
Journal of Archaeological Science - Arts & Sciences Pages · A new system for computing long-bone...
16
A new system for computing long-bone fusion age profiles in Sus scrofa * Melinda A. Zeder a, * , Ximena Lemoine b , Sebastian Payne c a Program in Human Ecology and Archaeobiology, Department of Anthropology, National Museum of Natural History, Smithsonian Institution, USA b Department of Anthropology, Washington University of St. Louis, St. Louis, USA c English Heritage, UK article info Article history: Received 6 November 2014 Received in revised form 22 December 2014 Accepted 24 December 2014 Available online 3 January 2015 Keywords: Aging Sus scrofa Pigs Harvest profiles Epiphyseal fusion abstract In this paper we present the results of a study of post-cranial fusion in pigs (Sus scrofa) and propose a new system for the construction of harvest profiles of pigs based on epiphyseal fusion. The study examined post-crania of 40 Asian wild boar in museum and personal collections. It finds a regular pattern in the sequence of fusion of elements in this sample that also agrees with the fusion sequences of 56 European wild boar published in earlier studies. The fusion sequence of post-cranial elements is grouped into eleven different age classes (AeK). Comparison of the dentition based age classes assigned to 38 of the wild boar studied here and in an earlier study (Lemoine et al., 2014) shows a close corre- spondence between dental and fusion based age classes. Although the age at death of these specimens is not known, it is possible to assign age estimates for the fusion based age classes defined here based on the relatively secure age estimates for the dentition based age classes. A comparison of the fusion based harvest profile for a large assemblage of pig remains from the Epipaleolithic site of Hallan Çemi (southeastern Anatolia) constructed using the system proposed here with dentition based profiles using the three systems proposed in Lemoine et al. shows a very close correspondence, especially in the younger age classes. We conclude with a consideration of the strengths and weaknesses of fusion based and dentition based harvest profiles finding that when taphonomic conditions permit fusion based harvest profiles are a valuable tool for understanding ancient exploitation strategies, especially when used in tandem with dentition based profiles. Published by Elsevier Ltd. 1. Introduction The value of harvest profiles for understanding ancient animal exploitation strategies has long been recognized (Dyson, 1953; Perkins, 1964; Hole et al., 1969; Redding, 1981; Payne, 1973; Stiner, 1990; Zeder, 1991). The importance of high resolution har- vest profiles in tracking the transition from hunting to herding has become especially apparent in recent years with the recognition that subtle shifts in the ages and sexes of harvested animals pre- cedes the manifestation of archaeologically detectable morpho- logical change in managed animals by hundreds, if not thousands of years (Helmer, 2008; Vigne et al., 2012; Zeder, 2006a, 2008). Considerable advances have been made in our ability to construct fine-grained harvest profiles in a variety of animals, with caprines (sheep and goats) being a primary focus of this work (Jones, 2006; Moran and O'Connor, 1994; Popkin et al., 2012; Zeder, 2006b). In contrast, our ability to reconstruct harvest profiles in another pri- mary livestock species e the pig (Sus scrofa), has lagged far behind. This is especially true for pig harvest profiles based on long bone fusion patterns. With the exception of early papers by Bull and Payne (1982) and van Wijngaarden-Bakker and Maliepaard (1982), and a more recent publication by Bridault et al. (2000), the primary data used to fix the sequence and age of fusion for pig long bones are restricted to a handful of sources of uncertain, and in some cases questionable, value (Habermehl, 1975; Lesbre, 1897/ 1898; Reiland, 1978; Silver, 1969; see Legge, nd). Confusion exists over how the moment of fusion was defined in these sources, how observations were made (direct vs. radiometric observation), and, most significantly, the identity and size of the populations used in making determinations of the sequence and timing of long bone * Funding: National Geographic Society, Wenner Gren Foundation; Smithsonian Institution. * Corresponding author. Tel.: þ1 703 626 9118. E-mail address: [email protected] (M.A. Zeder). Contents lists available at ScienceDirect Journal of Archaeological Science journal homepage: http://www.elsevier.com/locate/jas http://dx.doi.org/10.1016/j.jas.2014.12.017 0305-4403/Published by Elsevier Ltd. Journal of Archaeological Science 55 (2015) 135e150
Transcript of Journal of Archaeological Science - Arts & Sciences Pages · A new system for computing long-bone...

lable at ScienceDirect
Journal of Archaeological Science 55 (2015) 135e150
Contents lists avai
Journal of Archaeological Science
journal homepage: http : / /www.elsevier .com/locate/ jas
A new system for computing long-bone fusion age profiles in Susscrofa*
Melinda A. Zeder a, *, Ximena Lemoine b, Sebastian Payne c
a Program in Human Ecology and Archaeobiology, Department of Anthropology, National Museum of Natural History, Smithsonian Institution, USAb Department of Anthropology, Washington University of St. Louis, St. Louis, USAc English Heritage, UK
a r t i c l e i n f o
Article history:Received 6 November 2014Received in revised form22 December 2014Accepted 24 December 2014Available online 3 January 2015
Keywords:AgingSus scrofaPigsHarvest profilesEpiphyseal fusion
* Funding: National Geographic Society, Wenner GInstitution.* Corresponding author. Tel.: þ1 703 626 9118.
E-mail address: [email protected] (M.A. Zeder).
http://dx.doi.org/10.1016/j.jas.2014.12.0170305-4403/Published by Elsevier Ltd.
a b s t r a c t
In this paper we present the results of a study of post-cranial fusion in pigs (Sus scrofa) and propose anew system for the construction of harvest profiles of pigs based on epiphyseal fusion. The studyexamined post-crania of 40 Asian wild boar in museum and personal collections. It finds a regularpattern in the sequence of fusion of elements in this sample that also agrees with the fusion sequences of56 European wild boar published in earlier studies. The fusion sequence of post-cranial elements isgrouped into eleven different age classes (AeK). Comparison of the dentition based age classes assignedto 38 of the wild boar studied here and in an earlier study (Lemoine et al., 2014) shows a close corre-spondence between dental and fusion based age classes. Although the age at death of these specimens isnot known, it is possible to assign age estimates for the fusion based age classes defined here based onthe relatively secure age estimates for the dentition based age classes. A comparison of the fusion basedharvest profile for a large assemblage of pig remains from the Epipaleolithic site of Hallan Çemi(southeastern Anatolia) constructed using the system proposed here with dentition based profiles usingthe three systems proposed in Lemoine et al. shows a very close correspondence, especially in theyounger age classes. We conclude with a consideration of the strengths and weaknesses of fusion basedand dentition based harvest profiles finding that when taphonomic conditions permit fusion basedharvest profiles are a valuable tool for understanding ancient exploitation strategies, especially whenused in tandem with dentition based profiles.
Published by Elsevier Ltd.
1. Introduction
The value of harvest profiles for understanding ancient animalexploitation strategies has long been recognized (Dyson, 1953;Perkins, 1964; Hole et al., 1969; Redding, 1981; Payne, 1973;Stiner, 1990; Zeder, 1991). The importance of high resolution har-vest profiles in tracking the transition from hunting to herding hasbecome especially apparent in recent years with the recognitionthat subtle shifts in the ages and sexes of harvested animals pre-cedes the manifestation of archaeologically detectable morpho-logical change inmanaged animals by hundreds, if not thousands ofyears (Helmer, 2008; Vigne et al., 2012; Zeder, 2006a, 2008).
ren Foundation; Smithsonian
Considerable advances have been made in our ability to constructfine-grained harvest profiles in a variety of animals, with caprines(sheep and goats) being a primary focus of this work (Jones, 2006;Moran and O'Connor, 1994; Popkin et al., 2012; Zeder, 2006b). Incontrast, our ability to reconstruct harvest profiles in another pri-mary livestock species e the pig (Sus scrofa), has lagged far behind.This is especially true for pig harvest profiles based on long bonefusion patterns. With the exception of early papers by Bull andPayne (1982) and van Wijngaarden-Bakker and Maliepaard(1982), and a more recent publication by Bridault et al. (2000), theprimary data used to fix the sequence and age of fusion for pig longbones are restricted to a handful of sources of uncertain, and insome cases questionable, value (Habermehl, 1975; Lesbre, 1897/1898; Reiland, 1978; Silver, 1969; see Legge, nd). Confusion existsover how the moment of fusion was defined in these sources, howobservations were made (direct vs. radiometric observation), and,most significantly, the identity and size of the populations used inmaking determinations of the sequence and timing of long bone

M.A. Zeder et al. / Journal of Archaeological Science 55 (2015) 135e150136
fusion (Legge, nd). As a result, long bone harvest profiles based onthese sources lack both the resolution and the accuracy needed todetect critical shifts in harvest strategies that mark the transitionfrom hunting to herding in pigs or that reflect the very differentapproaches to pig husbandry in the past.
This paper is the second in a series that seeks to redress currentlimitations in methods for reconstructing harvest profiles in pigs.The first of these (Lemoine et al., 2014) presents a new system forcomputing dentition based age profiles based on a study of 91paired mandibles and maxillae of modern Asian wild boar. Thepresent paper focuses on a new system for computing pigepiphyseal fusion age profiles based on the study of the post-crania of 40 Asian wild boar. This paper also examines the corre-lation between this fusion aging system and the earlier describeddental aging system. A future paper will consider the degree andthe timing of the onset of detectable sexual dimorphism in theteeth and post-crania of this large sample of modern boars. Theultimate goal of this work is to provide a robust and empiricallygrounded system for the construction of high resolution sex-specific harvest profiles in pigs capable of discriminating amongvarious exploitation strategies in the harvest of both free-livingand managed pigs.
2. Method: a revision
2.1. The modern sample
As in the earlier study, the modern specimens utilized in thisresearch derive from the Field Museum of Natural History inChicago (FMNH), the Division of Mammals of the NationalMuseum of Natural History, Smithsonian Institution, in Washing-ton D.C. (NMNH) and Payne's personal collection of Turkish wildboar (SP). A total of 40 specimens were included in this study(Table S1), consisting of 25 females, 14 males, and one specimen ofunknown sex. Age at death of the specimens is not known.Specimens derive primarily from West Asia (Iran, Iraq, andTurkey), with four of the 40 specimens from India. Also as in theprevious study, all specimens have accompanying documentationthat confirms their status as free-living individuals. The smallernumber of specimens included in this study of long bone fusionthan in the previous study of dental eruption and wear (40 vs. 91specimens) reflects the common (and regrettable) practice of onlyretaining the skulls of collected specimens whose post-crania arediscarded. Observations of dental eruption and wear were recor-ded on 38 of the 40 specimens included in this study, making itpossible to compare the two aging systems resulting from thestudy of these specimens.
Once again, the broad geographic range of the sample (at leastseven collecting localities in four countries) is tempered by thelarge sample of wild boar from forests around Kızılcahamam incentral Anatolia in Turkey collected by Payne during the winter of1974e1975 (Bull and Payne, 1982). The large geographic range ofthe sample allows for an examination of variability in fusion se-quences among different populations of wild boar. At the sametime, the number of specimens from a single population of animalswith presumably the same diet and environmental context pro-vides an anchor for the patterns observed in more geographicallyand temporally dispersed specimens. As before we decided not toinclude domestic specimens in this study to avoid any confoundinginfluences of improved breeding, dietary supplements, or nutri-tional differences on epiphyseal fusion. As we state in our earlierpaper on dental eruption and wear, while it is possible that suchdifferences may affect the timing of long-bone fusion, we believethat the sequence of fusion will remain constant regardless of diet.
2.2. Scoring
Post-cranial elements examined in this study include the atlas(dorsal and ventral aspects), axis (cranial and distal), pelvis (ace-tabulum), scapula (coracoid and glenoid fossa), humerus (proximaland distal articulations), radius (proximal and distal articulations),ulna (proximal and distal articulations and fusion to radius), femur(proximal and distal articulations), tibia (proximal and distal ar-ticulations), fibula (proximal and distal articulations), metapodials(distal articulation), and first and second phalanges (proximal ar-ticulations) (Table S2). Articulations examined were scored as un-fused (1) when the epiphysis was separate from the diaphysis or, incases where cartilage remained, when a clear separation betweenthe epiphysis and diaphysis could be observed. Elements werescored as fusing (2) when there was evidence of bone knitting, butwith visible openings in the fusion plates remaining. The score ofearly fusion (3) was reserved for instances when the fusion wascomplete but a clear line of fusion was visible. Elements werescored as fully fused (4) when this line was indistinct or absent.Elements of very young animals (i.e. very small elements of poroustexture) were scored as neo-natal (0).
Specimens in the Payne collection were lacking humeri andtibiae (which had been removed for another study and have notyet been returned). When these elements were unfused, epiph-yses remained with the rest of the specimens and were scoreddirectly. For specimens in which the epiphyses of these boneswere fused, and for one specimen of the original collection thatwas unavailable for study (SP350), the fusion data reported inBull and Payne (1982) was used here instead. Scores taken fromBull and Payne are indicated in red in Table S2. This table alsonotes when the scoring done for this study differs from thosepublished in Bull and Payne. The original scores are indicated inred and presented in parentheses next to the score recorded forthis study.
2.3. Defining age classes
Once scored, a table was constructed with skeletal elements incolumns organized in order of fusion and specimens presented inrows organized by degree of fusion across the various elements(Fig. 1). There are clear and consistent patterns in the sequence offusion across the scored post-cranial elements, with distinctgroupings in the timing of fusion of various elements. These pat-terns are highlighted in Fig. 1 by shading elements that are eitherfusing (2) or in early fusion (3) in gray. Given the difficulty of dis-tinguishing between these two fusion phases in bones with sub-stantial amount of cartilage and other tissue, it is possible that someelements recorded as fusing were actually in early fusion and somerecorded as in early fusion were, in fact, fusing. It is also importantto note that this ‘early fusion’ phase was not recognized in Bull andPayne, so there may be some inconsistencies in scoring of missingelements in the Payne collection for fusion status was taken fromBull and Payne.
Based on these fusion patterns for the 40 specimens included inthis study it is possible to define 11 different age classes (Fig. 1):
Age Class A: Age class A is based on one neo-natal specimen.Age Class B: Three specimens grouped together in age class Bwhose post-crania were, like the specimen in age class A, allunfused but were clearly a little older than the neo-natal spec-imen in age class A.Age Class C: Age class C captures the fusion of the neural archand dens of the axis and the two halves of the atlas. Of the twoelements in this age class, the neural arch and dens of the axisseem to fuse slightly earlier than the atlas.

Fig. 1. Fusion stages for modern Sus specimens. 0 ¼ neonatal, 1 ¼ unfused, 2 ¼ fusing, 3 ¼ recently fused, 4 ¼ fully fused, - ¼ no data, (x) data from Bull and Payne that differs fromcurrent analysis. ID # from Table S1, Dr ¼ Dorsal, V ¼ Ventral, Art ¼ Articulation, Cor ¼ Coracoid, P ¼ Proximal, D ¼ Distal, ME ¼ Medial Epicondyle, T ¼ Trochanter, H ¼ Head, Tu ¼Tubercle.
M.A. Zeder et al. / Journal of Archaeological Science 55 (2015) 135e150 137
Age Class D: Three specimens were grouped together in age classD, defined by the fusion of the three elements that make up thepelvis (the pubis, ilium, and ischium) at the acetabulum, thefusion of the glenoid cavity and coracoid process of the scapula,and the fusion of the proximal articulation of the radius. Of theseelements, the glenoid cavity of the scapula and the pelvis seemto fuse earlier than the coracoid process of the scapula. Fusion ofthe proximal radius seems to be a bit later, but is still distinctfrom the next age class.Age Class E: Age class E captures the fusion of the second pha-lanx and both the trochlea and the medial epicondyle of thedistal end of the humerus. Of these elements, the second pha-lanx seems to fuse a bit earlier than the distal humerus. Distalhumerus scores for the Payne specimens in this, and all subse-quent age classes were not observed directly for this study, buttaken from Bull and Payne (1982). It is possible, then, that someof these specimens might have been classified as in early fusionrather than fusing when following the protocols of this study.Moreover, since Bull and Payne did not distinguish between thefusion of the trochlea and the medial epicondyle of the distalhumerus, the Bull and Payne scores were assigned only to thedistal trochlea. Our impression from the specimens in our study,and from our analysis of archaeological collections, is that theclosure of the medial epicondyle lags behind the fusion of thedistal trochlea, with closure of this fusion plate possibly occur-ring in age class F.Age Class F: Fusion of the first phalanx and the distal tibiacoincide and form the basis for age class F. While it is possiblethat the first phalanx may fuse a bit earlier than the distal tibia,the timing of the fusion of these two elements seems quitedistinct from that of the earlier fusing second phalanx and distaltrochlea of the humerus and the fusion of the elements includedin age class G. As with the distal humerus, specimens from thePayne collection with fusing or fully fused distal tibia were notobserved directly for this study and their fusion scores weretaken from Bull and Payne.
Age Class G: The next bones to fuse are themetapodials, with thetiming of the fusion of all four metacarpals and metatarsalscoinciding with one another. The fusion of the distal fibula oc-curs about this time, though the timing of fusion for thiselement may be a bit later than in the metapodials.Age Class H: The later fusing elements show more overlap inthe timing of their fusion. However, the specimens examinedhere suggest that the apex of the calcaneum and both the greattrochanter and head of the proximal femur fuse slightly laterthan the elements in age class G and earlier than those in thefinal age class H. Of the components of the proximal femur, thegreat trochanter seems to fuse somewhat earlier than thehead.Age Class I: This large group of post-cranial elements includesthe posterior end of the axis, the distal radius, the proximal anddistal ulna, the proximal tibia and the tibial tubercle, the prox-imal fibula, and both the great trochanter and head of theproximal humerus. Once again, scores for the fusion of theproximal tibia and humerus in specimens from the Paynecollection were taken from Bull and Payne. In addition, since allbut the fusion of the atlas and axis of specimen SP350 werederived from this earlier study, it cannot be certain whetherelements recorded as fully fused (4) might have been scored asearly fusion (3) in this study, and, especially, if the proximal tibiascored as fusing (2) by Bull and Paynemight have been judged tobe in early fusion following the protocols adopted in this study.Age Class J: Age class J is reserved for specimens where all ar-ticulations are fully fused and are, therefore, older than speci-mens in age class I. The radius and ulna in these specimens,however, remain unfused.Age Class K: Fusion of the radius to the ulna, especially in thedistal portions of these bones was observed in four of the FMNHspecimens. The fusion of these two elements, then, defines thisfinal age class. It is possible that this final fusion stage is more ofan idiopathic feature in some older individuals rather than adistinct and final fusion stage.
M.A. Zeder et al. / Journal of Archaeological Science 55 (2015) 135e150138
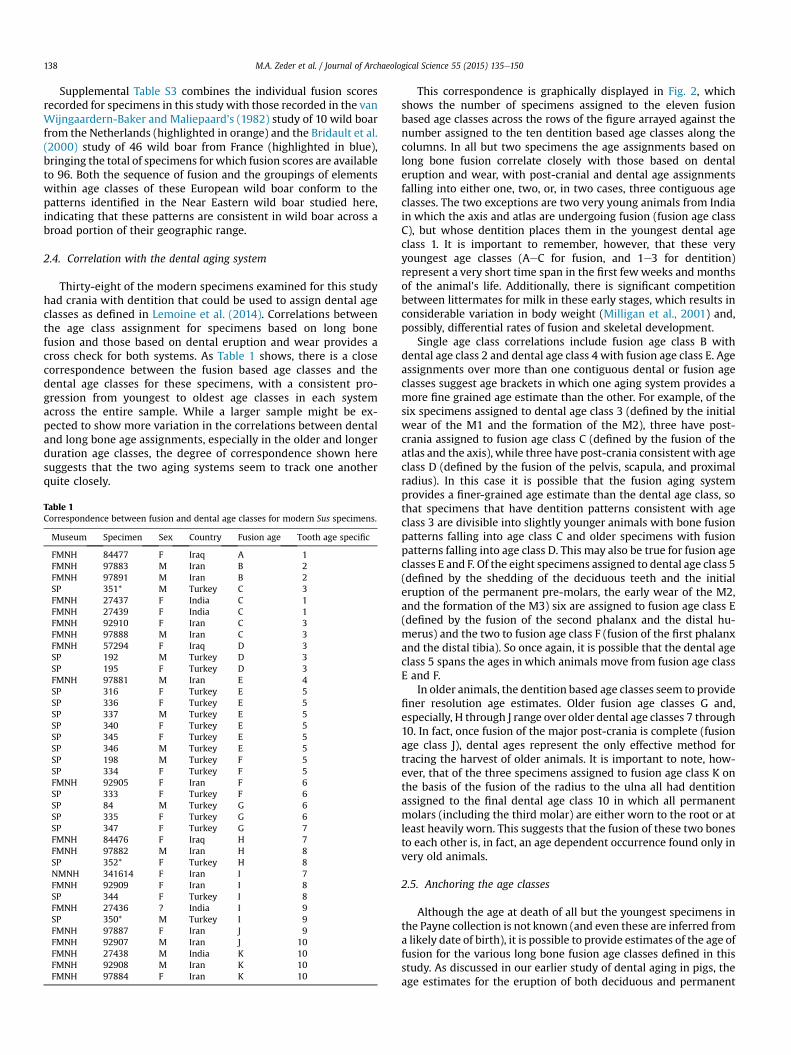
Supplemental Table S3 combines the individual fusion scoresrecorded for specimens in this study with those recorded in the vanWijngaardern-Baker and Maliepaard's (1982) study of 10 wild boarfrom the Netherlands (highlighted in orange) and the Bridault et al.(2000) study of 46 wild boar from France (highlighted in blue),bringing the total of specimens for which fusion scores are availableto 96. Both the sequence of fusion and the groupings of elementswithin age classes of these European wild boar conform to thepatterns identified in the Near Eastern wild boar studied here,indicating that these patterns are consistent in wild boar across abroad portion of their geographic range.
2.4. Correlation with the dental aging system
Thirty-eight of the modern specimens examined for this studyhad crania with dentition that could be used to assign dental ageclasses as defined in Lemoine et al. (2014). Correlations betweenthe age class assignment for specimens based on long bonefusion and those based on dental eruption and wear provides across check for both systems. As Table 1 shows, there is a closecorrespondence between the fusion based age classes and thedental age classes for these specimens, with a consistent pro-gression from youngest to oldest age classes in each systemacross the entire sample. While a larger sample might be ex-pected to show more variation in the correlations between dentaland long bone age assignments, especially in the older and longerduration age classes, the degree of correspondence shown heresuggests that the two aging systems seem to track one anotherquite closely.
Table 1Correspondence between fusion and dental age classes for modern Sus specimens.
Museum Specimen Sex Country Fusion age Tooth age specific
FMNH 84477 F Iraq A 1FMNH 97883 M Iran B 2FMNH 97891 M Iran B 2SP 351* M Turkey C 3FMNH 27437 F India C 1FMNH 27439 F India C 1FMNH 92910 F Iran C 3FMNH 97888 M Iran C 3FMNH 57294 F Iraq D 3SP 192 M Turkey D 3SP 195 F Turkey D 3FMNH 97881 M Iran E 4SP 316 F Turkey E 5SP 336 F Turkey E 5SP 337 M Turkey E 5SP 340 F Turkey E 5SP 345 F Turkey E 5SP 346 M Turkey E 5SP 198 M Turkey F 5SP 334 F Turkey F 5FMNH 92905 F Iran F 6SP 333 F Turkey F 6SP 84 M Turkey G 6SP 335 F Turkey G 6SP 347 F Turkey G 7FMNH 84476 F Iraq H 7FMNH 97882 M Iran H 8SP 352* F Turkey H 8NMNH 341614 F Iran I 7FMNH 92909 F Iran I 8SP 344 F Turkey I 8FMNH 27436 ? India I 9SP 350* M Turkey I 9FMNH 97887 F Iran J 9FMNH 92907 M Iran J 10FMNH 27438 M India K 10FMNH 92908 M Iran K 10FMNH 97884 F Iran K 10
This correspondence is graphically displayed in Fig. 2, whichshows the number of specimens assigned to the eleven fusionbased age classes across the rows of the figure arrayed against thenumber assigned to the ten dentition based age classes along thecolumns. In all but two specimens the age assignments based onlong bone fusion correlate closely with those based on dentaleruption and wear, with post-cranial and dental age assignmentsfalling into either one, two, or, in two cases, three contiguous ageclasses. The two exceptions are two very young animals from Indiain which the axis and atlas are undergoing fusion (fusion age classC), but whose dentition places them in the youngest dental ageclass 1. It is important to remember, however, that these veryyoungest age classes (AeC for fusion, and 1e3 for dentition)represent a very short time span in the first few weeks and monthsof the animal's life. Additionally, there is significant competitionbetween littermates for milk in these early stages, which results inconsiderable variation in body weight (Milligan et al., 2001) and,possibly, differential rates of fusion and skeletal development.
Single age class correlations include fusion age class B withdental age class 2 and dental age class 4 with fusion age class E. Ageassignments over more than one contiguous dental or fusion ageclasses suggest age brackets in which one aging system provides amore fine grained age estimate than the other. For example, of thesix specimens assigned to dental age class 3 (defined by the initialwear of the M1 and the formation of the M2), three have post-crania assigned to fusion age class C (defined by the fusion of theatlas and the axis), while three have post-crania consistent with ageclass D (defined by the fusion of the pelvis, scapula, and proximalradius). In this case it is possible that the fusion aging systemprovides a finer-grained age estimate than the dental age class, sothat specimens that have dentition patterns consistent with ageclass 3 are divisible into slightly younger animals with bone fusionpatterns falling into age class C and older specimens with fusionpatterns falling into age class D. This may also be true for fusion ageclasses E and F. Of the eight specimens assigned to dental age class 5(defined by the shedding of the deciduous teeth and the initialeruption of the permanent pre-molars, the early wear of the M2,and the formation of the M3) six are assigned to fusion age class E(defined by the fusion of the second phalanx and the distal hu-merus) and the two to fusion age class F (fusion of the first phalanxand the distal tibia). So once again, it is possible that the dental ageclass 5 spans the ages in which animals move from fusion age classE and F.
In older animals, the dentition based age classes seem to providefiner resolution age estimates. Older fusion age classes G and,especially, H through J range over older dental age classes 7 through10. In fact, once fusion of the major post-crania is complete (fusionage class J), dental ages represent the only effective method fortracing the harvest of older animals. It is important to note, how-ever, that of the three specimens assigned to fusion age class K onthe basis of the fusion of the radius to the ulna all had dentitionassigned to the final dental age class 10 in which all permanentmolars (including the third molar) are either worn to the root or atleast heavily worn. This suggests that the fusion of these two bonesto each other is, in fact, an age dependent occurrence found only invery old animals.
2.5. Anchoring the age classes
Although the age at death of all but the youngest specimens inthe Payne collection is not known (and even these are inferred froma likely date of birth), it is possible to provide estimates of the age offusion for the various long bone fusion age classes defined in thisstudy. As discussed in our earlier study of dental aging in pigs, theage estimates for the eruption of both deciduous and permanent

Fig. 2. Correlation between dental and fusion age classes shown as numbers of specimens.
M.A. Zeder et al. / Journal of Archaeological Science 55 (2015) 135e150 139
teeth are relatively secure. There is close agreement in studies ofthe eruption of deciduous teeth in two studies of wild captive pigsof known age by and Matschke (1967) and Anezaki (2009). More-over, an assessment of a wide range of sources citing eruption agesin pigs by Legge (nd) shows close agreement in the timing oferuption of permanent molars, especially the M1 and M2, in wild,feral, and domestic pigse calling into question the assumption thatthe timing of eruption varies in domestic and wild pigs. In ourearlier study this consistency allowed us to fix age estimates to thedentition based age classes that were defined primarily by theeruption of various deciduous and permanent teeth (dental ageclasses 1e6 in our system). Determining the age of older andbroader dental age classes defined primarily by tooth wear is lesssecure, but the correlation between the wear patterns of two veryold boars of known age from the Anezaki study with our dental ageclass 9 allowed us to anchor this penultimate stage to an age esti-mate of the order of 96 months. With this anchor we could thenprovide approximate estimates of the age ranges of age classes 7and 8 in our system, with an age estimate of greater than 96months (8 years) for our final age class 10, which is close to themaximum life span of wild boars of 9e10 years (De Magalh~aes andCosta, 2009; Jazierski, 1977).
Given the strong correlation between the dental age classesdefined in our earlier study and the fusion age classes definedhere, it is possible to assign age estimates to the fusion based ageclasses. Column two of Table 2 lists the ten dental age stages andtheir age estimates from Lemoine et al. (2014). The last column ofthe table lists the corresponding fusion age classes based on thecorrelation between the dental age and fusion age assignmentsof the specimens included in this study. Table 3 presents theeleven fusion age classes defined here, the elements whosefusion delimits each age class, the corresponding dental ageclasses, and an age estimate for each fusion age class extrapo-lated from the dental age classes estimates. Estimates for these
Table 2Comparison of dental and fusion age classes.
Dental age class specific Dental age estimate Fusion age class
1 �1 mo A2 3e5 mos B3 6e8 mos C,D4 8e12 mos E5 12e18 mos E,F6 18e30 mos F,G7 30e52 mos G,H,I8 52e72 mos H,I9 72e96 mos I,J10 >96 mos J,K
older age classes are likely to be less secure given the likelyhigher degree of variation in tooth wear on which the dentalages classes are based.
Based on the correlation of two specimens with dental age class1, age class A is assigned an age of equal to or less than 1 month, anassignment consistent with the neo-natal condition of the skele-tons of these specimens. Age class B which includes slightly olderspecimens that still show no sign of post-cranial bone fusion, andwhich correlates exclusively with dental age class 2, is assigned anage estimate of 3e5 months. Specimens assigned to age class C(fusion of atlas and axis) and age class D (fusion of the pelvis,scapula, and proximal radius) have dentition assigned to dental ageclass 3. In this case the 6e8 month age estimate for this dental ageclass is split between the two fusion age classes with age class Cassigned an age estimate of 6e7 months and age class D estimatedat between 7 and 8 months.
A similar approach is used in making age estimates for otherfusion age classes. Thus age class E (second phalanx and distalhumerus) that overlaps with dental ages 4 and 5 receives an ageestimate of 8e18 months. The age estimate for age class F (firstphalanx and distal tibia) that overlaps with dental ages 5 and 6 isset at 18e24 months, while an estimate of 24e36 months is set forage class G (distal metapodials and distal fibula). Age class H(calcaneum and proximal femur), which overlaps with dental ageclasses 7 and 8, receives an age estimate of 36e48 months.
The broader overlap of I, the final age class for epiphyseal fusion(distal radius, distal femur, proximal tibia, distal and proximal ulna,and proximalfibula), with age classes 7 through 9 is trimmed a bit toan estimate of 48e60 months. Age class J in which all epiphyses arefused but the radius and ulna remain separate is estimated to span60e96 months, while the correlation of specimens in fusion ageclass K in which the radius and the ulna fuse together with dentalage class 10 results in an age estimate of greater than 96 months.
Table 3Fusion age estimates extrapolated from dental ages.
Fusionage class
Bones fusing Dental ageclass specific
Fusion age estimatefrom dental age
A Neo-nate 1 �1 moB None 2 3e5 mosC Atlas & Axis 3 6e7 mosD Pelvis, Scapula, P. Radius 3 7e8 mosE 2 Phalanx, D. Humerus 4e5 8e18 mosF 1 Phalanx, D. Tibia 5e6 18e24 mosG D. Metapodials, D. Fibula 6e7 24e36 mosH Calcaneum, P. Femur 7e8 36e48 mosI D. Radius, D. Femur, P. Tibia, D
& P. Ulna, P. Fibula, P. Humerus7e9 48e60 mos
J All fused 9e10 60e96 mosK Radius & Ulna 10 >96 mos

M.A. Zeder et al. / Journal of Archaeological Science 55 (2015) 135e150140
2.6. Comparison with other long bone fusion systems
The fusion aging system proposed here is compared to otherfusion based aging systems in Table 4. The first three columns ofTable 4 list the post-cranial elements in the order of their fusion asobserved in the 40 wild boar specimens included in this study (andconfirmed by the fusion patterns of the 56 additional specimensfrom other studies, see Table S3), the corresponding age class asdefined here, and the age estimate as extrapolated from dental ageestimates. The next columns contain the age estimates for eachelement used in eight different fusion-aging schemes. The firstseven of these are systems that were devised independently of oneanother. The Bull and Payne system, as stated earlier, is based onmodern wild boar from Turkey of unknown but estimated age,specimens that are also included in the sample studied here. Lesbre(1897/8) is a very early study of long bone fusion in domestic ani-mals. Silver (1969) is a well known source for data on dental andlong bone fusion aging widely used by archaeozoologists tocompute age profiles for both domestic animals and their wildprogenitors. It is, however, perhaps the least reliable source forthese data as both the sequences and timing of element fusion (andtooth eruption) cited here are based on an amalgam of largelyunreferenced sources (Legge, nd). The Habermehl (1975) age esti-mates are similarly drawn from multiple, but better referenced,studies of skeletal fusion in domestic livestock. The Reiland (1978)age estimates are based on a radiometric study of a large number ofdomestic Swedish Landrace and Yorkshire breeds of pigs. The vanWijngaardern-Baker and Maliepaard (1982) age estimates, asnoted above, are derived from their study of wild boar from threedifferent repositories in the Netherlands, based on correlationswith specimen dental ages following Habermehl (1975). TheBridault et al. (2000) estimates are based on a study of a largenumber of wild boar from a single population living in northernFrance, with correlating dental ages followingMatschke (1967). TheHongo and Meadow (1998) system is not based on an independentstudy, but is instead a compilation drawing from Bull and Payne,Silver, Habermehl, and B€ok€onyi (1972).
Table 4Comparison of Zeder and Lemoine age estimates to other systems.
Elementsfusing
Zeder &lemoineage class
Zeder,lemoine,& Payne
Bull andPayne 1982.
Lesbre1897/8.
Silver1969.
Neo-natal A �1 mo e e e
Young B 3e5 mos e e e
Axis C 6e7 mos e e e
Atlas C 6e7 mos e e e
Pelvis D 7e8 mos 7e11 mos 12 mos 12 mosScapula D 7e8 mos 7e11 mos 12 mos 12 mosP. Radius D 7e8 mos 19e23 mos 12 mos 12 mos2 Phalanx E 8e18 mos 19e23 mos 12 mos 12 mosD. Humerus E 8e18 mos 19e23 mos 12 mos 12 mos1 Phalanx F 18e24 mos 19e23 mos 24 mos 24 mosD. Tibia F 18e24 mos 19e23 mos 24 mos 24 mosMetatarsal G 24e36 mos 31e35 mos 24 mos 30 mosMetacarpal G 24e36 mos 31e35 mos 24 mos 24 mosD. Fibula G 24e36 mos 31e35 mos 24e30 mos 30 mosCalcaneum H 36e48 mos >35 mos 24e30 mos 27 mosP. Femur H 36e48 mos 31e35 mos 36e42 mos 42 mosD. Radius I 48e60 mos >35 mos 42 mos 42 mosP. Ulna I 48e60 mos >35 mos 36 mos 36e42 mosD. Femur I 48e60 mos >35 mos 42 mos 42 mosP. Tibia I 48e60 mos >35 mos 42 mos 42 mosD. Ulna I 48e60 mos >35 mos 42 mos 36e42 mosP. Fibula I 48e60 mos >35 mos 42 mos 42 mosP. Humerus I 48e60 mos >35 mos 42 mos 42 mosRadius & Ulna K >96 mos e e e
The correspondence among the various aging schemes isreasonably close in most cases, especially considering vagaries ofsample size, inter-observer variability, and the shortage of securelyaged specimens. In all of the younger age classes for which there arecorollaries in other systems (age classes CeH) our estimates eitheroverlap with or closely abut estimates of other schemes. The majorexception is the aging scheme proposed by Reiland, in which age offusion estimates are consistently younger than other sources. It ishard to know if this outlier system is indicative of an acceleratedpace of bone fusion in the breeds used in this study or simplyrepresents unreliable data. Reiland's observations of bone fusionand tooth eruption were based on radiographic images, which mayalso account for these inconsistencies.
Our estimates for the final age classes in our system e ageclasses IeK, vary more significantly from other schemes. Our esti-mate of 48e60months for the fusion of the large group of elementsin age class I, in particular, is both somewhat older and broader thanestimates of most other systems, which variably put the fusion ofthese bones at either greater than 35 months, 36 months, 42months, or between 36 and 42 months. We believe, however, thatour estimate is justified on two grounds. First the pattern of earlierfusion for the calcaneum and the proximal femur (an elementusually included in other systems with this group of late fusingbones) suggests that there is an additional age class between thefusion of elements that most systems put between 24 and 30months and those that fuse after 30 months of age. The later andbroader age assignment for these final fusing elements is alsoconsistent with the correlation between specimens assigned to thisfusion age class and their corresponding dental age class assign-ments. With the exception of one borderline specimen (NMNH341614, ID # 22 in Fig. 1), two of the remaining four fusion age classI specimens are assigned to dental age class 8 (52e72 months) andtwo to dental age class 9 (72e96 months as anchored by Anezaki,2009). Extending this range up to 96 months (the maximum ageestimate for dentition age class 9) is far too broad, but it seemsreasonable that the time span over which this large group of ele-ments fuses is broader than the six-month 36e42 month age
Habermehl1975.
Reiland1978.
Van Wijgaarden-Bakkerand Maliepaard 1982.
Bridaultet al.,2000.
Hongo andMeadow1998.
e e e e e
e e e e e
e e e e e
e e e 7e13 mos e
12 mos e 12e19 mos 9e13 mos 12 mos12 mos e 12e19 mos 7e13 mos 12 mos12 mos 10 mos 12e19 mos 9e13 mos 12 mos12 mos <4 mos 19e22 mos 9e18 mos 12 mos12 mos 10 mos 12e19 mos 9e21 mos 12 mos24 mos <4 mos 19e22 mos 17e21 mos 24e30 mos24 mos 12 mos 22e31 mos 17e25 mos 24e30 mos24 mos 12e14 mos 22e31 mos 21e? mos 24e30 mos24 mos 12e14 mos 22e31 mos 21e? mos 24e30 mos24e30 mos e 22e31 mos 25e? mos 24e30 mos24e30 mos e 31e43 mos 26e? mos 24e30 mos42 mos 18 mos 31e43 mos 26e? mos 36e42 mos42 mos 20 mos >43 mos e 36e42 mos36 mos e 31e43 mos e 36e42 mos42 mos e 31e43 mos e 36e42 mos42 mos 20 mos 31e43 mos e 36e42 mos42 mos 20 mos >43 mos e 36e42 mos42 mos 18 mos 31e43 mos e e
42 mos 18 mos ca. 43 mos e 36e42 mose e e e e

M.A. Zeder et al. / Journal of Archaeological Science 55 (2015) 135e150 141
estimate of other systems. Expanding the range for fusion of thislarge group of bones to 12 months, from 48 e 60 months (4e5years) allows for a more staggered period of fusion of these ele-ments and accommodates both the fusion sequence patternsobserved here and the correlation between fusion and dentalattrition in the specimens in our sample.
Our age class J for specimens in which all epiphyses are fullyfused but the radius and ulna remain separate has no corollary inother studies, other than to be indicative of animals older than theage of fusion of the last fusing bones (in other systems variably putat greater than 35 months to greater than 42 months, and here atbetween 48 and 60 months). Nor is this age class very useful inconstructing age profiles for archaeological assemblages, unlessapplied to a complete skeleton of an animal that meets the con-ditions for inclusion in this age class (i.e. with complete fusion of allpost-cranial elements). But an age estimate of between 60 and 96months for these specimens seems reasonable, especially giventheir correlationwith dentition assigned to the final two dental ageclasses 9 and 10. The final age class K, defined by the fusion of theradius to the ulna and set here at older than 96 months, also has nocorollary in other systems and may be of limited utility in archae-ological assemblages. However, as wewill see below, it can be usedas a final anchoring point for these profiles in assemblages thatcontain the bones of very old animals in which these two boneshave fused together.
3. Application
3.1. The archaeological sample
In Lemoine et al. (2014) we demonstrated the efficacy of theproposed dental aging system by applying the system to three NearEastern assemblages where pig exploitation patterns are expectedto have varied. This comparison allowed us to assess the ability ofthe proposed dental aging system to detect differences in pigexploitation in three very different contexts: a Late Epipaleolithiccommunity of foragers on the threshold of initial animal manage-ment, a small Halafian agricultural village in which pigs were likelymanaged in a generalized way, and Bronze Age urban site wherethere was evidence for a highly specialized agricultural economythat included a possible internal meat producing system involvingpigs.
In this paper, we focus on one of these sites, Hallan Çemi, a smallsite in southeastern Anatolia that was occupied for a brief period oftime between circa 11,700e11,500 cal. BP (Starkovich and Stiner,2009). One of the earliest such communities in the easternTaurus/Zagros Arc (Peasnall, 2000) there is both botanical andfaunal evidence that the site was occupied year round by a smallcommunity of foragers (Rosenberg et al., 1998). Based on an earlierstudy of a portion of the faunal remains from the site, Redding andRosenberg argued that a kind of incipient pig management wasbeing practiced (Redding, 2005; Redding and Rosenberg, 1998,2000; Rosenberg et al., 1998).
Salvage excavations conducted in the early 1990s recovered anastonishing 2 metric tons of animal bones (Redding and Rosenberg,2000; Redding, personal communication), including an extraordi-narily large assemblage of Sus remains (c. 9000 NISP in Zeder andLemoine's recent analysis of the Hallan Çemi assemblage). In addi-tion to its very large assemblage of Sus remains, therewas also bettercontrol over recovery techniques at this site than at the other twosites featured in our earlier study. Although conducted as a rescueprogram in advance of flooding from dam construction, excavationsat the site placed a strong emphasis on the recovery of faunal re-mains. In addition to rigorous protocols for hand collection of faunalremains; 25% of all deposits at the sitewere dry-screened through a
5 mmmesh (Peasnall, 2000; Rosenberg et al., 1998), ensuring ahigher recovery of unfused epiphyses that are more likely to beoverlooked under less intensive recovery protocols. Although notpossible with this sample, ideally data from the screened and un-screened samples should be compared to check and, if necessary,correct for recovery loss in the unscreened part of the sample. Webelieve, however, that recovery was good enough that the datapresented here have not been compromised by partial screening.Moreover, the tight temporal frameworkof the site and its small size(estimated as less than 0.5 ha and with no more than 10e15 smallstructures at any one time) makes it unlikely that there were majordifferences in pig exploitation practices either over time or indifferent sectors at the site, reducing the confounding effect of suchdifferences in the interpretation of demographic profiles con-structed on the total assemblage of pig remains recovered from thesite. The preservation of animal bone at Hallan Çemi is extraordi-nary. There are appear to be no dogs in the assemblage and the lackof carnivore gnawing on the bones confirms that this commonsource of bias in later archaeozoological assemblages from the re-gion is not an issue here. Moreover, while researchers using some-what different recording protocols analyzed the other twoassemblages included in the earlier study, the analysis of fusion anddental eruption and wear patterns of the Sus remains from HallanÇemi were recorded by Zeder and Lemoine using the protocolsdeveloped here as part of an ongoing study of this large assemblage,assuring greater consistency in analytical practices.
The goal of this application is not to evaluate earlier claims forinitial pig management at the site. This will be the focus of a laterpaper. Instead, this application will look first for internal consis-tencies in the fusion patterns of elements, both in terms of theproposed sequence of fusion and the grouping of elements intolarger age classes. Next, we will compare the age profiles based onlong bone fusion of pig post-crania from the site to the early con-structed dental age profiles presented in Lemoine et al. to assess thecorrespondence between these two methods for constructing pigage profiles.
3.2. The process
Computation of long bone fusion scores follows the protocoldescribed in Zeder, 1991, which is itself derived from Redding, 1981.This protocol involves summing the number of unfused (u), fusing(g), and fused (f) elements for each of post-cranial bones includedin the system. These sums are computed using simple NISP for eachelement with some correction to assure that fragments clearlycoming from the same bone are not counted separately. Someprotocols for computing fusion based age profiles use a correctedmeasure of quantity (MNI or a more strictly applied MNE) incomputing fusion scores to minimize the risk of double countingelements from the same individual (Stiner et al., 2014). Here wetook care to assure that shafts and epiphyses in a single prove-nience unit that were scored for fusion represented different ele-ments, employing a kind of on-the-fly MNE adjustment tominimize the possibility of double counting the same element.Effort was also made to refit epiphyses to diaphyses and to makesure that fragments of either epiphyses or diaphyses were clearlyfrom different elements before they were scored for fusion. Thisapproach was facilitated by the excellent preservation and gener-ally large fragment size of the Hallan Çemi assemblage that made iteasy to score elements as MNEs. Some researchers may wish toadopt stricter protocols to avoid the possibility of double countingof unfused shafts and epiphyses or over counting unfused elementsat the expense of fused ones, especially where excavation units aresmall and highly fragmented (i.e. by counting only epiphyses ordiaphyses or employing some kind of weighting measure).
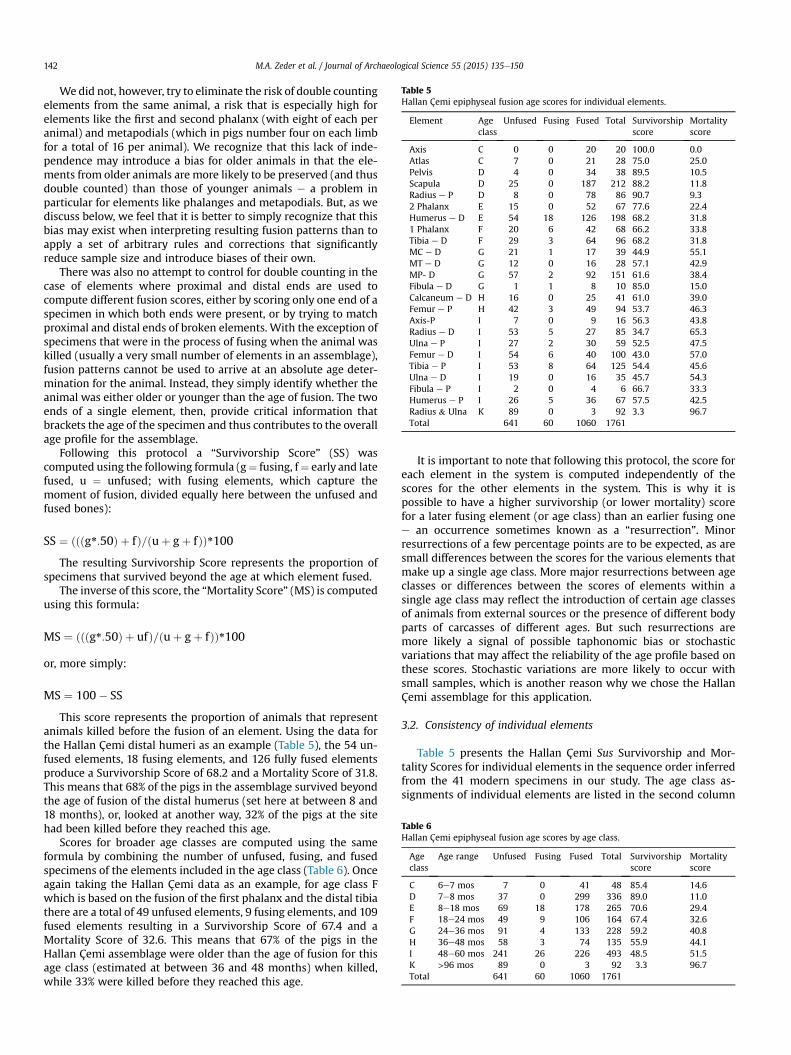
Table 5Hallan Çemi epiphyseal fusion age scores for individual elements.
Element Ageclass
Unfused Fusing Fused Total Survivorshipscore
Mortalityscore
Axis C 0 0 20 20 100.0 0.0Atlas C 7 0 21 28 75.0 25.0Pelvis D 4 0 34 38 89.5 10.5Scapula D 25 0 187 212 88.2 11.8Radius e P D 8 0 78 86 90.7 9.32 Phalanx E 15 0 52 67 77.6 22.4Humerus e D E 54 18 126 198 68.2 31.81 Phalanx F 20 6 42 68 66.2 33.8Tibia e D F 29 3 64 96 68.2 31.8MC e D G 21 1 17 39 44.9 55.1MT e D G 12 0 16 28 57.1 42.9MP- D G 57 2 92 151 61.6 38.4Fibula e D G 1 1 8 10 85.0 15.0Calcaneum e D H 16 0 25 41 61.0 39.0Femur e P H 42 3 49 94 53.7 46.3Axis-P I 7 0 9 16 56.3 43.8Radius e D I 53 5 27 85 34.7 65.3Ulna e P I 27 2 30 59 52.5 47.5Femur e D I 54 6 40 100 43.0 57.0Tibia e P I 53 8 64 125 54.4 45.6Ulna e D I 19 0 16 35 45.7 54.3Fibula e P I 2 0 4 6 66.7 33.3Humerus e P I 26 5 36 67 57.5 42.5Radius & Ulna K 89 0 3 92 3.3 96.7Total 641 60 1060 1761
Table 6Hallan Çemi epiphyseal fusion age scores by age class.
Ageclass
Age range Unfused Fusing Fused Total Survivorshipscore
Mortalityscore
C 6e7 mos 7 0 41 48 85.4 14.6D 7e8 mos 37 0 299 336 89.0 11.0E 8e18 mos 69 18 178 265 70.6 29.4F 18e24 mos 49 9 106 164 67.4 32.6G 24e36 mos 91 4 133 228 59.2 40.8H 36e48 mos 58 3 74 135 55.9 44.1I 48e60 mos 241 26 226 493 48.5 51.5K >96 mos 89 0 3 92 3.3 96.7Total 641 60 1060 1761
M.A. Zeder et al. / Journal of Archaeological Science 55 (2015) 135e150142
Wedid not, however, try to eliminate the risk of double countingelements from the same animal, a risk that is especially high forelements like the first and second phalanx (with eight of each peranimal) and metapodials (which in pigs number four on each limbfor a total of 16 per animal). We recognize that this lack of inde-pendence may introduce a bias for older animals in that the ele-ments from older animals are more likely to be preserved (and thusdouble counted) than those of younger animals e a problem inparticular for elements like phalanges and metapodials. But, as wediscuss below, we feel that it is better to simply recognize that thisbias may exist when interpreting resulting fusion patterns than toapply a set of arbitrary rules and corrections that significantlyreduce sample size and introduce biases of their own.
There was also no attempt to control for double counting in thecase of elements where proximal and distal ends are used tocompute different fusion scores, either by scoring only one end of aspecimen in which both ends were present, or by trying to matchproximal and distal ends of broken elements. With the exception ofspecimens that were in the process of fusing when the animal waskilled (usually a very small number of elements in an assemblage),fusion patterns cannot be used to arrive at an absolute age deter-mination for the animal. Instead, they simply identify whether theanimal was either older or younger than the age of fusion. The twoends of a single element, then, provide critical information thatbrackets the age of the specimen and thus contributes to the overallage profile for the assemblage.
Following this protocol a “Survivorship Score” (SS) wascomputed using the following formula (g¼ fusing, f¼ early and latefused, u ¼ unfused; with fusing elements, which capture themoment of fusion, divided equally here between the unfused andfused bones):
SS ¼ ðððg*:50Þ þ fÞ=ðuþ gþ fÞÞ*100The resulting Survivorship Score represents the proportion of
specimens that survived beyond the age at which element fused.The inverse of this score, the “Mortality Score” (MS) is computed
using this formula:
MS ¼ ðððg*:50Þ þ ufÞ=ðuþ gþ fÞÞ*100
or, more simply:
MS ¼ 100� SS
This score represents the proportion of animals that representanimals killed before the fusion of an element. Using the data forthe Hallan Çemi distal humeri as an example (Table 5), the 54 un-fused elements, 18 fusing elements, and 126 fully fused elementsproduce a Survivorship Score of 68.2 and a Mortality Score of 31.8.This means that 68% of the pigs in the assemblage survived beyondthe age of fusion of the distal humerus (set here at between 8 and18 months), or, looked at another way, 32% of the pigs at the sitehad been killed before they reached this age.
Scores for broader age classes are computed using the sameformula by combining the number of unfused, fusing, and fusedspecimens of the elements included in the age class (Table 6). Onceagain taking the Hallan Çemi data as an example, for age class Fwhich is based on the fusion of the first phalanx and the distal tibiathere are a total of 49 unfused elements, 9 fusing elements, and 109fused elements resulting in a Survivorship Score of 67.4 and aMortality Score of 32.6. This means that 67% of the pigs in theHallan Çemi assemblage were older than the age of fusion for thisage class (estimated at between 36 and 48 months) when killed,while 33% were killed before they reached this age.
It is important to note that following this protocol, the score foreach element in the system is computed independently of thescores for the other elements in the system. This is why it ispossible to have a higher survivorship (or lower mortality) scorefor a later fusing element (or age class) than an earlier fusing onee an occurrence sometimes known as a “resurrection”. Minorresurrections of a few percentage points are to be expected, as aresmall differences between the scores for the various elements thatmake up a single age class. More major resurrections between ageclasses or differences between the scores of elements within asingle age class may reflect the introduction of certain age classesof animals from external sources or the presence of different bodyparts of carcasses of different ages. But such resurrections aremore likely a signal of possible taphonomic bias or stochasticvariations that may affect the reliability of the age profile based onthese scores. Stochastic variations are more likely to occur withsmall samples, which is another reason why we chose the HallanÇemi assemblage for this application.
3.2. Consistency of individual elements
Table 5 presents the Hallan Çemi Sus Survivorship and Mor-tality Scores for individual elements in the sequence order inferredfrom the 41 modern specimens in our study. The age class as-signments of individual elements are listed in the second column

M.A. Zeder et al. / Journal of Archaeological Science 55 (2015) 135e150 143
of the table. The table also provides the counts of unfused, fusingand fused elements, plus the total number of elements used tocompute the individual scores for each element. Fig. 3 presents abar graph of the Survivorship Scores for each element with the ageclass grouping indicated by different color bars. This graph makesit easy to distinguish discrepancies in the scores of elementswithin the same age class and between elements assigned todifferent age classes. Since age classes A and B are reserved forvery young animals with no bones fused, scores cannot becomputed for these age classes and they are not shown in eitherTables 5 and 6 or Fig. 3. The same is true for age class I which isreserved for animals in which epiphyseal fusion is complete butthe radius and ulna remain separate.
Marked discrepancies in scores of both the distal and proximalfibulae for their respective age classes are almost certainly attrib-utable to small sample size for this slender and friable bone. Onlyten fibulae contributed to the scores for distal fibula fusion and sixfor computing the score for the proximal fusion. The large 25 pointdiscrepancy between the scores of the two elements in age class Ccomprised of the axis (SS¼ 100) and the atlas (SS¼ 75) may also beattributable to taphonomic issues; the bones of very young animalsin this age class (estimated here at between 6 and 7 months) maynot preserve well, with the denser atlas bone more likely to bepreserved in young animals than the smaller more fragile cranialportion of the axis. But the scores for these two elements may alsobe affected by slight above-mentioned differences in the timing offusion of these two bones. Thus while the resultant scores for theseelements may not be as reliable as those based on later fusingbones, it seems reasonable to keep both these elements in thecomputation of the age profile for the Hallan Çemi pigs.
Similarly, the nine point differential between the scores for thesecond phalanx and the distal humerus in age class E may reflectthe difference in the timing of the fusion of the distal articulation ofthe humerus and the closure of the fusion plate on the medialepicondyle discussed above. If, as suggested, the fusion of this partof the distal humerus is more closely aligned with the fusion of theelements included in the next age class, then the scores for theHallan Çemi distal humeri computed here (where fusion patternsfor these two parts of the distal epiphysis were not distinguishedfrom one another) might be expected to be closer to those of ele-ments in this next age class.
Fig. 3. Survivorship scores for individual elements in the Ha
The eight point difference between the scores for the calcaneumand the proximal femur that make up age class H may, in likefashion, be an artifact of the failure of our coding system todifferentiate between the trochanter and the head of the femurwhen recording proximal fusion for this bone. As above, ourmodern sample suggests that the head of the femur fuses some-what later than the trochanter e perhaps in line with the laterfusing elements in age class I. Since femur heads are denser andmore likely to be preserved than the trochanter, it is likely thatheads dominate in the elements that make up this score. Onceagain, future recording protocols should differentiate betweenthese two parts of the proximal femur when recording fusion data.
It is also interesting to note that the difference between thecalcaneum score and those of the most elements that comprise ageclass I, up to 26 points in the case of the distal radius, adds supportfor the decision to include this element in a younger age class thanthis latter group. Similarly, the agreement of the proximal radiuswith the other two elements in this group and the 13 point dif-ference between the scores for the proximal radius and the ele-ments in the next age class lends additional support for theinclusion of this element within age class D, rather than with thenext group of fusing bones (age class E) as is done in some agingsystem.
Another anomaly among the elements thatmake up age class I isthe distal radius in which the large number of unfused distal endsresults in scores that indicate much lower survivorship, and highmortality, than the other elements that make up this age class. Thedistal radius is a hardy, dense element with a higher likelihood ofpreservation and recovery than most other elements in its group(i.e. the proximal tibia, proximal ulna, and the proximal humerus).Keeping this element in the sample of elements that comprise thisgroup, despite this anomalous score, takes advantage of this higherrecovery potential and combats the bias for the preservation ofolder animals with fully fused epiphyses that may be especially feltamong the more friable bones in this age class.
Differences between the scores of metacarpals, metatarsals, andundifferentiated metapodials also merit some discussion. Meta-carpals and metatarsals in the sample are bones which could beidentified to one of the four metacarpals or metatarsals (bones withtheir proximal ends intact) and that had enough of their distal endspresent to be able to gage the state of fusion. The metapodials
llan Çemi assemblage in order of fusion and age classes.

Table 7Hallan Çemi epiphyseal fusion age scores without fibulae.
Ageclass
Age range Unfused Fusing Fused Total Survivorshipscore
Mortalityscore
C 6e7 mos 7 0 41 48 85.4 14.6D 7e8 mos 37 0 299 336 89.0 11.0E 8e18 mos 69 18 178 265 70.6 29.4F 18e24 mos 49 9 106 164 67.4 32.6G 24e36 mos 90 3 125 218 58.0 42.0H 36e48 mos 58 3 74 135 55.9 44.1I 48e60 mos 239 26 222 487 48.3 51.7K >96 mos 88 0 3 91 3.3 96.7Total 633 60 1051 1744
Fig. 4. Survivorship scores for epiphyseal age classes in the Hallan Çemi assemblage.
M.A. Zeder et al. / Journal of Archaeological Science 55 (2015) 135e150144
represent elements without proximal ends that would enable fineridentification but with distal ends sufficiently intact to gage fusion.Specimens belonging to the smaller second and fifth metapodialscould be distinguished from the larger third and fourth meta-podials and were so identified in the analysis, but are combinedhere.
As above, no effort was made to try to control for doublecountingmetapodials from the same individual by focusing on onlyone of the many metapodials in the pig's skeleton (only counting3rd metacarpals for example). The break down of metapodials usedto construct the scores for this element adds support for this de-cision. The number of metacarpals and metatarsals that could beboth identified to number and assessed for distal fusion is small. Ofthe 117 metacarpals identified in our analysis, only 39 had intactenough distal portions to be used to compute a fusion score for thiselement. Focusing on only the most numerous metacarpal wouldhave been reduced our sample to three specimens, two if only oneside was used. Similarly, of the 100 metatarsals identified, only 28could be used to compute a fusion score. If only the most numerousside of the most numerous metatarsal was used to compute thisscore the sample size for this element would have been reduced tothree. In contrast, the majority of the metapodial sample comesfrom undifferentiated metapodials, with 151 of the total 192 met-apodials identified yielding information on distal fusion. Applyingstrict rules that prevent the eventuality of double counting bonesfrom a single animal in this case would have reduced our overallsample from a robust 218 bones to a small and unreliable sample ofthree specimens. And this is for the huge Hallan Çemi Sus assem-blage! It is far better, in our opinion, to accept the fact that therewill be some double counting in the construction of these fusionage profiles that may favor certain ages of animals over others, thanto adopt a more puritanical protocol that results in a pure but small,unrepresentative, and ultimately uninformative sample.
The scores for the metatarsals and the undifferentiated meta-podials are close, within 4 points of one another. The metacarpalsare 13e17 points different from those of the metatarsals and met-apodials suggesting a lower survivorship (and higher mortality) ofanimals at the age at which this bone fuses (set here at between 24and 36 months). Given the likelihood of higher survivorship ofmetapodials of older animals, it seems best to leave these elementsin the sample used to compute an overall score for this element.
Finally, it is hard to say how accurate the score for the final ageclass K, defined by the fusion of the radius and the ulna is. But itdoes provide a means of at least detecting the presence of very oldpigs in an assemblage. Scores for this age class in the Hallan Çemiassemblage indicate that only 3% of the pigs in the assemblagewerevery elderly, or, conversely, that 97% of the animals consumed byHallan Çemi residents had not reached the very advanced age atwhich this bone fuses.
3.3. Consistency of age classes
Table 7 presents the scores for element groups into age classesshown in Table 6 without the distal fibula in age class G and theproximal fibula in age class I, a correction that, given the smallnumber of fibulae in the assemblage, has little impact on theresultant scores. Fig. 4 shows these corrected survivorship scores inthe form of a bar graph. With the exception of the three-point‘resurrection’ between youngest age class C and D, there is asteady decrease in survivorship over the eight age classes. Using theage estimates discussed above, the fusion based ageprofile indicatesthat around 10% of the pigs consumed by Hallan Çemi residentswere younger than one year when theywere killed, with a 20% dropin survivorship between age classes D and E, from about 8 to 18months of age. This is followed by only relatively minor declines in
survivorship over the course of the next several age classes (FeI),with almost 50% of the pigs in the Hallan Çemi assemblage olderthan age class I (estimated here at between48 and60months)whenthey were killed. The low survivorship for the final age class K,reserved for very old animals greater than 96 months of age, sug-gests that 50% of the pigs consumed at Hallan Çemi were primeadult animals between the ages of about 4.5e8 years.
3.4. Comparability of fused based and dentition based age profiles
When attempting to compare dentition based and fusion basedage profiles, it is important to remember that there are funda-mental differences in how age is estimated in each system and howthe resultant age profile based on these estimates is computed. Asdiscussed above, fusion based age profiles are composed of anumber of independent computations that estimate the pro-portions of animals in an assemblage that were both younger thanand older than a certain age when they were killed. In contrast, agedeterminations made using dental eruption and wear patternsarrive at an estimate of the proportion of animals in an assemblagethat were killed at a certain age, providing an overview of the in-tensity of culling at each of the different age classes in the profile.The difference in the way in which dentition and fusion basedsystems reconstruct ages at death makes it inherently difficult todirectly compare age profiles produced by the two systems.Dentition age data can, however, be converted into more compa-rable survivorship profiles by sequentially subtracting the differentmortality estimates from one another e starting with subtractingthe youngest mortality score from 100% and then each successive
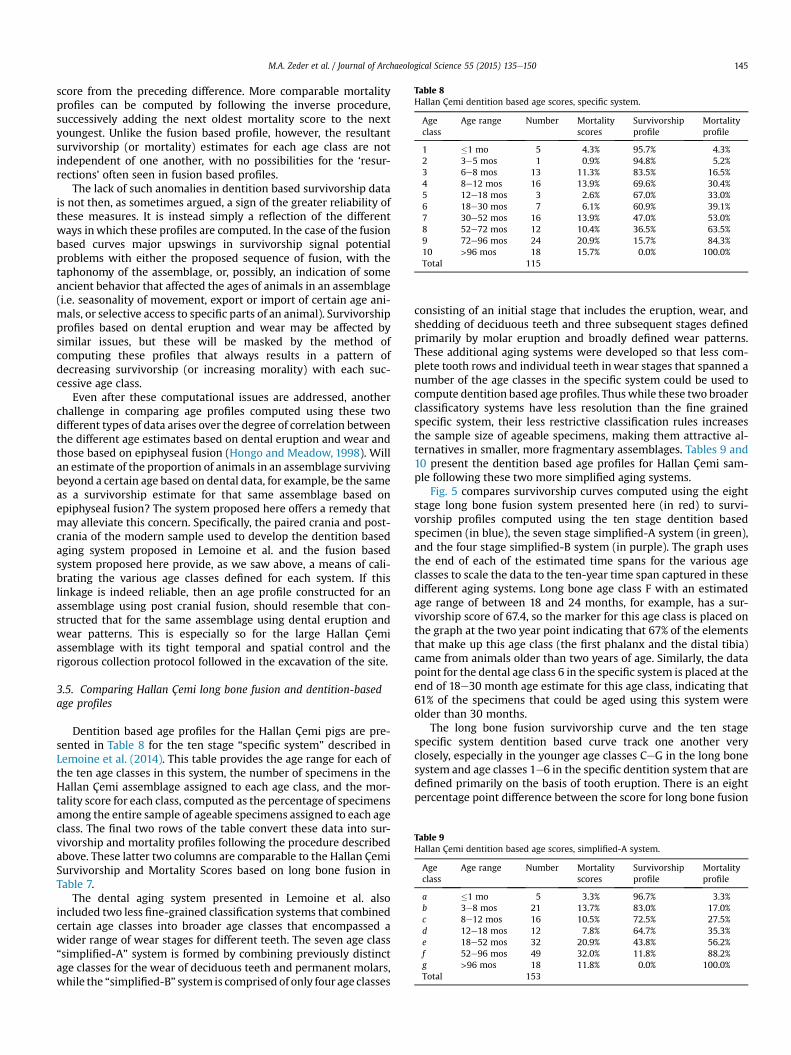
Table 8Hallan Çemi dentition based age scores, specific system.
Ageclass
Age range Number Mortalityscores
Survivorshipprofile
Mortalityprofile
1 �1 mo 5 4.3% 95.7% 4.3%2 3e5 mos 1 0.9% 94.8% 5.2%3 6e8 mos 13 11.3% 83.5% 16.5%4 8e12 mos 16 13.9% 69.6% 30.4%5 12e18 mos 3 2.6% 67.0% 33.0%6 18e30 mos 7 6.1% 60.9% 39.1%7 30e52 mos 16 13.9% 47.0% 53.0%8 52e72 mos 12 10.4% 36.5% 63.5%9 72e96 mos 24 20.9% 15.7% 84.3%10 >96 mos 18 15.7% 0.0% 100.0%Total 115
Table 9Hallan Çemi dentition based age scores, simplified-A system.
Ageclass
Age range Number Mortalityscores
Survivorshipprofile
Mortalityprofile
a �1 mo 5 3.3% 96.7% 3.3%b 3e8 mos 21 13.7% 83.0% 17.0%c 8e12 mos 16 10.5% 72.5% 27.5%d 12e18 mos 12 7.8% 64.7% 35.3%e 18e52 mos 32 20.9% 43.8% 56.2%f 52e96 mos 49 32.0% 11.8% 88.2%g >96 mos 18 11.8% 0.0% 100.0%Total 153
M.A. Zeder et al. / Journal of Archaeological Science 55 (2015) 135e150 145
score from the preceding difference. More comparable mortalityprofiles can be computed by following the inverse procedure,successively adding the next oldest mortality score to the nextyoungest. Unlike the fusion based profile, however, the resultantsurvivorship (or mortality) estimates for each age class are notindependent of one another, with no possibilities for the ‘resur-rections’ often seen in fusion based profiles.
The lack of such anomalies in dentition based survivorship datais not then, as sometimes argued, a sign of the greater reliability ofthese measures. It is instead simply a reflection of the differentways inwhich these profiles are computed. In the case of the fusionbased curves major upswings in survivorship signal potentialproblems with either the proposed sequence of fusion, with thetaphonomy of the assemblage, or, possibly, an indication of someancient behavior that affected the ages of animals in an assemblage(i.e. seasonality of movement, export or import of certain age ani-mals, or selective access to specific parts of an animal). Survivorshipprofiles based on dental eruption and wear may be affected bysimilar issues, but these will be masked by the method ofcomputing these profiles that always results in a pattern ofdecreasing survivorship (or increasing morality) with each suc-cessive age class.
Even after these computational issues are addressed, anotherchallenge in comparing age profiles computed using these twodifferent types of data arises over the degree of correlation betweenthe different age estimates based on dental eruption and wear andthose based on epiphyseal fusion (Hongo and Meadow, 1998). Willan estimate of the proportion of animals in an assemblage survivingbeyond a certain age based on dental data, for example, be the sameas a survivorship estimate for that same assemblage based onepiphyseal fusion? The system proposed here offers a remedy thatmay alleviate this concern. Specifically, the paired crania and post-crania of the modern sample used to develop the dentition basedaging system proposed in Lemoine et al. and the fusion basedsystem proposed here provide, as we saw above, a means of cali-brating the various age classes defined for each system. If thislinkage is indeed reliable, then an age profile constructed for anassemblage using post cranial fusion, should resemble that con-structed that for the same assemblage using dental eruption andwear patterns. This is especially so for the large Hallan Çemiassemblage with its tight temporal and spatial control and therigorous collection protocol followed in the excavation of the site.
3.5. Comparing Hallan Çemi long bone fusion and dentition-basedage profiles
Dentition based age profiles for the Hallan Çemi pigs are pre-sented in Table 8 for the ten stage “specific system” described inLemoine et al. (2014). This table provides the age range for each ofthe ten age classes in this system, the number of specimens in theHallan Çemi assemblage assigned to each age class, and the mor-tality score for each class, computed as the percentage of specimensamong the entire sample of ageable specimens assigned to each ageclass. The final two rows of the table convert these data into sur-vivorship and mortality profiles following the procedure describedabove. These latter two columns are comparable to the Hallan ÇemiSurvivorship and Mortality Scores based on long bone fusion inTable 7.
The dental aging system presented in Lemoine et al. alsoincluded two less fine-grained classification systems that combinedcertain age classes into broader age classes that encompassed awider range of wear stages for different teeth. The seven age class“simplified-A” system is formed by combining previously distinctage classes for the wear of deciduous teeth and permanent molars,while the “simplified-B” system is comprised of only four age classes
consisting of an initial stage that includes the eruption, wear, andshedding of deciduous teeth and three subsequent stages definedprimarily by molar eruption and broadly defined wear patterns.These additional aging systems were developed so that less com-plete tooth rows and individual teeth inwear stages that spanned anumber of the age classes in the specific system could be used tocompute dentition based age profiles. Thuswhile these two broaderclassificatory systems have less resolution than the fine grainedspecific system, their less restrictive classification rules increasesthe sample size of ageable specimens, making them attractive al-ternatives in smaller, more fragmentary assemblages. Tables 9 and10 present the dentition based age profiles for Hallan Çemi sam-ple following these two more simplified aging systems.
Fig. 5 compares survivorship curves computed using the eightstage long bone fusion system presented here (in red) to survi-vorship profiles computed using the ten stage dentition basedspecimen (in blue), the seven stage simplified-A system (in green),and the four stage simplified-B system (in purple). The graph usesthe end of each of the estimated time spans for the various ageclasses to scale the data to the ten-year time span captured in thesedifferent aging systems. Long bone age class F with an estimatedage range of between 18 and 24 months, for example, has a sur-vivorship score of 67.4, so the marker for this age class is placed onthe graph at the two year point indicating that 67% of the elementsthat make up this age class (the first phalanx and the distal tibia)came from animals older than two years of age. Similarly, the datapoint for the dental age class 6 in the specific system is placed at theend of 18e30 month age estimate for this age class, indicating that61% of the specimens that could be aged using this system wereolder than 30 months.
The long bone fusion survivorship curve and the ten stagespecific system dentition based curve track one another veryclosely, especially in the younger age classes CeG in the long bonesystem and age classes 1e6 in the specific dentition system that aredefined primarily on the basis of tooth eruption. There is an eightpercentage point difference between the score for long bone fusion
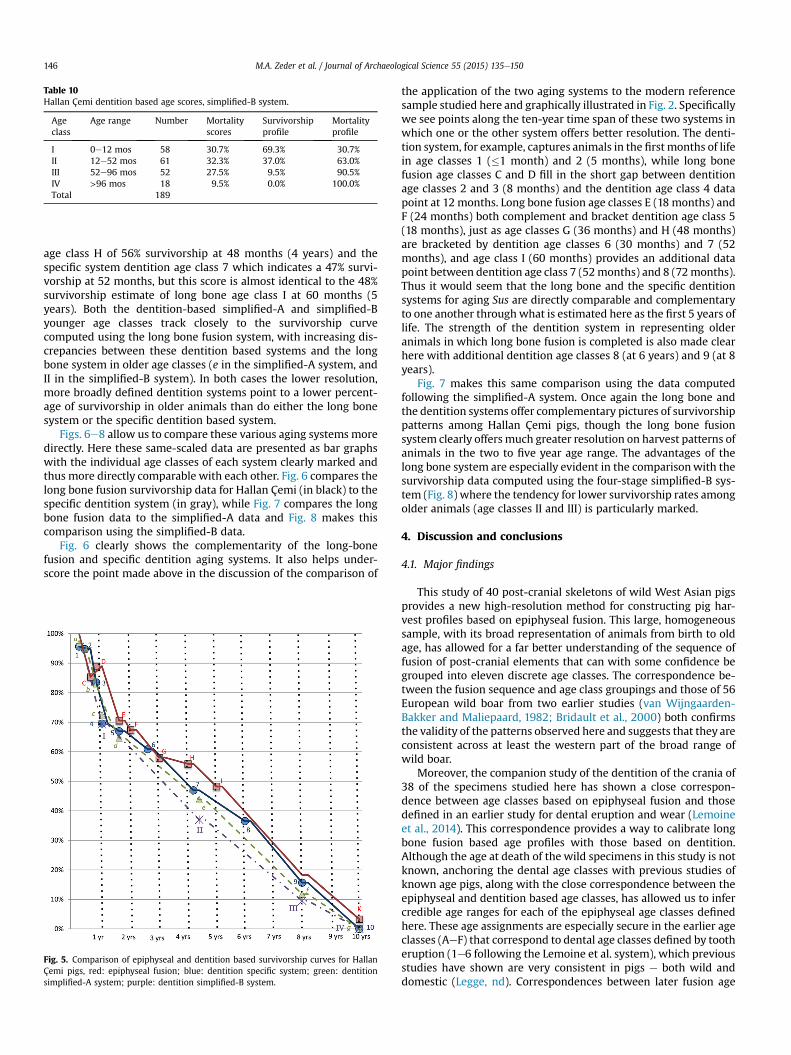
Table 10Hallan Çemi dentition based age scores, simplified-B system.
Ageclass
Age range Number Mortalityscores
Survivorshipprofile
Mortalityprofile
I 0e12 mos 58 30.7% 69.3% 30.7%II 12e52 mos 61 32.3% 37.0% 63.0%III 52e96 mos 52 27.5% 9.5% 90.5%IV >96 mos 18 9.5% 0.0% 100.0%Total 189
M.A. Zeder et al. / Journal of Archaeological Science 55 (2015) 135e150146
age class H of 56% survivorship at 48 months (4 years) and thespecific system dentition age class 7 which indicates a 47% survi-vorship at 52 months, but this score is almost identical to the 48%survivorship estimate of long bone age class I at 60 months (5years). Both the dentition-based simplified-A and simplified-Byounger age classes track closely to the survivorship curvecomputed using the long bone fusion system, with increasing dis-crepancies between these dentition based systems and the longbone system in older age classes (e in the simplified-A system, andII in the simplified-B system). In both cases the lower resolution,more broadly defined dentition systems point to a lower percent-age of survivorship in older animals than do either the long bonesystem or the specific dentition based system.
Figs. 6e8 allow us to compare these various aging systems moredirectly. Here these same-scaled data are presented as bar graphswith the individual age classes of each system clearly marked andthus more directly comparable with each other. Fig. 6 compares thelong bone fusion survivorship data for Hallan Çemi (in black) to thespecific dentition system (in gray), while Fig. 7 compares the longbone fusion data to the simplified-A data and Fig. 8 makes thiscomparison using the simplified-B data.
Fig. 6 clearly shows the complementarity of the long-bonefusion and specific dentition aging systems. It also helps under-score the point made above in the discussion of the comparison of
Fig. 5. Comparison of epiphyseal and dentition based survivorship curves for HallanÇemi pigs, red: epiphyseal fusion; blue: dentition specific system; green: dentitionsimplified-A system; purple: dentition simplified-B system.
the application of the two aging systems to the modern referencesample studied here and graphically illustrated in Fig. 2. Specificallywe see points along the ten-year time span of these two systems inwhich one or the other system offers better resolution. The denti-tion system, for example, captures animals in the first months of lifein age classes 1 (�1 month) and 2 (5 months), while long bonefusion age classes C and D fill in the short gap between dentitionage classes 2 and 3 (8 months) and the dentition age class 4 datapoint at 12 months. Long bone fusion age classes E (18 months) andF (24 months) both complement and bracket dentition age class 5(18 months), just as age classes G (36 months) and H (48 months)are bracketed by dentition age classes 6 (30 months) and 7 (52months), and age class I (60 months) provides an additional datapoint between dentition age class 7 (52months) and 8 (72months).Thus it would seem that the long bone and the specific dentitionsystems for aging Sus are directly comparable and complementaryto one another through what is estimated here as the first 5 years oflife. The strength of the dentition system in representing olderanimals in which long bone fusion is completed is also made clearhere with additional dentition age classes 8 (at 6 years) and 9 (at 8years).
Fig. 7 makes this same comparison using the data computedfollowing the simplified-A system. Once again the long bone andthe dentition systems offer complementary pictures of survivorshippatterns among Hallan Çemi pigs, though the long bone fusionsystem clearly offers much greater resolution on harvest patterns ofanimals in the two to five year age range. The advantages of thelong bone system are especially evident in the comparisonwith thesurvivorship data computed using the four-stage simplified-B sys-tem (Fig. 8) where the tendency for lower survivorship rates amongolder animals (age classes II and III) is particularly marked.
4. Discussion and conclusions
4.1. Major findings
This study of 40 post-cranial skeletons of wild West Asian pigsprovides a new high-resolution method for constructing pig har-vest profiles based on epiphyseal fusion. This large, homogeneoussample, with its broad representation of animals from birth to oldage, has allowed for a far better understanding of the sequence offusion of post-cranial elements that can with some confidence begrouped into eleven discrete age classes. The correspondence be-tween the fusion sequence and age class groupings and those of 56European wild boar from two earlier studies (van Wijngaarden-Bakker and Maliepaard, 1982; Bridault et al., 2000) both confirmsthe validity of the patterns observed here and suggests that they areconsistent across at least the western part of the broad range ofwild boar.
Moreover, the companion study of the dentition of the crania of38 of the specimens studied here has shown a close correspon-dence between age classes based on epiphyseal fusion and thosedefined in an earlier study for dental eruption and wear (Lemoineet al., 2014). This correspondence provides a way to calibrate longbone fusion based age profiles with those based on dentition.Although the age at death of the wild specimens in this study is notknown, anchoring the dental age classes with previous studies ofknown age pigs, along with the close correspondence between theepiphyseal and dentition based age classes, has allowed us to infercredible age ranges for each of the epiphyseal age classes definedhere. These age assignments are especially secure in the earlier ageclasses (AeF) that correspond to dental age classes defined by tootheruption (1e6 following the Lemoine et al. system), which previousstudies have shown are very consistent in pigs e both wild anddomestic (Legge, nd). Correspondences between later fusion age
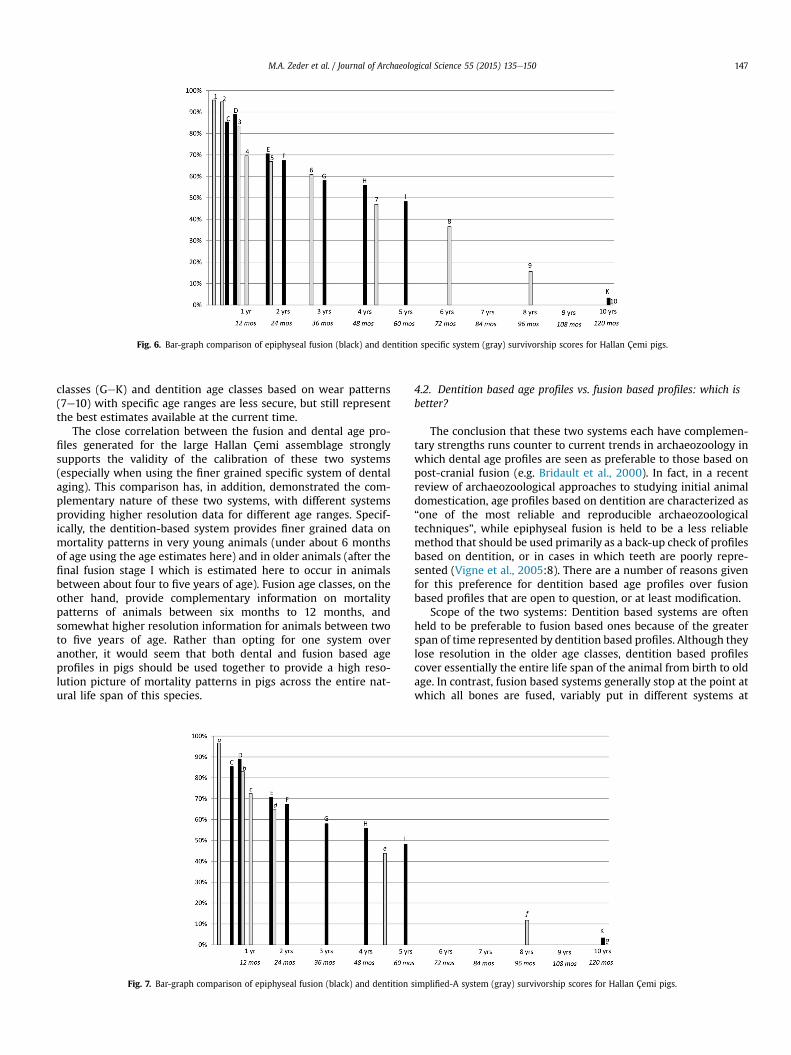
Fig. 6. Bar-graph comparison of epiphyseal fusion (black) and dentition specific system (gray) survivorship scores for Hallan Çemi pigs.
M.A. Zeder et al. / Journal of Archaeological Science 55 (2015) 135e150 147
classes (GeK) and dentition age classes based on wear patterns(7e10) with specific age ranges are less secure, but still representthe best estimates available at the current time.
The close correlation between the fusion and dental age pro-files generated for the large Hallan Çemi assemblage stronglysupports the validity of the calibration of these two systems(especially when using the finer grained specific system of dentalaging). This comparison has, in addition, demonstrated the com-plementary nature of these two systems, with different systemsproviding higher resolution data for different age ranges. Specif-ically, the dentition-based system provides finer grained data onmortality patterns in very young animals (under about 6 monthsof age using the age estimates here) and in older animals (after thefinal fusion stage I which is estimated here to occur in animalsbetween about four to five years of age). Fusion age classes, on theother hand, provide complementary information on mortalitypatterns of animals between six months to 12 months, andsomewhat higher resolution information for animals between twoto five years of age. Rather than opting for one system overanother, it would seem that both dental and fusion based ageprofiles in pigs should be used together to provide a high reso-lution picture of mortality patterns in pigs across the entire nat-ural life span of this species.
Fig. 7. Bar-graph comparison of epiphyseal fusion (black) and dentition s
4.2. Dentition based age profiles vs. fusion based profiles: which isbetter?
The conclusion that these two systems each have complemen-tary strengths runs counter to current trends in archaeozoology inwhich dental age profiles are seen as preferable to those based onpost-cranial fusion (e.g. Bridault et al., 2000). In fact, in a recentreview of archaeozoological approaches to studying initial animaldomestication, age profiles based on dentition are characterized as“one of the most reliable and reproducible archaeozoologicaltechniques”, while epiphyseal fusion is held to be a less reliablemethod that should be used primarily as a back-up check of profilesbased on dentition, or in cases in which teeth are poorly repre-sented (Vigne et al., 2005:8). There are a number of reasons givenfor this preference for dentition based age profiles over fusionbased profiles that are open to question, or at least modification.
Scope of the two systems: Dentition based systems are oftenheld to be preferable to fusion based ones because of the greaterspan of time represented by dentition based profiles. Although theylose resolution in the older age classes, dentition based profilescover essentially the entire life span of the animal from birth to oldage. In contrast, fusion based systems generally stop at the point atwhich all bones are fused, variably put in different systems at
implified-A system (gray) survivorship scores for Hallan Çemi pigs.
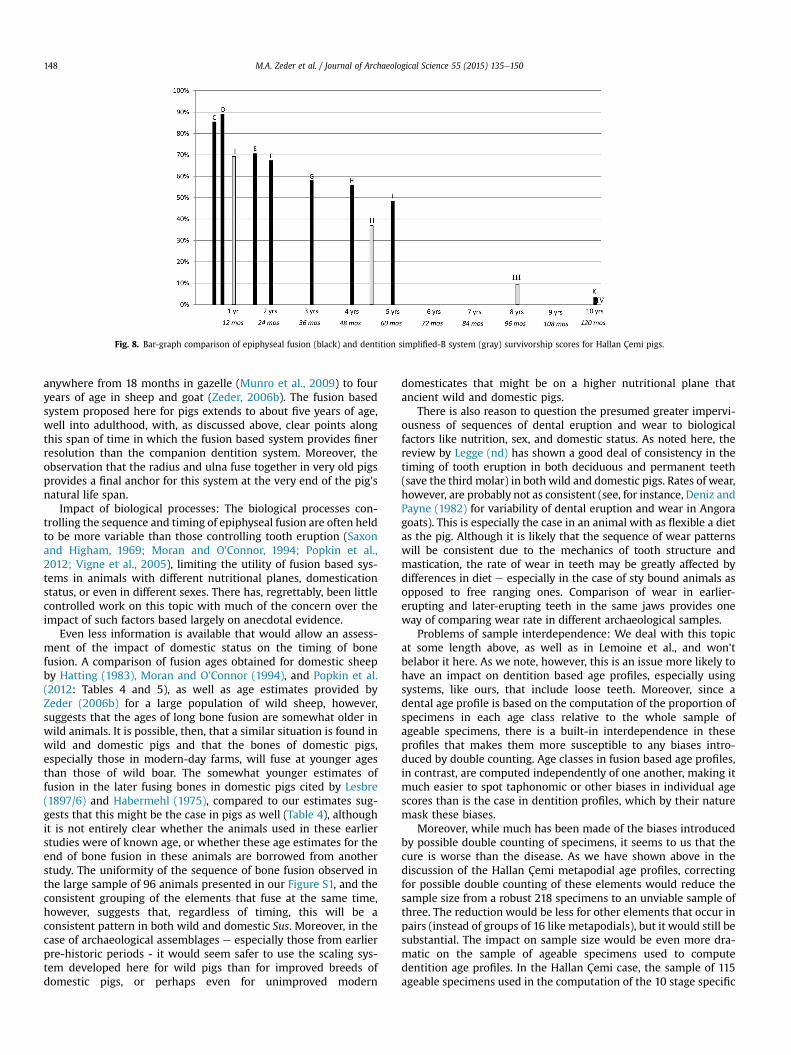
Fig. 8. Bar-graph comparison of epiphyseal fusion (black) and dentition simplified-B system (gray) survivorship scores for Hallan Çemi pigs.
M.A. Zeder et al. / Journal of Archaeological Science 55 (2015) 135e150148
anywhere from 18 months in gazelle (Munro et al., 2009) to fouryears of age in sheep and goat (Zeder, 2006b). The fusion basedsystem proposed here for pigs extends to about five years of age,well into adulthood, with, as discussed above, clear points alongthis span of time in which the fusion based system provides finerresolution than the companion dentition system. Moreover, theobservation that the radius and ulna fuse together in very old pigsprovides a final anchor for this system at the very end of the pig'snatural life span.
Impact of biological processes: The biological processes con-trolling the sequence and timing of epiphyseal fusion are often heldto be more variable than those controlling tooth eruption (Saxonand Higham, 1969; Moran and O'Connor, 1994; Popkin et al.,2012; Vigne et al., 2005), limiting the utility of fusion based sys-tems in animals with different nutritional planes, domesticationstatus, or even in different sexes. There has, regrettably, been littlecontrolled work on this topic with much of the concern over theimpact of such factors based largely on anecdotal evidence.
Even less information is available that would allow an assess-ment of the impact of domestic status on the timing of bonefusion. A comparison of fusion ages obtained for domestic sheepby Hatting (1983), Moran and O'Connor (1994), and Popkin et al.(2012: Tables 4 and 5), as well as age estimates provided byZeder (2006b) for a large population of wild sheep, however,suggests that the ages of long bone fusion are somewhat older inwild animals. It is possible, then, that a similar situation is found inwild and domestic pigs and that the bones of domestic pigs,especially those in modern-day farms, will fuse at younger agesthan those of wild boar. The somewhat younger estimates offusion in the later fusing bones in domestic pigs cited by Lesbre(1897/6) and Habermehl (1975), compared to our estimates sug-gests that this might be the case in pigs as well (Table 4), althoughit is not entirely clear whether the animals used in these earlierstudies were of known age, or whether these age estimates for theend of bone fusion in these animals are borrowed from anotherstudy. The uniformity of the sequence of bone fusion observed inthe large sample of 96 animals presented in our Figure S1, and theconsistent grouping of the elements that fuse at the same time,however, suggests that, regardless of timing, this will be aconsistent pattern in both wild and domestic Sus. Moreover, in thecase of archaeological assemblages e especially those from earlierpre-historic periods - it would seem safer to use the scaling sys-tem developed here for wild pigs than for improved breeds ofdomestic pigs, or perhaps even for unimproved modern
domesticates that might be on a higher nutritional plane thatancient wild and domestic pigs.
There is also reason to question the presumed greater impervi-ousness of sequences of dental eruption and wear to biologicalfactors like nutrition, sex, and domestic status. As noted here, thereview by Legge (nd) has shown a good deal of consistency in thetiming of tooth eruption in both deciduous and permanent teeth(save the third molar) in both wild and domestic pigs. Rates of wear,however, are probably not as consistent (see, for instance, Deniz andPayne (1982) for variability of dental eruption and wear in Angoragoats). This is especially the case in an animal with as flexible a dietas the pig. Although it is likely that the sequence of wear patternswill be consistent due to the mechanics of tooth structure andmastication, the rate of wear in teeth may be greatly affected bydifferences in diet e especially in the case of sty bound animals asopposed to free ranging ones. Comparison of wear in earlier-erupting and later-erupting teeth in the same jaws provides oneway of comparing wear rate in different archaeological samples.
Problems of sample interdependence: We deal with this topicat some length above, as well as in Lemoine et al., and won'tbelabor it here. As we note, however, this is an issue more likely tohave an impact on dentition based age profiles, especially usingsystems, like ours, that include loose teeth. Moreover, since adental age profile is based on the computation of the proportion ofspecimens in each age class relative to the whole sample ofageable specimens, there is a built-in interdependence in theseprofiles that makes them more susceptible to any biases intro-duced by double counting. Age classes in fusion based age profiles,in contrast, are computed independently of one another, making itmuch easier to spot taphonomic or other biases in individual agescores than is the case in dentition profiles, which by their naturemask these biases.
Moreover, while much has been made of the biases introducedby possible double counting of specimens, it seems to us that thecure is worse than the disease. As we have shown above in thediscussion of the Hallan Çemi metapodial age profiles, correctingfor possible double counting of these elements would reduce thesample size from a robust 218 specimens to an unviable sample ofthree. The reduction would be less for other elements that occur inpairs (instead of groups of 16 like metapodials), but it would still besubstantial. The impact on sample size would be even more dra-matic on the sample of ageable specimens used to computedentition age profiles. In the Hallan Çemi case, the sample of 115ageable specimens used in the computation of the 10 stage specific

M.A. Zeder et al. / Journal of Archaeological Science 55 (2015) 135e150 149
dental age profile (Table 8) would be reduced to 15 specimens ifonly mandibular tooth rows from one side of the jaw were used.
Problems of interdependence and double counting are besthandled by taking simple precautions when analyzing assemblagese working to reunite loose teeth from the same or nearby prove-nience into single tooth rows, and taking care to base assessments offusion on MNEs within single proveniences. Beyond controlling fordouble counting of bones within the same or conjoined pro-veniences, it seems to us that eliminating specimens from moredistant excavation units on the remote possibility that they mightconceivably belong to the same individual runs the risk of removing alarge number of viable specimens from the sample used to computethe age profile. The likelihood, after all, of teeth or post-cranial ele-ments from single animals popping up in spatially, and perhaps eventemporally, different deposits across an excavation seems small.
Taphonomy: Concerns arising over taphonomic biases againstthe preservation of the bones of young animals are also often citedas a reason why age profiles based on epiphyseal fusion is lessreliable than those based on dental eruption and wear (Payne,1972; Munson, 2000; Watson, 1978). And while such biasesalmost certainly exist (especially in assemblages that are highlyfragmented, subjected to substantial carnivore gnawing, orcollected without the benefit of screening), it seems to us thatdental elements are also vulnerable to taphonomic bias. The de-ciduous teeth and tooth germs of younger animals are particularlysusceptible to breakage or to being overlooked in handpickedsamples. Our earlier study (Lemoine et al., 2014) showed that ageprofiles computed using only loose teeth indicated a substantiallyhigher mortality among younger animals than did those computedusing tooth rows. This suggests that the jaws of younger animalsare more likely to be fragmented than those of older animals,freeing their teeth from the surrounding bone and making themeven more vulnerable to destruction or oversight. If one only usesmandibular teeth in jaws to compute the dentition based ageprofile (as is the case inmany systems that followGrant,1982), thenthis bias against younger animals will be even more keenly felt.
Once again, rather than throwing up ones hands and proclaimingthat taphonomic biases have rendered age profiles (dentition orfusion based) unreliable, a more productive approach would be totake some common sense precautions to try to limit these potentialbiases. Certainly taking the degree of post-depositional movementand destruction into account would seem essential, as would beingmindful of the degree of care with which the sample was recovered.Cross-checking between the correspondence of dentition and fusionbasedageprofiles is anotherway to look for thesebiases, as is lookingfor agreement in the fusion scores for elements within and betweenage classes as was done here. Comparing the MNE or MNI counts ofmandibles andmaxillae against those of post-cranial elements usedto compute fusion based profiles would seem another useful way ofsorting out biases arising from differential destruction of assem-blages, or even those stemming from ancient butchery or bone usepractices that differentially affect elements used to compute ageprofiles. In general, any element (dental or post-cranial) with arelatively low MNI count is unlikely to give reliable data and shouldbe either excluded altogether or viewed with caution.
Also on the topic of taphonomy, it seems instructive to comparethe very different size of the samples that could be used to computedental and fusion based age profiles in the extraordinarily largeHallan Çemi sample, 115 specimens for the specific dental profile vs.1744 for the fusionbasedprofile. Post-cranial elements ingeneral, dueto their larger numbers in the skeleton and their greater resilience topre and post-depositional destructive forces, stand a better chance ofbeing recovered in larger numbers from archaeological sites thandental elements (especially in the absence of extensive carnivoregnawing). We know of few other assemblages of archaeological Sus
that are as large as that from Hallan Çemi, especially if one considersthe temporal and spatial homogeneity of this sample. Other Sus as-semblages are typicallymuch smaller,with aneven smaller sample ofageable teeth relative to long bones. So just on sheer numbers ofspecimens that can beused to compute the age profile, itwould seemthat the fusion-based system has the upper hand. It is possible,however, that intensive carnivore gnawing may have a particularlysevere impacton thepreservationofpost-cranial elements, especiallythe greasy long bones of pigs, resulting in an assemblage dominatedby dental elements. But once again, such biases can be detected usingthe common sense methods mentioned above to evaluate therespective reliability of dental and long bone fusion age profiles.
Sex: One clear potential advantage of fusion based age profilesover dentition based ones in animals withmarked degrees of sexualdimorphism in body size is that they can be used to computeseparate age profiles for male and female animals (Zeder, 2001).Since both hunters and herders take sex as well as age into accountin culling strategies, the ability to compute sex-specific harvestprofiles offers special insight into these strategies. Sex-specificculling patterns are particularly important for monitoring theinitial steps toward animal domestication which are marked byshifts in the ages and sexes of animals harvested as the goal of thesestrategies move frommaximizing short term returns from the deadanimal to the long term viability of the living herd (Meadow, 1989;Zeder, 2006a) e shifts in strategy that long precede any detectablemorphological change in the animals subjected to these differentstrategies (Zeder, 2012). Zeder has developed methods forcomputing fusion based sex-specific harvest profiles in sheep andgoats that have been effectively used tomonitor the transition frommore classical hunting strategies to those consistent with herdingin the central Zagros region of the Near East (Zeder, 2001). Denti-tion based age profiles cannot be used for this purpose. Settingaside the lack of reliable methods for distinguishing between theteeth of sheep and goats (especially for molars and mandibularbone, see Zeder and Pilaar, 2009) that renders dentition based ageprofiles less useful in caprines, the teeth of these animals simply donot show any of the marked degree of dimorphism seen in the longbones of goats and to a slightly lesser degree in sheep.
Post-cranial elements of pigs are also held to behighly dimorphic(Albarella et al., 2006; Ervynck et al., 2001), something demon-strated by an early publication by Payne and Bull (1988) on themodern Turkish sample included in the present study. Preliminaryresults of our study of the bones and teeth ofmodernwild boar fromWest Asia confirm this widely held belief, with marked differencesin the length, breadth, and depth measurements of almost all ele-ments ofmale and female animals beginning as early as our age classD (7e8 months) and clearly expressed differences in animals in ageclass E (8e18 months) and older. On the other hand (and contra thesuggestion in Evin et al., 2013), with the exception of canines, thereis no evidence of any dimorphism in the size of the teeth ofmale andfemale pigs in our sample. Exploring these patterns, and applyingthem to the construction of sex-specific age profiles in pigs, will bethe focus of the third in this series of papers.
Fusion based age profiles are, then, an important tool capableof providing a high resolution picture of prehistoric prey/harveststrategies in pigs and other animals. Used carefully in conjunc-tion with dentition based profiles they hold the potential ofshedding new light on a wide variety of issues e from thetransition from hunting to herding to the development ofeconomically differentiated management strategies. We hopethat our efforts in this paper on epiphyseal fusion in pigs and ourearlier contribution on dentition based age profiles, along with afuture third paper on sexual dimorphism in pigs, will contributeuseful new methods for achieving these goals in the study ofancient pig exploitation.

M.A. Zeder et al. / Journal of Archaeological Science 55 (2015) 135e150150
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.jas.2014.12.017.
References
Albarella, U., Dobney, K., Rowley-Conwy, P., 2006. The domestication of the pig (Susscrofa): newchallengesandapproaches. In: Zeder,M.A., Bradley,D., Emschwiller, E.,Smith, B.D. (Eds.), Documenting Domestication: New Genetic and ArchaeologicalApproaches. University of California Press, Berkeley, CA, pp. 209e227.
Anezaki, T., 2009. Estimating age at death in Jomon Japanese wild boar (Sus ScrofaLeucomystax) based on the timing of molar eruption in recent comparativesamples. Mamm. Soc. Jpn. 34 (2), 53e63.
B€ok€onyi, S., 1972. Zoological evidence for seasonal or permanent occupation ofprehistoric settlements. In: Ucko, P.J., Tringham, R., Dimbleby, G.W. (Eds.), Man,Settlement, and Urbanism. Duckworth, London, pp. 121e126.
Bridault, A., Vigne, J.-D., Horard-Herbin, M.-P., Pell�e, E., Fiquet, P., Mashkour, M.,2000. Wild boar e age at death estimates: the relevance of new modern datafor archaeological skeletal material. 1. presentation on the corpus. dental andepiphyseal fusion ages. Ibex J. Mt. Ecol. 5, 11e18.
Bull, G., Payne, S., 1982. Tooth eruption and epiphysial fusion in pigs and wild boar.In: Wilson, B., Grigson, C., Payne, S. (Eds.), Ageing and Sexing Animal Bonesfrom Archaeological Sites, BAR British Series, vol. 109, pp. 55e71.
De Magalh~aes, J., Costa, J., 2009. A database of vertebrate longevity records and theirrelation to other life-history traits. Sch. Biolog. Sci. 22, 1770e1774.
Deniz, E., Payne, S., 1982. Eruption and wear in the mandibular dentition as a guideto ageing Turkish Angora goats. In: Wilson, B., Grigson, C., Payne, S. (Eds.),Ageing and Sexing Animal Bones from Archaeological Sites, British Archaeo-logical Reports, British Series, vol. 109. BAR, Oxford, pp. 153e206.
Dyson, R.H., 1953. Archaeology and the domestication of animals in the Old World.Am. Anthropol. 55, 661e673.
Ervynck, A., Dobney, K., Hongo, H., Meadow, R., 2001. Born free? New evidence forthe status of Sus scrofa at neolithic Çay€onü Tepesi (Southeastern Anatolia,Turkey). Pal�eorient 27, 47e73.
Evin, A., Cucchi, T., Cardini, A., Strand Vidarsdottir, U., Larson, G., Dobney, K., 2013.The long and winding road: identifying pig domestication through molar sizeand shape. J. Archaeol. Sci. 40, 735e743.
Grant, A., 1982. The use of tooth wear as a guide to the domestic ungulates. In:Wilson, B., Grigson, C., Payne, S. (Eds.), Ageing and Sexing Animal Bones fromArchaeological Sites, BAR British Series, vol. 109, pp. 91e108.
Habermehl, K.H., 1975. Die altersbestimmung bei hausund labortieren. Parey, Berlin& Hamburg.
Hatting, T., 1983. Osteological investigations on Ovis aries L. Videnskabelige Med-delelser fra Dansk Naturhistorisk Forening. Bind 144, 115e135.
Helmer, D., 2008. Revision de la faune de Cafer H€oyük (Malatya, Turquie): Apportsdes methods de l'analyse des m�elanges et se l'analyse de Kernal �a la mise enevidence de la domestication. In: Vila, E., Goucherin, L., Choyke, A.,Buitenhuis, H. (Eds.), Archaeozoology of the Near East VIII. Travaux de la Maisonde l'Orient et de la M�editerran�ee (TMO), Lyon, pp. 169e195.
Hole, F., Flannery, K.V., Neely, J.A., 1969. Prehistory and Human Ecology on the DehLuran Plain: Memoirs of the Museum of Anthropology. The University ofMichigan Press, Ann Arbor.
Hongo, H., Meadow, R.H., 1998. Pig exploitation at neolithic Cay€onü Tepesi(Southeastern Anatolia). In: Nelson, S.M. (Ed.), Ancestors for the Pigs: Pigs inPrehistory. MASCA, Philadelphia, pp. 77e98.
Jezierski, W., 1977. Longevity and mortality rate in a population of wild boar. ActaTheriol. 22/24, 337e348.
Jones, G.C., 2006. Tooth eruption and wear observed in live sheep from Butser Hill,the Cotswold Farm Park and five farms in the Pentland Hille, UK. In: Ruscillo, D.(Ed.), Recent Advances in Aging and Sexing Animal Bones. Durham: 9th ICAZConference, pp. 155e178.
Legge, A.J., nd. ‘Practice with Science’: molar tooth eruption ages in domestic, feral,and wild pigs (Sus scrofa). Int. J. Osteoarchaeol. (unpublished manuscript).
Lemoine, X., Zeder, M.a., Bishop, K., Rufolo, S., 2014. A new system for computingdentition-based age profiles in Sus scrofa. J. Archaeol. Sci. 40, 179e193.
Lesbre, F.-X., 1897/1898. Contribution a l'�etude de l'ossification du squelette desmammif�eres domestiques. In: Annales de la Societ�e d'Agriculture, pp. 1e106.Sciences, et Industrie de Lyon. S�er. 7, 5.
Matschke, G.H., 1967. Aging European wild hogs by dentition. J. Wildl. Manag. 31,109e113.
Meadow, R.H., 1989. Osteological evidence for the process of animal domestication.In: Clutton-Brock, J. (Ed.), The Walking Larder: Patterns of Domestication,Pastoralism, and Predation. Unwin Publications, London, pp. 80e90.
Milligan, B.N., Fraser, D., Kramer, D.L., 2001. Birth weight variation in the domesticpig: effects on offspring survival, weight gain, and suckling behavior. Appl.Anim. Behav. Sci. 73, 179e191.
Moran,N.C., O'Connor, T.P.,1994.Ageattribution indomestic sheepbyskeletal anddentalmaturation: a pilot study of available sources. Int. J. Osteoarchaeol. 4, 267e285.
Munro, N.D., Bar-Oz, G., Stutz, A.J., 2009. Aging mountain gazelle (Gazella gazella):refining methods of tooth eruption and wear and bone fusion. J. Archaeol. Sci.36, 952e963.
Munson, P.J., 2000. Age-correlated differential destruction of bones and its effect onarchaeological mortality profiles of domestic sheep and goat. J. Archaeol. Sci. 27,405e416.
Payne, S., 1972. The interpretation of bone samples from archaeological sites. In:Higgs, E. (Ed.), Papers in Economic Prehistory. Cambridge University Press,Cambridge, pp. 49e64.
Payne, S., 1973. Kill-off patterns in sheep and goats: the mandibles from Asvan Kale.Anatol. Stud. 23, 281e303.
Payne, S., Bull, G., 1988. Components of variation in measurements of pig bones andteeth, and the use of measurements to distinguish wild from domestic pig re-mains. Archaeozoologia 2, 27e66.
Peasnall, B.L., 2000. The Round-houseHorizonalong the Taurus/ZagrosArc: a Synthesisof Recent Excavations of Late Epipaleolithic and Early Aceramic Sites in South-eastern Anatolia and Northern Iraq (PhD dissertation). University of Pennsylvania.
Perkins, D.P., 1964. Prehistoric fauna from Shanidar, Iraq. Science 144, 1565e1566.Popkin, P.R.W., Baker, P., Worley, F., Payne, S., Hammon, A., 2012. The Sheep Project
(1): determining skeletal growth, timing of epiphyseal fusion and morpho-metric variation in unimproved Shetland sheep of known age, sex, castrationstatus and nutrition. J. Archaeol. Sci. 39, 1775e1792.
Redding, R.W., 1981. The faunal remains. In: Wright, H.T. (Ed.), An Early Town on theDeh Luran Plain: Excavations at Tepe Farukhabad, Memoirs of the Museum ofAnthropology, Number 13. University of Michigan, Ann Arbor, pp. 23e261.
Redding, R.W., 2005. Breaking the mold, a consideration of variation in the evolu-tion of animal domestication. In: Vigne, J.-D., Helmer, D. (Eds.), The First Steps ofAnimal Domestication: New Zooarchaeological Approaches. Oxbow Books,Oxford, pp. 41e48.
Redding, R.W., Rosenberg, M., 1998. Ancestral pigs: a New (Guinea) model for pigdomestication. In: Nelson, S.M. (Ed.), Ancestors for the Pigs: Pigs in Prehistory.MASCA, Philadelphia, pp. 65e76.
Redding, R.W., Rosenberg, M., 2000. Hallan Çemi and early village organization inEastern Anatolia. In: Kuijt, Ian (Ed.), Life in Neolithic Farming Communities.Plenum Publishers, New York, pp. 39e52.
Reiland, S., 1978. Growth and skeletal development in the pig. Acta Radiol. Suppl.358, 15e22.
Rosenberg, M., Nesbitt, R., Redding, R.W., Peasnall, B.L., 1998. Hallan Çemi, pighusbandry, and post-Pleistocene adaptations along the Taurus-Zagros Arc(Turkey). Pal�eorient 24, 25e41.
Saxon, A., Higham, C., 1969. A new research method for economic prehistorians.Am. Antiq. 34, 303e311.
Silver, I.A., 1969. The ageing of domestic animals. In: Brothwell, D., Higgs, E.S. (Eds.),Science in Archaeology. Thames and Hudson, London, pp. 283e302.
Starkovich, B.M., Stiner, M.C., 2009. Hallan Çemi Tepesi: high-ranked gameexploitation alongside intensive seed processing at the Epipaleolithic-Neolithictransition in southeastern Turkey. Anthropozoologica 44, 41e61.
Stiner, M.C., 1990. The use of mortality patterns in archaeological studies of Hom-inid predatory adaptations. J. Anthropol. Archaeol. 305e351.
Stiner, M.C., Buitenhuis, H., Duru, G., Kuhn, S.L., Mentzer, S.M., Munro, N.D.,P€ollath, N., Quade, J., Tsartsidou, G., €Ozbașaran, 2014. A forager-herder trade-off, from broad spectrum hunting to sheep management at Așıklı H€oyük,Turkey. Proc. Natl. Acad. Sci. 11, 8404e8409.
Van Wijngaarden-Bakker, L.H., Maliepaard, C.H., 1982. Leeftijdsbepaling aan hetskelet van het wilde zwijn Sus scrofa Linnaeus, 1758. Lutra 25, 30e37.
Vigne, J.-D., Helmer, D., Peter, J., 2005. New archaeological approaches to trace thefirst steps of animal domestication: general presentation, reflections, and pro-posals. In: The First Steps of Animal Domestication: New Archaeological Ap-proaches. Oxbow Books, Oxford, pp. 1e16.
Vigne, J.-D., Brios, F., Zazzo, A., Wilcox, G., Cucchi, T., Thi�ebault, S., Carr�ere, I.,Franel, Y., Touquet, R., Martin, C., Moreau, C., Comby, C., Guilane, J., 2012. Firstwave of cultivators spread to Cyprus at least 10,600 years ago. Proc. Natl. Acad.Sci. U. S. A. 109 (22), 8445e8449.
Watson, J.P.N., 1978. The interpretation of epiphyseal data. In: Brothwell, D.R.,Thomas, K.D., Clutton-Brock, J. (Eds.), Research Problems in Zooarchaeology.Institute of Archaeology, Occasional Publications 3, London, pp. 97e100.
Zeder, M.A., 1991. Feeding Cities: Specialized Animal Economy in the Ancient NearEast. Smithsonian Institution Press, Washington, DC.
Zeder, M.A., 2001. A metrical analysis of a collection of modern goats (Capra hircusaegargus and C. h. hircus) from Iran and Iraq: implications for the study ofcaprine domestication. J. Archaeol. Sci. 28, 61e79.
Zeder, M.A., 2006a. Archaeological approaches to documenting animal domestica-tion. In: Zeder, M.A., Decker-Walters, D., Bradley, D., Smith, B.D. (Eds.), Doc-umenting Domestication: New Genetic and Archaeological Paradigms.University of California Press, Berkeley, pp. 209e227.
Zeder,M.A., 2006b. Reconciling rates of long bone fusion and tooth eruption andwearin sheep (Ovis) and goat (Capra). In: Ruscillo, D. (Ed.), Recent Advances in Agingand Sexing Animal Bones. Durham: 9th ICAZ Conference, pp. 87e118.
Zeder, M.A., 2008. Animal domestication in the Zagros: an update and directions forfuture research. In: Vila, E., Goucherin, L., Choyke, A., Buitenhuis, H. (Eds.),Archaeozoology of the Near East VIII. Travaux de la Maison de l'Orient et de laM�editerran�ee (TMO), Lyon, pp. 243e278.
Zeder, M.A., 2012. The domestication of animals. J. Anthropol. Res. 68 (2), 161e190.Zeder, M.A., Pilaar, S., 2009. Assessing the reliability of criteria used to identify
mandibles and mandibular teeth in sheep, Ovis, and goats, Capra. J. Archaeol.Sci. 37, 3225e3242.


















