
Kinetic and thermodynamic framework for P4-P6 RNA … preferred folding pathway involves initial ......
10
Kinetic and thermodynamic framework for P4-P6 RNA reveals tertiary motif modularity and modulation of the folding preferred pathway Namita Bisaria a,1 , Max Greenfeld a,b,1 , Charles Limouse c , Dmitri S. Pavlichin c , Hideo Mabuchi c , and Daniel Herschlag a,b,d,2 a Department of Biochemistry, Stanford University, Stanford, CA 94305; b Department of Chemical Engineering, Stanford University, Stanford, CA 94305; c Department of Applied Physics, Stanford University, Stanford, CA 94305; and d Department of Chemistry, Stanford University, Stanford, CA 94305 Edited by Jennifer A. Doudna, University of California, Berkeley, CA, and approved June 21, 2016 (received for review December 18, 2015) The past decade has seen a wealth of 3D structural information about complex structured RNAs and identification of functional intermediates. Nevertheless, developing a complete and predictive understanding of the folding and function of these RNAs in biology will require connection of individual rate and equilibrium constants to structural changes that occur in individual folding steps and further relating these steps to the properties and behavior of isolated, simplified systems. To accomplish these goals we used the consider- able structural knowledge of the folded, unfolded, and intermediate states of P4-P6 RNA. We enumerated structural states and possible folding transitions and determined rate and equilibrium constants for the transitions between these states using single-molecule FRET with a series of mutant P4-P6 variants. Comparisons with simplified con- structs containing an isolated tertiary contact suggest that a given tertiary interaction has a stereotyped rate for breaking that may help identify structural transitions within complex RNAs and simplify the prediction of folding kinetics and thermodynamics for structured RNAs from their parts. The preferred folding pathway involves initial formation of the proximal tertiary contact. However, this preference was only ∼10 fold and could be reversed by a single point mutation, indicating that a model akin to a protein-folding contact order model will not suffice to describe RNA folding. Instead, our results suggest a strong analogy with a modified RNA diffusion-collision model in which tertiary elements within preformed secondary structures collide, with the success of these collisions dependent on whether the tertiary elements are in their rare binding-competent conformations. RNA folding | single-molecule FRET | kinetics | folding pathways | RNA tertiary motifs S tructured RNAs play central roles in biology, in the splicing and alternative splicing of eukaryotic pre-mRNAs, in the synthesis of proteins and their transport, and in chromosome maintenance (1–4). Beyond the RNAs and RNA–protein ma- chines involved in these processes, it has been increasingly rec- ognized that Nature has taken extensive advantage of RNA at multiple levels of gene regulation, and considerable current ef- forts are focused on uncovering the pathways and molecular mechanisms that underlie the functions of small RNAs and long noncoding RNAs (2, 5– 8). The pervasive functions of RNA in bio- logy underscore the importance of understanding RNA’s funda- mental physical properties and, ultimately, of using this understanding to predict the kinetics and thermodynamics of folding and confor- mational transitions responsible for RNA function. Decades of characterization of RNA folding and structure have led to generalizable principles and provided biological in- sights. The observations that RNA encodes genetic information, forms highly stable local structures, and can catalyze reactions provided support for the possibility that early life evolved using functional RNAs (9–12). This high stability was also recognized to lead to misfolding. Indeed, highly stable misfolded tRNAs were identified in the 1960s (13–15), and many subsequent studies supported a strong propensity of RNA to misfold (16–20). These observations and consideration of the physical properties of RNA (21) led to the “RNA chaperone hypothesis” (22), which pre- dicted the existence of RNA chaperones to prevent and resolve deleterious misfolded states in vivo (20, 23–28). Due to its complexity, most studies of RNA tertiary folding have been descriptive. Studies to date have identified misfolded intermediates (29–32), revealed multiple folding pathways (33, 34) and functional states (35), and led to the general perspective that RNA folds on a rugged energetic landscape (18). Never- theless, these conclusions drawn from qualitative studies have been limited and, even at times, misleading. For example, evidence for new intermediates arising upon mutation and for changes in conditions that supported a rugged landscape for RNA folding in- stead has been shown, upon placement of results within the context of a kinetic folding framework, to represent formation of the same intermediate with altered stability (36). Additionally, an alter- native base-pairing of the P3 helix was implicated in forming a long-lived misfolded state of the Tetrahymena group I intron (30) but could be isolated and probed only once the kinetic folding framework was established. Subsequent studies showed that the alternative P3 helix biased partitioning toward the long-lived misfolded state but that the final state contains the native P3 helix with its kinetic stability arising from an altered folding to- pology that requires near-complete unfolding to rectify (37, 38). The above examples highlight the importance and power of quantitative analysis of individual reaction steps within complex Significance Many biological processes, including splicing, translation, and genome maintenance, require structured RNAs to fold into complex three-dimensional shapes. Our current understanding of these processes is based on distilling principles from de- scriptive folding studies. Moving toward predictive models will require coupling observed structural changes with kinetic and thermodynamic measurements. We have dissected P4-P6 RNA folding through distinct structural states and measured the rate and equilibrium constants for transitions between these states. Common kinetics found for RNA tertiary elements embedded in different structural contexts may help develop predictive folding models. Also, our results suggest that RNA folding may be well described by a model analogous to the diffusion-collision model for protein folding. Author contributions: N.B., M.G., C.L., and D.H. designed research; N.B., M.G., and C.L. performed research; H.M. contributed new reagents/analytic tools; N.B., M.G., C.L., and D.S.P. analyzed data; and N.B., M.G., and D.H. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. Data deposition: The single-molecule FRET data in a standardized format for reanalysis (single-molecule dataset or SMD) reported in this paper can be found at https://figshare. com/articles/Bisaria2016_PNAS_smFRETSMD_zip/3504470. 1 N.B. and M.G. contributed equally to this work. 2 To whom correspondence should be addressed. Email: [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1525082113/-/DCSupplemental. E4956–E4965 | PNAS | Published online August 4, 2016 www.pnas.org/cgi/doi/10.1073/pnas.1525082113
Transcript of Kinetic and thermodynamic framework for P4-P6 RNA … preferred folding pathway involves initial ......

Kinetic and thermodynamic framework for P4-P6 RNAreveals tertiary motif modularity and modulation ofthe folding preferred pathwayNamita Bisariaa,1, Max Greenfelda,b,1, Charles Limousec, Dmitri S. Pavlichinc, Hideo Mabuchic, and Daniel Herschlaga,b,d,2
aDepartment of Biochemistry, Stanford University, Stanford, CA 94305; bDepartment of Chemical Engineering, Stanford University, Stanford, CA 94305;cDepartment of Applied Physics, Stanford University, Stanford, CA 94305; and dDepartment of Chemistry, Stanford University, Stanford, CA 94305
Edited by Jennifer A. Doudna, University of California, Berkeley, CA, and approved June 21, 2016 (received for review December 18, 2015)
The past decade has seen a wealth of 3D structural informationabout complex structured RNAs and identification of functionalintermediates. Nevertheless, developing a complete and predictiveunderstanding of the folding and function of these RNAs in biologywill require connection of individual rate and equilibrium constants tostructural changes that occur in individual folding steps and furtherrelating these steps to the properties and behavior of isolated,simplified systems. To accomplish these goals we used the consider-able structural knowledge of the folded, unfolded, and intermediatestates of P4-P6 RNA. We enumerated structural states and possiblefolding transitions and determined rate and equilibrium constants forthe transitions between these states using single-molecule FRET witha series of mutant P4-P6 variants. Comparisons with simplified con-structs containing an isolated tertiary contact suggest that a giventertiary interaction has a stereotyped rate for breaking that may helpidentify structural transitions within complex RNAs and simplify theprediction of folding kinetics and thermodynamics for structuredRNAs from their parts. The preferred folding pathway involves initialformation of the proximal tertiary contact. However, this preferencewas only ∼10 fold and could be reversed by a single point mutation,indicating that a model akin to a protein-folding contact order modelwill not suffice to describe RNA folding. Instead, our results suggest astrong analogywith amodified RNA diffusion-collisionmodel inwhichtertiary elements within preformed secondary structures collide, withthe success of these collisions dependent on whether the tertiaryelements are in their rare binding-competent conformations.
RNA folding | single-molecule FRET | kinetics | folding pathways |RNA tertiary motifs
Structured RNAs play central roles in biology, in the splicingand alternative splicing of eukaryotic pre-mRNAs, in the
synthesis of proteins and their transport, and in chromosomemaintenance (1–4). Beyond the RNAs and RNA–protein ma-chines involved in these processes, it has been increasingly rec-ognized that Nature has taken extensive advantage of RNA atmultiple levels of gene regulation, and considerable current ef-forts are focused on uncovering the pathways and molecularmechanisms that underlie the functions of small RNAs and longnoncoding RNAs (2, 5–8). The pervasive functions of RNA in bio-logy underscore the importance of understanding RNA’s funda-mental physical properties and, ultimately, of using this understandingto predict the kinetics and thermodynamics of folding and confor-mational transitions responsible for RNA function.Decades of characterization of RNA folding and structure
have led to generalizable principles and provided biological in-sights. The observations that RNA encodes genetic information,forms highly stable local structures, and can catalyze reactionsprovided support for the possibility that early life evolved usingfunctional RNAs (9–12). This high stability was also recognizedto lead to misfolding. Indeed, highly stable misfolded tRNAswere identified in the 1960s (13–15), and many subsequent studiessupported a strong propensity of RNA to misfold (16–20). Theseobservations and consideration of the physical properties of RNA
(21) led to the “RNA chaperone hypothesis” (22), which pre-dicted the existence of RNA chaperones to prevent and resolvedeleterious misfolded states in vivo (20, 23–28).Due to its complexity, most studies of RNA tertiary folding
have been descriptive. Studies to date have identified misfoldedintermediates (29–32), revealed multiple folding pathways (33,34) and functional states (35), and led to the general perspectivethat RNA folds on a rugged energetic landscape (18). Never-theless, these conclusions drawn from qualitative studies havebeen limited and, even at times, misleading. For example, evidencefor new intermediates arising upon mutation and for changes inconditions that supported a rugged landscape for RNA folding in-stead has been shown, upon placement of results within the contextof a kinetic folding framework, to represent formation of the sameintermediate with altered stability (36). Additionally, an alter-native base-pairing of the P3 helix was implicated in forming along-lived misfolded state of the Tetrahymena group I intron (30)but could be isolated and probed only once the kinetic foldingframework was established. Subsequent studies showed that thealternative P3 helix biased partitioning toward the long-livedmisfolded state but that the final state contains the native P3helix with its kinetic stability arising from an altered folding to-pology that requires near-complete unfolding to rectify (37, 38).The above examples highlight the importance and power of
quantitative analysis of individual reaction steps within complex
Significance
Many biological processes, including splicing, translation, andgenome maintenance, require structured RNAs to fold intocomplex three-dimensional shapes. Our current understandingof these processes is based on distilling principles from de-scriptive folding studies. Moving toward predictive models willrequire coupling observed structural changes with kinetic andthermodynamic measurements. We have dissected P4-P6 RNAfolding through distinct structural states and measured the rateand equilibrium constants for transitions between these states.Common kinetics found for RNA tertiary elements embedded indifferent structural contexts may help develop predictive foldingmodels. Also, our results suggest that RNA folding may be welldescribed by a model analogous to the diffusion-collision modelfor protein folding.
Author contributions: N.B., M.G., C.L., and D.H. designed research; N.B., M.G., and C.L.performed research; H.M. contributed new reagents/analytic tools; N.B., M.G., C.L., andD.S.P. analyzed data; and N.B., M.G., and D.H. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Data deposition: The single-molecule FRET data in a standardized format for reanalysis(single-molecule dataset or SMD) reported in this paper can be found at https://figshare.com/articles/Bisaria2016_PNAS_smFRETSMD_zip/3504470.1N.B. and M.G. contributed equally to this work.2To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1525082113/-/DCSupplemental.
E4956–E4965 | PNAS | Published online August 4, 2016 www.pnas.org/cgi/doi/10.1073/pnas.1525082113

processes. Such studies have been instrumental in many areas ofbiology and biochemistry—providing kinetics and frameworks toconsolidate existing knowledge, aid the design of new experi-mental tests, anchor the interpretation of new results, and allowmore precise and predictive models to be built (39–43). Thepower of this information is fully manifest when accompanied bystructural information sufficient to allow molecular, and evenbiological, interpretation of kinetic and thermodynamic behavior(44, 45). However, the folding of RNAs, such as the group Iribozymes, RNase P RNA, and ribosomal RNAs, is extremelycomplex, with many structural elements, folding intermediates,and pathways, and there is limited structural information aboutthese intermediates, precluding them as foundational models toconnect kinetic and thermodynamic measurements with specificstructural changes during folding.
Decades of work on P4-P6 RNA (Fig. 1) have produced anX-ray structure of the folded state and rich structural in-formation that delineate a set of potential folding intermediatesand folding pathways (Fig. 1C) (47–57). Moreover, functionalwork has uncovered sets of mutations that allow ablation andisolation of these folding intermediates with high confidence (49,51, 53, 56, 58–60). The control afforded by this well-behaved,established system provides an opportunity to deepen under-standing and develop and test generalizable principles beyondthis model system, provided that a detailed framework is in placeto provide context for interpretation. Here, we use single-moleculeFRET (smFRET) with a series of P4-P6 mutants to isolate andmeasure specific folding transitions, extending beyond measure-ments of overall folding rates (52–55, 61–65) and constructing akinetic and thermodynamic framework for P4-P6 RNA. Thisframework defines a preferred folding pathway, allows this path-way and the partitioning of states to be interpreted in structuralterms, helps to uncover the origin of the effects of mutations onfolding rates and pathways, and suggests a generalized frameworkfor considering RNA tertiary folding.A further major goal in RNA folding, beyond a description of
the kinetics and thermodynamics of individual structural transi-tions, is to obtain quantitative and predictive algorithms forRNA tertiary structure, akin to the predictive rules for RNAsecondary structure formation. Although this problem is complex,several observations suggest that such a goal may be achievabledue to a modular architecture and folding behavior of RNA (66,67). This perspective is based on the observations that struc-tures of tertiary contacts and junctions found in different RNAstructures can be overlaid with little deviation (68–70) and thatthese substructures, or motifs, can be “cut and pasted” from onestructural context to another with function retained (67–69, 71–75).From these observations, it has been proposed that it may be pos-sible to predict the thermodynamics and kinetics of RNA tertiarystructure formation from the energetic contributions of these motifsand other components of RNA structure (67). However, despitethe structural similarity and transferability of motifs, there is asyet no quantitative assessment of whether the energetic propertiesof tertiary structure elements are the same in different structuralcontexts—a basic requirement for the viability of an energetic re-construction of RNA tertiary structure from its components.The P4-P6 kinetic and thermodynamic folding framework we
derive provides an opportunity to assess whether components ofRNA tertiary structure are energetically modular. Specifically,the P4-P6 framework isolates the formation and breaking of eachof its two tertiary contacts, the tetraloop/tetraloop receptor andthe metal core/metal-core receptor, allowing us to compare therate constants for breaking these tertiary contacts in P4-P6 withthose in simplified RNA constructs with each tertiary motifalone. We find kinetic properties of these motifs that are in-dependent of structural context and thus may be transferrable,thereby providing an early step toward a predictive, energeticmodel for RNA tertiary structure.
Results and DiscussionWe first developed the P4-P6 folding framework from structuralstates previously identified and measured the rate and equilibriumconstants for transitions between these states with smFRET. Wethen used this framework to isolate the formation and dissolutionof particular tertiary contacts to test whether the kinetics prop-erties of this structural transition are transferable to other RNAcontexts. Finally, we probed properties of the framework, in par-ticular the flux through folding pathways in response to mutation,that suggest features of RNA folding that are analogous to simpleprotein-folding models and may be general.
Building and Determining a Kinetic and Thermodynamic Frameworkfor P4-P6 RNA.Building the structural states for the P4-P6 framework.We first built onthe large body of work on P4-P6 RNA (e.g., refs. 47–56, 59, 60,65 and 76–82) to construct a framework of structural states, and
Fig. 1. P4-P6 RNA and proposed framework. (A) P4-P6 crystal structure (PDB IDcode 1GID). Tertiary contacts are colored as follows: tetraloop (TL, light blue),tetraloop receptor (TLR, dark blue), metal core (MC, light green), metal-corereceptor (MCR, dark green). Crystallographically and functionally defined Mg2+
ions bound by theMC are shown as gold spheres (46, 50, 82). (B) P4-P6 secondarystructure and variants used in this study to isolate specific folding intermediates(see also Fig. 2A). Tertiary contacts are colored as inA. The positions of the donorand acceptor dyes are shown. (C) Proposed thermodynamic and kinetic frame-work based on previously identified structural intermediates. P4-P6 is shown in aschematic depiction, color-coded as in A and B, and tertiary contacts are shownthroughout as a green swatch and a blue swatch for the MC/MCR and TL/TLR,respectively. Intermediates include the following: Alt P5cU, unfolded state with analternative P5c secondary structure; Alt P5cITL/TLR, intermediate with the TL/TLRinteraction formed and the alternative P5c secondary structure; U, unfoldedstate with the native P5c secondary structure (states not notated “Alt P5c” havenative P5c secondary structure); ITL/TLR, intermediate with only the TL/TLR in-teraction formed; MU, unfolded state with the MC formed through binding oftwo Mg2+ ions (depicted as two bound gold spheres); MITL/TLR, intermediate inwhich the TL/TLR interaction and the MC are formed; IMC/MCR, intermediate withthe MC/MCR interaction formed; and F, the native folded state in which bothtertiary contacts are formed (49, 56, 60, 77, 82, 118).
Bisaria et al. PNAS | Published online August 4, 2016 | E4957
BIOCH
EMISTR
YPN
ASPL
US
we then probed transitions between these states to establish a ki-netic and thermodynamic folding framework.The P456 helical stack of P4-P6 (Fig. 1B, Right) connects to
the P5abc helices (Fig. 1B, Left) via the two-way J5/5a junction,which enables a 180-degree turn that allows the P5abc helix tobe roughly parallel to the P456 helices (Fig. 1A). This parallelconfiguration is enforced by two tertiary contacts of P4-P6, themetal core/metal-core receptor (MC/MCR) and the tetraloop/tetraloop receptor (TL/TLR). TL/TLR tertiary motifs are foundin many structured RNAs (70), and, whereas the MC/MCR maybe unique to this RNA (50), specific divalent binding sites arecommon in tertiary structures (83–87). In addition, many structuredRNAs have junctions that, like J5/5a, exhibit distinct bendingproperties (51, 68, 88, 89).The folding of P4-P6 has been followed in multiple studies,
with folding rate constants of 2–30 s−1 observed across a rangeof conditions (52–55, 61–65). However, interpretation of theseoverall rate constants has been stymied by a lack of knowledge ofwhich individual folding steps were followed and rate limiting.Fig. 1C diagrams potential intermediates in the folding of P4-P6 suggested by prior structural work (47–57). States differ inwhether an alternative secondary structure is present (Fig. 1C,Alt P5c, red), whether Mg2+ ions are bound to the metal core(Fig. 1C, gold circles), and whether each tertiary contact isformed (Fig. 1C, MC/MCR, green and TL/TLR, blue). NMR,small angle X-ray scattering, and chemical probing data withWT and mutant P4-P6 RNAs provide evidence for each ofthese structural states and conditions or mutations that alloweach species to predominate (49, 51, 53, 56, 58–60).Determining the P4-P6 kinetic and thermodynamic framework. We stud-ied the folding of the P4-P6 constructs depicted in Fig. 1Bby smFRET.smFRET allows determination of forward and reverse reaction
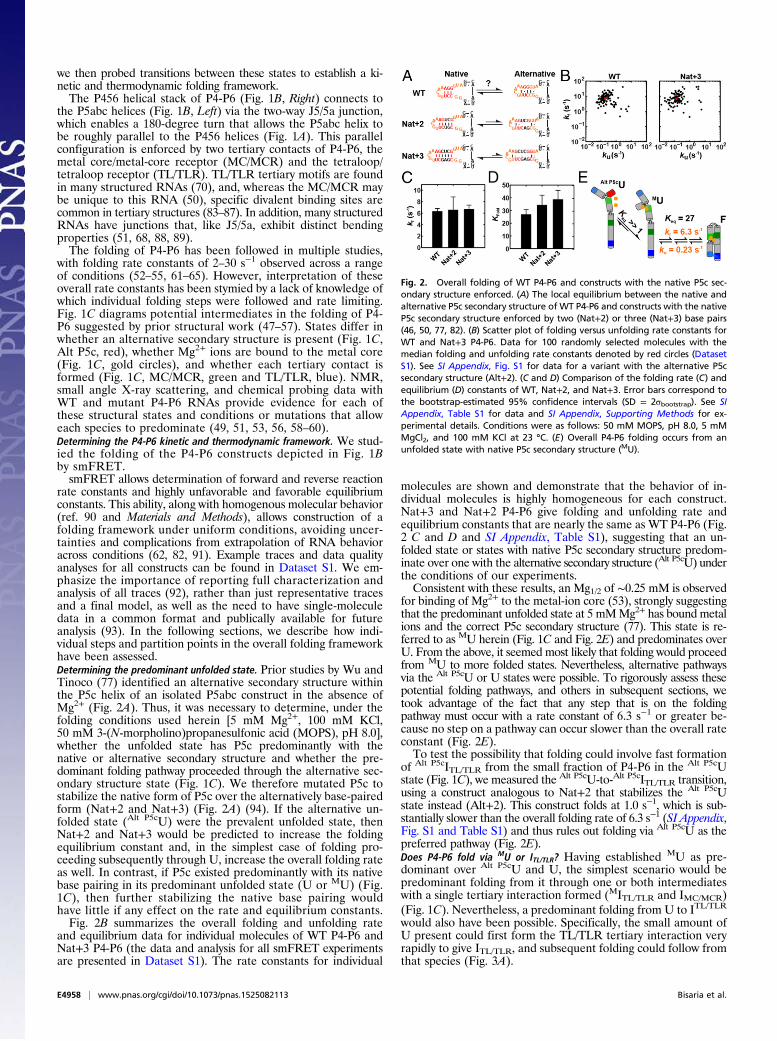
rate constants and highly unfavorable and favorable equilibriumconstants. This ability, along with homogenous molecular behavior(ref. 90 and Materials and Methods), allows construction of afolding framework under uniform conditions, avoiding uncer-tainties and complications from extrapolation of RNA behavioracross conditions (62, 82, 91). Example traces and data qualityanalyses for all constructs can be found in Dataset S1. We em-phasize the importance of reporting full characterization andanalysis of all traces (92), rather than just representative tracesand a final model, as well as the need to have single-moleculedata in a common format and publically available for futureanalysis (93). In the following sections, we describe how indi-vidual steps and partition points in the overall folding frameworkhave been assessed.Determining the predominant unfolded state. Prior studies by Wu andTinoco (77) identified an alternative secondary structure withinthe P5c helix of an isolated P5abc construct in the absence ofMg2+ (Fig. 2A). Thus, it was necessary to determine, under thefolding conditions used herein [5 mM Mg2+, 100 mM KCl,50 mM 3-(N-morpholino)propanesulfonic acid (MOPS), pH 8.0],whether the unfolded state has P5c predominantly with thenative or alternative secondary structure and whether the pre-dominant folding pathway proceeded through the alternative sec-ondary structure state (Fig. 1C). We therefore mutated P5c tostabilize the native form of P5c over the alternatively base-pairedform (Nat+2 and Nat+3) (Fig. 2A) (94). If the alternative un-folded state (Alt P5cU) were the prevalent unfolded state, thenNat+2 and Nat+3 would be predicted to increase the foldingequilibrium constant and, in the simplest case of folding pro-ceeding subsequently through U, increase the overall folding rateas well. In contrast, if P5c existed predominantly with its nativebase pairing in its predominant unfolded state (U or MU) (Fig.1C), then further stabilizing the native base pairing wouldhave little if any effect on the rate and equilibrium constants.Fig. 2B summarizes the overall folding and unfolding rate
and equilibrium data for individual molecules of WT P4-P6 andNat+3 P4-P6 (the data and analysis for all smFRET experimentsare presented in Dataset S1). The rate constants for individual
molecules are shown and demonstrate that the behavior of in-dividual molecules is highly homogeneous for each construct.Nat+3 and Nat+2 P4-P6 give folding and unfolding rate andequilibrium constants that are nearly the same as WT P4-P6 (Fig.2 C and D and SI Appendix, Table S1), suggesting that an un-folded state or states with native P5c secondary structure predom-inate over one with the alternative secondary structure (Alt P5cU) underthe conditions of our experiments.Consistent with these results, an Mg1/2 of ∼0.25 mM is observed
for binding of Mg2+ to the metal-ion core (53), strongly suggestingthat the predominant unfolded state at 5 mMMg2+ has bound metalions and the correct P5c secondary structure (77). This state is re-ferred to as MU herein (Fig. 1C and Fig. 2E) and predominates overU. From the above, it seemed most likely that folding would proceedfrom MU to more folded states. Nevertheless, alternative pathwaysvia the Alt P5cU or U states were possible. To rigorously assess thesepotential folding pathways, and others in subsequent sections, wetook advantage of the fact that any step that is on the foldingpathway must occur with a rate constant of 6.3 s−1 or greater be-cause no step on a pathway can occur slower than the overall rateconstant (Fig. 2E).To test the possibility that folding could involve fast formation
of Alt P5cITL/TLR from the small fraction of P4-P6 in the Alt P5cUstate (Fig. 1C), we measured the Alt P5cU-to-Alt P5cITL/TLR transition,using a construct analogous to Nat+2 that stabilizes the Alt P5cUstate instead (Alt+2). This construct folds at 1.0 s−1, which is sub-stantially slower than the overall folding rate of 6.3 s−1 (SI Appendix,Fig. S1 and Table S1) and thus rules out folding via Alt P5cU as thepreferred pathway (Fig. 2E).Does P4-P6 fold via MU or ITL/TLR? Having established MU as pre-dominant over Alt P5cU and U, the simplest scenario would bepredominant folding from it through one or both intermediateswith a single tertiary interaction formed (MITL/TLR and IMC/MCR)(Fig. 1C). Nevertheless, a predominant folding from U to ITL/TLR
would also have been possible. Specifically, the small amount ofU present could first form the TL/TLR tertiary interaction veryrapidly to give ITL/TLR, and subsequent folding could follow fromthat species (Fig. 3A).
Fig. 2. Overall folding of WT P4-P6 and constructs with the native P5c sec-ondary structure enforced. (A) The local equilibrium between the native andalternative P5c secondary structure of WT P4-P6 and constructs with the nativeP5c secondary structure enforced by two (Nat+2) or three (Nat+3) base pairs(46, 50, 77, 82). (B) Scatter plot of folding versus unfolding rate constants forWT and Nat+3 P4-P6. Data for 100 randomly selected molecules with themedian folding and unfolding rate constants denoted by red circles (DatasetS1). See SI Appendix, Fig. S1 for data for a variant with the alternative P5csecondary structure (Alt+2). (C and D) Comparison of the folding rate (C) andequilibrium (D) constants of WT, Nat+2, and Nat+3. Error bars correspond tothe bootstrap-estimated 95% confidence intervals (SD = 2σbootstrap). See SIAppendix, Table S1 for data and SI Appendix, Supporting Methods for ex-perimental details. Conditions were as follows: 50 mM MOPS, pH 8.0, 5 mMMgCl2, and 100 mM KCl at 23 °C. (E) Overall P4-P6 folding occurs from anunfolded state with native P5c secondary structure (MU).
E4958 | www.pnas.org/cgi/doi/10.1073/pnas.1525082113 Bisaria et al.

We isolated the folding of U to ITL/TLR by mutating residues inthe so-called A-rich bulge, an internal loop that binds Mg2+ ions;and these mutations were previously shown to prevent Mg2+
binding (Fig. 1B, ArichU) (56). Fig. 3B shows the folding andunfolding of individual ArichU RNA molecules, and Fig. 3Ccompares the folding rate constants for this mutant, an addi-tional mutant that prevents Mg2+ binding (A186U) (58), and WTP4-P6. These mutants can fold only to ITL/TLR whereas WTP4-P6 folds to its final state (F) with both contacts under theseconditions (56). The MC mutants fold ∼10-fold slower than WT(Fig. 3C and SI Appendix, Table S1). Because an overall processcannot occur faster than its slowest step, a pathway involving arare state such as U would need to be correspondingly fasterthan the overall folding rate. Thus, these data strongly suggestthat a pathway via ITL/TLR is not significantly traversed in WTP4-P6 folding—i.e., this pathway is not kinetically competent.The observation that folding via ITL/LTR is not kinetcally
competent requires that folding of U to MU be at least as fast asthe overall folding rate constant of 6.3 s−1 because this step is onthe pathway that must be traversed (Fig. 3D). Although oursmFRET assay cannot detect the U-to-MU transition, because thereis no significant FRET difference between these states, prior
time-resolved hydroxyl radical footprinting results (53) suggest thatMg2+ binding is fast relative to overall P4-P6 folding, with a rateconstant of >50 s−1 (Fig. 3D and SI Appendix, SI Text), furthersupporting the preferred folding pathway from U to MU.Does folding proceed from MU via MITL/TLR or IMC/MCR? Once Mg2+ ionsare bound to the metal core (MC) to give MU, two possible in-termediates can form next, MITL/TLR or IMC/MCR (Fig. 1C and Fig.4A). We distinguished between these potential folding pathwaysfrom MU using mutations to isolate the transitions to MITL/TLR andIMC/MCR (Fig. 4A). To isolate the MU-to-IMC/MCR transition, wemutated the tetraloop (TL) and tetraloop receptor (TLR) (Fig. 1B).The folding of these mutants is as fast as that for overall folding(Fig. 4B, dashed line), and the same results were obtained formultiple constructs with mutations in the TL or TLR (Fig. 4D andSI Appendix, Table S1).These results meet a necessary criterion for folding proceeding
through the IMC/MCR intermediate but alone are not sufficientto establish this pathway over pathways through the MITL/TLRintermediate (Fig. 4A): i.e., a downstream step subsequent toformation of the MITL/TLR intermediate could be slow and therebyrender this path kinetically incompetent. Thus, we used MCRmutations previously shown to affect the formation of theMC/MCR (Fig. 1B) (79) to evaluate the potential for foldingthrough the MITL/TLR intermediate (Fig. 4A). Each of the threeMCR mutations led to different levels of destabilization (Fig.4C and SI Appendix, Table S1), suggesting that the two leastdestabilizing mutants did not fully ablate the MC/MCR. Be-cause the MC/MCR motif involves A-minor interactions be-tween adenosine residues and 2′-hydroxyl groups of residues inWatson–Crick base pairs, it may be particularly refractory to fullablation (50, 79, 95, 96). Nevertheless, the most destabilizingmutant, G212U, has rate constants that are the same as variantswith mutations in the MC that remove Mg2+ binding and ablatethe MC/MCR (56)(SI Appendix, Table S1) (G212U vs. ArichUand A186U), suggesting that the G212U variant also ablatesthe MC/MCR interaction. The G212U mutant and other MCRmutants gave folding rate constants slower than overall folding(Fig. 4D, dashed line vs. MCR mutants), providing evidence thatthe pathway with TL/TLR formation before MC/MCR forma-tion is not kinetically competent (Fig. 4E).The MC and MCR tertiary contact partners that form first in
folding are closer in secondary structure than the TL and TLR andare also closer to the major bend in the molecule mediated by theJ5/5a junction (Fig. 1A) (49). This observation is reminiscent ofthe correlation over four orders of magnitude of folding rates withthe closeness of tertiary contact partners in primary sequence for asubset of proteins (97). This concept has also been explored in,and suggested for, RNA folding, but only overall reaction rateswere followed and effects were modest in scale (98, 99). Herein,we observed a pathway preference of only ∼10-fold (Figs. 4E and5), despite the close positioning of the MC and MCR tertiaryelements (Fig. 1), and, as shown below, this preference can bereversed by a single point mutation. These results underscore theneed for more complex descriptions for the RNA folding process,and our results further suggest a general framework that may beappropriate (Analysis of Mutational Effects on Folding Pathways).The overall kinetic and thermodynamic framework for P4-P6 folding. Fig.5 presents the rate and equilibrium constants for individual re-action steps in the P4-P6 folding framework, combining the re-sults described above with additional results and inferences fromthe following section (Modularity of Tertiary Contact UnfoldingKinetics). Our findings can be summarized as follows: The MUstate predominates over the Alt P5cU and U states, and foldingdoes not proceed through these less populated unfolded statesbecause folding to Alt P5cITL/TLR and ITL/TLR from Alt P5cU and U,respectively, is slower than overall folding. MU proceeds pref-erentially to IMC/MCR, with a rate constant similar to that foroverall folding, whereas folding to MITL/TLR from MU is not ki-netically competent. Thus, the final step in the preferred foldingpathway is formation of F from IMC/MCR.
Fig. 3. Does the folding pathway proceed via formation of the tetraloop/tetraloop receptor (TL/TLR) or the metal core (MC)? (A) Folding from U couldoccur either through ITL/TLR or
MU. (B) Scatter plot of the folding versus unfoldingrate constants of the ArichU mutant (Fig. 1C) that isolates the U-to-ITL/TLR tran-sition. (C) Comparison of the folding rate constants of WT, ArichU, and A186UP4-P6 in Mg2+ and Ba2+. Prior work has shown that Ba2+ does not bind to themetal core so the same folding behavior is predicted for WT P4-P6 and theArichU and A186U mutants in Ba2+ (81). This expectation was met, suggestingthat there are no unintended effects from the above mutations. Errors as inFig. 2. Conditions were as follows: 50 mM MOPS, pH 8.0, 5 mM MgCl2 or BaCl2,and 100mMKCl at 23 °C. (D) P4-P6 folds predominantly via formation of the MUintermediate (green arrows). The equilibrium and kinetic values shown for theU-to-ITL/TLR pathway are average values from A186U and ArichU variants (SIAppendix, Table S1). Estimation of the limit for the U-to-MU transition of >50 s−1
was determined from previous literature data (SI Appendix, SI Text).
Bisaria et al. PNAS | Published online August 4, 2016 | E4959
BIOCH
EMISTR
YPN
ASPL
US

We could not measure the conversion of IMC/MCR to F directlybecause IMC/MCR does not accumulate sufficiently under conditionsin which there is also significant formation of F. Nevertheless, wecould calculate the equilibrium constant for the folding of IMC/MCRto F based on the values of the thermodynamic cycle in Fig. 5 in-volving the overall folding equilibrium and the folding equilibriumbetween MU and IMC/MCR (Keq = [F]/[IMC/MCR] = 190) (Fig. 5) (andsee Eq. 2 in SI Appendix, Supporting Methods). In addition, com-monalities in conformational steps described in the next sectionallow us to estimate rate constants for this step (Fig. 5, rate con-stants in parentheses and square brackets) and complete the P4-P6folding framework.
Modularity of Tertiary Contact Unfolding Kinetics. As noted in theIntroduction, structured RNAs seem to be modular, made upof motifs that are seen across multiple RNAs, and are often struc-turally superimposable (68–70). A straightforward, but hitherto un-tested, extension of this perspective would be that these structuralmotifs are also energetically modular.What is meant by “energetic modularity” and what would its
signature be? One of course would not expect all structures thatcontain a given motif to have the same overall folding equilib-rium. However, the rate constant for breaking of a particulartertiary motif could be independent of its surroundings: i.e., thesame across different structured RNAs. Although more com-plex scenarios are possible, a common unfolding rate constantacross different RNAs would be expected if a motif has thesame structure in different RNAs and if the transition state forits dissolution entails disruption of interactions within the motif
rather than distal rearrangements that induce disruption and doso differentially in different RNAs. The experiments describedbelow use our folding framework to test this kinetic prediction ofenergetic modularity.We determined the rate constants for formation and dissolu-
tion of the two tertiary contacts, the TL/TLR and the MC/MCR,in P4-P6 and in engineered constructs that isolate the folding ofeach tertiary contact (100–102). These engineered single-moleculeFRET constructs, developed by Pardi, Nesbitt, and coworkers(100–102), contain a single tertiary contact connected by a simplifiedjunction (Fig. 6A and SI Appendix, Fig. S3) (TL/TLRiso and MC/MCRiso) and allow us to obtain rate constants for unfolding ofeach contact alone.We measured the unfolding and folding rate constants for the
simplified TL/TLRiso construct previously studied (100) as wellas several related TL/TLRiso constructs (SI Appendix, Fig. S2),and we compared these values with those for P4-P6 constructsthat form only the TL/TLR tertiary contact (ArichU, Fig. 6B). Theunfolding rate constants measured for three TL/TLRiso constructsand a mutant P4-P6 construct were nearly the same over an Mg2+
concentration range of ∼10-fold. Given the very shallow Mg2+
dependences, this correspondence likely holds over a considerablywider range (Fig. 6B, Left, open vs. closed symbols). In contrast,the folding rate constants varied considerably across this range ofMg2+ (Fig. 6B, Right), presumably reflecting the different proba-bilities of aligning the TL and TLR for formation (Implications).We extended the comparisons of TL/TLR unfolding rates to
include additional P4-P6 variants (Fig. 6C). For four TL/TLRisoconstructs (SI Appendix, Fig. S2) and three P4-P6 variants lacking
Fig. 4. Does folding proceed from MU via formation of the tetraloop/tetraloop receptor (TL/TLR) or the metal core/metal-core receptor (MC/MCR)?(A) From MU, folding can proceed through MITL/TLR or IMC/MCR. (B) Scatter plotof the folding versus unfolding rate constants for the TL-AllU (blue), L5B(green), and J6a/6b BP (orange) mutants that destabilizes the TL/TLR; and(C) MCR variants C109U (blue), G212C (green), and G212U (orange) that de-stabilize the formation of the MC/MCR contact (SI Appendix, Fig. S5).(D) Comparison of the folding rate constants of WT P4-P6 with those of the TL,TLR, and MCR mutants. Triple mut, the A225U/A226U/C223U variant. Thedotted gray lines in C and D indicate the rate constant for WT P4-P6 folding.The values are from SI Appendix, Table S1, and the error estimates andconditions are as in Fig. 3. (E ) Preferential folding via IMC/MCR. The rateconstants for the MU-to-MITL/TLR transition are for the G212U mutant (SIAppendix, Table S1) because this mutant has rate constants and behaviorsimilar to MC mutations that ablate the MC/MCR interaction (56). The rateconstants for the MU-to-IMC/MCR transition are averages of the values forthe TL and TLR variants in D.
Fig. 5. Thermodynamic and kinetic framework of P4-P6 folding (5 mMMgCl2, 100 mM KCl, 23 °C). The preferred folding pathway is depicted withthe green arrows. Values for each step are taken from results in Fig. 3D, Fig.4E, and SI Appendix, Fig. S1. The value in the square bracket for F→IMC/MCR isthe average of the unfolding rate constants determined for constructs thatisolate the TL/TLR (SI Appendix, Table S2). The value in the square bracket forF → MITL/TLR is the average of unfolding rate constants for constructs thatisolate the MC/MCR (P4-P6, MU⇌IMC/MCR) (SI Appendix, Table S1); (MC/MCRiso, 5 mM Mg2+) (SI Appendix, Table S3). The value for Alt P5cITL/TLR→
Alt P5c
U was corrected by a factor of two to account for the uniform twofold slowerunfolding in 5 mM Ba2+ relative to 5 mM Mg2+ (Fig. 6C and SI Appendix, TableS1) because the experiment was carried out with Ba2+ to prevent Mg2+ occu-pancy of the metal core (MC) (81). Values in parentheses were calculated fromcompletion of the thermodynamic cycle (SI Appendix, Supporting Methods).
E4960 | www.pnas.org/cgi/doi/10.1073/pnas.1525082113 Bisaria et al.
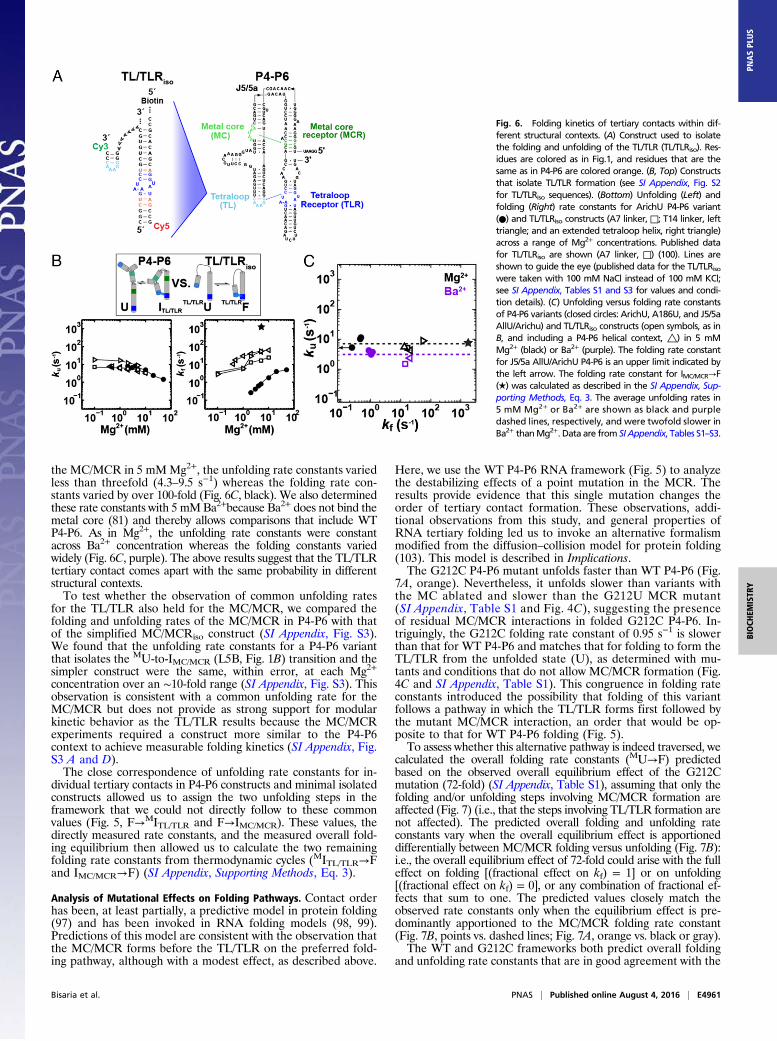
the MC/MCR in 5 mMMg2+, the unfolding rate constants variedless than threefold (4.3–9.5 s−1) whereas the folding rate con-stants varied by over 100-fold (Fig. 6C, black). We also determinedthese rate constants with 5 mMBa2+because Ba2+ does not bind themetal core (81) and thereby allows comparisons that include WTP4-P6. As in Mg2+, the unfolding rate constants were constantacross Ba2+ concentration whereas the folding constants variedwidely (Fig. 6C, purple). The above results suggest that the TL/TLRtertiary contact comes apart with the same probability in differentstructural contexts.To test whether the observation of common unfolding rates
for the TL/TLR also held for the MC/MCR, we compared thefolding and unfolding rates of the MC/MCR in P4-P6 with thatof the simplified MC/MCRiso construct (SI Appendix, Fig. S3).We found that the unfolding rate constants for a P4-P6 variantthat isolates the MU-to-IMC/MCR (L5B, Fig. 1B) transition and thesimpler construct were the same, within error, at each Mg2+concentration over an ∼10-fold range (SI Appendix, Fig. S3). Thisobservation is consistent with a common unfolding rate for theMC/MCR but does not provide as strong support for modularkinetic behavior as the TL/TLR results because the MC/MCRexperiments required a construct more similar to the P4-P6context to achieve measurable folding kinetics (SI Appendix, Fig.S3 A and D).The close correspondence of unfolding rate constants for in-
dividual tertiary contacts in P4-P6 constructs and minimal isolatedconstructs allowed us to assign the two unfolding steps in theframework that we could not directly follow to these commonvalues (Fig. 5, F→MITL/TLR and F→IMC/MCR). These values, thedirectly measured rate constants, and the measured overall fold-ing equilibrium then allowed us to calculate the two remainingfolding rate constants from thermodynamic cycles (MITL/TLR→Fand IMC/MCR→F) (SI Appendix, Supporting Methods, Eq. 3).
Analysis of Mutational Effects on Folding Pathways. Contact orderhas been, at least partially, a predictive model in protein folding(97) and has been invoked in RNA folding models (98, 99).Predictions of this model are consistent with the observation thatthe MC/MCR forms before the TL/TLR on the preferred fold-ing pathway, although with a modest effect, as described above.
Here, we use the WT P4-P6 RNA framework (Fig. 5) to analyzethe destabilizing effects of a point mutation in the MCR. Theresults provide evidence that this single mutation changes theorder of tertiary contact formation. These observations, addi-tional observations from this study, and general properties ofRNA tertiary folding led us to invoke an alternative formalismmodified from the diffusion–collision model for protein folding(103). This model is described in Implications.The G212C P4-P6 mutant unfolds faster than WT P4-P6 (Fig.
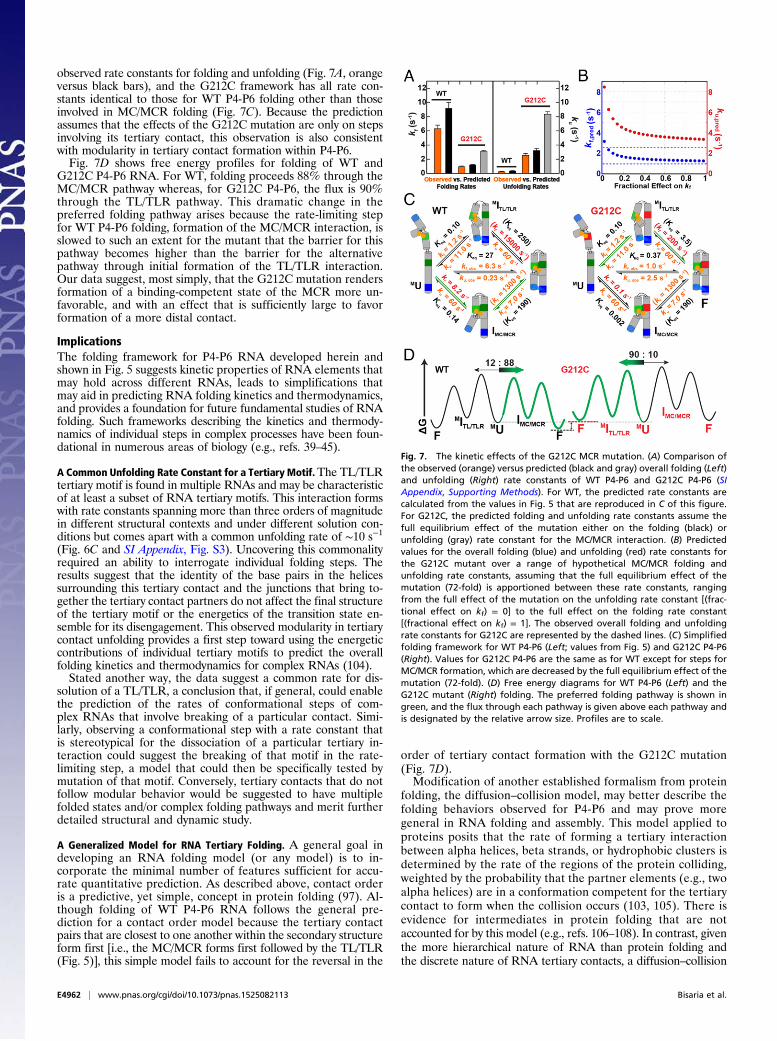
7A, orange). Nevertheless, it unfolds slower than variants withthe MC ablated and slower than the G212U MCR mutant(SI Appendix, Table S1 and Fig. 4C), suggesting the presenceof residual MC/MCR interactions in folded G212C P4-P6. In-triguingly, the G212C folding rate constant of 0.95 s−1 is slowerthan that for WT P4-P6 and matches that for folding to form theTL/TLR from the unfolded state (U), as determined with mu-tants and conditions that do not allow MC/MCR formation (Fig.4C and SI Appendix, Table S1). This congruence in folding rateconstants introduced the possibility that folding of this variantfollows a pathway in which the TL/TLR forms first followed bythe mutant MC/MCR interaction, an order that would be op-posite to that for WT P4-P6 folding (Fig. 5).To assess whether this alternative pathway is indeed traversed, we
calculated the overall folding rate constants (MU→F) predictedbased on the observed overall equilibrium effect of the G212Cmutation (72-fold) (SI Appendix, Table S1), assuming that only thefolding and/or unfolding steps involving MC/MCR formation areaffected (Fig. 7) (i.e., that the steps involving TL/TLR formation arenot affected). The predicted overall folding and unfolding rateconstants vary when the overall equilibrium effect is apportioneddifferentially between MC/MCR folding versus unfolding (Fig. 7B):i.e., the overall equilibrium effect of 72-fold could arise with the fulleffect on folding [(fractional effect on kf) = 1] or on unfolding[(fractional effect on kf) = 0], or any combination of fractional ef-fects that sum to one. The predicted values closely match theobserved rate constants only when the equilibrium effect is pre-dominantly apportioned to the MC/MCR folding rate constant(Fig. 7B, points vs. dashed lines; Fig. 7A, orange vs. black or gray).The WT and G212C frameworks both predict overall folding
and unfolding rate constants that are in good agreement with the
Fig. 6. Folding kinetics of tertiary contacts within dif-ferent structural contexts. (A) Construct used to isolatethe folding and unfolding of the TL/TLR (TL/TLRiso). Res-idues are colored as in Fig.1, and residues that are thesame as in P4-P6 are colored orange. (B, Top) Constructsthat isolate TL/TLR formation (see SI Appendix, Fig. S2for TL/TLRiso sequences). (Bottom) Unfolding (Left) andfolding (Right) rate constants for ArichU P4-P6 variant(●) and TL/TLRiso constructs (A7 linker, □; T14 linker, lefttriangle; and an extended tetraloop helix, right triangle)across a range of Mg2+ concentrations. Published datafor TL/TLRiso are shown (A7 linker, □) (100). Lines areshown to guide the eye (published data for the TL/TLRisowere taken with 100 mM NaCl instead of 100 mM KCl;see SI Appendix, Tables S1 and S3 for values and condi-tion details). (C) Unfolding versus folding rate constantsof P4-P6 variants (closed circles: ArichU, A186U, and J5/5aAllU/Arichu) and TL/TLRiso constructs (open symbols, as inB, and including a P4-P6 helical context, △) in 5 mMMg2+ (black) or Ba2+ (purple). The folding rate constantfor J5/5a AllU/ArichU P4-P6 is an upper limit indicated bythe left arrow. The folding rate constant for IMC/MCR→F(★) was calculated as described in the SI Appendix, Sup-porting Methods, Eq. 3. The average unfolding rates in5 mM Mg2+ or Ba2+ are shown as black and purpledashed lines, respectively, and were twofold slower inBa2+ thanMg2+. Data are from SI Appendix, Tables S1–S3.
Bisaria et al. PNAS | Published online August 4, 2016 | E4961
BIOCH
EMISTR
YPN
ASPL
US
observed rate constants for folding and unfolding (Fig. 7A, orangeversus black bars), and the G212C framework has all rate con-stants identical to those for WT P4-P6 folding other than thoseinvolved in MC/MCR folding (Fig. 7C). Because the predictionassumes that the effects of the G212C mutation are only on stepsinvolving its tertiary contact, this observation is also consistentwith modularity in tertiary contact formation within P4-P6.Fig. 7D shows free energy profiles for folding of WT and
G212C P4-P6 RNA. For WT, folding proceeds 88% through theMC/MCR pathway whereas, for G212C P4-P6, the flux is 90%through the TL/TLR pathway. This dramatic change in thepreferred folding pathway arises because the rate-limiting stepfor WT P4-P6 folding, formation of the MC/MCR interaction, isslowed to such an extent for the mutant that the barrier for thispathway becomes higher than the barrier for the alternativepathway through initial formation of the TL/TLR interaction.Our data suggest, most simply, that the G212C mutation rendersformation of a binding-competent state of the MCR more un-favorable, and with an effect that is sufficiently large to favorformation of a more distal contact.
ImplicationsThe folding framework for P4-P6 RNA developed herein andshown in Fig. 5 suggests kinetic properties of RNA elements thatmay hold across different RNAs, leads to simplifications thatmay aid in predicting RNA folding kinetics and thermodynamics,and provides a foundation for future fundamental studies of RNAfolding. Such frameworks describing the kinetics and thermody-namics of individual steps in complex processes have been foun-dational in numerous areas of biology (e.g., refs. 39–45).
A Common Unfolding Rate Constant for a Tertiary Motif.The TL/TLRtertiary motif is found in multiple RNAs and may be characteristicof at least a subset of RNA tertiary motifs. This interaction formswith rate constants spanning more than three orders of magnitudein different structural contexts and under different solution con-ditions but comes apart with a common unfolding rate of ∼10 s−1
(Fig. 6C and SI Appendix, Fig. S3). Uncovering this commonalityrequired an ability to interrogate individual folding steps. Theresults suggest that the identity of the base pairs in the helicessurrounding this tertiary contact and the junctions that bring to-gether the tertiary contact partners do not affect the final structureof the tertiary motif or the energetics of the transition state en-semble for its disengagement. This observed modularity in tertiarycontact unfolding provides a first step toward using the energeticcontributions of individual tertiary motifs to predict the overallfolding kinetics and thermodynamics for complex RNAs (104).Stated another way, the data suggest a common rate for dis-
solution of a TL/TLR, a conclusion that, if general, could enablethe prediction of the rates of conformational steps of com-plex RNAs that involve breaking of a particular contact. Simi-larly, observing a conformational step with a rate constant thatis stereotypical for the dissociation of a particular tertiary in-teraction could suggest the breaking of that motif in the rate-limiting step, a model that could then be specifically tested bymutation of that motif. Conversely, tertiary contacts that do notfollow modular behavior would be suggested to have multiplefolded states and/or complex folding pathways and merit furtherdetailed structural and dynamic study.
A Generalized Model for RNA Tertiary Folding. A general goal indeveloping an RNA folding model (or any model) is to in-corporate the minimal number of features sufficient for accu-rate quantitative prediction. As described above, contact orderis a predictive, yet simple, concept in protein folding (97). Al-though folding of WT P4-P6 RNA follows the general pre-diction for a contact order model because the tertiary contactpairs that are closest to one another within the secondary structureform first [i.e., the MC/MCR forms first followed by the TL/TLR(Fig. 5)], this simple model fails to account for the reversal in the
order of tertiary contact formation with the G212C mutation(Fig. 7D).Modification of another established formalism from protein
folding, the diffusion–collision model, may better describe thefolding behaviors observed for P4-P6 and may prove moregeneral in RNA folding and assembly. This model applied toproteins posits that the rate of forming a tertiary interactionbetween alpha helices, beta strands, or hydrophobic clusters isdetermined by the rate of the regions of the protein colliding,weighted by the probability that the partner elements (e.g., twoalpha helices) are in a conformation competent for the tertiarycontact to form when the collision occurs (103, 105). There isevidence for intermediates in protein folding that are notaccounted for by this model (e.g., refs. 106–108). In contrast, giventhe more hierarchical nature of RNA than protein folding andthe discrete nature of RNA tertiary contacts, a diffusion–collision
Fig. 7. The kinetic effects of the G212C MCR mutation. (A) Comparison ofthe observed (orange) versus predicted (black and gray) overall folding (Left)and unfolding (Right) rate constants of WT P4-P6 and G212C P4-P6 (SIAppendix, Supporting Methods). For WT, the predicted rate constants arecalculated from the values in Fig. 5 that are reproduced in C of this figure.For G212C, the predicted folding and unfolding rate constants assume thefull equilibrium effect of the mutation either on the folding (black) orunfolding (gray) rate constant for the MC/MCR interaction. (B) Predictedvalues for the overall folding (blue) and unfolding (red) rate constants forthe G212C mutant over a range of hypothetical MC/MCR folding andunfolding rate constants, assuming that the full equilibrium effect of themutation (72-fold) is apportioned between these rate constants, rangingfrom the full effect of the mutation on the unfolding rate constant [(frac-tional effect on kf) = 0] to the full effect on the folding rate constant[(fractional effect on kf) = 1]. The observed overall folding and unfoldingrate constants for G212C are represented by the dashed lines. (C) Simplifiedfolding framework for WT P4-P6 (Left; values from Fig. 5) and G212C P4-P6(Right). Values for G212C P4-P6 are the same as for WT except for steps forMC/MCR formation, which are decreased by the full equilibrium effect of themutation (72-fold). (D) Free energy diagrams for WT P4-P6 (Left) and theG212C mutant (Right) folding. The preferred folding pathway is shown ingreen, and the flux through each pathway is given above each pathway andis designated by the relative arrow size. Profiles are to scale.
E4962 | www.pnas.org/cgi/doi/10.1073/pnas.1525082113 Bisaria et al.

model may be more readily and quantitatively applied to RNA, aselaborated below.RNA secondary is preformed and stable, and RNA tertiary
contact partners come together due to diffusion of the RNAhelices they are embedded in (66, 104, 109). RNA tertiary con-tact formation also typically requires local unfavorable confor-mational rearrangements (104, 110–115) such that the rates offormation of these interactions are orders of magnitude slowerthan diffusional collision (104, 116).In this way, RNA tertiary elements are potentially analogous
to protein secondary structure elements in the diffusion–collisionmodel. (Long-range secondary structure is also involved in RNA3D structure formation and can be considered along with tertiarycontacts in this model.) The probability that RNA tertiary con-tact partners associate is determined by (i) the conformationalproperties of the helices and the junctions that connect the ter-tiary elements and thus determine their collisional frequenciesand (ii) the properties of the tertiary elements that determinethe probability that both partners are in binding-competentconformations and the lifetimes of these states, relative to thelifetime of the collisional state. This process is akin to the needfor protein secondary structure elements to be formed to allowtertiary structure formation in the original diffusion–collisionmodel (103).Our P4-P6 folding results can be described in terms of this
RNA diffusion–collision model. Formation of one of P4-P6’stertiary interactions increases the rate constant (i.e., the proba-bility) of forming the other, presumably because the first contactgreatly restricts the conformational space explored by the secondtertiary contact elements and thereby increases their collisionalprobability and thus the rate of forming the second contact (Fig.5). Cooperativity, a fundamental feature of the folding of bi-ological macromolecules, is an emergent property of this in-creased collisional probability (60, 104).Mutations in J5/5a, which forms a bend in P4-P6 (Fig. 1 A and
B) (49, 51), slow formation of the TL/TLR contact, and linkingthe TL and TLR elements by a flexible single strand increasesthe folding rate; in contrast, neither alteration affects the unfoldingrate (Fig. 6C and SI Appendix, Table S2), consistent with differ-ent linkages imparting different collisional probabilities but notaffecting the contacts themselves. Collisional probabilities forRNA, as a polyelectrolyte, are also expected to be dependent onionic conditions, and accordingly we observe a steeper salt de-pendence on the rate constant for TL/TLR formation for themore highly charged P4-P6 construct than the TL/TLRiso con-structs (Fig. 6B and SI Appendix, Table S3). Finally, our resultssuggest that the G212C mutation decreases the probability of theMCR adopting its tertiary-competent conformation and therebyslows formation of the MC/MCR and reverses the order of ter-tiary contact formation in folding (Fig. 7D).We have established a P4-P6 RNA folding framework and
determined the effects of mutations within this RNA and alsocompared its folding to dramatically simplified constructs. Thedata emerging from these studies have allowed us to describe thefolding of P4-P6 in terms of an RNA diffusion–collision model.Although this description is, to our knowledge, the first suchdescription, there is no reason to think that such a descriptionwill not be generalizable because the model encompasses struc-tural features that are common to folded RNAs. Certainly, addi-tional complexities are expected, including parallel folding pathwaysfrom distinct secondary structure transitions, secondary structuretransitions during tertiary folding, and influences from proteinand other ligand binding. Thus, it will be of great interest to de-termine whether the RNA diffusion–collision model can be modifiedand adapted to describe this full range of behaviors and proper-ties and whether it provides a foundation for building a quan-titative and predictive understanding of RNA from the propertiesof its components.
Materials and MethodsFurther details of themethods are provided in SI Appendix, SupportingMethods.
Preparation of P4-P6 smFRET (smP4-P6) Variants. The smP4-P6 variants wereprepared by a five-piece splinted ligation with protocols slightly altered fromthose described previously (60, 90, 117). Prior studies revealed that standardlaboratory protocols introduce covalent differences in the RNA andthat these modifications can affect the folding of individual molecules(90). Alterations in the protocol include substituting a synthetically derived3′ and 5′ piece and removing intermediate gel purification and UV shadowingsteps (SI Appendix, Supporting Methods). These changes increase the yieldand throughput of the ligation procedure while producing RNA with thesame purity, kinetics and thermodynamics as assayed by smFRET and com-pared with RNAs prepared by differed methods (SI Appendix, Fig. S4).
Preparation of Isolated Constructs. Synthetic RNAs (SI Appendix, Tables S4and S5) for each isolated construct were purchased from IDT with a 3′ pri-mary amine, labeled with Cy3/Cy5 or Cy3b/Atto647N dyes with an NHS-esterreaction, and gel-purified.
smFRET Experiments. smFRET measurements were performed on a custom-built prism-based total internal reflection microscope. Buffers used in smFRETmeasurements contained 1–50 mM MgCl2 or 5 mM BaCl2, 50 mM MOPS,pH 8.0, 100 mM KCl, 2 mg/mL glucose, 1.8 mM Trolox, 100 units/mL glucoseoxidase (Sigma-Aldrich), and 1,000 units/mL catalase (Sigma-Aldrich). De-pending on the folding equilibrium and change in FRET for each molecule,smFRET data were collected at an acquisition rate between 50 and 310frames per second. Data were analyzed using the SMART data analysispackage (92). See Dataset S1 for the imaging parameters and buffer con-ditions for each measurement reported herein. Before imaging, P4-P6molecules were annealed to a DNA oligo containing a 5′ biotin (5′ biotin-AACCAAAATCAACCTAAAACTTACACA-3′) at 50 °C for 15 min and cooled at0.1 °C/min to room temperature. Samples were diluted to ∼10–100 pM forimaging. Prior smFRET studies have shown that the unfolded states ofP4-P6 have very low FRET (∼0.2) whereas states with the TL/TLR formed,including the fully folded state, have a high FRET value (∼0.8) and the in-termediate with only the MC/MCR formed has an intermediate FRET value(∼0.5) (Fig. 1C) (60). Also, prior P4-P6 smFRET work revealed molecularheterogeneity from covalent damage caused by standard RNA treatmentsand showed that this damage could be avoided to yield highly homoge-neous populations (90).
Data Analysis. Thermodynamic and kinetic parameters were inferred fromsingle-molecule FRET traces that were fit with a hidden Markov model(HMM)-based algorithm to a two-state model with a single unfolded (lowFRET = 0.16) and single folded (high FRET = 0.45 or 0.8) state to extracttransition probabilities, taking into account the noise observed in each in-tensity channel (92).
A subset of 20 traces for each construct and condition were also fit to athree-state linear model. In each case, the datawere found to fit better to thetwo-state model than to a three-state model based on the difference in theBayesian information criterion (BIC) (92), which includes a statistical penaltyfor including additional states, with the following exceptions that do notaffect the conclusions of this work: WT P4-P6 folding with a Cy3b/Atto674Ndye pair at 1 mM Mg, and MC/MCRiso at 5 and 10 mM Mg2+. (Example tracescan be found in Dataset S1.) For WT P4-P6 folding, this behavior may berelated to the dye dynamics because the apparent three-state behavior is notseen in variants with the Cy3/Cy5 dye pair. For MC/MCRiso, multistate behavior isseen only at low Mg2+ concentration, and not above 10 mM Mg2+. This con-struct has a lower folded FRET value than constructs that form the TL/TLRand fast transitions at lower [Mg2+], so that the signal-to-noise is relativelylow; thus, the apparent three-state behavior may arise from noise in thedata. We emphasize that elimination of the above data with potentialthree-state behavior would not change any of the conclusions of this work.We also note that, in general, best-fit models are sensitive to the penalty usedfor additional variables and the data quality and number of transitions (92).
Standard criteria for the identification of single-molecule traces wereused: (i ) single-step photobleaching, (ii) anticorrelated donor and accep-tor channels, (iii ) total intensity consistent with a single molecule, and(iv) stable total intensity. The folding rate constant reported for the J5/5a-AllU/ArichU P4-P6 variant is a limit because most traces for this moleculeshowed no folding behavior whereas traces that contained transitions hadon average fewer than two transitions, preventing reliable estimation ofthe folding rate constant (92). All rate and equilibrium constants reportedherein are the mean of the log distribution of the values for each molecule(118), and all other folding parameters are the mean of the distributionof each parameter for each molecule. Error estimates for all measure-ments, including the predictions of the overall folding and unfolding rate
Bisaria et al. PNAS | Published online August 4, 2016 | E4963
BIOCH
EMISTR
YPN
ASPL
US

constants for WT and G212C, were determined by 95% confidence inter-vals of the mean from bootstrapping the parameters from individualmolecules. Because the bootstrapped errors are asymmetric, the maxi-mum of the two values is reported as the error for simplicity in the maintext. Both errors are reported in SI Appendix. Signal-to-noise (SNR)cutoffs of 1 and 0.25 for molecules with high FRET states of 0.8 and 0.45,respectively, were used to minimize use of molecules for which fittingerrors contributed disproportionately to the inferred rate constants.Evaluation of smFRET data for each construct is presented in Dataset S1.
All smFRET data are available online for reanalysis in a standard single-molecule dataset (SMD) format (93).
ACKNOWLEDGMENTS. We thank Julie Fiore and David Nesbitt for gener-ously sharing their design and unpublished data on the isolated MC/MCRsmFRET construct; and the D.H. laboratory and Geeta Narlikar for helpfuldiscussions and for critical advice throughout this work. This work wassupported by National Institutes of Health Grant P01GM066275. N.B. wassupported by an NSF graduate fellowship. M.G. was supported in part by NIHBiotechnology Training Grant GM008412.
1. Warf MB, Berglund JA (2010) Role of RNA structure in regulating pre-mRNA splicing.Trends Biochem Sci 35(3):169–178.
2. Rinn JL, Chang HY (2012) Genome regulation by long noncoding RNAs. Annu RevBiochem 81(1):145–166.
3. Mercer TR, Mattick JS (2013) Structure and function of long noncoding RNAs inepigenetic regulation. Nat Struct Mol Biol 20(3):300–307.
4. Cech TR, Steitz JA (2014) The noncoding RNA revolution-trashing old rules to forgenew ones. Cell 157(1):77–94.
5. Staley JP, Guthrie C (1998) Mechanical devices of the spliceosome: Motors, clocks,springs, and things. Cell 92(3):315–326.
6. Moore MJ (2005) From birth to death: The complex lives of eukaryotic mRNAs.Science 309(5740):1514–1518.
7. Carthew RW, Sontheimer EJ (2009) Origins and mechanisms of miRNAs and siRNAs.Cell 136(4):642–655.
8. Bartel DP (2009) MicroRNAs: Target recognition and regulatory functions. Cell136(2):215–233.
9. Orgel LE (1968) Evolution of the genetic apparatus. J Mol Biol 38(3):381–393.10. Woese C (1967) The Genetic Code: The Molecular Basis for Genetic Expression
(Harper & Row, New York).11. Crick FH (1968) The origin of the genetic code. J Mol Biol 38(3):367–379.12. Gilbert W (1986) Origin of life: The RNA world. Nature 319(6055):618.13. Lindahl T, Adams A, Fresco JR (1966) Renaturation of transfer ribonucleic acids
through site binding of magnesium. Proc Natl Acad Sci USA 55(4):941–948.14. Adams A, Lindahl T, Fresco JR (1967) Conformational differences between the bi-
ologically active and inactive forms of a transfer ribonucleic acid. Proc Natl Acad SciUSA 57(6):1684–1691.
15. Ishida T, Sueoka N (1968) Elimination of magnesium ions as an absolute requirementfor the native conformation of tryptophan transfer ribonucleic acid. J Mol Biol 37(2):313–316.
16. Walstrum SA, Uhlenbeck OC (1990) The self-splicing RNA of Tetrahymena is trappedin a less active conformation by gel purification. Biochemistry 29(46):10573–10576.
17. Uhlenbeck OC (1995) Keeping RNA happy. RNA 1(1):4–6.18. Treiber DK, Williamson JR (1999) Exposing the kinetic traps in RNA folding. Curr
Opin Struct Biol 9(3):339–345.19. Shcherbakova I, Mitra S, Laederach A, Brenowitz M (2008) Energy barriers, path-
ways, and dynamics during folding of large, multidomain RNAs. Curr Opin Chem Biol12(6):655–666.
20. Russell R (2008) RNA misfolding and the action of chaperones. Front Biosci 13:1–20.21. Sigler PB (1975) An analysis of the structure of tRNA. Annu Rev Biophys Bioeng 4(00):
477–527.22. Herschlag D (1995) RNA chaperones and the RNA folding problem. J Biol Chem
270(36):20871–20874.23. Jiang W, Hou Y, Inouye M (1997) CspA, the major cold-shock protein of Escherichia
coli, is an RNA chaperone. J Biol Chem 272(1):196–202.24. Semrad K, Green R, Schroeder R (2004) RNA chaperone activity of large ribosomal
subunit proteins from Escherichia coli. RNA 10(12):1855–1860.25. Lorsch JR (2002) RNA chaperones exist and DEAD box proteins get a life. Cell 109(7):
797–800.26. Schroeder R, Barta A, Semrad K (2004) Strategies for RNA folding and assembly. Nat
Rev Mol Cell Biol 5(11):908–919.27. Woodson SA (2010) Taming free energy landscapes with RNA chaperones. RNA Biol
7(6):677–686.28. Rein A (2010) Nucleic acid chaperone activity of retroviral Gag proteins. RNA Biol
7(6):700–705.29. Woodson SA, Cech TR (1991) Alternative secondary structures in the 5′ exon affect
both forward and reverse self-splicing of the Tetrahymena intervening sequenceRNA. Biochemistry 30(8):2042–2050.
30. Pan J, Woodson SA (1998) Folding intermediates of a self-splicing RNA: Mispairing ofthe catalytic core. J Mol Biol 280(4):597–609.
31. Treiber DK, Rook MS, Zarrinkar PP, Williamson JR (1998) Kinetic intermediatestrapped by native interactions in RNA folding. Science 279(5358):1943–1946.
32. Chadalavada DM, Senchak SE, Bevilacqua PC (2002) The folding pathway of thegenomic hepatitis delta virus ribozyme is dominated by slow folding of the pseu-doknots. J Mol Biol 317(4):559–575.
33. Russell R, et al. (2002) Exploring the folding landscape of a structured RNA. Proc NatlAcad Sci USA 99(1):155–160.
34. Pan J, Deras ML, Woodson SA (2000) Fast folding of a ribozyme by stabilizing coreinteractions: Evidence for multiple folding pathways in RNA. J Mol Biol 296(1):133–144.
35. Solomatin SV, Greenfeld M, Chu S, Herschlag D (2010) Multiple native statesreveal persistent ruggedness of an RNA folding landscape. Nature 463(7281):681–684.
36. Russell R, Herschlag D (2001) Probing the folding landscape of the Tetrahymenaribozyme: Commitment to form the native conformation is late in the foldingpathway. J Mol Biol 308(5):839–851.
37. Russell R, et al. (2006) The paradoxical behavior of a highly structured misfoldedintermediate in RNA folding. J Mol Biol 363(2):531–544.
38. Mitchell D, 3rd, Jarmoskaite I, Seval N, Seifert S, Russell R (2013) The long-range P3helix of the Tetrahymena ribozyme is disrupted during folding between the nativeand misfolded conformations. J Mol Biol 425(15):2670–2686.
39. Albery WJ, Knowles JR (1976) Free-energy profile of the reaction catalyzed by tri-osephosphate isomerase. Biochemistry 15(25):5627–5631.
40. Gresser MJ, Myers JA, Boyer PD (1982) Catalytic site cooperativity of beef heartmitochondrial F1 adenosine triphosphatase: Correlations of initial velocity, boundintermediate, and oxygen exchange measurements with an alternating three-sitemodel. J Biol Chem 257(20):12030–12038.
41. Fierke CA, Johnson KA, Benkovic SJ (1987) Construction and evaluation of the kineticscheme associated with dihydrofolate reductase from Escherichia coli. Biochemistry26(13):4085–4092.
42. Patel SS, Wong I, Johnson KA (1991) Pre-steady-state kinetic analysis of processiveDNA replication including complete characterization of an exonuclease-deficientmutant. Biochemistry 30(2):511–525.
43. Ma YZ, Taylor EW (1997) Kinetic mechanism of a monomeric kinesin construct. J BiolChem 272(2):717–723.
44. Mullins RD, Heuser JA, Pollard TD (1998) The interaction of Arp2/3 complex withactin: Nucleation, high affinity pointed end capping, and formation of branchingnetworks of filaments. Proc Natl Acad Sci USA 95(11):6181–6186.
45. Zhang X, Rashid R, Wang K, Shan S-O (2010) Sequential checkpoints govern sub-strate selection during cotranslational protein targeting. Science 328(5979):757–760.
46. Das R, Travers KJ, Bai Y, Herschlag D (2005) Determining the Mg2+ stoichiometry forfolding an RNA metal ion core. J Am Chem Soc 127(23):8272–8273.
47. Murphy FL, Cech TR (1993) An independently folding domain of RNA tertiarystructure within the Tetrahymena ribozyme. Biochemistry 32(20):5291–5300.
48. Murphy FL, Wang YH, Griffith JD, Cech TR (1994) Coaxially stacked RNA helices in thecatalytic center of the Tetrahymena ribozyme. Science 265(5179):1709–1712.
49. Cate JH, et al. (1996) Crystal structure of a group I ribozyme domain: Principles ofRNA packing. Science 273(5282):1678–1685.
50. Cate JH, Hanna RL, Doudna JA (1997) A magnesium ion core at the heart of a ri-bozyme domain. Nat Struct Biol 4(7):553–558.
51. Szewczak AA, Cech TR (1997) An RNA internal loop acts as a hinge to facilitate ri-bozyme folding and catalysis. RNA 3(8):838–849.
52. Sclavi B, Sullivan M, Chance MR, Brenowitz M, Woodson SA (1998) RNA folding atmillisecond intervals by synchrotron hydroxyl radical footprinting. Science 279(5358):1940–1943.
53. Deras ML, Brenowitz M, Ralston CY, Chance MR, Woodson SA (2000) Folding mecha-nism of the Tetrahymena ribozyme P4-P6 domain. Biochemistry 39(36):10975–10985.
54. Silverman SK, Deras ML, Woodson SA, Scaringe SA, Cech TR (2000) Multiple foldingpathways for the P4-P6 RNA domain. Biochemistry 39(40):12465–12475.
55. Silverman SK, Cech TR (2001) An early transition state for folding of the P4-P6 RNAdomain. RNA 7(2):161–166.
56. Takamoto K, et al. (2004) Principles of RNA compaction: Insights from the equilib-rium folding pathway of the P4-P6 RNA domain in monovalent cations. J Mol Biol343(5):1195–1206.
57. Das R, Baker D (2007) Automated de novo prediction of native-like RNA tertiarystructures. Proc Natl Acad Sci USA 104(37):14664–14669.
58. Murphy FL, Cech TR (1994) GAAA tetraloop and conserved bulge stabilize tertiarystructure of a group I intron domain. J Mol Biol 236(1):49–63.
59. Das R, et al. (2008) Structural inference of native and partially folded RNA by high-throughput contact mapping. Proc Natl Acad Sci USA 105(11):4144–4149.
60. Sattin BD, Zhao W, Travers K, Chu S, Herschlag D (2008) Direct measurement oftertiary contact cooperativity in RNA folding. J Am Chem Soc 130(19):6085–6087.
61. Silverman SK, Cech TR (1999) RNA tertiary folding monitored by fluorescence ofcovalently attached pyrene. Biochemistry 38(43):14224–14237.
62. Young BT, Silverman SK (2002) The GAAA tetraloop-receptor interaction contributesdifferentially to folding thermodynamics and kinetics for the P4-P6 RNA domain.Biochemistry 41(41):12271–12276.
63. Lee TH, et al. (2007) Measuring the folding transition time of single RNA molecules.Biophys J 92(9):3275–3283.
64. Lamb J, et al. (2008) Reconstructing three-dimensional shape envelopes from time-resolved small-angle X-ray scattering data. J Appl Cryst 41(Pt 6):1046–1052.
65. Schlatterer JC, et al. (2008) Hinge stiffness is a barrier to RNA folding. J Mol Biol379(4):859–870.
66. Leontis NB, Westhof E (2003) Analysis of RNA motifs. Curr Opin Struct Biol 13(3):300–308.
E4964 | www.pnas.org/cgi/doi/10.1073/pnas.1525082113 Bisaria et al.

67. Tinoco I, Jr, Bustamante C (1999) How RNA folds. J Mol Biol 293(2):271–281.68. Klein DJ, Schmeing TM, Moore PB, Steitz TA (2001) The kink-turn: A new RNA sec-
ondary structure motif. EMBO J 20(15):4214–4221.69. Lukavsky PJ, Kim I, Otto GA, Puglisi JD (2003) Structure of HCV IRES domain II de-
termined by NMR. Nat Struct Biol 10(12):1033–1038.70. Wu L, Chai D, Fraser ME, Zimmerly S (2012) Structural variation and uniformity
among tetraloop-receptor interactions and other loop-helix interactions in RNAcrystal structures. PLoS One 7(11):e49225.
71. Ishikawa J, Fujita Y, Maeda Y, Furuta H, Ikawa Y (2011) GNRA/receptor interactingmodules: Versatile modular units for natural and artificial RNA architectures.Methods 54(2):226–238.
72. Costa M, Michel F (1995) Frequent use of the same tertiary motif by self-foldingRNAs. EMBO J 14(6):1276–1285.
73. Geary C, Baudrey S, Jaeger L (2008) Comprehensive features of natural and in vitroselected GNRA tetraloop-binding receptors. Nucleic Acids Res 36(4):1138–1152.
74. Nissen P, Ippolito JA, Ban N, Moore PB, Steitz TA (2001) RNA tertiary interactions in thelarge ribosomal subunit: The A-minor motif. Proc Natl Acad Sci USA 98(9):4899–4903.
75. Nagaswamy U, Fox GE (2002) Frequent occurrence of the T-loop RNA folding motifin ribosomal RNAs. RNA 8(9):1112–1119.
76. Basu S, et al. (1998) A specific monovalent metal ion integral to the AA platform ofthe RNA tetraloop receptor. Nat Struct Biol 5(11):986–992.
77. Wu M, Tinoco I, Jr (1998) RNA folding causes secondary structure rearrangement.Proc Natl Acad Sci USA 95(20):11555–11560.
78. Uchida T, He Q, Ralston CY, Brenowitz M, Chance MR (2002) Linkage of monovalentand divalent ion binding in the folding of the P4-P6 domain of the Tetrahymenaribozyme. Biochemistry 41(18):5799–5806.
79. Battle DJ, Doudna JA (2002) Specificity of RNA-RNA helix recognition. Proc Natl AcadSci USA 99(18):11676–11681.
80. Schwans JP, Cortez CN, Olvera JM, Piccirilli JA (2003) 2′-mercaptonucleotide in-terference reveals regions of close packing within folded RNA molecules. J Am ChemSoc 125(33):10012–10018.
81. Travers KJ, Boyd N, Herschlag D (2007) Low specificity of metal ion binding in themetal ion core of a folded RNA. RNA 13(8):1205–1213.
82. Frederiksen JK, Li N-S, Das R, Herschlag D, Piccirilli JA (2012) Metal-ion rescue re-visited: Biochemical detection of site-bound metal ions important for RNA folding.RNA 18(6):1123–1141.
83. Wang S, Karbstein K, Peracchi A, Beigelman L, Herschlag D (1999) Identification ofthe hammerhead ribozyme metal ion binding site responsible for rescue of thedeleterious effect of a cleavage site phosphorothioate. Biochemistry 38(43):14363–14378.
84. Shan So, Yoshida A, Sun S, Piccirilli JA, Herschlag D (1999) Three metal ions at theactive site of the Tetrahymena group I ribozyme. Proc Natl Acad Sci USA 96(22):12299–12304.
85. Maderia M, Hunsicker LM, DeRose VJ (2000) Metal-phosphate interactions in thehammerhead ribozyme observed by 31P NMR and phosphorothioate substitutions.Biochemistry 39(40):12113–12120.
86. Murray JB, Dunham CM, Scott WG (2002) A pH-dependent conformational change,rather than the chemical step, appears to be rate-limiting in the hammerhead ri-bozyme cleavage reaction. J Mol Biol 315(2):121–130.
87. Pyle AM (2002) Metal ions in the structure and function of RNA. J Biol Inorg Chem7(7-8):679–690.
88. Thomson JB, Lilley DM (1999) The influence of junction conformation on RNAcleavage by the hairpin ribozyme in its natural junction form. RNA 5(2):180–187.
89. Dibrov SM, McLean J, Parsons J, Hermann T (2011) Self-assembling RNA square. ProcNatl Acad Sci USA 108(16):6405–6408.
90. Greenfeld M, Solomatin SV, Herschlag D (2011) Removal of covalent heterogeneityreveals simple folding behavior for P4-P6 RNA. J Biol Chem 286(22):19872–19879.
91. Leipply D, Draper DE (2010) Dependence of RNA tertiary structural stability on Mg2+
concentration: Interpretation of the Hill equation and coefficient. Biochemistry49(9):1843–1853.
92. Greenfeld M, Pavlichin DS, Mabuchi H, Herschlag D (2012) Single Molecule AnalysisResearch Tool (SMART): An integrated approach for analyzing single molecule data.PLoS One 7(2):e30024.
93. Greenfeld M, et al. (2015) Single-molecule dataset (SMD): A generalized storageformat for raw and processed single-molecule data. BMC Bioinformatics 16:3.
94. Silverman SK, Zheng M, Wu M, Tinoco I, Jr, Cech TR (1999) Quantifying the energeticinterplay of RNA tertiary and secondary structure interactions. RNA 5(12):1665–1674.
95. Cate JH, Doudna JA (1996) Metal-binding sites in the major groove of a large ri-bozyme domain. Structure 4(10):1221–1229.
96. Silverman SK, Cech TR (1999) Energetics and cooperativity of tertiary hydrogenbonds in RNA structure. Biochemistry 38(27):8691–8702.
97. Plaxco KW, Simons KT, Baker D (1998) Contact order, transition state placement andthe refolding rates of single domain proteins. J Mol Biol 277(4):985–994.
98. Sosnick TR, Pan T (2004) Reduced contact order and RNA folding rates. J Mol Biol342(5):1359–1365.
99. Heilman-Miller SL, Woodson SA (2003) Perturbed folding kinetics of circularly per-muted RNAs with altered topology. J Mol Biol 328(2):385–394.
100. Hodak JH, Downey CD, Fiore JL, Pardi A, Nesbitt DJ (2005) Docking kinetics andequilibrium of a GAAA tetraloop-receptor motif probed by single-molecule FRET.Proc Natl Acad Sci USA 102(30):10505–10510.
101. Downey CD, et al. (2006) Metal ion dependence, thermodynamics, and kinetics forintramolecular docking of a GAAA tetraloop and receptor connected by a flexiblelinker. Biochemistry 45(11):3664–3673.
102. Fiore JL, Holmstrom ED, Nesbitt DJ (2012) Entropic origin of Mg2+-facilitated RNAfolding. Proc Natl Acad Sci USA 109(8):2902–2907.
103. Karplus M, Weaver DL (1976) Protein-folding dynamics. Nature 260(5550):404–406.104. Herschlag D, Allred BE, Gowrishankar S (2015) From static to dynamic: The need for
structural ensembles and a predictive model of RNA folding and function. Curr OpinStruct Biol 30:125–133.
105. Karplus M, Weaver DL (1994) Protein folding dynamics: The diffusion-collision modeland experimental data. Protein Sci 3(4):650–668.
106. Ohgushi M, Wada A (1983) ‘Molten-globule state’: A compact form of globularproteins with mobile side-chains. FEBS Lett 164(1):21–24.
107. Baldwin RL, Rose GD (2013) Molten globules, entropy-driven conformational changeand protein folding. Curr Opin Struct Biol 23(1):4–10.
108. Bachmann A, Wildemann D, Praetorius F, Fischer G, Kiefhaber T (2011) Mappingbackbone and side-chain interactions in the transition state of a coupled proteinfolding and binding reaction. Proc Natl Acad Sci USA 108(10):3952–3957.
109. Chu VB, et al. (2009) Do conformational biases of simple helical junctions influenceRNA folding stability and specificity? RNA 15(12):2195–2205.
110. Butcher SE, Dieckmann T, Feigon J (1997) Solution structure of a GAAA tetraloopreceptor RNA. EMBO J 16(24):7490–7499.
111. Al-Hashimi HM, Walter NG (2008) RNA dynamics: It is about time. Curr Opin StructBiol 18(3):321–329.
112. Jiang F, Kumar RA, Jones RA, Patel DJ (1996) Structural basis of RNA folding andrecognition in an AMP-RNA aptamer complex. Nature 382(6587):183–186.
113. Allain FH, et al. (1996) Specificity of ribonucleoprotein interaction determined byRNA folding during complex formulation. Nature 380(6575):646–650.
114. Puglisi JD, Tan R, Calnan BJ, Frankel AD, Williamson JR (1992) Conformation of theTAR RNA-arginine complex by NMR spectroscopy. Science 257(5066):76–80.
115. Sussman D, Nix JC, Wilson C (2000) The structural basis for molecular recognition bythe vitamin B 12 RNA aptamer. Nat Struct Biol 7(1):53–57.
116. Karbstein K, Herschlag D (2003) Extraordinarily slow binding of guanosine to theTetrahymena group I ribozyme: Implications for RNA preorganization and function.Proc Natl Acad Sci USA 100(5):2300–2305.
117. Solomatin SV, Greenfeld M, Herschlag D (2011) Implications of molecular hetero-geneity for the cooperativity of biological macromolecules.Nat Struct Mol Biol 18(6):732–734.
118. McKinney SA, Joo C, Ha T (2006) Analysis of single-molecule FRET trajectories usinghidden Markov modeling. Biophys J 91(5):1941–1951.
Bisaria et al. PNAS | Published online August 4, 2016 | E4965
BIOCH
EMISTR
YPN
ASPL
US


















