
Isolation andPartial Characterization ofOuterandInner …jb.asm.org/content/145/1/596.full.pdf ·...
9
JOURNAL OF BACTERIOLOGY, Jan. 1981, p. 596-604 0021-9193/81/010596-09$02.00/0 Vol. 145, No. 1 Isolation and Partial Characterization of Outer and Inner Membranes from Encapsulated Haemophilus influenzae Type b MARILYN R. LOEB,`* ARTHUR L. ZACHARY,2 AND DAVID H. SMITH' Department of Pediatrics, University of Rochester Medical Center, Rochester, New York 14642,' and Department of Biological Chemistry, University of Maryland Medical School, Baltimore, Maryland 212012 A method has been developed to separate the cell envelope of encapsulated (type b) Haemophilus influenzae into its outer and inner membrane components with procedures that avoided two problems encountered in fractionation of this envelope: (i) the tendency of the outer and inner membranes to hybridize and (ii) the tendency of the apparently fragile inner membrane to fragment into difficulty sedimentable units. Log phase cells, whose lipids were radioactively labeled, were lysed by passage throdgh a French press. The lysate was applied to a discontin- uous sucrose gradient, and envelope-rich material was collected by centrifugation onto a cushion of dense sucrose under carefully controlled conditions. This material was then further fractionated by isopycnic centrifugation in a sucrose gradient to yield four membrane fractions which were partially characterized. On the basis of their radioactivity, buoyant density, ultrastructure, polypeptide composition, and content of phospholipid, protein, lipopolysaccharide, and suc- cinic dehydrogenase, these fractions were identified as follows: fraction 1, outer membrane vesicles with very little inner membrane contamination (<4%); fraction 2, outer membrane vesicles containing entrapped inner membrane; fraction 3, a protein-rich fraction of inner membrane; fraction 4, a protein-poor fraction of inner membrane. Fractions 3 and 4 contained about 25% outer membrane contam- ination. Haemophilus influenzae is a gram-negative pathogen that occurs in both unencapsulated and encapsulated forms. Of the six encapsulated types (types a through f), type b is most often associated with systemic invasive disease and is a pathogen of serious consequence, especially for young children (31). Whereas the well-charac- terized type b capsule (8) has been recognized as playing a role in the pathogenesis of H. influ- enzae (1), little is known of the significance of other cell surface components, i.e., outer mem- brane proteins and lipopolysaccharide (LPS), in the disease process. Recently, this laboratory demonstrated that infected humans produce antibodies to envelope proteins (R. A. Insel et al., submitted for publi- cation). As a first step toward identifying which envelope proteins are immunogenic, and which, if any, are critical to pathogenesis, we have, as described below, isolated, fractionated, and par- tially characterized the cell envelope of a single strain of encapsulated (type b) H. influenzae. Studies by others have shown similarities be- tween the envelopes of H. influenzae and those of other gram-negative bacteria. As seen in elec- tron micrographs of stationary phase cells of both type b and untypable strains, the envelope consists of an inner and outer membrane (10, 29). Outer membranes isolated by established techniques from an unencapsulated strain, Rd, have a typical polypeptide composition (35). In addition, strain Rd cells have a periplasmic space (27), shed single bilayer vesicles (presum- ably of outer membrane) during growth (17), and appear to contain a specific binding protein in their envelopes, in this case, for homologous double-stranded DNA (9). However, previous studies also revealed some differences. H. influenzae type b appears to be considerably more permeable to ampicillin than Escherichia coli (23) and contains LPS with a very low content (less than 1%) of 2-keto-3-deox- yoctonate (12). Also, H. influenzae undergoes transformation with high efficiency under shift- down nutritional conditions (13), indicating an ability to generate functions for uptake of DNA and for the transfer of this DNA to the cyto- plasm. Not surprisingly, the envelope composi- tion in the competent state is altered (35). In contrast, transformation of the enteric bacteria occurs by a different mechanism and is less efficient (6). In attempting to fractionate the envelopes from rapidly growing log phase H. influenzae 596 on June 5, 2018 by guest http://jb.asm.org/ Downloaded from
Transcript of Isolation andPartial Characterization ofOuterandInner …jb.asm.org/content/145/1/596.full.pdf ·...

JOURNAL OF BACTERIOLOGY, Jan. 1981, p. 596-6040021-9193/81/010596-09$02.00/0
Vol. 145, No. 1
Isolation and Partial Characterization of Outer and InnerMembranes from Encapsulated Haemophilus influenzae
Type bMARILYN R. LOEB,`* ARTHUR L. ZACHARY,2 AND DAVID H. SMITH'
Department of Pediatrics, University ofRochester Medical Center, Rochester, New York 14642,' andDepartment ofBiological Chemistry, University ofMaryland Medical School, Baltimore, Maryland 212012
A method has been developed to separate the cell envelope of encapsulated(type b) Haemophilus influenzae into its outer and inner membrane componentswith procedures that avoided two problems encountered in fractionation of thisenvelope: (i) the tendency of the outer and inner membranes to hybridize and (ii)the tendency of the apparently fragile inner membrane to fragment into difficultysedimentable units. Log phase cells, whose lipids were radioactively labeled, werelysed by passage throdgh a French press. The lysate was applied to a discontin-uous sucrose gradient, and envelope-rich material was collected by centrifugationonto a cushion of dense sucrose under carefully controlled conditions. Thismaterial was then further fractionated by isopycnic centrifugation in a sucrosegradient to yield four membrane fractions which were partially characterized. Onthe basis of their radioactivity, buoyant density, ultrastructure, polypeptidecomposition, and content of phospholipid, protein, lipopolysaccharide, and suc-cinic dehydrogenase, these fractions were identified as follows: fraction 1, outermembrane vesicles with very little inner membrane contamination (<4%); fraction2, outer membrane vesicles containing entrapped inner membrane; fraction 3, aprotein-rich fraction of inner membrane; fraction 4, a protein-poor fraction ofinner membrane. Fractions 3 and 4 contained about 25% outer membrane contam-ination.
Haemophilus influenzae is a gram-negativepathogen that occurs in both unencapsulatedand encapsulated forms. Of the six encapsulatedtypes (types a through f), type b is most oftenassociated with systemic invasive disease and isa pathogen of serious consequence, especially foryoung children (31). Whereas the well-charac-terized type b capsule (8) has been recognized asplaying a role in the pathogenesis of H. influ-enzae (1), little is known of the significance ofother cell surface components, i.e., outer mem-brane proteins and lipopolysaccharide (LPS), inthe disease process.
Recently, this laboratory demonstrated thatinfected humans produce antibodies to envelopeproteins (R. A. Insel et al., submitted for publi-cation). As a first step toward identifying whichenvelope proteins are immunogenic, and which,if any, are critical to pathogenesis, we have, asdescribed below, isolated, fractionated, and par-tially characterized the cell envelope of a singlestrain of encapsulated (type b) H. influenzae.
Studies by others have shown similarities be-tween the envelopes of H. influenzae and thoseof other gram-negative bacteria. As seen in elec-tron micrographs of stationary phase cells ofboth type b and untypable strains, the envelope
consists of an inner and outer membrane (10,29). Outer membranes isolated by establishedtechniques from an unencapsulated strain, Rd,have a typical polypeptide composition (35). Inaddition, strain Rd cells have a periplasmicspace (27), shed single bilayer vesicles (presum-ably of outer membrane) during growth (17),and appear to contain a specific binding proteinin their envelopes, in this case, for homologousdouble-stranded DNA (9).However, previous studies also revealed some
differences. H. influenzae type b appears to beconsiderably more permeable to ampicillin thanEscherichia coli (23) and contains LPS with avery low content (less than 1%) of 2-keto-3-deox-yoctonate (12). Also, H. influenzae undergoestransformation with high efficiency under shift-down nutritional conditions (13), indicating anability to generate functions for uptake of DNAand for the transfer of this DNA to the cyto-plasm. Not surprisingly, the envelope composi-tion in the competent state is altered (35). Incontrast, transformation of the enteric bacteriaoccurs by a different mechanism and is lessefficient (6).
In attempting to fractionate the envelopesfrom rapidly growing log phase H. influenzae
596
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

ENVELOPE OF ENCAPSULATED H. INFLUENZAE 597
possessing type b capsules, we found that thetechniques developed for use with the Rd strain(35) and with enteric bacteria (18, 24) had to bemodified, primarily because the outer and innermembranes tended to hydridize and the innermembranes were unusually fragile. This repo:ctdescribes the techniques we developed for sep-aration of outer and inner membranes of type bH. influenzae and the partial characterization cfthose membranes.
MATERIALS AND METHODS
Media. Solid (1.1% agar) and liquid media consistedof brain heart infusion (BHI, Difco Laboratories) sup-
plemented with 2 ug of NAD per ml and 10 ,ug ofhemin per ml. The NAD stock solution (1 mg/ml) was
sterilized by filtration. The hemin stock solution (1mg/ml in 0.27 M Tris containing 1 mg of histidinehydrochloride per ml) was sterilized by heating for 15min at 65°C with constant stirring.
Cell culture. H. influenzae type b, strain Eag waskept at -70°C in skim milk. A "parent" plate was
made by partially defrosting a skim milk culture andstreaking a loopful onto solid medium. After incuba-tion at 37°C for 18 h, the plate was stored in the coldand used as a source of inoculum for up to 4 days. Aninoculum plate, made by streaking cells from the par-
ent plate onto solid medium, was incubated for about18 h. BHI medium (10 ml) was added to the plate, andthe cells were suspended and then diluted with me-
dium to an optical density of 0.65 to 0.85 at 600 nm. A15-ml portion of this suspension was used to inoculate1 liter of liquid medium. The cells were then incubatedat 37°C, with vigorous aeration provided by shaking,and harvested after about 4 h at an optical density of0.45 at 600 am. The viable count was 5 x 108 cells per
ml, and the doubling time was about 33 min.Radioactive labeling of cells. [2-'4C]acetate (25
to 100 MlCi/liter, 58 Ci/mol; Amersham Corp.) was
added to the medium at the start of cell growth.Carrier was not needed because the rate of incorpo-ration of isotope paralleled the rate of increase in cell
mass. The isotope was incorporated almost exclusively(at least 92%) into fatty acids (see below), and henceit was used a marker for membranes. With 25 ,iCi, 1.2x 106 cpm were incorporated per liter (5 x 10"1 cells).Membrane preparation. Unless stated otherwise,
all procedures were performed at 4°C, and the bufferused was 0.05M Tris hydrochloride, pH 7.8. Cells from1 liter of culture were collected by centrifugation at10,000 x g for 15 min and washed with 120 and then60 ml of buffer. The pellet was stored at -200C untilneeded, but usually just overnight. The pellet was
defrosted and suspended in 12 ml of buffer, and deox-yribonuclease I and ribonuclease A (Worthington Bio-chemical Corp.) were added (final concentration, 100
yg/ml); the cells were passed twice through a Frenchpressure cell (Aminco) at 8,000 lb/in2. Unbroken cellswere removed by centrifugation at 12,000 x g for 5min. The membranes were partially purified by lay-ering the lysate onto two discontinuous sucrose gra-
dients (6 ml of lysate per gradient), consisting of 8 ml
of 2.02 M sucrose in buffer overlaid with 10 ml of 0.44
M sucrose in buffer, and then centrifuging the gra-dients in a 60 Ti rotor at 60,000 rpm (255,000 x g) fora total of 75 min in a Beckman L2-65B centrifuge. Thebrake was used to stop the rotor. This procedureconcentrated the membranes at the interface of thetwo sucrose solutions. (Note: These conditions arecritical to the separation of membrane-rich materialfrom contaminating cytosol. Other rotors and othercentrifuges will require different conditions of centrif-ugation.) The top layer was carefully removed with aPasteur pipette, and the middle (membrane) layer(about 2.5 ml per gradient) was collected. Collectionwas performed in a darkened room, the membranelayer being visualized, due to its light scattering ability,by lighting from above with a flashlight. The mem-brane layers from the two prepurification gradientswere combined, diluted with buffer to 13 ml, andlayered onto four linear sucrose gradients (3 ml ofmembrane per gradient) of 2.02 M sucrose to 0.99 Msucrose in buffer formed on top of a 2-ml layer of 2.02M sucrose in buffer. The gradients were centrifuged at27,000 rpm (96,000 x g) in an SW27 rotor for 65 h,since centrifugation for less time yielded poorer reso-lution of membrane fractions. A total of 20 drop frac-tions were collected from the bottom of each tube,yielding about 30 fractions per gradient. The distri-bution of membranes was determined by analyzingeach fraction for radioactivity, succinic dehydrogenase(SDH), and optical density at 260 and 280 nm. Use offreshly grown rather than frozen cells did not alter theresults.Membranes were obtained for ultrastructural and
chemical analysis by pooling selected gradient frac-tions which were then diluted with about 4 volumes ofbuffer and pelleted by centrifuging at 255,000 x g for3 h. The membrane pellets were suspended in 1 ml ofwater by forcing them through hypodermic needles ofincreasingly smaller sizes (18, 21, and 26 gauge). Aportion was assayed for SDH. Another portion, forsubsequent sodium dodecyl sulfate-polyacrylamide gelelectrophoresis (SDS-PAGE), was added (2:1, vol/vol)to buffer consisting of 1 volume of 0.5 M Tris (pH 6.8),1 volume of glycerol, and 1 volume of 10% SDS; thissolution was frozen at -20°C. The remainder wasfrozen at -20°C and later assayed for total phosphate,phospholipid phosphate, nucleic acid phosphate, andratio of hydroxy fatty acids to total fatty acids.SDS-PAGE. SDS-PAGE was performed according
to Laemmli (19) by using 1.5-mm thick 10% polyacryl-amide slab gels, with the following changes: the finalconcentrations of N,N,N',N'-tetramethylenediamineand ammonium persulfate were 0.05% and 0.1%, re-spectively. Unless noted otherwise, the membranesamples were solubilized in sample buffer, containing3% 2-mercapthethanol, by heating at 100°C for 5 min.Gels were electrophoresed at 25 mA, fixed, stainedwith Coomassie blue R, and destained (11).
Analytical procedures. Protein was determinedby the procedure of Lowry et al. (22). SDH wasdetermined by the procedure of Osborn et al. (24),except that the assay was done in the presence of 1%Triton X-100. This detergent increased the activity ofthe samples by about threefold, equally across thegradient as well as in the unfractionated membranes.The activity was calculated in relative units as optical
VOL. 145, 1981
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

598 LOEB, ZACHARY, AND SMITH
density change per 5 min. The presence of type bcapsule was determined qualitatively by counterim-munoelectrophoresis (15). Phosphate was determinedby the method ofAmes (2). The relative concentrationof LPS was determined by measuring the ratio ofhydroxy fatty acids to total fatty acids. This assay wasbased on the demonstrated presence of /8-hydroxy-myristic acid in LPS of type b H. influenzae (A.Flesher, Ph.D. thesis, Boston University, 1977) and itslow levels in lipids extracted by the procedure of Blighand Dyer (4) (data not shown). Samples were hydro-lyzed with 4 N KOH (25), acidified with 6 N HCO, andextracted three times with diethyl ether. The etherextract was subjected to thin-layer chromatographyon Bakerflex silica gel IB2 sheets (9.5 cm long) in asolvent system that separates hydroxy fatty acids froma band containing saturated and unsaturated fattyacids (5); the relative radioactivity in each band wasdetermined by counting 5-mm strips of the chromat-ogram. The phospholipid phosphorus content of eachmembrane fraction was determined by assaying thechloroform-soluble layer obtained after extraction induplicate by the procedure of Bligh and Dyer (4), withphospholipid being calculated on the basis of a molec-ular weight of 700 per mole of phosphorus. Nucleicacid content of membrane fractions was calculatedfrom the phosphorus content of hot trichloroaceticacid-soluble phosphate after removal of cold acid-sol-uble and organic solvent-soluble material (26). Thedistribution of isotope into cellular components wasdetermined by the procedure of Roberts et al. (26)with the following results: 0.9% of the isotope incor-porated appeared in the cold trichloroacetic acid-sol-uble fraction, 77% appeared in the combined ethanoland ethanol-diethylether-soluble fractions; 1% ap-peared in the hot acid-soluble fraction, 5% was releasedfrom the remaining residue by pronase, and 15% wasreleased from that residue as petroleum ether-solublematerial after hydrolysis with 4 N KOH for 5 h andsubsequent acidification. Hence, at least 92% of labelwas incorporated into lipid.
Electron microscropy. Samples were fixed for 1h at room temperature in 2% glutaraldehyde bufferedto pH 7.4 with 0.2M sodium cacodylate. After a bufferrinse, the samples were fixed an additional 3 h at roomtemperature in 1% aqueous OsO4. The samples weredehydrated, by using a graded acetone series, andembedded in Epon 812. Thin sections were stainedwith uranyl acetate (1.5% wt/vol) and lead citrate (33)before examination in an electron microscope (HitachiHU-11-B, 75 kV). Measurement of membrane vesicleprofile diameters were made in the longest vesicleprofile dimension, and the values reported representthe range observed for at least 20 randomly chosenvesicle profiles.
RESULTS
Fractionation ofmembranes. Electron mi-crographs of thin sections of H. infiuenzae re-
vealed an envelope typical of gram-negative ba-cilli (data not shown; 7, 10, 29). Nevertheless,difficulties were encountered in attempts to sep-arate and isolate outer and inner membranes byusing established procedures (3, 17, 20, 22, 30) or
modifications of those procedures in which, forexample, concentrations of buffers and Mg2",types of buffers, and incubation with nucleaseswere varied. Inner membrane was found alonewith outer membrane in a heavy region (i.e.,buoyant density, 1.21 g/ml) of the gradient, or
inner membrane appeared in a light region ofthe gradient, but was either heavily contami-nated with outer membrane or obtained in verylow yields.The cause of these problems became obvious
when the membrane fractionation was followedquantitatively by using cells carrying radioactivelabel exclusively in their fatty acids. Use of son-ication or too high a pressure in the French pressresulted in fragmentation ofthe inner membraneinto small pieces that did not readily sediment(see below). In addition, the collection of the cellenvelope as a packed pellet by ultracentrifuga-tion of the lysate interfered with the subsequentseparation of outer and inner membrane upon
isopycnic centrifugation of this envelope mate-rial (data not shown).We avoided these difficulties by briefly cen-
tifuging the lysate through a discontinuous su-crose gradient onto a layer of 2.02 M sucrose.This separated envelope-rich material from therest of the lysate, as seen by the distribution oftotal radioactivity, SDH and material absorbingat 280 and 260 nm in the top, bottom, and middlelayers of this prepurification gradient (Table 1).At least 75% of the radioactivity and enzymeactivity, but less than 30% of the total UV-ab-sorbing material in the cell lysate appear in themiddle layer. The low OD2so/OD2ws ratio in thetop layer indicates that the bulk of the cellularnucleic acid is separated from the middle layer.Isopycnic centrifugation of the membrane-en-riched fraction resolved the envelope into fourfractions (Fig. 1A) with the following properties.Fraction 1, tubes 8 through 10, had a buoyantdensity of 1.23 g/ml and a small proportion (lessthan 4%) of the total SDH activity. Fraction 2,tubes 11 through 14, had a buoyant density of1.22 to 1.20 g/ml and also differed from fraction1 by having a higher proportion of the totalSDH. Although fractions 1 and 2 representedthe heavy and light sides, respectively, ofa singlepeak, data presented below justified their sepa-ration into 2 fractions. Fraction 3, tubes 16
TABLE 1. Prepurification ofmembranes% of total ODml
Layer OD280Lcpm ODm ODmw SDH 0Dm
Top 20 72 79 20 0.41Middle 76 26 19 78 0.61Bottom 5 2 2 2 0.59
J. BACTERIOL.
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

ENVELOPE OF ENCAPSULATED H. INFLUENZAE 599
through 22, had a buoyant density of 1.18 g/mlat its peak and most of the SDH activity. Frac-tion 4, tubes 23 through 27, was a small fractionwith a buoyant density of 1.15 g/ml and only a
small proportion of the SDH. At least 75% of theenzyme activity applied to the gradient was re-
covered. Type b capsule, assayed qualitatively,appeared in fractions 8 to 32 (data not shown).
Different results were obtained when the pre-purification step was omitted and the cell lysatewas subjected directly to isopycnic centrifuga-
1.0
000.4 %201
BOTTOM 10 20 30 10 20 30
TUBE NO.
FIG. 1. (A) Isopycnic centrifugation ofprepurifiedcell lysate. (B) Isopycnic centrifugation of total celllysate. Each gradient represents about 180 ml ofcells.Symbols: (-) cpm per 0.05 ml; (- - -) SDH units per0.3 ml; (.... ) OD2Ls, uncorrected for light scattering.
tion, thus allowing all of the membrane to beaccounted for (Fig. 1B). Notably, the amount ofmaterial in fraction 4 was increased. This sug-gested that the membrane material remaining inthe top layer of the prepurification gradient(Table 1) consists largely of fraction 4 material,and that loss of fraction 4 in the prepurificationstep was due to the small size of some of itscomponents. However, comparison ofthe opticaldensity profiles of Figures 1A and 1B revealedthat all fractions were purified by the prepurifi-cation step.Chemical characterization of the four
fractions. To further characterize the four frac-tions resolved on isopycnic gradients (Fig. 1A),they were each collected by ultracentrifugationand analyzed. Ninety-nine percent of the radio-activity in fractions 1 and 2 was pelleted after 1h of centrifugation at 255,000 x g. Fractions 3and 4 required 3 h to pellet 95% of the radioac-tivity, an indication of their relatively smallersize. The pellet of fraction 1 was nearly colorless,the pellets of fractions 2 and 4 had some yellowcolor, and the pellet of fraction 3 had the yellow-brown color common to membranes containingcytochromes.The biochemical properties of these four frac-
tions (Table 2) can be summarized as follows.Fraction 1 contained the largest proportion ofLPS, and the lowest absolute amounts and spe-
cific activity of SDH. Fraction 2 was similar tofraction 1, but contained more SDH. Fraction 3had considerably less LPS than fractions 1 and2 and was highly enriched in SDH. Fraction 4,a very small fraction, was low in both SDH andLPS.As determined by SDS-PAGE, fractions 1 and
2 had a very similar polypeptide composition(Fig. 2, lanes 1 and 2, respectively). Fraction 2possessed some faint bands not present in frac-tion 1 but found in greater concentrations infractions 3 and 4 (lanes 3 and 4). Fraction 3 hada unique set of proteins in addition to containing
TABLE 2. Composition of the four membrane fractions'Ratio (hy-
Fraction Protein (mg/ml) Phospholipid Phosphorus SDH (U/ml) acids: total LPS (%)bfatty acids)
1 2.5 (26) 1.3 (29) 0.097 (31) 0.4 (3) 0.126 402 4.0 (41) 1.9 (43) 0.134 (42) 2.1 (14) 0.096 493 2.8 (29) 0.84 (19) 0.065 (20) 11.7 (77) 0.034 84 0.50 (5) 0.39 (9) 0.022 (7) 0.7 (5) 0.027 3
a Data are from duplicate determinations in two experiments. Figures in parentheses indicate percents oftotal.
b Calculated as follows for each fraction:
hydroxy fatty acids cpm in fractiontotal fatty acids cpm in all fractions
VOL. 145, 1981
III :-., 0
CY0 -,-"- z
..- a
& co.t.0
;_
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

600 LOEB, ZACHARY, AND SMITH
some of the bands present in the first two frac-tions. Fraction 4 was simpler in compositionthan fraction 3 and was enriched in some of the
- -
a-_4_
FIG. 2. SDS-PAGE of the four membrane fr-ac-tions. Lanes 1 through 4 correspond to fr-actions 1through 4; 30 pg ofprotein were applied to each lane.
J. BACTERIOL.
proteins present in fraction 3.The pattern of polypeptide composition of
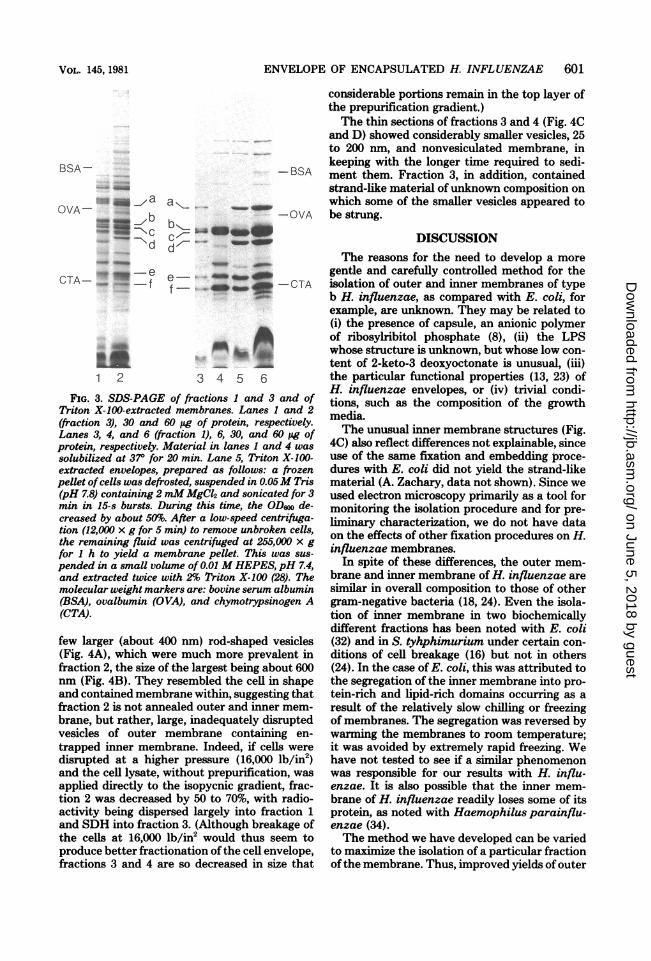
fraction 1 was typical of outer membranes (3,18), and was as follows. (i) Its composition wasrelatively simple with about six polypeptides,labeled a through f, predominating (see Fig. 3,lane 6). These proteins, visible even when smallamounts of protein were applied to the gel (Fig.3, lane 3) had the following approximate molec-ular weights: a, 46,000; b, 38,000; c, 37,000; d,34,000; e, 28,000; and f, 26,000. In addition, therewas another major protein, of 15,000 molecularweight, and a faint-staining band with the mo-bility of free lipoprotein (14) that were visible on15% gels (data not shown). Further studies arein progress to determine if the latter actually islipoprotein and if the faint staining is due to itslow concentration or to interference of stainingby LPS (21). (ii) Fraction 1 contained two heat-modifiable polypeptides, proteins a and d. Thus,gels of samples solubilized at 37°C for 20 mininstead of at 1000C for 5 min did not showproteins a and d (Fig. 3, lane 4), but did displaynew bands. (iii) Extraction of cell envelopes ofH. influenzae with 2% Triton X-100 (28) re-moved inner membrane proteins and yielded apreparation with a polypeptide pattern resem-bling fraction 1 (Fig. 3, lane 5); however, poly-peptide b was present in reduced amounts.The polypeptide composition of fraction 3 had
a pattern similar to the inner membrane of E.coli or Salmonella typhimurium, i.e., there weremany bands, the majority having a molecularweight above 30,000 (Fig. 3, lane 2). From thedata in Fig. 3 and the LPS data (Table 2), weestimated the extent of contamination of frac-tion 3 by fraction 1 or 2 to be about 25%.The above results suggested the following
fraction compositions: Fraction 1, outer mem-brane that was only slightly contaminated withinner membrane; fraction 2, a mixture of outerand inner membrane with the former predomi-nating; and fraction 3, largely inner membranewith about a 25%-contamination with outermembrane. Fraction 4 was an anomaly; it had abuoyant density typical of inner membrane butonly low levels of SDH, and, as indicated bypreliminary data, of D-lactic dehydrogenase andNAD dehydrogenase. However, it did containpolypeptides that are a subclass of fraction 3polypeptides. Based on these data, we suggestthat fraction 4 is derived from the inner mem-brane.Ultrastructure of the four fractions. Each
of the fractions could be easily distinguished inthe thin-section electron micrographs shown inFigure 4A, B, C, and D. Fraction 1 consistedlargely of single ovoid-shaped membrane vesi-cles, 100 to 200 nm in diameter. There were a
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
ENVELOPE OF ENCAPSULATED H. INFLUENZAE 601
BSA -
-, ai, a
OVA-
CTA-_ _e2- _ f
1 2
a ,_ _w 4-OVA
-r _b_,,_.q o
de' - -_ef
ap4CT-CA
3 4 5 6
FIG. 3. SDS-PAGE of fractions I and 3 and ofTriton X-100-extracted membranes. Lanes 1 and 2(fraction 3), 30 and 60 pg of protein, respectively.Lanes 3, 4, and 6 (fraction 1), 6, 30, and 60 pg ofprotein, respectively. Material in lanes 1 and 4 was
solubilized at 37° for 20 min. Lane 5, Triton X-100-extracted envelopes, prepared as follows: a frozenpellet ofcells was defrosted, suspended in 0.05M Tris(pH 7.8) containing 2 mM MgCl2 and sonicated for 3min in 15-s bursts. During this time, the OD600 de-creased by about 50%. After a low-speed centrifuga-tion (12,000 x g for 5 min) to remove unbroken cells,the remaining fluid was centrifuged at 255,000 x gfor 1 h to yield a membrane pellet. This was sus-
pended in a small volume of 0.01 M HEPES, pH 7.4,and extracted twice with 2% Triton X-100 (28). Themolecular weight markers are: bovine serum albumin(BSA), ovalbumin (OVA), and chymotrypsinogen A(CTA).
few larger (about 400 nm) rod-shaped vesicles(Fig. 4A), which were much more prevalent infraction 2, the size of the largest being about 600nm (Fig. 4B). They resembled the cell in shapeand contained membrane within, suggesting thatfraction 2 is not annealed outer and inner mem-brane, but rather, large, inadequately disruptedvesicles of outer membrane containing en-
trapped inner membrane. Indeed, if cells weredisrupted at a higher pressure (16,000 lb/in2)and the cell lysate, without prepurification, was
applied directly to the isopycnic gradient, frac-tion 2 was decreased by 50 to 70%, with radio-activity being dispersed largely into fraction 1and SDH into fraction 3. (Although breakage ofthe cells at 16,000 lb/in2 would thus seem toproduce better fractionation of the cell envelope,fractions 3 and 4 are so decreased in size that
considerable portions remain in the top layer ofthe prepurification gradient.)The thin sections of fractions 3 and 4 (Fig. 4C
and D) showed considerably smaller vesicles, 25to 200 nm, and nonvesiculated membrane, inkeeping with the longer time required to sedi-ment them. Fraction 3, in addition, containedstrand-like material ofunknown composition onwhich some of the smaller vesicles appeared tobe strung.
DISCUSSIONThe reasons for the need to develop a more
gentle and carefully controlled method for theisolation of outer and inner membranes of typeb H. influenzae, as compared with E. coli, forexample, are unknown. They may be related to(i) the presence of capsule, an anionic polymerof ribosylribitol phosphate (8), (ii) the LPSwhose structure is unknown, but whose low con-tent of 2-keto-3 deoxyoctonate is unusual, (iii)the particular functional properties (13, 23) ofH. influenzae envelopes, or (iv) trivial condi-tions, such as the composition of the growthmedia.The unusual inner membrane structures (Fig.
40) also reflect differences not explainable, sinceuse of the same fixation and embedding proce-dures with E. coli did not yield the strand-likematerial (A. Zachary, data not shown). Since weused electron microscopy primarily as a tool formonitoring the isolation procedure and for pre-liminary characterization, we do not have dataon the effects of other fixation procedures on H.influenzae membranes.
In spite of these differences, the outer mem-brane and inner membrane of H. influenzae aresimilar in overall composition to those of othergram-negative bacteria (18, 24). Even the isola-tion of inner membrane in two biochemicallydifferent fractions has been noted with E. coli(32) and in S. tyhphimurium under certain con-ditions of cell breakage (16) but not in others(24). In the case of E. coli, this was attributed tothe segregation of the inner membrane into pro-tein-rich and lipid-rich domains occurring as aresult of the relatively slow chilling or freezingof membranes. The segregation was reversed bywarming the membranes to room temperature;it was avoided by extremely rapid freezing. Wehave not tested to see if a similar phenomenonwas responsible for our results with H. influ-enzae. It is also possible that the inner mem-brane of H. influenzae readily loses some of itsprotein, as noted with Haemophilus parainflu-enzae (34).The method we have developed can be varied
to maximize the isolation of a particular fractionofthe membrane. Thus, improved yields of outer
VOL. 145, 1981
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

602 LOEB, ZACHARY, AND SMITH
i.
faction4.Scale bar equals 200 nm.
membrane free of inner membrane can be ob-tained by breaking the cells at 16,000 lb/in2prepurifying the lysate and centrifuging the pre-purified material by sedimentation for 16 h on alinear sucrose gradient. Under these conditions,fraction 2 will be decreased in amount, and theinner membrane either will be left behind in theprepurification gradient or will only partiallysediment through the linear gradient. Isolationof inner membrane free of outer membrane andcytoplasmic constituents would be more diffi-cult, but could be approached by breaking the
cells at 8,000 lb/in2, subjecting the total lysateto isopycnic centrifugation for 65 h and thencollecting fractions 3 and 4 by ultracentrifuga-tion for 3 h. We did try to further purify fraction3 as obtained in Fig. 1A by applying it to asecond isopycnic gradient; we obtained a smallpeak in the fraction 2 region of the gradient anda larger peak of fraction 3. However, we did notsubsequently try to collect and characterize thepurified fraction 3. We also found that our basicprocedure gave the same results if 0.01 M N-2-hydroxyethylpiperazine-N'-2'-ethanesulfonic
J. BACTERIOL.
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

ENVELOPE OF ENCAPSULATED H. INFLUENZAE 603
acid (HEPES), pH 7.4, or 0.01 M HEPES-0.002M EDTA, pH 7.4, were used throughout.The latter might be helpful in obtaining frac-tions 3 and 4 with less contamination of cyto-plasmic material.
In summary, this report describes a methodfor fractionating the cell envelope of type b H.influenzae into four fractions and their partialcharacterization. Fraction 1, representing almostpure outer membrane, will be useful in our
planned studies on the role of outer membraneproteins in pathogenesis.
ACKNOWLEDGMENTS
We appreciate the excellent technical assistance of ThomasJ. M. McLaughlin and Suzanne Connett and the typing as-sistance of Mary Henning and Susan Brightman. We thankour colleagues Porter Anderson and Richard Insel for review-ing this manuscript.
This work was supported by Public Health Service grantAI-14411 from the National Institute of Allergy and InfectiousDiseases to D.H.S.
LITERATURE CITED
1. Alexander, H. E. 1965. The Hemophilus group, p. 724.In R. J. Dubos, and J. G. Hirsch (ed.), Bacterial andmycotic infections of man. J. B. Lippincott Co. Phila-delphia.
2. Ames, B. N. 1966. Assay of inorganic phosphate, totalphosphate and phosphatases. Methods Enzymol. 8:115-118.
3. Ames, G. F.-L., E. N. Spudich, and H. Nikaido. 1974.Protein composition of the outer membrane of Salmo-nella typhimurium: effect of lipopolysaccharide muta-tions. J. Bacteriol. 117:406-416.
4. Bligh, E. G., and W. J. Dyer. 1959. A rapid method oftotal lipid extraction and purification. Can. J. Biochem.Physiol. 37:911-917.
5. Braun, V., and K. Rehn. 1969. Chemical characteriza-tion, spatial distribution and function of a lipoprotein(murein-lipoprotein) of the E. coli cell wall. The specificeffect of trypsin on membrane structure. Eur. J. Bio-chem. 10:426-438.
6. Cosloy, S. D., and M. Oishi. 1973. Genetic transforma-tion in Escherichia coli K12. Proc. Natl. Acad. Sci.U.S.A. 70:84-87.
7. Costerton, J. W., J. M. Ingram, and K.-J. Cheng.1974. Structure and function of the cell envelope ofgram-negative bacteria. Bacteriol Rev. 38:87-110.
8. Crisel, R. M., R. S. Baker, and D. E. Dorman. 1975.Capsular polymer of Haemophilus influenzae, type b.I. Structural characterization of the capsular polymerof strain Eagan. J. Biol. Chem. 250:4926-4930.
9. Deich, R. A., and H. 0. Smith. 1980. Mechanism ofhomospecific DNA uptake in Haemophilus influenzaetransformation. Mol. Gen. Genet. 177:369-374.
10. Doern, G. V., and F. L. A. Buckmire. 1976. Ultrastruc-tural characterization of capsulated Haemophilus influ-enzae type b and two spontaneous nontypable mutants.J. Bacteriol. 127:523-535.
11. Fairbanks, G., T. L. Steck, and D. F. H. Wallach. 1971.Electrophoretic analysis of the major polypeptides ofthe human erythrocyte membrane. Biochemistry 10:2606-2617.
12. Flesher, A., and R. A. Insel. 1978. Characterization oflipopolysaccharide of Haemophilius influenzae. J. Inf.Dis. 138:719-730.
13. Herriott, R. M., E. Y. Meyer, M. Vogt, and M. Modan.
1970. Defined medium for growth of Haemophilus in-fluenzae. J. Bacteriol. 101:513-516.
14. Hirashima, A., H. C. Wu, P. S. Venkateswaren, andM. Inouye. 1973. Two forms of a structural lipoproteinin the envelope of Escherichia coli. J. Biol. Chem. 248:5654-5659.
15. Ingram, D. L., P. Anderson, and D. H. Smith. 1972.Countercurrent immunoelectrophoresis in the diagnosisof systemic diseases caused by Hemophilus influenzaetype b. J. Pediat. 81:1156-1159.
16. Jones, N. C., and M. J. Osborn. 1977. Translocation ofphospholipids between the outer and inner membranesof Salmonella typhimurium. J. Biol. Chem. 252:7405-7412.
17. Kahn, M., M. Concino, R. Gromkova, and S. Goodgal.1979. DNA binding activity of vesicles produced bycompetence deficient mutants of Haemophilus. Bio-chem. Biophys. Res. Comm. 87:764-772.
18. Koplow, J., and H. Goldfine. 1974. Alterations in theouter membrane of the cell envelope of heptose-defi-cient mutants of Escherichia coli. J. Bacteriol. 117:527-543.
19. Laemmli, U. K. 1970. Cleavage of structural proteinsduring the assembly of the head of bacteriophage T4.Nature (London) 227:680-685.
20. Loeb, M. R., and J. Kilner. 1978. Release of a specialfraction of the outer membrane from both growing andphage T4-infected Escherichia coli B. Biochim. Bio-phys. Acta 514:117-127.
21. Loeb, M. R., and J. Kilner. 1979. Lipopolysaccharideinterferes with the staining of lipoprotein on polyacryl-amide gels. Biochim. Biophys. Acta 544:676-679.
22. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with the Folinphenol reagent. J. Biol. Chem. 193:265-275.
23. Medeiros, A. A., and T. F. O'Brien. 1975. Ampicillin-resistant Haemophilus influenzae tybe b possessing aTEM-type 8-lactamase but little permeability barrierto ampicillin. Lancet i:716-719.
24. Osborn, M. J., J. E. Gander, E. Parisi, and J. Carson.1972. Mechanism of assembly of the outer membraneof Salmonella typhimurium. Isolation and characteri-zation of cytoplasmic and outer membrane. J. Biol.Chem. 247:3962-3972.
25. Rick, P. D., L. W.-M. Fung, C. Ho, and M. J. Osborn.1977. Lipid A mutants of Salmonella typhimurium.Purification and characterization of a lipid precursorproduced by a mutant in 3-deoxy-D-mannooctuloson-ate-8-phosphate synthetase. J. Biol. Chem. 252:4904-4912.
26. Roberts, R. B., D. B. Cowie, P. H. Abelson, E. T.Bolton, and R. J. Britten. 1963. Studies of biosyn-thesis in Escherichia coli. Carnegie Institute of Wash-ington publication 607, Washington, D.C.
27. Rodden, J. L., and J. J. Scocca. 1972. Purification andproperties of cyclic phosphodiesterase; 3'-nucleotidase,a periplasmic enzyme of Hemophilus influenzae. Arch.Biochem. Biophys. 153:837-844.
28. Schnaitman, C. A. 1971. Solubilization of the cytoplas-mic membrane of Escherichia coli by Triton X-100. J.Bacteriol. 108:545-552.
29. Sherwin, R. P., and Wilkins, J. 1973. The ultrastructureof Hemophilus influenzae, p. 143-151. In S. H. Sell(ed.), Hemophilus influenzae. Vanderbilt UniversityPress, Nashville, Tenn.
30. Simon, L. D., T. J. M. McLaughlin, D. Snover, J. Ou,C. Grisham, and M. Loeb. 1975. E. coli membranelipid alteration affecting T4 capsid morphogenesis. Na-ture (London) 256:379-383.
31. Turk, D. C., and J. R. May. 1967. Haemophilus influ-enzae, p. 24ff. The English University Press, London.
32. VanHeerikhuisen, H., E. Kwak, E. F. J. VanBruggen,and B. Witholt. 1975. Characterization of a low density
VOL. 145, 1981
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

604 LOEB, ZACHARY, AND SMITH
cytoplasmic membrane subfraction isolated from Esch-erichia coli. Biochim. Biophys. Acta 413:177-191.
33. Venable, J. H., and R. Coggeshall. 1965. A simplifiedlead citrate stain for use in electron microscopy. J. CellBiol. 25:407-408.
34. White, D. C., and L. Smith. 1962. Hematin enzymes of
J. BACTERIOL.
Hemophilus parainfluenzae. J. Biol. Chem. 237:1332-1336.
35. Zoon, K. C., and J. J. Scocca. 1975. Constitution of thecell envelope of Haemophilus influenzae in relation tocompetence for genetic transformation. J. Bacteriol.123:666-677.
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from


















