noshock (Seligman & Rosellini, unpublished data).(The inescapable shock group did not learn.)
sive. However, it should be recognized thatthe Weiss et al. logic demands that repeatedexposure to the same shock conditions asemployed in learned helplessness experimentsprevents a deficit on a task known to besensitive to the degree of control whichorganisms have over initial shock experience.
Thus we have each independently repeatedthe Weiss et al. (1975) experiment with theshock parameters normally used in ourlaboratories and with test tasks known to besensitive to learned helplessness of control-lability effects. Rosellini and Seligman (un-published data) gave one group 15 dailysessions of 1.0-mA inescapable shocks. Asecond group received no treatment. Allthree groups were then tested in an FR-3level pressing task. Maier employed 10 daysof exposure to either escapable, inescapable,or no shocks in a wheel-turn apparatus andthen tested his subjects in an FR-2 shuttlingtask. The shock level was 1.0 mA. Theresults of these experiments can be seen inFigures 10 and 11. It is clear that repeatedexposure to 1.0-mA shocks does not reducethe deficit seen in tasks known to be sensi-tive to learned helplessness effects.
Many of the comments made thus far arealso appropriate to Experiments 10 through12. Recall that in these experiments Weisset al. (1975) found that pharmacologicaldepletion of monoamines produced poor FR-1 shuttle box performance 30 minutes later,and that pharmacological blockade of mono-amine degradation prevented the effects ofinescapable shock 30 minutes later in a FR-1shuttle box task. This is consistent with thenotion that Weiss et al. are studying a motordebilitation mediated by monoamines, but asalready noted, this may be a very differentphenomenon from learned helplessness.
We conclude that the experiments pre-sented by Weiss et alt do not strongly sup-port the motor activation deficit hypothesisas an explanation of the learned helplessnesseffect. It should be stressed that we arenot arguing that the learned helplessnesshypothesis can account for the data presentedby Weiss et al. We think it likely thatWeiss et al. were quite correct in theirinterpretation of their own data. Weisset al. have demonstrated a number of veryinteresting effects, but they may not be
C9
35
30
25
20J
15
10-
5"
INESC.-ESC.-^CONTROL
• i^ i i i i1 2 3 4 5 6
BLOCKS OF 5 TRIALSFIGURE 11. Mean FR-2 shuttle box response
latencies for rats given 10 days of escapable, yokedinescapable, or no shocks (Maier, unpublished data,1975). (The inescapable shock subjects failed tolearn.)

LEARNED HELPLESSNESS: THEORY AND EVIDENCE 37
closely related to learned helplessness effects.The basic similarity is that in both casesanimals failed to escape shock, but theremay be many roots to producing such adeficit, and they may not reduce to one ex-planation. The deficit produced by expos-ure to extremely traumatic events as usedby Weiss et al. may be produced by a verydifferent mechanism from the deficits pro-duced by exposure to much less traumaticuncontrollable aversive events in the learnedhelplessness experiments.
Other evidence. There are a number offindings not considered by Weiss et al.which are difficult for the motor activationdeficit hypothesis 'to explain. Many of thesehave already been described and only a briefdiscussion is necessary.
1. We have already described experi-ments by Maier and Testa (1975) whichindicate that although inescapably shockedrats are poor at learning FR-2 shuttling,they will learn as well as controls when thereis a brief break in shock after the first re-sponse of the FR-2. In an analogous fash-ion, inescapably shocked rats readily learnedFR-1 shuttling when shock terminated im-mediately after the response, but failed tolearn when shock termination was brieflydelayed. The motor activation deficit hy-pothesis cannot explain why the behavior ofinescapably shocked subjects is so stronglycontrolled by the nature of the contingencybetween responding and shock terminationin the test task. The same amount of motoractivity is required to perform an FR-2 re-sponse whether or not there is a brief breakin shock, but the outcomes are very differ-ent. Similarly, the amount of motor activityrequired to make a single crossing of a shut-tle box is the same whether or not shocktermination is slightly delayed, but the out-come is dependent on this factor. Again,mention should be made that Weiss et al.restricted their explanation to learned help-lessness in the dog, but this is ad hoc and itdoes not seem justifiable to restrict consider-ation to only a small part of the data.
2. Recall that Seligman and Maier (1967)found that prior experience with controllable
shock in a shuttle box eliminated the inter-fering effects of subsequent inescapableshocks on shuttle box performance. Themotor activation deficit hypothesis is able toaccount for this immunizing effect by argu-ing that pretraining in the shuttle box reducesthe amount of motor behavior required dur-ing testing because the subject does nothave to search for the correct response—ithas already been trained. We have pre-sented new data (p. 26) indicating that priorexperience with controllable shock has animmunizing effect even when such prior ex-perience is given with a different responsefrom that later used in testing. The motoractivation deficit hypothesis cannot explainthis effect. Prior training to turn a wheelwith the paws should not reduce the amountof motor activity required to find the corre'ctresponse in a shuttle box.
3. Seligman, Maier, and Geer (1968) andSeligman and Beagley (1975) found thatescape deficits in dogs and rats already fail-ing to escape could be eliminated by forcibleexposure to the escape contingency. It ishard to see how the motor deficit hypothesiscan account for this therapy effect. Doesforcible exposure to the escape contingencyproduce a rapid recovery of norepinephrinelevels? It is interesting to note the Abram-son and Seligman (unpublished data) havefound that forcible exposure to the escapecontingency eliminates the behavioral deficiteven in rats whose norepinephrine levelshave been depleted by injection of a-Methyl-para-tyrosine.
4. We have several times referred to thefact that the learned helplessness effect doesnot have a time course in dogs when thedogs are given a number of sessions of in-escapable shock (Seligman & Groves, 1970)and may not have a time course in rats atall (Seligman & Beagley, 1975). This isdirectly opposed to the expectations of themotor deficit hypothesis.
5. Hannum et al. (in press) found thatrats given four sessions,of inescapable shockshortly after weaning failed to learn to escapeas adults. Such a failure did not result ifthe shocks were escapable. Do four sessions

38 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
of inescapable shock shortly after weaningproduce a permanent depletion of norepi-nephrine ?
6. Although Weiss et al. (1975) haveexplicitly limited the score of their hy-pothesis to learned helplessness experimentsemploying electric shocks to animals (even,on an ad hoc basis, to dogs), mention shouldbe made that there are a large number ofexperiments outside this domain which canbe integrated by the learned helplessnesshypothesis but not by the motor activationdeficit hypothesis. Examples are the effectsof exposure to inescapable noise and un-solvable problems on problem-solving be-havior in humans (see p. 13), and the pos-sible analogues to shock-induced learnedhelplessness produced by delivery of appeti-tive events independently of behavior (see p.12) in animals.
CONCLUSIONS
We feel that the burden of the evidence isthat the motor theories here discussed cannotaccount for the learned helplessness phe-nomenon. It seems to us that the centralshortcoming of these theories is that theyare performance theories. That is, theyclaim that inescapably shocked subjects arelater less likely to emit the test task response,not that they are less likely to learn from anexposure to the contingency between re-sponding and relief if they do respond. Thelearned helplessness hypothesis also arguesfor a performance deficit (the reduced incen-tive motivation proposition), but in additionargues that learning will be undermined (theassociative interference proposition). Wefeel that the Maier and Testa (1975) ex-periments (see p. 24) offer conclusive evi-dence that exposure to inescapable shockhas an effect on associative processes as wellas on motivational processes.
This should not be taken to mean that wefeel that the processes discussed by Brace-well and Black (1974), Anisman and Wal-ler (1973), and Weiss et al. (1975) do notexist and do not influence behavior. Organ-isms can, of course, learn motor responses inone situation that are able to interfere withthe acquisition of motor responses in a dif-
ferent situation. Further, the data pre-sented by Weiss et al. do convincingly pointto the conclusion that severe trauma inducesa transient motor deficit perhaps mediatedby norepinephrine depletion. What we dis-pute is that these processes are sufficient toaccount for all or even many of the learnedhelplessness effects. We also dispute thatthese hypotheses handle the data better thanthe learned helplessness hypothesis, and wefeel that a stronger case has been presentedfor the learned helplessness hypothesis.
It should be noted, however, that thelearned helplessness hypothesis is not with-out its problems. We will list those thatloom largest.
1. The time course of learned helplessnesseffects has yet to be satisfactorily explained.Recall that one session of inescapable shockproduces only a transient effect in dogs, whilefour sessions of inescapable shock seem toproduce a more permanent effect. More-over, there are data to indicate that thelearned helplessness effect may be perma-nent in rats even after one session of inescap-able shock.
This pattern of results is difficult to ex-plain. The learned helplessness theorycould argue (see pp. 30-31) that previousexperience controlling events proactively in-terferes with the memory that shock is ines-capable. Proactive interference is known toincrease with time from learning and todecrease with additional training, and so thedog effects can be explained. But, why dorats show a permanent effect after one ses-sion? A possibility concerns the rearingconditions of our subjects. Our dogs werenot laboratory- or cage-reared, while the ratswe used were raised in cages. Perhaps thisreduced the likelihood that the rats en-countered many immunizing experienceswith controllable events. Consistent withthe argument, Seligman and Groves (1970)found that only two sessions of inescapableshock were required to produce a permanentlearned helplessness effect in cage-rearedbeagles while four sessions were required toproduce a permanent effect in mongrelsraised outside the laboratory. However,such an explanation of the time course effects
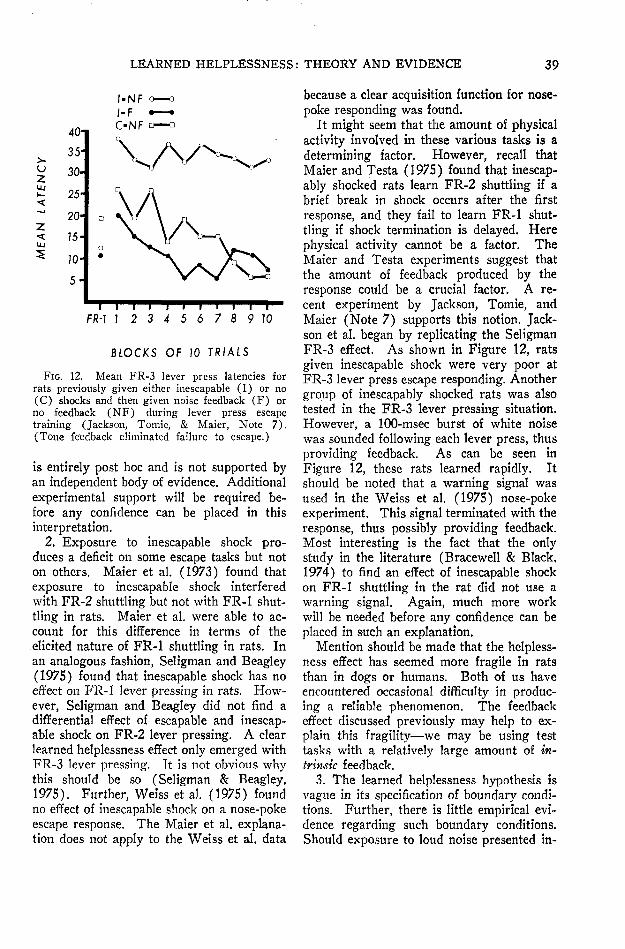
LEARNED HELPLESSNESS: THEORY AND EVIDENCE 39
uzLUI—<• 1
Z
40'
35'
30-\
25
20
IS-
10-
5
FR-1 1 2 3 4 5 6 7 8 9 10
BLOCKS OF 10 T R I A L S
FIG. 12. Mean FR-3 lever press latencies forrats previously given either inescapable (I) or no(C) shocks and then given noise feedback (F) orno feedback (NF) during lever press escapetraining (Jackson, Tomie, & Maier, Note 7).(Tone feedback eliminated failure to escape.)
is entirely post hoc and is not supported byan independent body of evidence. Additionalexperimental support will be required be-fore any confidence can be placed in thisinterpretation.
2. Exposure to inescapable shock pro-duces a deficit on some escape tasks but noton others. Maier et al. (1973) found thatexposure to inescapable shock interferedwith FR-2 shuttling but not with FR-1 shut-tling in rats. Maier et al. were able to ac-count for this difference in terms of theelicited nature of FR-1 shuttling in rats. Inan analogous fashion, Seligman and Beagley(1975) found that inescapable shock has noeffect on FR-1 lever pressing in rats. How-ever, Seligman and Beagley did not find adifferential effect of escapable and inescap-able shock on FR-2 lever pressing. A clearlearned helplessness effect only emerged withFR-3 lever pressing. It is not obvious whythis should be so (Seligman & Beagley,1975). Further, Weiss et al. (1975) foundno effect of inescapable shock on a nose-pokeescape response. The Maier et al. explana-tion does not apply to the Weiss et al. data
because a clear acquisition function for nose-poke responding was found.
It might seem that the amount of physicalactivity involved in these various tasks is adetermining factor. However, recall thatMaier and Testa (1975) found that inescap-ably shocked rats learn FR-2 shuttling if abrief break in shock occurs after the firstresponse, and they fail to learn FR-1 shut-tling if shock termination is delayed. Herephysical activity cannot be a factor. TheMaier and Testa experiments suggest thatthe amount of feedback produced by theresponse could be a crucial factor. A re-cent experiment by Jackson, Tomie, andMaier (Note 7) supports this notion. Jack-son et al. began by replicating the SeligmanFR-3 effect. As shown in Figure 12, ratsgiven inescapable shock were very poor atFR-3 lever press escape responding. Anothergroup of inescapably shocked rats was alsotested in the FR-3 lever pressing situation.However, a 100-msec burst of white noisewas sounded following each lever press, thusproviding feedback. As can be seen inFigure 12, these rats learned rapidly. Itshould be noted that a warning signal wasused in the Weiss et al. (1975) nose-pokeexperiment. This signal terminated with theresponse, thus possibly providing feedback.Most interesting is the fact that the onlystudy in the literature (Bracewell & Black,1974) to find an effect of inescapable shockon FR-1 shuttling in the rat did not use awarning signal. Again, much more workwill be needed before any confidence can beplaced in such an explanation.
Mention should be made that the helpless-ness effect has seemed more fragile in ratsthan in dogs or humans. Both of us haveencountered occasional difficulty in produc-ing a reliable phenomenon. The feedbackeffect discussed previously may help to ex-plain this fragility—we may be using testtasks with a relatively large amount of in-trinsic feedback.
3. The learned helplessness hypothesis isvague in its specification of boundary condi-tions. Further, there is little empirical evi-dence regarding such boundary conditions.Should exposure to loud noise presented in-

40 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
dependently of behavior interfere with theacquisition of responses that escape electricshock? Should exposure to uncontrollableshock interfere with the acquisition of re-sponses that procure food ? Should exposureto noncontingent food interfere with the ac-quisition of responses that terminate shock?Should exposure to uncontrollable shockinterfere with Pavlovian conditioning ? Manyfurther questions of this sort could easily beposed. The learned helplessness hypothesisdoes not make any predictions regarding theresults of such experiments. Further, thereare very few experiments directed at answer-ing these questions. The learned helpless-ness hypothesis will have to become morespecific, and experiments designed to de-lineate these boundary conditions will haveto be conducted.
4. A final problem concerns the relation-ship between the subject's perception of in-dependence between responding and out-come and the conditions which lead to sucha perception (see p. 17). In our experi-ments we have arranged conditions in whichthe relationship between responding and out-come is objectively independent. However,subjects do not always respond to randomrelationships as if they were random • (e.g.,Bruner & Revusky, 1961; Hake & Hyman,1953; Naylor & Clark, 1968), and it is wellknown that subjective probabilities do notreflect objective probabilities with accuracy(cf. Kahneman & Tversky, 1972). Thus wewill have to specify the conditions underwhich the perception of independence devel-ops—only these conditions should lead tolearned helplessness.
Before closing a few comments are inorder.
Tt must seem to the reader that a greatdeal of theoretical confusion surrounds thelearned helplessness phenomenon. A num-ber of investigators have proposed alterna-tive accounts of the same behavioral phe-nomenon and have presented experimentalevidence which they feel supports their posi-tion and refutes the learned helplessness hy-pothesis. We, on the other hand, havequestioned the implications of these experi-
ments for the learned helplessness hypothe-sis. Why all this confusion? We feel thata large part of this confusion stems from anerror of inference noted previously. Thelogical structure of many of the experimentswhich have been presented as inconsistentwith the learned helplessness hypothesis havethe following form: (a) A procedure isfound which produces an escape deficit, (b)this procedure is hard to interpret in termsof the learning of independence between re-sponding and shock termination, and (c)therefore the learned helplessness hypothesisis contradicted. This seems to us to be poorlogic. The learned helplessness hypothesisdoes not argue that all escape deficits areproduced by learning that responding andshock termination are independent. Thereare undoubtedly many ways to produce poorescape performance and there is no reason toexpect that all of these operate through asingle mechanism.
Just as there may be many different waysto produce escape-learning deficits, exposureto inescapable shock may itself produce avariety of changes in the organism, not justone. Under some conditions, organismsmight learn a motor response, stress mightbe induced, and the organism might learnthat its behavior is independent of shocktermination. A complete explanation of theinfluence of exposure to inescapable aversiveevents on later behavior might involve aconsideration of each of these factors. Thuswe do not view the various theoretical posi-tions that have been here discussed as incom-patible: They might be complementary.The existence of one of these processes doesnot imply the nonexistence of the others.We feel that we have presented convincingevidence for the reality of the processesspecified by the learned helplessness hypothe-sis; however, this evidence does not implythat other processes do not also occur.
It might seem that this makes the learnedhelplessness hypothesis difficult to test, butthis is not so. A variety of predictions made
8 Thanks go to Christopher Peterson for thesereferences.

LEARNED HELPLESSNESS: THEORY AND EVIDENCE 41
by the learned helplessness hypothesis havebeen tested and confirmed (e.g., Hiroto &Seligman, 1975; Maier, 1970; Maier et al.,1973; Maier & Testa, 1975; Overmier &Seligman, 1967; Seligman & Beagley, 1975;Seligman & Groves, 1970; Seligman &Maier, 1967; Seligman, Maier, & Geer,1968; Testa, Juraska, & Maier, 1974) Fail-ures of these predictions would have beenevidence against the learned helplessnessposition. In addition there are many newways in which the hypothesis could be tested.For example the logic, if not the execution,underlying Experiments 8 and 9 of Weiss etal. (1975) seems to us to be adequate to testthe learned helplessness position. The com-ing years should see a number of such testsand a more detailed elaboration of the learnedhelplessness hypothesis.
Finally, we would like to say a word aboutthe cognitive nature of our position. Thelearned helplessness hypothesis has beenstated in cognitive languages, whereas mostof the alternative views have been stated inS-R language. We have found it difficultto even approach the sort of phenomena thatwe have tried to explain within an S-Rframework, and have found the cognitivetheorizing to be more fruitful and to reflectmore accurately those processes that we feelto be reflected in behavior. Many other in-vestigators of learning and motivation inlower organisms seem to have reached asimilar conclusion (e.g., Bolles, 1972; Irwin,1971; Kamin, 1969; Seligman & Johnston,1973; Terry & Wagner, 1975; Wagner,Rudy, & Whitlow, 1973). The next fewyears should determine the value of such anapproach.
REFERENCES NOTES
1. Miller, W., & Seligman, M. E. P. Depression,learned helplessness, and the perception of re-inforcement. Manuscript submitted for publica-tion, 1975.
2. James, W. H. DeKalb survey tests: Studentopinion survey form I-E, I. Author's copyright,
1963.3. Anderson, D. C, Schwendiman, G., & Payne, R.
Prior unsignalled inescapable shock trauma inthe rat. Paper presented at the meeting of theWestern Psychological Association, San Diego,March 1968.
4. Carder, B., & Leaf, R. Personal communication,1973.
5. Weisman, R. Personal communication, 1974.6. Weiss, J. M., Pohorecky, L. A., Dorros, K.,
Williams, S., Emmel, D., Whittlesey, M., &Case, E. Coping behavior and brain norepi-nephrine turnover. Paper presented at the meet-ing of the Eastern Psychological Association,Washington, D.C., May 1973.
7. Jackson, R. L., Tomie, A., & Maier, S. F.Learned helplessness in rats: Effects of re-sponse produced feedback. Paper presented atRocky Mountain Psychological Association,Salt Lake City, May 1975.
REFERENCES
Amsel, A., Rashotte, N. E., & MacKinnon, J. R.Partial reinforcement effects within subject andbetween subjects. Psychological Monographs,1966, 80 (20, Whole No. 628).
Anderson, D. C., Cole, J., & McVaugh, W. Varia-tions in unsignaled inescapable preshock as de-terminants of responses to punishment. Journalof Comparative and Physiological Psychology,1968, 65 (3, Pt. 2).
Anderson, D. C., & Paden, P. Passive avoidanceresponse learning as a function of prior tumblingtrauma. Psychonomic Science, 1966, 4, 129-130.
Anisman, H. Effects of pretraining compatible andincompatible responses on subsequent one-wayand shuttle-avoidance performance in rats. Jour-nal of Comparative and Physiological Psychology,1973, 82, 95-104.
Anisman, H., & Waller, T. G. Facilitative anddisruptive effect of prior exposure to shock onsubsequent avoidance performance. Journal ofComparative and Physiological Psychology, 1972,78, 113-122.
Anisman, H., & Waller, T. G. Effects of inescap-able shock on subsequent avoidance performance:Role of response repertoire changes. BehavioralBiology, 1973, 9, 331-355.
Averill, J. R. Personal control over aversivestimuli and its relationship to stress. Psycho-logical Bulletin, 1973, SO, 286-303.
Averill, J. R., & Rosenn, M. Vigilant and non-vigilant coping strategies and psychophysiologicalstress reactions during the anticipation of electricshock. Journal of Personality and Social Psy-chology, 1972, 23, 128-141. _
Bainbridge, P. L. Learning in the rat: Effect ofearly experience with an unsolvable problem.Journal of Comparative and Physiological Psy-chology, 1973, 82, 301-307.
Bandler, R. J., Jr., Madaras, G. R., & Bern, D. J.Self-observation as a source of pain perception.Journal of Personality and Social Psychology,1968, 9, 205-209.
Behrend, E. R., & Bitterman, M. E. Sidmanavoidance in the fish. Journal of the Experi-mental Analysis of Behavior, 1963, 13, 229-242.

42 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
Bintz, J. Between- and within-subject effect ofshock intensity on avoidance in goldfish (Caras-sius auratus). Journal of Comparative andPhysiological Psychology, 1971, 75, 92-97.
Black, A. H., Young, G. A., & Batenchuk, C.Avoidance training of hippocampal theta wavesin flaxedilized dogs and its relation to move-ment. Journal of Comparative and PhysiologicalPsychology, 1970, 70, 15-24.
Bolles, R. C. Species-specific defense reactions inavoidance learning. Psychological Review, 1970,77, 32-48.
Bolles, R. C. Reinforcement, expectancy, andlearning, Psychological Review, 1972, 79, 394-409.
Bracewell, R. J., & Black, A. H. The effects ofrestraint and noncontingent pre-shock on sub-sequent escape learning in the rat. Learning andMotivation, 1974, 5, 53-69.
Brady, J. V., Porter, R. W., Conrad, D. G., &Mason, J. W. Avoidance behavior and thedevelopment of gastroduodenal ulcers. Journalof the Experimental Analysis of Behavior, 1958,1, 69-72.
Braud, W., Wepmann, B., & Russo, D. Task andspecies generality of the "helplessness" phe-nomenon. Psychonomic Science, 1969, 16, 154-155.
Brimer, C. J., & Kamin, L. J. Disinhibition, habitu-ation, sensitization and the conditioned emotionalresponse. Journal of Comparative and Physio-logical Psychology, 1963, 56, 508-516.
Brookshire, K. H., Littman, R. A., & Stewart, C.N. Residue of shock trauma in the white rat:A three factor theory. Psychological Mono-graphs, 1961, 75, (10, Whole No. 514).
Brown, P., & Jenkins, H. M. Autoshaping of thepigeons key-peck. Journal of the ExperimentalAnalysis of Behavior, 1968, 11, 1-8.
Bruner, A., & Revusky, S. H. Collateral be-havior in humans. Journal of the ExperimentalAnalysis of Behavior, 1961, 4, 349-350.
Catania, A. C. Elicitation, reinforcement, andstimulus control. In R. Glaser (Ed.), Thenature of reinforcement. New York: AcademicPress, 1971.
Church, R. M. Response suppression. In B. A.Campbell & R. M. Church (Eds.), Punishmentand aversive behavior. New York: Appleton-Century-Crofts, 1969.
Church, R. M., LoLordo, V. M., Overmier, J. B.,Solomon, R. L., & Turner, L. H. Cardiac re-sponses to shock in curarized dogs: Effects ofshock intensity and duration, warning signal,and prior experience with shock. Journal ofComparative and Physiological Psychology, 1966,62, 1-7.
Cohen, P. S. Interference effects of escapableshock upon subsequent acquisition of escape-avoidance responding. Journal of Comparativeand Physiological Psychology, 1970, 71, 484-486.
Corah, N. L., & Boffa, J. Perceived control, self-observation, and response to aversive stimula-tion. Journal of Personality and Social Psy-chology, 1970, 16, 1-4.
D'Amato, M. Delayed matching and short-termmemory in monkeys. In G. H. Bower, (Ed.),The psychology of learning and motivation (Vol.7). New York: Academic Press, 1973.
Desiderate, O., & Newman, A. Conditioned sup-pression produced in rats by tones paired withescapable or inescapable shock. Journal of Com-parative and Physiological Psychology, 1971, 77,427-443.
De Toledo, L., & Black, A. H. Effects of pre-shock on subsequent avoidance conditioning.Journal of Comparative and Physiological Psy-chology, 1967, 63, 493-499.
Dinsmoor, J. A., & Campbell, S. L. Escape-from-shock training following exposure to inescap-able shock. Psychological Reports, 1956, 2, 43-49. (a)
Dinsmoor, J. A., & Campbell, S. L. Level of cur-rent and time between sessions as factors in"adaptation" to shock. Psychological Reports1956,2,441-444. (b)
Dweck, C. S. The role of expectations and attribu-tions in the alleviation of learned helplessness.Journal of Personality and Social Psychology,1975, 31, 674-685.
Dweck, C. S., & Repucci, N. D. Learned helpless-ness and reinforcement responsibility in children.Journal of Personality and Social Psychology,1973, 25,109-116.
Elliot, R. Tonic heart rate: Experiments on theeffects of collative variables lead to an< hy-pothesis about its motivational significance. Jour-nal of Personality and Social Psychology, 1969,#211-228.
Engberg, L. A., Hansen, G., Welker, R. L., &Thomas, D. R. Acquisition of key-pecking viaautoshaping as a function of prior experience:"Learned laziness"? Science, 1973, 178, 1002-1004.
Ferster, C. B., & Skinner, B. F. Schedules of Re-inforcement. New York: Appleton-Century-Crofts, 1957.
Fosco, F., & Geer, J. H. Effects of gaining con-trol over aversive stimuli after differing amountsof no control. Psychological Reports, 1971, 29,1153-1154.
Frumkin, K., & Brookshire, K. H. Conditionedfear training and later avoidance learning ingoldfish. Psychonomic Science, 1969, 16, 159-160.
Gamzu, E. R., Williams, D. A., & Schwartz, BPitfalls of organism and concept: "Learnedlaziness"? Science, 1973, 101, 367-368.
Geer, J. H., Davison, G. C., & Gatchel, R. I.Reduction of stress in humans through non-veridical perceived control of aversive stimula-tion. Journal of Personality and Social Psy-chology, 1970, 16, 731-738.

LEARNED HELPLESSNESS: THEORY AND EVIDENCE 43
Geer, J. H., & Maisel, E. Evaluating the effectsof the prediction-control confound. Journal ofPersonality and Social Psychology, 1972, 23, 314-319.
Gibbon, J., Berryman, R., & Thompson, R. L.Contingency spaces and measures in classical andinstrumental conditioning. Journal of the Ex-perimental Analysis of Behavior, 1974, 21, 585-605.
Glass, D. C., & Singer, J. E. Urban stress: Ex-periments on noise and social stressors. NewYork: Academic Press, 1972.
Goeckner, D. J,, Greenough, W. T., & Maier, S.F. Escape learning deficit after overcrowdedrearing in rats: Test of a helplessness hypothe-sis. Bulletin of the Psychonomic Society, 1974,3, 54-56.
Hake, H. W., & Hyman, R. Perception of thestatistical structure of a random series of binarysymbols. Journal of Experimental Psychology,1953, 45, 64-74.
Hannum, R. D., Rosellini, R. A., & Seligman, M.E. P. Retention and immunization of learnedhelplessness from weaning to adulthood. Devel-opmental Psychology, in press.
Hearst, E. Stress induced breakdown of an ap-petitive discrimination. Journal of the Experi-mental Analysis of Behavior, 1965, 8, 135-146.
Herman, Z. The effects of noradrenaline on rat'sbehaviour. Psychopharmacologia, 1970, 16, 369-374.
Hiroto, D. S. Locus of control and learned help-lessness. Journal of Experimental Psychology,1974, 102, 187-193.
Hiroto, D. S., & Seligman, M. E. P. Generalityof learned helplessness in man. Journal ofPersonality and Social Psychology, 1975, 31, 311-327.
Hokanson, J. E., DeGood, D. E., Forrest, M. S.,& Brittain, T. M. Availability of avoidance be-haviors in modulating vascular-stress responses.Journal of Personality and Social Psychology,1971,19, 60-68.
Honig, W. K. (Ed.). Operant conditioning: Re-search and application. New York: Appleton-Century-Crofts, 1966.
Irwin, F. W. Intentional behavior and motiva-tion: A cognitive theory, Philadelphia: Lip-pincott, 1971.
Kahneman, D., & Tversky, A. Subjective prob-ability: A judgment of representativeness.Cognitive Psychology, 1972, 3, 430-454.
Kamin, L. J. Predictability, surprise, attention,and conditioning. In B. A. Campbell & R. M.Church (Eds.), Punishment and aversive be-havior, New York: Appleton-Century-Crofts,1969.
Kelley, H. H. Attribution theory in social psy-chology. In D. Levine (Ed.), Nebraska Sym-posium on Motivation. Lincoln: University ofNebraska Press, 1967.
Kelley, H. H. Causal schemata and the attribu-tion process. Morristown, N. J.: General Learn-ing Press, 1972.
Kemler, D., & Shepp, B. The learning and trans-fer of dimensional relevance and irrelevance inchildren. Journal of Experimental Psychology,1971, 90, 120-127.
Klein, D. C., Fencil-Morse, E., & Seligman, M. E.P. Depression, learned helplessness, and the at-tribution of failure. Journal of Personality andSocial Psychology, in press.
Krantz, D. S., Glass, D. C., & Snyder, M. L.Helplessness, stress level, and the coronary pronebehavior pattern. Journal of Experimental SocialPsychology, 1974, 10, 284-300.
Langer, E. J. The illusion of control. Unpub-lished doctoral dissertation, Yale University,1974.
Lazarus, R. S. Psychological stress and the copingprocess. New York: McGraw-Hill, 1966.
Lefcourt, H. N. Internal versus external controlof reinforcement: A review. PsychologicalBulletin, 1966, 65, 206-220.
Liddell, H. S., James, W. T., & Anderson, O. D.The comparative physiology of the conditionedmotor reflex: Based on experiments with thepig, dog, sheep, goat and rabbit. ComparativePhysiology Monograph, 1934, 11(1}, Serial No.89.
Lindner, M. Hereditary and environmental in-fluences upon resistance to stress. Unpublisheddoctoral dissertation, University of Pennsylvania,1968.
Looney, T. A., & Cohen, P. S. Retardation ofjump-up escape responding in rats pretreatedwith different frequencies of noncontingentelectric shock. Journal of Comparative andPhysiological Psychology, 1972, 78, 317-322.
Loucks, R. B. The experimental delimitation ofneural structures essential for learning: Theattempt to condition striped muscle responseswith faradization of the sigmoid gyri. Journalof Psychology, 1935, 1, 5-44.
MacKintosh, N. J. Stimulus selection: Learningto ignore stimuli that predict no change in rein-forcement. In R. A. Hindle & J. Stevenson-Hinde (Eds.), Constraints on learning. NewYork: Academic Press, 1973.
Maier, S. F. Failure to escape traumatic shock:Incompatible skeletal motor response or learnedhelplessness? Learning and Motivation, 1970,1, 157-170.
Maier, S. F., Albin, R. W., & Testa, T. J. Fail-ure to learn to escape in rats previously exposedto inescapable shock depends on nature of escaperesponse. Journal of Comparative and Physio-logical Psychology, 1973, 85, 581-592.
Maier, S. F., Allaway, T. A., & Gleitman, H.Proactive inhibition in rats after prior reversal:A critique of the spontaneous recovery hypotlie-sis. Psychonomic Science, 1967, 9, 63-65.

44 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
Maier, S. F., Anderson, C., & Lieberman, D. A.Influence of control of shock on subsequentshock-elicited aggression. Journal of Compara-tive and Physiological Psychology, 1972, 81, 94-100.
Maier, S. F., & Gleitman, H. Proactive interfer-ence in rats. Psychonomic Science, 1967, 7, 25-26.
Maier, S. F., Seligman, M. E. P., & Solomon, R.L. Pavlovian fear conditioning and learnedhelplessness. In B. A. Campbell & R. M. Church(Eds.), Punishment. New York: Appleton-Century-Crofts, 1969.
Maier, S. F., & Testa, T. J. Failure to learn toescape by rats previously exposed to inescapableshock is partly produced by associative inter-ference. Journal of Comparative and Physio-logical Psychology, 1975, 88, 554-564.
Masserman, J. H. Behavior and neurosis. Chicago:University of Chicago Press, 1943.
Masserman, J. H. The principle of uncertainty inNeurotegenesis. In H. D. Kimmel (Ed.),Experimental psychopathology. New York:Academic Press, 1971.
McCulloch, T. L., & Bruner, J. S. The effects ofelectric shock upon subsequent learning in therat. Journal of Psychology, 1939, 7, 333-336.
Mellgren, R. L., & Ost, J. W. P. Discriminativestimulus preexposure and learning of an operantdiscrimination in the rat. Journal of Compara-tive and Physiological Psychology, 1971, 77, 179-187.
Miller, S., & Konorski, J. Sur une forme particu-liere des reflexes conditionals. Comptes rendushebdomadaires des stances et memoires de laSociete de Biologie et filiales, 1928, 99, 1155-1157.
Miller, W., & Seligman, M. E. P. Depression inhumans. Journal of Abnormal Psychology, 1975,84,228-238.
Miller, W., & Weiss, J. M. Effects of somatic orvisceral responses to punishment. In B. A.Campbell & R. M. Church (Eds.), Punishmentand aversive behavior. New York: Appleton-Century-Crofts, 1969.
Moore, B. R. The role of directed Pavlovian re-actions in simple instrumental learning in thepigeon. In R. A. Hinde & J. Stevenson-Hinde(Eds.), Constraints on learning. New York:Academic Press, 1973.
Moot, S. A., Ceballa, R. P., & Crabtree, J. M.Instrumental control and ulcertation in rats.Journal of Comparative and Physiological Psy-chology, 1970, 71, 405-410.
Mowrer, O. H., & Viek, P. An experimentalanalogue of fear from a sense of helplessness.Journal of Abnormal and Social Psychology,1948, 43, 193-200.
Mullin, A. D., & Mogenson, G. J. Effects of fearconditioning on avoidance learning. Psychologi-cal Reports, 1963, 13, 707-710.
Naylor, J. C., & Clark, R. D. Intuitive interfer-
ence strategies in interval learning tasks as afunction of validity magnitude and sign. Organ-isational Behavior and Human Performance.1968, 3, 378-399.
Overmier, J. B. Interference with avoidance be-havior: Failure to avoid traumatic shock. Jour-nal of Experimental Psychology, 1968, 78, 340-343.
Overmier, J. B., & Seligman, M. E. P. Effects ofinescapable shock upon subsequent escape andavoidance learning. Journal of Comparative andPhysiological Psychology, 1967, 63, 28-33.
Padilla, A. M. Effects of prior and interpolatedshock exposures on subsequent avoidance learn-ing by goldfish. Psychological Reports, 1973,32, 451-456.
Padilla, A. M., Padilla, C., Ketterer, T., & Gia-colone, D. Inescapable shocks and subsequentavoidance conditioning in goldfish (Carrasiusauratus). Psychonomic Science, 1970, 20, 295-296.
Pavlov, I. P. Conditioned reflexes. New York:Dover, 1927.
Payne, R. J. Alterations of Sidman avoidancebaselines by CS paired with avoidable or un-avoidable shock. Psychological Reports, 1972,31, 291-294.
Payne, R., Anderson, D. C., & Murcurio, J. Pre-shock-produced alterations in pain-elicited fight-ing. Journal of Comparative and PhysiologicalPsychology, 1970, 71, 258-266.
Pervin, L. A. The need to predict and controlunder conditions of threat. Journal of Person-ality, 1963, 31, 570-585.
Powell, D. A., & Creer, T. L. Interaction of devel-opmental and environmental variables in shock-elicited aggression. Journal of Comparative andPhysiological Psychology, 1969, 69, 219-225.
Racinskas, J. R. Maladaptive consequences of lossor lack of control over aversive events. Un-published doctoral dissertation, Waterloo Uni-versity, Ontario, Canada, 1971.
Rech, R. H., Bovys, H. K., & Moore, K. Altera-tions in behavior and brain catecholamine levelsin rats treated with a-methyltyrosine. Journalof Pharmacology and Experimental Therapeutics,1966, 153, 412-419.
Rescorla, R. A. Pavlovian conditioning and itsproper control procedures. Psychological Re-view, 1967, 74, 71-79.
Rescorla, R. A. Probability of shock in the pres-ence and absence of the CS in fear conditioning.Journal of Comparative and Physiological Psy-chology, 1968, 66, 1-5.
Rodin, J. Overcrowding, perceived choice andresponse to controllable and uncontrollable out-comes. Journal of Experimental Social Psy-chology, in press.
Rosellini, R., & Seligman, M. E. P. Learned help-lessness and escape from frustration. Journalof Experimental Psychology: Animal BehaviorProcesses, 1975, 1, 149-158.

LEARNED HELPLESSNESS: THEORY AND EVIDENCE 45
Roth, S. The effects of experimentally inducedexpectancies of control: Facilitation of control-ling behavior or learned helplessness? Unpub-lished doctoral dissertation, Northwestern Uni-versity, 1973.
Roth, S., & Bootzin, R. R. Effects of experi-mentally induced expectancies of external con-trol: An investigation of learned helplessness.Journal of Personality and Social Psychology,1974, 29, 253-264.
Roth, S., & Kubal, L. The effects of noncontingentreinforcement on tasks of differing importance:Facilitation and learned helplessness effects.Journal of Personality and Social Psychology,1975, 32, 680-691.
Rotter, J. B. Generalized expectancies for internalversus external control of reinforcement. Psy-chological Monographs, 1966, 80, (1, Whole No.609).
Schwartz, B., & Williams, D. R. Two differentkinds of key peck in the pigeon: Some prop-erties of responses maintained by negative andpositive response-reinforcer contingencies. Jour-nal of the Experimental Analysis of Behavior,1972, 18, 201-216.
Segal, D. S., Knapp, S., Kuczenski, R. T., &Mandell, A. J., The effects of environmentalisolation on behavior and regional rat braintyrosine hydroxylase and tryptophan hydroxylaseactivities. Behavioral Biology, 1973, 8, 47-53.
Seligman, M. E. P. Helplessness. On depression,development, and death. San Francisco: W. H.Freeman, 1975.
Seligman, M. E. P., & Beagley, G. Learned help-lessness in the rat. Journal of Comparative andPhysiological Psychology, 1975, 88, 534-541.
Seligman, M. E. P., & Groves, D. Non-transientlearned helplessness. Psychonomic Science, 1970,19,191-192.
Seligman, M. E. P., & Johnston, J. C. A cognitivetheory of avoidance learning. In F. J. Mc-Guigan & D. B. Lumsden (Eds.), Contemporaryapproaches to conditioning and learning. Wash-ington, D. C.: V. H. Winston, 1973.
Seligman, M. E. P., & Maier, S. F. Failure toescape traumatic shock. Journal of ExperimentalPsychology, 1967, 74, 1-9.
Seligman, M. E. P., Maier, S. F., & Geer, J. Thealleviation of learned helplessness in the dog.Journal of Abnormal and Social Psychology,1968, 73, 256-262.
Seligman, M. E. P., Maier, S. F., & Solomon, R.L. Unpredictable and uncontrollable aversiveevents. In F. R. Brush (Ed.), Aversive condi-tioning and learning. New York: AcademicPress, 1971.
Seligman, M. E. P., Rosellini, R. A., & Kozak, _M.Learned helplessness in the rat: Reversibility,time course, and immunization. Journal of Com-parative and Physiological Psychology, 1975, 88,542-547.
Seward, J. P., & Humphrey, G. L. Avoidance learn-ing as a function of pretraining in the cat.Journal of Comparative and Physiological Psy-chology, 1967, 63, 338-341.
Sines, J. O., Cleeland, C., & Adkins, J. The be-havior of normal and stomach lesion susceptiblerats in several learning situations. Journal ofGenetic Psychology, 1963, 102, 91-94.
Spear, N. E. Retrieval of memory in animals.Psychological Review, 1973, 80, 163-194.
Stone, E. A. Behavioral and neurochemical ef-fects of acute swim stress are due to hypothermia.Life Sciences, 1970, 9, 877-888. (a)
Stone, E. A. Swim-stress-induced inactivity: Re-lation to body temperature and brain norepi-nephrine, and effects of d-amphetamine. Psy-chosomatic Medicine, 1970, 32, 51-59. (b)
Stotland, E. The psychology of hope. San Fran-cisco: Jossey-Bass, 1969.
Stotland, E., & Blumenthal, A. The reduction ofanxiety as a result of the expectation of makinga choice. Canadian Journal of Psychology, 1964,18, 139-145.
Stroebel, C. F. Biologic rhythm correlates ofdisturbed behavior in the rhesus monkey. Biblio-theca Primatologica, 1969, 9, 91-105.
Terry, W. S., & Wagner, A. R, Short-termmemory for "surprising" versus "expected" un-conditioned stimuli in Pavlovian conditioning.Journal of Experimental Psychology: AnimalBehavior Process, 1975, 1, 122-134.
Testa, T. J., Juraska, J. M., & Maier, S. F. Priorexposure to inescapable electric shock in ratseffects extinction behavior after the successfulacquisition of an escape response. Learning andMotivation, 1974, 5, 380-392.
Thomas, D. R., Freeman, F., Svinicki, J. G.,Burr, D. E. S., & Lyons, J. Effects of extra-dimensional training on stimulus generalization.Journal of Experimental Psychology Mono-graph, 1970, 83, (1, Pt. 2).
Thomas, E., & Baiter, A. Learned helplessness:Amelioration of symptoms by cholinergic blockadeof the septum. Science, in press.
Thornton, J. W., & Jacobs, P. D. Learned help-,lessness in human subjects. Journal of Experi-mental Psychology, 1971, 87, 369-372.
Wagner, A. R. Stimulus selection and a "modi-fied continuity theory". In G. H. Bower & J. T.Spence (Eds.), The psychology of learning andmotivation (Vol. 3). New York: AcademicPress, 1969.
Wagner, A. R., Rudy, J. W., & Whitlow, J. W.Rehearsal in animal conditioning. Journal ofExperimental Psychology, 1973, 97, 407-426.
Weiner, B., Frieze, I., Kukla, A., Reed, L., Rest,S., & Rosenbaum, R. M. Perceiving the causesof success and failure. Morristown, N. J.: Gen-eral Learning Press, 1971.
Weiss, J. M. Effects of coping responses on stress.Journal of Comparative and Physiological Psy-chology, 1968, 65, 251-260.

46 STEVEN F. MAIER AND MARTIN E. P. SELIGMAN
Weiss, J. M. Effects of coping behavior in dif-ferent warning signal conditions on stresspathology in rats. Journal of Comparative andPhysiological Psychology, 1971, 77, 1-13. (a)
Weiss, J. M. Effects of coping behavior with andwithout a feedback signal on stress pathology inrats. Journal of Comparative and PhysiologicalPsychology, 1971, 77, 22-30. (b)
Weiss, J. M. Effects of punishing the copingresponse (conflict) on stress pathology in rats.Journal of Comparative and Physiological Psy-chology, 1971, 77, 14-21. (c)
Weiss, J. M., Glazer, H. I., & Pohorecky, L. A.Coping behavior and neurochemical changes: Analternative explanation for the original "learnedhelplessness" experiments. In Relevance of thepsychopathological animal model to the human.New York: Plenum Press, 197S.
Weiss, J. M., Krieckhaus, E. E., & Conte, R.Effects of fear conditioning on subsequent avoid-ance behavior. Journal of Comparative andPhysiological Psychology, 1968, 65, 413-421.
Weiss, J. M., Stone, E. A., & Harrell, N. Copingbehavior and brain norepinephrine in rats. Jour-nal of Comparative and Physiological Psy-chology, 1970, 72, 153-160.
Welker, R. E. "Learned lasiness": A replicationand extension. Unpublished doctoral disserta-tion, University of Colorado, 1974.
Woodbury, C. B. A note on "passive" condition-ing. Journal of General Psychology, 1942, 27,359-361.
Zielinski, K., & Soltysik, S. The effect of pretrain-ing on the acquisition and extinction of the avoid-ance reflex. Acta Blologiae Experlmentalls,1964, 24, 73-87.
Zigmond, M., & Harvey, J. Resistance to centralnorepinephrine depletion and decreased mortalityin rats chronically exposed to electric foot shock.Journal of Neuro-Vlsceral Relations, 1970, 31,373-381.
(Received June 30, 1975)


















