
Isolation Intact and Functional Mesophyll Sheath ... · percentage intactness ofchloroplasts...
7
Plant Physiol. (1979) 63, 821-827 0032-0889/79/63/0821/07/$00.50/0 Isolation of Intact and Functional Chloroplasts from Mesophyll and Bundle Sheath Protoplasts of the C4 Plant Paniclum miliaceum 1 Received for publication October 12, 1978 and in revised form January 3, 1979 GERALD E. EDWARDS Department of Horticulture, University of Wisconsin, Madison, Wisconsin 53706 Ross MCC. LILLEY Department of Biology, The University of Wollongong, P.O. Box 1144, Wollongong, N.S. W 2500 Australia STEWART CRAIG AND MARSHALL D. HATCH Division of Plant Industry, CSIRO, P.O. Box 1600, Canberra City, A. C. T 2601 Australia ABSTRACT A procedure is described for isolating and purifying mesophyll proto- plasts and bundle sheath protoplasts of the C4 plant Panicum milaceum. Following enzymic digestion of leaf tissue, nesophyll protoplasts and bundle sheath protoplasts are released and purified by density centrifuga- ti The lower density of mesophyll protoplasts aflowed rapid separation of the two protoplast types. Evidence for separation of mesophyll proto- plasts and bundle sheath protoplasts (up to 95% purity) is provided from light microscopy (based on size difference in both chloroplasts and proto- plasts), levels of marker enzymes in the preparations (i.e. pyruvate, Pi dikinase and phosphoenolpyruvate carboxylase for mesophyll and ribuose- 1,5-bisphosphate carboxylase for bundle sheath), and differences in sub- strate-dependent 02 evolution by chloroplasts isolated from protoplasts. Chloroplasts were isolated from protoplasts by several passages of the protoplasts through a 20-micrometer nylon mesh. Mesophyll chloroplasts were judged approximately 90 to 95% intact and bundle sheath chloroplasts 80 to 90% intact based on retention of chloroplast marker enzymes and the ferricyanide test for intactness. It was necessary to include 10 millimolar MgC2 in media for osmotically shocking the chloroplasts in order to obtain maximum and linear rates of ferricyanide-dependent 02 evolution. Chloroplasts isolated from mesophyll protoplast preparations had low rates of Light-dependent 02 evolution in the presence of 10 millimolar NaHCO3 (0.13 micromoles per milligram chlorophyll per minute) in com- parison to bundle sheath chloroplasts (1 to 2.5 micromoles per milligram chlorophyll per minute). The mesophyll chloroplasts catalyze high rates of 3-phosphoglycerate-dependent 02 evolution (2 to 4 micromoles per mill gram chlorophyll per minute). Orthophosphate but not phosphoenolpyru- vate inhibited the 3-phosphoglycerate-dependent 02 evolution by the mes- ophyll chloroplasts. Rates of 02 evolution were much higher with mesophyll than with bundle sheath chloroplasts in the presence of pyruvate plus oxaloacetate. The results are discussed in relation to the proposed function of these chloroplasts during C4 photosynthesis. Previous studies have established that there is differential com- partmentation of photosynthetic functions between mesophyll and 'Research was performed at Division of Plant Industry, CSIRO, Can- berra City, Australia in part while G. E. Edwards was on study leave under a Guggenheim Fellowship. Partial support was provided by Na- tional Science Foundation Grant PCM 77-09384 to G. E. E. bundle sheath cells of C4 plants (see 12 for review). While C4 photosynthesis provides a mechanism for high rates of carbon assimilation, rapid inter- and intracellular fluxes of certain metab- olites are required. Comparative studies on the regulation of photosynthetic metabolism and the mechanism of metabolite transport of mesophyll and bundle sheath chloroplasts are de- pendent on obtaining the respective organelles in a pure, intact, and functional state. As effective means of isolating intact mesophyll chloroplasts of both C3 and C4 plants is by gentle lysis of MP2 (4). Previously, enzymic digestion of leaf tissue of C4 plants resulted in release of MP and strands of bundle sheath cells attached to vascular strands. The bundle sheath cell wall was much more resistant to enzymic digestion due either to its greater thickness or differing composi- tion. Compared with bundle sheath strands, BSP would be a more convenient source of intact organelles and plasma membrane. In addition, contamination by membranes, enzymes, and organelles derived from vascular tissue would be eliminated. In the present study, procedures for isolating mesophyll and bundle sheath protoplasts of Panicum miliaceum were developed and the quality of chloroplasts isolated from protoplasts was evaluated. MATERIALS AND METHODS Plant Material and Protoplast Isolation. P. miliaceum, variety Sirocket-5, was grown in soil in a greenhouse within a day-night temperature range of 30 to 20 C. For isolation of MP and BSP, leaf tissue was taken 15 to 25 days after planting (leaves approxi- mately 70-130 mm in length). In older tissue the bundle sheath strands became resistant to enzymic digestion which resulted in isolation of MP and bundle sheath strands. Eight to 10 leaves at a time were sampled, stacked, and cut by hand into small segments about 0.7 mm in width. Approximately 8 g of leaf tissue were prepared and subjected to enzymic digestion as previously described (5, 18). The enzyme medium contained 0.5 M sorbitol, 1 mm CaCl2, 0.1% BSA, 2% cellulase (Onozuka 3S), and 0.3% pectinase (Macerozyme R-10) (pH 5.0). The tissue was incubated in a crystallizing dish in a 30 C water bath under low light (500 ft-c or about 15 nE cm-2 s-1 between 400 to 700 nm from a Phillips 250-w lamp) for 4.5 h (similar yields are obtained 2Abbreviations: MP: mesophyll protoplasts; BSP: Bundle sheath pro- toplasts; PGA: 3-phosphoglycerate; PEP: phosphoenolpyruvate; RuBP: ribulose 1,5-bisphosphate. 821 www.plantphysiol.org on January 26, 2020 - Published by Downloaded from Copyright © 1979 American Society of Plant Biologists. All rights reserved.
Transcript of Isolation Intact and Functional Mesophyll Sheath ... · percentage intactness ofchloroplasts...

Plant Physiol. (1979) 63, 821-8270032-0889/79/63/0821/07/$00.50/0
Isolation of Intact and Functional Chloroplasts from Mesophylland Bundle Sheath Protoplasts of the C4 Plant Paniclummiliaceum 1
Received for publication October 12, 1978 and in revised form January 3, 1979
GERALD E. EDWARDSDepartment of Horticulture, University of Wisconsin, Madison, Wisconsin 53706
Ross MCC. LILLEYDepartment of Biology, The University of Wollongong, P.O. Box 1144, Wollongong, N.S.W 2500 Australia
STEWART CRAIG AND MARSHALL D. HATCHDivision of Plant Industry, CSIRO, P.O. Box 1600, Canberra City, A. C. T 2601 Australia
ABSTRACT
A procedure is described for isolating and purifying mesophyll proto-plasts and bundle sheath protoplasts of the C4 plant Panicum milaceum.Following enzymic digestion of leaf tissue, nesophyll protoplasts andbundle sheath protoplasts are released and purified by density centrifuga-ti The lower density of mesophyll protoplasts aflowed rapid separationof the two protoplast types. Evidence for separation of mesophyll proto-plasts and bundle sheath protoplasts (up to 95% purity) is provided fromlight microscopy (based on size difference in both chloroplasts and proto-plasts), levels of marker enzymes in the preparations (i.e. pyruvate, Pidikinase and phosphoenolpyruvate carboxylase for mesophyll and ribuose-1,5-bisphosphate carboxylase for bundle sheath), and differences in sub-strate-dependent 02 evolution by chloroplasts isolated from protoplasts.
Chloroplasts were isolated from protoplasts by several passages of theprotoplasts through a 20-micrometer nylon mesh. Mesophyll chloroplastswere judged approximately 90 to 95% intact and bundle sheath chloroplasts80 to 90% intact based on retention of chloroplast marker enzymes and theferricyanide test for intactness. It was necessary to include 10 millimolarMgC2 in media for osmotically shocking the chloroplasts in order to obtainmaximum and linear rates of ferricyanide-dependent 02 evolution.
Chloroplasts isolated from mesophyll protoplast preparations had lowrates of Light-dependent 02 evolution in the presence of 10 millimolarNaHCO3 (0.13 micromoles per milligram chlorophyll per minute) in com-parison to bundle sheath chloroplasts (1 to 2.5 micromoles per milligramchlorophyll per minute). The mesophyll chloroplasts catalyze high rates of3-phosphoglycerate-dependent 02 evolution (2 to 4 micromoles per millgram chlorophyll per minute). Orthophosphate but not phosphoenolpyru-vate inhibited the 3-phosphoglycerate-dependent 02 evolution by the mes-ophyll chloroplasts. Rates of02 evolution were much higher with mesophyllthan with bundle sheath chloroplasts in the presence of pyruvate plusoxaloacetate. The results are discussed in relation to the proposed functionof these chloroplasts during C4 photosynthesis.
Previous studies have established that there is differential com-partmentation ofphotosynthetic functions between mesophyll and
'Research was performed at Division of Plant Industry, CSIRO, Can-berra City, Australia in part while G. E. Edwards was on study leaveunder a Guggenheim Fellowship. Partial support was provided by Na-tional Science Foundation Grant PCM 77-09384 to G. E. E.
bundle sheath cells of C4 plants (see 12 for review). While C4photosynthesis provides a mechanism for high rates of carbonassimilation, rapid inter- and intracellular fluxes of certain metab-olites are required. Comparative studies on the regulation ofphotosynthetic metabolism and the mechanism of metabolitetransport of mesophyll and bundle sheath chloroplasts are de-pendent on obtaining the respective organelles in a pure, intact,and functional state.As effective means of isolating intact mesophyll chloroplasts of
both C3 and C4 plants is by gentle lysis of MP2 (4). Previously,enzymic digestion of leaf tissue of C4 plants resulted in release ofMP and strands ofbundle sheath cells attached to vascular strands.The bundle sheath cell wall was much more resistant to enzymicdigestion due either to its greater thickness or differing composi-tion. Compared with bundle sheath strands, BSP would be a moreconvenient source of intact organelles and plasma membrane. Inaddition, contamination by membranes, enzymes, and organellesderived from vascular tissue would be eliminated.
In the present study, procedures for isolating mesophyll andbundle sheath protoplasts of Panicum miliaceum were developedand the quality of chloroplasts isolated from protoplasts wasevaluated.
MATERIALS AND METHODS
Plant Material and Protoplast Isolation. P. miliaceum, varietySirocket-5, was grown in soil in a greenhouse within a day-nighttemperature range of 30 to 20 C. For isolation of MP and BSP,leaf tissue was taken 15 to 25 days after planting (leaves approxi-mately 70-130 mm in length). In older tissue the bundle sheathstrands became resistant to enzymic digestion which resulted inisolation of MP and bundle sheath strands.
Eight to 10 leaves at a time were sampled, stacked, and cut byhand into small segments about 0.7 mm in width. Approximately8 g of leaf tissue were prepared and subjected to enzymic digestionas previously described (5, 18). The enzyme medium contained0.5 M sorbitol, 1 mm CaCl2, 0.1% BSA, 2% cellulase (Onozuka 3S),and 0.3% pectinase (Macerozyme R-10) (pH 5.0). The tissue wasincubated in a crystallizing dish in a 30 C water bath under lowlight (500 ft-c or about 15 nE cm-2 s-1 between 400 to 700 nmfrom a Phillips 250-w lamp) for 4.5 h (similar yields are obtained
2Abbreviations: MP: mesophyll protoplasts; BSP: Bundle sheath pro-toplasts; PGA: 3-phosphoglycerate; PEP: phosphoenolpyruvate; RuBP:ribulose 1,5-bisphosphate.
821 www.plantphysiol.orgon January 26, 2020 - Published by Downloaded from Copyright © 1979 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 63, 1979
by digestion for 16 h at 18 C, although the shorter incubationperiod was used in the present study). Following incubation, theenzyme medium was gently discarded and the leaf segmentswashed several times with 0.5 M sorbitol, 1 mm CaCl2, and 5 mMHepes-KOH (pH 7.0) (hereafter referred to as the sorbitol me-dium). After each washing, the extract was filtered through acoarse mesh nylon kitchen strainer (1-mm apertures) and a layerof cheese cloth (apertures approximately 200 ,um). The filtrate wascentrifuged for 2 min at 300g and the supernatant discarded.
Protoplast Purification. I. The protoplast types were easy torecognize by light microscopy (see under "Results") which aidedin developing procedures for their separation. The BSP are gen-erally larger and have a higher density than MP. Initially, thecrude protoplast pellet was resuspended in four tubes (15 x 100mm) each containing 4 ml of 0.5 M sucrose, I mm CaCl2, 5 mMHepes-KOH (pH 7.0). This was overlaid by 2 ml of the sorbitolmedium. After centrifugation for 5 min at 300g, the MP parti-tioned at the interphase between the layer of sucrose and sorbitol.The BSP sedimented to the bottom of the tubes and these wereresuspended in a medium containing 0.5 M sucrose, I mm CaCl2,5 mM Hepes-KOH (pH 7.0), and 9.1% dextran T40 (I g dextranT40/10 ml). This mixture was overlaid with 2 ml of the sorbitolmedium and, after centrifugation at 300g for 5 min, the BSPcollected at the interphase between the layer of sucrose-dextranT4o and sorbitol. The MP were washed once and resuspended inthe sorbitol medium. The BSP fraction contained some very small,dense MP (assays of levels of PEP carboxylase verified that thesewere MP). These MP were partially removed by suspending theBSP preparation in 0.5M sucrose, 1 mm CaC12, 5 mm Hepes-KOH(pH 7.0), and centrifuging at 200g for 5 min. The pellet was thenresuspended in the sorbitol medium. This procedure for purifica-tion resulted in MP preparations which were about 95% pure andBSP preparations about 80%o pure based on cross-contaminationas indicated by photosynthetic marker enzymes of the protoplasttypes.
Protoplast Purification. II. A modification of the first purifica-tion procedure in the latter part of the study resulted in anincreased purity of the BSP preparation up to approximately 95%.In this method the crude protoplast pellet was resuspended in 0.5M sucrose, 5mM Hepes-KOH (pH 7.0), and 1 mrm CaCl2 with 13%dextran T40 (1.5 g/10 ml). Four ml of this suspension were addedto each of four tubes. Two ml layers of 0.5M sucrose, 5 mM Hepes-KOH (pH 7.0), and 10.7% dextran T40 (1.2 g dextran T40/10 ml)and then 2 ml of the sorbitol medium were added on top of thesesolutions. After centrifugation at 300g for 5 min, a mixture of MPand BSP appeared at the upper interphase (between the sorbitollayer and sucrose-dextran layer). The bands of protoplasts werecollected with a Pasteur pipette and brought to a volume of 8 mlwith the sorbitol medium. Four ml of this protoplast suspensionwere layered into two tubes over 10 ml of 0.4M sucrose, 0.1 Msorbitol, 5 mm Hepes-KOH (pH 7.0), and 1 mM CaCl2 andcentrifuged for 2 min at 300g. The supernatant contained MP andthe pellet was an enriched preparation of BSP. The MP from thesupernatant fraction were diluted with an equal volume of thesorbitol medium and centrifuged at 300g for 2 min. The pelletwas resuspended in the sorbitol medium and taken as the MPpurification.The BSP pellet was resuspended in 4 ml of sorbitol medium
and the purification repeated by layering on top of 10 ml of themedium containing 0.4M sucrose and 0.1 M sorbitol (describedabove) and centrifuging at 300g for 1 min. The pellet was thentaken as the BSP preparation. This purification procedure takesadvantage of both size and density differences between the pro-toplast types and tends to eliminate any smaller and relativelydense MP from contaminating the BSP fraction. In either purifi-cation procedure from about 10 g of leaf tissue, the yield of MPon a Chl basis was 600 to 1,000,ug and that of BSP about 200 to400 ,Lg. Since there is about 1.8 mg Chl/g leaf tissue, the total
yield of protoplasts from the leaf is roughly 5%. The remainingChl is in undigested bundle sheath strands and broken protoplasts.After isolation, the protoplasts were stored at 0 C.
Light Microscopy. Protoplasts were fixed in 3% glutaraldehydewith 25 mm Hepes buffer (pH 7.0), and 0.3 M sorbitol followed by2% OS04 in 25 mm phosphate buffer. Samples were then washedfour times with the phosphate buffer and photographed, in buffer,with the light microscope using Kodak Panatomic X film.
Isolation of Chloroplasts from Protoplasts. Aliquots from theprotoplast preparations were centrifuged and resuspended in anappropriate medium (see under "Results") for chloroplast isola-tion. Protoplasts equivalent to 100 to 200 ,ig Chl in 0.4 to 0.8 mlof isolation media were broken by several passages through a 20-,um nylon mesh as previously described (5, 22). With less concen-trated suspensions, it becomes difficult to break all of the proto-plasts. Chloroplasts were then isolated by centrifugation at 300gfor 90s. The resuspended pellet, which contained essentially all ofthe Chl of the protoplasts extract, was used for determiningpercentage intactness of chloroplasts according to the ferricyanidetest and for measuring substrate-dependent 02 evolution.Enzyme Assays. Cross-contamination in the preparation of
mesophyll and bundle sheath protoplasts was routinely checkedby osmotically shocking samples of the protoplasts and assayingmarker enzymes (PEP carboxylase, pyruvate, Pi dikinase formesophyll and RuBP carboxylase for bundle sheath) in the totalprotoplast extract. Pyruvate, Pi dikinase (for mesophyll chloro-plasts) and RuBP carboxylase (for bundle sheath chloroplasts)were also used to evaluate the degree of purity and intactness ofchloroplasts isolated from protoplasts. The activity of a bundlesheath marker enzyme in MP preparations divided by the activityof the enzyme in pure bundle sheath cells times 100 would givethe percentage contamination. Likewise, measurement of a markerenzyme for mesophyll cells would provide an estimate of MPcontamination of BSP. With activity expressed per unit Chl, thiswil then give the percentage contamination on a Chl basis. Sincethe activity of marker enzymes in completely pure cells wasunknown, the degree of purity was estimated as follows:
% Contamination of mesophyll protoplasts = (100)(B)
where A and B equal the activity of the bundle sheath markerenzyme in the mesophyll and bundle sheath protoplast prepara-tion, respectively.
% Contamination of bundle sheath protoplast preparation = (C) (100)
where C and D equal the activity of the marker enzyme formesophyll cells in the BSP and MP preparation, respectively. Thisis a satisfactory method when the preparations are relatively pure.The error is negligible when the respective cross-contaminationsare 10%o or less and only slightly overestimated with a cross-contamination up to 20o.
In the experiment reported (Table I), both the levels of enzymesin the protoplast preparations and the percentage intactness of thechloroplast based on retention of marker enzymes were deter-mined. Aliquots of the mesophyll and bundle sheath protoplastswere pelleted and resuspended in the isolation medium containing5 mM Tricine, 3 nm DTT, 3 mM MgCl2, and 0.3M sorbitol (pH8.3). Enzymes were assayed in protoplast extracts, chloroplasts,and supernatant fractions isolated as described above. For assayof RuBP carboxylase, a 0.l-ml aliquot was diluted to 0.3ml with5 mM Tricine, 3 mm DTT, and 3 mM MgCl2 (pH 8.3). Then 10,ulof 5% digitonin was added to each to ensure complete breakage ofchloroplasts and the mixture incubated for 5 min at 30 C prior toassay. For PEP carboxylase, an aliquot of the samples was addedtol/lo volume of 0.5M Hepes-KOH (pH 7.0) for activation(11).For pyruvate, Pi dikinase,50-IL aliquots were added to 10 ,u 25
822 EDWARDS ET AL.
www.plantphysiol.orgon January 26, 2020 - Published by Downloaded from Copyright © 1979 American Society of Plant Biologists. All rights reserved.

CHLOROPLASTS FROM PROTOPLASTS OF A C4 PLANT
Table I. Level of pyruvate,Pi dikinase, PEP carboxylase, and RuBP carboxylase in mesophyll and bundlesheath protoplast extracts, chloroplasts, and supernatant fractions.
Values in parentheses give the percentage enzyme retention by the chloroplasts.
Extract Pyruvate,Pi dikinase PEP carboxylase RuBP carboxylase
pmol Pmol mg'1 imol jmol mg'1 jmol pmol mg-1ml1 minila Chl min'1 ml-I min1I Chl min-I ml-1 min-I Chl min-1
I. MesophyllProtoplast extract 0.93 3.3 7.45 26.5 0.052 0.185Chloroplast (200 g
pellet) 0.80 (92) 2.96 0.19 (3) -- 0.060 0.22Supernatant 0.07 -- 5.9 -- 0.0042 --
II. Bundle Sheath bProtoplast extract 0.051 0.26c 0.24 1.22 0.72 3.7Chloroplast (200 g
pellet) 0.046 0.21c 0.05 __ 0.65 (89) 3.0Supernatant 0.040 -- 0.12 -- 0.08 --
aPart of the protoplast extract was assayed directly and part separated into chloroplast and supernatantfractions by differential centrifugation. In the latter case, the chloroplasts were resuspended in thesame volume as the supernatant so that relative comparisons could then be made on a per volume basis inthe original preparation.
bIn a separate mechanical preparation of bundle sheath cells of Panicun miZiaceum, the level of PEP carboxy-lase in the cell extract was 0.45 ijmol mg-1 Chl min-1.
cCorrecting for small blank rate (minus protoplast extract) in the assay system, these values reduce to 0.09and 0.05 imol mg'1 Chl minm', indicating mesophyll contamination of BSP of 3%.
mM Tricine (pH 8.3), 50 mim DTT, and 50 mm MgCl2 plus 2.5 ,ulof 50 mm K2HPO4. To activate the enzyme, samples were gassedwith N2 and incubated for approximately 2 h at 25 C before assay.
After activation pretreatments described above RuBP carbox-ylase (21), PEP carboxylase (11), and pyruvate, Pi dikinase (1)were assayed as previously described except that the pyruvate, Pidikinase assay mixture included 5.5 mm (NH4)2SO4. The temper-atures of the assays were 30 C for RuBP carboxylase and 22 C forall other enzymes.02 Evolution by Chloroplasts. Ferricyanide-dependent 02 evo-
lution was measured polarographically at 30 C in a twin Clark-type electrode system similar to that described by Delieu andWalker (3). The ferricyanide test on original and osmoticallyshocked preparations was used as a method to determine percent-age intactness of chloroplast preparations as already described(19). Using the method of osmotic shock previously used withspinach (19), the chloroplasts of P. miliaceum gave poor andnonlinear rates of ferricyanide-dependent O2 evolution (with orwithout the uncoupler NH4Cl). By this method, chloroplasts weresuspended in 0.3 M sorbitol, I mM MgCI2, 1 mm MnCl2, 2 mMEDTA, and 50 nmi Hepes-KOH (pH 7.5). The chloroplasts (75pl) were then osmotically shocked by addition to 0.5 ml of water.After 1 min, addition of 0.5 ml of 0.6 M sorbitol, 2 mm MgCl2, 2mM MnCl2, 4 mm EDTA, and 100 mm Hepes-KOH (pH 7.5)restored the original osmotic conditions. This procedure was mod-ified to include 10 mM MgCl2 in the osmotic shock medium.Ferricyanide rates were then linear, stimulated approximately 2-fold by addition of 2.5 mm NH4Cl with maximum rates of 4 to 6JMol 02 evolved mg-' Chl min-'.
Substrate-dependent O2 evolution with intact chloroplasts wasmeasured polarographically at 30 C with a twin Rank electrodesystem (Rank Bros., Bottisham, Cambridge, U.K.). The assayscontained 20 to 30 ug Chi in a volume of 1.8 ml. Light wasprovided by a Voightlander VP 135 projector with an illuminanceof 8,000 ft-c at the surface of the cuvette (approximately 140-150nE cm-2 s-' at 400-700 nm).
Chl. Chl was determined by the method of Arnon (2) in 80%o
acetone or according to Wintermans and De Mots in 96% ethanol(24). The two methods give similar results.
RESULTS AND DISCUSSION
LIGHT MICROSCOPY
P. miliaceum is an NAD-malic enzyme type C4 species charac-terized anatomically by a centripetal location of the bundle sheathchloroplasts. Like many species of this C4 subgroup, the bundlesheath chloroplasts are considerably larger than the mesophyllchloroplasts (7, 10).
Figure I shows preparations of MP and BSP of P. miliaceum.In general, both the bundle sheath protoplasts and their chloro-plasts are larger than those of the mesophyll preparations. Mostof the freshly prepared and fixed BSP showed an asymmetricorientation of chloroplasts characteristic of their position in vivo.This feature is clearly evident in only a small proportion of theseprotoplasts when they are randomly orientated and photographedin narrow plain of focus. Both size and density differences in thetwo types of protoplasts allowed their separation (see under "Ma-terials and Methods"). The reason for the greater density of BSPis uncertain although it might be due to a relatively lower volumeof vacuole per unit volume of cytoplasm than in the mesophyllcells. The tissue was in a rapid state of growth and was sampledearly in the morning so that little starch accumulation in eithermesophyll or bundle sheath chloroplasts was expected.
Previously, enzymic digestion of C4 species resulted in theisolation of MP and bundle sheath strands (e.g. ref. 14). P.miliaceum is the first C4 species from which BSP have beenisolated. Recently, mesophyll and BSP were readily isolated frommature leaves ofAmaranthus graecizans using a similar procedure(Ku and Edwards, unpublished). Differences in the compositionof bundle sheath cell walls among C4 species and in the commer-cial sources of cellulase and pectinase will likely be importantfactors in further efforts to isolate BSP from other C4 species.
Plant Physiol. Vol. 63, 1979 823
www.plantphysiol.orgon January 26, 2020 - Published by Downloaded from Copyright © 1979 American Society of Plant Biologists. All rights reserved.

EDWARDS ET AL. Plant Physiol. Vol. 63, 1979
*i I.S7 ..
,-t s e
I
.-
(. Nr i:*.4,.Pi
eff - I }2
4t. 4w
+n s '4.,' X .ei.r'~~-tt@ * .
J7,
*t \ >
. c4' rs, }
FIG. 1. Light micrograph of mesophyll protoplasts of P. miliaceum (upper figure). Light micrograph of bundle sheath protoplasts of P. miliaceum(lower figure). Bar represents 30,m.
PURITY OF PREPARATIONS
Besides light microscopy, the purity of the preparations wastested by assaying marker enzymes for MP and BSP. In theexperiment of Table I, MP were about 95% pure based on cross-contamination by RuBP carboxylase
0.185 jAmol mg-' Chl min-' in mesophyll preparation3.7 ,umol mg-' Chl min-' in bundle sheath preparation (
(also see under "Materials and Methods"). Likewise, BSP wasabout 93% pure based on cross-contamination by pyruvate, Pidikinase and PEP carboxylase. If corrections are applied for apyruvate, Pi dikinase assay blank and PEP carboxylase activityassociated with bundle sheath cells (see Table I footnotes), BSPpurity was about 97%.
INTACTNESS OF CHLOROPLASTS ISOLATED FROM PROTOPLASTS
Retention of Chloroplast Marker Enzymes. Pyruvate, Pi diki-nase is localized in the stroma of C4 mesophyll chloroplasts (8, 9),and its retention by chloroplasts prepared from MP indicates thatthe chloroplasts are about 90%o intact. Likewise, bundle sheath
chloroplasts were considered about 90%o intact based on the reten-tion of RuBP carboxylase in the 200g chloroplast pellet (Table I).PEP carboxylase, previously concluded to be a cytoplasmic en-zyme (8, 9), was associated with the supernatant fraction frommesophyll protoplast extracts confirming its extrachloroplasticlocation (Table I).
Fernicyanide Test. The impermeability of ferricyanide to intactchloroplasts can be used as a method ofdetermining the intactnessof a chloroplast preparation (19). Ferricyanide-dependent 02 ev-olution is determined before and after osmotic shock. Accordingto this method, chloroplasts prepared from protoplasts of P.miliaceum (protoplast preparation as in Table I) were 98% and82% intact for mesophyll and bundle sheath chloroplasts, respec-
tively. Both retention of chloroplast marker enzymes and theferricyanide test indicate that high proportions of intact chloro-plasts can be obtained from both protoplast types. Slight discrep-ancies between the two methods probably occur due to the use ofseparate isolations. Reducing conditions (inclusion of DTT) wereconsidered essential for extraction for the enzyme assays (Table I)while DTT was eliminated from the preparations for ferricyanidemeasurements in order to avoid its interference with the assay offerricyanide-dependent 02 evolution.
824
t' -4,
r, frwIIN "I".40.
IN
.r7.1
1 0%T..,
A'. .
p1\itt
.1 A
...7.
..." WM.,?.1911k.
w.4
www.plantphysiol.orgon January 26, 2020 - Published by Downloaded from Copyright © 1979 American Society of Plant Biologists. All rights reserved.
CHLOROPLASTS FROM PROTOPLASTS OF A C4 PLANT
SUBSTRATE-DEPENDENT 02 EVOLUTION
Chloroplasts isolated from MP had very low rates of 02 evolu-tion in the light with or without HCO3- whereas bundle sheathchloroplasts had relatively high rates of 02 evolution with HCO3as a substrate (Table II). These results are consistent with previousevidence that bundle sheath but not mesophyll cells contain theCalvin pathway enzymes and capacity for net carbon assimilation(4, 12).The mesophyll chloroplasts show a high capacity for PGA-
dependent 02 evolution supporting previous conclusions that C4mesophyll chloroplasts have the capacity to reduce PGA to triose-P (12). Light-dependent 02 evolution in the presence of oxaloac-etate was severalfold higher in mesophyll than in bundle sheathchloroplasts. This was stimulated by pyruvate only in mesophyllpreparations. Maximum rates of noncycic electron flow dependon both ATP and NADPH being utilized. In mesophyll chloro-plasts, limitation of NADPH-dependent oxaloacetate reductionvia NADP malate dehydrogenase would be relieved by pyruvate-mediated ATP utilization via pyruvate, Pi dikinase (4, 17). Thus,the levels of substrate-dependent 02 evolution by the preparationwith bicarbonate, PGA, and oxaloacetate + pyruvate furthersubstantiate the differing photosynthetic functions of these chlo-roplast types.
Photosynthesis by the isolated bundle sheath chloroplasts was
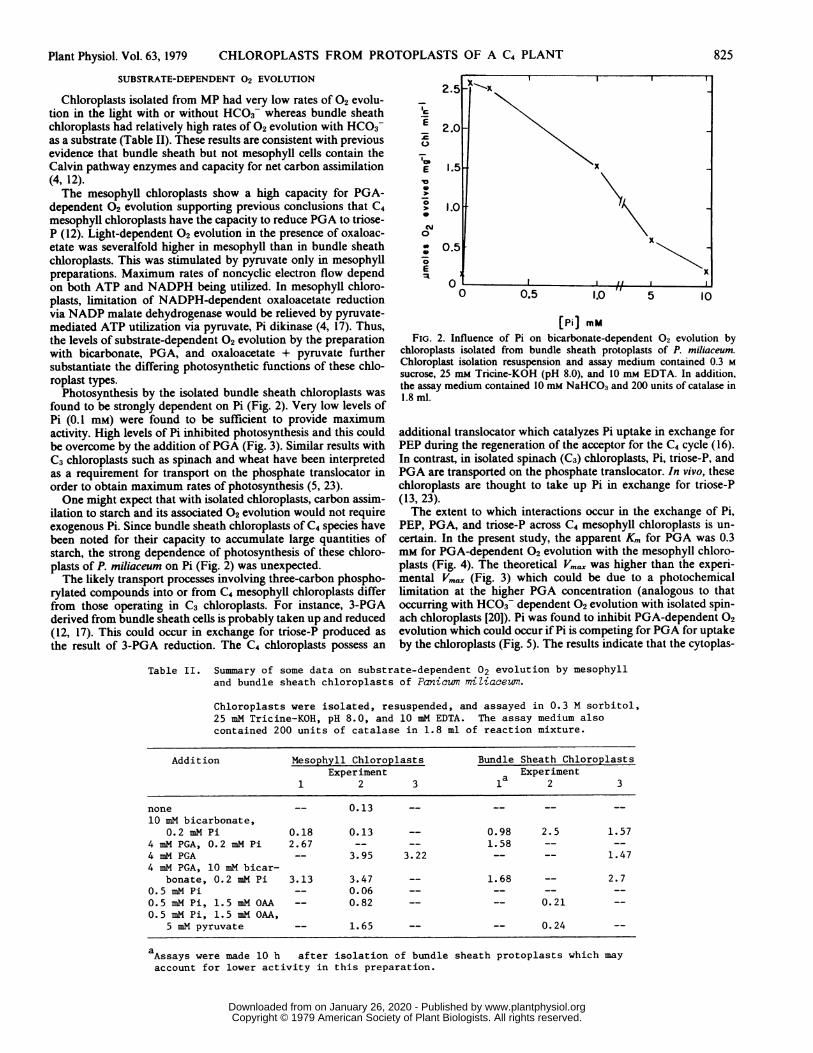
found to be strongly dependent on Pi (Fig. 2). Very low levels ofPi (0.1 mM) were found to be sufficient to provide maximumactivity. High levels of Pi inhibited photosynthesis and this couldbe overcome by the addition ofPGA (Fig. 3). Similar results withC3 chloroplasts such as spinach and wheat have been interpretedas a requirement for transport on the phosphate translocator inorder to obtain maximum rates of photosynthesis (5, 23).One might expect that with isolated chloroplasts, carbon assim-
ilation to starch and its associated 02 evolution would not requireexogenous Pi. Since bundle sheath chloroplasts of C4 species havebeen noted for their capacity to accumulate large quantities ofstarch, the strong dependence of photosynthesis of these chloro-plasts of P. miliaceum on Pi (Fig. 2) was unexpected.The likely transport processes involving three-carbon phospho-
rylated compounds into or from C4 mesophyll chloroplasts differfrom those operating in C3 chloroplasts. For instance, 3-PGAderived from bundle sheath cells is probably taken up and reduced(12, 17). This could occur in exchange for triose-P produced as
the result of 3-PGA reduction. The C4 chloroplasts possess an
2.0
E 0.5 X
10
0
o 5x
0
E x
11 ' I0 0.5 1.0 5 10
[Pi] mm
FIG. 2. Influence of Pi on bicarbonate-dependent 02 evolution bychloroplasts isolated from bundle sheath protoplasts of P. miliaceum.Chloroplast isolation resuspension and assay medium contained 0.3 M
sucrose, 25 mM Tricine-KOH (pH 8.0), and 10 mm EDTA. In addition,the assay medium contained 10 mM NaHCO3 and 200 units of catalase in1.8 ml.
additional translocator which catalyzes Pi uptake in exchange forPEP during the regeneration of the acceptor for the C4 cycle (16).In contrast, in isolated spinach (C3) chloroplasts, Pi, triose-P, andPGA are transported on the phosphate translocator. In vivo, thesechloroplasts are thought to take up Pi in exchange for triose-P(13, 23).The extent to which interactions occur in the exchange of Pi,
PEP, PGA, and triose-P across C4 mesophyll chloroplasts is un-
certain. In the present study, the apparent Km for PGA was 0.3mm for PGA-dependent 02 evolution with the mesophyll chloro-plasts (Fig. 4). The theoretical Vm. was higher than the experi-mental V,,. (Fig. 3) which could be due to a photochemicallimitation at the higher PGA concentration (analogous to thatoccurring with HC03- dependent 02 evolution with isolated spin-ach chloroplasts [20]). Pi was found to inhibit PGA-dependent 02
evolution which could occur if Pi is competing for PGA for uptakeby the chloroplasts (Fig. 5). The results indicate that the cytoplas-
Table II. Summary of some data on substrate-dependent 02 evolution by mesophylland bundle sheath chloroplasts of Panicum miZiaceum.
Chloroplasts were isolated, resuspended, and assayed in 0.3 M sorbitol,25 mM Tricine-KOH, pH 8.0, and 10 mM EDTA. The assay medium alsocontained 200 units of catalase in 1.8 ml of reaction mixture.
Addition Mesophyll Chloroplasts Bundle Sheath ChloroplastsExperiment a Experiment
1 2 3 1 2 3
none -- 0.13 -- -- -- --
10 mM bicarbonate,0.2 mM Pi 0.18 0.13 -- 0.98 2.5 1.57
4 mM PGA, 0.2 mM Pi 2.67 -- -- 1.58 -- --4 mM PGA -- 3.95 3.22 -- -- 1.474 mM PGA, 10 mM bicar-
bonate, 0.2 mM Pi 3.13 3.47 -- 1.68 -- 2.70.5 mM Pi -- 0.06 -- -- -- --
0.5 mM Pi, 1.5 mM OAA -- 0.82 -- -- 0.21 --0.5 mM Pi, 1.5 mM OAA,
5 mM pyruvate -- 1.65 -- -- 0.24 --
aAssays were made 10 h after isolation of bundle sheath protoplasts which mayaccount for lower activity in this preparation.
825Plant Physiol. Vol. 63, 1979
www.plantphysiol.orgon January 26, 2020 - Published by Downloaded from Copyright © 1979 American Society of Plant Biologists. All rights reserved.

EDWARDS ET AL. Plant Physiol. Vol. 63, 1979
.0
0
0
4,)
0
Ec
0 5 10 15 20 25 30
MinutesFIG. 3. Light-dependent 02 evolution by bundle sheath chloroplasts of P. miliaceum. Chloroplasts were isolated, resuspended, and assayed in 0.3 M
sorbitol, 25 mM Tricine-KOH (pH 8.0), and 10 mM EDTA. In addition, the assay medium contained 10 mM NaHCO3, 200 units of catalase/1.8 ml and(a) 0.05 mM Pi, (b) 4 mM Pi + addition of PGA as indicated. Numbers in parentheses indicate rate of 02 evolution as ,umol mg-' Chl min-'.
826
www.plantphysiol.orgon January 26, 2020 - Published by Downloaded from Copyright © 1979 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 63, 1979 CHLOROPLASTS FROM PROTOPLASTS OF A C4 PLANT
/K 0.9E
K
E 2.0 0.6
E II x x.
> 0.35) K Km 0.31mMC\Jo 1.0 Vmax 4.3 jjrnoles mg-1
Chi minf'
o -40 4 8
0 2 3 4
[PGA] mMFIG. 4. Influence of PGA concentration on rate of light-dependent 02
evolution by mesophyll chloroplasts of P. miliaceum. See Table II forisolation and assay media. Inset shows a double reciprocal plot of rate ofphotosynthesis versus PGA concentration.
3.0
E
0
,,, 2.0E-o>.0
&N 1.0
E:a
0 5 10[Pi] mM
FIG. 5. Influence of Pi concentration on rate of PGA-dependent 02evolution with mesophyUl chloroplasts of P miliaceum. See Table II fordescription of isolation and assay medium. In addition, the assay mediumcontained I mm PGA.
Table III. PGA-dependent 02 evolution with chloroplastsisolated from mesophyll protoplasts ofPanicum miliaceum. See Table III for compo-nents of the isolation and assay medium.
Addition Rateimol mg1I Chl min1
(Preparation 1)none 0.131 mM PGA 2.105 mM PEP 0.071 mM PGA, 5 mM PEP 2.30
(Preparation 2)0.3 mM PGA 2.300.3 mM PGA, 3 mM PEP 2.450.5 mM PGA 3.220.5 mM PGA, 1 mM pyridoxal phosphate 0.080.5 mM PGA, 5 mM Pi 1.38
827
02 evolution. However, as shown in Table III, up to 5 mm PEP,or concentrations of PEP 5- to 10-fold higher than that of PGA,had no effect on PGA-dependent O2 evolution. This suggests thatPEP and PGA transport occur on separate carriers. Pyridoxalphosphate has been found to be a strong inhibitor ofthe phosphatetranslocator in spinach chloroplasts (6). Likewise, pyridoxal phos-phate was very inhibitory to PGA-dependent O2 evolution withthe C4 mesophyll chloroplasts (Table III) which could be due toinhibition of carrier-mediated transport.Some evidence exists for specific carrier-mediated transport of
PEP, Pi, and pyruvate with C4 mesophyll chloroplasts (15, 16).Further direct studies on exchange of a number of metabolitesacross envelopes of both C4 mesophyll and bundle sheath chlo-roplasts will be required to understand the transport mechanismsand their regulation in C4 photosynthesis.
Acknowledgments-The technical assistance of Ms. B. Beckenham, Ms. Celia Miller. and Mr.Mark Schmitt is gratefully acknowledged.
LITERATURE CITED
1. ANDREWS TJ, MD HATCH 1969 Properties and mechanisms of action of pyruvate. Pi dikinasefrom leaves. Biochem J 114: 117-125
2. ARNON DI 1949 Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris.Plant Physiol 24: 1-15
3. DELIEU T, DA WALKER 1972 An improved cathode for the measurement of photosyntheticoxygen evolution by isolated chloroplasts. New Phytol 71: 201-225
4. EDWARDs GE, SC HUBER 1978 Usefulness of isolated cells and protoplasts for photosyntheticstudies. In DO Hall, J Coombs, TW Goodwin, eds. IV International Congress on Photosyn-thesis. Biochemical Society. London, pp 95-106
5. EDWARDS GE, SP ROBINSON. NJC TYLER. DA WALKER 1978 Photosynthesis by isolatedprotoplasts, protoplast extracts, and chloroplasts of wheat. Influence of orthophosphate,pyrophosphate, and adenylates. Plant Physiol 62: 313-319
6. FLUGGE Ul, HW HELDT 1977 Specific labeling of a protein involved in phosphate transport ofchloroplasts by pyridoxal-5'-phosphate. FEBS Lett 82: 29-33
7. GUTIERREZ M, VE GRACEN, GE EDWARDS 1974 Biochemical and cytological relationships inC4 plants. Planta 119: 279-300
8. GUTIERREZ M, SC HUBER, SB Ku. R KANAI, GE EDWARDS 1975 Intracellular carbonmetabolism in mesophyll cells of C4 plants. In M Arnon, ed, III International Congress onPhotosynthesis Research. Elsevier, Amsterdam, pp 1219-1320
9. HATCH MD, T KAGAWA 1973 Enzymes and functional capacities of mesophyll chloroplastsfrom plants with C,-pathway photosynthesis. Arch Biochem Biophys 159: 842-853
10. HATCH MD, T KAGAWA, S CRAIG 1975 Subdivision of C4-pathway species based on differingC4 acid decarboxylating systems and ultrastructural features. Aust J Plant Physiol 2: 111-128
11. HATCH MD, IR OLIVER 1978 Activation and inactivation of phosphoenolpyruvate carboxyla-tion in kaf extracts from C4 species. Aust J Plant Physiol 5: 571-580
12. HATCH MD, CB OSMOND 1976 Compartmentation and transport in C4 photosynthesis. In CRStocking, U Heber, eds, Transport in Plants. Encyclopedia of Plant Physiol, New Series. Vol3. Springer-Verlag, New York, pp 144-184
13. HELDT HW, L RAPLEY 1970 Specific transport of inorganic phosphate, 3-phosphoglycerate,and dihydroxyacetonephosphate, and of dicarboxylates across the inner membrane ofspinach chloroplasts. FEBS Lett 10: 143-148
14. HUBER SC, GE EDWARDS 1975 An evaluation of some parameters required for the enzymaticisolation of cells and protoplasts with CO2 fixation capacity from C3 and C4 grasses. PhysiolPlant 35: 203-209
15. HUBER SC, GE EDWARDS 1977 Transport in C4 mesophyll chloroplasts. Characterization ofthe pyruvate carrier. Biochim Biophys Acta 462: 583-602
16. HUBER SC, GE EDWARDS 1977 Transport in C4 mesophyll chloroplasts. Evidence for anexchange of inorganic phosphate and phosphoenolpyruvate. Biochim Biophys Acta 462:603-612
17. KAGAWA T, MD HATCH 1974 Light-dependent metabolism ofcarbon compounds by mesophyllchloroplasts from plants with the C4 pathway of photosynthesis. Aust J Plant Physiol 1: 51-64
18. KANEI R, GE EDWARDS 1973 Separation of mesophyll protoplasts and bundle sheath cellsfrom maize leaves for photosynthetic studies. Plant Physiol 51: 1133-1137
19. LILLEY R MCC, MP FITZGERALD, KG RIENns, DA WALKER 1975 Criteria of intactness andthe photosynthetic activity of spinach chloroplast preparations. New Phytol 75: 1-10
20. LILLEY R MCC, DA WALKER 1975 Carbon dioxide assimilation by leaves, isolated chloroplasts,and ribulose bisphosphate carboxylase from spinach. Plant Physiol 55: 1087-1092
21. LORIMER GH, MR BADGER, TJ ANDREWS 1977 D-Ribulose-1,5-bisphosphate-carboxylase-oxygenase. Improved methods for the activation and assay of catalytic activities. AnalBiochem 78: 66-75
22. RATHNAM CKM. GE EDWARDS 1976 Protoplasts as a tool for isolating photosyntheticallyactive chloroplasts from grass leaves. Plant Cell Physiol 17: 177-186
23. WALKER DA, A HEROLD 1977 Can the chloroplast support photosynthesis unaided? In YFujita, S Katoh, K Shibata, S Miyachi, eds, Photosynthetic Organelles: Structure andFunction. Plant Cell Physiol (special issue), 1-7
24. WINTERMANS JFGM, A DE MOTS 1965 Spectrophotometric characteristics of chlorophyll andtheir pheophytins in ethanol. Biochim Biophys Acta 104: 448-453
x~~~~~~~~~~~~~~~~
mic Pi concentration could influence PGA reduction by themesophyll chloroplasts in vivo.
If PGA and PEP were transported on a common carrier in theenvelope of C4 chloroplasts, PEP should inhibit PGA-dependent
www.plantphysiol.orgon January 26, 2020 - Published by Downloaded from Copyright © 1979 American Society of Plant Biologists. All rights reserved.














![Outstanding Balkan River landscapes a basis for wise ...smallpdf.com].pdf2 1. Hydromorphological intactness of rivers There are four classes characterising the different levels of](https://static.fdocuments.in/doc/165x107/607055892f8058439e131268/outstanding-balkan-river-landscapes-a-basis-for-wise-smallpdfcompdf-2-1.jpg)



