
Iron Reductases from Pseudomonas aeruginosa - Journal of
6
JOURNAL OF BACTERIOLOGY, Jan. 1980, p. 199-204 0021-9193/80/01-0199/06$02.00/0 Vol. 141, No. 1 Iron Reductases from Pseudomonas aeruginosa CHARLES D. COX Department of Microbiology, University of Iowa, Iowa City, Iowa 52242 Cell-free extracts of Pseudomonas aeruginosa contain enzyme activities which reduce Fe(III) to Fe(II) when iron is provided in certain chelates, but not when the iron is uncomplexed. Iron reductase activities for two substrates, ferripy- ochelin and ferric citrate, appear to be separate enzymes because of differences in heat stabilities, in locations in fractions of cell-free extracts, in reductant specific- ity, and in apparent sizes during gel filtration chromatography. Ferric citrate iron reductase is an extremely labile activity found in the cytoplasmic fraction, and ferripyochelin iron reductase is a more stable activity found in the periplasmic as well as cytoplasmic fraction of extracts. A small amount of activity detectable in the membrane fraction seemed to be loosely associated with the membranes. Although both enzymes have highest activity with reduced nicotinamide adenine dinucleotide, reduced glutathione also worked with ferripyochelin iron reductase. In addition, oxygen caused an irreversible loss of a percentage of the ferripyochelin iron reductase following sparge of reaction mixtures, whereas the reductase for ferric citrate was not appreciably affected by oxygen. Siderophores are produced by microorga- nisms for the purposes of binding extracellular iron and mediating the transport of Fe(EI). However, the iron must also be released from complex with the siderophore to allow synthesis of iron-containing celllar constituents. An es- terase enzyme has been described from Esche- richia coli which hydrolyzes the cyclic trimer of 2,3-dihydroxybenzoylserine (enterochelin) and has a theoretical function in iron release, since mutants lacking the esterase activity accumulate ferrienterochelin, but cannot grow unless sup- plied with an alternative source of iron (11, 16). Another mechanism of iron release was demon- strated in Mycobacterium smegmatis for iron complexed with mycobactin, a siderophore pro- duced by mycobacteria (1). Ferrimycobactin re- ductase catalyzed the reduction of Fe(III) and release from the siderophore of Fe(ll), and was assayed by using a chelator that formed a col- ored complex with Fe(ll). The enzyme was found in the soluble portion of cell extracts and utilized electrons from both NADH and NADPH. Another enzyme from Paracoccus de- nitrificans used electrons from NADH or NADPH to reduce Fe(III) from a catechol sid- erophore produced by this bacterium (18). En- zymes from Neurospora crassa and Aspergillus fwnigatus in the soluble portions of cell extracts were described as reducing iron from complex with various hydroxamate siderophores (6). E. coli is capable of utilizing ferrc citrate as well as ferrienterochelin in the transport of iron (8). Iron reductase activity for ferric citrate has also been reported in the membrane fractions of E. coli as well as Spirillum itersonii and Staph- ylococcus aureus (3). The enzyme from S. iter- sonii reduced iron using electrons from NADH or succinate via reactions of the respiratory chain. The enzyme found in S. aureus used electrons from NADH or L-glycerol 3-phosphate to reduce iron from ferric citrate (12). The activ- ities of both enzymes were reversibly inhibited by oxygen, which drained the electrons used anaerobically by the iron reductase from the respiratory chain. The alternative electron ac- ceptor, nitrate, also inhibited iron reductase ac- tivity in membrane preparations from cells of S. aureus induced for nitrate respiration (12). Pyochelin is a siderophore which can be iso- lated from culture media of Pseudomonas aeruginosa (2). This is probably the same com- pound studied by Liu and Shokrani and called by them pyochelin A (13). Both ferripyochelin and ferric citrate are used as substrates for iron transport in P. aeruginosa (manuscript submit- ted for publication). The present paper describes iron reductase activities which reduce iron from complexes with pyochelin and citrate. The activ- ities appear to reside on separate proteins and will be referred to as ferripyochelin iron reduc- tase and ferric citrate iron reductase. METHODS AND MATERIALS Bacteria and culture conditions. P. aeruginsa, strain PAO-1 (ATCC 15692), was obtained from the American Type Culture Collection and was main- tained on slants of brain heart infusion agar with monthly transfers. Bacteria were grown in succinate minimal medium containing 20 mM sodium succinate, 199 on December 26, 2018 by guest http://jb.asm.org/ Downloaded from
Transcript of Iron Reductases from Pseudomonas aeruginosa - Journal of

JOURNAL OF BACTERIOLOGY, Jan. 1980, p. 199-2040021-9193/80/01-0199/06$02.00/0
Vol. 141, No. 1
Iron Reductases from Pseudomonas aeruginosaCHARLES D. COX
Department ofMicrobiology, University of Iowa, Iowa City, Iowa 52242
Cell-free extracts ofPseudomonas aeruginosa contain enzyme activities whichreduce Fe(III) to Fe(II) when iron is provided in certain chelates, but not whenthe iron is uncomplexed. Iron reductase activities for two substrates, ferripy-ochelin and ferric citrate, appear to be separate enzymes because of differences inheat stabilities, in locations in fractions of cell-free extracts, in reductant specific-ity, and in apparent sizes during gel filtration chromatography. Ferric citrate ironreductase is an extremely labile activity found in the cytoplasmic fraction, andferripyochelin iron reductase is a more stable activity found in the periplasmic aswell as cytoplasmic fraction of extracts. A small amount of activity detectable inthe membrane fraction seemed to be loosely associated with the membranes.Although both enzymes have highest activity with reduced nicotinamide adeninedinucleotide, reduced glutathione also worked with ferripyochelin iron reductase.In addition, oxygen caused an irreversible loss ofa percentage ofthe ferripyocheliniron reductase following sparge of reaction mixtures, whereas the reductase forferric citrate was not appreciably affected by oxygen.
Siderophores are produced by microorga-nisms for the purposes of binding extracellulariron and mediating the transport of Fe(EI).However, the iron must also be released fromcomplex with the siderophore to allow synthesisof iron-containing celllar constituents. An es-terase enzyme has been described from Esche-richia coli which hydrolyzes the cyclic trimer of2,3-dihydroxybenzoylserine (enterochelin) andhas a theoretical function in iron release, sincemutants lacking the esterase activity accumulateferrienterochelin, but cannot grow unless sup-plied with an alternative source of iron (11, 16).Another mechanism of iron release was demon-strated in Mycobacterium smegmatis for ironcomplexed with mycobactin, a siderophore pro-duced by mycobacteria (1). Ferrimycobactin re-ductase catalyzed the reduction of Fe(III) andrelease from the siderophore of Fe(ll), and wasassayed by using a chelator that formed a col-ored complex with Fe(ll). The enzyme wasfound in the soluble portion of cell extracts andutilized electrons from both NADH andNADPH. Another enzyme from Paracoccus de-nitrificans used electrons from NADH orNADPH to reduce Fe(III) from a catechol sid-erophore produced by this bacterium (18). En-zymes from Neurospora crassa and Aspergillusfwnigatus in the soluble portions of cell extractswere described as reducing iron from complexwith various hydroxamate siderophores (6).
E. coli is capable of utilizing ferrc citrate aswell as ferrienterochelin in the transport of iron(8). Iron reductase activity for ferric citrate hasalso been reported in the membrane fractions of
E. coli as well as Spirillum itersonii and Staph-ylococcus aureus (3). The enzyme from S. iter-sonii reduced iron using electrons from NADHor succinate via reactions of the respiratorychain. The enzyme found in S. aureus usedelectrons from NADH or L-glycerol 3-phosphateto reduce iron from ferric citrate (12). The activ-ities of both enzymes were reversibly inhibitedby oxygen, which drained the electrons usedanaerobically by the iron reductase from therespiratory chain. The alternative electron ac-ceptor, nitrate, also inhibited iron reductase ac-tivity in membrane preparations from cells of S.aureus induced for nitrate respiration (12).
Pyochelin is a siderophore which can be iso-lated from culture media of Pseudomonasaeruginosa (2). This is probably the same com-pound studied by Liu and Shokrani and calledby them pyochelin A (13). Both ferripyochelinand ferric citrate are used as substrates for irontransport in P. aeruginosa (manuscript submit-ted for publication). The present paper describesiron reductase activities which reduce iron fromcomplexes with pyochelin and citrate. The activ-ities appear to reside on separate proteins andwill be referred to as ferripyochelin iron reduc-tase and ferric citrate iron reductase.
METHODS AND MATERIALSBacteria and culture conditions. P. aeruginsa,
strain PAO-1 (ATCC 15692), was obtained from theAmerican Type Culture Collection and was main-tained on slants of brain heart infusion agar withmonthly transfers. Bacteria were grown in succinateminimal medium containing 20mM sodium succinate,
199
on Decem
ber 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

200 Cox
4 mM NH4Cl, 0.4 mM MgSO4, 1.0 mM K2SO4, 1 JMZnCl2, 1 uM MnCl2, 1 ,uM CaSO4, and 4mM potassiumphosphate buffer (pH 7.4). Some studies involved cellsgrown in 0.5% Casamino Acids containing 0.4 mMMgSO4. The media were autoclaved without phos-phate buffer or minerals, which, along with FeCl3 andferric citrate, were made bacteria free by passagethrough a membrane filter of 0.45-,um pore size (Mil-lipore) and added to media before inoculation. Cellswere harvested in late logarithmic phase of growth bycentrifugation at room temperature. The cells werewashed in 0.01 M Tris-hydrochloride (pH 7.6) with 1mM MgCl2 (TM buffer).
Preparation ofextracts. Suspensions of cells weretreated for release of periplasmic contents by themethod of Stinson et al. (17). Washed cells were sus-pended (14 mg [dry weight] of cels per ml) in 0.2 MMgC12 with 0.05 M Tris-hydrochloride buffer at pH8.5, and the mixture was stirred for 1 h at 240C. Aftercentrifugation at 5,900 x g, the supernatant fluid wassaved as "shock fluid," and the cell pellet was sus-pended in distilled water and stirred for 1 h at 240C.This suspension was centrifuged at 5,900 x g, and thesupernatant was combined with the first. The com-bined shock fluids were concentrated with an AmiconPM-10 membrane and dialyzed against Tris-hydro-chloride (0.01 M, pH 7.6) for 20 h at 40C. Viablebacteria were determined before and after the shockprocedure by plating dilutions on 1% tryptic soy agar.The cell pellet resulting from the preparation of the
shock fluid was suspended in TM buffer, and the cellswere ruptured at 16,000 lb/in2 with a French pressurecell (Aminco). The ruptured cell suspensions werecentrifuged at 20,000 x g at 40C to remove whole cellsand cell debris. The resulting supernatants were cen-trifuged at 130,000 x g for 2 h to obtain fractions richin membrane constituents (pellet) and in soluble orcytoplasmic constituents (supernatant). The pelletswere suspended in minimal volumes of 0.01 M Tris-hydrochloride, and both fractions were dialyzedagainst the same buffer. The fractions were made cellfree by pressurized filtrations through 0.45-Lm poresize filters.
Preparations of substrates. Ferric citrate wasdissolved in distilled water and made to a final con-centration of 10 mg/ml at a pH of 7.5. Ferric chloridewas made 0.1 M in distilled water and allowed toequilibrate so that after 1 day there was no detectableFe(ll).
Pyochelin (siderophore without iron) was extractedfrom culture media into ethyl acetate (2), concentratedby rotary evaporation, and made into the iron chelate(ferripyochelin) by adding FeCI3 until the solutionchanged to wine red and then to brown. Ferripyochelinwas extracted with multiple additions of ethyl acetate,leaving a brown-colored water layer. Unchelated ironwas removed by concentrating ferripyochelin in ethylacetate and extracting the organic solvent phase with0.01 M HCI. Ferripyochelin was precipitated by addingheptane to the ethyl acetate solution in a 3:2 ratio.The dried precipitate was dissolved in ethanol andpurified by preparative thin-layer chromatography onsilicic acid (Adsorbosil-5, Applied Sciences Laborato-ries) in chloroform-acetic acid-ethanol (90:5:5). Thered band at Rf 0.22 was scraped and eluted from silicic
J. BACTERIOL.
acid in methanol. Ferripyochelin was dried, dissolvedin ethyl acetate, and precipitated and washed withheptane. The precipitate was dried under vacuum for6 h, weighed, and stored at 40C.
Iron concentrations in ferric citrate and ferripy-ochelin solutions were determined by an assay fornonheme iron (5). The concentration of iron in 10 mgof ferric citrate per ml was 28.6 mM, and that in 10 mgof ferripyochelin per ml was 14.6 mM. The molecularweight of ferripyochelin was determined from titra-tions of pyochelin with iron to be approximately 700.Using this value to determine the molarity of ferripy-ochelin yielded 14.2 mM in a solution containing 10mg/ml. This molecular weight estimate has also beenconfirmed by mass spectroscopy (data not shown).Enzyme assays. Iron reductase activity was meas-
ured by the method of Dailey and Lascelles (3). Ironchelate was added first, followed by the ferrozine re-agent in 0.01 M Tris buffer. Spontaneous reduction ofiron was measured, and the cell extract was added in20- to 100-1il volumes. Controls were conducted withvaried sequences of reaction constituents. Assays forferric citrate iron reductase yielded maximal activitieswhen reductant was added before ferric citrate. Thereactions at 240C were monitored at 562 nm on a 0.1absorbance scale in a model 124 Perkin-Elmer double-beam spectrophotometer. Reference cuvettes con-tained all of the reaction components except reduc-tant. Nonenzymatic reduction of iron was the slowestin polystyrene cuvettes, but glass Thunberg cuvetteswere used in assays to determine effects of oxygen onenzymatic activity. Assays involving large numbers ofdeterminations were conducted by measuring differ-ences in absorbance at times of incubation at 240C ina Gilford model 240 spectrophotometer with continualreference to reactions containing no extract and reac-tions containing no reductant.A variety of enzyme assays were used in the deter-
mination of the location of iron reductase activities incell fractions. Diaphorase activities measured in theseextracts were equivalent in rates with NADH and withNADPH and appear similar to the DT diaphorasedescribed by Ernster (7). Sensitive assays were con-ducted by fluorometric determination of resazurin re-duction in dialyzed extracts as described by Guilbaultand Kramer (9). The activities of diaphorase are ex-pressed in nanomoles of resorufin accumulating perminute per milligram of protein, using NADH as elec-tron donor. Malate dehydrogenase (EC 1.1.1.37) wasused as another marker of the soluble fraction of cellextracts and was measured by fluorometric determi-nation ofNADH accumulation using the assay condi-tions of Yoshida (21). Succinate dehydrogenase (EC1.3.99.1) was used as an indicator of the presence ofmembrane-bound enzymes and was determined by thespectrophotometric method of Veeger et al. (19). Fer-ricyanide was measured at 420 nm, and an extinctioncoefficient of 1.03 x 103 M` cm-' was used to measureactivity. The assay conditions for glucose dehydrogen-ase (EC 1.1.99.10) in particulate fractions of pseudo-monads by Hauge (10) measured dichlorophenolin-dophenol reduction. Alkaline phosphatase has beenfound in the periplasm of P. aeruginosa (4) and wasassayed using the conditions employed by Malamyand Horecker (15) with the exception that 1 M Tris
on Decem
ber 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 141, 1980
buffer (pH 8.0) with 1 mM MgCl2 was used.Protein was determined according to the method of
Lowry et al. (14) in particulate and soluble fractionsand by the Coomassie blue assay (BioRad) in solublefractions. Reduced cofactors, glutathione, and ferro-zine were obtained from Sigma.
Measurements of enzyme stability were made byadding extract to buffer and ferrozine reagent pre-heated to 60°C and incubating the mixtures for specifictimes. At the end ofthe incubation times, the solutionswere rapidly cooled in an ice bath and equilibrated to24°C in a water bath. Substrate and reductant wereadded, and the reactions were measured spectropho-tometrically.
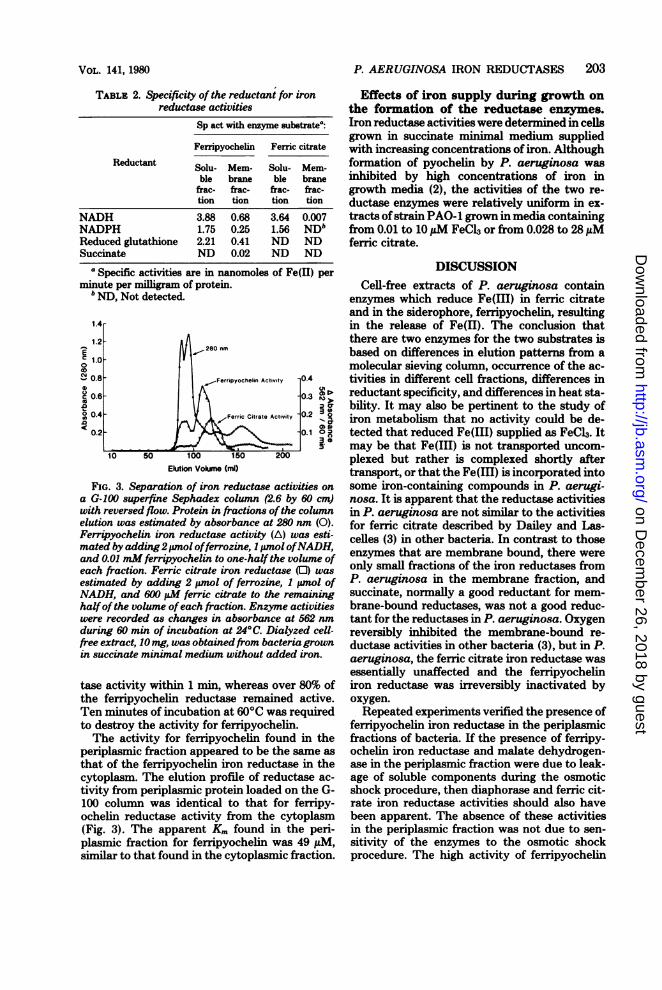
Chromatography of reductase activities was con-ducted at 4°C on a G-100 Sephadex superfine column(2.6 by 60 cm) equilibrated with 0.05 M Tris-hydro-chloride (pH 7.6) with 0.1 M KCl. Flow was againstgravity at a rate of 1.5 ml per 10 min, and DextranBlue 2000 was used to mark the void volume elution.Column fractions were measured for absorbance at280 nm, and then the volumes in the tubes were splitequally and separate assay mixtures were added to thetubes: ferric citrate to one set and ferripyochelin tothe other. Each set of tubes was measured for changesin absorbance at 562 nm between 10 and 70 min ofincubation. The differences in these readings weretaken as estimations of the respective activities of thetwo reductase enzymes in the fractions.
RESULTSAssay ofiron reductases. Several iron-con-
taining compounds were used in iron reductaseassays. The insoluble nature of Fe(III) at pH 7.4imposed limitations on the concentrations ofunchelated iron that could be used in assays, butno reduction was detected when 0.2 ,umol ofFeCl3 per ml was incubated with NADH andcell-free extract of P. aeruginosa (curve a, Fig.1). Utilization of iron from ferripyochelin (2)indicated the presence of a mechanism for re-lease of the iron from this complex, and therewas reduction of the Fe(III) and release of Fe(ll)from ferripyochelin when mixed with fresh, di-alyzed extracts (curve b, Fig. 1). No reductionwas apparent without added NADH (curve c,Fig. 1). Anaerobic conditions were not requiredfor maximal activity of this enzyme, but a 5-ssparge with oxygen (arrow, curve b, Fig. 1)caused a decrease in specific activity from 3.54to 2.83 nmol/min per mg of protein. This effectwas reproducible, with an average inhibitoryeffect of 25.1%, and was irreversible.
Fe(III) from ferric citrate was also not reducedin cell-free extracts in the absence of reductant(curve a, Fig. 2), but there was reduction afteraddition of NADH (first arrow, curve b, Fig. 2).Anaerobic conditions were not required for max-imal activity, and a 5-s sparge with oxygencaused an apparent increase in activity. How-ever, repeated analyses indicated that there was
P. AERUGINOSA IRON REDUCTASES 201
Tin (mi)
FIG. 1. Tracing of a ferripyochelin iron reductasereaction conducted with a dialyzed cell-free extractfrom P. aeruginosa. The reaction mixture contained58.3 pg ofprotein per ml with (a) 0.2mM FeCl3 and Irnol of NADH; (b) 0.31 mM ferripyochelin and Ipmol ofNADH; or (c) 0.31 mMferripyochelin lackingNADH. All reactions were initiated with nitrogen-flushed buffer in Thunberg cuvettes containing 2.4 mlcomposed of 100 ltmol of Tris-hydrochloride (pH 7.4)and 2 unol offerrozine. NADH was added to assays(curves a and b) at the first arrow, and a 5-s oxygensparge was conducted at the second arrow on curveb. Cell-free extract was obtained from bacteria grownin succinate minimal medium without added iron.
.061
.05
9CY .04
* .03
i .02.0
.01
02
NADH
1k- b
O 2 4 6 8 10 12 14Thw (mb)
FIG. 2. Tracing of a ferric citrate iron reductasereaction conducted with a dialyzed cell-free extractofP. aeruginosa. The reaction mixture contained 58.3pg ofprotein per ml with (a) 600 M ferric citrate andno NADH or (b) 600 pM ferric citrate with 1 tumol ofNADH. All reactions were initiated in nitrogen-flushed buffer in Thunberg cuvettes containing 2.4 mlcomposed of 100 jmol of Tri-hydrochloride (pH 7.4)and 2pmol offerrozine. NADHwas added at the firstarrow, and a 5-s oxygen sparge was conducted at thesecond arrow. Cell-free extract was obtained frombacteria grown in succinate minimal medium withoutadded iron.
no detectable effect with oxygen, and the appar-ent increase in activity was an event occurringwith or without oxygen over the subsequentminutes of assay. The aeration was strenuous inboth Fig. 1 and Fig. 2, and the period of time forthe return to steady rates was apparently due tobubbles leaving solution. Both ferric citrate and
on Decem
ber 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

202 COx
ferripyochelin iron reductases were affected byoxygen in ways that were different from thosereported for ferric citrate reductases from otherbacteria (3).Locations of activities. There was no loss
of viability in populations of bacteria subjectedto osmotic shock for the release of componentsof the periplasm. Alkaline phosphatase activitywas present in highest activity in the periplasmicfraction (Table 1). The absence of diaphoraseactivity from this fraction indicated that therewas no contamination of periplasm with cyto-plasmic components. Malic dehydrogenase wasfound in highest activity in the cytoplasmic frac-tion, but there was also activity in the otherfractions. Glucose and succinate dehydrogenaseactivities were used as markers for the mem-brane-rich fraction, and although high activitieswere found in this fraction, approximately 10%of both activities were in the soluble fraction.
Ferripyochelin iron reductase was present inthe periplasmic fraction where negligible ferriccitrate iron reductase was detected. Both reduc-tase activities were in high levels in the cyto-plasmic fraction. The presence of both activitiesin low amounts in the membrane fraction wasfurther studied by washing the membranes inbuffer and again collecting the membrane frac-tions by centrifugation. Both activities appearedagain in the supernatants (soluble fractions),indicating a possible loose attachment to themembranes. The activities of the enzymes inmembrane and soluble fractions were identical;the apparent Km values were 56 ttM for ferripy-ochelin and 303 ,uM for ferric citrate.
Specificity of the reductant. Table 2 dis-plays the abilities of various reduced compoundsand succinate to serve as electron donors for thetwo iron reductase activities in soluble and mem-brane fractions. NADH was routinely the bestreductant; NADPH worked approxinately 30 to
40% as well. Reduced glutathione was also veryeffective in donating electrons to the ferripy-ochelin iron reductase, but not to ferric citrateiron reductase. The periplasmic activity reduc-ing iron from ferripyochelin was identical inspecificity for reductant to the soluble activity.Many other amino acids, carbohydrates, andcarboxylic acids were tested and demonstratedno electron-donating function for the reductases.Other reducing agents (ascorbate and dithio-threitol) reduced iron from the complexes as
rapidly with extract as without extract.Separate activities of ferric citrate and
ferripyochelin reductases. The apparent dif-ferences in location of the ferripyochelin andferric citrate reductase activities suggested theexistence of separate enzymes and not one en-
zyme with multiple substrates. There was a
small but reproducible difference in the elutionprofile of the two activities from a G-100 Seph-adex superfine column. Effluent fractions result-ing from crude, dialyzed extract applied to thecolumn were measured for absorbance at 280nm (solid line, Fig. 3), and then the fractionswere split and reagents were added to the twosets oftubes for reductase assays for ferric citrateand ferripyochelin. Both activities appeared inlater fractions containing proteins ofsmaller size,and there was a significant amount of activityfor ferripyochelin eluting in the void volume,possibly with particulate material or as an ag-
gregate of activity eluting later.The separate nature of the two enzymes was
also suggested by differences in stabilities of thetwo activities. Optimal activity of ferric citrateiron reductase was found only in fresh extracts,whereas the activity of ferripyochelin iron re-
ductase could be measured over a period ofapproximately 1 week before there was signifi-cant decay in activity. Heating cell-free extractat 60°C destroyed the ferric citrate iron reduc-
TABLE 1. Specific activities ofmarker enzymes and iron reductase enzymes in different cell fractionsSp act of enzymes
Cell fraction Alkalinephos- Diah Malate de- e- Glucose dehy- Ferripy- Ferric cit-
phatase' raseb hydrogen- hydrogenased drogenase' ironuc rate ironawe~~ ~ ~ ~ ~ ion u-reductase
Soluble 5.41 x 10-3 3.19 28.35 8.0 x 10-3 1.82 x 10-4 3.88 3.64Membrane ND' 0.63 16.14 8.6 x 10-2 3.57 x 10-3 0.68 0.28Periplasm 4.77 x 10-2 ND 9.20 ND ND 2.29 0.08
aUnits per milligram of protein (1 unit - 1 micromole ofp-nitrophenol formed per hour).b Nanomoles of resorufin formed per minute per milligram of protein.Nanomoles of,NADH formed per minute per milligram of protein.
d Units per milligram of protein (1 unit = 1 micromole of succinate oxidized per minute).' Units per milligram of protein (1 unit = 1 micromole of glucose oxidized per minute).f Nanomoles of Fe(II) formed per minute per mlliram of protein.' ND, Not detected.
J. BACTERIOL.
on Decem
ber 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
VOL. 141, 1980
TABLE 2. Specificity of the reductant for ironreductase activities
Sp act with enzyme substrate':
Feripyochelin Feric citrate
Reductant Solu- Mem- Solu- Mem-
ble brane ble branefrac- frac- frac- frac-tion tion tion tion
NADH 3.88 0.68 3.64 0.007NADPH 1.75 0.25 1.56 NDbReduced glutathione 2.21 0.41 ND NDSuccinate ND 0.02 ND ND
'Specific activities are in nanomoles of Fe(ll) perminute per milligram of protein.
b ND, Not detected.
1.4r
.--280 nm
| Ferripyochelin Activity 0.4
-0.3
Ferric Citrate Activity -0.2
0.1 0 z
S. (
10 50 100 150 200Elution Volume (ml)
FIG. 3. Separation of iron reductase activities ona G-100 superfine Sephadex column (2.6 by 60 cm)with reversed flow. Protein in fractions ofthe columnelution was estimated by absorbance at 280 nm (0).Ferripyochelin iron reductase activity (A) was esti-mated by adding2,pmol offerrozine, 1 pmol ofNADH,and 0.01 mM ferripyochelin to one-half the volume ofeach fraction. Ferric citrate iron reductase (0) wasestimated by adding 2 pmol of ferrozine, I pnol ofNADH, and 600 EM ferric citrate to the remaininghalfof the volume ofeach fraction. Enzyme activitieswere recorded as changes in absorbance at 562 nmduring 60 min of incubation at 24°C. Dialyzed cell-free extract, 10 mg, was obtainedfrom bacteria grownin succinate minimal medium without added iron.
tase activity within 1 min, whereas over 80% ofthe ferripyochelin reductase remained active.Ten minutes of incubation at 60°C was requiredto destroy the activity for ferripyochelin.The activity for ferripyochelin found in the
periplasmic fraction appeared to be the same as
that of the ferripyochelin iron reductase in thecytoplasm. The elution profile of reductase ac-
tivity from periplasmic protein loaded on the G-100 column was identical to that for ferripy-ochelin reductase activity from the cytoplasm(Fig. 3). The apparent Km found in the peri-plasmic fraction for ferripyochelin was 49 MuM,similar to that found in the cytoplasmic fraction.
P. AERUGINOSA IRON REDUCTASES 203
Effects of iron supply during growth onthe formation of the reductase enzymes.Iron reductase activities were determined in celisgrown in succinate minimal medium suppliedwith increasing concentrations of iron. Althoughformation of pyochelin by P. aeruginosa wasinhibited by high concentrations of iron ingrowth media (2), the activities of the two re-ductase enzymes were relatively uniform in ex-tracts ofstrain PAO-1 grown in media containingfrom 0.01 to 10 ,M FeCl3 or from 0.028 to 28,uMferric citrate.
DISCUSSIONCell-free extracts of P. aeruginosa contain
enzymes which reduce Fe(III) in ferric citrateand in the siderophore, ferripyochelin, resultingin the release of Fe(II). The conclusion thatthere are two enzymes for the two substrates isbased on differences in elution patterns from amolecular sieving column, occurrence of the ac-tivities in different cell fractions, differences inreductant specificity, and differences in heat sta-bility. It may also be pertinent to the study ofiron metabolism that no activity could be de-tected that reduced Fe(III) supplied as FeCl3. Itmay be that Fe(III) is not transported uncom-plexed but rather is complexed shortly aftertransport, or that the Fe(Ill) is incorporated intosome iron-containing compounds in P. aerugi-nosa. It is apparent that the reductase activitiesin P. aeruginosa are not similar to the activitiesfor ferric citrate described by Dailey and Las-celles (3) in other bacteria. In contrast to thoseenzymes that are membrane bound, there wereonly small fractions of the iron reductases fromP. aeruginosa in the membrane fraction, andsuccinate, normally a good reductant for mem-brane-bound reductases, was not a good reduc-tant for the reductases in P. aeruginosa. Oxygenreversibly inhibited the membrane-bound re-ductase activities in other bacteria (3), but in P.aeruginosa, the ferric citrate iron reductase wasessentially unaffected and the ferripyocheliniron reductase was irreversibly inactivated byoxygen.Repeated experiments verified the presence of
ferripyochelin iron reductase in the periplasmicfractions of bacteria. If the presence of ferripy-ochelin iron reductase and malate dehydrogen-ase in the periplasmic fraction were due to leak-age of soluble components during the osmoticshock procedure, then diaphorase and ferric cit-rate iron reductase activities should also havebeen apparent. The absence of these activitiesin the periplasmic fraction was not due to sen-sitivity of the enzymes to the osmotic shockprocedure. The high activity of ferripyochelin
1.2
O 1.00
SM 0.8aC 0.6
o 04< 0.2
on Decem
ber 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

J. BACTERIOL.
iron reductase in the periplasm raises specula-tion about the release of reduced iron from thesiderophore prior to membrane transport andthe subsequent transport of Fe(II). It is unclearhow the periplasmic enzyme could interact withreductant, but a fraction of the malate dehydro-genase activity was also found in the periplasmicfraction, and there may be NADH-generatingmechanisms available for this reductase.The survey of possible reductants for the sol-
uble ferripyochelin iron reductase revealed thatNADH was most effective, but that reducedglutathione also served as an effective electrondonor. The physiological importance of this re-ductant to enzyme activity is unknown, but flu-orometric examination of NADH oxidation inextracts with reduced glutathione and flavin ad-enine dinucleotide revealed glutathione reduc-tase activity in the soluble fraction of extracts ofP. aeruginosa.The lack of control of the reductase activities
by iron supplied in high concentrations ingrowth media was an unexpected finding. P.aeruginosa has a high requirement for iron (20),and a relatively high concentration of iron (10suM FeCl3) was found necessary to inhibit for-mation of pyochelin (2). Ferric chloride ingrowth media at pH 7.4 may not constitute areadily available source of iron, and even at highconcentrations of FeC13 bacteria must transportand incorporate extracellular Fe(III). Althoughthis may explain the lack of iron control on thesynthesis of ferripyochelin iron reductase, lackof control by ferric citrate, a soluble source ofiron, suggests alternatively that reductase en-zymes may not be affected by extracellular ironconcentrations.Methods are being investigated for the puri-
fication of large amounts of reductase enzymesto answer remaining questions. However, futureresearch must first solve the problem of stabilityof both enzymes, because activities survive onlya few weeks in frozen extracts.
ACKNOWLEDGMENTSThis investigation was supported by Public Health Service
grant AI-13120 from the National Institute of Allergy andInfectious Diseases.
LITERATURE CITED1. Brown, K. A., and C. Ratledge. 1975. Iron transport in
Mycobacterium smegmatis: ferrimycobactin reductase(NAD(P)H-ferrimycobactin oxidoreductase), the en-
zyme releasing iron from its carrier. FEBS Lett. 53:262-266.
2. Cox, C. D., and RI Graham. 1979. Isolation of an iron-binding compound from Pseudomonas aeruginosa. J.Bacteriol. 137:357-364.
3. Dailey, H. A., and J. Lascelles. 1977. Reduction of ironand synthesis of protoheme by Spirillum itersonii andother organisms. J. Bacteriol. 129:815-820.
4. Day, D. F., and J. M. Ingram. 1973. Purification andcharacterization of Pseudomonas aeruginosa alkalinephosphatase. Can. J. Bacteriol. 19:1225-1233.
5. Doeg, K. A., and D. M. Ziegler. 1962. Simplified meth-ods for the estimation of iron in mitochondria andsubmitochondrial fractions. Arch. Biochem. Biophys.97:37-40.
6. Ernst, J. F., and G. Winkelmann. 1977. Enzymaticrelease of iron from sideramines in fungi, NADH: sid-eramine oxidoreductase in Neurospora crassa.Biochim. Biophys. Acta 500:27-41.
7. Ernster, L 1967. DT diaphorase. Methods Enzymol. 10:309-317.
8. Frost, G., and J. Rosenberg. 1973. The inducible citrate-dependent iron transport system in Escherichia coliK12. Biochim. Biophys. Acta 330:90-101.
9. Guilbault, G. G., and D. N. Kramer. 1965. Fluorometricprocedure for measuring the activity of dehydrogenase.Anal. Chem. 37:1219-1221.
10. Hauge, J. G. 1966. Glucose dehydrogenases-particulate.Methods Enzymol. 9:92-98.
11. Langman, L., I. G. Young, G. E. Frost, H. Rosenberg,and F. Gibson. 1972. Enterochelin system for irontransport in Escherichia coli: mutations affecting fenicenterochelin esterase. J. Bacteriol. 112:1142-1149.
12. Lascelles, J., and K. A. Burke. 1978. Reduction of ferriciron by L-lactate and DL-glycerol-3-phosphate in mem-brane preparations from Staphylococcus aureus andinteractions with the nitrate reductase system. J. Bac-teriol. 134:585-589.
13. Liu, P. V., and F. Shokrani. 1978. Biological activitiesof pyochelins: iron-chelating agents of Pseudomonasaeruginosa. Infect. Immun. 22:878-90.
14. Lowry, O.IL, N. J. Rosebrough, A. L Farr, and R J.Randall. 1951. Protein measurement with the Folinphenol reagent. J. Biol. Chem. 193:265-275.
15. Malamy, M., and B. L. Horecker. 1966. Alkaline phos-phatase. Methods Enzymol. 9:639-642.
16. O'Brien, L. G., and F. Gibson. 1971. Enterochelin hy-drolysis and iron metabolism in Escherichia coli.Biochim. Biophys. Acta 237:537-549.
17. Stinson, M. W., M. A. Cohen, and J. M. Merrick. 1976.Isolation of dicarboxylic acid- and glucose-binding pro-teins from Pseudomonas aeruginosa. J. Bacteriol. 128:573-579.
18. Tait, G. H. 1975. The identification and biosynthesis ofsiderochromes formed by Micrococcus dentrificans.Biochem. J. 146:191-204.
19. Veeger, C., D. V. DerVartanian, and W. P. Zeyle-maker. 1969. Succinate dehydrogenase. Methods En-zymol. 13:81-90.
20. Waring, N. S., and C. H. Werkman. 1942. Iron require-ments of heterotrophic bacteria. Arch. Biochem. 1:425-433.
21. Yoshida, A. 1969. L-Malate dehydrogenase from BaciUussubtilis. Methods Enzymol. 13:141-145.
204 Cox
on Decem
ber 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from


















