
Lamina Propria CD4+LAP+ Regulatory T Cells Are Increased ... · Alessandro Zandri,a Alessia Butera,...
8
Copyright © 2015 European Crohn’s and Colitis Organisation (ECCO). Published by Oxford University Press. All rights reserved. For permissions, please email: [email protected] 346 Journal of Crohn's and Colitis, 2016, 346–353 doi:10.1093/ecco-jcc/jjv216 Advance Access publication November 20, 2015 Original Article Original Article Lamina Propria CD4+LAP+ Regulatory T Cells Are Increased in Active Ulcerative Colitis but Show Increased IL-17 Expression and Reduced Suppressor Activity Antonella D’Ambrosio, a Andrea Cossu, a,g Antonello Amendola, a Alessandro Zandri, a Alessia Butera, a Massimo Sanchez, b Mauro Biffoni, c Annamaria Pronio, d Chiara Montesani, d Anna Kohn, e Roberta Pica, f Monica Boirivant a a Department of Infectious, Parasitic and Immune-Mediated Diseases, Istituto Superiore di Sanita, Roma, Italy b Department of Cell Biology and Neurosciences, Istituto Superiore di Sanità, Rome, Italy c Department of Hematology, Oncology and Molecular Medicine, Istituto Superiore di Sanità, Rome, Italy d Department of General Surgery, ‘P. Stefanini’, University ‘Sapienza’, Roma, Italy e Division of Gastroenterology, Azienda Ospedaliera S. Camillo-Forlanini, Roma, Italy f IBD, GE Unit, Sandro Pertini Hospital, Roma, Italy g Department of Internal Medicine and Medical Specialties,, University ‘Sapienza,’ Roma, Italy Corresponding author: Monica Boirivant, MD, Department of Infectious, Parasitic and Immune- Mediated Diseases, Istituto Superiore di Sanità, Viale R. Elena, 299 00161, Roma, Italy. Tel: 39-06-49902976; fax: 39-06-49902931; email: [email protected] Abstract Background: A CD4+CD25- regulatory T cell population expressing the surface TGF-β in its latent form LAP+ [latency associated peptide] cells was proved to be protective in experimental colitis and to be suppressive of human peripheral blood [PB] T proliferation. We investigated the frequency and function of lamina propria [LP] CD4+LAP+ T cells in inflammatory bowel disease [IBD] patients. Methods: Specimens from patients undergoing colonoscopy or bowel resection for IBD and colonic cancer were used as source of lamina propria mononuclear cells [LPMC]. The ulcerative colitis [UC] group was divided according to endoscopic activity evaluated with modified Baron Score. IL-17, IFN-γ, IL-10, LAP, and Foxp3 expression in CD3+CD8- [CD4] or CD3+/CD4+ gated cell population was assessed by immunofluorescence. The ability of FACS-sorted LP CD3+CD8-[CD4] LAP+CD25- to inhibit stimulated autologous PB CD3+CD8-[CD4] LAP- CD25- cells proliferation was assessed. Results: LP CD4LAP+ cells were significantly increased, when compared with controls, in active UC patients and not in Crohn’s disease patients. The majority of LP CD4+LAP+ cells were Foxp3-. The percentage of IL-17+ cells in LP CD3+CD8-[CD4] LAP+ cells was significantly higher in active UC patients when compared with controls. LP CD3+CD8-[CD4]LAP+CD25- isolated from UC patients showed reduced or no ability to inhibit autologous PB CD3+CD8-[CD4]LAP-CD25- cell proliferation when compared with controls. Removal of IL-17+ cells from LP CD3+CD8-[CD4] LAP+ cells increases their suppressive ability. Conclusions: The percentage of LP CD4LAP+ cells is increased in active UC, showing reduced suppressor activity due to their increased proportion of intracellular IL-17 expression. Key Words: Inflammatory bowel disease; regulatory CD4+LAP+ cells; IL-17
Transcript of Lamina Propria CD4+LAP+ Regulatory T Cells Are Increased ... · Alessandro Zandri,a Alessia Butera,...

Copyright © 2015 European Crohn’s and Colitis Organisation (ECCO). Published by Oxford University Press. All rights reserved. For permissions, please email: [email protected]
346
Journal of Crohn's and Colitis, 2016, 346–353doi:10.1093/ecco-jcc/jjv216
Advance Access publication November 20, 2015Original Article
Original Article
Lamina Propria CD4+LAP+ Regulatory T Cells Are Increased in Active Ulcerative Colitis but Show Increased IL-17 Expression and Reduced Suppressor ActivityAntonella D’Ambrosio,a Andrea Cossu,a,g Antonello Amendola,a Alessandro Zandri,a Alessia Butera,a Massimo Sanchez,b Mauro Biffoni,c Annamaria Pronio,d Chiara Montesani,d Anna Kohn,e Roberta Pica,f Monica Boirivanta
aDepartment of Infectious, Parasitic and Immune-Mediated Diseases, Istituto Superiore di Sanita, Roma, Italy bDepartment of Cell Biology and Neurosciences, Istituto Superiore di Sanità, Rome, Italy cDepartment of Hematology, Oncology and Molecular Medicine, Istituto Superiore di Sanità, Rome, Italy dDepartment of General Surgery, ‘P. Stefanini’, University ‘Sapienza’, Roma, Italy eDivision of Gastroenterology, Azienda Ospedaliera S. Camillo-Forlanini, Roma, Italy fIBD, GE Unit, Sandro Pertini Hospital, Roma, Italy gDepartment of Internal Medicine and Medical Specialties,, University ‘Sapienza,’ Roma, Italy
Corresponding author: Monica Boirivant, MD, Department of Infectious, Parasitic and Immune- Mediated Diseases, Istituto Superiore di Sanità, Viale R. Elena, 299 00161, Roma, Italy. Tel: 39-06-49902976; fax: 39-06-49902931; email: [email protected]
Abstract
Background: A CD4+CD25- regulatory T cell population expressing the surface TGF-β in its latent form LAP+ [latency associated peptide] cells was proved to be protective in experimental colitis and to be suppressive of human peripheral blood [PB] T proliferation. We investigated the frequency and function of lamina propria [LP] CD4+LAP+ T cells in inflammatory bowel disease [IBD] patients.Methods: Specimens from patients undergoing colonoscopy or bowel resection for IBD and colonic cancer were used as source of lamina propria mononuclear cells [LPMC]. The ulcerative colitis [UC] group was divided according to endoscopic activity evaluated with modified Baron Score. IL-17, IFN-γ, IL-10, LAP, and Foxp3 expression in CD3+CD8- [CD4] or CD3+/CD4+ gated cell population was assessed by immunofluorescence. The ability of FACS-sorted LP CD3+CD8-[CD4] LAP+CD25- to inhibit stimulated autologous PB CD3+CD8-[CD4] LAP- CD25- cells proliferation was assessed.Results: LP CD4LAP+ cells were significantly increased, when compared with controls, in active UC patients and not in Crohn’s disease patients. The majority of LP CD4+LAP+ cells were Foxp3-. The percentage of IL-17+ cells in LP CD3+CD8-[CD4] LAP+ cells was significantly higher in active UC patients when compared with controls. LP CD3+CD8-[CD4]LAP+CD25- isolated from UC patients showed reduced or no ability to inhibit autologous PB CD3+CD8-[CD4]LAP-CD25- cell proliferation when compared with controls. Removal of IL-17+ cells from LP CD3+CD8-[CD4] LAP+ cells increases their suppressive ability.Conclusions: The percentage of LP CD4LAP+ cells is increased in active UC, showing reduced suppressor activity due to their increased proportion of intracellular IL-17 expression.
Key Words: Inflammatory bowel disease; regulatory CD4+LAP+ cells; IL-17

Lamina Propria CD4 LAP+ Regulatory Cells in IBD 347
1. Introduction
Inflammatory bowel disease [IBD] results from an exaggerated T cell response to gut microbiota in genetically predisposed hosts. This response is not adequately counteracted by regulatory CD4+Foxp3+ T cells [Treg], although increased.1 We have recently described both in mice with experimental colitis2 and in humans with ileal pouch-anal anastomosis [IPAA] for ulcerative colitis [UC]3 the occurrence of another type of regulatory T cell in the intestinal lamina propria. This cell is a CD4+ T cell that bears surface TGF-β linked to its latency associated peptide [LAP+ cells] but does not express Foxp3. These cells are increased, in mice, in the presence of increased intes-tinal permeability and during the homeostatic response to a tran-sient increase of intestinal permeability.4,5 Their in vivo regulatory activity has been proven by the fact that they contribute to the pre-vention of 2,4,6-Trinitrobenzenesulfonic acid [TNBS] and adoptive transfer colitis in mice.5,6 Their contribution was found to be crucial for the protection from TNBS colitis, since transfer of CD4+LAP+ depleted cells enriched with CD4+ Foxp3+ cells were not able to prevent TNBS colitis in recipient mice.5 Notable characteristics of these cells are that in mice they require IL-10 and TLR2 for their in vivo expansion2,5 and they express LAP independently of Foxp3 in the presence of TGF-β.7 Initial description of their properties in humans showed that CD4+LAP+ peripheral T cells produce IL-10 and TGF-β and are expanded in vitro by polyclonal stimulation in the presence of IL-2 and IL-8.8 In the present study, we investigated the frequency and function of lamina propria CD4+LAP+ in IBD. We found that they were selectively increased in endoscopically active UC patients, showing a significant increase of IL-17 expres-sion when compared with controls. Functional studies demon-strated that LP CD3+CD8-[ CD4]LAP+CD25- cells isolated from UC patients showed reduced or no ability to inhibit autologous PB CD3+CD8-[CD4]LAP-CD25- cell proliferation when compared with controls. Depletion of CD3+CD8-[CD4] LAP+ IL-17+ cells increases the suppressor ability of CD4 LAP+ cells.
2. Materials and Methods
2.1. Patients and tissues2.1.1. Surgical specimensSurgical specimens were taken from macroscopically inflamed areas of 5 ilea and 5 colons of a total of 10 patients affected by Crohn’s disease, and from colonic macroscopically inflamed areas of 11 patients affected by ulcerative colitis. Mucosal samples were also collected from macroscopically unaffected ileal or colonic areas [6 ileal specimens, 11 colonic specimens] of a total of 17 patients undergoing surgery for colonic cancer.
2.1.2. Biopsy specimensEndoscopic mucosal biopsies were obtained from a total of 23 UC patients [13 males, 10 females, mean age: 48.8 ± 4.5 year] and from a total of 19 control subjects undergoing colonoscopy for colonic cancer screening and suspected functional bowel disorders in which histology confirmed the absence of inflammatory changes. Biopsies were taken from inflamed areas in UC patients with active disease, from previously inflamed areas in UC patients with endoscopic remission [anamnestic information] and in matched areas in con-trol subjects. Diagnosis of Crohn’s disease and ulcerative colitis was ascertained according to established criteria, and site and extent of the disease were confirmed by histology.
2.1.3. PatientsIn UC patients, disease activity was assessed by endoscopic modified Baron Score [mBs].9 Accordingly, patients with an mBs >1 were con-sidered as active patients [12 patients] and 11 patients were consid-ered as in remission. At time of surgery/endoscopy none of the active patients was treated with cyclosporine, azathioprine, or anti-TNF antibodies. Two UC patients in remission were receivinging azathio-prine. In UC patients, disease duration was 11 ± 2.5 years with no difference between active and inactive patients
2.2. Ethical considerationsAll participants provided written informed consent. Ethical approval was provided by the Istituto Superiore di Sanità’s Ethical Committee, reference: Pre-C-899/14.
2.3. Cells isolationLPMC were isolated from freshly resected mucosa or biopsies using a previously described DDT-EDTA collagenase method.3,10 Briefly, strips of mucosa [6–8 g total weight] or biopsies were washed in HBBS free of calcium and magnesium [HBSS-CMF; Hyclone, Europe LTD, Cramlington, UK]. They were then incubated in HBSS-CMF containing 1 mmol/l DTT [Sigma Chemical Co., St. Louis, MO] and antibiotics [penicillin, 100 U/ml; streptomycin, 100 mg/ml; gentamicin, 50 mg/ml; and fungizone, 25 mg/ml] for 15 min [surgical specimens] or 5 min [biopsies] at room temperature. After three washings in HBSS-CMF, the mucosal strips or the biopsies were chopped into pieces and incubated in HBSS-CMF containing 0.75 mmol/l EDTA, 10 mmol/l HEPES buffer, and antibiotics, three times for 45 min each [surgical specimens] or for15 min [biopsies] at 37°C in a humid 5% CO2 atmosphere to remove epithelial cells. After two washes, the pieces were incubated for 10–13 h [surgical specimens] or 2h [biopsies] at 37°C in a humid 5% CO2 atmosphere in complete medium [RPMI 1640 plus 10 mM HEPES buffer, 2 mM l-glutamine, 10% heat-inactivated FCS [Hyclone], and antibiotics] containing 25 U/ml collagenase V [Sigma-Aldrich, Milan, Italy] and 100 µg/ml of DNase [Roche Diagnostics, Mannheim, Germany]. After incubation, the supernatant was collected and washed twice in HBSS-CMF, and LPMC were isolated using a Ficoll–Paque Plus gradient [Amersham Pharmacia Biotech, Uppsala]
2.4. Immunofluorescence studies2.4.1. Antibodies and reagentsAPC Cy7-labelled anti-CD3, PE/Cy7-labelled anti-IL-10, and isotype-matched Ig control were obtained from Biolegend [San Diego, CA, USA]. PE CF594-labelled anti-CD8, V450-labelled anti-IFN-γ and isotype-matched Ig control were obtained from Becton Dickinson Horizon [St José, CA, USA]. APC-labelled anti-CD3, APC-labelled anti-CD8, FITC-labelled anti-CD8, PE-labelled anti-CD25, FITC-labelled anti-CD25, FITC-labelled anti-CD4, FITC-labelled anti-CD161, isotype-matched Ig controls, and Monensin solution [Golgi StopTM], were obtained from BD Pharmingen [St José, CA, USA]. PerCP-labelled anti-LAP [TGF-β1] and isotype-matched Ig control were obtained from R&D Systems [Minneapolis, MN, USA]. APC-labelled anti-Foxp3, Foxp3 staining buffer set, PE-labelled anti-IL-17, and isotype-matched Ig control were obtained from eBioscience [San Diego, CA, USA]. LIVE/DEAD® Fixable Aqua Dead Cell Stain Kit and SYTOX® blue dead cell stain were obtained from Life Technologies [Carlsbad, CA, USA ]. Phorbol-12-myristate-acetate [PMA] and ionomycin were obtained from Sigma-Aldrich.

348 A. D’Ambrosio et al.
2.4.2. Immunofluorescence stainingFor frequency evaluation of CD4+LAP+, represented in Figure 1, isolated LPMC were stained with anti-human-CD3, anti-human-CD4, and anti-human LAP [TGF-β1]. In preliminary experiments, we established that the percentages of LAP+ and Foxp3+ cells were not different before and after PMA-ionomicyn stimulation in the presence of Golgi Stop. Therefore, for evaluation of LAP and Foxp3 expression in combination with intracellular cytokine, LPMC iso-lated from biopsies or surgical specimens were cultured for 4 h in complete medium with PMA [50 ng/ml; Sigma-Aldrich] ionomycin [1 µg/ml; Sigma-Aldrich], and Monensin [0.66 µl/ml GolgiStop; BD Pharmingen]. After stimulation, cells were washed and labelled with LIVE/DEAD® Fixable Aqua Dead Cell Stain for 30 min. At the end of the incubation period, cells were washed and labelled with Ab against surface antigens. Due to the down-modulation of CD4 expression after PMA-ionomycin stimulation, CD8 staining was preferred. Consequently, cells were stained with anti-human-CD3, anti-human CD8, anti-human LAP [TGF-β1], and incubated for 30 min. Cells were then washed, fixed, and permeabilised with fixation/permeabilisation solution. After 40 min, cells were washed with permeabilisation buffer and labelled with anti-human IL-10, anti-human IFN-γ, anti-human IL-17, anti-human Foxp3, or iso-type-matched Ig control for 30 min. Cells were then washed twice with permeabilisation buffer solution and the percentage of viable fluorescent cells was quantified using a FACSAria [BD Biosciences, Milan, Italy]. CD4+ cells were defined as CD3+CD8- cells.
Since CD161+ cell population has been reported to be increased in UC11,12 and indirectly shown to contain sulfatide-reactive Type II NK-T cells,12 in a subset of experiments we also evaluated CD161+ and CD161+ LAP+ cells. As previously reported,11,12 we observed a higher percentage of CD161+ cells in UC than in control samples in CD3+CD8-[CD4] LP cells; however, we observed very few, if any, CD161+LAP+ cells in controls or in active UC samples, suggesting that the great majority of LAP+ cells are not CD161+.
2.3. Cell populations’ purificationIn experiments involving cell-sorting, LPMC and PBMC were stained with anti-CD3, anti-CD8, anti-LAP [TGF-β1], and anti-CD25 abs
and different T cell populations were sorted using a FACSAria Cell sorter [BD Biosciences]. Dead and dying cells were excluded with SYTOX® blue dead cell stain [Life Technologies]. In some experi-ments, LP cells were expanded by in vitro stimulation with anti-CD3/CD28 coated beads [T cell Activation/expansion Kit, Miltenyi Biotech] according to the manufacturer’s instructions. At the end of the culture period, LP CD3+CD8-[CD4]LAP+CD25-IL-17- T cells were sorted after IL17-producing LAP+CD25- LP T cells were identified using the IL-17secretion Assay-Detection Kit [Miltenyi Biotech] according to the manufacturer’s instructions. After exclu-sion of CD25+ cells, the percentages of Foxp3+ cells in sorted LP CD3+CD8-[CD4]LAP+CD25- cell population ranged from 1.5% to 3% and from 0.2% to 1.3% in sorted PB LAP-cells.
2.5. Suppression assayCFSE- labelled PB CD3+CD8-[CD4]LAP-CD25- cells stimulated with aCD3aCD28 and aCD2 coated beads [ ratio 1 bead:2 cells] were cultured in round-bottom 96-well plates for 5 days in the absence or in the presence of autologous LP CD3+CD8-[CD4] LAP+CD25- cells at 2:1 [responder: regulatory ] ratio. Proliferation of PB CD3+CD8-[CD4]LAP-CD25- cells was assessed by the CFSE fading method. The ability of sorted LP CD3+CD8-[CD4] LAP+CD25- cells to inhibit PB CD3+CD8-[CD4]LAP-CD25- cell proliferation was defined as percentage suppression and was cal-culated by the following formula: [proliferation of PB CD3+CD8-[CD4]LAP-CD25- cells] minus [proliferation PB CD3+CD8-[CD4] LAP-CD25- cells co-cultured with LP CD3+CD8-[CD4] LAP+CD25- cells] divided by [proliferation of PB CD3+CD8-[CD4]LAP-CD25- cells ] multiplied by 100. In some experiments, suppressor ability of LP CD3+CD8-[CD4]LAP+CD25-IL-17- T cells was evaluated.
2.6. Statistical analysisData were analysed in the GraphPad Prism statistical PC program [GraphPad Software, San Diego, CA, USA] using the Mann–Whitney U test and Spearman correlation. A level of p < 0.05 was considered statistically significant.
30
20
p<0.005
LAP(% in CD3+CD4+ gated LPMC)
10
LA
P (%
in C
D3+
CD
4+ g
ated
LPM
C)
0
Contro
l ileu
m
CD ileum
Contro
l colo
n
CD colon UC
A B Control colon
1.3%
1.6% 0.9%
0.3%13.4%
Control ileumCou
nt
CD ileum
CD colon UC
Figure 1. Frequency of lamina propria [LP] CD4+LAP+ regulatory T cells is increased in involved tissue of UC and not in involved tissue of CD patients. LPMC were isolated by enzymatic procedure from macroscopically disease-involved tissue of surgical specimens of patients with IBD undergoing intestinal resection. Control LPMC were isolated from surgical specimens of patients undergoing intestinal resection for non-inflammatory conditions. Frequency of LAP+ was evaluated by immunofluorescence in LP CD3+/CD4+ gated cells. A: Frequency of LAP+ cells in controls, CD, and UC surgical specimens. In each series, a line shows the median value. B: Representative immunofluorescence histograms. Markers were fixed according to isotype controls. CD: Crohn’s Disease, UC: ulcerative colitis; LAP, latency associated peptide; LPMC, lamina propria mononuclear cells; IBD, inflammatory bowel disease.
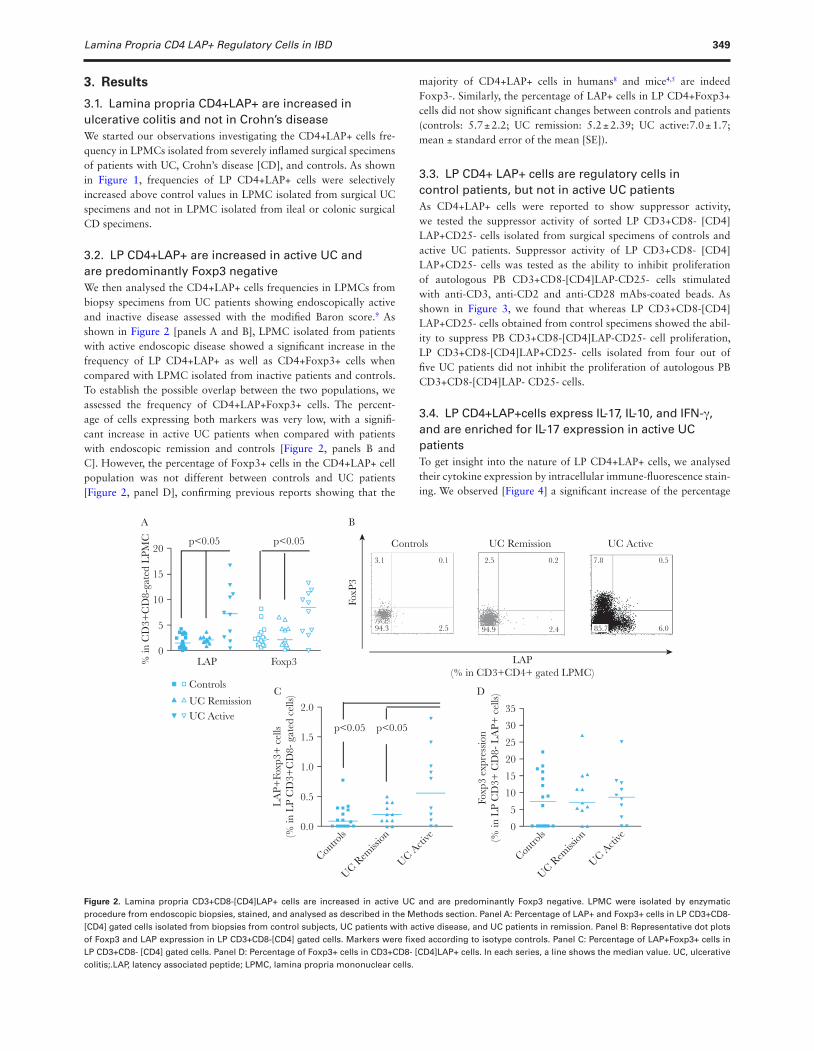
Lamina Propria CD4 LAP+ Regulatory Cells in IBD 349
3. Results
3.1. Lamina propria CD4+LAP+ are increased in ulcerative colitis and not in Crohn’s diseaseWe started our observations investigating the CD4+LAP+ cells fre-quency in LPMCs isolated from severely inflamed surgical specimens of patients with UC, Crohn’s disease [CD], and controls. As shown in Figure 1, frequencies of LP CD4+LAP+ cells were selectively increased above control values in LPMC isolated from surgical UC specimens and not in LPMC isolated from ileal or colonic surgical CD specimens.
3.2. LP CD4+LAP+ are increased in active UC and are predominantly Foxp3 negativeWe then analysed the CD4+LAP+ cells frequencies in LPMCs from biopsy specimens from UC patients showing endoscopically active and inactive disease assessed with the modified Baron score.9 As shown in Figure 2 [panels A and B], LPMC isolated from patients with active endoscopic disease showed a significant increase in the frequency of LP CD4+LAP+ as well as CD4+Foxp3+ cells when compared with LPMC isolated from inactive patients and controls. To establish the possible overlap between the two populations, we assessed the frequency of CD4+LAP+Foxp3+ cells. The percent-age of cells expressing both markers was very low, with a signifi-cant increase in active UC patients when compared with patients with endoscopic remission and controls [Figure 2, panels B and C]. However, the percentage of Foxp3+ cells in the CD4+LAP+ cell population was not different between controls and UC patients [Figure 2, panel D], confirming previous reports showing that the
majority of CD4+LAP+ cells in humans8 and mice4,5 are indeed Foxp3-. Similarly, the percentage of LAP+ cells in LP CD4+Foxp3+ cells did not show significant changes between controls and patients (controls: 5.7 ± 2.2; UC remission: 5.2 ± 2.39; UC active:7.0 ± 1.7; mean ± standard error of the mean [SE]).
3.3. LP CD4+ LAP+ cells are regulatory cells in control patients, but not in active UC patientsAs CD4+LAP+ cells were reported to show suppressor activity, we tested the suppressor activity of sorted LP CD3+CD8- [CD4] LAP+CD25- cells isolated from surgical specimens of controls and active UC patients. Suppressor activity of LP CD3+CD8- [CD4] LAP+CD25- cells was tested as the ability to inhibit proliferation of autologous PB CD3+CD8-[CD4]LAP-CD25- cells stimulated with anti-CD3, anti-CD2 and anti-CD28 mAbs-coated beads. As shown in Figure 3, we found that whereas LP CD3+CD8-[CD4] LAP+CD25- cells obtained from control specimens showed the abil-ity to suppress PB CD3+CD8-[CD4]LAP-CD25- cell proliferation, LP CD3+CD8-[CD4]LAP+CD25- cells isolated from four out of five UC patients did not inhibit the proliferation of autologous PB CD3+CD8-[CD4]LAP- CD25- cells.
3.4. LP CD4+LAP+cells express IL-17, IL-10, and IFN-γ, and are enriched for IL-17 expression in active UC patientsTo get insight into the nature of LP CD4+LAP+ cells, we analysed their cytokine expression by intracellular immune-fluorescence stain-ing. We observed [Figure 4] a significant increase of the percentage
LAP(% in CD3+CD4+ gated LPMC)
0
FoxP
3
Controls UC Remission UC Active
5
10
% in
CD
3+C
D8-
gate
d L
PMC p<0.05
LAP Foxp3
p<0.05
15
20
0.0
Contro
ls
UC Rem
ission
UC Acti
ve
Contro
ls
UC Rem
ission
UC Acti
ve
0.5
LA
P+Fo
xp3+
cel
ls(%
in L
P C
D3+
CD
8- g
ated
cel
ls)
Foxp
3 ex
pres
sion
(% in
LP
CD
3+ C
D8-
LA
P+ c
ells)
1.0
1.5
2.0
p<0.05 p<0.05
0
10
5
15
20
35
Controls
UC RemissionUC Active
25
30
3.1 0.1
94.3 2.5
2.5 7.8 0.5
85.7 6.0
0.2
94.9 2.4
A B
C D
Figure 2. Lamina propria CD3+CD8-[CD4]LAP+ cells are increased in active UC and are predominantly Foxp3 negative. LPMC were isolated by enzymatic procedure from endoscopic biopsies, stained, and analysed as described in the Methods section. Panel A: Percentage of LAP+ and Foxp3+ cells in LP CD3+CD8- [CD4] gated cells isolated from biopsies from control subjects, UC patients with active disease, and UC patients in remission. Panel B: Representative dot plots of Foxp3 and LAP expression in LP CD3+CD8-[CD4] gated cells. Markers were fixed according to isotype controls. Panel C: Percentage of LAP+Foxp3+ cells in LP CD3+CD8- [CD4] gated cells. Panel D: Percentage of Foxp3+ cells in CD3+CD8- [CD4]LAP+ cells. In each series, a line shows the median value. UC, ulcerative colitis;.LAP, latency associated peptide; LPMC, lamina propria mononuclear cells.
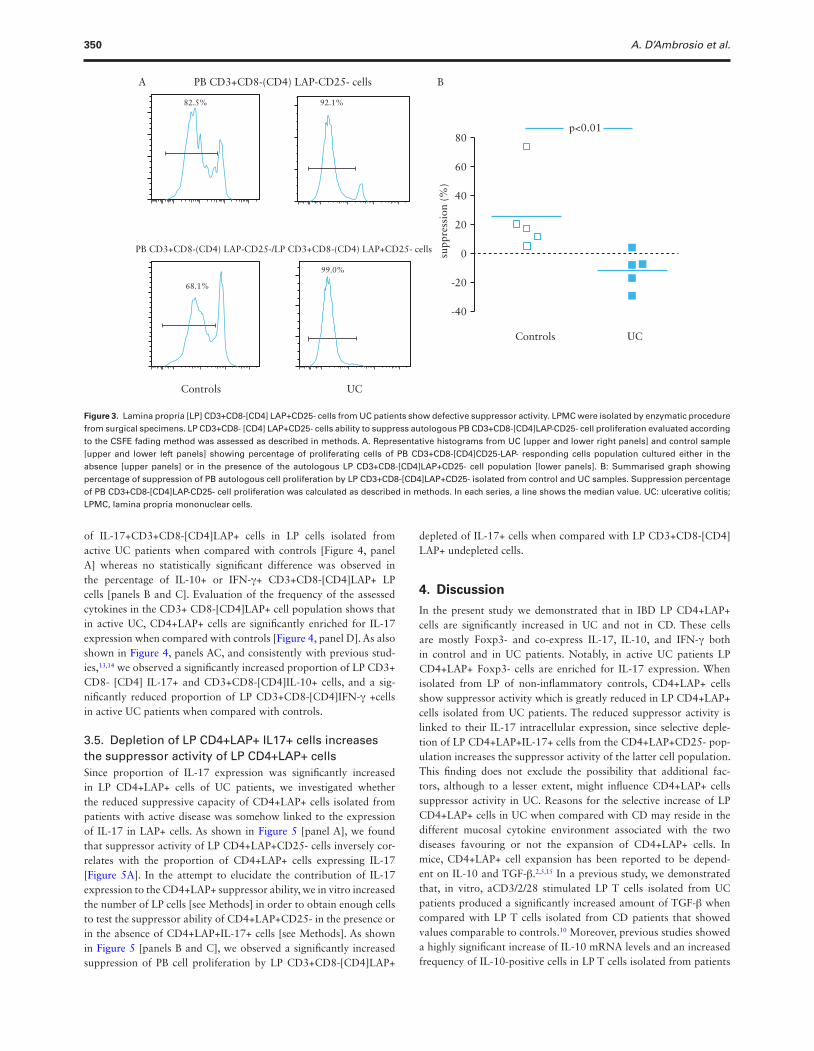
350 A. D’Ambrosio et al.
of IL-17+CD3+CD8-[CD4]LAP+ cells in LP cells isolated from active UC patients when compared with controls [Figure 4, panel A] whereas no statistically significant difference was observed in the percentage of IL-10+ or IFN-γ+ CD3+CD8-[CD4]LAP+ LP cells [panels B and C]. Evaluation of the frequency of the assessed cytokines in the CD3+ CD8-[CD4]LAP+ cell population shows that in active UC, CD4+LAP+ cells are significantly enriched for IL-17 expression when compared with controls [Figure 4, panel D]. As also shown in Figure 4, panels AC, and consistently with previous stud-ies,13,14 we observed a significantly increased proportion of LP CD3+ CD8- [CD4] IL-17+ and CD3+CD8-[CD4]IL-10+ cells, and a sig-nificantly reduced proportion of LP CD3+CD8-[CD4]IFN-γ +cells in active UC patients when compared with controls.
3.5. Depletion of LP CD4+LAP+ IL17+ cells increases the suppressor activity of LP CD4+LAP+ cellsSince proportion of IL-17 expression was significantly increased in LP CD4+LAP+ cells of UC patients, we investigated whether the reduced suppressive capacity of CD4+LAP+ cells isolated from patients with active disease was somehow linked to the expression of IL-17 in LAP+ cells. As shown in Figure 5 [panel A], we found that suppressor activity of LP CD4+LAP+CD25- cells inversely cor-relates with the proportion of CD4+LAP+ cells expressing IL-17 [Figure 5A]. In the attempt to elucidate the contribution of IL-17 expression to the CD4+LAP+ suppressor ability, we in vitro increased the number of LP cells [see Methods] in order to obtain enough cells to test the suppressor ability of CD4+LAP+CD25- in the presence or in the absence of CD4+LAP+IL-17+ cells [see Methods]. As shown in Figure 5 [panels B and C], we observed a significantly increased suppression of PB cell proliferation by LP CD3+CD8-[CD4]LAP+
depleted of IL-17+ cells when compared with LP CD3+CD8-[CD4] LAP+ undepleted cells.
4. Discussion
In the present study we demonstrated that in IBD LP CD4+LAP+ cells are significantly increased in UC and not in CD. These cells are mostly Foxp3- and co-express IL-17, IL-10, and IFN-γ both in control and in UC patients. Notably, in active UC patients LP CD4+LAP+ Foxp3- cells are enriched for IL-17 expression. When isolated from LP of non-inflammatory controls, CD4+LAP+ cells show suppressor activity which is greatly reduced in LP CD4+LAP+ cells isolated from UC patients. The reduced suppressor activity is linked to their IL-17 intracellular expression, since selective deple-tion of LP CD4+LAP+IL-17+ cells from the CD4+LAP+CD25- pop-ulation increases the suppressor activity of the latter cell population. This finding does not exclude the possibility that additional fac-tors, although to a lesser extent, might influence CD4+LAP+ cells suppressor activity in UC. Reasons for the selective increase of LP CD4+LAP+ cells in UC when compared with CD may reside in the different mucosal cytokine environment associated with the two diseases favouring or not the expansion of CD4+LAP+ cells. In mice, CD4+LAP+ cell expansion has been reported to be depend-ent on IL-10 and TGF-β.2,5,15 In a previous study, we demonstrated that, in vitro, aCD3/2/28 stimulated LP T cells isolated from UC patients produced a significantly increased amount of TGF-β when compared with LP T cells isolated from CD patients that showed values comparable to controls.10 Moreover, previous studies showed a highly significant increase of IL-10 mRNA levels and an increased frequency of IL-10-positive cells in LP T cells isolated from patients
PB CD3+CD8-(CD4) LAP-CD25- cells
PB CD3+CD8-(CD4) LAP-CD25-/LP CD3+CD8-(CD4) LAP+CD25- cells
82.5% 92.1%
Controls UC
-40
-20
0
20
p<0.01
supp
ress
ion
(%)
Controls UC
40
60
80
68.1%
99.0%
A B
Figure 3. Lamina propria [LP] CD3+CD8-[CD4] LAP+CD25- cells from UC patients show defective suppressor activity. LPMC were isolated by enzymatic procedure from surgical specimens. LP CD3+CD8- [CD4] LAP+CD25- cells ability to suppress autologous PB CD3+CD8-[CD4]LAP-CD25- cell proliferation evaluated according to the CSFE fading method was assessed as described in methods. A. Representative histograms from UC [upper and lower right panels] and control sample [upper and lower left panels] showing percentage of proliferating cells of PB CD3+CD8-[CD4]CD25-LAP- responding cells population cultured either in the absence [upper panels] or in the presence of the autologous LP CD3+CD8-[CD4]LAP+CD25- cell population [lower panels]. B: Summarised graph showing percentage of suppression of PB autologous cell proliferation by LP CD3+CD8-[CD4]LAP+CD25- isolated from control and UC samples. Suppression percentage of PB CD3+CD8-[CD4]LAP-CD25- cell proliferation was calculated as described in methods. In each series, a line shows the median value. UC: ulcerative colitis; LPMC, lamina propria mononuclear cells.
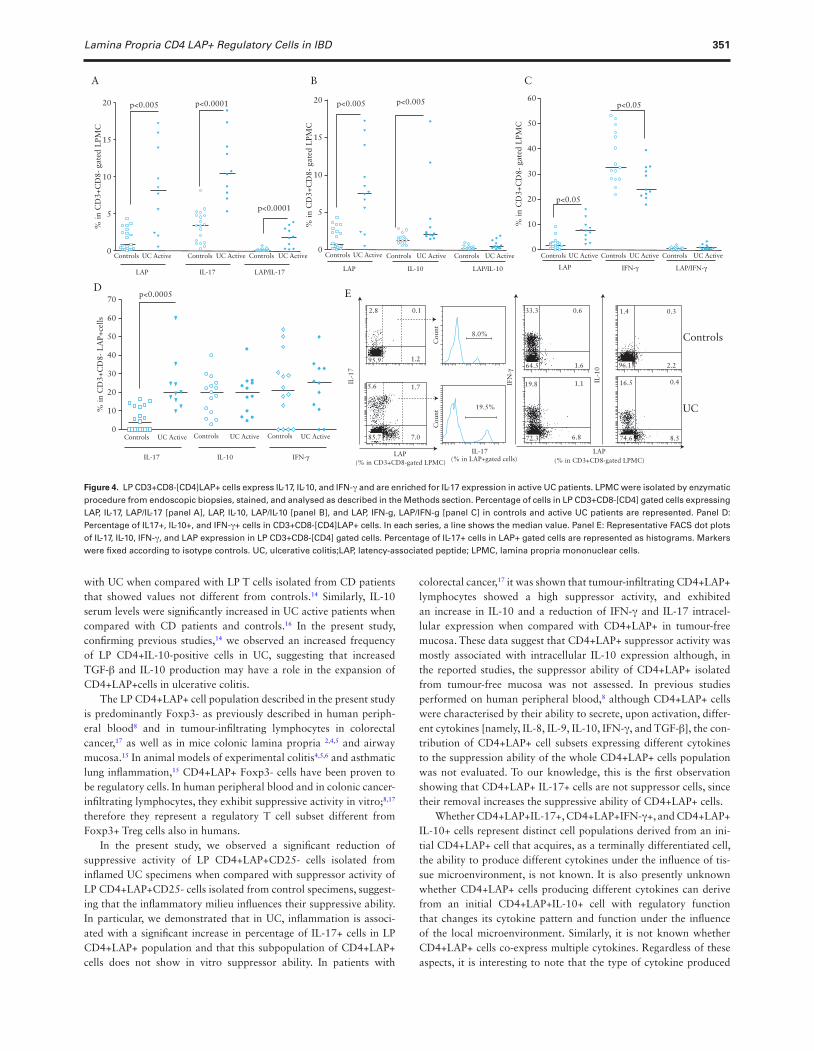
Lamina Propria CD4 LAP+ Regulatory Cells in IBD 351
with UC when compared with LP T cells isolated from CD patients that showed values not different from controls.14 Similarly, IL-10 serum levels were significantly increased in UC active patients when compared with CD patients and controls.16 In the present study, confirming previous studies,14 we observed an increased frequency of LP CD4+IL-10-positive cells in UC, suggesting that increased TGF-β and IL-10 production may have a role in the expansion of CD4+LAP+cells in ulcerative colitis.
The LP CD4+LAP+ cell population described in the present study is predominantly Foxp3- as previously described in human periph-eral blood8 and in tumour-infiltrating lymphocytes in colorectal cancer,17 as well as in mice colonic lamina propria 2,4,5 and airway mucosa.15 In animal models of experimental colitis4,5,6 and asthmatic lung inflammation,15 CD4+LAP+ Foxp3- cells have been proven to be regulatory cells. In human peripheral blood and in colonic cancer-infiltrating lymphocytes, they exhibit suppressive activity in vitro;8,17 therefore they represent a regulatory T cell subset different from Foxp3+ Treg cells also in humans.
In the present study, we observed a significant reduction of suppressive activity of LP CD4+LAP+CD25- cells isolated from inflamed UC specimens when compared with suppressor activity of LP CD4+LAP+CD25- cells isolated from control specimens, suggest-ing that the inflammatory milieu influences their suppressive ability. In particular, we demonstrated that in UC, inflammation is associ-ated with a significant increase in percentage of IL-17+ cells in LP CD4+LAP+ population and that this subpopulation of CD4+LAP+ cells does not show in vitro suppressor ability. In patients with
colorectal cancer,17 it was shown that tumour-infiltrating CD4+LAP+ lymphocytes showed a high suppressor activity, and exhibited an increase in IL-10 and a reduction of IFN-γ and IL-17 intracel-lular expression when compared with CD4+LAP+ in tumour-free mucosa. These data suggest that CD4+LAP+ suppressor activity was mostly associated with intracellular IL-10 expression although, in the reported studies, the suppressor ability of CD4+LAP+ isolated from tumour-free mucosa was not assessed. In previous studies performed on human peripheral blood,8 although CD4+LAP+ cells were characterised by their ability to secrete, upon activation, differ-ent cytokines [namely, IL-8, IL-9, IL-10, IFN-γ, and TGF-β], the con-tribution of CD4+LAP+ cell subsets expressing different cytokines to the suppression ability of the whole CD4+LAP+ cells population was not evaluated. To our knowledge, this is the first observation showing that CD4+LAP+ IL-17+ cells are not suppressor cells, since their removal increases the suppressive ability of CD4+LAP+ cells.
Whether CD4+LAP+IL-17+, CD4+LAP+IFN-γ+, and CD4+LAP+ IL-10+ cells represent distinct cell populations derived from an ini-tial CD4+LAP+ cell that acquires, as a terminally differentiated cell, the ability to produce different cytokines under the influence of tis-sue microenvironment, is not known. It is also presently unknown whether CD4+LAP+ cells producing different cytokines can derive from an initial CD4+LAP+IL-10+ cell with regulatory function that changes its cytokine pattern and function under the influence of the local microenvironment. Similarly, it is not known whether CD4+LAP+ cells co-express multiple cytokines. Regardless of these aspects, it is interesting to note that the type of cytokine produced
BA
20 p<0.005 p<0.0001
p<0.0001
15
10
% in
CD
3+C
D8-
gat
ed L
PMC
5
0Controls UC Active
LAP IL-17
LAP(% in CD3+CD8-gated LPMC) (% in CD3+CD8-gated LPMC)(% in LAP+gated cells)
LAPIL-17
IL-1
0
LAP/IL-17
Controls UC Active Controls UC Active
D Ep<0.0005
10
20
30
40
50
60
70
% in
CD
3+C
D8-
LA
P+ce
lls
0Controls UC Active
IL-17 IL-10
Controls UC Active Controls UC Active
20 p<0.005 p<0.005
15
10
% in
CD
3+C
D8-
gat
ed L
PMC
5
0Controls UC Active
LAP IL-10 LAP/IL-10
Controls UC Active Controls UC Active
C
60
50
40
30
20
p<0.05
p<0.05
10% in
CD
3+C
D8-
gat
ed L
PMC
0Controls UC Active
LAP IFN-γ
IFN
-γ
IFN-γ
LAP/IFN-γ
Controls UC Active
Controls
UC
Controls UC Active
8.0%
2.8 33.3 0.6 0.3
0.4
2.2
8.5
16.5
1.4
1.6
1.119.8
6.8
5.6 1.7
7.085.7
95.964.5 96.1
74.672.3
0.1
1.2
19.5%
Cou
nt
IL-1
7
Cou
nt
Figure 4. LP CD3+CD8-[CD4]LAP+ cells express IL-17, IL-10, and IFN-γ and are enriched for IL-17 expression in active UC patients. LPMC were isolated by enzymatic procedure from endoscopic biopsies, stained, and analysed as described in the Methods section. Percentage of cells in LP CD3+CD8-[CD4] gated cells expressing LAP, IL-17, LAP/IL-17 [panel A], LAP, IL-10, LAP/IL-10 [panel B], and LAP, IFN-g, LAP/IFN-g [panel C] in controls and active UC patients are represented. Panel D: Percentage of IL17+, IL-10+, and IFN-γ+ cells in CD3+CD8-[CD4]LAP+ cells. In each series, a line shows the median value. Panel E: Representative FACS dot plots of IL-17, IL-10, IFN-γ, and LAP expression in LP CD3+CD8-[CD4] gated cells. Percentage of IL-17+ cells in LAP+ gated cells are represented as histograms. Markers were fixed according to isotype controls. UC, ulcerative colitis;LAP, latency-associated peptide; LPMC, lamina propria mononuclear cells.

352 A. D’Ambrosio et al.
by CD4+LAP+ are all influenced by TGF-β itself, suggesting a pos-sible autocrine mechanism of cytokine production regulation.18,19,20 Further studies focused on CD4+ LAP+ cells biology are necessary to clarify these issues.
Although we did not investigate the possible co-expression of different cytokines in LAP+ cells, from our results it appears that, at least in UC, the expression of IL-17 in CD4LAP+ cells reduces the overall ability of CD4LAP+ cells to inhibit autologous cell prolifera-tion independently of concomitant co-expression of other cytokines in the CD4LAP+IL-17+ cells. The reduced suppressor activity of CD4LAP+ regulatory cells in UC reinforces the differences observed in the regulatory cell subsets between UC and CD, where a distinct population of Foxp3+ IL-17-producing CD4 T cells maintaining Treg-typical suppressor activity in vitro has been described in CD, but not in UC patients or healthy controls.21 The above reported differences between CD and UC patients raise the possibility that the increased IL-17 production observed both in CD and UC may have different pathogenic relevance in the two diseases, being more
relevant for the genesis and perpetuation of inflammation in UC as opposed to CD. This hypothesis is supported by recent studies that showed a significantly increased percentage of circulating Th17 cells in patients with active UC when compared with patients with CD22 and the ability of serum IL-17A levels to reflect clinical disease sever-ity and predict the course of disease in patients with ulcerative coli-tis.23 Therefore, in CD, the reported failure of anti-IL-17 antibody administration to ameliorate the disease24 might have been associ-ated, at least in part, with a protective role of IL-17+/Foxp3+ T cell populations. The above reported differences between CD and UC patients highlight possible different outcomes in the two diseases of therapeutic strategies aimed to specifically block Th17 polarisation and function.
FundingThis study was supported by funds from the Italian Ministry of Health [grant n° 524].
A
C
B
D
a a+b
CD
25 F
ITC
CD
25 F
ITC
IL-1
7 PE
IL-1
7 PE
CD
8 A
PCC
D8
APC
CD3 APC-Cy7
CD3 APC-Cy7 CD3 APC-Cy7
LAP PerCP-Cy5.5
LAP PerCP-Cy5.5
LAP PerCP-Cy5.5
(CD4)CD25-LAP-(CD4)
(CD4)(CD4)CD25-
(CD4)CD25-LAP+IL-17+/-
(CD4)CD25-LAP+IL-17-
a (responders)
b
c
PBMC
LPMC
CFSE
a+c
40
30
20
10
25
20
15
10
5
0
–5
–10–25
–30
a+b a+c
–40 –30 –20 –10suppression (%)
supp
ress
ion
(%)
IL-1
7+ c
ells
(%
in L
P C
D3+
CD
8-(C
D4)
LA
P+ c
ells
)
0 10 20
Controls
UC
r = –0.96p = 0.002
0
43.4%
0 102
103
104
105
102
0–1
2710
310
410
5
102
0–191 103
104
105
0 102
103
104
105
0 102
103
104
105
54.7%34.6%
102
0–1
3410
310
410
5
102
0–157 103
104
105
102
0–1
1810
310
410
5
102
0–92 103
104
105
102
0–1
0010
310
410
5
102
0–92 103
104
105
102
0–1
3710
310
410
5
102
0–213 103
104
105
102
0–1
3710
310
410
5
102
0–213 103
104
105
Figure 5. Depletion of LP CD3+CD8-[CD4]LAP+IL-17+ increases suppressor ability of LP CD3+CD8-[CD4]LAP+ cells. A: Correlation between percentage of suppression and percentage of IL-17+ cells in LP CD3+CD8-[ CD4]LAP+ cells. LPMC were isolated from surgical specimens. B: Immune-fluorescence sorting strategy. PBMC were sorted to obtain [CD4]CD25-LAP- cells [a]. LPMC were sorted to obtain [CD4]CD25-LAP+IL-17+/- cells [b] and [CD4]CD25-LAP+IL-17- cells [c]. C: Percentage of suppression of sorted PB [CD4] CD25- LAP-cell proliferation [a] by LP [CD4] CD25- LAP+ IL-17+/- cells [b] [■, □] and by LP [CD4] LAP+CD25- cell population depleted of LAP+IL-17 producing cells [c] [▲, △]. LPMC were isolated by enzymatic procedure from endoscopic biopsies of three control subjects [closed symbols] and two UC active patients [open symbols] and expanded by in vitro stimulation with anti-CD3/CD28 coated beads [see Methods]. At the end of the culture period, after IL17-producing LP T cells were identified using the IL-17 secretion Assay-Detection Kit [see Methods], cells were immune-fluorescence sorted as in panel B and each fraction was co-cultured with autologous sorted PB CD4+CD25- LAP- cells stained with CFSE and stimulated with aCD3/2/28 coated beads. Percentage of suppression of PB cells proliferation was evaluated as in Figure 3. D. Representative histograms of PB [CD4]CD25-LAP- cells proliferation cultured in the absence of LP cells [a], in the presence of LP [CD4]CD25-LAP+ IL-17+/-cells [a b], or in the presence of LP [CD4]CD25-LAP+ IL-17- cells [a + c]. Percentage of proliferating cells is indicated. UC, ulcerative colitis;.LAP, latency associated peptide; LPMC, lamina propria mononuclear cells; PBMC, peripheral blood mononuclear cells.

Lamina Propria CD4 LAP+ Regulatory Cells in IBD 353
Conflict of InterestThe authors declare no conflict of interest.
Author ContributionsStudy conception and design: MB. Acquisition of data: AD, A C, AA, AZ, AB, M, MBiffoni, AP, CM, AK, RP. Analysis and interpretation of data: AD, AC, AA, AZ, AB, MS, MB, AP, CM, AK, RP, MB. Drafting of manuscript: MB. Critical revision: AD, AC, AA, AZ, AB, MS, MB, AP, CM, AK, RP. All authors gave final approval of the version to be submitted.
This study was presented in part at the Digestive Disease Week [DDW] 2014, Chicago IL.
References 1. Gibson DJ, Ryan EJ, Doherty GA. Keeping the bowel regular: the emerg-
ing role of Treg as a therapeutic target in inflammatory bowel disease. Inflamm Bowel Dis 2013;19:2716–24.
2. Di Giacinto C, Marinaro M, Sanchez M, Strober W, Boirivant M. Pro-biotics ameliorate recurrent Th1-mediated murine colitis by inducing IL-10 and IL-10-dependent TGF-beta-bearing regulatory cells. J Immunol 2005;15:3237–46.
3. Pronio A, Montesani C, Butteroni C, et al. Probiotic administration in patients with ileal pouch-anal anastomosis for ulcerative colitis is associated with expansion of mucosal regulatory cells. Inflamm Bowel Dis 2008;14:662–8.
4. Amendola A, Butera A, Sanchez M, Strober W, Boirivant M. Nod2 defi-ciency is associated with an increased mucosal immunoregulatory response to commensal microorganisms. Mucosal Immunol 2014;7:391–404.
5. Amendola A, Butera A, Sanchez M, et al. A transient breach in the epithe-lial barrier leads to regulatory T-cell generation and resistance to experi-mental colitis. Gastroenterology 2008;135:1612–23.
6. Oida T, Zhang X, Goto M, et al. CD4+CD25- T cells that express latency-associated peptide on the surface suppress CD4+CD45RBhigh-induced coli-tis by a TGF-beta-dependent mechanism. J Immunol 2003;170:2516–22.
7. Oida T, Weiner HL. TGF-β induces surface LAP expression on murine CD4 T cells independent of Foxp3 induction. PLoS One 2010;5:e15523.
8. Gandhi R, Farez MF, Wang Y, Kozoriz D, Quintana FJ, Weiner HL. Cutting edge: human latency-associated peptide+ T cells: a novel regulatory T cell subset. J Immunol 2010;184:4620–4.
9. Feagan BG, Greenberg GR, Wild G, et al. Treatment of ulcerative colitis with a humanized antibody to the alpha4beta7 integrin. N Engl J Med 2005;352:2499–507.
10. Del Zotto B, Mumolo G, Pronio AM, Montesani C, Tersigni R, Boirivant M. TGF-beta1 production in inflammatory bowel disease: differing pro-
duction patterns in Crohn’s disease and ulcerative colitis. Clin Exp Immu-nol 2003;134:120–6.
11. Fuss IJ, Heller F, Boirivant M, et al. Nonclassical CD1d-restricted NK T cells that produce IL-13 characterize an atypical Th2 response in ulcera-tive colitis. J Clin Invest 2004;113:1490–7.
12. Fuss IJ, Joshi B, Yang Z, et al. IL-13Rα2-bearing, type II NKT cells reac-tive to sulfatide self-antigen populate the mucosa of ulcerative colitis. Gut 2014; 63:1728–36.
13. Kobayashi T, Okamoto S, Hisamatsu T, et al. IL23 differentially regu-lates the Th1/Th17 balance in ulcerative colitis and Crohn’s disease. Gut 2008;57:1682–9.
14. Melgar S, Yeung MM-W, Bas A, et al. Over-expression of interleukin 10 in mucosal T cells of patients with active ulcerative colitis. Clin Exp Immu-nol 2003;134:127–37.
15. Duan W, So T, Mehta AK, Choi H, Croft M. Inducible CD4+LAP+Foxp3- regulatory T cells suppress allergic inflammation. J Immunol 2011;187:6499–507.
16. Szkaradkiewicz A, Marciniak R, Chudzicka-Strugała I, et al. Proinflam-matory cytokines and IL-10 in inflammatory bowel disease and colorectal cancer patients. Arch Immunol Ther Exp [Warsz] 2009;57:291–4.
17. Scurr M, Ladell K, Besneux M, et al. Highly prevalent colorectal cancer-infiltrating LAP⁺ Foxp3⁻ T cells exhibit more potent immunosuppressive activity than Foxp3⁺ regulatory T cells. Mucosal Immunol 2014;7:428–39.
18. Oh SA, Li MO. TGF-β: guardian of T cell function. J Immunol 2013;191:3973–9.
19. Geginat J, Paroni M, Maglie S, et al. Plasticity of human CD4 T cell sub-sets. Front Immunol 2014;5:630.
20. Huss DJ, Winger RC, Cox GM, et al. TGF-β signaling via Smad4 drives IL-10 production in effector Th1 cells and reduces T-cell trafficking in EAE. Eur J Immunol 2011;41:2987–96.
21. Hovhannisyan Z, Treatman J, Littman DR, Mayer L. Characteriza-tion of IL-17-producing regulatory T cells in inflamed intestinal mucosa from patients with inflammatory bowel diseases. Gastroenterology 2011;140:957–65.
22. Dong Z, Du L, Xu X, et al. Aberrant expression of circulating Th17, Th1 and Tc1 cells in patients with active and inactive ulcerative colitis. Int J Mol Med 2013;31:989–97.
23. Ohman L, Dahlén R, Isaksson S, et al. Serum IL-17A in newly diag-nosed treatment-naive patients with ulcerative colitis reflects clinical disease severity and predicts the course of disease. Inflamm Bowel Dis 2013;19:2433–9.
24. Hueber W, Sands BE, Lewitzky S, et al. Secukinumab, a human anti-IL-17A monoclonal antibody, for moderate to severe Crohn’s disease: unex-pected results of a randomised, double-blind placebo-controlled trial. Gut 2012;61:1693–700.


















