
Its Entomopatogenico
11
Nematology , 2000, Vol. 2(5), 477-487 Molecular characterisation of Heterorhabditis indica isolates from India, Kenya, Indonesia and Cuba Colin M. STACK 1 , Subbana G. E ASWARAMOORTHY 2 , Usha K. M ETHA 2 , Martin J. D OWNES 1 , Christine T. GRIFFIN 1 and Ann M. BURNELL 1, * 1 The Institute for Bioengineeringand Agroecology , Departme nt of Biology, National Unive rsity of Ir eland, Maynooth, Co. Kildare, Ir eland 2 Sugarc ane Br eeding Institute , Indian Council for Agric ultural Resear ch, Coimbator e, T amil Nadu 641 007, India Accepted for publication:20 November 1999 Summary – Isolates of Heterorhabditis were identied as H. indica using the follow ing molecular diagnostic features: hybridisation to a H. indica speci c satellite DNA p robe; AluI and MboI restriction proles of the rDN A ITS PCR product and the AluI prole of the rDN A IGS PCR product. The Kenyan isolates represent a distinct subgroup of H. indica . These isolates lacked one of the two Hinf I restriction sites which are present in the r DN A ITS product of al l the other isolates tested and they also differed from other H. indica isolates in their rDNA IGS HaeIII restriction prole. T he Indian isolates are interfertile. The Keny an isolates are interfertile b ut only one Kenyan isolate, Ki3, produced viable progeny when crossed with H. indica LN2. T he four Ind onesian isolates are interfertile, but only one Indo nesianisolate (INA H1) producedviable hybri ds wh en crossed with H. indica LN2. INA H1 was also interfertile with the Ken yan isolate Ki3. Résumé – Caractérisation moléculaire d’isolats d ’Heterorhabd itis indica provenant d’Inde, du Kenya, d’Indonésie et de Cuba – Des isolats d’ Heterorhabditis ont été identiés comme H. indica par l’utilisation des techniques de caractérisation moléculaire suivant es: hybridation avec une sonde spécique du DN A satellite de H. indica , produits des prols de restriction par PCR de l’ITS du rDNA par AluI et MboI et produit de PCR de l’IGS du rDNA par AluI. Les isolats keniyans constituent un sous-groupe distinct d’ H. indica . Un des deux sites de restrictionde Hinf I, présent dans les pr oduits de l’ITS du rDNA de tous les autres isolats étudiés, est absent dans ces isolats qui différaient ég alement dans leurs pro ls de res triction de l’IGS du rDNA par HaeIII. Les is olats d’Inde sont interfertiles. Les isolats kenyans sont inter-fertilesmais un seul de ces isolats, Ki3, a produit une descendance viable aprè s croisement avec H. indica LN2. Les quatre isolats indonésiens sont interfertiles,mais un seul d’entre eux (INA H1) a produit des hybrides via bles après croisement avec H. indica LN2. IN A H1 a été également interfertile avec l ’isolat kenyan Ki3. Keywords – crossbreeding, DN A probe, ento mopathogenic nematode, molecular diagnostics, rDN A ITS, rD NA IGS , satellite DNA. Entomopathogenic nematodes (EPN) of the families Steinernematidae and Heterorhabditidae, together with thei r symbioti c bacte ria Xenorhabdusspp.and Photorhab- dus sp., respectively, are currently being mass produced commerci all y andused to cont rola vari etyof soil- dwelling insect pest s in Europe, USA, Aus tralia and China (see re- view s by K aya and Gaugl er, 1993; Ehle rs,1996).The bio- logical c ontrol potential of EPN has s timulated numerous surveys in an effort to nd new indigenous isolates and possibly also new species of Heterorhabditis and Stein- ernema (reviewed by Hominick et al. , 1996). Rapid and reliable diagnostic tests are required for species identi- cation in s uch surv eys and DN A ngerprintingtechniques * Correspondinga uthor, e-mail: ann.burnell@may.ie are now becoming more widely used as a rst screen to determine the s pecies composition of ne wly isolated EPN collections. These molecular approaches can then be s up- plemented by morphological, morphometric and cross- breeding techniques to conrm the identication of pu- tative new s pecies (reviewe d by Hominick et al., 1997). Although Steinernema and Heterorhabditis share many similarities in their mode of life and morphology, these similari ties result from convergent ev olution and are not indicative of a close phylogenetic relationship between the two families(Poinar, 1993; Sudhaus, 1993; Blaxt er et al., 1998). Steinernema also appears to be more species rich than is Heterorhabditis . Twenty two Steinernema c ® Koninklijke Brill NV , Leiden, 2000 477
-
Upload
alexandre-almeida -
Category
Documents
-
view
212 -
download
0
Transcript of Its Entomopatogenico

862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 111
Nematology 2000 Vol 2(5) 477-487
Molecular characterisation of Heterorhabditis indica isolates from
India Kenya Indonesia and Cub
Colin M STACK 1 Subbana G EASWARAMOORTHY 2 Usha K METHA 2 Martin J DOWNES
Christine T GRIFFIN 1 and Ann M BURNELL 1
1 The Institute for Bioengineering and Agroecology Department of Biology National University of Ireland Maynoo
Co Kildare Irela2 Sugarcane Breeding Institute Indian Council for Agricultural Research Coimbatore Tamil Nadu 641 007 Ind
Accepted for publication 20 November 19
Summary ndash Isolates of Heterorhabditis were identied as H indica using the following molecular diagnostic features hybridisatto a H indica specic satellite DNA probe AluI and MboI restriction proles of the rDNA ITS PCR product and the AluI prole of trDNA IGS PCR product The Kenyan isolates represent a distinct subgroup of H indica These isolates lacked one of the two Hin
restriction sites which are present in the rDNA ITS product of al l the other isolates tested and they also differed from other H ind
isolates in their rDNA IGS HaeIII restriction prole The Indian isolates are interfertile The Kenyan isolates are interfertile but onone Kenyan isolate Ki3 produced viable progeny when crossed with H indica LN2 The four Indonesian isolates are interfertile bonly one Indonesianisolate(INA H1) producedviable hybrids when crossed with H indica LN2 INA H1 was also interfertile with tKenyan isolate Ki3
Reacutesumeacute ndash Caracteacuterisation moleacuteculaire drsquoisolats d rsquoHeterorhabditis indica provenant drsquoInde du Kenya drsquoIndoneacutesie et de Cub
ndash Des isolats drsquo Heterorhabditis ont eacuteteacute identieacutes comme H indica par lrsquoutilisation des techniques de caracteacuterisation moleacuteculaisuivantes hybridation avec une sonde speacutecique du DNA satellite de H indica produits des prols de restriction par PCR de lrsquoITdu rDNA par AluI et MboI et produit de PCR de lrsquoIGS du rDNA par AluI Les isolats keniyans constituent un sous-groupe distindrsquo H indica Un des deux sites de restrictionde Hinf I preacutesent dans les produits de lrsquoITS du rDNA de tous les autres isolats eacutetudieacutes eabsent dans ces isolats qui diffeacuteraient eacutegalement dans leurs prols de restriction de lrsquoIGS du rDNA par HaeIII Les isolats drsquoInde sointerfertiles Les isolats kenyans sont inter-fertilesmais un seul de ces isolats Ki3 a produit une descendance viable apregraves croiseme
avec H indica LN2 Les quatre isolats indoneacutesiens sont interfertiles mais un seul drsquoentre eux (INA H1) a produit des hybrides viablapregraves croisement avec H indica LN2 INA H1 a eacuteteacute eacutegalement interfertile avec lrsquoisolat kenyan Ki3
Keywords ndash crossbreeding DNA probe entomopathogenicnematode molecular diagnostics rDNA ITS rDNA IGS satellite DNA
Entomopathogenic nematodes (EPN) of the familiesSteinernematidae and Heterorhabditidae together withtheir symbiotic bacteria Xenorhabdussppand Photorhab-
dus sp respectively are currently being mass producedcommercially andused to controla varietyof soil-dwellinginsect pests in Europe USA Australia and China (see re-views by Kaya and Gaugler 1993 Ehlers 1996)The bio-
logical control potential of EPN has stimulated numeroussurveys in an effort to nd new indigenous isolates andpossibly also new species of Heterorhabditis and Stein-
ernema (reviewed by Hominick et al 1996) Rapid andreliable diagnostic tests are required for species identi-cation in such surveys and DNA ngerprintingtechniques
Correspondingauthor e-mail annburnellmayie
are now becoming more widely used as a rst screen determine the species compositionof newly isolated EPcollections These molecular approaches can then be suplemented by morphological morphometric and crosbreeding techniques to conrm the identication of ptative new species (reviewed by Hominick et al 1997)
Although Steinernema and Heterorhabditis share ma
similarities in their mode of life and morphology thesimilarities result from convergent evolution and are nindicative of a close phylogenetic relationship betwethe two families (Poinar 1993 Sudhaus 1993 Blaxteral 1998) Steinernema also appears to be more specirich than is Heterorhabditis Twenty two Steinernem
creg Koninklijke Brill NV Leiden 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 211
CM Stack et al
species are recognised by Hominick et al (1997) Adamset al (1998) carried out a phylogenetic analysis basedon rDNA internal transcribed spacer 1 DNA sequencesof nine described species and one putative species of Heterorhabditis and they suggested that three pairs of sister taxa may be conspecic thereby delimiting six
species of Heterorhabditis The study of Adams et al
(1998) did not include H brevicaudis(Liu 1994)Current biogeographicdata suggests that two species of
Heterorhabditis H indica and H bacteriophora have aglobal distribution H indica occurs widely in the trop-ics and subtropics having been isolated in southern In-dia (Poinar et al 1992) Sri Lanka (Amarasinghe et al1994) peninsular Malaysia (Mason et al 1996) Indone-sia (Grifn et al 1999b) the Caribbean region (ArteagaHernandez amp MraacuteIumlcek 1984 Joyce et al 1994a Gre-nier et al 1996a Constant et al 1998) Egypt (Gre-
nier et al 1996a) and in subtropical and warm temperatezones in Japan (Yoshida et al 1998) Curran and Driver(1994) presented data for HaeIII restriction digests of therDNA intergenic spacer region of a range of tropical iso-lates of Heterorhabditis from north Australia Egypt theCaribbean region Florida USA and Hawaii They dis-tinguished between two restriction proles designated D1and D1a This distinctionbetween the D1 and D1a proleswas based on a size polymorphism of the large HaeIII re-striction fragment from the rDNA IGS fragment
H bacteriophora occurs in regions of continental and
Mediterranean climate in both the northern and south-ern hemispheres (reviewed by Hominick et al 1996)To date H megidis has been recorded only from thenorthern hemisphere (Poinar et al 1987 Smits et al1991 Miduturi et al 1996 Menti et al 1997 Yoshidaet al 1998 Grifn et al 1999a) where it typically hasa more northerly and more restricted distribution than H bacteriophora although H megidis can be locallycommon in coastal regions of North West Europe (Grif-n et al 1999a) The remaining described species of Heterorhabditis appear to have a more restricted distri-bution H zealandica has been isolated in New Zealandand Tasmania (Wouts 1979 Poinar 1990 Curran ampDriver 1994) H marelatus has been isolated so far onlyin Oregon and California USA (Liu amp Berry 1996Stock 1997) H brevicaudis has been isolated in southeast China (Liu 1994) and H argentinensis in Argentina(Stock 1993) although it is possible that H argentinen-
sis and H bacteriophora may be conspecic (Adams et
al 1998) Heterorhabditis species distributions can alsobe inuenced by altitude (Constant et al 1998) soil type
(Kaya 1990 Grifn et al 1994) and vegetation cov(Strong et al 1996)
We report here the isolation of new isolates of H indi
from India and Kenya We have utilised a range of moleular diagnostic techniquesin conjunctionwith crossbreeing in the identication of these new H indica isolat
and of isolates of H indica previously isolated from Idia Indonesiaand Cuba Our data indicatethat the Hae
rDNA IGS restriction prole designated D1a by Currand Driver(1994)also occursin the H indica typespeciLN2 and that the Kenyan isolates represent a distinct sugroup within the D1a group of H indica
Materials and methods
NEMATODE ISOLATES
H indica LN2 the H indica type species (Poinaral 1992) is maintained at the Sugarcane Breeding Instute (SBI) Coimbatore India It was originally isolatfrom soil samples collected at Ramanathapuram 20 knorth of Coimbatore Tamil Nadu Coimbatore (at 11deg
latitude and 77degE longitude) is 120 km from the Arabisea 270 km from the Indian ocean and 310 km from tBay of Bengal The soil samples(soil type red loam) wecollected from fallow land (the previous crop was mosoon sorghum) and were baited in the laboratory with tborer (Scirpophaga excerptalis Pyralidae Lepidopte
larvae LN2B mdash in situ baiting with S excerptalis larvwas carried out in a eld at SomayanurCoimbatorefrowhich groundnuts had been harvested The soil type wred loam The S excerptalis larvae were placed indiviually in small plastic lids covered with brass wire mesThe insecttraps were left in the soil for four days and wethen returned to the laboratory at SBI and checked for nmatode infection LN4 mdash this isolate was obtained froan infected white grub ( Holotrichia serrata Scarabadae Coleoptera) larva collected from a sugarcane eldThirupattur25 km north of CoimbatoreThe soil type wheavy clay
In collaboration with personnel from the Kenya Agcultural Research Institute (KARI) 21 soil samples eaof about 500 g were collected from ve locations coastal Kenya and two locations inland in August 199One sample was taken near Tambia village four from tWAUKARI centre at Mtwapa one from coral-based soat an elevated (8 m) coastal site near Mombasa 13 frovegetation fringing a beach at Kanamai one from far iland in the south-west near Lolgorien and one from ne
478 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 311
Isolates of Heterorhabditisindi
the Sand river in the same area as the last The sampleswere own to NUI Maynoothwhere each sample was di-vided and each half was baited with ve late instar Galle-
ria mellonella larvae and incubated at 28degC Heterorhab-
ditis isolates were recovered only at Kanamai in a veg-
etation belt extending 15 m inland from the beach edgeBioluminescent cadavers( indicating the presence of Het-
erorhabditis) were recovered from two of the 13 samplestaken from there
Four isolates were collected in a survey of ve Indone-sian islands by Grifn et al (1999b)
Extracted DNA samples from four isolates displayingthe D1a IGS restriction prole(FLGS10 JAM23JAM79ST09 ES10) were obtainedfrom Dr Felice Driver (CSIROCanberra Australia) The source and geographicorigin of the other Heterorhabditis species and isolates included in
this study are listed in Table 1
MOLECULAR CHARACTERISATION
DNA was isolated according to Smits et al (1991) Tinternal transcribed spacer (ITS) and intergenic spac(IGS) regions of the rDNA cistron were amplied bmeans of the polymerase chain reaction (PCR) as dscribed by Joyce et al (1994a b) Amplication proucts were digested with restriction endonucleases folloing the manufacturerrsquos instructions using 5-12 m l PCproduct in a 15 m l reaction volume The entire digewas loaded on a 2 agarose gel and electrophoresed 1 acute TBE at 5 Vcm for 35 h Restriction fragments wevisualised by ethidium bromide staining
The dot blot procedure utilised the H indica speciespecic satellite DNA probeused by Grenier et al (1996GenomicDNA(100ng)sampleswere denaturedbyadd1 M NaOH and 200 mM EDTA pH 82 to each samp
to give a nal concentration of 04 M NaOH 10 m
Table 1 Source and geographic origin of the Heterorhabditis species and isolates included in this study
Species Isolate Geographic Origin Source
H indica LN2 Coimbatore India Easwaramoorthy1
H indica LN2B Coimbatore India This study H indica LN4 Coimbatore India This study H indica Ki3 Kanami Kenya This study H indica K4A Kanami Kenya This study H indica INA H1 West Java Indonesia Grifn et al (1999 H indica INA H9 Ambon Indonesia Grifn et al (1999 H indica INA H17 Seram Indonesia Grifn et al (1999 H indica INA H23 Moluccas Indonesia Grifn et al (1999 H indica P2M Artemisia Cuba Mracek2
H indica D1 Darwin Australia Bedding3
H indica FLGS10 Florida USA Curran amp Driver3
H indica JAM23 Jamaica Curran amp Driver H indica JAM79 Jamaica Curran amp Driver H indica ST09 Virgin Islands Curran amp Driver H indica ES10 Egypt Curran amp Driver H bacteriophora HP88 Utah USA Akhurst3
H zealandica NZH3 New Zealand Bedding3
H marelatus OH-10 Oregon USA Liu4
H hepialus Bodega Bay California USA Stock5 H megidis HL81 Leeuwarden The Netherlands Westerman6
H lsquoIrish typersquo K122 Wexford Ireland Grifn et al (1994
1 Sugar Cane Breeding Institute Coimbatore India2 Institute of Entomology Czech Academy of Sciences Ceske BudejoviceCzech Republic3 CSIRO Canberra Australia4 Oregon State University OR USA5 University of California Davis CA USA6 Van Hall Instituut Leeuwarden The Netherlands
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 411
CM Stack et al
EDTA and the samples were then boiled for 10 minin a water bath The samples were then transferred byvacuum suction onto a positively charged nylon mem-brane (Amersham Life Sciences Ltd Amersham Buck-inghamshire UK) in a slot blot apparatus (Schleicheramp Schuell D37582 Dassel Germany) as per manufac-
turerrsquos instructions The DNA was then xed onto thenylon membrane by UV cross-linking using StratagenersquosStratalinker (Stratagene La Jolla CA 92037 USA) Therecombinant pUC plasmids Hi12 and HP88s9 which con-tained respectively the H indica and H bacteriophora
satellite DNA monomers were obtained from Dr PierreAbad INRA Antibes France and amplied using PCRThe PCR primers used to amplify the H indica monomerfrom the Hi12 plasmid were 5cent-CTGAAGCACTTGGGA-CAGAGC-3cent and 5cent-CTCCTCGTTGAGGACGGGAGT-3cent (Abadon et al 1998Grenier pers comm) The H bac-
teriophora monomer was amplied from the HP88s9plasmid using the following PCR primers 5cent-AGCTATG-CCAGAATGATCGCC-3cent and5cent-AGATTCTCTGTACG-ATGAGTA-3cent (Grenier et al 1996b Grenier perscomm) DNA was amplied using the following condi-tions one cycle of 94degC for 5 min was followed by 35cycles of denaturation at 94degC for 05 min annealing at52degC for 1 min and extension at 72degC for 2 min with anal cycle of extension at 72degC for 5 min
The probes were labelled using the ECL direct nucleicacid detection system (Amersham) following the man-ufacturerrsquos instructions Hybridisations were conducted
at 42degC overnight After hybridisation the lters werewashed rst with 05 acute SSC for 40 min at 42degC and thenwith 01acute SSC for 5 min at room temperatureAfter post-hybridisation washes lters were exposed to Hyperim-ECL (Amersham) following the manufacturerrsquos instruc-tions The membrane was then stripped and reprobed us-ing an 18S rDNA probe as a loading control The 18Sprobe was obtained by PCR from H indica LN2 genomicDNA using the primers 18SR2B 5cent-TACAAAGGGCAG-GGACGTATT-3cent and 18S12 5cent-GGCGATCAGATACC-GCCCTAGTT-3cent (TO Powers pers comm)
CROSS-BREEDING TESTS
Cross-breedingstudieswere carried outas describedbyDix et al (1992) with the following controls being set upfor each cross virginity test mdash 20 virgin females wereplaced on a lipid agar plate that had been inoculated andpre-incubated with the primary form of the LN2 bacteriaself-cross mdash ten virgin females and ten males of thesame isolate were placed on lipid agar plates containing
the bacterial symbiont The result of any cross betwedifferent isolates was taken as valid only if there were progeny in the virginity test and there were progeny in tself-cross At least ten second generation virgin femalwere used for each cross
Results
RESTRICTION PROFILES OF THE RDNA INTERNAL
TRANSCRIBER SPACER (ITS) REGION
All of the tropical isolates of Heterorhabditis yielda ca 1 kb fragment upon PCR amplication with the ITprimers These amplication products were digested wthe diagnostic restriction endonucleases used by Joy
et al (1994a) for species diagnosis in Heterorhabdi(viz AluI Hinf I and MboI) When digested with Al
(Fig 1) the Indian isolates LN2B and LN4 shared tsame restriction prole as the H indica LN2 type specias did the Indonesian isolates the P2M isolate froCuba and the Kenyan isolates When digested with Mbo
the Kenyan isolates shared the same restriction pro
Fig 1 Alu I restriction digests of the PCR amplication produ
of the rDNA internal transcribed spacer region of Heterorhditis isolates separated on a 2 agarose gel and stained w
ethidium bromide M 1 kb marker 1 H indica LN2 2 P2
3 LN2B 4 LN4 5 INA H23 6 INA H9 7 INA H17 8 I
H1 9 Ki3 10 K4A 11 HeterorhabditislsquoIrish typersquo K122 1
H megidis HL81
480 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 511
Isolates of Heterorhabditisindi
as H indica LN2 and P2M (Fig 2) The Indonesianisolates also displayed the same MboI restriction proleas H indica LN2 (data not shown) When digested with Hinf I (Fig 3) the Indian Indonesian and Cuban isolatesbut not the Kenyan isolates possessed the same restrictionprole as H indica LN2 The Kenyan isolates have only
a single Hinf I restriction site in the ITS rDNA fragmentyielding two restriction fragments of ca 620 and 450 bpunlikeall the other H indica isolates testedwhich possesstwo Hinf I sites in this region yielding three restrictionfragments of ca 450 360 and 240 bp
Fig 2 Mbo I restriction digests of the PCR amplication prod-
ucts of the rDNA internal transcribed spacer region of Het-erorhabditis isolates separated on a 2 agarose gel and stained
with ethidium bromide M 1 kb marker 1 H indica LN22 P2M 3 Ki3 4 K4A 5 H bacteriophora HP88
RESTRICTION PROFILES OF THE RDNA INTERGENIC
SPACER (IGS) REGION
The PCR amplication products for the IGS rDNA rgion of the tropical isolates varied in size from 15 to 17 kb The amplied IGS rDNA fragments were r
stricted with the endonucleases AluI and HaeIII A dianostic AluI restriction pattern was obtained for H indi
LN2 and this was shared by all the Indian IndonesiaCuban and Kenyan isolates (Fig 4) and the D1 and Dtype isolates(Fig 6) When HaeIII was used to digest tIGS rDNA region H indica LN2 displayed a distinctive fragment prole of 740 280 190 160 and 120 (Fig 5) and with the exception of the Kenyan isolatefour of these fragments were shared by the other tropcal isolates The Kenyan isolates Ki3 (lane 9) and K4(lane 10) had a distinct restriction prole yielding Hae
fragments of 700 280 (a doublet) 185 and 120 bp Tlargest H indica HaeIII fragment was highlypolymorphin size between the tropical isolates ranging in size fro700 to 850 bp with some isolates having fragments of itermediate size ca 740 bp Size polymorphismin thisframent was the basis on which the types D1 and D1a werecognised by Curran and Driver (1994) The HaeIII prles of the other three species of Heterorhabditis includin Fig 5 were all distinctly different from that of H i
dica As can be seen from Fig 6 the HaeIII rDNA IGrestriction prole designated D1a by Curran and Driv
(1994)(lanes 4-7) is similar to that of H indica LN2 bthe HaeIII pattern of the Kenyan isolates (Fig 6 laneFig 5 lanes 910) is unique
Fig 3 Hinf I restriction digests of the PCR amplication products of the rDNA int ernal transcribed spacer region of Heterorhabdisolates separated on a 2 agarose gel and stained with ethidium bromide M 1 kb marker 1 LN2 2 P2M 3 LN2B 4 LN
5 INA H23 6 INA H9 7 INA H17 8 INA H1 9 Ki3 10 K4A 11HeterorhabditislsquoIrish typersquo K122 12 H megidis HL81
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 611
CM Stack et al
Fig 4 Alu I restriction digests of the PCR amplication products of the r DNA intergenic spacer region of Heterorhabditis isolat
separated on a 2 agarose gel and stained with ethidium bromide M 1kb size marker 1 LN2 2 P2M 3 LN2B 4 LN5 INA H23 6 INA H9 7 INA H17 8 INA H1 9 Ki3 10 K4A 11 Heterorhabditis lsquoIrish typersquo K122 12 H megidis HL8
13 H bacteriophora HP88
Fig 5 Hae III restriction digests of the PCR amplication products of the rDNA intergenic spacer region of Heterorhabditis isolat
separated on a 2 agarose gel and stained with ethidiumbromide M 1kb size marker 1 LN2 2 P2M 3 LN2B 4 LN4 5 INA H
6 INA H9 7 INA H9 8 INA H1 9 Ki3 10 K4A 11HeterorhabditislsquoIrish typersquo K122 12 H megidis HL81 13 H bacterioph HP88
USE OF H INDICA SPECIES S PECIFIC SATELLITE DNAPROBE
When Southern blots of total genomic DNA of thetropical isolates were probed with the H indica species-specic satellite DNA probe described by Abadon et al
(1998) the probe hybridised only with the isolates clas-sied from PCR analysis as being H indica including
the Kenyan isolates Ki3 and K4A (Fig 7A) No hybrisation was detected by this probe to any of the othspecies included as controls on the Southern blot Thblot was then reprobed with the H bacteriophora specisatellite DNA probe described by Grenier et al (199and this probe hybridised only with the HP88 isolate H bacteriophora included on the Southern blot (data nshown)
482 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 711
Isolates of Heterorhabditisindi
Fig 6 Hae III and Alu I restriction digests of the PCR amplication products of the rDNA intergenic spacer region of Heterorhabdisolates including isolates designated type D1a by Curran and Driver (1994) M 1kb size marker 1 H indica LN2 2 D1 3 K4
4 ES10 5 JAM79 6 ST09 7 FLGS10 M 1 kb marker 9 H indica LN2 10 D1 11 ES10 12 JAM79 13 ST09 14 FLGS1
Lanes 1 to 7 contain Hae III digests and lanes 9 to 14 contain Alu I digests The digests were separated on a 2 agarose gel and stain
with ethidium bromide
Table 2 The results of cross-breeding experiments of six Het-erorhabditis isolates from India Indonesia and Kenya
Female Male
H indica LN 4 INA H1 INA H23 Ki3 K4ALN2India India Indonesia Indonesia Kenya Kenya
H indica + + + 0 + 0LN2
LN 4 + + 0 0 + +INA H1 + + + + + 0
INA H23 0 0 + + 0 0Ki3 + + + 0 + +K4A 0 0 0 0 + +
+ cross resulted in fertile progeny0 no progeny detected
CROSS BREEDING ANALYSIS
Selectedcross-breedingresults are presented in Table2All the Indian isolates are interfertile The Kenyan iso-
lates which differed from H indica in their Hinf I ITSrestriction prole and their HaeIII IGS restriction proleare interfertile amongst each other but only one Kenyanisolate Ki3 produced viable progeny when crossed withthe H indica LN2 type species The Indonesian isolatesare interfertile but surprisingly only one of the Indone-sian isolates tested (INA H1) produced viable hybridswhen crossed with H indica LN2 The INA H1 isolatewas also interfertile with the Kenyan isolate Ki3
Discussion
The results presented here conrm and extend prevous studies which show that a combination of molecudiagnostictoolscan be reliably used for species identiction in Heterorhabditis Isolates of Heterorhabditis froIndia Kenya Indonesia and Cuba hybridised to the H i
dica specic satellite DNA probe (Grenier et al 199Abadon et al 1998) H indica specic DNA restrictiproles were also obtained for all the tropical isolat
of Heterorhabditis investigated here when the ITS rDNregion was restricted using the diagnostic restriction ezymes AluI and MboI (Joyce et al 1994a) The utilof restriction digests of the rDNA ITS region in specidiagnosis of EPN has been conrmed in several studi(Joyce et al 1994a b Miduturi et al 1996 Hominickal 1997 Yoshida et al 1998 Grifn et al 1999a Pa
jav et al 1999) Restriction of the rDNA IGS region wi AluI also yielded a H indica specic restriction prole fall the tropical isolates tested in this study
The PCR amplication product which we obtained fthe H indica rDNA IGS varied in size from 12 to 16 kHominick et al (1997) also observed length heterogenity in the rDNA IGS region among geographic isolatof single Steinernema spp and they suggested that thheterogeneity may make restriction proles of this DNfragment unreliable for species identication The lengheterogeneity which we detected in the H indica IGPCR amplication product did not affect the AluI restrtion prole of the product and a clear-cut seven restrictifragment pattern was common to all the H indica isolat
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 811
CM Stack et al
Fig 7 A Slot blot analysis of genomic DNA (100 ng) of the
Heterorhabditis isolates using a H indica specic satellite DNA
probe B The membrane used for Fig 7A was stripped and
reprobed using an rDNA 18S probe as a loading control A1
H indica LN2 A2 D1 A3 P2M A4 LN2B A5 LN4 A6
INA H23 B1 INA H9 B2 INA H17 B3 INA H1 B4 Ki3
B5 K4A B6 H bacteriophora HP88 C1 H zealandica NZH3
C2 H marelatus OH-10 C3 H hepialus Bodega Bay C4
Heterorhabditis lsquoIrish typersquo K122
tested When the IGS region was digested with HaeIII H indica LN2 displayed a distinctive ve fragment pro-le All of the H indica isolates with the exception of the Kenyan isolates also displayed a ve fragment pro-le Four of these fragments were conserved among theisolates but the largest fragment (of ca 740 in H indica
LN2) was polymorphicamong the H indica isolates Thisfragment which seems to be associated with the lengthpolymorphism in the H indica IGS region is the basisof the D1D1a polymorphism observed by Curran and
Driver (1994) In most species studied to date the rDNIGS contains tandem arrays of subrepeats (Gerbi 198Williams et al 1990 Vahidi amp Honda 1991 Novakal 1993 Linares et al 1994 Crease 1995) This rsults in the length of the IGS region being variable bobetween and within species The length heterogeneity
the H indica IGS PCR product did not affect the AluI rstriction prole of this fragment which suggests that Al
has a restriction site within a tandem repetitive unit in trDNA IGS of H indica In contrast HaeIII appears nto cut within this tandem repeat region with the result thone of the HaeIII restriction fragments is polymorphiclength presumably depending on the number of AluI rpeat units which it contains
The HaeIII rDNA IGS restriction prole of the Disolate is similar to that of H indica LN2 but the Hae
pattern of the Kenyan isolates is unique The large Hae
fragment of the Kenyan isolate K4A is the same size that of D1a thus the Kenyan isolates appear to represea distinct subgroup within D1a Tropical isolates froDarwin (North Australia) Puerto Rico Hawaii the VirgIslands and Egypt were found by Curran and Driv(1994) to possess the D1 prole while the D1a prowas described by these authors from isolates collectin Puerto Rico Jamaica Florida the Virgin Islands aEgypt Comparison of the D1 and D1a HaeIII and Al
rDNA IGS restriction proles with those of H indi
LN2 indicates that the isolates designated D1 and D
by Curran and Driver (1994) belong to H indicaAn important step towards achieving an effective nmatode bacterium complex for pest control is to seek nurally occurring endemic EPN isolates as such isolatare likely to possess physiological traits that are adaptto local climatic and ecological conditions The Kenyisolates described in this study clearly belong to a distinsub-group of H indica and it is probable that these islates also share a distinctive phenotype The two Kenyisolates are interfertile but only one of these isolates (Kiproduced viable progeny when crossed with the H indi
LN2 type species All of the Indonesian Heterorhabdi
isolates were indistinguishable from H indica in the dagnostic molecular tests and they were interfertile wieach other but only one of the Indonesian isolates (INH1) was interfertile with H indica LN2 and INA H1 walso interfertile with the Ki3 isolate from Kenya Theresults suggest that although H indica has a global dtribution in tropical and subtropical regions of the worgeneow within the species may be quite restricted Whave previously observedreproductiveincompatibilityb
484 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 911
Isolates of Heterorhabditisindi
tween isolates of H bacteriophora from Europe (Grifnet al 1999a) and from North America (Dix et al un-publ) We have not determined whether this reproductiveisolation is caused by genetic means or is the result of cy-toplasmic factors Cytoplasmic factors such as endosym-biont bacteria (see reviews by Werren 1997 Johanowicz
amp Hoy 1998) and chromosomal factors such as trans-posons (reviewed by Kidwell 1990 Petrov et al 1995)have been shown to be frequent causes of reproductivein-compatibility in arthropods Endosymbiont bacteria haverecently been shown to cause reproductiveincompatibilityin larial nematodes (Hoerauf et al 1999) Since the re-productive incompatibilityobserved in this study does notyield clear groups of compatible and incompatiblestrainsand in view of the consistencyof the DNA based diagnos-tic tests any assignment of biological species within the H indica group would be injudicious at present Adams
(1988) evaluates phylogeneticand biologicalspecies con-cepts in the delimitationof species in the H bacteriophora
groupThe global distribution of H indica throughout the
tropics and subtropics suggests that H indica possessesa range of phenotypic characters which give it an advan-tage in this climatic zone Such phenotypes might includeadaptationto high temperatures desiccation tolerance andgood dispersal ability It is unlikely that the widespreaddistribution of H indica in the tropics arises because thisspecies was extant before the continents began breakingup and drifting The best supported hypothesis of phylo-genetic relationships among Heterorhabditis species in aphylogeny derived from the rDNA ITS 1 DNA sequencedata puts H indica as the most ancient Heterorhabditis
lineage (Adams et al 1998) However the phylogenetictree for Heterorhabditis and Steinernema of Reid (1994)shows that the genetic distance between Heterorhabdi-
tis species is considerably less than that between Stein-
ernema species and given the small genetic distance de-tected by Reid (1994) between the most divergent of the Heterorhabditis species in his study (a D value of ca
004 as calculated by the method of Nei and Li 1979) it
seems unlikely that H indica speciated before the Juras-sicCretaceousbreak up of Pangaea(an estimated 200 mil-lion years ago)
Acknowledgement
We acknowledge the support of the European Commu-nity STD-3 Programme (TS3 CT 940273)
References
ABADON M GRENIER E LAUMOND C amp ABAD (1998) A species-specic satellite DNA from the enmopathogenic nematode Heterorhabditis indicus Geno
41 148-153
ADAMS BJ (1998) Species concepts and the evolutionparadigm in modern nematology Journal of Nematology 31-21
ADAMS BJ BURNELL AM amp POWERS TO (1998)phylogenetic analysis of the genus Heterorhabditis(NemaRhabditidae) based on internal transcribed spacer 1 DNsequence data Journal of Nematology 30 22-39
AMARASINGHE LD HOMINICK WM BRISCOE Bamp REI D AP (1994) Occurrence and distribution entomopathogenic nematodes in Sri Lanka Journal
Helminthology 68 277-286ARTEAGA HERNANDEZ E M amp MRAacute IumlCE K Z (1984) H
terorhabditis heliothidis a parasite of insect pests in CubFolia Parasitologia(Praha) 31 11-17
BLAXTER ML DE LE Y P GAREY J R LIU LXSCHELDEMAN P VIERSTRAETE A VANFLETERE
JR MACKEY LY DORRIS M FRISSE LM VID
JT amp THOMAS WK (1998) A molecular evolutionaframework for the phylum Nematoda Nature 392 71-75
CONSTANT P MARCHAY L F ISHER-LE SAU X MBRIAND-PANOMA S amp MAULEacuteON H (1998)Naturalocurrence of entomopathogenic nematodes( RhabditidaSteernematidae and Heterorhabditidae) in Guadeloupe islanFundamental and Applied Nematology 21 667-672
CREASE TJ (1995) Ribosomal DNA evolution at tpopulation-level mdash nucleotide variation in intergenic spaarrays of Daphnia pulex Genetics 141 1327-1337
CURRAN J amp DRIVER F (1994) Molecular taxonomy of Hterorhabditis In Burnell AM Ehlers R-U amp Masson J(Eds) Genetics of Entomopathogenic Nematode-Bacteri
Complexes Luxembourg European Commission PublicatiEUR 15681 EN pp 178-187
DIX I BURNELL A M GRIFFIN CT JOYCE SAamp DOWNES MJ (1992) The identication of biologispecies in the genus Heterorhabditis (Nematoda Heterhabditae) by cross-breeding second generation amphimicadults Parasitology104 509-518
EHLERS R-U (1996) Current and future use of nematodin biocontrolpractice and commercial aspects with regardregulatory policy issues Biocontrol Science and Technolo
6 303-317GERBI SA (1985) Evolution of ribosomal DNA In MacI
tyre RJ (Ed) Molecular evolutionary genetics New YoNY USA Plenum Press pp 419-517
GRENIER E BONIFASSI E ABAD P amp LAUMOND (1996a) Use of species specic satellite DNAs as dianostic probes in the identication of Steinernematidae a
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1011
CM Stack et al
Heterorhabditidae entomopathogenic nematodes Parasitol-
ogy 113 483-489GRENIER E LAUMOND C amp ABAD P (1996b) Molecu-
lar characterisationof two species-specic tandemly repeatedDNAs from entomopathogenic nematodes Steinernema and
Heterorhabditis(NematodaRhabditida) Molecularand Bio-
chemical Parasitology 83 47-56GRIFFIN CT JOYCE S A DIX I BURNELL AM amp
DOWNES MJ (1994)Characterisationof the entomopatho-genic nematode Heterorhabditis (Nematoda Heterorhabdi-tidae) from Ireland and Britain by molecular and cross-breeding techniques and the occurrence of the genus in theseislands Fundamental and Applied Nematology 17 245-253
GRIFFIN CT DIX I JOYCE SA BURNELL AM ampDOWNES MJ (1999a) Isolation and characterisation of
Heterorhabditis spp from Hungary Estonia and Denmark Nematology 1 321-332
GRIFFIN CT CHAERANI R FALLON D REI D AP
amp DOWNES MJ (1999b) Occurrence and distribution of the entomopathogenic nematodes Steinernema spp and Het-
erorhabditis indica in Indonesia Journal of Helminthology
74 143-150HOERAUF A NISSENPAHLE K SCHMETZ C HEN-
KLEDUHRSEN K BLAXTER ML BUTTNER DWGALLIN MY ALQAOUD KM LUCIUS R amp FLEIS-CHER B (1999) Tetracycline therapy targets intracellularbacteria in the larial nematode Litomososoides sigmodontis
and results in larial infertility Journal of Clinical Investiga-
tion 103 11-17HOMINICK WM REID AP BOHAN DA amp BRISCOE
BR (1996) Entomopathogenicnematodes biodiversity ge-ographicaldistributionand conventionon biological diversity
Biocontrol Science and Technology 6 317-332HOMINICK WM BRISCOE BR DE L PIN O FG HEN G
J HUN T DJ KOZODOY E MRAacute IumlCE K Z NGUYENKB REI D AP SPIRIDONOV S STOCK P STURHAN D WATURU C amp YOSHIDA M (1997) Biosystematics of entomopathogenic nematodes current status protocols anddenitions Journal of Helminthology 71 271-298
JOHANOWICZ D L amp HOY MA (1998) The manipula-tion of arthropodreproductionby Wolbachia endosymbiontsFlorida Entomologist 81 310-317
JOYCE S A BURNELL A M amp P OWERS TO (1994a)The characterisationof Heterorhabditisisolatesby restrictionenzyme analysis of PCR amplied fragments of mtDNA andrDNA genes Journal of Nematology 26 260-270
JOYCE SA REI D A amp CURRAN J (1994b) Applicationof the polymerase chain reaction (PCR) methods to the iden-tication of entomopathogenicnematodes In Burnell AMEhlers R-U amp Masson JP (Eds) Genetics of entomopatho-
genic nematode-bacterium complexes Luxembourg Euro-pean Commission Publication EUR 15681 EN pp 178-187
KAYA H K (1990)Soil ecology In Gaugler R amp Kaya H(Eds) Entomopathogenic nematodes in biological contr
Boca Raton FL USA CRC Press pp 93-115KAYA HK amp GAUGLER R (1993) Entomopathogenic n
matodes Annual Review of Entomology 38 181-206KIDWELL M G (1990) Evolutionaryaspects of hybrid dysg
nesis in Drosophila Canadian Journal of Zoology 68 1711726LINARES AR BOWEN T amp DOVER GA (1994) A
pects of non-random turnover involved in the concerted evlution of intergenic spacers within the ribosomal DNA
Drosophila melanogaster Journal of Molecular Evoluti
39 151-159LIU J (1994) [A new species of the genus Heterorhabd
from China (Rhabditida Heterorhabditidae)] Acta Zoota
nomica Sinica 19 268-272LIU J amp BERRY RE (1996) Heterorhabditis marela
nsp (Rhabditida Heterorhabditidae) from Oregon Journ
of Invertebrate Pathology
67 48-54MASON JM RAZAK AR amp WRIGHT DJ (1996) Trecoveryof entomopathogenicnematodes from selectedarewithin Peninsular Malaysia Journal of Helminthology 7303-307
MENTI H WRIGHT DJ amp PERRY RN (1997)Desiccatsurvival of populations of the entomopathogenic nematodSteinernema feltiaeand Heterorhabditismegidis from Greeand the UK Journal of Helminthology 71 41-46
MIDUTURI JS MOENS M HOMINICK WM BRISCO
B R amp RE ID AP (1996) Naturally occurring enmopathogenic nematodes in the province of West-FlandeBelgium Journal of Helminthology 70 319-327
NE I M amp LI W-H (1979) Mathematical model for studygenetic variation in terms of restriction endonucleases P
ceedings of the National Academy of Sciences of the Uni
States 76 5269-5273NOVAK EM DE MELLO MP GOMES HBM
GALINDO I GUEVARA P RAMIREZ JL amp DA SILV
RI A JF (1993) Repetitive sequences in the ribosomal intgenic spacer of Trypanosoma cruzi Molecular and Bioche
ical Parasitology 60 273-280PAMJAV H TRIGA D BUZAS Z VELLAI T LUCSKA
A ADAMS B REI D AP BURNELL AM GRIFF
C GLAZER I KLEIN MG amp FODOR A (1999) Nov
application of PhastSystem polyacrylamide gel electrophosis using restriction fragment length polymorphism-intertranscribed spacer patterns of individuals for molecular idetication of entomopathogenic nematodes Electrophore
201266-1273PETROV DA SCHUTZMAN J L HARTL D L amp L
ZOVSKAYA ER ( 1995) Diverse transposable elements amobilised in hybrid dysgenesis in Drosophila virilis P
ceedings of the National Academy of Sciences of the Uni
States 92 8050-8054
486 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1111
Isolates of Heterorhabditisindi
POINAR GO JR (1990) Taxonomy and biology of Stein-ernematidae and HeterorhabditidaeIn Gaugler R amp KayaHK (Eds) Entomopathogenic nematodes in biological con-
trol Boca Raton FL USA CRC Press pp 23-64POINAR GO JR (1993) Origins and phylogenetic relation-
ships of the entomophilic rhabditids Heterorhabditis and
Steinernema Fundamental and Applied Nematology16 333-338POINAR GO JR JACKSON T amp KLEIN M (1987) He-
terorhabditis megidis sp n (Heterorhabditidae Rhabditida)parasitic in the Japanese beetlePopilliajaponica(ScarabidaeColeoptera) in Ohio Proceedings of the Helminthological
Society of Washington 53 53-59POINAR GO JR KARUNAKAR GK amp DAVID H (1992)
Heterorhabditis indicus n sp (Rhabditida Nematoda) fromIndia Separation of Heterorhabditis spp by infective juve-niles Fundamental and Applied Nematology 15 467-472
REI D AP (1994) Molecular taxonomy of Steinernema InBurnell AM Ehlers R-U amp Masson JP (Eds) Genetics
of entomopathogenic nematode-bacterium complexes Lux-embourg European Commission Publication EUR 15681EN pp 178-187
SMITS P GROENEN JTM amp DERAAY G (1991) Charac-terisation of Heterorhabditis isolates using DNA restrictionlength polymorphism Revue de Neacutematologie 14 445-453
STOCK SP (1993) A new species of thegenus Heterorhabdi-
tis Poinar 1976 (Nematoda Heterorhabditidae) parasitizingGraphognathus sp larvae (Coteoptera Curculionidae) fromArgentina Research and Reviews in Parasitology 53 103-107
STOCK SP (1997) Heterorhabditis hepialus Stock Strong amp
Gardner 1996a junior synonym of H marelatus Liu amp Berry1996 (Rhabditida Rhabditidae) with a redescription of thespecies Nematologica 43 455-463
STOCK SP STRONG D amp GARDNER S (1996) Identiction of Heterorhabditis (Nematoda Heterorhabditidae)froCalifornia with a new species isolated from the larvae of tghost moth Hepialius [sic] californicus (Lepidoptera Healidae) from Bodega Bay Natural Reserve Fundamental a
Applied Nematology 19 585-592
STRONG DR KAYA HK WHIPPLE AV CHILD ALKRAIG S BONDONNE M DYE R K amp MARON J(1996) Entomopathogenic nematodes natural enemiesroot feeding caterpillarson bush lupine Oecologica 108 345
SUDHAUS W (1993) Die mittels symbiontischerBakterienetomopathogenen Nematoden Gattungen Heterorhabditis aSteinernema sind keine Schwestertaxa Verhandlungen d
Deutschen Zoologischen Gesellschaft 86 146VAHIDI H amp HONDA BM (1991)Repeats and subrepeats
the intergenic spacer of rDNA from the nematode Meloid
gyne arenaria Molecular and General Genetics 227 33
336WERREN JH (1997) Biology of Wolbachia Annual Revi
of Entomology 42 587-609WILLIAMS S M ROBBINS L G CLUSTER PD A
LARD RW amp STROBECK C (1990) Superstructureof t Drosophila ribosomal gene family Proceedings of the N
tional Academy of Sciences of the United States 87 3153160
WOUTS WM (1979) The biology and life cycle ofNew Zealand population of Heterorhabditis heliothidis (Hterorhabditidae)Nematologica 25 191-202
YOSHIDA M REID AP BRISCOE BR amp HOMINIC
WM (1998) Survey of entomopathogenic nematod(Rhabditida Steinernematidae and Heterorhabditidae) Japan Fundamental and Applied Nematology 21 185-198
Vol 2(5) 2000 4

862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 211
CM Stack et al
species are recognised by Hominick et al (1997) Adamset al (1998) carried out a phylogenetic analysis basedon rDNA internal transcribed spacer 1 DNA sequencesof nine described species and one putative species of Heterorhabditis and they suggested that three pairs of sister taxa may be conspecic thereby delimiting six
species of Heterorhabditis The study of Adams et al
(1998) did not include H brevicaudis(Liu 1994)Current biogeographicdata suggests that two species of
Heterorhabditis H indica and H bacteriophora have aglobal distribution H indica occurs widely in the trop-ics and subtropics having been isolated in southern In-dia (Poinar et al 1992) Sri Lanka (Amarasinghe et al1994) peninsular Malaysia (Mason et al 1996) Indone-sia (Grifn et al 1999b) the Caribbean region (ArteagaHernandez amp MraacuteIumlcek 1984 Joyce et al 1994a Gre-nier et al 1996a Constant et al 1998) Egypt (Gre-
nier et al 1996a) and in subtropical and warm temperatezones in Japan (Yoshida et al 1998) Curran and Driver(1994) presented data for HaeIII restriction digests of therDNA intergenic spacer region of a range of tropical iso-lates of Heterorhabditis from north Australia Egypt theCaribbean region Florida USA and Hawaii They dis-tinguished between two restriction proles designated D1and D1a This distinctionbetween the D1 and D1a proleswas based on a size polymorphism of the large HaeIII re-striction fragment from the rDNA IGS fragment
H bacteriophora occurs in regions of continental and
Mediterranean climate in both the northern and south-ern hemispheres (reviewed by Hominick et al 1996)To date H megidis has been recorded only from thenorthern hemisphere (Poinar et al 1987 Smits et al1991 Miduturi et al 1996 Menti et al 1997 Yoshidaet al 1998 Grifn et al 1999a) where it typically hasa more northerly and more restricted distribution than H bacteriophora although H megidis can be locallycommon in coastal regions of North West Europe (Grif-n et al 1999a) The remaining described species of Heterorhabditis appear to have a more restricted distri-bution H zealandica has been isolated in New Zealandand Tasmania (Wouts 1979 Poinar 1990 Curran ampDriver 1994) H marelatus has been isolated so far onlyin Oregon and California USA (Liu amp Berry 1996Stock 1997) H brevicaudis has been isolated in southeast China (Liu 1994) and H argentinensis in Argentina(Stock 1993) although it is possible that H argentinen-
sis and H bacteriophora may be conspecic (Adams et
al 1998) Heterorhabditis species distributions can alsobe inuenced by altitude (Constant et al 1998) soil type
(Kaya 1990 Grifn et al 1994) and vegetation cov(Strong et al 1996)
We report here the isolation of new isolates of H indi
from India and Kenya We have utilised a range of moleular diagnostic techniquesin conjunctionwith crossbreeing in the identication of these new H indica isolat
and of isolates of H indica previously isolated from Idia Indonesiaand Cuba Our data indicatethat the Hae
rDNA IGS restriction prole designated D1a by Currand Driver(1994)also occursin the H indica typespeciLN2 and that the Kenyan isolates represent a distinct sugroup within the D1a group of H indica
Materials and methods
NEMATODE ISOLATES
H indica LN2 the H indica type species (Poinaral 1992) is maintained at the Sugarcane Breeding Instute (SBI) Coimbatore India It was originally isolatfrom soil samples collected at Ramanathapuram 20 knorth of Coimbatore Tamil Nadu Coimbatore (at 11deg
latitude and 77degE longitude) is 120 km from the Arabisea 270 km from the Indian ocean and 310 km from tBay of Bengal The soil samples(soil type red loam) wecollected from fallow land (the previous crop was mosoon sorghum) and were baited in the laboratory with tborer (Scirpophaga excerptalis Pyralidae Lepidopte
larvae LN2B mdash in situ baiting with S excerptalis larvwas carried out in a eld at SomayanurCoimbatorefrowhich groundnuts had been harvested The soil type wred loam The S excerptalis larvae were placed indiviually in small plastic lids covered with brass wire mesThe insecttraps were left in the soil for four days and wethen returned to the laboratory at SBI and checked for nmatode infection LN4 mdash this isolate was obtained froan infected white grub ( Holotrichia serrata Scarabadae Coleoptera) larva collected from a sugarcane eldThirupattur25 km north of CoimbatoreThe soil type wheavy clay
In collaboration with personnel from the Kenya Agcultural Research Institute (KARI) 21 soil samples eaof about 500 g were collected from ve locations coastal Kenya and two locations inland in August 199One sample was taken near Tambia village four from tWAUKARI centre at Mtwapa one from coral-based soat an elevated (8 m) coastal site near Mombasa 13 frovegetation fringing a beach at Kanamai one from far iland in the south-west near Lolgorien and one from ne
478 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 311
Isolates of Heterorhabditisindi
the Sand river in the same area as the last The sampleswere own to NUI Maynoothwhere each sample was di-vided and each half was baited with ve late instar Galle-
ria mellonella larvae and incubated at 28degC Heterorhab-
ditis isolates were recovered only at Kanamai in a veg-
etation belt extending 15 m inland from the beach edgeBioluminescent cadavers( indicating the presence of Het-
erorhabditis) were recovered from two of the 13 samplestaken from there
Four isolates were collected in a survey of ve Indone-sian islands by Grifn et al (1999b)
Extracted DNA samples from four isolates displayingthe D1a IGS restriction prole(FLGS10 JAM23JAM79ST09 ES10) were obtainedfrom Dr Felice Driver (CSIROCanberra Australia) The source and geographicorigin of the other Heterorhabditis species and isolates included in
this study are listed in Table 1
MOLECULAR CHARACTERISATION
DNA was isolated according to Smits et al (1991) Tinternal transcribed spacer (ITS) and intergenic spac(IGS) regions of the rDNA cistron were amplied bmeans of the polymerase chain reaction (PCR) as dscribed by Joyce et al (1994a b) Amplication proucts were digested with restriction endonucleases folloing the manufacturerrsquos instructions using 5-12 m l PCproduct in a 15 m l reaction volume The entire digewas loaded on a 2 agarose gel and electrophoresed 1 acute TBE at 5 Vcm for 35 h Restriction fragments wevisualised by ethidium bromide staining
The dot blot procedure utilised the H indica speciespecic satellite DNA probeused by Grenier et al (1996GenomicDNA(100ng)sampleswere denaturedbyadd1 M NaOH and 200 mM EDTA pH 82 to each samp
to give a nal concentration of 04 M NaOH 10 m
Table 1 Source and geographic origin of the Heterorhabditis species and isolates included in this study
Species Isolate Geographic Origin Source
H indica LN2 Coimbatore India Easwaramoorthy1
H indica LN2B Coimbatore India This study H indica LN4 Coimbatore India This study H indica Ki3 Kanami Kenya This study H indica K4A Kanami Kenya This study H indica INA H1 West Java Indonesia Grifn et al (1999 H indica INA H9 Ambon Indonesia Grifn et al (1999 H indica INA H17 Seram Indonesia Grifn et al (1999 H indica INA H23 Moluccas Indonesia Grifn et al (1999 H indica P2M Artemisia Cuba Mracek2
H indica D1 Darwin Australia Bedding3
H indica FLGS10 Florida USA Curran amp Driver3
H indica JAM23 Jamaica Curran amp Driver H indica JAM79 Jamaica Curran amp Driver H indica ST09 Virgin Islands Curran amp Driver H indica ES10 Egypt Curran amp Driver H bacteriophora HP88 Utah USA Akhurst3
H zealandica NZH3 New Zealand Bedding3
H marelatus OH-10 Oregon USA Liu4
H hepialus Bodega Bay California USA Stock5 H megidis HL81 Leeuwarden The Netherlands Westerman6
H lsquoIrish typersquo K122 Wexford Ireland Grifn et al (1994
1 Sugar Cane Breeding Institute Coimbatore India2 Institute of Entomology Czech Academy of Sciences Ceske BudejoviceCzech Republic3 CSIRO Canberra Australia4 Oregon State University OR USA5 University of California Davis CA USA6 Van Hall Instituut Leeuwarden The Netherlands
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 411
CM Stack et al
EDTA and the samples were then boiled for 10 minin a water bath The samples were then transferred byvacuum suction onto a positively charged nylon mem-brane (Amersham Life Sciences Ltd Amersham Buck-inghamshire UK) in a slot blot apparatus (Schleicheramp Schuell D37582 Dassel Germany) as per manufac-
turerrsquos instructions The DNA was then xed onto thenylon membrane by UV cross-linking using StratagenersquosStratalinker (Stratagene La Jolla CA 92037 USA) Therecombinant pUC plasmids Hi12 and HP88s9 which con-tained respectively the H indica and H bacteriophora
satellite DNA monomers were obtained from Dr PierreAbad INRA Antibes France and amplied using PCRThe PCR primers used to amplify the H indica monomerfrom the Hi12 plasmid were 5cent-CTGAAGCACTTGGGA-CAGAGC-3cent and 5cent-CTCCTCGTTGAGGACGGGAGT-3cent (Abadon et al 1998Grenier pers comm) The H bac-
teriophora monomer was amplied from the HP88s9plasmid using the following PCR primers 5cent-AGCTATG-CCAGAATGATCGCC-3cent and5cent-AGATTCTCTGTACG-ATGAGTA-3cent (Grenier et al 1996b Grenier perscomm) DNA was amplied using the following condi-tions one cycle of 94degC for 5 min was followed by 35cycles of denaturation at 94degC for 05 min annealing at52degC for 1 min and extension at 72degC for 2 min with anal cycle of extension at 72degC for 5 min
The probes were labelled using the ECL direct nucleicacid detection system (Amersham) following the man-ufacturerrsquos instructions Hybridisations were conducted
at 42degC overnight After hybridisation the lters werewashed rst with 05 acute SSC for 40 min at 42degC and thenwith 01acute SSC for 5 min at room temperatureAfter post-hybridisation washes lters were exposed to Hyperim-ECL (Amersham) following the manufacturerrsquos instruc-tions The membrane was then stripped and reprobed us-ing an 18S rDNA probe as a loading control The 18Sprobe was obtained by PCR from H indica LN2 genomicDNA using the primers 18SR2B 5cent-TACAAAGGGCAG-GGACGTATT-3cent and 18S12 5cent-GGCGATCAGATACC-GCCCTAGTT-3cent (TO Powers pers comm)
CROSS-BREEDING TESTS
Cross-breedingstudieswere carried outas describedbyDix et al (1992) with the following controls being set upfor each cross virginity test mdash 20 virgin females wereplaced on a lipid agar plate that had been inoculated andpre-incubated with the primary form of the LN2 bacteriaself-cross mdash ten virgin females and ten males of thesame isolate were placed on lipid agar plates containing
the bacterial symbiont The result of any cross betwedifferent isolates was taken as valid only if there were progeny in the virginity test and there were progeny in tself-cross At least ten second generation virgin femalwere used for each cross
Results
RESTRICTION PROFILES OF THE RDNA INTERNAL
TRANSCRIBER SPACER (ITS) REGION
All of the tropical isolates of Heterorhabditis yielda ca 1 kb fragment upon PCR amplication with the ITprimers These amplication products were digested wthe diagnostic restriction endonucleases used by Joy
et al (1994a) for species diagnosis in Heterorhabdi(viz AluI Hinf I and MboI) When digested with Al
(Fig 1) the Indian isolates LN2B and LN4 shared tsame restriction prole as the H indica LN2 type specias did the Indonesian isolates the P2M isolate froCuba and the Kenyan isolates When digested with Mbo
the Kenyan isolates shared the same restriction pro
Fig 1 Alu I restriction digests of the PCR amplication produ
of the rDNA internal transcribed spacer region of Heterorhditis isolates separated on a 2 agarose gel and stained w
ethidium bromide M 1 kb marker 1 H indica LN2 2 P2
3 LN2B 4 LN4 5 INA H23 6 INA H9 7 INA H17 8 I
H1 9 Ki3 10 K4A 11 HeterorhabditislsquoIrish typersquo K122 1
H megidis HL81
480 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 511
Isolates of Heterorhabditisindi
as H indica LN2 and P2M (Fig 2) The Indonesianisolates also displayed the same MboI restriction proleas H indica LN2 (data not shown) When digested with Hinf I (Fig 3) the Indian Indonesian and Cuban isolatesbut not the Kenyan isolates possessed the same restrictionprole as H indica LN2 The Kenyan isolates have only
a single Hinf I restriction site in the ITS rDNA fragmentyielding two restriction fragments of ca 620 and 450 bpunlikeall the other H indica isolates testedwhich possesstwo Hinf I sites in this region yielding three restrictionfragments of ca 450 360 and 240 bp
Fig 2 Mbo I restriction digests of the PCR amplication prod-
ucts of the rDNA internal transcribed spacer region of Het-erorhabditis isolates separated on a 2 agarose gel and stained
with ethidium bromide M 1 kb marker 1 H indica LN22 P2M 3 Ki3 4 K4A 5 H bacteriophora HP88
RESTRICTION PROFILES OF THE RDNA INTERGENIC
SPACER (IGS) REGION
The PCR amplication products for the IGS rDNA rgion of the tropical isolates varied in size from 15 to 17 kb The amplied IGS rDNA fragments were r
stricted with the endonucleases AluI and HaeIII A dianostic AluI restriction pattern was obtained for H indi
LN2 and this was shared by all the Indian IndonesiaCuban and Kenyan isolates (Fig 4) and the D1 and Dtype isolates(Fig 6) When HaeIII was used to digest tIGS rDNA region H indica LN2 displayed a distinctive fragment prole of 740 280 190 160 and 120 (Fig 5) and with the exception of the Kenyan isolatefour of these fragments were shared by the other tropcal isolates The Kenyan isolates Ki3 (lane 9) and K4(lane 10) had a distinct restriction prole yielding Hae
fragments of 700 280 (a doublet) 185 and 120 bp Tlargest H indica HaeIII fragment was highlypolymorphin size between the tropical isolates ranging in size fro700 to 850 bp with some isolates having fragments of itermediate size ca 740 bp Size polymorphismin thisframent was the basis on which the types D1 and D1a werecognised by Curran and Driver (1994) The HaeIII prles of the other three species of Heterorhabditis includin Fig 5 were all distinctly different from that of H i
dica As can be seen from Fig 6 the HaeIII rDNA IGrestriction prole designated D1a by Curran and Driv
(1994)(lanes 4-7) is similar to that of H indica LN2 bthe HaeIII pattern of the Kenyan isolates (Fig 6 laneFig 5 lanes 910) is unique
Fig 3 Hinf I restriction digests of the PCR amplication products of the rDNA int ernal transcribed spacer region of Heterorhabdisolates separated on a 2 agarose gel and stained with ethidium bromide M 1 kb marker 1 LN2 2 P2M 3 LN2B 4 LN
5 INA H23 6 INA H9 7 INA H17 8 INA H1 9 Ki3 10 K4A 11HeterorhabditislsquoIrish typersquo K122 12 H megidis HL81
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 611
CM Stack et al
Fig 4 Alu I restriction digests of the PCR amplication products of the r DNA intergenic spacer region of Heterorhabditis isolat
separated on a 2 agarose gel and stained with ethidium bromide M 1kb size marker 1 LN2 2 P2M 3 LN2B 4 LN5 INA H23 6 INA H9 7 INA H17 8 INA H1 9 Ki3 10 K4A 11 Heterorhabditis lsquoIrish typersquo K122 12 H megidis HL8
13 H bacteriophora HP88
Fig 5 Hae III restriction digests of the PCR amplication products of the rDNA intergenic spacer region of Heterorhabditis isolat
separated on a 2 agarose gel and stained with ethidiumbromide M 1kb size marker 1 LN2 2 P2M 3 LN2B 4 LN4 5 INA H
6 INA H9 7 INA H9 8 INA H1 9 Ki3 10 K4A 11HeterorhabditislsquoIrish typersquo K122 12 H megidis HL81 13 H bacterioph HP88
USE OF H INDICA SPECIES S PECIFIC SATELLITE DNAPROBE
When Southern blots of total genomic DNA of thetropical isolates were probed with the H indica species-specic satellite DNA probe described by Abadon et al
(1998) the probe hybridised only with the isolates clas-sied from PCR analysis as being H indica including
the Kenyan isolates Ki3 and K4A (Fig 7A) No hybrisation was detected by this probe to any of the othspecies included as controls on the Southern blot Thblot was then reprobed with the H bacteriophora specisatellite DNA probe described by Grenier et al (199and this probe hybridised only with the HP88 isolate H bacteriophora included on the Southern blot (data nshown)
482 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 711
Isolates of Heterorhabditisindi
Fig 6 Hae III and Alu I restriction digests of the PCR amplication products of the rDNA intergenic spacer region of Heterorhabdisolates including isolates designated type D1a by Curran and Driver (1994) M 1kb size marker 1 H indica LN2 2 D1 3 K4
4 ES10 5 JAM79 6 ST09 7 FLGS10 M 1 kb marker 9 H indica LN2 10 D1 11 ES10 12 JAM79 13 ST09 14 FLGS1
Lanes 1 to 7 contain Hae III digests and lanes 9 to 14 contain Alu I digests The digests were separated on a 2 agarose gel and stain
with ethidium bromide
Table 2 The results of cross-breeding experiments of six Het-erorhabditis isolates from India Indonesia and Kenya
Female Male
H indica LN 4 INA H1 INA H23 Ki3 K4ALN2India India Indonesia Indonesia Kenya Kenya
H indica + + + 0 + 0LN2
LN 4 + + 0 0 + +INA H1 + + + + + 0
INA H23 0 0 + + 0 0Ki3 + + + 0 + +K4A 0 0 0 0 + +
+ cross resulted in fertile progeny0 no progeny detected
CROSS BREEDING ANALYSIS
Selectedcross-breedingresults are presented in Table2All the Indian isolates are interfertile The Kenyan iso-
lates which differed from H indica in their Hinf I ITSrestriction prole and their HaeIII IGS restriction proleare interfertile amongst each other but only one Kenyanisolate Ki3 produced viable progeny when crossed withthe H indica LN2 type species The Indonesian isolatesare interfertile but surprisingly only one of the Indone-sian isolates tested (INA H1) produced viable hybridswhen crossed with H indica LN2 The INA H1 isolatewas also interfertile with the Kenyan isolate Ki3
Discussion
The results presented here conrm and extend prevous studies which show that a combination of molecudiagnostictoolscan be reliably used for species identiction in Heterorhabditis Isolates of Heterorhabditis froIndia Kenya Indonesia and Cuba hybridised to the H i
dica specic satellite DNA probe (Grenier et al 199Abadon et al 1998) H indica specic DNA restrictiproles were also obtained for all the tropical isolat
of Heterorhabditis investigated here when the ITS rDNregion was restricted using the diagnostic restriction ezymes AluI and MboI (Joyce et al 1994a) The utilof restriction digests of the rDNA ITS region in specidiagnosis of EPN has been conrmed in several studi(Joyce et al 1994a b Miduturi et al 1996 Hominickal 1997 Yoshida et al 1998 Grifn et al 1999a Pa
jav et al 1999) Restriction of the rDNA IGS region wi AluI also yielded a H indica specic restriction prole fall the tropical isolates tested in this study
The PCR amplication product which we obtained fthe H indica rDNA IGS varied in size from 12 to 16 kHominick et al (1997) also observed length heterogenity in the rDNA IGS region among geographic isolatof single Steinernema spp and they suggested that thheterogeneity may make restriction proles of this DNfragment unreliable for species identication The lengheterogeneity which we detected in the H indica IGPCR amplication product did not affect the AluI restrtion prole of the product and a clear-cut seven restrictifragment pattern was common to all the H indica isolat
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 811
CM Stack et al
Fig 7 A Slot blot analysis of genomic DNA (100 ng) of the
Heterorhabditis isolates using a H indica specic satellite DNA
probe B The membrane used for Fig 7A was stripped and
reprobed using an rDNA 18S probe as a loading control A1
H indica LN2 A2 D1 A3 P2M A4 LN2B A5 LN4 A6
INA H23 B1 INA H9 B2 INA H17 B3 INA H1 B4 Ki3
B5 K4A B6 H bacteriophora HP88 C1 H zealandica NZH3
C2 H marelatus OH-10 C3 H hepialus Bodega Bay C4
Heterorhabditis lsquoIrish typersquo K122
tested When the IGS region was digested with HaeIII H indica LN2 displayed a distinctive ve fragment pro-le All of the H indica isolates with the exception of the Kenyan isolates also displayed a ve fragment pro-le Four of these fragments were conserved among theisolates but the largest fragment (of ca 740 in H indica
LN2) was polymorphicamong the H indica isolates Thisfragment which seems to be associated with the lengthpolymorphism in the H indica IGS region is the basisof the D1D1a polymorphism observed by Curran and
Driver (1994) In most species studied to date the rDNIGS contains tandem arrays of subrepeats (Gerbi 198Williams et al 1990 Vahidi amp Honda 1991 Novakal 1993 Linares et al 1994 Crease 1995) This rsults in the length of the IGS region being variable bobetween and within species The length heterogeneity
the H indica IGS PCR product did not affect the AluI rstriction prole of this fragment which suggests that Al
has a restriction site within a tandem repetitive unit in trDNA IGS of H indica In contrast HaeIII appears nto cut within this tandem repeat region with the result thone of the HaeIII restriction fragments is polymorphiclength presumably depending on the number of AluI rpeat units which it contains
The HaeIII rDNA IGS restriction prole of the Disolate is similar to that of H indica LN2 but the Hae
pattern of the Kenyan isolates is unique The large Hae
fragment of the Kenyan isolate K4A is the same size that of D1a thus the Kenyan isolates appear to represea distinct subgroup within D1a Tropical isolates froDarwin (North Australia) Puerto Rico Hawaii the VirgIslands and Egypt were found by Curran and Driv(1994) to possess the D1 prole while the D1a prowas described by these authors from isolates collectin Puerto Rico Jamaica Florida the Virgin Islands aEgypt Comparison of the D1 and D1a HaeIII and Al
rDNA IGS restriction proles with those of H indi
LN2 indicates that the isolates designated D1 and D
by Curran and Driver (1994) belong to H indicaAn important step towards achieving an effective nmatode bacterium complex for pest control is to seek nurally occurring endemic EPN isolates as such isolatare likely to possess physiological traits that are adaptto local climatic and ecological conditions The Kenyisolates described in this study clearly belong to a distinsub-group of H indica and it is probable that these islates also share a distinctive phenotype The two Kenyisolates are interfertile but only one of these isolates (Kiproduced viable progeny when crossed with the H indi
LN2 type species All of the Indonesian Heterorhabdi
isolates were indistinguishable from H indica in the dagnostic molecular tests and they were interfertile wieach other but only one of the Indonesian isolates (INH1) was interfertile with H indica LN2 and INA H1 walso interfertile with the Ki3 isolate from Kenya Theresults suggest that although H indica has a global dtribution in tropical and subtropical regions of the worgeneow within the species may be quite restricted Whave previously observedreproductiveincompatibilityb
484 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 911
Isolates of Heterorhabditisindi
tween isolates of H bacteriophora from Europe (Grifnet al 1999a) and from North America (Dix et al un-publ) We have not determined whether this reproductiveisolation is caused by genetic means or is the result of cy-toplasmic factors Cytoplasmic factors such as endosym-biont bacteria (see reviews by Werren 1997 Johanowicz
amp Hoy 1998) and chromosomal factors such as trans-posons (reviewed by Kidwell 1990 Petrov et al 1995)have been shown to be frequent causes of reproductivein-compatibility in arthropods Endosymbiont bacteria haverecently been shown to cause reproductiveincompatibilityin larial nematodes (Hoerauf et al 1999) Since the re-productive incompatibilityobserved in this study does notyield clear groups of compatible and incompatiblestrainsand in view of the consistencyof the DNA based diagnos-tic tests any assignment of biological species within the H indica group would be injudicious at present Adams
(1988) evaluates phylogeneticand biologicalspecies con-cepts in the delimitationof species in the H bacteriophora
groupThe global distribution of H indica throughout the
tropics and subtropics suggests that H indica possessesa range of phenotypic characters which give it an advan-tage in this climatic zone Such phenotypes might includeadaptationto high temperatures desiccation tolerance andgood dispersal ability It is unlikely that the widespreaddistribution of H indica in the tropics arises because thisspecies was extant before the continents began breakingup and drifting The best supported hypothesis of phylo-genetic relationships among Heterorhabditis species in aphylogeny derived from the rDNA ITS 1 DNA sequencedata puts H indica as the most ancient Heterorhabditis
lineage (Adams et al 1998) However the phylogenetictree for Heterorhabditis and Steinernema of Reid (1994)shows that the genetic distance between Heterorhabdi-
tis species is considerably less than that between Stein-
ernema species and given the small genetic distance de-tected by Reid (1994) between the most divergent of the Heterorhabditis species in his study (a D value of ca
004 as calculated by the method of Nei and Li 1979) it
seems unlikely that H indica speciated before the Juras-sicCretaceousbreak up of Pangaea(an estimated 200 mil-lion years ago)
Acknowledgement
We acknowledge the support of the European Commu-nity STD-3 Programme (TS3 CT 940273)
References
ABADON M GRENIER E LAUMOND C amp ABAD (1998) A species-specic satellite DNA from the enmopathogenic nematode Heterorhabditis indicus Geno
41 148-153
ADAMS BJ (1998) Species concepts and the evolutionparadigm in modern nematology Journal of Nematology 31-21
ADAMS BJ BURNELL AM amp POWERS TO (1998)phylogenetic analysis of the genus Heterorhabditis(NemaRhabditidae) based on internal transcribed spacer 1 DNsequence data Journal of Nematology 30 22-39
AMARASINGHE LD HOMINICK WM BRISCOE Bamp REI D AP (1994) Occurrence and distribution entomopathogenic nematodes in Sri Lanka Journal
Helminthology 68 277-286ARTEAGA HERNANDEZ E M amp MRAacute IumlCE K Z (1984) H
terorhabditis heliothidis a parasite of insect pests in CubFolia Parasitologia(Praha) 31 11-17
BLAXTER ML DE LE Y P GAREY J R LIU LXSCHELDEMAN P VIERSTRAETE A VANFLETERE
JR MACKEY LY DORRIS M FRISSE LM VID
JT amp THOMAS WK (1998) A molecular evolutionaframework for the phylum Nematoda Nature 392 71-75
CONSTANT P MARCHAY L F ISHER-LE SAU X MBRIAND-PANOMA S amp MAULEacuteON H (1998)Naturalocurrence of entomopathogenic nematodes( RhabditidaSteernematidae and Heterorhabditidae) in Guadeloupe islanFundamental and Applied Nematology 21 667-672
CREASE TJ (1995) Ribosomal DNA evolution at tpopulation-level mdash nucleotide variation in intergenic spaarrays of Daphnia pulex Genetics 141 1327-1337
CURRAN J amp DRIVER F (1994) Molecular taxonomy of Hterorhabditis In Burnell AM Ehlers R-U amp Masson J(Eds) Genetics of Entomopathogenic Nematode-Bacteri
Complexes Luxembourg European Commission PublicatiEUR 15681 EN pp 178-187
DIX I BURNELL A M GRIFFIN CT JOYCE SAamp DOWNES MJ (1992) The identication of biologispecies in the genus Heterorhabditis (Nematoda Heterhabditae) by cross-breeding second generation amphimicadults Parasitology104 509-518
EHLERS R-U (1996) Current and future use of nematodin biocontrolpractice and commercial aspects with regardregulatory policy issues Biocontrol Science and Technolo
6 303-317GERBI SA (1985) Evolution of ribosomal DNA In MacI
tyre RJ (Ed) Molecular evolutionary genetics New YoNY USA Plenum Press pp 419-517
GRENIER E BONIFASSI E ABAD P amp LAUMOND (1996a) Use of species specic satellite DNAs as dianostic probes in the identication of Steinernematidae a
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1011
CM Stack et al
Heterorhabditidae entomopathogenic nematodes Parasitol-
ogy 113 483-489GRENIER E LAUMOND C amp ABAD P (1996b) Molecu-
lar characterisationof two species-specic tandemly repeatedDNAs from entomopathogenic nematodes Steinernema and
Heterorhabditis(NematodaRhabditida) Molecularand Bio-
chemical Parasitology 83 47-56GRIFFIN CT JOYCE S A DIX I BURNELL AM amp
DOWNES MJ (1994)Characterisationof the entomopatho-genic nematode Heterorhabditis (Nematoda Heterorhabdi-tidae) from Ireland and Britain by molecular and cross-breeding techniques and the occurrence of the genus in theseislands Fundamental and Applied Nematology 17 245-253
GRIFFIN CT DIX I JOYCE SA BURNELL AM ampDOWNES MJ (1999a) Isolation and characterisation of
Heterorhabditis spp from Hungary Estonia and Denmark Nematology 1 321-332
GRIFFIN CT CHAERANI R FALLON D REI D AP
amp DOWNES MJ (1999b) Occurrence and distribution of the entomopathogenic nematodes Steinernema spp and Het-
erorhabditis indica in Indonesia Journal of Helminthology
74 143-150HOERAUF A NISSENPAHLE K SCHMETZ C HEN-
KLEDUHRSEN K BLAXTER ML BUTTNER DWGALLIN MY ALQAOUD KM LUCIUS R amp FLEIS-CHER B (1999) Tetracycline therapy targets intracellularbacteria in the larial nematode Litomososoides sigmodontis
and results in larial infertility Journal of Clinical Investiga-
tion 103 11-17HOMINICK WM REID AP BOHAN DA amp BRISCOE
BR (1996) Entomopathogenicnematodes biodiversity ge-ographicaldistributionand conventionon biological diversity
Biocontrol Science and Technology 6 317-332HOMINICK WM BRISCOE BR DE L PIN O FG HEN G
J HUN T DJ KOZODOY E MRAacute IumlCE K Z NGUYENKB REI D AP SPIRIDONOV S STOCK P STURHAN D WATURU C amp YOSHIDA M (1997) Biosystematics of entomopathogenic nematodes current status protocols anddenitions Journal of Helminthology 71 271-298
JOHANOWICZ D L amp HOY MA (1998) The manipula-tion of arthropodreproductionby Wolbachia endosymbiontsFlorida Entomologist 81 310-317
JOYCE S A BURNELL A M amp P OWERS TO (1994a)The characterisationof Heterorhabditisisolatesby restrictionenzyme analysis of PCR amplied fragments of mtDNA andrDNA genes Journal of Nematology 26 260-270
JOYCE SA REI D A amp CURRAN J (1994b) Applicationof the polymerase chain reaction (PCR) methods to the iden-tication of entomopathogenicnematodes In Burnell AMEhlers R-U amp Masson JP (Eds) Genetics of entomopatho-
genic nematode-bacterium complexes Luxembourg Euro-pean Commission Publication EUR 15681 EN pp 178-187
KAYA H K (1990)Soil ecology In Gaugler R amp Kaya H(Eds) Entomopathogenic nematodes in biological contr
Boca Raton FL USA CRC Press pp 93-115KAYA HK amp GAUGLER R (1993) Entomopathogenic n
matodes Annual Review of Entomology 38 181-206KIDWELL M G (1990) Evolutionaryaspects of hybrid dysg
nesis in Drosophila Canadian Journal of Zoology 68 1711726LINARES AR BOWEN T amp DOVER GA (1994) A
pects of non-random turnover involved in the concerted evlution of intergenic spacers within the ribosomal DNA
Drosophila melanogaster Journal of Molecular Evoluti
39 151-159LIU J (1994) [A new species of the genus Heterorhabd
from China (Rhabditida Heterorhabditidae)] Acta Zoota
nomica Sinica 19 268-272LIU J amp BERRY RE (1996) Heterorhabditis marela
nsp (Rhabditida Heterorhabditidae) from Oregon Journ
of Invertebrate Pathology
67 48-54MASON JM RAZAK AR amp WRIGHT DJ (1996) Trecoveryof entomopathogenicnematodes from selectedarewithin Peninsular Malaysia Journal of Helminthology 7303-307
MENTI H WRIGHT DJ amp PERRY RN (1997)Desiccatsurvival of populations of the entomopathogenic nematodSteinernema feltiaeand Heterorhabditismegidis from Greeand the UK Journal of Helminthology 71 41-46
MIDUTURI JS MOENS M HOMINICK WM BRISCO
B R amp RE ID AP (1996) Naturally occurring enmopathogenic nematodes in the province of West-FlandeBelgium Journal of Helminthology 70 319-327
NE I M amp LI W-H (1979) Mathematical model for studygenetic variation in terms of restriction endonucleases P
ceedings of the National Academy of Sciences of the Uni
States 76 5269-5273NOVAK EM DE MELLO MP GOMES HBM
GALINDO I GUEVARA P RAMIREZ JL amp DA SILV
RI A JF (1993) Repetitive sequences in the ribosomal intgenic spacer of Trypanosoma cruzi Molecular and Bioche
ical Parasitology 60 273-280PAMJAV H TRIGA D BUZAS Z VELLAI T LUCSKA
A ADAMS B REI D AP BURNELL AM GRIFF
C GLAZER I KLEIN MG amp FODOR A (1999) Nov
application of PhastSystem polyacrylamide gel electrophosis using restriction fragment length polymorphism-intertranscribed spacer patterns of individuals for molecular idetication of entomopathogenic nematodes Electrophore
201266-1273PETROV DA SCHUTZMAN J L HARTL D L amp L
ZOVSKAYA ER ( 1995) Diverse transposable elements amobilised in hybrid dysgenesis in Drosophila virilis P
ceedings of the National Academy of Sciences of the Uni
States 92 8050-8054
486 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1111
Isolates of Heterorhabditisindi
POINAR GO JR (1990) Taxonomy and biology of Stein-ernematidae and HeterorhabditidaeIn Gaugler R amp KayaHK (Eds) Entomopathogenic nematodes in biological con-
trol Boca Raton FL USA CRC Press pp 23-64POINAR GO JR (1993) Origins and phylogenetic relation-
ships of the entomophilic rhabditids Heterorhabditis and
Steinernema Fundamental and Applied Nematology16 333-338POINAR GO JR JACKSON T amp KLEIN M (1987) He-
terorhabditis megidis sp n (Heterorhabditidae Rhabditida)parasitic in the Japanese beetlePopilliajaponica(ScarabidaeColeoptera) in Ohio Proceedings of the Helminthological
Society of Washington 53 53-59POINAR GO JR KARUNAKAR GK amp DAVID H (1992)
Heterorhabditis indicus n sp (Rhabditida Nematoda) fromIndia Separation of Heterorhabditis spp by infective juve-niles Fundamental and Applied Nematology 15 467-472
REI D AP (1994) Molecular taxonomy of Steinernema InBurnell AM Ehlers R-U amp Masson JP (Eds) Genetics
of entomopathogenic nematode-bacterium complexes Lux-embourg European Commission Publication EUR 15681EN pp 178-187
SMITS P GROENEN JTM amp DERAAY G (1991) Charac-terisation of Heterorhabditis isolates using DNA restrictionlength polymorphism Revue de Neacutematologie 14 445-453
STOCK SP (1993) A new species of thegenus Heterorhabdi-
tis Poinar 1976 (Nematoda Heterorhabditidae) parasitizingGraphognathus sp larvae (Coteoptera Curculionidae) fromArgentina Research and Reviews in Parasitology 53 103-107
STOCK SP (1997) Heterorhabditis hepialus Stock Strong amp
Gardner 1996a junior synonym of H marelatus Liu amp Berry1996 (Rhabditida Rhabditidae) with a redescription of thespecies Nematologica 43 455-463
STOCK SP STRONG D amp GARDNER S (1996) Identiction of Heterorhabditis (Nematoda Heterorhabditidae)froCalifornia with a new species isolated from the larvae of tghost moth Hepialius [sic] californicus (Lepidoptera Healidae) from Bodega Bay Natural Reserve Fundamental a
Applied Nematology 19 585-592
STRONG DR KAYA HK WHIPPLE AV CHILD ALKRAIG S BONDONNE M DYE R K amp MARON J(1996) Entomopathogenic nematodes natural enemiesroot feeding caterpillarson bush lupine Oecologica 108 345
SUDHAUS W (1993) Die mittels symbiontischerBakterienetomopathogenen Nematoden Gattungen Heterorhabditis aSteinernema sind keine Schwestertaxa Verhandlungen d
Deutschen Zoologischen Gesellschaft 86 146VAHIDI H amp HONDA BM (1991)Repeats and subrepeats
the intergenic spacer of rDNA from the nematode Meloid
gyne arenaria Molecular and General Genetics 227 33
336WERREN JH (1997) Biology of Wolbachia Annual Revi
of Entomology 42 587-609WILLIAMS S M ROBBINS L G CLUSTER PD A
LARD RW amp STROBECK C (1990) Superstructureof t Drosophila ribosomal gene family Proceedings of the N
tional Academy of Sciences of the United States 87 3153160
WOUTS WM (1979) The biology and life cycle ofNew Zealand population of Heterorhabditis heliothidis (Hterorhabditidae)Nematologica 25 191-202
YOSHIDA M REID AP BRISCOE BR amp HOMINIC
WM (1998) Survey of entomopathogenic nematod(Rhabditida Steinernematidae and Heterorhabditidae) Japan Fundamental and Applied Nematology 21 185-198
Vol 2(5) 2000 4

862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 311
Isolates of Heterorhabditisindi
the Sand river in the same area as the last The sampleswere own to NUI Maynoothwhere each sample was di-vided and each half was baited with ve late instar Galle-
ria mellonella larvae and incubated at 28degC Heterorhab-
ditis isolates were recovered only at Kanamai in a veg-
etation belt extending 15 m inland from the beach edgeBioluminescent cadavers( indicating the presence of Het-
erorhabditis) were recovered from two of the 13 samplestaken from there
Four isolates were collected in a survey of ve Indone-sian islands by Grifn et al (1999b)
Extracted DNA samples from four isolates displayingthe D1a IGS restriction prole(FLGS10 JAM23JAM79ST09 ES10) were obtainedfrom Dr Felice Driver (CSIROCanberra Australia) The source and geographicorigin of the other Heterorhabditis species and isolates included in
this study are listed in Table 1
MOLECULAR CHARACTERISATION
DNA was isolated according to Smits et al (1991) Tinternal transcribed spacer (ITS) and intergenic spac(IGS) regions of the rDNA cistron were amplied bmeans of the polymerase chain reaction (PCR) as dscribed by Joyce et al (1994a b) Amplication proucts were digested with restriction endonucleases folloing the manufacturerrsquos instructions using 5-12 m l PCproduct in a 15 m l reaction volume The entire digewas loaded on a 2 agarose gel and electrophoresed 1 acute TBE at 5 Vcm for 35 h Restriction fragments wevisualised by ethidium bromide staining
The dot blot procedure utilised the H indica speciespecic satellite DNA probeused by Grenier et al (1996GenomicDNA(100ng)sampleswere denaturedbyadd1 M NaOH and 200 mM EDTA pH 82 to each samp
to give a nal concentration of 04 M NaOH 10 m
Table 1 Source and geographic origin of the Heterorhabditis species and isolates included in this study
Species Isolate Geographic Origin Source
H indica LN2 Coimbatore India Easwaramoorthy1
H indica LN2B Coimbatore India This study H indica LN4 Coimbatore India This study H indica Ki3 Kanami Kenya This study H indica K4A Kanami Kenya This study H indica INA H1 West Java Indonesia Grifn et al (1999 H indica INA H9 Ambon Indonesia Grifn et al (1999 H indica INA H17 Seram Indonesia Grifn et al (1999 H indica INA H23 Moluccas Indonesia Grifn et al (1999 H indica P2M Artemisia Cuba Mracek2
H indica D1 Darwin Australia Bedding3
H indica FLGS10 Florida USA Curran amp Driver3
H indica JAM23 Jamaica Curran amp Driver H indica JAM79 Jamaica Curran amp Driver H indica ST09 Virgin Islands Curran amp Driver H indica ES10 Egypt Curran amp Driver H bacteriophora HP88 Utah USA Akhurst3
H zealandica NZH3 New Zealand Bedding3
H marelatus OH-10 Oregon USA Liu4
H hepialus Bodega Bay California USA Stock5 H megidis HL81 Leeuwarden The Netherlands Westerman6
H lsquoIrish typersquo K122 Wexford Ireland Grifn et al (1994
1 Sugar Cane Breeding Institute Coimbatore India2 Institute of Entomology Czech Academy of Sciences Ceske BudejoviceCzech Republic3 CSIRO Canberra Australia4 Oregon State University OR USA5 University of California Davis CA USA6 Van Hall Instituut Leeuwarden The Netherlands
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 411
CM Stack et al
EDTA and the samples were then boiled for 10 minin a water bath The samples were then transferred byvacuum suction onto a positively charged nylon mem-brane (Amersham Life Sciences Ltd Amersham Buck-inghamshire UK) in a slot blot apparatus (Schleicheramp Schuell D37582 Dassel Germany) as per manufac-
turerrsquos instructions The DNA was then xed onto thenylon membrane by UV cross-linking using StratagenersquosStratalinker (Stratagene La Jolla CA 92037 USA) Therecombinant pUC plasmids Hi12 and HP88s9 which con-tained respectively the H indica and H bacteriophora
satellite DNA monomers were obtained from Dr PierreAbad INRA Antibes France and amplied using PCRThe PCR primers used to amplify the H indica monomerfrom the Hi12 plasmid were 5cent-CTGAAGCACTTGGGA-CAGAGC-3cent and 5cent-CTCCTCGTTGAGGACGGGAGT-3cent (Abadon et al 1998Grenier pers comm) The H bac-
teriophora monomer was amplied from the HP88s9plasmid using the following PCR primers 5cent-AGCTATG-CCAGAATGATCGCC-3cent and5cent-AGATTCTCTGTACG-ATGAGTA-3cent (Grenier et al 1996b Grenier perscomm) DNA was amplied using the following condi-tions one cycle of 94degC for 5 min was followed by 35cycles of denaturation at 94degC for 05 min annealing at52degC for 1 min and extension at 72degC for 2 min with anal cycle of extension at 72degC for 5 min
The probes were labelled using the ECL direct nucleicacid detection system (Amersham) following the man-ufacturerrsquos instructions Hybridisations were conducted
at 42degC overnight After hybridisation the lters werewashed rst with 05 acute SSC for 40 min at 42degC and thenwith 01acute SSC for 5 min at room temperatureAfter post-hybridisation washes lters were exposed to Hyperim-ECL (Amersham) following the manufacturerrsquos instruc-tions The membrane was then stripped and reprobed us-ing an 18S rDNA probe as a loading control The 18Sprobe was obtained by PCR from H indica LN2 genomicDNA using the primers 18SR2B 5cent-TACAAAGGGCAG-GGACGTATT-3cent and 18S12 5cent-GGCGATCAGATACC-GCCCTAGTT-3cent (TO Powers pers comm)
CROSS-BREEDING TESTS
Cross-breedingstudieswere carried outas describedbyDix et al (1992) with the following controls being set upfor each cross virginity test mdash 20 virgin females wereplaced on a lipid agar plate that had been inoculated andpre-incubated with the primary form of the LN2 bacteriaself-cross mdash ten virgin females and ten males of thesame isolate were placed on lipid agar plates containing
the bacterial symbiont The result of any cross betwedifferent isolates was taken as valid only if there were progeny in the virginity test and there were progeny in tself-cross At least ten second generation virgin femalwere used for each cross
Results
RESTRICTION PROFILES OF THE RDNA INTERNAL
TRANSCRIBER SPACER (ITS) REGION
All of the tropical isolates of Heterorhabditis yielda ca 1 kb fragment upon PCR amplication with the ITprimers These amplication products were digested wthe diagnostic restriction endonucleases used by Joy
et al (1994a) for species diagnosis in Heterorhabdi(viz AluI Hinf I and MboI) When digested with Al
(Fig 1) the Indian isolates LN2B and LN4 shared tsame restriction prole as the H indica LN2 type specias did the Indonesian isolates the P2M isolate froCuba and the Kenyan isolates When digested with Mbo
the Kenyan isolates shared the same restriction pro
Fig 1 Alu I restriction digests of the PCR amplication produ
of the rDNA internal transcribed spacer region of Heterorhditis isolates separated on a 2 agarose gel and stained w
ethidium bromide M 1 kb marker 1 H indica LN2 2 P2
3 LN2B 4 LN4 5 INA H23 6 INA H9 7 INA H17 8 I
H1 9 Ki3 10 K4A 11 HeterorhabditislsquoIrish typersquo K122 1
H megidis HL81
480 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 511
Isolates of Heterorhabditisindi
as H indica LN2 and P2M (Fig 2) The Indonesianisolates also displayed the same MboI restriction proleas H indica LN2 (data not shown) When digested with Hinf I (Fig 3) the Indian Indonesian and Cuban isolatesbut not the Kenyan isolates possessed the same restrictionprole as H indica LN2 The Kenyan isolates have only
a single Hinf I restriction site in the ITS rDNA fragmentyielding two restriction fragments of ca 620 and 450 bpunlikeall the other H indica isolates testedwhich possesstwo Hinf I sites in this region yielding three restrictionfragments of ca 450 360 and 240 bp
Fig 2 Mbo I restriction digests of the PCR amplication prod-
ucts of the rDNA internal transcribed spacer region of Het-erorhabditis isolates separated on a 2 agarose gel and stained
with ethidium bromide M 1 kb marker 1 H indica LN22 P2M 3 Ki3 4 K4A 5 H bacteriophora HP88
RESTRICTION PROFILES OF THE RDNA INTERGENIC
SPACER (IGS) REGION
The PCR amplication products for the IGS rDNA rgion of the tropical isolates varied in size from 15 to 17 kb The amplied IGS rDNA fragments were r
stricted with the endonucleases AluI and HaeIII A dianostic AluI restriction pattern was obtained for H indi
LN2 and this was shared by all the Indian IndonesiaCuban and Kenyan isolates (Fig 4) and the D1 and Dtype isolates(Fig 6) When HaeIII was used to digest tIGS rDNA region H indica LN2 displayed a distinctive fragment prole of 740 280 190 160 and 120 (Fig 5) and with the exception of the Kenyan isolatefour of these fragments were shared by the other tropcal isolates The Kenyan isolates Ki3 (lane 9) and K4(lane 10) had a distinct restriction prole yielding Hae
fragments of 700 280 (a doublet) 185 and 120 bp Tlargest H indica HaeIII fragment was highlypolymorphin size between the tropical isolates ranging in size fro700 to 850 bp with some isolates having fragments of itermediate size ca 740 bp Size polymorphismin thisframent was the basis on which the types D1 and D1a werecognised by Curran and Driver (1994) The HaeIII prles of the other three species of Heterorhabditis includin Fig 5 were all distinctly different from that of H i
dica As can be seen from Fig 6 the HaeIII rDNA IGrestriction prole designated D1a by Curran and Driv
(1994)(lanes 4-7) is similar to that of H indica LN2 bthe HaeIII pattern of the Kenyan isolates (Fig 6 laneFig 5 lanes 910) is unique
Fig 3 Hinf I restriction digests of the PCR amplication products of the rDNA int ernal transcribed spacer region of Heterorhabdisolates separated on a 2 agarose gel and stained with ethidium bromide M 1 kb marker 1 LN2 2 P2M 3 LN2B 4 LN
5 INA H23 6 INA H9 7 INA H17 8 INA H1 9 Ki3 10 K4A 11HeterorhabditislsquoIrish typersquo K122 12 H megidis HL81
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 611
CM Stack et al
Fig 4 Alu I restriction digests of the PCR amplication products of the r DNA intergenic spacer region of Heterorhabditis isolat
separated on a 2 agarose gel and stained with ethidium bromide M 1kb size marker 1 LN2 2 P2M 3 LN2B 4 LN5 INA H23 6 INA H9 7 INA H17 8 INA H1 9 Ki3 10 K4A 11 Heterorhabditis lsquoIrish typersquo K122 12 H megidis HL8
13 H bacteriophora HP88
Fig 5 Hae III restriction digests of the PCR amplication products of the rDNA intergenic spacer region of Heterorhabditis isolat
separated on a 2 agarose gel and stained with ethidiumbromide M 1kb size marker 1 LN2 2 P2M 3 LN2B 4 LN4 5 INA H
6 INA H9 7 INA H9 8 INA H1 9 Ki3 10 K4A 11HeterorhabditislsquoIrish typersquo K122 12 H megidis HL81 13 H bacterioph HP88
USE OF H INDICA SPECIES S PECIFIC SATELLITE DNAPROBE
When Southern blots of total genomic DNA of thetropical isolates were probed with the H indica species-specic satellite DNA probe described by Abadon et al
(1998) the probe hybridised only with the isolates clas-sied from PCR analysis as being H indica including
the Kenyan isolates Ki3 and K4A (Fig 7A) No hybrisation was detected by this probe to any of the othspecies included as controls on the Southern blot Thblot was then reprobed with the H bacteriophora specisatellite DNA probe described by Grenier et al (199and this probe hybridised only with the HP88 isolate H bacteriophora included on the Southern blot (data nshown)
482 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 711
Isolates of Heterorhabditisindi
Fig 6 Hae III and Alu I restriction digests of the PCR amplication products of the rDNA intergenic spacer region of Heterorhabdisolates including isolates designated type D1a by Curran and Driver (1994) M 1kb size marker 1 H indica LN2 2 D1 3 K4
4 ES10 5 JAM79 6 ST09 7 FLGS10 M 1 kb marker 9 H indica LN2 10 D1 11 ES10 12 JAM79 13 ST09 14 FLGS1
Lanes 1 to 7 contain Hae III digests and lanes 9 to 14 contain Alu I digests The digests were separated on a 2 agarose gel and stain
with ethidium bromide
Table 2 The results of cross-breeding experiments of six Het-erorhabditis isolates from India Indonesia and Kenya
Female Male
H indica LN 4 INA H1 INA H23 Ki3 K4ALN2India India Indonesia Indonesia Kenya Kenya
H indica + + + 0 + 0LN2
LN 4 + + 0 0 + +INA H1 + + + + + 0
INA H23 0 0 + + 0 0Ki3 + + + 0 + +K4A 0 0 0 0 + +
+ cross resulted in fertile progeny0 no progeny detected
CROSS BREEDING ANALYSIS
Selectedcross-breedingresults are presented in Table2All the Indian isolates are interfertile The Kenyan iso-
lates which differed from H indica in their Hinf I ITSrestriction prole and their HaeIII IGS restriction proleare interfertile amongst each other but only one Kenyanisolate Ki3 produced viable progeny when crossed withthe H indica LN2 type species The Indonesian isolatesare interfertile but surprisingly only one of the Indone-sian isolates tested (INA H1) produced viable hybridswhen crossed with H indica LN2 The INA H1 isolatewas also interfertile with the Kenyan isolate Ki3
Discussion
The results presented here conrm and extend prevous studies which show that a combination of molecudiagnostictoolscan be reliably used for species identiction in Heterorhabditis Isolates of Heterorhabditis froIndia Kenya Indonesia and Cuba hybridised to the H i
dica specic satellite DNA probe (Grenier et al 199Abadon et al 1998) H indica specic DNA restrictiproles were also obtained for all the tropical isolat
of Heterorhabditis investigated here when the ITS rDNregion was restricted using the diagnostic restriction ezymes AluI and MboI (Joyce et al 1994a) The utilof restriction digests of the rDNA ITS region in specidiagnosis of EPN has been conrmed in several studi(Joyce et al 1994a b Miduturi et al 1996 Hominickal 1997 Yoshida et al 1998 Grifn et al 1999a Pa
jav et al 1999) Restriction of the rDNA IGS region wi AluI also yielded a H indica specic restriction prole fall the tropical isolates tested in this study
The PCR amplication product which we obtained fthe H indica rDNA IGS varied in size from 12 to 16 kHominick et al (1997) also observed length heterogenity in the rDNA IGS region among geographic isolatof single Steinernema spp and they suggested that thheterogeneity may make restriction proles of this DNfragment unreliable for species identication The lengheterogeneity which we detected in the H indica IGPCR amplication product did not affect the AluI restrtion prole of the product and a clear-cut seven restrictifragment pattern was common to all the H indica isolat
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 811
CM Stack et al
Fig 7 A Slot blot analysis of genomic DNA (100 ng) of the
Heterorhabditis isolates using a H indica specic satellite DNA
probe B The membrane used for Fig 7A was stripped and
reprobed using an rDNA 18S probe as a loading control A1
H indica LN2 A2 D1 A3 P2M A4 LN2B A5 LN4 A6
INA H23 B1 INA H9 B2 INA H17 B3 INA H1 B4 Ki3
B5 K4A B6 H bacteriophora HP88 C1 H zealandica NZH3
C2 H marelatus OH-10 C3 H hepialus Bodega Bay C4
Heterorhabditis lsquoIrish typersquo K122
tested When the IGS region was digested with HaeIII H indica LN2 displayed a distinctive ve fragment pro-le All of the H indica isolates with the exception of the Kenyan isolates also displayed a ve fragment pro-le Four of these fragments were conserved among theisolates but the largest fragment (of ca 740 in H indica
LN2) was polymorphicamong the H indica isolates Thisfragment which seems to be associated with the lengthpolymorphism in the H indica IGS region is the basisof the D1D1a polymorphism observed by Curran and
Driver (1994) In most species studied to date the rDNIGS contains tandem arrays of subrepeats (Gerbi 198Williams et al 1990 Vahidi amp Honda 1991 Novakal 1993 Linares et al 1994 Crease 1995) This rsults in the length of the IGS region being variable bobetween and within species The length heterogeneity
the H indica IGS PCR product did not affect the AluI rstriction prole of this fragment which suggests that Al
has a restriction site within a tandem repetitive unit in trDNA IGS of H indica In contrast HaeIII appears nto cut within this tandem repeat region with the result thone of the HaeIII restriction fragments is polymorphiclength presumably depending on the number of AluI rpeat units which it contains
The HaeIII rDNA IGS restriction prole of the Disolate is similar to that of H indica LN2 but the Hae
pattern of the Kenyan isolates is unique The large Hae
fragment of the Kenyan isolate K4A is the same size that of D1a thus the Kenyan isolates appear to represea distinct subgroup within D1a Tropical isolates froDarwin (North Australia) Puerto Rico Hawaii the VirgIslands and Egypt were found by Curran and Driv(1994) to possess the D1 prole while the D1a prowas described by these authors from isolates collectin Puerto Rico Jamaica Florida the Virgin Islands aEgypt Comparison of the D1 and D1a HaeIII and Al
rDNA IGS restriction proles with those of H indi
LN2 indicates that the isolates designated D1 and D
by Curran and Driver (1994) belong to H indicaAn important step towards achieving an effective nmatode bacterium complex for pest control is to seek nurally occurring endemic EPN isolates as such isolatare likely to possess physiological traits that are adaptto local climatic and ecological conditions The Kenyisolates described in this study clearly belong to a distinsub-group of H indica and it is probable that these islates also share a distinctive phenotype The two Kenyisolates are interfertile but only one of these isolates (Kiproduced viable progeny when crossed with the H indi
LN2 type species All of the Indonesian Heterorhabdi
isolates were indistinguishable from H indica in the dagnostic molecular tests and they were interfertile wieach other but only one of the Indonesian isolates (INH1) was interfertile with H indica LN2 and INA H1 walso interfertile with the Ki3 isolate from Kenya Theresults suggest that although H indica has a global dtribution in tropical and subtropical regions of the worgeneow within the species may be quite restricted Whave previously observedreproductiveincompatibilityb
484 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 911
Isolates of Heterorhabditisindi
tween isolates of H bacteriophora from Europe (Grifnet al 1999a) and from North America (Dix et al un-publ) We have not determined whether this reproductiveisolation is caused by genetic means or is the result of cy-toplasmic factors Cytoplasmic factors such as endosym-biont bacteria (see reviews by Werren 1997 Johanowicz
amp Hoy 1998) and chromosomal factors such as trans-posons (reviewed by Kidwell 1990 Petrov et al 1995)have been shown to be frequent causes of reproductivein-compatibility in arthropods Endosymbiont bacteria haverecently been shown to cause reproductiveincompatibilityin larial nematodes (Hoerauf et al 1999) Since the re-productive incompatibilityobserved in this study does notyield clear groups of compatible and incompatiblestrainsand in view of the consistencyof the DNA based diagnos-tic tests any assignment of biological species within the H indica group would be injudicious at present Adams
(1988) evaluates phylogeneticand biologicalspecies con-cepts in the delimitationof species in the H bacteriophora
groupThe global distribution of H indica throughout the
tropics and subtropics suggests that H indica possessesa range of phenotypic characters which give it an advan-tage in this climatic zone Such phenotypes might includeadaptationto high temperatures desiccation tolerance andgood dispersal ability It is unlikely that the widespreaddistribution of H indica in the tropics arises because thisspecies was extant before the continents began breakingup and drifting The best supported hypothesis of phylo-genetic relationships among Heterorhabditis species in aphylogeny derived from the rDNA ITS 1 DNA sequencedata puts H indica as the most ancient Heterorhabditis
lineage (Adams et al 1998) However the phylogenetictree for Heterorhabditis and Steinernema of Reid (1994)shows that the genetic distance between Heterorhabdi-
tis species is considerably less than that between Stein-
ernema species and given the small genetic distance de-tected by Reid (1994) between the most divergent of the Heterorhabditis species in his study (a D value of ca
004 as calculated by the method of Nei and Li 1979) it
seems unlikely that H indica speciated before the Juras-sicCretaceousbreak up of Pangaea(an estimated 200 mil-lion years ago)
Acknowledgement
We acknowledge the support of the European Commu-nity STD-3 Programme (TS3 CT 940273)
References
ABADON M GRENIER E LAUMOND C amp ABAD (1998) A species-specic satellite DNA from the enmopathogenic nematode Heterorhabditis indicus Geno
41 148-153
ADAMS BJ (1998) Species concepts and the evolutionparadigm in modern nematology Journal of Nematology 31-21
ADAMS BJ BURNELL AM amp POWERS TO (1998)phylogenetic analysis of the genus Heterorhabditis(NemaRhabditidae) based on internal transcribed spacer 1 DNsequence data Journal of Nematology 30 22-39
AMARASINGHE LD HOMINICK WM BRISCOE Bamp REI D AP (1994) Occurrence and distribution entomopathogenic nematodes in Sri Lanka Journal
Helminthology 68 277-286ARTEAGA HERNANDEZ E M amp MRAacute IumlCE K Z (1984) H
terorhabditis heliothidis a parasite of insect pests in CubFolia Parasitologia(Praha) 31 11-17
BLAXTER ML DE LE Y P GAREY J R LIU LXSCHELDEMAN P VIERSTRAETE A VANFLETERE
JR MACKEY LY DORRIS M FRISSE LM VID
JT amp THOMAS WK (1998) A molecular evolutionaframework for the phylum Nematoda Nature 392 71-75
CONSTANT P MARCHAY L F ISHER-LE SAU X MBRIAND-PANOMA S amp MAULEacuteON H (1998)Naturalocurrence of entomopathogenic nematodes( RhabditidaSteernematidae and Heterorhabditidae) in Guadeloupe islanFundamental and Applied Nematology 21 667-672
CREASE TJ (1995) Ribosomal DNA evolution at tpopulation-level mdash nucleotide variation in intergenic spaarrays of Daphnia pulex Genetics 141 1327-1337
CURRAN J amp DRIVER F (1994) Molecular taxonomy of Hterorhabditis In Burnell AM Ehlers R-U amp Masson J(Eds) Genetics of Entomopathogenic Nematode-Bacteri
Complexes Luxembourg European Commission PublicatiEUR 15681 EN pp 178-187
DIX I BURNELL A M GRIFFIN CT JOYCE SAamp DOWNES MJ (1992) The identication of biologispecies in the genus Heterorhabditis (Nematoda Heterhabditae) by cross-breeding second generation amphimicadults Parasitology104 509-518
EHLERS R-U (1996) Current and future use of nematodin biocontrolpractice and commercial aspects with regardregulatory policy issues Biocontrol Science and Technolo
6 303-317GERBI SA (1985) Evolution of ribosomal DNA In MacI
tyre RJ (Ed) Molecular evolutionary genetics New YoNY USA Plenum Press pp 419-517
GRENIER E BONIFASSI E ABAD P amp LAUMOND (1996a) Use of species specic satellite DNAs as dianostic probes in the identication of Steinernematidae a
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1011
CM Stack et al
Heterorhabditidae entomopathogenic nematodes Parasitol-
ogy 113 483-489GRENIER E LAUMOND C amp ABAD P (1996b) Molecu-
lar characterisationof two species-specic tandemly repeatedDNAs from entomopathogenic nematodes Steinernema and
Heterorhabditis(NematodaRhabditida) Molecularand Bio-
chemical Parasitology 83 47-56GRIFFIN CT JOYCE S A DIX I BURNELL AM amp
DOWNES MJ (1994)Characterisationof the entomopatho-genic nematode Heterorhabditis (Nematoda Heterorhabdi-tidae) from Ireland and Britain by molecular and cross-breeding techniques and the occurrence of the genus in theseislands Fundamental and Applied Nematology 17 245-253
GRIFFIN CT DIX I JOYCE SA BURNELL AM ampDOWNES MJ (1999a) Isolation and characterisation of
Heterorhabditis spp from Hungary Estonia and Denmark Nematology 1 321-332
GRIFFIN CT CHAERANI R FALLON D REI D AP
amp DOWNES MJ (1999b) Occurrence and distribution of the entomopathogenic nematodes Steinernema spp and Het-
erorhabditis indica in Indonesia Journal of Helminthology
74 143-150HOERAUF A NISSENPAHLE K SCHMETZ C HEN-
KLEDUHRSEN K BLAXTER ML BUTTNER DWGALLIN MY ALQAOUD KM LUCIUS R amp FLEIS-CHER B (1999) Tetracycline therapy targets intracellularbacteria in the larial nematode Litomososoides sigmodontis
and results in larial infertility Journal of Clinical Investiga-
tion 103 11-17HOMINICK WM REID AP BOHAN DA amp BRISCOE
BR (1996) Entomopathogenicnematodes biodiversity ge-ographicaldistributionand conventionon biological diversity
Biocontrol Science and Technology 6 317-332HOMINICK WM BRISCOE BR DE L PIN O FG HEN G
J HUN T DJ KOZODOY E MRAacute IumlCE K Z NGUYENKB REI D AP SPIRIDONOV S STOCK P STURHAN D WATURU C amp YOSHIDA M (1997) Biosystematics of entomopathogenic nematodes current status protocols anddenitions Journal of Helminthology 71 271-298
JOHANOWICZ D L amp HOY MA (1998) The manipula-tion of arthropodreproductionby Wolbachia endosymbiontsFlorida Entomologist 81 310-317
JOYCE S A BURNELL A M amp P OWERS TO (1994a)The characterisationof Heterorhabditisisolatesby restrictionenzyme analysis of PCR amplied fragments of mtDNA andrDNA genes Journal of Nematology 26 260-270
JOYCE SA REI D A amp CURRAN J (1994b) Applicationof the polymerase chain reaction (PCR) methods to the iden-tication of entomopathogenicnematodes In Burnell AMEhlers R-U amp Masson JP (Eds) Genetics of entomopatho-
genic nematode-bacterium complexes Luxembourg Euro-pean Commission Publication EUR 15681 EN pp 178-187
KAYA H K (1990)Soil ecology In Gaugler R amp Kaya H(Eds) Entomopathogenic nematodes in biological contr
Boca Raton FL USA CRC Press pp 93-115KAYA HK amp GAUGLER R (1993) Entomopathogenic n
matodes Annual Review of Entomology 38 181-206KIDWELL M G (1990) Evolutionaryaspects of hybrid dysg
nesis in Drosophila Canadian Journal of Zoology 68 1711726LINARES AR BOWEN T amp DOVER GA (1994) A
pects of non-random turnover involved in the concerted evlution of intergenic spacers within the ribosomal DNA
Drosophila melanogaster Journal of Molecular Evoluti
39 151-159LIU J (1994) [A new species of the genus Heterorhabd
from China (Rhabditida Heterorhabditidae)] Acta Zoota
nomica Sinica 19 268-272LIU J amp BERRY RE (1996) Heterorhabditis marela
nsp (Rhabditida Heterorhabditidae) from Oregon Journ
of Invertebrate Pathology
67 48-54MASON JM RAZAK AR amp WRIGHT DJ (1996) Trecoveryof entomopathogenicnematodes from selectedarewithin Peninsular Malaysia Journal of Helminthology 7303-307
MENTI H WRIGHT DJ amp PERRY RN (1997)Desiccatsurvival of populations of the entomopathogenic nematodSteinernema feltiaeand Heterorhabditismegidis from Greeand the UK Journal of Helminthology 71 41-46
MIDUTURI JS MOENS M HOMINICK WM BRISCO
B R amp RE ID AP (1996) Naturally occurring enmopathogenic nematodes in the province of West-FlandeBelgium Journal of Helminthology 70 319-327
NE I M amp LI W-H (1979) Mathematical model for studygenetic variation in terms of restriction endonucleases P
ceedings of the National Academy of Sciences of the Uni
States 76 5269-5273NOVAK EM DE MELLO MP GOMES HBM
GALINDO I GUEVARA P RAMIREZ JL amp DA SILV
RI A JF (1993) Repetitive sequences in the ribosomal intgenic spacer of Trypanosoma cruzi Molecular and Bioche
ical Parasitology 60 273-280PAMJAV H TRIGA D BUZAS Z VELLAI T LUCSKA
A ADAMS B REI D AP BURNELL AM GRIFF
C GLAZER I KLEIN MG amp FODOR A (1999) Nov
application of PhastSystem polyacrylamide gel electrophosis using restriction fragment length polymorphism-intertranscribed spacer patterns of individuals for molecular idetication of entomopathogenic nematodes Electrophore
201266-1273PETROV DA SCHUTZMAN J L HARTL D L amp L
ZOVSKAYA ER ( 1995) Diverse transposable elements amobilised in hybrid dysgenesis in Drosophila virilis P
ceedings of the National Academy of Sciences of the Uni
States 92 8050-8054
486 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1111
Isolates of Heterorhabditisindi
POINAR GO JR (1990) Taxonomy and biology of Stein-ernematidae and HeterorhabditidaeIn Gaugler R amp KayaHK (Eds) Entomopathogenic nematodes in biological con-
trol Boca Raton FL USA CRC Press pp 23-64POINAR GO JR (1993) Origins and phylogenetic relation-
ships of the entomophilic rhabditids Heterorhabditis and
Steinernema Fundamental and Applied Nematology16 333-338POINAR GO JR JACKSON T amp KLEIN M (1987) He-
terorhabditis megidis sp n (Heterorhabditidae Rhabditida)parasitic in the Japanese beetlePopilliajaponica(ScarabidaeColeoptera) in Ohio Proceedings of the Helminthological
Society of Washington 53 53-59POINAR GO JR KARUNAKAR GK amp DAVID H (1992)
Heterorhabditis indicus n sp (Rhabditida Nematoda) fromIndia Separation of Heterorhabditis spp by infective juve-niles Fundamental and Applied Nematology 15 467-472
REI D AP (1994) Molecular taxonomy of Steinernema InBurnell AM Ehlers R-U amp Masson JP (Eds) Genetics
of entomopathogenic nematode-bacterium complexes Lux-embourg European Commission Publication EUR 15681EN pp 178-187
SMITS P GROENEN JTM amp DERAAY G (1991) Charac-terisation of Heterorhabditis isolates using DNA restrictionlength polymorphism Revue de Neacutematologie 14 445-453
STOCK SP (1993) A new species of thegenus Heterorhabdi-
tis Poinar 1976 (Nematoda Heterorhabditidae) parasitizingGraphognathus sp larvae (Coteoptera Curculionidae) fromArgentina Research and Reviews in Parasitology 53 103-107
STOCK SP (1997) Heterorhabditis hepialus Stock Strong amp
Gardner 1996a junior synonym of H marelatus Liu amp Berry1996 (Rhabditida Rhabditidae) with a redescription of thespecies Nematologica 43 455-463
STOCK SP STRONG D amp GARDNER S (1996) Identiction of Heterorhabditis (Nematoda Heterorhabditidae)froCalifornia with a new species isolated from the larvae of tghost moth Hepialius [sic] californicus (Lepidoptera Healidae) from Bodega Bay Natural Reserve Fundamental a
Applied Nematology 19 585-592
STRONG DR KAYA HK WHIPPLE AV CHILD ALKRAIG S BONDONNE M DYE R K amp MARON J(1996) Entomopathogenic nematodes natural enemiesroot feeding caterpillarson bush lupine Oecologica 108 345
SUDHAUS W (1993) Die mittels symbiontischerBakterienetomopathogenen Nematoden Gattungen Heterorhabditis aSteinernema sind keine Schwestertaxa Verhandlungen d
Deutschen Zoologischen Gesellschaft 86 146VAHIDI H amp HONDA BM (1991)Repeats and subrepeats
the intergenic spacer of rDNA from the nematode Meloid
gyne arenaria Molecular and General Genetics 227 33
336WERREN JH (1997) Biology of Wolbachia Annual Revi
of Entomology 42 587-609WILLIAMS S M ROBBINS L G CLUSTER PD A
LARD RW amp STROBECK C (1990) Superstructureof t Drosophila ribosomal gene family Proceedings of the N
tional Academy of Sciences of the United States 87 3153160
WOUTS WM (1979) The biology and life cycle ofNew Zealand population of Heterorhabditis heliothidis (Hterorhabditidae)Nematologica 25 191-202
YOSHIDA M REID AP BRISCOE BR amp HOMINIC
WM (1998) Survey of entomopathogenic nematod(Rhabditida Steinernematidae and Heterorhabditidae) Japan Fundamental and Applied Nematology 21 185-198
Vol 2(5) 2000 4

862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 411
CM Stack et al
EDTA and the samples were then boiled for 10 minin a water bath The samples were then transferred byvacuum suction onto a positively charged nylon mem-brane (Amersham Life Sciences Ltd Amersham Buck-inghamshire UK) in a slot blot apparatus (Schleicheramp Schuell D37582 Dassel Germany) as per manufac-
turerrsquos instructions The DNA was then xed onto thenylon membrane by UV cross-linking using StratagenersquosStratalinker (Stratagene La Jolla CA 92037 USA) Therecombinant pUC plasmids Hi12 and HP88s9 which con-tained respectively the H indica and H bacteriophora
satellite DNA monomers were obtained from Dr PierreAbad INRA Antibes France and amplied using PCRThe PCR primers used to amplify the H indica monomerfrom the Hi12 plasmid were 5cent-CTGAAGCACTTGGGA-CAGAGC-3cent and 5cent-CTCCTCGTTGAGGACGGGAGT-3cent (Abadon et al 1998Grenier pers comm) The H bac-
teriophora monomer was amplied from the HP88s9plasmid using the following PCR primers 5cent-AGCTATG-CCAGAATGATCGCC-3cent and5cent-AGATTCTCTGTACG-ATGAGTA-3cent (Grenier et al 1996b Grenier perscomm) DNA was amplied using the following condi-tions one cycle of 94degC for 5 min was followed by 35cycles of denaturation at 94degC for 05 min annealing at52degC for 1 min and extension at 72degC for 2 min with anal cycle of extension at 72degC for 5 min
The probes were labelled using the ECL direct nucleicacid detection system (Amersham) following the man-ufacturerrsquos instructions Hybridisations were conducted
at 42degC overnight After hybridisation the lters werewashed rst with 05 acute SSC for 40 min at 42degC and thenwith 01acute SSC for 5 min at room temperatureAfter post-hybridisation washes lters were exposed to Hyperim-ECL (Amersham) following the manufacturerrsquos instruc-tions The membrane was then stripped and reprobed us-ing an 18S rDNA probe as a loading control The 18Sprobe was obtained by PCR from H indica LN2 genomicDNA using the primers 18SR2B 5cent-TACAAAGGGCAG-GGACGTATT-3cent and 18S12 5cent-GGCGATCAGATACC-GCCCTAGTT-3cent (TO Powers pers comm)
CROSS-BREEDING TESTS
Cross-breedingstudieswere carried outas describedbyDix et al (1992) with the following controls being set upfor each cross virginity test mdash 20 virgin females wereplaced on a lipid agar plate that had been inoculated andpre-incubated with the primary form of the LN2 bacteriaself-cross mdash ten virgin females and ten males of thesame isolate were placed on lipid agar plates containing
the bacterial symbiont The result of any cross betwedifferent isolates was taken as valid only if there were progeny in the virginity test and there were progeny in tself-cross At least ten second generation virgin femalwere used for each cross
Results
RESTRICTION PROFILES OF THE RDNA INTERNAL
TRANSCRIBER SPACER (ITS) REGION
All of the tropical isolates of Heterorhabditis yielda ca 1 kb fragment upon PCR amplication with the ITprimers These amplication products were digested wthe diagnostic restriction endonucleases used by Joy
et al (1994a) for species diagnosis in Heterorhabdi(viz AluI Hinf I and MboI) When digested with Al
(Fig 1) the Indian isolates LN2B and LN4 shared tsame restriction prole as the H indica LN2 type specias did the Indonesian isolates the P2M isolate froCuba and the Kenyan isolates When digested with Mbo
the Kenyan isolates shared the same restriction pro
Fig 1 Alu I restriction digests of the PCR amplication produ
of the rDNA internal transcribed spacer region of Heterorhditis isolates separated on a 2 agarose gel and stained w
ethidium bromide M 1 kb marker 1 H indica LN2 2 P2
3 LN2B 4 LN4 5 INA H23 6 INA H9 7 INA H17 8 I
H1 9 Ki3 10 K4A 11 HeterorhabditislsquoIrish typersquo K122 1
H megidis HL81
480 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 511
Isolates of Heterorhabditisindi
as H indica LN2 and P2M (Fig 2) The Indonesianisolates also displayed the same MboI restriction proleas H indica LN2 (data not shown) When digested with Hinf I (Fig 3) the Indian Indonesian and Cuban isolatesbut not the Kenyan isolates possessed the same restrictionprole as H indica LN2 The Kenyan isolates have only
a single Hinf I restriction site in the ITS rDNA fragmentyielding two restriction fragments of ca 620 and 450 bpunlikeall the other H indica isolates testedwhich possesstwo Hinf I sites in this region yielding three restrictionfragments of ca 450 360 and 240 bp
Fig 2 Mbo I restriction digests of the PCR amplication prod-
ucts of the rDNA internal transcribed spacer region of Het-erorhabditis isolates separated on a 2 agarose gel and stained
with ethidium bromide M 1 kb marker 1 H indica LN22 P2M 3 Ki3 4 K4A 5 H bacteriophora HP88
RESTRICTION PROFILES OF THE RDNA INTERGENIC
SPACER (IGS) REGION
The PCR amplication products for the IGS rDNA rgion of the tropical isolates varied in size from 15 to 17 kb The amplied IGS rDNA fragments were r
stricted with the endonucleases AluI and HaeIII A dianostic AluI restriction pattern was obtained for H indi
LN2 and this was shared by all the Indian IndonesiaCuban and Kenyan isolates (Fig 4) and the D1 and Dtype isolates(Fig 6) When HaeIII was used to digest tIGS rDNA region H indica LN2 displayed a distinctive fragment prole of 740 280 190 160 and 120 (Fig 5) and with the exception of the Kenyan isolatefour of these fragments were shared by the other tropcal isolates The Kenyan isolates Ki3 (lane 9) and K4(lane 10) had a distinct restriction prole yielding Hae
fragments of 700 280 (a doublet) 185 and 120 bp Tlargest H indica HaeIII fragment was highlypolymorphin size between the tropical isolates ranging in size fro700 to 850 bp with some isolates having fragments of itermediate size ca 740 bp Size polymorphismin thisframent was the basis on which the types D1 and D1a werecognised by Curran and Driver (1994) The HaeIII prles of the other three species of Heterorhabditis includin Fig 5 were all distinctly different from that of H i
dica As can be seen from Fig 6 the HaeIII rDNA IGrestriction prole designated D1a by Curran and Driv
(1994)(lanes 4-7) is similar to that of H indica LN2 bthe HaeIII pattern of the Kenyan isolates (Fig 6 laneFig 5 lanes 910) is unique
Fig 3 Hinf I restriction digests of the PCR amplication products of the rDNA int ernal transcribed spacer region of Heterorhabdisolates separated on a 2 agarose gel and stained with ethidium bromide M 1 kb marker 1 LN2 2 P2M 3 LN2B 4 LN
5 INA H23 6 INA H9 7 INA H17 8 INA H1 9 Ki3 10 K4A 11HeterorhabditislsquoIrish typersquo K122 12 H megidis HL81
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 611
CM Stack et al
Fig 4 Alu I restriction digests of the PCR amplication products of the r DNA intergenic spacer region of Heterorhabditis isolat
separated on a 2 agarose gel and stained with ethidium bromide M 1kb size marker 1 LN2 2 P2M 3 LN2B 4 LN5 INA H23 6 INA H9 7 INA H17 8 INA H1 9 Ki3 10 K4A 11 Heterorhabditis lsquoIrish typersquo K122 12 H megidis HL8
13 H bacteriophora HP88
Fig 5 Hae III restriction digests of the PCR amplication products of the rDNA intergenic spacer region of Heterorhabditis isolat
separated on a 2 agarose gel and stained with ethidiumbromide M 1kb size marker 1 LN2 2 P2M 3 LN2B 4 LN4 5 INA H
6 INA H9 7 INA H9 8 INA H1 9 Ki3 10 K4A 11HeterorhabditislsquoIrish typersquo K122 12 H megidis HL81 13 H bacterioph HP88
USE OF H INDICA SPECIES S PECIFIC SATELLITE DNAPROBE
When Southern blots of total genomic DNA of thetropical isolates were probed with the H indica species-specic satellite DNA probe described by Abadon et al
(1998) the probe hybridised only with the isolates clas-sied from PCR analysis as being H indica including
the Kenyan isolates Ki3 and K4A (Fig 7A) No hybrisation was detected by this probe to any of the othspecies included as controls on the Southern blot Thblot was then reprobed with the H bacteriophora specisatellite DNA probe described by Grenier et al (199and this probe hybridised only with the HP88 isolate H bacteriophora included on the Southern blot (data nshown)
482 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 711
Isolates of Heterorhabditisindi
Fig 6 Hae III and Alu I restriction digests of the PCR amplication products of the rDNA intergenic spacer region of Heterorhabdisolates including isolates designated type D1a by Curran and Driver (1994) M 1kb size marker 1 H indica LN2 2 D1 3 K4
4 ES10 5 JAM79 6 ST09 7 FLGS10 M 1 kb marker 9 H indica LN2 10 D1 11 ES10 12 JAM79 13 ST09 14 FLGS1
Lanes 1 to 7 contain Hae III digests and lanes 9 to 14 contain Alu I digests The digests were separated on a 2 agarose gel and stain
with ethidium bromide
Table 2 The results of cross-breeding experiments of six Het-erorhabditis isolates from India Indonesia and Kenya
Female Male
H indica LN 4 INA H1 INA H23 Ki3 K4ALN2India India Indonesia Indonesia Kenya Kenya
H indica + + + 0 + 0LN2
LN 4 + + 0 0 + +INA H1 + + + + + 0
INA H23 0 0 + + 0 0Ki3 + + + 0 + +K4A 0 0 0 0 + +
+ cross resulted in fertile progeny0 no progeny detected
CROSS BREEDING ANALYSIS
Selectedcross-breedingresults are presented in Table2All the Indian isolates are interfertile The Kenyan iso-
lates which differed from H indica in their Hinf I ITSrestriction prole and their HaeIII IGS restriction proleare interfertile amongst each other but only one Kenyanisolate Ki3 produced viable progeny when crossed withthe H indica LN2 type species The Indonesian isolatesare interfertile but surprisingly only one of the Indone-sian isolates tested (INA H1) produced viable hybridswhen crossed with H indica LN2 The INA H1 isolatewas also interfertile with the Kenyan isolate Ki3
Discussion
The results presented here conrm and extend prevous studies which show that a combination of molecudiagnostictoolscan be reliably used for species identiction in Heterorhabditis Isolates of Heterorhabditis froIndia Kenya Indonesia and Cuba hybridised to the H i
dica specic satellite DNA probe (Grenier et al 199Abadon et al 1998) H indica specic DNA restrictiproles were also obtained for all the tropical isolat
of Heterorhabditis investigated here when the ITS rDNregion was restricted using the diagnostic restriction ezymes AluI and MboI (Joyce et al 1994a) The utilof restriction digests of the rDNA ITS region in specidiagnosis of EPN has been conrmed in several studi(Joyce et al 1994a b Miduturi et al 1996 Hominickal 1997 Yoshida et al 1998 Grifn et al 1999a Pa
jav et al 1999) Restriction of the rDNA IGS region wi AluI also yielded a H indica specic restriction prole fall the tropical isolates tested in this study
The PCR amplication product which we obtained fthe H indica rDNA IGS varied in size from 12 to 16 kHominick et al (1997) also observed length heterogenity in the rDNA IGS region among geographic isolatof single Steinernema spp and they suggested that thheterogeneity may make restriction proles of this DNfragment unreliable for species identication The lengheterogeneity which we detected in the H indica IGPCR amplication product did not affect the AluI restrtion prole of the product and a clear-cut seven restrictifragment pattern was common to all the H indica isolat
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 811
CM Stack et al
Fig 7 A Slot blot analysis of genomic DNA (100 ng) of the
Heterorhabditis isolates using a H indica specic satellite DNA
probe B The membrane used for Fig 7A was stripped and
reprobed using an rDNA 18S probe as a loading control A1
H indica LN2 A2 D1 A3 P2M A4 LN2B A5 LN4 A6
INA H23 B1 INA H9 B2 INA H17 B3 INA H1 B4 Ki3
B5 K4A B6 H bacteriophora HP88 C1 H zealandica NZH3
C2 H marelatus OH-10 C3 H hepialus Bodega Bay C4
Heterorhabditis lsquoIrish typersquo K122
tested When the IGS region was digested with HaeIII H indica LN2 displayed a distinctive ve fragment pro-le All of the H indica isolates with the exception of the Kenyan isolates also displayed a ve fragment pro-le Four of these fragments were conserved among theisolates but the largest fragment (of ca 740 in H indica
LN2) was polymorphicamong the H indica isolates Thisfragment which seems to be associated with the lengthpolymorphism in the H indica IGS region is the basisof the D1D1a polymorphism observed by Curran and
Driver (1994) In most species studied to date the rDNIGS contains tandem arrays of subrepeats (Gerbi 198Williams et al 1990 Vahidi amp Honda 1991 Novakal 1993 Linares et al 1994 Crease 1995) This rsults in the length of the IGS region being variable bobetween and within species The length heterogeneity
the H indica IGS PCR product did not affect the AluI rstriction prole of this fragment which suggests that Al
has a restriction site within a tandem repetitive unit in trDNA IGS of H indica In contrast HaeIII appears nto cut within this tandem repeat region with the result thone of the HaeIII restriction fragments is polymorphiclength presumably depending on the number of AluI rpeat units which it contains
The HaeIII rDNA IGS restriction prole of the Disolate is similar to that of H indica LN2 but the Hae
pattern of the Kenyan isolates is unique The large Hae
fragment of the Kenyan isolate K4A is the same size that of D1a thus the Kenyan isolates appear to represea distinct subgroup within D1a Tropical isolates froDarwin (North Australia) Puerto Rico Hawaii the VirgIslands and Egypt were found by Curran and Driv(1994) to possess the D1 prole while the D1a prowas described by these authors from isolates collectin Puerto Rico Jamaica Florida the Virgin Islands aEgypt Comparison of the D1 and D1a HaeIII and Al
rDNA IGS restriction proles with those of H indi
LN2 indicates that the isolates designated D1 and D
by Curran and Driver (1994) belong to H indicaAn important step towards achieving an effective nmatode bacterium complex for pest control is to seek nurally occurring endemic EPN isolates as such isolatare likely to possess physiological traits that are adaptto local climatic and ecological conditions The Kenyisolates described in this study clearly belong to a distinsub-group of H indica and it is probable that these islates also share a distinctive phenotype The two Kenyisolates are interfertile but only one of these isolates (Kiproduced viable progeny when crossed with the H indi
LN2 type species All of the Indonesian Heterorhabdi
isolates were indistinguishable from H indica in the dagnostic molecular tests and they were interfertile wieach other but only one of the Indonesian isolates (INH1) was interfertile with H indica LN2 and INA H1 walso interfertile with the Ki3 isolate from Kenya Theresults suggest that although H indica has a global dtribution in tropical and subtropical regions of the worgeneow within the species may be quite restricted Whave previously observedreproductiveincompatibilityb
484 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 911
Isolates of Heterorhabditisindi
tween isolates of H bacteriophora from Europe (Grifnet al 1999a) and from North America (Dix et al un-publ) We have not determined whether this reproductiveisolation is caused by genetic means or is the result of cy-toplasmic factors Cytoplasmic factors such as endosym-biont bacteria (see reviews by Werren 1997 Johanowicz
amp Hoy 1998) and chromosomal factors such as trans-posons (reviewed by Kidwell 1990 Petrov et al 1995)have been shown to be frequent causes of reproductivein-compatibility in arthropods Endosymbiont bacteria haverecently been shown to cause reproductiveincompatibilityin larial nematodes (Hoerauf et al 1999) Since the re-productive incompatibilityobserved in this study does notyield clear groups of compatible and incompatiblestrainsand in view of the consistencyof the DNA based diagnos-tic tests any assignment of biological species within the H indica group would be injudicious at present Adams
(1988) evaluates phylogeneticand biologicalspecies con-cepts in the delimitationof species in the H bacteriophora
groupThe global distribution of H indica throughout the
tropics and subtropics suggests that H indica possessesa range of phenotypic characters which give it an advan-tage in this climatic zone Such phenotypes might includeadaptationto high temperatures desiccation tolerance andgood dispersal ability It is unlikely that the widespreaddistribution of H indica in the tropics arises because thisspecies was extant before the continents began breakingup and drifting The best supported hypothesis of phylo-genetic relationships among Heterorhabditis species in aphylogeny derived from the rDNA ITS 1 DNA sequencedata puts H indica as the most ancient Heterorhabditis
lineage (Adams et al 1998) However the phylogenetictree for Heterorhabditis and Steinernema of Reid (1994)shows that the genetic distance between Heterorhabdi-
tis species is considerably less than that between Stein-
ernema species and given the small genetic distance de-tected by Reid (1994) between the most divergent of the Heterorhabditis species in his study (a D value of ca
004 as calculated by the method of Nei and Li 1979) it
seems unlikely that H indica speciated before the Juras-sicCretaceousbreak up of Pangaea(an estimated 200 mil-lion years ago)
Acknowledgement
We acknowledge the support of the European Commu-nity STD-3 Programme (TS3 CT 940273)
References
ABADON M GRENIER E LAUMOND C amp ABAD (1998) A species-specic satellite DNA from the enmopathogenic nematode Heterorhabditis indicus Geno
41 148-153
ADAMS BJ (1998) Species concepts and the evolutionparadigm in modern nematology Journal of Nematology 31-21
ADAMS BJ BURNELL AM amp POWERS TO (1998)phylogenetic analysis of the genus Heterorhabditis(NemaRhabditidae) based on internal transcribed spacer 1 DNsequence data Journal of Nematology 30 22-39
AMARASINGHE LD HOMINICK WM BRISCOE Bamp REI D AP (1994) Occurrence and distribution entomopathogenic nematodes in Sri Lanka Journal
Helminthology 68 277-286ARTEAGA HERNANDEZ E M amp MRAacute IumlCE K Z (1984) H
terorhabditis heliothidis a parasite of insect pests in CubFolia Parasitologia(Praha) 31 11-17
BLAXTER ML DE LE Y P GAREY J R LIU LXSCHELDEMAN P VIERSTRAETE A VANFLETERE
JR MACKEY LY DORRIS M FRISSE LM VID
JT amp THOMAS WK (1998) A molecular evolutionaframework for the phylum Nematoda Nature 392 71-75
CONSTANT P MARCHAY L F ISHER-LE SAU X MBRIAND-PANOMA S amp MAULEacuteON H (1998)Naturalocurrence of entomopathogenic nematodes( RhabditidaSteernematidae and Heterorhabditidae) in Guadeloupe islanFundamental and Applied Nematology 21 667-672
CREASE TJ (1995) Ribosomal DNA evolution at tpopulation-level mdash nucleotide variation in intergenic spaarrays of Daphnia pulex Genetics 141 1327-1337
CURRAN J amp DRIVER F (1994) Molecular taxonomy of Hterorhabditis In Burnell AM Ehlers R-U amp Masson J(Eds) Genetics of Entomopathogenic Nematode-Bacteri
Complexes Luxembourg European Commission PublicatiEUR 15681 EN pp 178-187
DIX I BURNELL A M GRIFFIN CT JOYCE SAamp DOWNES MJ (1992) The identication of biologispecies in the genus Heterorhabditis (Nematoda Heterhabditae) by cross-breeding second generation amphimicadults Parasitology104 509-518
EHLERS R-U (1996) Current and future use of nematodin biocontrolpractice and commercial aspects with regardregulatory policy issues Biocontrol Science and Technolo
6 303-317GERBI SA (1985) Evolution of ribosomal DNA In MacI
tyre RJ (Ed) Molecular evolutionary genetics New YoNY USA Plenum Press pp 419-517
GRENIER E BONIFASSI E ABAD P amp LAUMOND (1996a) Use of species specic satellite DNAs as dianostic probes in the identication of Steinernematidae a
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1011
CM Stack et al
Heterorhabditidae entomopathogenic nematodes Parasitol-
ogy 113 483-489GRENIER E LAUMOND C amp ABAD P (1996b) Molecu-
lar characterisationof two species-specic tandemly repeatedDNAs from entomopathogenic nematodes Steinernema and
Heterorhabditis(NematodaRhabditida) Molecularand Bio-
chemical Parasitology 83 47-56GRIFFIN CT JOYCE S A DIX I BURNELL AM amp
DOWNES MJ (1994)Characterisationof the entomopatho-genic nematode Heterorhabditis (Nematoda Heterorhabdi-tidae) from Ireland and Britain by molecular and cross-breeding techniques and the occurrence of the genus in theseislands Fundamental and Applied Nematology 17 245-253
GRIFFIN CT DIX I JOYCE SA BURNELL AM ampDOWNES MJ (1999a) Isolation and characterisation of
Heterorhabditis spp from Hungary Estonia and Denmark Nematology 1 321-332
GRIFFIN CT CHAERANI R FALLON D REI D AP
amp DOWNES MJ (1999b) Occurrence and distribution of the entomopathogenic nematodes Steinernema spp and Het-
erorhabditis indica in Indonesia Journal of Helminthology
74 143-150HOERAUF A NISSENPAHLE K SCHMETZ C HEN-
KLEDUHRSEN K BLAXTER ML BUTTNER DWGALLIN MY ALQAOUD KM LUCIUS R amp FLEIS-CHER B (1999) Tetracycline therapy targets intracellularbacteria in the larial nematode Litomososoides sigmodontis
and results in larial infertility Journal of Clinical Investiga-
tion 103 11-17HOMINICK WM REID AP BOHAN DA amp BRISCOE
BR (1996) Entomopathogenicnematodes biodiversity ge-ographicaldistributionand conventionon biological diversity
Biocontrol Science and Technology 6 317-332HOMINICK WM BRISCOE BR DE L PIN O FG HEN G
J HUN T DJ KOZODOY E MRAacute IumlCE K Z NGUYENKB REI D AP SPIRIDONOV S STOCK P STURHAN D WATURU C amp YOSHIDA M (1997) Biosystematics of entomopathogenic nematodes current status protocols anddenitions Journal of Helminthology 71 271-298
JOHANOWICZ D L amp HOY MA (1998) The manipula-tion of arthropodreproductionby Wolbachia endosymbiontsFlorida Entomologist 81 310-317
JOYCE S A BURNELL A M amp P OWERS TO (1994a)The characterisationof Heterorhabditisisolatesby restrictionenzyme analysis of PCR amplied fragments of mtDNA andrDNA genes Journal of Nematology 26 260-270
JOYCE SA REI D A amp CURRAN J (1994b) Applicationof the polymerase chain reaction (PCR) methods to the iden-tication of entomopathogenicnematodes In Burnell AMEhlers R-U amp Masson JP (Eds) Genetics of entomopatho-
genic nematode-bacterium complexes Luxembourg Euro-pean Commission Publication EUR 15681 EN pp 178-187
KAYA H K (1990)Soil ecology In Gaugler R amp Kaya H(Eds) Entomopathogenic nematodes in biological contr
Boca Raton FL USA CRC Press pp 93-115KAYA HK amp GAUGLER R (1993) Entomopathogenic n
matodes Annual Review of Entomology 38 181-206KIDWELL M G (1990) Evolutionaryaspects of hybrid dysg
nesis in Drosophila Canadian Journal of Zoology 68 1711726LINARES AR BOWEN T amp DOVER GA (1994) A
pects of non-random turnover involved in the concerted evlution of intergenic spacers within the ribosomal DNA
Drosophila melanogaster Journal of Molecular Evoluti
39 151-159LIU J (1994) [A new species of the genus Heterorhabd
from China (Rhabditida Heterorhabditidae)] Acta Zoota
nomica Sinica 19 268-272LIU J amp BERRY RE (1996) Heterorhabditis marela
nsp (Rhabditida Heterorhabditidae) from Oregon Journ
of Invertebrate Pathology
67 48-54MASON JM RAZAK AR amp WRIGHT DJ (1996) Trecoveryof entomopathogenicnematodes from selectedarewithin Peninsular Malaysia Journal of Helminthology 7303-307
MENTI H WRIGHT DJ amp PERRY RN (1997)Desiccatsurvival of populations of the entomopathogenic nematodSteinernema feltiaeand Heterorhabditismegidis from Greeand the UK Journal of Helminthology 71 41-46
MIDUTURI JS MOENS M HOMINICK WM BRISCO
B R amp RE ID AP (1996) Naturally occurring enmopathogenic nematodes in the province of West-FlandeBelgium Journal of Helminthology 70 319-327
NE I M amp LI W-H (1979) Mathematical model for studygenetic variation in terms of restriction endonucleases P
ceedings of the National Academy of Sciences of the Uni
States 76 5269-5273NOVAK EM DE MELLO MP GOMES HBM
GALINDO I GUEVARA P RAMIREZ JL amp DA SILV
RI A JF (1993) Repetitive sequences in the ribosomal intgenic spacer of Trypanosoma cruzi Molecular and Bioche
ical Parasitology 60 273-280PAMJAV H TRIGA D BUZAS Z VELLAI T LUCSKA
A ADAMS B REI D AP BURNELL AM GRIFF
C GLAZER I KLEIN MG amp FODOR A (1999) Nov
application of PhastSystem polyacrylamide gel electrophosis using restriction fragment length polymorphism-intertranscribed spacer patterns of individuals for molecular idetication of entomopathogenic nematodes Electrophore
201266-1273PETROV DA SCHUTZMAN J L HARTL D L amp L
ZOVSKAYA ER ( 1995) Diverse transposable elements amobilised in hybrid dysgenesis in Drosophila virilis P
ceedings of the National Academy of Sciences of the Uni
States 92 8050-8054
486 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1111
Isolates of Heterorhabditisindi
POINAR GO JR (1990) Taxonomy and biology of Stein-ernematidae and HeterorhabditidaeIn Gaugler R amp KayaHK (Eds) Entomopathogenic nematodes in biological con-
trol Boca Raton FL USA CRC Press pp 23-64POINAR GO JR (1993) Origins and phylogenetic relation-
ships of the entomophilic rhabditids Heterorhabditis and
Steinernema Fundamental and Applied Nematology16 333-338POINAR GO JR JACKSON T amp KLEIN M (1987) He-
terorhabditis megidis sp n (Heterorhabditidae Rhabditida)parasitic in the Japanese beetlePopilliajaponica(ScarabidaeColeoptera) in Ohio Proceedings of the Helminthological
Society of Washington 53 53-59POINAR GO JR KARUNAKAR GK amp DAVID H (1992)
Heterorhabditis indicus n sp (Rhabditida Nematoda) fromIndia Separation of Heterorhabditis spp by infective juve-niles Fundamental and Applied Nematology 15 467-472
REI D AP (1994) Molecular taxonomy of Steinernema InBurnell AM Ehlers R-U amp Masson JP (Eds) Genetics
of entomopathogenic nematode-bacterium complexes Lux-embourg European Commission Publication EUR 15681EN pp 178-187
SMITS P GROENEN JTM amp DERAAY G (1991) Charac-terisation of Heterorhabditis isolates using DNA restrictionlength polymorphism Revue de Neacutematologie 14 445-453
STOCK SP (1993) A new species of thegenus Heterorhabdi-
tis Poinar 1976 (Nematoda Heterorhabditidae) parasitizingGraphognathus sp larvae (Coteoptera Curculionidae) fromArgentina Research and Reviews in Parasitology 53 103-107
STOCK SP (1997) Heterorhabditis hepialus Stock Strong amp
Gardner 1996a junior synonym of H marelatus Liu amp Berry1996 (Rhabditida Rhabditidae) with a redescription of thespecies Nematologica 43 455-463
STOCK SP STRONG D amp GARDNER S (1996) Identiction of Heterorhabditis (Nematoda Heterorhabditidae)froCalifornia with a new species isolated from the larvae of tghost moth Hepialius [sic] californicus (Lepidoptera Healidae) from Bodega Bay Natural Reserve Fundamental a
Applied Nematology 19 585-592
STRONG DR KAYA HK WHIPPLE AV CHILD ALKRAIG S BONDONNE M DYE R K amp MARON J(1996) Entomopathogenic nematodes natural enemiesroot feeding caterpillarson bush lupine Oecologica 108 345
SUDHAUS W (1993) Die mittels symbiontischerBakterienetomopathogenen Nematoden Gattungen Heterorhabditis aSteinernema sind keine Schwestertaxa Verhandlungen d
Deutschen Zoologischen Gesellschaft 86 146VAHIDI H amp HONDA BM (1991)Repeats and subrepeats
the intergenic spacer of rDNA from the nematode Meloid
gyne arenaria Molecular and General Genetics 227 33
336WERREN JH (1997) Biology of Wolbachia Annual Revi
of Entomology 42 587-609WILLIAMS S M ROBBINS L G CLUSTER PD A
LARD RW amp STROBECK C (1990) Superstructureof t Drosophila ribosomal gene family Proceedings of the N
tional Academy of Sciences of the United States 87 3153160
WOUTS WM (1979) The biology and life cycle ofNew Zealand population of Heterorhabditis heliothidis (Hterorhabditidae)Nematologica 25 191-202
YOSHIDA M REID AP BRISCOE BR amp HOMINIC
WM (1998) Survey of entomopathogenic nematod(Rhabditida Steinernematidae and Heterorhabditidae) Japan Fundamental and Applied Nematology 21 185-198
Vol 2(5) 2000 4
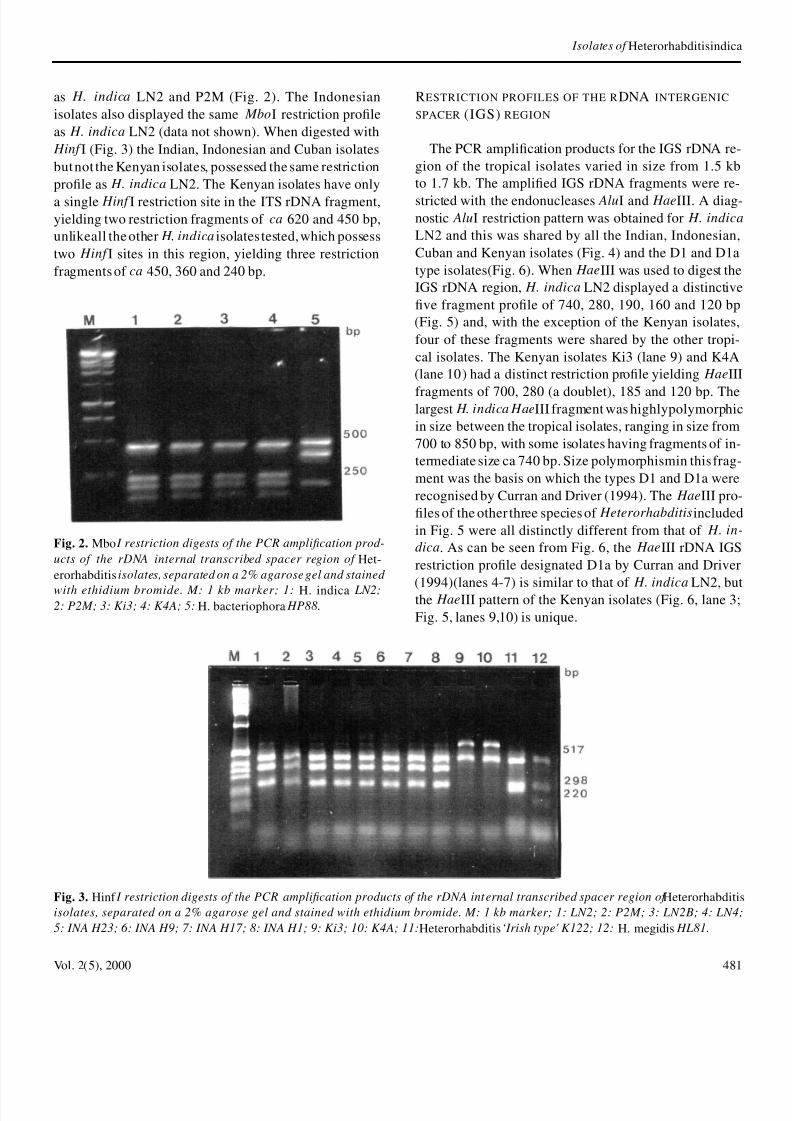
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 511
Isolates of Heterorhabditisindi
as H indica LN2 and P2M (Fig 2) The Indonesianisolates also displayed the same MboI restriction proleas H indica LN2 (data not shown) When digested with Hinf I (Fig 3) the Indian Indonesian and Cuban isolatesbut not the Kenyan isolates possessed the same restrictionprole as H indica LN2 The Kenyan isolates have only
a single Hinf I restriction site in the ITS rDNA fragmentyielding two restriction fragments of ca 620 and 450 bpunlikeall the other H indica isolates testedwhich possesstwo Hinf I sites in this region yielding three restrictionfragments of ca 450 360 and 240 bp
Fig 2 Mbo I restriction digests of the PCR amplication prod-
ucts of the rDNA internal transcribed spacer region of Het-erorhabditis isolates separated on a 2 agarose gel and stained
with ethidium bromide M 1 kb marker 1 H indica LN22 P2M 3 Ki3 4 K4A 5 H bacteriophora HP88
RESTRICTION PROFILES OF THE RDNA INTERGENIC
SPACER (IGS) REGION
The PCR amplication products for the IGS rDNA rgion of the tropical isolates varied in size from 15 to 17 kb The amplied IGS rDNA fragments were r
stricted with the endonucleases AluI and HaeIII A dianostic AluI restriction pattern was obtained for H indi
LN2 and this was shared by all the Indian IndonesiaCuban and Kenyan isolates (Fig 4) and the D1 and Dtype isolates(Fig 6) When HaeIII was used to digest tIGS rDNA region H indica LN2 displayed a distinctive fragment prole of 740 280 190 160 and 120 (Fig 5) and with the exception of the Kenyan isolatefour of these fragments were shared by the other tropcal isolates The Kenyan isolates Ki3 (lane 9) and K4(lane 10) had a distinct restriction prole yielding Hae
fragments of 700 280 (a doublet) 185 and 120 bp Tlargest H indica HaeIII fragment was highlypolymorphin size between the tropical isolates ranging in size fro700 to 850 bp with some isolates having fragments of itermediate size ca 740 bp Size polymorphismin thisframent was the basis on which the types D1 and D1a werecognised by Curran and Driver (1994) The HaeIII prles of the other three species of Heterorhabditis includin Fig 5 were all distinctly different from that of H i
dica As can be seen from Fig 6 the HaeIII rDNA IGrestriction prole designated D1a by Curran and Driv
(1994)(lanes 4-7) is similar to that of H indica LN2 bthe HaeIII pattern of the Kenyan isolates (Fig 6 laneFig 5 lanes 910) is unique
Fig 3 Hinf I restriction digests of the PCR amplication products of the rDNA int ernal transcribed spacer region of Heterorhabdisolates separated on a 2 agarose gel and stained with ethidium bromide M 1 kb marker 1 LN2 2 P2M 3 LN2B 4 LN
5 INA H23 6 INA H9 7 INA H17 8 INA H1 9 Ki3 10 K4A 11HeterorhabditislsquoIrish typersquo K122 12 H megidis HL81
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 611
CM Stack et al
Fig 4 Alu I restriction digests of the PCR amplication products of the r DNA intergenic spacer region of Heterorhabditis isolat
separated on a 2 agarose gel and stained with ethidium bromide M 1kb size marker 1 LN2 2 P2M 3 LN2B 4 LN5 INA H23 6 INA H9 7 INA H17 8 INA H1 9 Ki3 10 K4A 11 Heterorhabditis lsquoIrish typersquo K122 12 H megidis HL8
13 H bacteriophora HP88
Fig 5 Hae III restriction digests of the PCR amplication products of the rDNA intergenic spacer region of Heterorhabditis isolat
separated on a 2 agarose gel and stained with ethidiumbromide M 1kb size marker 1 LN2 2 P2M 3 LN2B 4 LN4 5 INA H
6 INA H9 7 INA H9 8 INA H1 9 Ki3 10 K4A 11HeterorhabditislsquoIrish typersquo K122 12 H megidis HL81 13 H bacterioph HP88
USE OF H INDICA SPECIES S PECIFIC SATELLITE DNAPROBE
When Southern blots of total genomic DNA of thetropical isolates were probed with the H indica species-specic satellite DNA probe described by Abadon et al
(1998) the probe hybridised only with the isolates clas-sied from PCR analysis as being H indica including
the Kenyan isolates Ki3 and K4A (Fig 7A) No hybrisation was detected by this probe to any of the othspecies included as controls on the Southern blot Thblot was then reprobed with the H bacteriophora specisatellite DNA probe described by Grenier et al (199and this probe hybridised only with the HP88 isolate H bacteriophora included on the Southern blot (data nshown)
482 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 711
Isolates of Heterorhabditisindi
Fig 6 Hae III and Alu I restriction digests of the PCR amplication products of the rDNA intergenic spacer region of Heterorhabdisolates including isolates designated type D1a by Curran and Driver (1994) M 1kb size marker 1 H indica LN2 2 D1 3 K4
4 ES10 5 JAM79 6 ST09 7 FLGS10 M 1 kb marker 9 H indica LN2 10 D1 11 ES10 12 JAM79 13 ST09 14 FLGS1
Lanes 1 to 7 contain Hae III digests and lanes 9 to 14 contain Alu I digests The digests were separated on a 2 agarose gel and stain
with ethidium bromide
Table 2 The results of cross-breeding experiments of six Het-erorhabditis isolates from India Indonesia and Kenya
Female Male
H indica LN 4 INA H1 INA H23 Ki3 K4ALN2India India Indonesia Indonesia Kenya Kenya
H indica + + + 0 + 0LN2
LN 4 + + 0 0 + +INA H1 + + + + + 0
INA H23 0 0 + + 0 0Ki3 + + + 0 + +K4A 0 0 0 0 + +
+ cross resulted in fertile progeny0 no progeny detected
CROSS BREEDING ANALYSIS
Selectedcross-breedingresults are presented in Table2All the Indian isolates are interfertile The Kenyan iso-
lates which differed from H indica in their Hinf I ITSrestriction prole and their HaeIII IGS restriction proleare interfertile amongst each other but only one Kenyanisolate Ki3 produced viable progeny when crossed withthe H indica LN2 type species The Indonesian isolatesare interfertile but surprisingly only one of the Indone-sian isolates tested (INA H1) produced viable hybridswhen crossed with H indica LN2 The INA H1 isolatewas also interfertile with the Kenyan isolate Ki3
Discussion
The results presented here conrm and extend prevous studies which show that a combination of molecudiagnostictoolscan be reliably used for species identiction in Heterorhabditis Isolates of Heterorhabditis froIndia Kenya Indonesia and Cuba hybridised to the H i
dica specic satellite DNA probe (Grenier et al 199Abadon et al 1998) H indica specic DNA restrictiproles were also obtained for all the tropical isolat
of Heterorhabditis investigated here when the ITS rDNregion was restricted using the diagnostic restriction ezymes AluI and MboI (Joyce et al 1994a) The utilof restriction digests of the rDNA ITS region in specidiagnosis of EPN has been conrmed in several studi(Joyce et al 1994a b Miduturi et al 1996 Hominickal 1997 Yoshida et al 1998 Grifn et al 1999a Pa
jav et al 1999) Restriction of the rDNA IGS region wi AluI also yielded a H indica specic restriction prole fall the tropical isolates tested in this study
The PCR amplication product which we obtained fthe H indica rDNA IGS varied in size from 12 to 16 kHominick et al (1997) also observed length heterogenity in the rDNA IGS region among geographic isolatof single Steinernema spp and they suggested that thheterogeneity may make restriction proles of this DNfragment unreliable for species identication The lengheterogeneity which we detected in the H indica IGPCR amplication product did not affect the AluI restrtion prole of the product and a clear-cut seven restrictifragment pattern was common to all the H indica isolat
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 811
CM Stack et al
Fig 7 A Slot blot analysis of genomic DNA (100 ng) of the
Heterorhabditis isolates using a H indica specic satellite DNA
probe B The membrane used for Fig 7A was stripped and
reprobed using an rDNA 18S probe as a loading control A1
H indica LN2 A2 D1 A3 P2M A4 LN2B A5 LN4 A6
INA H23 B1 INA H9 B2 INA H17 B3 INA H1 B4 Ki3
B5 K4A B6 H bacteriophora HP88 C1 H zealandica NZH3
C2 H marelatus OH-10 C3 H hepialus Bodega Bay C4
Heterorhabditis lsquoIrish typersquo K122
tested When the IGS region was digested with HaeIII H indica LN2 displayed a distinctive ve fragment pro-le All of the H indica isolates with the exception of the Kenyan isolates also displayed a ve fragment pro-le Four of these fragments were conserved among theisolates but the largest fragment (of ca 740 in H indica
LN2) was polymorphicamong the H indica isolates Thisfragment which seems to be associated with the lengthpolymorphism in the H indica IGS region is the basisof the D1D1a polymorphism observed by Curran and
Driver (1994) In most species studied to date the rDNIGS contains tandem arrays of subrepeats (Gerbi 198Williams et al 1990 Vahidi amp Honda 1991 Novakal 1993 Linares et al 1994 Crease 1995) This rsults in the length of the IGS region being variable bobetween and within species The length heterogeneity
the H indica IGS PCR product did not affect the AluI rstriction prole of this fragment which suggests that Al
has a restriction site within a tandem repetitive unit in trDNA IGS of H indica In contrast HaeIII appears nto cut within this tandem repeat region with the result thone of the HaeIII restriction fragments is polymorphiclength presumably depending on the number of AluI rpeat units which it contains
The HaeIII rDNA IGS restriction prole of the Disolate is similar to that of H indica LN2 but the Hae
pattern of the Kenyan isolates is unique The large Hae
fragment of the Kenyan isolate K4A is the same size that of D1a thus the Kenyan isolates appear to represea distinct subgroup within D1a Tropical isolates froDarwin (North Australia) Puerto Rico Hawaii the VirgIslands and Egypt were found by Curran and Driv(1994) to possess the D1 prole while the D1a prowas described by these authors from isolates collectin Puerto Rico Jamaica Florida the Virgin Islands aEgypt Comparison of the D1 and D1a HaeIII and Al
rDNA IGS restriction proles with those of H indi
LN2 indicates that the isolates designated D1 and D
by Curran and Driver (1994) belong to H indicaAn important step towards achieving an effective nmatode bacterium complex for pest control is to seek nurally occurring endemic EPN isolates as such isolatare likely to possess physiological traits that are adaptto local climatic and ecological conditions The Kenyisolates described in this study clearly belong to a distinsub-group of H indica and it is probable that these islates also share a distinctive phenotype The two Kenyisolates are interfertile but only one of these isolates (Kiproduced viable progeny when crossed with the H indi
LN2 type species All of the Indonesian Heterorhabdi
isolates were indistinguishable from H indica in the dagnostic molecular tests and they were interfertile wieach other but only one of the Indonesian isolates (INH1) was interfertile with H indica LN2 and INA H1 walso interfertile with the Ki3 isolate from Kenya Theresults suggest that although H indica has a global dtribution in tropical and subtropical regions of the worgeneow within the species may be quite restricted Whave previously observedreproductiveincompatibilityb
484 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 911
Isolates of Heterorhabditisindi
tween isolates of H bacteriophora from Europe (Grifnet al 1999a) and from North America (Dix et al un-publ) We have not determined whether this reproductiveisolation is caused by genetic means or is the result of cy-toplasmic factors Cytoplasmic factors such as endosym-biont bacteria (see reviews by Werren 1997 Johanowicz
amp Hoy 1998) and chromosomal factors such as trans-posons (reviewed by Kidwell 1990 Petrov et al 1995)have been shown to be frequent causes of reproductivein-compatibility in arthropods Endosymbiont bacteria haverecently been shown to cause reproductiveincompatibilityin larial nematodes (Hoerauf et al 1999) Since the re-productive incompatibilityobserved in this study does notyield clear groups of compatible and incompatiblestrainsand in view of the consistencyof the DNA based diagnos-tic tests any assignment of biological species within the H indica group would be injudicious at present Adams
(1988) evaluates phylogeneticand biologicalspecies con-cepts in the delimitationof species in the H bacteriophora
groupThe global distribution of H indica throughout the
tropics and subtropics suggests that H indica possessesa range of phenotypic characters which give it an advan-tage in this climatic zone Such phenotypes might includeadaptationto high temperatures desiccation tolerance andgood dispersal ability It is unlikely that the widespreaddistribution of H indica in the tropics arises because thisspecies was extant before the continents began breakingup and drifting The best supported hypothesis of phylo-genetic relationships among Heterorhabditis species in aphylogeny derived from the rDNA ITS 1 DNA sequencedata puts H indica as the most ancient Heterorhabditis
lineage (Adams et al 1998) However the phylogenetictree for Heterorhabditis and Steinernema of Reid (1994)shows that the genetic distance between Heterorhabdi-
tis species is considerably less than that between Stein-
ernema species and given the small genetic distance de-tected by Reid (1994) between the most divergent of the Heterorhabditis species in his study (a D value of ca
004 as calculated by the method of Nei and Li 1979) it
seems unlikely that H indica speciated before the Juras-sicCretaceousbreak up of Pangaea(an estimated 200 mil-lion years ago)
Acknowledgement
We acknowledge the support of the European Commu-nity STD-3 Programme (TS3 CT 940273)
References
ABADON M GRENIER E LAUMOND C amp ABAD (1998) A species-specic satellite DNA from the enmopathogenic nematode Heterorhabditis indicus Geno
41 148-153
ADAMS BJ (1998) Species concepts and the evolutionparadigm in modern nematology Journal of Nematology 31-21
ADAMS BJ BURNELL AM amp POWERS TO (1998)phylogenetic analysis of the genus Heterorhabditis(NemaRhabditidae) based on internal transcribed spacer 1 DNsequence data Journal of Nematology 30 22-39
AMARASINGHE LD HOMINICK WM BRISCOE Bamp REI D AP (1994) Occurrence and distribution entomopathogenic nematodes in Sri Lanka Journal
Helminthology 68 277-286ARTEAGA HERNANDEZ E M amp MRAacute IumlCE K Z (1984) H
terorhabditis heliothidis a parasite of insect pests in CubFolia Parasitologia(Praha) 31 11-17
BLAXTER ML DE LE Y P GAREY J R LIU LXSCHELDEMAN P VIERSTRAETE A VANFLETERE
JR MACKEY LY DORRIS M FRISSE LM VID
JT amp THOMAS WK (1998) A molecular evolutionaframework for the phylum Nematoda Nature 392 71-75
CONSTANT P MARCHAY L F ISHER-LE SAU X MBRIAND-PANOMA S amp MAULEacuteON H (1998)Naturalocurrence of entomopathogenic nematodes( RhabditidaSteernematidae and Heterorhabditidae) in Guadeloupe islanFundamental and Applied Nematology 21 667-672
CREASE TJ (1995) Ribosomal DNA evolution at tpopulation-level mdash nucleotide variation in intergenic spaarrays of Daphnia pulex Genetics 141 1327-1337
CURRAN J amp DRIVER F (1994) Molecular taxonomy of Hterorhabditis In Burnell AM Ehlers R-U amp Masson J(Eds) Genetics of Entomopathogenic Nematode-Bacteri
Complexes Luxembourg European Commission PublicatiEUR 15681 EN pp 178-187
DIX I BURNELL A M GRIFFIN CT JOYCE SAamp DOWNES MJ (1992) The identication of biologispecies in the genus Heterorhabditis (Nematoda Heterhabditae) by cross-breeding second generation amphimicadults Parasitology104 509-518
EHLERS R-U (1996) Current and future use of nematodin biocontrolpractice and commercial aspects with regardregulatory policy issues Biocontrol Science and Technolo
6 303-317GERBI SA (1985) Evolution of ribosomal DNA In MacI
tyre RJ (Ed) Molecular evolutionary genetics New YoNY USA Plenum Press pp 419-517
GRENIER E BONIFASSI E ABAD P amp LAUMOND (1996a) Use of species specic satellite DNAs as dianostic probes in the identication of Steinernematidae a
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1011
CM Stack et al
Heterorhabditidae entomopathogenic nematodes Parasitol-
ogy 113 483-489GRENIER E LAUMOND C amp ABAD P (1996b) Molecu-
lar characterisationof two species-specic tandemly repeatedDNAs from entomopathogenic nematodes Steinernema and
Heterorhabditis(NematodaRhabditida) Molecularand Bio-
chemical Parasitology 83 47-56GRIFFIN CT JOYCE S A DIX I BURNELL AM amp
DOWNES MJ (1994)Characterisationof the entomopatho-genic nematode Heterorhabditis (Nematoda Heterorhabdi-tidae) from Ireland and Britain by molecular and cross-breeding techniques and the occurrence of the genus in theseislands Fundamental and Applied Nematology 17 245-253
GRIFFIN CT DIX I JOYCE SA BURNELL AM ampDOWNES MJ (1999a) Isolation and characterisation of
Heterorhabditis spp from Hungary Estonia and Denmark Nematology 1 321-332
GRIFFIN CT CHAERANI R FALLON D REI D AP
amp DOWNES MJ (1999b) Occurrence and distribution of the entomopathogenic nematodes Steinernema spp and Het-
erorhabditis indica in Indonesia Journal of Helminthology
74 143-150HOERAUF A NISSENPAHLE K SCHMETZ C HEN-
KLEDUHRSEN K BLAXTER ML BUTTNER DWGALLIN MY ALQAOUD KM LUCIUS R amp FLEIS-CHER B (1999) Tetracycline therapy targets intracellularbacteria in the larial nematode Litomososoides sigmodontis
and results in larial infertility Journal of Clinical Investiga-
tion 103 11-17HOMINICK WM REID AP BOHAN DA amp BRISCOE
BR (1996) Entomopathogenicnematodes biodiversity ge-ographicaldistributionand conventionon biological diversity
Biocontrol Science and Technology 6 317-332HOMINICK WM BRISCOE BR DE L PIN O FG HEN G
J HUN T DJ KOZODOY E MRAacute IumlCE K Z NGUYENKB REI D AP SPIRIDONOV S STOCK P STURHAN D WATURU C amp YOSHIDA M (1997) Biosystematics of entomopathogenic nematodes current status protocols anddenitions Journal of Helminthology 71 271-298
JOHANOWICZ D L amp HOY MA (1998) The manipula-tion of arthropodreproductionby Wolbachia endosymbiontsFlorida Entomologist 81 310-317
JOYCE S A BURNELL A M amp P OWERS TO (1994a)The characterisationof Heterorhabditisisolatesby restrictionenzyme analysis of PCR amplied fragments of mtDNA andrDNA genes Journal of Nematology 26 260-270
JOYCE SA REI D A amp CURRAN J (1994b) Applicationof the polymerase chain reaction (PCR) methods to the iden-tication of entomopathogenicnematodes In Burnell AMEhlers R-U amp Masson JP (Eds) Genetics of entomopatho-
genic nematode-bacterium complexes Luxembourg Euro-pean Commission Publication EUR 15681 EN pp 178-187
KAYA H K (1990)Soil ecology In Gaugler R amp Kaya H(Eds) Entomopathogenic nematodes in biological contr
Boca Raton FL USA CRC Press pp 93-115KAYA HK amp GAUGLER R (1993) Entomopathogenic n
matodes Annual Review of Entomology 38 181-206KIDWELL M G (1990) Evolutionaryaspects of hybrid dysg
nesis in Drosophila Canadian Journal of Zoology 68 1711726LINARES AR BOWEN T amp DOVER GA (1994) A
pects of non-random turnover involved in the concerted evlution of intergenic spacers within the ribosomal DNA
Drosophila melanogaster Journal of Molecular Evoluti
39 151-159LIU J (1994) [A new species of the genus Heterorhabd
from China (Rhabditida Heterorhabditidae)] Acta Zoota
nomica Sinica 19 268-272LIU J amp BERRY RE (1996) Heterorhabditis marela
nsp (Rhabditida Heterorhabditidae) from Oregon Journ
of Invertebrate Pathology
67 48-54MASON JM RAZAK AR amp WRIGHT DJ (1996) Trecoveryof entomopathogenicnematodes from selectedarewithin Peninsular Malaysia Journal of Helminthology 7303-307
MENTI H WRIGHT DJ amp PERRY RN (1997)Desiccatsurvival of populations of the entomopathogenic nematodSteinernema feltiaeand Heterorhabditismegidis from Greeand the UK Journal of Helminthology 71 41-46
MIDUTURI JS MOENS M HOMINICK WM BRISCO
B R amp RE ID AP (1996) Naturally occurring enmopathogenic nematodes in the province of West-FlandeBelgium Journal of Helminthology 70 319-327
NE I M amp LI W-H (1979) Mathematical model for studygenetic variation in terms of restriction endonucleases P
ceedings of the National Academy of Sciences of the Uni
States 76 5269-5273NOVAK EM DE MELLO MP GOMES HBM
GALINDO I GUEVARA P RAMIREZ JL amp DA SILV
RI A JF (1993) Repetitive sequences in the ribosomal intgenic spacer of Trypanosoma cruzi Molecular and Bioche
ical Parasitology 60 273-280PAMJAV H TRIGA D BUZAS Z VELLAI T LUCSKA
A ADAMS B REI D AP BURNELL AM GRIFF
C GLAZER I KLEIN MG amp FODOR A (1999) Nov
application of PhastSystem polyacrylamide gel electrophosis using restriction fragment length polymorphism-intertranscribed spacer patterns of individuals for molecular idetication of entomopathogenic nematodes Electrophore
201266-1273PETROV DA SCHUTZMAN J L HARTL D L amp L
ZOVSKAYA ER ( 1995) Diverse transposable elements amobilised in hybrid dysgenesis in Drosophila virilis P
ceedings of the National Academy of Sciences of the Uni
States 92 8050-8054
486 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1111
Isolates of Heterorhabditisindi
POINAR GO JR (1990) Taxonomy and biology of Stein-ernematidae and HeterorhabditidaeIn Gaugler R amp KayaHK (Eds) Entomopathogenic nematodes in biological con-
trol Boca Raton FL USA CRC Press pp 23-64POINAR GO JR (1993) Origins and phylogenetic relation-
ships of the entomophilic rhabditids Heterorhabditis and
Steinernema Fundamental and Applied Nematology16 333-338POINAR GO JR JACKSON T amp KLEIN M (1987) He-
terorhabditis megidis sp n (Heterorhabditidae Rhabditida)parasitic in the Japanese beetlePopilliajaponica(ScarabidaeColeoptera) in Ohio Proceedings of the Helminthological
Society of Washington 53 53-59POINAR GO JR KARUNAKAR GK amp DAVID H (1992)
Heterorhabditis indicus n sp (Rhabditida Nematoda) fromIndia Separation of Heterorhabditis spp by infective juve-niles Fundamental and Applied Nematology 15 467-472
REI D AP (1994) Molecular taxonomy of Steinernema InBurnell AM Ehlers R-U amp Masson JP (Eds) Genetics
of entomopathogenic nematode-bacterium complexes Lux-embourg European Commission Publication EUR 15681EN pp 178-187
SMITS P GROENEN JTM amp DERAAY G (1991) Charac-terisation of Heterorhabditis isolates using DNA restrictionlength polymorphism Revue de Neacutematologie 14 445-453
STOCK SP (1993) A new species of thegenus Heterorhabdi-
tis Poinar 1976 (Nematoda Heterorhabditidae) parasitizingGraphognathus sp larvae (Coteoptera Curculionidae) fromArgentina Research and Reviews in Parasitology 53 103-107
STOCK SP (1997) Heterorhabditis hepialus Stock Strong amp
Gardner 1996a junior synonym of H marelatus Liu amp Berry1996 (Rhabditida Rhabditidae) with a redescription of thespecies Nematologica 43 455-463
STOCK SP STRONG D amp GARDNER S (1996) Identiction of Heterorhabditis (Nematoda Heterorhabditidae)froCalifornia with a new species isolated from the larvae of tghost moth Hepialius [sic] californicus (Lepidoptera Healidae) from Bodega Bay Natural Reserve Fundamental a
Applied Nematology 19 585-592
STRONG DR KAYA HK WHIPPLE AV CHILD ALKRAIG S BONDONNE M DYE R K amp MARON J(1996) Entomopathogenic nematodes natural enemiesroot feeding caterpillarson bush lupine Oecologica 108 345
SUDHAUS W (1993) Die mittels symbiontischerBakterienetomopathogenen Nematoden Gattungen Heterorhabditis aSteinernema sind keine Schwestertaxa Verhandlungen d
Deutschen Zoologischen Gesellschaft 86 146VAHIDI H amp HONDA BM (1991)Repeats and subrepeats
the intergenic spacer of rDNA from the nematode Meloid
gyne arenaria Molecular and General Genetics 227 33
336WERREN JH (1997) Biology of Wolbachia Annual Revi
of Entomology 42 587-609WILLIAMS S M ROBBINS L G CLUSTER PD A
LARD RW amp STROBECK C (1990) Superstructureof t Drosophila ribosomal gene family Proceedings of the N
tional Academy of Sciences of the United States 87 3153160
WOUTS WM (1979) The biology and life cycle ofNew Zealand population of Heterorhabditis heliothidis (Hterorhabditidae)Nematologica 25 191-202
YOSHIDA M REID AP BRISCOE BR amp HOMINIC
WM (1998) Survey of entomopathogenic nematod(Rhabditida Steinernematidae and Heterorhabditidae) Japan Fundamental and Applied Nematology 21 185-198
Vol 2(5) 2000 4
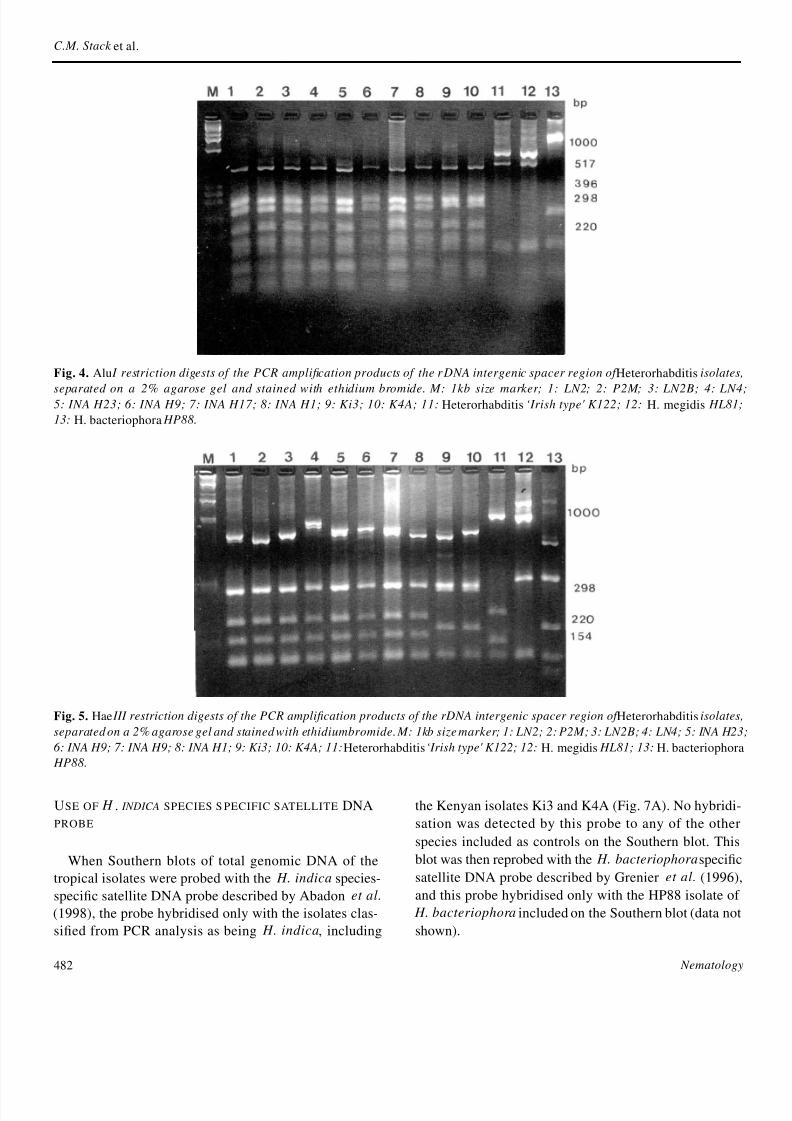
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 611
CM Stack et al
Fig 4 Alu I restriction digests of the PCR amplication products of the r DNA intergenic spacer region of Heterorhabditis isolat
separated on a 2 agarose gel and stained with ethidium bromide M 1kb size marker 1 LN2 2 P2M 3 LN2B 4 LN5 INA H23 6 INA H9 7 INA H17 8 INA H1 9 Ki3 10 K4A 11 Heterorhabditis lsquoIrish typersquo K122 12 H megidis HL8
13 H bacteriophora HP88
Fig 5 Hae III restriction digests of the PCR amplication products of the rDNA intergenic spacer region of Heterorhabditis isolat
separated on a 2 agarose gel and stained with ethidiumbromide M 1kb size marker 1 LN2 2 P2M 3 LN2B 4 LN4 5 INA H
6 INA H9 7 INA H9 8 INA H1 9 Ki3 10 K4A 11HeterorhabditislsquoIrish typersquo K122 12 H megidis HL81 13 H bacterioph HP88
USE OF H INDICA SPECIES S PECIFIC SATELLITE DNAPROBE
When Southern blots of total genomic DNA of thetropical isolates were probed with the H indica species-specic satellite DNA probe described by Abadon et al
(1998) the probe hybridised only with the isolates clas-sied from PCR analysis as being H indica including
the Kenyan isolates Ki3 and K4A (Fig 7A) No hybrisation was detected by this probe to any of the othspecies included as controls on the Southern blot Thblot was then reprobed with the H bacteriophora specisatellite DNA probe described by Grenier et al (199and this probe hybridised only with the HP88 isolate H bacteriophora included on the Southern blot (data nshown)
482 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 711
Isolates of Heterorhabditisindi
Fig 6 Hae III and Alu I restriction digests of the PCR amplication products of the rDNA intergenic spacer region of Heterorhabdisolates including isolates designated type D1a by Curran and Driver (1994) M 1kb size marker 1 H indica LN2 2 D1 3 K4
4 ES10 5 JAM79 6 ST09 7 FLGS10 M 1 kb marker 9 H indica LN2 10 D1 11 ES10 12 JAM79 13 ST09 14 FLGS1
Lanes 1 to 7 contain Hae III digests and lanes 9 to 14 contain Alu I digests The digests were separated on a 2 agarose gel and stain
with ethidium bromide
Table 2 The results of cross-breeding experiments of six Het-erorhabditis isolates from India Indonesia and Kenya
Female Male
H indica LN 4 INA H1 INA H23 Ki3 K4ALN2India India Indonesia Indonesia Kenya Kenya
H indica + + + 0 + 0LN2
LN 4 + + 0 0 + +INA H1 + + + + + 0
INA H23 0 0 + + 0 0Ki3 + + + 0 + +K4A 0 0 0 0 + +
+ cross resulted in fertile progeny0 no progeny detected
CROSS BREEDING ANALYSIS
Selectedcross-breedingresults are presented in Table2All the Indian isolates are interfertile The Kenyan iso-
lates which differed from H indica in their Hinf I ITSrestriction prole and their HaeIII IGS restriction proleare interfertile amongst each other but only one Kenyanisolate Ki3 produced viable progeny when crossed withthe H indica LN2 type species The Indonesian isolatesare interfertile but surprisingly only one of the Indone-sian isolates tested (INA H1) produced viable hybridswhen crossed with H indica LN2 The INA H1 isolatewas also interfertile with the Kenyan isolate Ki3
Discussion
The results presented here conrm and extend prevous studies which show that a combination of molecudiagnostictoolscan be reliably used for species identiction in Heterorhabditis Isolates of Heterorhabditis froIndia Kenya Indonesia and Cuba hybridised to the H i
dica specic satellite DNA probe (Grenier et al 199Abadon et al 1998) H indica specic DNA restrictiproles were also obtained for all the tropical isolat
of Heterorhabditis investigated here when the ITS rDNregion was restricted using the diagnostic restriction ezymes AluI and MboI (Joyce et al 1994a) The utilof restriction digests of the rDNA ITS region in specidiagnosis of EPN has been conrmed in several studi(Joyce et al 1994a b Miduturi et al 1996 Hominickal 1997 Yoshida et al 1998 Grifn et al 1999a Pa
jav et al 1999) Restriction of the rDNA IGS region wi AluI also yielded a H indica specic restriction prole fall the tropical isolates tested in this study
The PCR amplication product which we obtained fthe H indica rDNA IGS varied in size from 12 to 16 kHominick et al (1997) also observed length heterogenity in the rDNA IGS region among geographic isolatof single Steinernema spp and they suggested that thheterogeneity may make restriction proles of this DNfragment unreliable for species identication The lengheterogeneity which we detected in the H indica IGPCR amplication product did not affect the AluI restrtion prole of the product and a clear-cut seven restrictifragment pattern was common to all the H indica isolat
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 811
CM Stack et al
Fig 7 A Slot blot analysis of genomic DNA (100 ng) of the
Heterorhabditis isolates using a H indica specic satellite DNA
probe B The membrane used for Fig 7A was stripped and
reprobed using an rDNA 18S probe as a loading control A1
H indica LN2 A2 D1 A3 P2M A4 LN2B A5 LN4 A6
INA H23 B1 INA H9 B2 INA H17 B3 INA H1 B4 Ki3
B5 K4A B6 H bacteriophora HP88 C1 H zealandica NZH3
C2 H marelatus OH-10 C3 H hepialus Bodega Bay C4
Heterorhabditis lsquoIrish typersquo K122
tested When the IGS region was digested with HaeIII H indica LN2 displayed a distinctive ve fragment pro-le All of the H indica isolates with the exception of the Kenyan isolates also displayed a ve fragment pro-le Four of these fragments were conserved among theisolates but the largest fragment (of ca 740 in H indica
LN2) was polymorphicamong the H indica isolates Thisfragment which seems to be associated with the lengthpolymorphism in the H indica IGS region is the basisof the D1D1a polymorphism observed by Curran and
Driver (1994) In most species studied to date the rDNIGS contains tandem arrays of subrepeats (Gerbi 198Williams et al 1990 Vahidi amp Honda 1991 Novakal 1993 Linares et al 1994 Crease 1995) This rsults in the length of the IGS region being variable bobetween and within species The length heterogeneity
the H indica IGS PCR product did not affect the AluI rstriction prole of this fragment which suggests that Al
has a restriction site within a tandem repetitive unit in trDNA IGS of H indica In contrast HaeIII appears nto cut within this tandem repeat region with the result thone of the HaeIII restriction fragments is polymorphiclength presumably depending on the number of AluI rpeat units which it contains
The HaeIII rDNA IGS restriction prole of the Disolate is similar to that of H indica LN2 but the Hae
pattern of the Kenyan isolates is unique The large Hae
fragment of the Kenyan isolate K4A is the same size that of D1a thus the Kenyan isolates appear to represea distinct subgroup within D1a Tropical isolates froDarwin (North Australia) Puerto Rico Hawaii the VirgIslands and Egypt were found by Curran and Driv(1994) to possess the D1 prole while the D1a prowas described by these authors from isolates collectin Puerto Rico Jamaica Florida the Virgin Islands aEgypt Comparison of the D1 and D1a HaeIII and Al
rDNA IGS restriction proles with those of H indi
LN2 indicates that the isolates designated D1 and D
by Curran and Driver (1994) belong to H indicaAn important step towards achieving an effective nmatode bacterium complex for pest control is to seek nurally occurring endemic EPN isolates as such isolatare likely to possess physiological traits that are adaptto local climatic and ecological conditions The Kenyisolates described in this study clearly belong to a distinsub-group of H indica and it is probable that these islates also share a distinctive phenotype The two Kenyisolates are interfertile but only one of these isolates (Kiproduced viable progeny when crossed with the H indi
LN2 type species All of the Indonesian Heterorhabdi
isolates were indistinguishable from H indica in the dagnostic molecular tests and they were interfertile wieach other but only one of the Indonesian isolates (INH1) was interfertile with H indica LN2 and INA H1 walso interfertile with the Ki3 isolate from Kenya Theresults suggest that although H indica has a global dtribution in tropical and subtropical regions of the worgeneow within the species may be quite restricted Whave previously observedreproductiveincompatibilityb
484 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 911
Isolates of Heterorhabditisindi
tween isolates of H bacteriophora from Europe (Grifnet al 1999a) and from North America (Dix et al un-publ) We have not determined whether this reproductiveisolation is caused by genetic means or is the result of cy-toplasmic factors Cytoplasmic factors such as endosym-biont bacteria (see reviews by Werren 1997 Johanowicz
amp Hoy 1998) and chromosomal factors such as trans-posons (reviewed by Kidwell 1990 Petrov et al 1995)have been shown to be frequent causes of reproductivein-compatibility in arthropods Endosymbiont bacteria haverecently been shown to cause reproductiveincompatibilityin larial nematodes (Hoerauf et al 1999) Since the re-productive incompatibilityobserved in this study does notyield clear groups of compatible and incompatiblestrainsand in view of the consistencyof the DNA based diagnos-tic tests any assignment of biological species within the H indica group would be injudicious at present Adams
(1988) evaluates phylogeneticand biologicalspecies con-cepts in the delimitationof species in the H bacteriophora
groupThe global distribution of H indica throughout the
tropics and subtropics suggests that H indica possessesa range of phenotypic characters which give it an advan-tage in this climatic zone Such phenotypes might includeadaptationto high temperatures desiccation tolerance andgood dispersal ability It is unlikely that the widespreaddistribution of H indica in the tropics arises because thisspecies was extant before the continents began breakingup and drifting The best supported hypothesis of phylo-genetic relationships among Heterorhabditis species in aphylogeny derived from the rDNA ITS 1 DNA sequencedata puts H indica as the most ancient Heterorhabditis
lineage (Adams et al 1998) However the phylogenetictree for Heterorhabditis and Steinernema of Reid (1994)shows that the genetic distance between Heterorhabdi-
tis species is considerably less than that between Stein-
ernema species and given the small genetic distance de-tected by Reid (1994) between the most divergent of the Heterorhabditis species in his study (a D value of ca
004 as calculated by the method of Nei and Li 1979) it
seems unlikely that H indica speciated before the Juras-sicCretaceousbreak up of Pangaea(an estimated 200 mil-lion years ago)
Acknowledgement
We acknowledge the support of the European Commu-nity STD-3 Programme (TS3 CT 940273)
References
ABADON M GRENIER E LAUMOND C amp ABAD (1998) A species-specic satellite DNA from the enmopathogenic nematode Heterorhabditis indicus Geno
41 148-153
ADAMS BJ (1998) Species concepts and the evolutionparadigm in modern nematology Journal of Nematology 31-21
ADAMS BJ BURNELL AM amp POWERS TO (1998)phylogenetic analysis of the genus Heterorhabditis(NemaRhabditidae) based on internal transcribed spacer 1 DNsequence data Journal of Nematology 30 22-39
AMARASINGHE LD HOMINICK WM BRISCOE Bamp REI D AP (1994) Occurrence and distribution entomopathogenic nematodes in Sri Lanka Journal
Helminthology 68 277-286ARTEAGA HERNANDEZ E M amp MRAacute IumlCE K Z (1984) H
terorhabditis heliothidis a parasite of insect pests in CubFolia Parasitologia(Praha) 31 11-17
BLAXTER ML DE LE Y P GAREY J R LIU LXSCHELDEMAN P VIERSTRAETE A VANFLETERE
JR MACKEY LY DORRIS M FRISSE LM VID
JT amp THOMAS WK (1998) A molecular evolutionaframework for the phylum Nematoda Nature 392 71-75
CONSTANT P MARCHAY L F ISHER-LE SAU X MBRIAND-PANOMA S amp MAULEacuteON H (1998)Naturalocurrence of entomopathogenic nematodes( RhabditidaSteernematidae and Heterorhabditidae) in Guadeloupe islanFundamental and Applied Nematology 21 667-672
CREASE TJ (1995) Ribosomal DNA evolution at tpopulation-level mdash nucleotide variation in intergenic spaarrays of Daphnia pulex Genetics 141 1327-1337
CURRAN J amp DRIVER F (1994) Molecular taxonomy of Hterorhabditis In Burnell AM Ehlers R-U amp Masson J(Eds) Genetics of Entomopathogenic Nematode-Bacteri
Complexes Luxembourg European Commission PublicatiEUR 15681 EN pp 178-187
DIX I BURNELL A M GRIFFIN CT JOYCE SAamp DOWNES MJ (1992) The identication of biologispecies in the genus Heterorhabditis (Nematoda Heterhabditae) by cross-breeding second generation amphimicadults Parasitology104 509-518
EHLERS R-U (1996) Current and future use of nematodin biocontrolpractice and commercial aspects with regardregulatory policy issues Biocontrol Science and Technolo
6 303-317GERBI SA (1985) Evolution of ribosomal DNA In MacI
tyre RJ (Ed) Molecular evolutionary genetics New YoNY USA Plenum Press pp 419-517
GRENIER E BONIFASSI E ABAD P amp LAUMOND (1996a) Use of species specic satellite DNAs as dianostic probes in the identication of Steinernematidae a
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1011
CM Stack et al
Heterorhabditidae entomopathogenic nematodes Parasitol-
ogy 113 483-489GRENIER E LAUMOND C amp ABAD P (1996b) Molecu-
lar characterisationof two species-specic tandemly repeatedDNAs from entomopathogenic nematodes Steinernema and
Heterorhabditis(NematodaRhabditida) Molecularand Bio-
chemical Parasitology 83 47-56GRIFFIN CT JOYCE S A DIX I BURNELL AM amp
DOWNES MJ (1994)Characterisationof the entomopatho-genic nematode Heterorhabditis (Nematoda Heterorhabdi-tidae) from Ireland and Britain by molecular and cross-breeding techniques and the occurrence of the genus in theseislands Fundamental and Applied Nematology 17 245-253
GRIFFIN CT DIX I JOYCE SA BURNELL AM ampDOWNES MJ (1999a) Isolation and characterisation of
Heterorhabditis spp from Hungary Estonia and Denmark Nematology 1 321-332
GRIFFIN CT CHAERANI R FALLON D REI D AP
amp DOWNES MJ (1999b) Occurrence and distribution of the entomopathogenic nematodes Steinernema spp and Het-
erorhabditis indica in Indonesia Journal of Helminthology
74 143-150HOERAUF A NISSENPAHLE K SCHMETZ C HEN-
KLEDUHRSEN K BLAXTER ML BUTTNER DWGALLIN MY ALQAOUD KM LUCIUS R amp FLEIS-CHER B (1999) Tetracycline therapy targets intracellularbacteria in the larial nematode Litomososoides sigmodontis
and results in larial infertility Journal of Clinical Investiga-
tion 103 11-17HOMINICK WM REID AP BOHAN DA amp BRISCOE
BR (1996) Entomopathogenicnematodes biodiversity ge-ographicaldistributionand conventionon biological diversity
Biocontrol Science and Technology 6 317-332HOMINICK WM BRISCOE BR DE L PIN O FG HEN G
J HUN T DJ KOZODOY E MRAacute IumlCE K Z NGUYENKB REI D AP SPIRIDONOV S STOCK P STURHAN D WATURU C amp YOSHIDA M (1997) Biosystematics of entomopathogenic nematodes current status protocols anddenitions Journal of Helminthology 71 271-298
JOHANOWICZ D L amp HOY MA (1998) The manipula-tion of arthropodreproductionby Wolbachia endosymbiontsFlorida Entomologist 81 310-317
JOYCE S A BURNELL A M amp P OWERS TO (1994a)The characterisationof Heterorhabditisisolatesby restrictionenzyme analysis of PCR amplied fragments of mtDNA andrDNA genes Journal of Nematology 26 260-270
JOYCE SA REI D A amp CURRAN J (1994b) Applicationof the polymerase chain reaction (PCR) methods to the iden-tication of entomopathogenicnematodes In Burnell AMEhlers R-U amp Masson JP (Eds) Genetics of entomopatho-
genic nematode-bacterium complexes Luxembourg Euro-pean Commission Publication EUR 15681 EN pp 178-187
KAYA H K (1990)Soil ecology In Gaugler R amp Kaya H(Eds) Entomopathogenic nematodes in biological contr
Boca Raton FL USA CRC Press pp 93-115KAYA HK amp GAUGLER R (1993) Entomopathogenic n
matodes Annual Review of Entomology 38 181-206KIDWELL M G (1990) Evolutionaryaspects of hybrid dysg
nesis in Drosophila Canadian Journal of Zoology 68 1711726LINARES AR BOWEN T amp DOVER GA (1994) A
pects of non-random turnover involved in the concerted evlution of intergenic spacers within the ribosomal DNA
Drosophila melanogaster Journal of Molecular Evoluti
39 151-159LIU J (1994) [A new species of the genus Heterorhabd
from China (Rhabditida Heterorhabditidae)] Acta Zoota
nomica Sinica 19 268-272LIU J amp BERRY RE (1996) Heterorhabditis marela
nsp (Rhabditida Heterorhabditidae) from Oregon Journ
of Invertebrate Pathology
67 48-54MASON JM RAZAK AR amp WRIGHT DJ (1996) Trecoveryof entomopathogenicnematodes from selectedarewithin Peninsular Malaysia Journal of Helminthology 7303-307
MENTI H WRIGHT DJ amp PERRY RN (1997)Desiccatsurvival of populations of the entomopathogenic nematodSteinernema feltiaeand Heterorhabditismegidis from Greeand the UK Journal of Helminthology 71 41-46
MIDUTURI JS MOENS M HOMINICK WM BRISCO
B R amp RE ID AP (1996) Naturally occurring enmopathogenic nematodes in the province of West-FlandeBelgium Journal of Helminthology 70 319-327
NE I M amp LI W-H (1979) Mathematical model for studygenetic variation in terms of restriction endonucleases P
ceedings of the National Academy of Sciences of the Uni
States 76 5269-5273NOVAK EM DE MELLO MP GOMES HBM
GALINDO I GUEVARA P RAMIREZ JL amp DA SILV
RI A JF (1993) Repetitive sequences in the ribosomal intgenic spacer of Trypanosoma cruzi Molecular and Bioche
ical Parasitology 60 273-280PAMJAV H TRIGA D BUZAS Z VELLAI T LUCSKA
A ADAMS B REI D AP BURNELL AM GRIFF
C GLAZER I KLEIN MG amp FODOR A (1999) Nov
application of PhastSystem polyacrylamide gel electrophosis using restriction fragment length polymorphism-intertranscribed spacer patterns of individuals for molecular idetication of entomopathogenic nematodes Electrophore
201266-1273PETROV DA SCHUTZMAN J L HARTL D L amp L
ZOVSKAYA ER ( 1995) Diverse transposable elements amobilised in hybrid dysgenesis in Drosophila virilis P
ceedings of the National Academy of Sciences of the Uni
States 92 8050-8054
486 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1111
Isolates of Heterorhabditisindi
POINAR GO JR (1990) Taxonomy and biology of Stein-ernematidae and HeterorhabditidaeIn Gaugler R amp KayaHK (Eds) Entomopathogenic nematodes in biological con-
trol Boca Raton FL USA CRC Press pp 23-64POINAR GO JR (1993) Origins and phylogenetic relation-
ships of the entomophilic rhabditids Heterorhabditis and
Steinernema Fundamental and Applied Nematology16 333-338POINAR GO JR JACKSON T amp KLEIN M (1987) He-
terorhabditis megidis sp n (Heterorhabditidae Rhabditida)parasitic in the Japanese beetlePopilliajaponica(ScarabidaeColeoptera) in Ohio Proceedings of the Helminthological
Society of Washington 53 53-59POINAR GO JR KARUNAKAR GK amp DAVID H (1992)
Heterorhabditis indicus n sp (Rhabditida Nematoda) fromIndia Separation of Heterorhabditis spp by infective juve-niles Fundamental and Applied Nematology 15 467-472
REI D AP (1994) Molecular taxonomy of Steinernema InBurnell AM Ehlers R-U amp Masson JP (Eds) Genetics
of entomopathogenic nematode-bacterium complexes Lux-embourg European Commission Publication EUR 15681EN pp 178-187
SMITS P GROENEN JTM amp DERAAY G (1991) Charac-terisation of Heterorhabditis isolates using DNA restrictionlength polymorphism Revue de Neacutematologie 14 445-453
STOCK SP (1993) A new species of thegenus Heterorhabdi-
tis Poinar 1976 (Nematoda Heterorhabditidae) parasitizingGraphognathus sp larvae (Coteoptera Curculionidae) fromArgentina Research and Reviews in Parasitology 53 103-107
STOCK SP (1997) Heterorhabditis hepialus Stock Strong amp
Gardner 1996a junior synonym of H marelatus Liu amp Berry1996 (Rhabditida Rhabditidae) with a redescription of thespecies Nematologica 43 455-463
STOCK SP STRONG D amp GARDNER S (1996) Identiction of Heterorhabditis (Nematoda Heterorhabditidae)froCalifornia with a new species isolated from the larvae of tghost moth Hepialius [sic] californicus (Lepidoptera Healidae) from Bodega Bay Natural Reserve Fundamental a
Applied Nematology 19 585-592
STRONG DR KAYA HK WHIPPLE AV CHILD ALKRAIG S BONDONNE M DYE R K amp MARON J(1996) Entomopathogenic nematodes natural enemiesroot feeding caterpillarson bush lupine Oecologica 108 345
SUDHAUS W (1993) Die mittels symbiontischerBakterienetomopathogenen Nematoden Gattungen Heterorhabditis aSteinernema sind keine Schwestertaxa Verhandlungen d
Deutschen Zoologischen Gesellschaft 86 146VAHIDI H amp HONDA BM (1991)Repeats and subrepeats
the intergenic spacer of rDNA from the nematode Meloid
gyne arenaria Molecular and General Genetics 227 33
336WERREN JH (1997) Biology of Wolbachia Annual Revi
of Entomology 42 587-609WILLIAMS S M ROBBINS L G CLUSTER PD A
LARD RW amp STROBECK C (1990) Superstructureof t Drosophila ribosomal gene family Proceedings of the N
tional Academy of Sciences of the United States 87 3153160
WOUTS WM (1979) The biology and life cycle ofNew Zealand population of Heterorhabditis heliothidis (Hterorhabditidae)Nematologica 25 191-202
YOSHIDA M REID AP BRISCOE BR amp HOMINIC
WM (1998) Survey of entomopathogenic nematod(Rhabditida Steinernematidae and Heterorhabditidae) Japan Fundamental and Applied Nematology 21 185-198
Vol 2(5) 2000 4
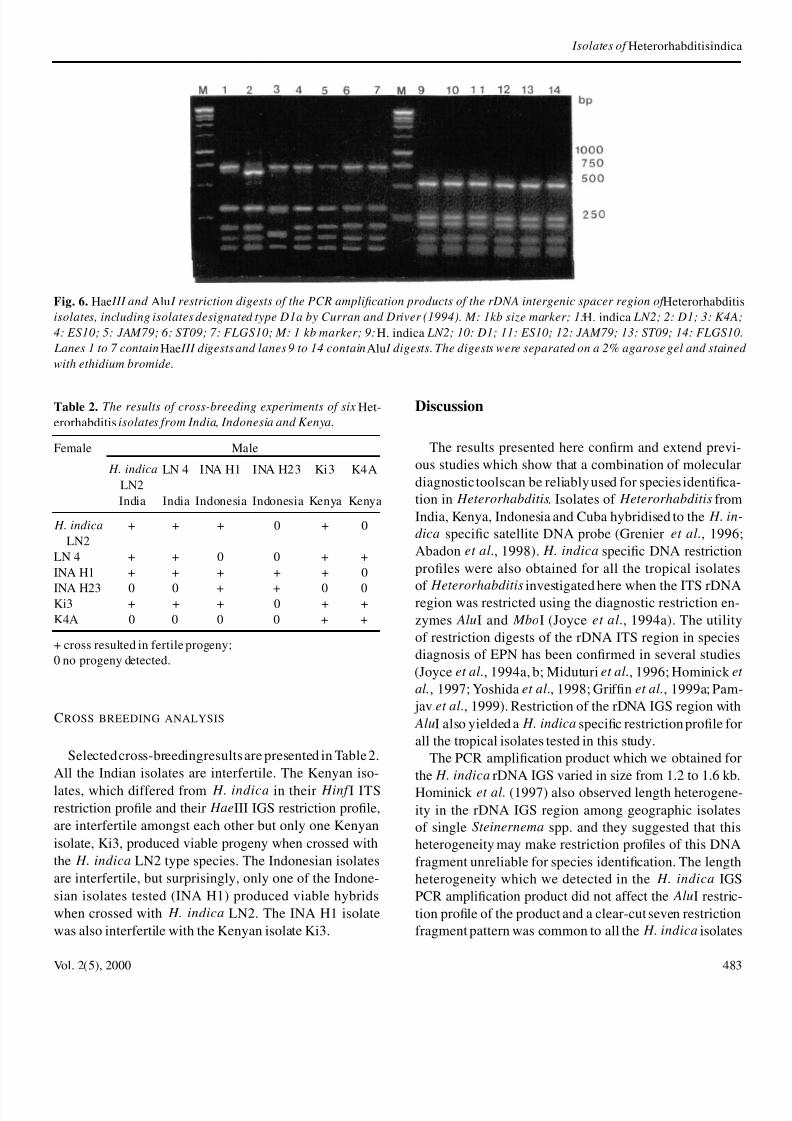
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 711
Isolates of Heterorhabditisindi
Fig 6 Hae III and Alu I restriction digests of the PCR amplication products of the rDNA intergenic spacer region of Heterorhabdisolates including isolates designated type D1a by Curran and Driver (1994) M 1kb size marker 1 H indica LN2 2 D1 3 K4
4 ES10 5 JAM79 6 ST09 7 FLGS10 M 1 kb marker 9 H indica LN2 10 D1 11 ES10 12 JAM79 13 ST09 14 FLGS1
Lanes 1 to 7 contain Hae III digests and lanes 9 to 14 contain Alu I digests The digests were separated on a 2 agarose gel and stain
with ethidium bromide
Table 2 The results of cross-breeding experiments of six Het-erorhabditis isolates from India Indonesia and Kenya
Female Male
H indica LN 4 INA H1 INA H23 Ki3 K4ALN2India India Indonesia Indonesia Kenya Kenya
H indica + + + 0 + 0LN2
LN 4 + + 0 0 + +INA H1 + + + + + 0
INA H23 0 0 + + 0 0Ki3 + + + 0 + +K4A 0 0 0 0 + +
+ cross resulted in fertile progeny0 no progeny detected
CROSS BREEDING ANALYSIS
Selectedcross-breedingresults are presented in Table2All the Indian isolates are interfertile The Kenyan iso-
lates which differed from H indica in their Hinf I ITSrestriction prole and their HaeIII IGS restriction proleare interfertile amongst each other but only one Kenyanisolate Ki3 produced viable progeny when crossed withthe H indica LN2 type species The Indonesian isolatesare interfertile but surprisingly only one of the Indone-sian isolates tested (INA H1) produced viable hybridswhen crossed with H indica LN2 The INA H1 isolatewas also interfertile with the Kenyan isolate Ki3
Discussion
The results presented here conrm and extend prevous studies which show that a combination of molecudiagnostictoolscan be reliably used for species identiction in Heterorhabditis Isolates of Heterorhabditis froIndia Kenya Indonesia and Cuba hybridised to the H i
dica specic satellite DNA probe (Grenier et al 199Abadon et al 1998) H indica specic DNA restrictiproles were also obtained for all the tropical isolat
of Heterorhabditis investigated here when the ITS rDNregion was restricted using the diagnostic restriction ezymes AluI and MboI (Joyce et al 1994a) The utilof restriction digests of the rDNA ITS region in specidiagnosis of EPN has been conrmed in several studi(Joyce et al 1994a b Miduturi et al 1996 Hominickal 1997 Yoshida et al 1998 Grifn et al 1999a Pa
jav et al 1999) Restriction of the rDNA IGS region wi AluI also yielded a H indica specic restriction prole fall the tropical isolates tested in this study
The PCR amplication product which we obtained fthe H indica rDNA IGS varied in size from 12 to 16 kHominick et al (1997) also observed length heterogenity in the rDNA IGS region among geographic isolatof single Steinernema spp and they suggested that thheterogeneity may make restriction proles of this DNfragment unreliable for species identication The lengheterogeneity which we detected in the H indica IGPCR amplication product did not affect the AluI restrtion prole of the product and a clear-cut seven restrictifragment pattern was common to all the H indica isolat
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 811
CM Stack et al
Fig 7 A Slot blot analysis of genomic DNA (100 ng) of the
Heterorhabditis isolates using a H indica specic satellite DNA
probe B The membrane used for Fig 7A was stripped and
reprobed using an rDNA 18S probe as a loading control A1
H indica LN2 A2 D1 A3 P2M A4 LN2B A5 LN4 A6
INA H23 B1 INA H9 B2 INA H17 B3 INA H1 B4 Ki3
B5 K4A B6 H bacteriophora HP88 C1 H zealandica NZH3
C2 H marelatus OH-10 C3 H hepialus Bodega Bay C4
Heterorhabditis lsquoIrish typersquo K122
tested When the IGS region was digested with HaeIII H indica LN2 displayed a distinctive ve fragment pro-le All of the H indica isolates with the exception of the Kenyan isolates also displayed a ve fragment pro-le Four of these fragments were conserved among theisolates but the largest fragment (of ca 740 in H indica
LN2) was polymorphicamong the H indica isolates Thisfragment which seems to be associated with the lengthpolymorphism in the H indica IGS region is the basisof the D1D1a polymorphism observed by Curran and
Driver (1994) In most species studied to date the rDNIGS contains tandem arrays of subrepeats (Gerbi 198Williams et al 1990 Vahidi amp Honda 1991 Novakal 1993 Linares et al 1994 Crease 1995) This rsults in the length of the IGS region being variable bobetween and within species The length heterogeneity
the H indica IGS PCR product did not affect the AluI rstriction prole of this fragment which suggests that Al
has a restriction site within a tandem repetitive unit in trDNA IGS of H indica In contrast HaeIII appears nto cut within this tandem repeat region with the result thone of the HaeIII restriction fragments is polymorphiclength presumably depending on the number of AluI rpeat units which it contains
The HaeIII rDNA IGS restriction prole of the Disolate is similar to that of H indica LN2 but the Hae
pattern of the Kenyan isolates is unique The large Hae
fragment of the Kenyan isolate K4A is the same size that of D1a thus the Kenyan isolates appear to represea distinct subgroup within D1a Tropical isolates froDarwin (North Australia) Puerto Rico Hawaii the VirgIslands and Egypt were found by Curran and Driv(1994) to possess the D1 prole while the D1a prowas described by these authors from isolates collectin Puerto Rico Jamaica Florida the Virgin Islands aEgypt Comparison of the D1 and D1a HaeIII and Al
rDNA IGS restriction proles with those of H indi
LN2 indicates that the isolates designated D1 and D
by Curran and Driver (1994) belong to H indicaAn important step towards achieving an effective nmatode bacterium complex for pest control is to seek nurally occurring endemic EPN isolates as such isolatare likely to possess physiological traits that are adaptto local climatic and ecological conditions The Kenyisolates described in this study clearly belong to a distinsub-group of H indica and it is probable that these islates also share a distinctive phenotype The two Kenyisolates are interfertile but only one of these isolates (Kiproduced viable progeny when crossed with the H indi
LN2 type species All of the Indonesian Heterorhabdi
isolates were indistinguishable from H indica in the dagnostic molecular tests and they were interfertile wieach other but only one of the Indonesian isolates (INH1) was interfertile with H indica LN2 and INA H1 walso interfertile with the Ki3 isolate from Kenya Theresults suggest that although H indica has a global dtribution in tropical and subtropical regions of the worgeneow within the species may be quite restricted Whave previously observedreproductiveincompatibilityb
484 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 911
Isolates of Heterorhabditisindi
tween isolates of H bacteriophora from Europe (Grifnet al 1999a) and from North America (Dix et al un-publ) We have not determined whether this reproductiveisolation is caused by genetic means or is the result of cy-toplasmic factors Cytoplasmic factors such as endosym-biont bacteria (see reviews by Werren 1997 Johanowicz
amp Hoy 1998) and chromosomal factors such as trans-posons (reviewed by Kidwell 1990 Petrov et al 1995)have been shown to be frequent causes of reproductivein-compatibility in arthropods Endosymbiont bacteria haverecently been shown to cause reproductiveincompatibilityin larial nematodes (Hoerauf et al 1999) Since the re-productive incompatibilityobserved in this study does notyield clear groups of compatible and incompatiblestrainsand in view of the consistencyof the DNA based diagnos-tic tests any assignment of biological species within the H indica group would be injudicious at present Adams
(1988) evaluates phylogeneticand biologicalspecies con-cepts in the delimitationof species in the H bacteriophora
groupThe global distribution of H indica throughout the
tropics and subtropics suggests that H indica possessesa range of phenotypic characters which give it an advan-tage in this climatic zone Such phenotypes might includeadaptationto high temperatures desiccation tolerance andgood dispersal ability It is unlikely that the widespreaddistribution of H indica in the tropics arises because thisspecies was extant before the continents began breakingup and drifting The best supported hypothesis of phylo-genetic relationships among Heterorhabditis species in aphylogeny derived from the rDNA ITS 1 DNA sequencedata puts H indica as the most ancient Heterorhabditis
lineage (Adams et al 1998) However the phylogenetictree for Heterorhabditis and Steinernema of Reid (1994)shows that the genetic distance between Heterorhabdi-
tis species is considerably less than that between Stein-
ernema species and given the small genetic distance de-tected by Reid (1994) between the most divergent of the Heterorhabditis species in his study (a D value of ca
004 as calculated by the method of Nei and Li 1979) it
seems unlikely that H indica speciated before the Juras-sicCretaceousbreak up of Pangaea(an estimated 200 mil-lion years ago)
Acknowledgement
We acknowledge the support of the European Commu-nity STD-3 Programme (TS3 CT 940273)
References
ABADON M GRENIER E LAUMOND C amp ABAD (1998) A species-specic satellite DNA from the enmopathogenic nematode Heterorhabditis indicus Geno
41 148-153
ADAMS BJ (1998) Species concepts and the evolutionparadigm in modern nematology Journal of Nematology 31-21
ADAMS BJ BURNELL AM amp POWERS TO (1998)phylogenetic analysis of the genus Heterorhabditis(NemaRhabditidae) based on internal transcribed spacer 1 DNsequence data Journal of Nematology 30 22-39
AMARASINGHE LD HOMINICK WM BRISCOE Bamp REI D AP (1994) Occurrence and distribution entomopathogenic nematodes in Sri Lanka Journal
Helminthology 68 277-286ARTEAGA HERNANDEZ E M amp MRAacute IumlCE K Z (1984) H
terorhabditis heliothidis a parasite of insect pests in CubFolia Parasitologia(Praha) 31 11-17
BLAXTER ML DE LE Y P GAREY J R LIU LXSCHELDEMAN P VIERSTRAETE A VANFLETERE
JR MACKEY LY DORRIS M FRISSE LM VID
JT amp THOMAS WK (1998) A molecular evolutionaframework for the phylum Nematoda Nature 392 71-75
CONSTANT P MARCHAY L F ISHER-LE SAU X MBRIAND-PANOMA S amp MAULEacuteON H (1998)Naturalocurrence of entomopathogenic nematodes( RhabditidaSteernematidae and Heterorhabditidae) in Guadeloupe islanFundamental and Applied Nematology 21 667-672
CREASE TJ (1995) Ribosomal DNA evolution at tpopulation-level mdash nucleotide variation in intergenic spaarrays of Daphnia pulex Genetics 141 1327-1337
CURRAN J amp DRIVER F (1994) Molecular taxonomy of Hterorhabditis In Burnell AM Ehlers R-U amp Masson J(Eds) Genetics of Entomopathogenic Nematode-Bacteri
Complexes Luxembourg European Commission PublicatiEUR 15681 EN pp 178-187
DIX I BURNELL A M GRIFFIN CT JOYCE SAamp DOWNES MJ (1992) The identication of biologispecies in the genus Heterorhabditis (Nematoda Heterhabditae) by cross-breeding second generation amphimicadults Parasitology104 509-518
EHLERS R-U (1996) Current and future use of nematodin biocontrolpractice and commercial aspects with regardregulatory policy issues Biocontrol Science and Technolo
6 303-317GERBI SA (1985) Evolution of ribosomal DNA In MacI
tyre RJ (Ed) Molecular evolutionary genetics New YoNY USA Plenum Press pp 419-517
GRENIER E BONIFASSI E ABAD P amp LAUMOND (1996a) Use of species specic satellite DNAs as dianostic probes in the identication of Steinernematidae a
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1011
CM Stack et al
Heterorhabditidae entomopathogenic nematodes Parasitol-
ogy 113 483-489GRENIER E LAUMOND C amp ABAD P (1996b) Molecu-
lar characterisationof two species-specic tandemly repeatedDNAs from entomopathogenic nematodes Steinernema and
Heterorhabditis(NematodaRhabditida) Molecularand Bio-
chemical Parasitology 83 47-56GRIFFIN CT JOYCE S A DIX I BURNELL AM amp
DOWNES MJ (1994)Characterisationof the entomopatho-genic nematode Heterorhabditis (Nematoda Heterorhabdi-tidae) from Ireland and Britain by molecular and cross-breeding techniques and the occurrence of the genus in theseislands Fundamental and Applied Nematology 17 245-253
GRIFFIN CT DIX I JOYCE SA BURNELL AM ampDOWNES MJ (1999a) Isolation and characterisation of
Heterorhabditis spp from Hungary Estonia and Denmark Nematology 1 321-332
GRIFFIN CT CHAERANI R FALLON D REI D AP
amp DOWNES MJ (1999b) Occurrence and distribution of the entomopathogenic nematodes Steinernema spp and Het-
erorhabditis indica in Indonesia Journal of Helminthology
74 143-150HOERAUF A NISSENPAHLE K SCHMETZ C HEN-
KLEDUHRSEN K BLAXTER ML BUTTNER DWGALLIN MY ALQAOUD KM LUCIUS R amp FLEIS-CHER B (1999) Tetracycline therapy targets intracellularbacteria in the larial nematode Litomososoides sigmodontis
and results in larial infertility Journal of Clinical Investiga-
tion 103 11-17HOMINICK WM REID AP BOHAN DA amp BRISCOE
BR (1996) Entomopathogenicnematodes biodiversity ge-ographicaldistributionand conventionon biological diversity
Biocontrol Science and Technology 6 317-332HOMINICK WM BRISCOE BR DE L PIN O FG HEN G
J HUN T DJ KOZODOY E MRAacute IumlCE K Z NGUYENKB REI D AP SPIRIDONOV S STOCK P STURHAN D WATURU C amp YOSHIDA M (1997) Biosystematics of entomopathogenic nematodes current status protocols anddenitions Journal of Helminthology 71 271-298
JOHANOWICZ D L amp HOY MA (1998) The manipula-tion of arthropodreproductionby Wolbachia endosymbiontsFlorida Entomologist 81 310-317
JOYCE S A BURNELL A M amp P OWERS TO (1994a)The characterisationof Heterorhabditisisolatesby restrictionenzyme analysis of PCR amplied fragments of mtDNA andrDNA genes Journal of Nematology 26 260-270
JOYCE SA REI D A amp CURRAN J (1994b) Applicationof the polymerase chain reaction (PCR) methods to the iden-tication of entomopathogenicnematodes In Burnell AMEhlers R-U amp Masson JP (Eds) Genetics of entomopatho-
genic nematode-bacterium complexes Luxembourg Euro-pean Commission Publication EUR 15681 EN pp 178-187
KAYA H K (1990)Soil ecology In Gaugler R amp Kaya H(Eds) Entomopathogenic nematodes in biological contr
Boca Raton FL USA CRC Press pp 93-115KAYA HK amp GAUGLER R (1993) Entomopathogenic n
matodes Annual Review of Entomology 38 181-206KIDWELL M G (1990) Evolutionaryaspects of hybrid dysg
nesis in Drosophila Canadian Journal of Zoology 68 1711726LINARES AR BOWEN T amp DOVER GA (1994) A
pects of non-random turnover involved in the concerted evlution of intergenic spacers within the ribosomal DNA
Drosophila melanogaster Journal of Molecular Evoluti
39 151-159LIU J (1994) [A new species of the genus Heterorhabd
from China (Rhabditida Heterorhabditidae)] Acta Zoota
nomica Sinica 19 268-272LIU J amp BERRY RE (1996) Heterorhabditis marela
nsp (Rhabditida Heterorhabditidae) from Oregon Journ
of Invertebrate Pathology
67 48-54MASON JM RAZAK AR amp WRIGHT DJ (1996) Trecoveryof entomopathogenicnematodes from selectedarewithin Peninsular Malaysia Journal of Helminthology 7303-307
MENTI H WRIGHT DJ amp PERRY RN (1997)Desiccatsurvival of populations of the entomopathogenic nematodSteinernema feltiaeand Heterorhabditismegidis from Greeand the UK Journal of Helminthology 71 41-46
MIDUTURI JS MOENS M HOMINICK WM BRISCO
B R amp RE ID AP (1996) Naturally occurring enmopathogenic nematodes in the province of West-FlandeBelgium Journal of Helminthology 70 319-327
NE I M amp LI W-H (1979) Mathematical model for studygenetic variation in terms of restriction endonucleases P
ceedings of the National Academy of Sciences of the Uni
States 76 5269-5273NOVAK EM DE MELLO MP GOMES HBM
GALINDO I GUEVARA P RAMIREZ JL amp DA SILV
RI A JF (1993) Repetitive sequences in the ribosomal intgenic spacer of Trypanosoma cruzi Molecular and Bioche
ical Parasitology 60 273-280PAMJAV H TRIGA D BUZAS Z VELLAI T LUCSKA
A ADAMS B REI D AP BURNELL AM GRIFF
C GLAZER I KLEIN MG amp FODOR A (1999) Nov
application of PhastSystem polyacrylamide gel electrophosis using restriction fragment length polymorphism-intertranscribed spacer patterns of individuals for molecular idetication of entomopathogenic nematodes Electrophore
201266-1273PETROV DA SCHUTZMAN J L HARTL D L amp L
ZOVSKAYA ER ( 1995) Diverse transposable elements amobilised in hybrid dysgenesis in Drosophila virilis P
ceedings of the National Academy of Sciences of the Uni
States 92 8050-8054
486 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1111
Isolates of Heterorhabditisindi
POINAR GO JR (1990) Taxonomy and biology of Stein-ernematidae and HeterorhabditidaeIn Gaugler R amp KayaHK (Eds) Entomopathogenic nematodes in biological con-
trol Boca Raton FL USA CRC Press pp 23-64POINAR GO JR (1993) Origins and phylogenetic relation-
ships of the entomophilic rhabditids Heterorhabditis and
Steinernema Fundamental and Applied Nematology16 333-338POINAR GO JR JACKSON T amp KLEIN M (1987) He-
terorhabditis megidis sp n (Heterorhabditidae Rhabditida)parasitic in the Japanese beetlePopilliajaponica(ScarabidaeColeoptera) in Ohio Proceedings of the Helminthological
Society of Washington 53 53-59POINAR GO JR KARUNAKAR GK amp DAVID H (1992)
Heterorhabditis indicus n sp (Rhabditida Nematoda) fromIndia Separation of Heterorhabditis spp by infective juve-niles Fundamental and Applied Nematology 15 467-472
REI D AP (1994) Molecular taxonomy of Steinernema InBurnell AM Ehlers R-U amp Masson JP (Eds) Genetics
of entomopathogenic nematode-bacterium complexes Lux-embourg European Commission Publication EUR 15681EN pp 178-187
SMITS P GROENEN JTM amp DERAAY G (1991) Charac-terisation of Heterorhabditis isolates using DNA restrictionlength polymorphism Revue de Neacutematologie 14 445-453
STOCK SP (1993) A new species of thegenus Heterorhabdi-
tis Poinar 1976 (Nematoda Heterorhabditidae) parasitizingGraphognathus sp larvae (Coteoptera Curculionidae) fromArgentina Research and Reviews in Parasitology 53 103-107
STOCK SP (1997) Heterorhabditis hepialus Stock Strong amp
Gardner 1996a junior synonym of H marelatus Liu amp Berry1996 (Rhabditida Rhabditidae) with a redescription of thespecies Nematologica 43 455-463
STOCK SP STRONG D amp GARDNER S (1996) Identiction of Heterorhabditis (Nematoda Heterorhabditidae)froCalifornia with a new species isolated from the larvae of tghost moth Hepialius [sic] californicus (Lepidoptera Healidae) from Bodega Bay Natural Reserve Fundamental a
Applied Nematology 19 585-592
STRONG DR KAYA HK WHIPPLE AV CHILD ALKRAIG S BONDONNE M DYE R K amp MARON J(1996) Entomopathogenic nematodes natural enemiesroot feeding caterpillarson bush lupine Oecologica 108 345
SUDHAUS W (1993) Die mittels symbiontischerBakterienetomopathogenen Nematoden Gattungen Heterorhabditis aSteinernema sind keine Schwestertaxa Verhandlungen d
Deutschen Zoologischen Gesellschaft 86 146VAHIDI H amp HONDA BM (1991)Repeats and subrepeats
the intergenic spacer of rDNA from the nematode Meloid
gyne arenaria Molecular and General Genetics 227 33
336WERREN JH (1997) Biology of Wolbachia Annual Revi
of Entomology 42 587-609WILLIAMS S M ROBBINS L G CLUSTER PD A
LARD RW amp STROBECK C (1990) Superstructureof t Drosophila ribosomal gene family Proceedings of the N
tional Academy of Sciences of the United States 87 3153160
WOUTS WM (1979) The biology and life cycle ofNew Zealand population of Heterorhabditis heliothidis (Hterorhabditidae)Nematologica 25 191-202
YOSHIDA M REID AP BRISCOE BR amp HOMINIC
WM (1998) Survey of entomopathogenic nematod(Rhabditida Steinernematidae and Heterorhabditidae) Japan Fundamental and Applied Nematology 21 185-198
Vol 2(5) 2000 4

862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 811
CM Stack et al
Fig 7 A Slot blot analysis of genomic DNA (100 ng) of the
Heterorhabditis isolates using a H indica specic satellite DNA
probe B The membrane used for Fig 7A was stripped and
reprobed using an rDNA 18S probe as a loading control A1
H indica LN2 A2 D1 A3 P2M A4 LN2B A5 LN4 A6
INA H23 B1 INA H9 B2 INA H17 B3 INA H1 B4 Ki3
B5 K4A B6 H bacteriophora HP88 C1 H zealandica NZH3
C2 H marelatus OH-10 C3 H hepialus Bodega Bay C4
Heterorhabditis lsquoIrish typersquo K122
tested When the IGS region was digested with HaeIII H indica LN2 displayed a distinctive ve fragment pro-le All of the H indica isolates with the exception of the Kenyan isolates also displayed a ve fragment pro-le Four of these fragments were conserved among theisolates but the largest fragment (of ca 740 in H indica
LN2) was polymorphicamong the H indica isolates Thisfragment which seems to be associated with the lengthpolymorphism in the H indica IGS region is the basisof the D1D1a polymorphism observed by Curran and
Driver (1994) In most species studied to date the rDNIGS contains tandem arrays of subrepeats (Gerbi 198Williams et al 1990 Vahidi amp Honda 1991 Novakal 1993 Linares et al 1994 Crease 1995) This rsults in the length of the IGS region being variable bobetween and within species The length heterogeneity
the H indica IGS PCR product did not affect the AluI rstriction prole of this fragment which suggests that Al
has a restriction site within a tandem repetitive unit in trDNA IGS of H indica In contrast HaeIII appears nto cut within this tandem repeat region with the result thone of the HaeIII restriction fragments is polymorphiclength presumably depending on the number of AluI rpeat units which it contains
The HaeIII rDNA IGS restriction prole of the Disolate is similar to that of H indica LN2 but the Hae
pattern of the Kenyan isolates is unique The large Hae
fragment of the Kenyan isolate K4A is the same size that of D1a thus the Kenyan isolates appear to represea distinct subgroup within D1a Tropical isolates froDarwin (North Australia) Puerto Rico Hawaii the VirgIslands and Egypt were found by Curran and Driv(1994) to possess the D1 prole while the D1a prowas described by these authors from isolates collectin Puerto Rico Jamaica Florida the Virgin Islands aEgypt Comparison of the D1 and D1a HaeIII and Al
rDNA IGS restriction proles with those of H indi
LN2 indicates that the isolates designated D1 and D
by Curran and Driver (1994) belong to H indicaAn important step towards achieving an effective nmatode bacterium complex for pest control is to seek nurally occurring endemic EPN isolates as such isolatare likely to possess physiological traits that are adaptto local climatic and ecological conditions The Kenyisolates described in this study clearly belong to a distinsub-group of H indica and it is probable that these islates also share a distinctive phenotype The two Kenyisolates are interfertile but only one of these isolates (Kiproduced viable progeny when crossed with the H indi
LN2 type species All of the Indonesian Heterorhabdi
isolates were indistinguishable from H indica in the dagnostic molecular tests and they were interfertile wieach other but only one of the Indonesian isolates (INH1) was interfertile with H indica LN2 and INA H1 walso interfertile with the Ki3 isolate from Kenya Theresults suggest that although H indica has a global dtribution in tropical and subtropical regions of the worgeneow within the species may be quite restricted Whave previously observedreproductiveincompatibilityb
484 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 911
Isolates of Heterorhabditisindi
tween isolates of H bacteriophora from Europe (Grifnet al 1999a) and from North America (Dix et al un-publ) We have not determined whether this reproductiveisolation is caused by genetic means or is the result of cy-toplasmic factors Cytoplasmic factors such as endosym-biont bacteria (see reviews by Werren 1997 Johanowicz
amp Hoy 1998) and chromosomal factors such as trans-posons (reviewed by Kidwell 1990 Petrov et al 1995)have been shown to be frequent causes of reproductivein-compatibility in arthropods Endosymbiont bacteria haverecently been shown to cause reproductiveincompatibilityin larial nematodes (Hoerauf et al 1999) Since the re-productive incompatibilityobserved in this study does notyield clear groups of compatible and incompatiblestrainsand in view of the consistencyof the DNA based diagnos-tic tests any assignment of biological species within the H indica group would be injudicious at present Adams
(1988) evaluates phylogeneticand biologicalspecies con-cepts in the delimitationof species in the H bacteriophora
groupThe global distribution of H indica throughout the
tropics and subtropics suggests that H indica possessesa range of phenotypic characters which give it an advan-tage in this climatic zone Such phenotypes might includeadaptationto high temperatures desiccation tolerance andgood dispersal ability It is unlikely that the widespreaddistribution of H indica in the tropics arises because thisspecies was extant before the continents began breakingup and drifting The best supported hypothesis of phylo-genetic relationships among Heterorhabditis species in aphylogeny derived from the rDNA ITS 1 DNA sequencedata puts H indica as the most ancient Heterorhabditis
lineage (Adams et al 1998) However the phylogenetictree for Heterorhabditis and Steinernema of Reid (1994)shows that the genetic distance between Heterorhabdi-
tis species is considerably less than that between Stein-
ernema species and given the small genetic distance de-tected by Reid (1994) between the most divergent of the Heterorhabditis species in his study (a D value of ca
004 as calculated by the method of Nei and Li 1979) it
seems unlikely that H indica speciated before the Juras-sicCretaceousbreak up of Pangaea(an estimated 200 mil-lion years ago)
Acknowledgement
We acknowledge the support of the European Commu-nity STD-3 Programme (TS3 CT 940273)
References
ABADON M GRENIER E LAUMOND C amp ABAD (1998) A species-specic satellite DNA from the enmopathogenic nematode Heterorhabditis indicus Geno
41 148-153
ADAMS BJ (1998) Species concepts and the evolutionparadigm in modern nematology Journal of Nematology 31-21
ADAMS BJ BURNELL AM amp POWERS TO (1998)phylogenetic analysis of the genus Heterorhabditis(NemaRhabditidae) based on internal transcribed spacer 1 DNsequence data Journal of Nematology 30 22-39
AMARASINGHE LD HOMINICK WM BRISCOE Bamp REI D AP (1994) Occurrence and distribution entomopathogenic nematodes in Sri Lanka Journal
Helminthology 68 277-286ARTEAGA HERNANDEZ E M amp MRAacute IumlCE K Z (1984) H
terorhabditis heliothidis a parasite of insect pests in CubFolia Parasitologia(Praha) 31 11-17
BLAXTER ML DE LE Y P GAREY J R LIU LXSCHELDEMAN P VIERSTRAETE A VANFLETERE
JR MACKEY LY DORRIS M FRISSE LM VID
JT amp THOMAS WK (1998) A molecular evolutionaframework for the phylum Nematoda Nature 392 71-75
CONSTANT P MARCHAY L F ISHER-LE SAU X MBRIAND-PANOMA S amp MAULEacuteON H (1998)Naturalocurrence of entomopathogenic nematodes( RhabditidaSteernematidae and Heterorhabditidae) in Guadeloupe islanFundamental and Applied Nematology 21 667-672
CREASE TJ (1995) Ribosomal DNA evolution at tpopulation-level mdash nucleotide variation in intergenic spaarrays of Daphnia pulex Genetics 141 1327-1337
CURRAN J amp DRIVER F (1994) Molecular taxonomy of Hterorhabditis In Burnell AM Ehlers R-U amp Masson J(Eds) Genetics of Entomopathogenic Nematode-Bacteri
Complexes Luxembourg European Commission PublicatiEUR 15681 EN pp 178-187
DIX I BURNELL A M GRIFFIN CT JOYCE SAamp DOWNES MJ (1992) The identication of biologispecies in the genus Heterorhabditis (Nematoda Heterhabditae) by cross-breeding second generation amphimicadults Parasitology104 509-518
EHLERS R-U (1996) Current and future use of nematodin biocontrolpractice and commercial aspects with regardregulatory policy issues Biocontrol Science and Technolo
6 303-317GERBI SA (1985) Evolution of ribosomal DNA In MacI
tyre RJ (Ed) Molecular evolutionary genetics New YoNY USA Plenum Press pp 419-517
GRENIER E BONIFASSI E ABAD P amp LAUMOND (1996a) Use of species specic satellite DNAs as dianostic probes in the identication of Steinernematidae a
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1011
CM Stack et al
Heterorhabditidae entomopathogenic nematodes Parasitol-
ogy 113 483-489GRENIER E LAUMOND C amp ABAD P (1996b) Molecu-
lar characterisationof two species-specic tandemly repeatedDNAs from entomopathogenic nematodes Steinernema and
Heterorhabditis(NematodaRhabditida) Molecularand Bio-
chemical Parasitology 83 47-56GRIFFIN CT JOYCE S A DIX I BURNELL AM amp
DOWNES MJ (1994)Characterisationof the entomopatho-genic nematode Heterorhabditis (Nematoda Heterorhabdi-tidae) from Ireland and Britain by molecular and cross-breeding techniques and the occurrence of the genus in theseislands Fundamental and Applied Nematology 17 245-253
GRIFFIN CT DIX I JOYCE SA BURNELL AM ampDOWNES MJ (1999a) Isolation and characterisation of
Heterorhabditis spp from Hungary Estonia and Denmark Nematology 1 321-332
GRIFFIN CT CHAERANI R FALLON D REI D AP
amp DOWNES MJ (1999b) Occurrence and distribution of the entomopathogenic nematodes Steinernema spp and Het-
erorhabditis indica in Indonesia Journal of Helminthology
74 143-150HOERAUF A NISSENPAHLE K SCHMETZ C HEN-
KLEDUHRSEN K BLAXTER ML BUTTNER DWGALLIN MY ALQAOUD KM LUCIUS R amp FLEIS-CHER B (1999) Tetracycline therapy targets intracellularbacteria in the larial nematode Litomososoides sigmodontis
and results in larial infertility Journal of Clinical Investiga-
tion 103 11-17HOMINICK WM REID AP BOHAN DA amp BRISCOE
BR (1996) Entomopathogenicnematodes biodiversity ge-ographicaldistributionand conventionon biological diversity
Biocontrol Science and Technology 6 317-332HOMINICK WM BRISCOE BR DE L PIN O FG HEN G
J HUN T DJ KOZODOY E MRAacute IumlCE K Z NGUYENKB REI D AP SPIRIDONOV S STOCK P STURHAN D WATURU C amp YOSHIDA M (1997) Biosystematics of entomopathogenic nematodes current status protocols anddenitions Journal of Helminthology 71 271-298
JOHANOWICZ D L amp HOY MA (1998) The manipula-tion of arthropodreproductionby Wolbachia endosymbiontsFlorida Entomologist 81 310-317
JOYCE S A BURNELL A M amp P OWERS TO (1994a)The characterisationof Heterorhabditisisolatesby restrictionenzyme analysis of PCR amplied fragments of mtDNA andrDNA genes Journal of Nematology 26 260-270
JOYCE SA REI D A amp CURRAN J (1994b) Applicationof the polymerase chain reaction (PCR) methods to the iden-tication of entomopathogenicnematodes In Burnell AMEhlers R-U amp Masson JP (Eds) Genetics of entomopatho-
genic nematode-bacterium complexes Luxembourg Euro-pean Commission Publication EUR 15681 EN pp 178-187
KAYA H K (1990)Soil ecology In Gaugler R amp Kaya H(Eds) Entomopathogenic nematodes in biological contr
Boca Raton FL USA CRC Press pp 93-115KAYA HK amp GAUGLER R (1993) Entomopathogenic n
matodes Annual Review of Entomology 38 181-206KIDWELL M G (1990) Evolutionaryaspects of hybrid dysg
nesis in Drosophila Canadian Journal of Zoology 68 1711726LINARES AR BOWEN T amp DOVER GA (1994) A
pects of non-random turnover involved in the concerted evlution of intergenic spacers within the ribosomal DNA
Drosophila melanogaster Journal of Molecular Evoluti
39 151-159LIU J (1994) [A new species of the genus Heterorhabd
from China (Rhabditida Heterorhabditidae)] Acta Zoota
nomica Sinica 19 268-272LIU J amp BERRY RE (1996) Heterorhabditis marela
nsp (Rhabditida Heterorhabditidae) from Oregon Journ
of Invertebrate Pathology
67 48-54MASON JM RAZAK AR amp WRIGHT DJ (1996) Trecoveryof entomopathogenicnematodes from selectedarewithin Peninsular Malaysia Journal of Helminthology 7303-307
MENTI H WRIGHT DJ amp PERRY RN (1997)Desiccatsurvival of populations of the entomopathogenic nematodSteinernema feltiaeand Heterorhabditismegidis from Greeand the UK Journal of Helminthology 71 41-46
MIDUTURI JS MOENS M HOMINICK WM BRISCO
B R amp RE ID AP (1996) Naturally occurring enmopathogenic nematodes in the province of West-FlandeBelgium Journal of Helminthology 70 319-327
NE I M amp LI W-H (1979) Mathematical model for studygenetic variation in terms of restriction endonucleases P
ceedings of the National Academy of Sciences of the Uni
States 76 5269-5273NOVAK EM DE MELLO MP GOMES HBM
GALINDO I GUEVARA P RAMIREZ JL amp DA SILV
RI A JF (1993) Repetitive sequences in the ribosomal intgenic spacer of Trypanosoma cruzi Molecular and Bioche
ical Parasitology 60 273-280PAMJAV H TRIGA D BUZAS Z VELLAI T LUCSKA
A ADAMS B REI D AP BURNELL AM GRIFF
C GLAZER I KLEIN MG amp FODOR A (1999) Nov
application of PhastSystem polyacrylamide gel electrophosis using restriction fragment length polymorphism-intertranscribed spacer patterns of individuals for molecular idetication of entomopathogenic nematodes Electrophore
201266-1273PETROV DA SCHUTZMAN J L HARTL D L amp L
ZOVSKAYA ER ( 1995) Diverse transposable elements amobilised in hybrid dysgenesis in Drosophila virilis P
ceedings of the National Academy of Sciences of the Uni
States 92 8050-8054
486 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1111
Isolates of Heterorhabditisindi
POINAR GO JR (1990) Taxonomy and biology of Stein-ernematidae and HeterorhabditidaeIn Gaugler R amp KayaHK (Eds) Entomopathogenic nematodes in biological con-
trol Boca Raton FL USA CRC Press pp 23-64POINAR GO JR (1993) Origins and phylogenetic relation-
ships of the entomophilic rhabditids Heterorhabditis and
Steinernema Fundamental and Applied Nematology16 333-338POINAR GO JR JACKSON T amp KLEIN M (1987) He-
terorhabditis megidis sp n (Heterorhabditidae Rhabditida)parasitic in the Japanese beetlePopilliajaponica(ScarabidaeColeoptera) in Ohio Proceedings of the Helminthological
Society of Washington 53 53-59POINAR GO JR KARUNAKAR GK amp DAVID H (1992)
Heterorhabditis indicus n sp (Rhabditida Nematoda) fromIndia Separation of Heterorhabditis spp by infective juve-niles Fundamental and Applied Nematology 15 467-472
REI D AP (1994) Molecular taxonomy of Steinernema InBurnell AM Ehlers R-U amp Masson JP (Eds) Genetics
of entomopathogenic nematode-bacterium complexes Lux-embourg European Commission Publication EUR 15681EN pp 178-187
SMITS P GROENEN JTM amp DERAAY G (1991) Charac-terisation of Heterorhabditis isolates using DNA restrictionlength polymorphism Revue de Neacutematologie 14 445-453
STOCK SP (1993) A new species of thegenus Heterorhabdi-
tis Poinar 1976 (Nematoda Heterorhabditidae) parasitizingGraphognathus sp larvae (Coteoptera Curculionidae) fromArgentina Research and Reviews in Parasitology 53 103-107
STOCK SP (1997) Heterorhabditis hepialus Stock Strong amp
Gardner 1996a junior synonym of H marelatus Liu amp Berry1996 (Rhabditida Rhabditidae) with a redescription of thespecies Nematologica 43 455-463
STOCK SP STRONG D amp GARDNER S (1996) Identiction of Heterorhabditis (Nematoda Heterorhabditidae)froCalifornia with a new species isolated from the larvae of tghost moth Hepialius [sic] californicus (Lepidoptera Healidae) from Bodega Bay Natural Reserve Fundamental a
Applied Nematology 19 585-592
STRONG DR KAYA HK WHIPPLE AV CHILD ALKRAIG S BONDONNE M DYE R K amp MARON J(1996) Entomopathogenic nematodes natural enemiesroot feeding caterpillarson bush lupine Oecologica 108 345
SUDHAUS W (1993) Die mittels symbiontischerBakterienetomopathogenen Nematoden Gattungen Heterorhabditis aSteinernema sind keine Schwestertaxa Verhandlungen d
Deutschen Zoologischen Gesellschaft 86 146VAHIDI H amp HONDA BM (1991)Repeats and subrepeats
the intergenic spacer of rDNA from the nematode Meloid
gyne arenaria Molecular and General Genetics 227 33
336WERREN JH (1997) Biology of Wolbachia Annual Revi
of Entomology 42 587-609WILLIAMS S M ROBBINS L G CLUSTER PD A
LARD RW amp STROBECK C (1990) Superstructureof t Drosophila ribosomal gene family Proceedings of the N
tional Academy of Sciences of the United States 87 3153160
WOUTS WM (1979) The biology and life cycle ofNew Zealand population of Heterorhabditis heliothidis (Hterorhabditidae)Nematologica 25 191-202
YOSHIDA M REID AP BRISCOE BR amp HOMINIC
WM (1998) Survey of entomopathogenic nematod(Rhabditida Steinernematidae and Heterorhabditidae) Japan Fundamental and Applied Nematology 21 185-198
Vol 2(5) 2000 4

862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 911
Isolates of Heterorhabditisindi
tween isolates of H bacteriophora from Europe (Grifnet al 1999a) and from North America (Dix et al un-publ) We have not determined whether this reproductiveisolation is caused by genetic means or is the result of cy-toplasmic factors Cytoplasmic factors such as endosym-biont bacteria (see reviews by Werren 1997 Johanowicz
amp Hoy 1998) and chromosomal factors such as trans-posons (reviewed by Kidwell 1990 Petrov et al 1995)have been shown to be frequent causes of reproductivein-compatibility in arthropods Endosymbiont bacteria haverecently been shown to cause reproductiveincompatibilityin larial nematodes (Hoerauf et al 1999) Since the re-productive incompatibilityobserved in this study does notyield clear groups of compatible and incompatiblestrainsand in view of the consistencyof the DNA based diagnos-tic tests any assignment of biological species within the H indica group would be injudicious at present Adams
(1988) evaluates phylogeneticand biologicalspecies con-cepts in the delimitationof species in the H bacteriophora
groupThe global distribution of H indica throughout the
tropics and subtropics suggests that H indica possessesa range of phenotypic characters which give it an advan-tage in this climatic zone Such phenotypes might includeadaptationto high temperatures desiccation tolerance andgood dispersal ability It is unlikely that the widespreaddistribution of H indica in the tropics arises because thisspecies was extant before the continents began breakingup and drifting The best supported hypothesis of phylo-genetic relationships among Heterorhabditis species in aphylogeny derived from the rDNA ITS 1 DNA sequencedata puts H indica as the most ancient Heterorhabditis
lineage (Adams et al 1998) However the phylogenetictree for Heterorhabditis and Steinernema of Reid (1994)shows that the genetic distance between Heterorhabdi-
tis species is considerably less than that between Stein-
ernema species and given the small genetic distance de-tected by Reid (1994) between the most divergent of the Heterorhabditis species in his study (a D value of ca
004 as calculated by the method of Nei and Li 1979) it
seems unlikely that H indica speciated before the Juras-sicCretaceousbreak up of Pangaea(an estimated 200 mil-lion years ago)
Acknowledgement
We acknowledge the support of the European Commu-nity STD-3 Programme (TS3 CT 940273)
References
ABADON M GRENIER E LAUMOND C amp ABAD (1998) A species-specic satellite DNA from the enmopathogenic nematode Heterorhabditis indicus Geno
41 148-153
ADAMS BJ (1998) Species concepts and the evolutionparadigm in modern nematology Journal of Nematology 31-21
ADAMS BJ BURNELL AM amp POWERS TO (1998)phylogenetic analysis of the genus Heterorhabditis(NemaRhabditidae) based on internal transcribed spacer 1 DNsequence data Journal of Nematology 30 22-39
AMARASINGHE LD HOMINICK WM BRISCOE Bamp REI D AP (1994) Occurrence and distribution entomopathogenic nematodes in Sri Lanka Journal
Helminthology 68 277-286ARTEAGA HERNANDEZ E M amp MRAacute IumlCE K Z (1984) H
terorhabditis heliothidis a parasite of insect pests in CubFolia Parasitologia(Praha) 31 11-17
BLAXTER ML DE LE Y P GAREY J R LIU LXSCHELDEMAN P VIERSTRAETE A VANFLETERE
JR MACKEY LY DORRIS M FRISSE LM VID
JT amp THOMAS WK (1998) A molecular evolutionaframework for the phylum Nematoda Nature 392 71-75
CONSTANT P MARCHAY L F ISHER-LE SAU X MBRIAND-PANOMA S amp MAULEacuteON H (1998)Naturalocurrence of entomopathogenic nematodes( RhabditidaSteernematidae and Heterorhabditidae) in Guadeloupe islanFundamental and Applied Nematology 21 667-672
CREASE TJ (1995) Ribosomal DNA evolution at tpopulation-level mdash nucleotide variation in intergenic spaarrays of Daphnia pulex Genetics 141 1327-1337
CURRAN J amp DRIVER F (1994) Molecular taxonomy of Hterorhabditis In Burnell AM Ehlers R-U amp Masson J(Eds) Genetics of Entomopathogenic Nematode-Bacteri
Complexes Luxembourg European Commission PublicatiEUR 15681 EN pp 178-187
DIX I BURNELL A M GRIFFIN CT JOYCE SAamp DOWNES MJ (1992) The identication of biologispecies in the genus Heterorhabditis (Nematoda Heterhabditae) by cross-breeding second generation amphimicadults Parasitology104 509-518
EHLERS R-U (1996) Current and future use of nematodin biocontrolpractice and commercial aspects with regardregulatory policy issues Biocontrol Science and Technolo
6 303-317GERBI SA (1985) Evolution of ribosomal DNA In MacI
tyre RJ (Ed) Molecular evolutionary genetics New YoNY USA Plenum Press pp 419-517
GRENIER E BONIFASSI E ABAD P amp LAUMOND (1996a) Use of species specic satellite DNAs as dianostic probes in the identication of Steinernematidae a
Vol 2(5) 2000 4
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1011
CM Stack et al
Heterorhabditidae entomopathogenic nematodes Parasitol-
ogy 113 483-489GRENIER E LAUMOND C amp ABAD P (1996b) Molecu-
lar characterisationof two species-specic tandemly repeatedDNAs from entomopathogenic nematodes Steinernema and
Heterorhabditis(NematodaRhabditida) Molecularand Bio-
chemical Parasitology 83 47-56GRIFFIN CT JOYCE S A DIX I BURNELL AM amp
DOWNES MJ (1994)Characterisationof the entomopatho-genic nematode Heterorhabditis (Nematoda Heterorhabdi-tidae) from Ireland and Britain by molecular and cross-breeding techniques and the occurrence of the genus in theseislands Fundamental and Applied Nematology 17 245-253
GRIFFIN CT DIX I JOYCE SA BURNELL AM ampDOWNES MJ (1999a) Isolation and characterisation of
Heterorhabditis spp from Hungary Estonia and Denmark Nematology 1 321-332
GRIFFIN CT CHAERANI R FALLON D REI D AP
amp DOWNES MJ (1999b) Occurrence and distribution of the entomopathogenic nematodes Steinernema spp and Het-
erorhabditis indica in Indonesia Journal of Helminthology
74 143-150HOERAUF A NISSENPAHLE K SCHMETZ C HEN-
KLEDUHRSEN K BLAXTER ML BUTTNER DWGALLIN MY ALQAOUD KM LUCIUS R amp FLEIS-CHER B (1999) Tetracycline therapy targets intracellularbacteria in the larial nematode Litomososoides sigmodontis
and results in larial infertility Journal of Clinical Investiga-
tion 103 11-17HOMINICK WM REID AP BOHAN DA amp BRISCOE
BR (1996) Entomopathogenicnematodes biodiversity ge-ographicaldistributionand conventionon biological diversity
Biocontrol Science and Technology 6 317-332HOMINICK WM BRISCOE BR DE L PIN O FG HEN G
J HUN T DJ KOZODOY E MRAacute IumlCE K Z NGUYENKB REI D AP SPIRIDONOV S STOCK P STURHAN D WATURU C amp YOSHIDA M (1997) Biosystematics of entomopathogenic nematodes current status protocols anddenitions Journal of Helminthology 71 271-298
JOHANOWICZ D L amp HOY MA (1998) The manipula-tion of arthropodreproductionby Wolbachia endosymbiontsFlorida Entomologist 81 310-317
JOYCE S A BURNELL A M amp P OWERS TO (1994a)The characterisationof Heterorhabditisisolatesby restrictionenzyme analysis of PCR amplied fragments of mtDNA andrDNA genes Journal of Nematology 26 260-270
JOYCE SA REI D A amp CURRAN J (1994b) Applicationof the polymerase chain reaction (PCR) methods to the iden-tication of entomopathogenicnematodes In Burnell AMEhlers R-U amp Masson JP (Eds) Genetics of entomopatho-
genic nematode-bacterium complexes Luxembourg Euro-pean Commission Publication EUR 15681 EN pp 178-187
KAYA H K (1990)Soil ecology In Gaugler R amp Kaya H(Eds) Entomopathogenic nematodes in biological contr
Boca Raton FL USA CRC Press pp 93-115KAYA HK amp GAUGLER R (1993) Entomopathogenic n
matodes Annual Review of Entomology 38 181-206KIDWELL M G (1990) Evolutionaryaspects of hybrid dysg
nesis in Drosophila Canadian Journal of Zoology 68 1711726LINARES AR BOWEN T amp DOVER GA (1994) A
pects of non-random turnover involved in the concerted evlution of intergenic spacers within the ribosomal DNA
Drosophila melanogaster Journal of Molecular Evoluti
39 151-159LIU J (1994) [A new species of the genus Heterorhabd
from China (Rhabditida Heterorhabditidae)] Acta Zoota
nomica Sinica 19 268-272LIU J amp BERRY RE (1996) Heterorhabditis marela
nsp (Rhabditida Heterorhabditidae) from Oregon Journ
of Invertebrate Pathology
67 48-54MASON JM RAZAK AR amp WRIGHT DJ (1996) Trecoveryof entomopathogenicnematodes from selectedarewithin Peninsular Malaysia Journal of Helminthology 7303-307
MENTI H WRIGHT DJ amp PERRY RN (1997)Desiccatsurvival of populations of the entomopathogenic nematodSteinernema feltiaeand Heterorhabditismegidis from Greeand the UK Journal of Helminthology 71 41-46
MIDUTURI JS MOENS M HOMINICK WM BRISCO
B R amp RE ID AP (1996) Naturally occurring enmopathogenic nematodes in the province of West-FlandeBelgium Journal of Helminthology 70 319-327
NE I M amp LI W-H (1979) Mathematical model for studygenetic variation in terms of restriction endonucleases P
ceedings of the National Academy of Sciences of the Uni
States 76 5269-5273NOVAK EM DE MELLO MP GOMES HBM
GALINDO I GUEVARA P RAMIREZ JL amp DA SILV
RI A JF (1993) Repetitive sequences in the ribosomal intgenic spacer of Trypanosoma cruzi Molecular and Bioche
ical Parasitology 60 273-280PAMJAV H TRIGA D BUZAS Z VELLAI T LUCSKA
A ADAMS B REI D AP BURNELL AM GRIFF
C GLAZER I KLEIN MG amp FODOR A (1999) Nov
application of PhastSystem polyacrylamide gel electrophosis using restriction fragment length polymorphism-intertranscribed spacer patterns of individuals for molecular idetication of entomopathogenic nematodes Electrophore
201266-1273PETROV DA SCHUTZMAN J L HARTL D L amp L
ZOVSKAYA ER ( 1995) Diverse transposable elements amobilised in hybrid dysgenesis in Drosophila virilis P
ceedings of the National Academy of Sciences of the Uni
States 92 8050-8054
486 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1111
Isolates of Heterorhabditisindi
POINAR GO JR (1990) Taxonomy and biology of Stein-ernematidae and HeterorhabditidaeIn Gaugler R amp KayaHK (Eds) Entomopathogenic nematodes in biological con-
trol Boca Raton FL USA CRC Press pp 23-64POINAR GO JR (1993) Origins and phylogenetic relation-
ships of the entomophilic rhabditids Heterorhabditis and
Steinernema Fundamental and Applied Nematology16 333-338POINAR GO JR JACKSON T amp KLEIN M (1987) He-
terorhabditis megidis sp n (Heterorhabditidae Rhabditida)parasitic in the Japanese beetlePopilliajaponica(ScarabidaeColeoptera) in Ohio Proceedings of the Helminthological
Society of Washington 53 53-59POINAR GO JR KARUNAKAR GK amp DAVID H (1992)
Heterorhabditis indicus n sp (Rhabditida Nematoda) fromIndia Separation of Heterorhabditis spp by infective juve-niles Fundamental and Applied Nematology 15 467-472
REI D AP (1994) Molecular taxonomy of Steinernema InBurnell AM Ehlers R-U amp Masson JP (Eds) Genetics
of entomopathogenic nematode-bacterium complexes Lux-embourg European Commission Publication EUR 15681EN pp 178-187
SMITS P GROENEN JTM amp DERAAY G (1991) Charac-terisation of Heterorhabditis isolates using DNA restrictionlength polymorphism Revue de Neacutematologie 14 445-453
STOCK SP (1993) A new species of thegenus Heterorhabdi-
tis Poinar 1976 (Nematoda Heterorhabditidae) parasitizingGraphognathus sp larvae (Coteoptera Curculionidae) fromArgentina Research and Reviews in Parasitology 53 103-107
STOCK SP (1997) Heterorhabditis hepialus Stock Strong amp
Gardner 1996a junior synonym of H marelatus Liu amp Berry1996 (Rhabditida Rhabditidae) with a redescription of thespecies Nematologica 43 455-463
STOCK SP STRONG D amp GARDNER S (1996) Identiction of Heterorhabditis (Nematoda Heterorhabditidae)froCalifornia with a new species isolated from the larvae of tghost moth Hepialius [sic] californicus (Lepidoptera Healidae) from Bodega Bay Natural Reserve Fundamental a
Applied Nematology 19 585-592
STRONG DR KAYA HK WHIPPLE AV CHILD ALKRAIG S BONDONNE M DYE R K amp MARON J(1996) Entomopathogenic nematodes natural enemiesroot feeding caterpillarson bush lupine Oecologica 108 345
SUDHAUS W (1993) Die mittels symbiontischerBakterienetomopathogenen Nematoden Gattungen Heterorhabditis aSteinernema sind keine Schwestertaxa Verhandlungen d
Deutschen Zoologischen Gesellschaft 86 146VAHIDI H amp HONDA BM (1991)Repeats and subrepeats
the intergenic spacer of rDNA from the nematode Meloid
gyne arenaria Molecular and General Genetics 227 33
336WERREN JH (1997) Biology of Wolbachia Annual Revi
of Entomology 42 587-609WILLIAMS S M ROBBINS L G CLUSTER PD A
LARD RW amp STROBECK C (1990) Superstructureof t Drosophila ribosomal gene family Proceedings of the N
tional Academy of Sciences of the United States 87 3153160
WOUTS WM (1979) The biology and life cycle ofNew Zealand population of Heterorhabditis heliothidis (Hterorhabditidae)Nematologica 25 191-202
YOSHIDA M REID AP BRISCOE BR amp HOMINIC
WM (1998) Survey of entomopathogenic nematod(Rhabditida Steinernematidae and Heterorhabditidae) Japan Fundamental and Applied Nematology 21 185-198
Vol 2(5) 2000 4

862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1011
CM Stack et al
Heterorhabditidae entomopathogenic nematodes Parasitol-
ogy 113 483-489GRENIER E LAUMOND C amp ABAD P (1996b) Molecu-
lar characterisationof two species-specic tandemly repeatedDNAs from entomopathogenic nematodes Steinernema and
Heterorhabditis(NematodaRhabditida) Molecularand Bio-
chemical Parasitology 83 47-56GRIFFIN CT JOYCE S A DIX I BURNELL AM amp
DOWNES MJ (1994)Characterisationof the entomopatho-genic nematode Heterorhabditis (Nematoda Heterorhabdi-tidae) from Ireland and Britain by molecular and cross-breeding techniques and the occurrence of the genus in theseislands Fundamental and Applied Nematology 17 245-253
GRIFFIN CT DIX I JOYCE SA BURNELL AM ampDOWNES MJ (1999a) Isolation and characterisation of
Heterorhabditis spp from Hungary Estonia and Denmark Nematology 1 321-332
GRIFFIN CT CHAERANI R FALLON D REI D AP
amp DOWNES MJ (1999b) Occurrence and distribution of the entomopathogenic nematodes Steinernema spp and Het-
erorhabditis indica in Indonesia Journal of Helminthology
74 143-150HOERAUF A NISSENPAHLE K SCHMETZ C HEN-
KLEDUHRSEN K BLAXTER ML BUTTNER DWGALLIN MY ALQAOUD KM LUCIUS R amp FLEIS-CHER B (1999) Tetracycline therapy targets intracellularbacteria in the larial nematode Litomososoides sigmodontis
and results in larial infertility Journal of Clinical Investiga-
tion 103 11-17HOMINICK WM REID AP BOHAN DA amp BRISCOE
BR (1996) Entomopathogenicnematodes biodiversity ge-ographicaldistributionand conventionon biological diversity
Biocontrol Science and Technology 6 317-332HOMINICK WM BRISCOE BR DE L PIN O FG HEN G
J HUN T DJ KOZODOY E MRAacute IumlCE K Z NGUYENKB REI D AP SPIRIDONOV S STOCK P STURHAN D WATURU C amp YOSHIDA M (1997) Biosystematics of entomopathogenic nematodes current status protocols anddenitions Journal of Helminthology 71 271-298
JOHANOWICZ D L amp HOY MA (1998) The manipula-tion of arthropodreproductionby Wolbachia endosymbiontsFlorida Entomologist 81 310-317
JOYCE S A BURNELL A M amp P OWERS TO (1994a)The characterisationof Heterorhabditisisolatesby restrictionenzyme analysis of PCR amplied fragments of mtDNA andrDNA genes Journal of Nematology 26 260-270
JOYCE SA REI D A amp CURRAN J (1994b) Applicationof the polymerase chain reaction (PCR) methods to the iden-tication of entomopathogenicnematodes In Burnell AMEhlers R-U amp Masson JP (Eds) Genetics of entomopatho-
genic nematode-bacterium complexes Luxembourg Euro-pean Commission Publication EUR 15681 EN pp 178-187
KAYA H K (1990)Soil ecology In Gaugler R amp Kaya H(Eds) Entomopathogenic nematodes in biological contr
Boca Raton FL USA CRC Press pp 93-115KAYA HK amp GAUGLER R (1993) Entomopathogenic n
matodes Annual Review of Entomology 38 181-206KIDWELL M G (1990) Evolutionaryaspects of hybrid dysg
nesis in Drosophila Canadian Journal of Zoology 68 1711726LINARES AR BOWEN T amp DOVER GA (1994) A
pects of non-random turnover involved in the concerted evlution of intergenic spacers within the ribosomal DNA
Drosophila melanogaster Journal of Molecular Evoluti
39 151-159LIU J (1994) [A new species of the genus Heterorhabd
from China (Rhabditida Heterorhabditidae)] Acta Zoota
nomica Sinica 19 268-272LIU J amp BERRY RE (1996) Heterorhabditis marela
nsp (Rhabditida Heterorhabditidae) from Oregon Journ
of Invertebrate Pathology
67 48-54MASON JM RAZAK AR amp WRIGHT DJ (1996) Trecoveryof entomopathogenicnematodes from selectedarewithin Peninsular Malaysia Journal of Helminthology 7303-307
MENTI H WRIGHT DJ amp PERRY RN (1997)Desiccatsurvival of populations of the entomopathogenic nematodSteinernema feltiaeand Heterorhabditismegidis from Greeand the UK Journal of Helminthology 71 41-46
MIDUTURI JS MOENS M HOMINICK WM BRISCO
B R amp RE ID AP (1996) Naturally occurring enmopathogenic nematodes in the province of West-FlandeBelgium Journal of Helminthology 70 319-327
NE I M amp LI W-H (1979) Mathematical model for studygenetic variation in terms of restriction endonucleases P
ceedings of the National Academy of Sciences of the Uni
States 76 5269-5273NOVAK EM DE MELLO MP GOMES HBM
GALINDO I GUEVARA P RAMIREZ JL amp DA SILV
RI A JF (1993) Repetitive sequences in the ribosomal intgenic spacer of Trypanosoma cruzi Molecular and Bioche
ical Parasitology 60 273-280PAMJAV H TRIGA D BUZAS Z VELLAI T LUCSKA
A ADAMS B REI D AP BURNELL AM GRIFF
C GLAZER I KLEIN MG amp FODOR A (1999) Nov
application of PhastSystem polyacrylamide gel electrophosis using restriction fragment length polymorphism-intertranscribed spacer patterns of individuals for molecular idetication of entomopathogenic nematodes Electrophore
201266-1273PETROV DA SCHUTZMAN J L HARTL D L amp L
ZOVSKAYA ER ( 1995) Diverse transposable elements amobilised in hybrid dysgenesis in Drosophila virilis P
ceedings of the National Academy of Sciences of the Uni
States 92 8050-8054
486 Nematolo
862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1111
Isolates of Heterorhabditisindi
POINAR GO JR (1990) Taxonomy and biology of Stein-ernematidae and HeterorhabditidaeIn Gaugler R amp KayaHK (Eds) Entomopathogenic nematodes in biological con-
trol Boca Raton FL USA CRC Press pp 23-64POINAR GO JR (1993) Origins and phylogenetic relation-
ships of the entomophilic rhabditids Heterorhabditis and
Steinernema Fundamental and Applied Nematology16 333-338POINAR GO JR JACKSON T amp KLEIN M (1987) He-
terorhabditis megidis sp n (Heterorhabditidae Rhabditida)parasitic in the Japanese beetlePopilliajaponica(ScarabidaeColeoptera) in Ohio Proceedings of the Helminthological
Society of Washington 53 53-59POINAR GO JR KARUNAKAR GK amp DAVID H (1992)
Heterorhabditis indicus n sp (Rhabditida Nematoda) fromIndia Separation of Heterorhabditis spp by infective juve-niles Fundamental and Applied Nematology 15 467-472
REI D AP (1994) Molecular taxonomy of Steinernema InBurnell AM Ehlers R-U amp Masson JP (Eds) Genetics
of entomopathogenic nematode-bacterium complexes Lux-embourg European Commission Publication EUR 15681EN pp 178-187
SMITS P GROENEN JTM amp DERAAY G (1991) Charac-terisation of Heterorhabditis isolates using DNA restrictionlength polymorphism Revue de Neacutematologie 14 445-453
STOCK SP (1993) A new species of thegenus Heterorhabdi-
tis Poinar 1976 (Nematoda Heterorhabditidae) parasitizingGraphognathus sp larvae (Coteoptera Curculionidae) fromArgentina Research and Reviews in Parasitology 53 103-107
STOCK SP (1997) Heterorhabditis hepialus Stock Strong amp
Gardner 1996a junior synonym of H marelatus Liu amp Berry1996 (Rhabditida Rhabditidae) with a redescription of thespecies Nematologica 43 455-463
STOCK SP STRONG D amp GARDNER S (1996) Identiction of Heterorhabditis (Nematoda Heterorhabditidae)froCalifornia with a new species isolated from the larvae of tghost moth Hepialius [sic] californicus (Lepidoptera Healidae) from Bodega Bay Natural Reserve Fundamental a
Applied Nematology 19 585-592
STRONG DR KAYA HK WHIPPLE AV CHILD ALKRAIG S BONDONNE M DYE R K amp MARON J(1996) Entomopathogenic nematodes natural enemiesroot feeding caterpillarson bush lupine Oecologica 108 345
SUDHAUS W (1993) Die mittels symbiontischerBakterienetomopathogenen Nematoden Gattungen Heterorhabditis aSteinernema sind keine Schwestertaxa Verhandlungen d
Deutschen Zoologischen Gesellschaft 86 146VAHIDI H amp HONDA BM (1991)Repeats and subrepeats
the intergenic spacer of rDNA from the nematode Meloid
gyne arenaria Molecular and General Genetics 227 33
336WERREN JH (1997) Biology of Wolbachia Annual Revi
of Entomology 42 587-609WILLIAMS S M ROBBINS L G CLUSTER PD A
LARD RW amp STROBECK C (1990) Superstructureof t Drosophila ribosomal gene family Proceedings of the N
tional Academy of Sciences of the United States 87 3153160
WOUTS WM (1979) The biology and life cycle ofNew Zealand population of Heterorhabditis heliothidis (Hterorhabditidae)Nematologica 25 191-202
YOSHIDA M REID AP BRISCOE BR amp HOMINIC
WM (1998) Survey of entomopathogenic nematod(Rhabditida Steinernematidae and Heterorhabditidae) Japan Fundamental and Applied Nematology 21 185-198
Vol 2(5) 2000 4

862019 Its Entomopatogenico
httpslidepdfcomreaderfullits-entomopatogenico 1111
Isolates of Heterorhabditisindi
POINAR GO JR (1990) Taxonomy and biology of Stein-ernematidae and HeterorhabditidaeIn Gaugler R amp KayaHK (Eds) Entomopathogenic nematodes in biological con-
trol Boca Raton FL USA CRC Press pp 23-64POINAR GO JR (1993) Origins and phylogenetic relation-
ships of the entomophilic rhabditids Heterorhabditis and
Steinernema Fundamental and Applied Nematology16 333-338POINAR GO JR JACKSON T amp KLEIN M (1987) He-
terorhabditis megidis sp n (Heterorhabditidae Rhabditida)parasitic in the Japanese beetlePopilliajaponica(ScarabidaeColeoptera) in Ohio Proceedings of the Helminthological
Society of Washington 53 53-59POINAR GO JR KARUNAKAR GK amp DAVID H (1992)
Heterorhabditis indicus n sp (Rhabditida Nematoda) fromIndia Separation of Heterorhabditis spp by infective juve-niles Fundamental and Applied Nematology 15 467-472
REI D AP (1994) Molecular taxonomy of Steinernema InBurnell AM Ehlers R-U amp Masson JP (Eds) Genetics
of entomopathogenic nematode-bacterium complexes Lux-embourg European Commission Publication EUR 15681EN pp 178-187
SMITS P GROENEN JTM amp DERAAY G (1991) Charac-terisation of Heterorhabditis isolates using DNA restrictionlength polymorphism Revue de Neacutematologie 14 445-453
STOCK SP (1993) A new species of thegenus Heterorhabdi-
tis Poinar 1976 (Nematoda Heterorhabditidae) parasitizingGraphognathus sp larvae (Coteoptera Curculionidae) fromArgentina Research and Reviews in Parasitology 53 103-107
STOCK SP (1997) Heterorhabditis hepialus Stock Strong amp
Gardner 1996a junior synonym of H marelatus Liu amp Berry1996 (Rhabditida Rhabditidae) with a redescription of thespecies Nematologica 43 455-463
STOCK SP STRONG D amp GARDNER S (1996) Identiction of Heterorhabditis (Nematoda Heterorhabditidae)froCalifornia with a new species isolated from the larvae of tghost moth Hepialius [sic] californicus (Lepidoptera Healidae) from Bodega Bay Natural Reserve Fundamental a
Applied Nematology 19 585-592
STRONG DR KAYA HK WHIPPLE AV CHILD ALKRAIG S BONDONNE M DYE R K amp MARON J(1996) Entomopathogenic nematodes natural enemiesroot feeding caterpillarson bush lupine Oecologica 108 345
SUDHAUS W (1993) Die mittels symbiontischerBakterienetomopathogenen Nematoden Gattungen Heterorhabditis aSteinernema sind keine Schwestertaxa Verhandlungen d
Deutschen Zoologischen Gesellschaft 86 146VAHIDI H amp HONDA BM (1991)Repeats and subrepeats
the intergenic spacer of rDNA from the nematode Meloid
gyne arenaria Molecular and General Genetics 227 33
336WERREN JH (1997) Biology of Wolbachia Annual Revi
of Entomology 42 587-609WILLIAMS S M ROBBINS L G CLUSTER PD A
LARD RW amp STROBECK C (1990) Superstructureof t Drosophila ribosomal gene family Proceedings of the N
tional Academy of Sciences of the United States 87 3153160
WOUTS WM (1979) The biology and life cycle ofNew Zealand population of Heterorhabditis heliothidis (Hterorhabditidae)Nematologica 25 191-202
YOSHIDA M REID AP BRISCOE BR amp HOMINIC
WM (1998) Survey of entomopathogenic nematod(Rhabditida Steinernematidae and Heterorhabditidae) Japan Fundamental and Applied Nematology 21 185-198
Vol 2(5) 2000 4


















