
Investigations into Seed Dormancy in Grevillea - Annals of Botany
16
Investigations into Seed Dormancy in Grevillea linearifolia, G. buxifolia and G. sericea: Anatomy and Histochemistry of the Seed Coat C. L. BRIGGS 1 , E. C. MORRIS 1, * and A. E. ASHFORD 2 1 Ecology Research Group, University of Western Sydney, Locked Bag 1797, Penrith South DC, NSW 1797, Australia and 2 School of Biological, Earth and Environmental Sciences, University of New South Wales, Sydney, NSW 2052, Australia Received: 29 March 2005 Returned for revision: 23 May 2005 Accepted: 5 July 2005 Published electronically: 12 September 2005 Background and Aims Seeds of east Australian Grevillea species generally recruit post-fire; previous work showed that the seed coat was the controller of dormancy in Grevillea linearifolia. Former studies on seed development in Grevillea have concentrated on embryology, with little information that would allow testing of hypotheses about the breaking of dormancy by fire-related cues. Our aim was to investigate structural and chemical characteristics of the seed coat that may be related to dormancy for three Grevillea species. Methods Seeds of Grevillea linearifolia, Grevillea buxifolia and Grevillea sericea were investigated using gross dissection, thin sectioning and histochemical staining. Water movement across the seed coat was tested for by determining the water content of embryos from imbibed and dry seeds of G. sericea. Penetration of intact seeds by Lucifer Yellow was used to test for internal barriers to diffusion of high-molecular-weight compounds. Key Results Two integuments were present in the seed coat: an outer testa, with exo-, meso- and endotestal (palisade) layers, and an inner tegmen of unlignified sclerenchyma. A hypostase at the chalazal end was a region of structural difference in the seed coat, and differed slightly among the three species. An internal cuticle was found on each side of the sclerenchyma layer. The embryos of imbibed seeds had a water content six times that of dry seeds. Barriers to diffusion of Lucifer Yellow existed at the exotestal and the endotestal/hypostase layers. Conclusions Several potential mechanisms of seed coat dormancy were identified. The embryo appeared to be completely surrounded by outer and inner barriers to diffusion of high-molecular-weight compounds. Phenolic compounds present in the exotesta could interfere with gas exchange. The sclerenchyma layer, together with strengthening in the endotestal and exotestal cells, could act as a mechanical constraint. Key words: Seed coat structure, Grevillea linearifolia, Grevillea buxifolia, Grevillea sericea, histochemistry, seed dormancy. INTRODUCTION All Grevillea species from south-east Australia produce a hard, dry seed that is shed from the parent plant after dehis- cence of the mature fruit (Olde, 1997; Auld and Denham, 1999). Seeds from most of these species possess an oil- filled body, the elaiosome (located at the chalazal end of the seed), that is attractive to many ant species and results in the removal of the seed into the nest of large ant species (Auld and Denham, 1999). Hence, many Grevillea seeds are incorporated into the seed bank where they lie dormant until recruitment in an immediate post-fire period (Auld and Tozer, 1995). Laboratory studies of the germination characteristics of these species have shown that fire-related signals such as smoke or heat shock stimulate germination (Edwards and Whelan 1995; Kenny, 2000; Morris, 2000). In addition, scarification of the seed coat increases germina- tion in some Grevillea species (G. barkylana; Edwards and Whelan, 1995), sometimes in combination with heat shock (G. buxifolia) or smoke (G. diffusa, G. mucronulata, G. speciosa; Morris, 2000). Stimulation of germination by treatments such as heat shock and scarification suggests that the seed coat plays a role in dormancy; this hypothesis was tested for G. linearifolia, a species that shows increased germination in response to smoke and heat shock (Morris, 2000). Removal of the seed coat in G. linearifolia results in ger- mination of all embryos, establishing the seed coat as the primary controller of dormancy in this species (Morris et al., 2000). Seed coat dormancy is a widespread phenomenon, and there are several mechanisms by which the seed coat can impose dormancy (Bewley and Black, 1994). The seed coat can (1) mechanically restrict germination of the embryo, (2) restrict the exit of germination inhibitors from the embryo, (3) contain germination inhibitors, (4) restrict water uptake and (5) restrict oxygen uptake. Evidence to date allows judgement to be made on some of these possible mechanisms for Grevillea species. The seed coat does not appear to restrict water uptake as unscarified seeds take up water as readily as do scarified seeds (Morris, 2000; Morris et al., 2000). Re-insertion of embryos into dissected seed coats does not inhibit germination of de-coated G. linearifolia (Morris et al., 2000), indicating that germination inhibitors are not released from the seed coat itself. In view of the importance of the seed coat in the dorm- ancy of these Grevillea species, an improved understanding of the structure of the seed coat is required before hypothe- ses about how germination treatments affect that structure can be tested (Egerton-Warburton, 1998). * For correspondence. E-mail [email protected] Annals of Botany 96: 965–980, 2005 doi:10.1093/aob/mci250, available online at www.aob.oxfordjournals.org Ó The Author 2005. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: [email protected] Downloaded from https://academic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022
Transcript of Investigations into Seed Dormancy in Grevillea - Annals of Botany

Investigations into Seed Dormancy in Grevillea linearifolia, G. buxifolia andG. sericea: Anatomy and Histochemistry of the Seed Coat
C. L. BRIGGS1, E. C. MORRIS1,* and A. E. ASHFORD2
1Ecology Research Group, University of Western Sydney, Locked Bag 1797, Penrith South DC, NSW 1797,
Australia and 2School of Biological, Earth and Environmental Sciences, University of New South Wales,
Sydney, NSW 2052, Australia
Received: 29 March 2005 Returned for revision: 23 May 2005 Accepted: 5 July 2005 Published electronically: 12 September 2005
� Background and Aims Seeds of east Australian Grevillea species generally recruit post-fire; previous work showedthat the seed coat was the controller of dormancy in Grevillea linearifolia. Former studies on seed development inGrevillea have concentrated on embryology, with little information that would allow testing of hypotheses about thebreaking of dormancy by fire-related cues. Our aim was to investigate structural and chemical characteristics of theseed coat that may be related to dormancy for three Grevillea species.� Methods Seeds of Grevillea linearifolia, Grevillea buxifolia and Grevillea sericea were investigated using grossdissection, thin sectioning and histochemical staining. Water movement across the seed coat was tested for bydetermining the water content of embryos from imbibed and dry seeds of G. sericea. Penetration of intact seeds byLucifer Yellow was used to test for internal barriers to diffusion of high-molecular-weight compounds.� Key Results Two integuments were present in the seed coat: an outer testa, with exo-, meso- and endotestal(palisade) layers, and an inner tegmen of unlignified sclerenchyma. A hypostase at the chalazal end was a region ofstructural difference in the seed coat, and differed slightly among the three species. An internal cuticle was found oneach side of the sclerenchyma layer. The embryos of imbibed seeds had a water content six times that of dry seeds.Barriers to diffusion of Lucifer Yellow existed at the exotestal and the endotestal/hypostase layers.� Conclusions Several potential mechanisms of seed coat dormancy were identified. The embryo appeared to becompletely surrounded by outer and inner barriers to diffusion of high-molecular-weight compounds. Phenoliccompounds present in the exotesta could interfere with gas exchange. The sclerenchyma layer, together withstrengthening in the endotestal and exotestal cells, could act as a mechanical constraint.
Key words: Seed coat structure, Grevillea linearifolia, Grevillea buxifolia, Grevillea sericea, histochemistry, seeddormancy.
INTRODUCTION
All Grevillea species from south-east Australia produce ahard, dry seed that is shed from the parent plant after dehis-cence of the mature fruit (Olde, 1997; Auld and Denham,1999). Seeds from most of these species possess an oil-filled body, the elaiosome (located at the chalazal end ofthe seed), that is attractive to many ant species and results inthe removal of the seed into the nest of large ant species(Auld and Denham, 1999). Hence, many Grevillea seedsare incorporated into the seed bank where they lie dormantuntil recruitment in an immediate post-fire period (Auldand Tozer, 1995). Laboratory studies of the germinationcharacteristics of these species have shown that fire-relatedsignals such as smoke or heat shock stimulate germination(Edwards and Whelan 1995; Kenny, 2000; Morris, 2000).In addition, scarification of the seed coat increases germina-tion in some Grevillea species (G. barkylana; Edwardsand Whelan, 1995), sometimes in combination with heatshock (G. buxifolia) or smoke (G. diffusa, G. mucronulata,G. speciosa; Morris, 2000).
Stimulation of germination by treatments such as heatshock and scarification suggests that the seed coat playsa role in dormancy; this hypothesis was tested forG. linearifolia, a species that shows increased germination
in response to smoke and heat shock (Morris, 2000).Removal of the seed coat in G. linearifolia results in ger-mination of all embryos, establishing the seed coat as theprimary controller of dormancy in this species (Morris et al.,2000).
Seed coat dormancy is a widespread phenomenon, andthere are several mechanisms by which the seed coat canimpose dormancy (Bewley and Black, 1994). The seed coatcan (1) mechanically restrict germination of the embryo,(2) restrict the exit of germination inhibitors from theembryo, (3) contain germination inhibitors, (4) restrictwater uptake and (5) restrict oxygen uptake. Evidence todate allows judgement to be made on some of these possiblemechanisms for Grevillea species. The seed coat does notappear to restrict water uptake as unscarified seeds takeup water as readily as do scarified seeds (Morris, 2000;Morris et al., 2000). Re-insertion of embryos into dissectedseed coats does not inhibit germination of de-coatedG. linearifolia (Morris et al., 2000), indicating thatgermination inhibitors are not released from the seed coatitself.
In view of the importance of the seed coat in the dorm-ancy of these Grevillea species, an improved understandingof the structure of the seed coat is required before hypothe-ses about how germination treatments affect that structurecan be tested (Egerton-Warburton, 1998).* For correspondence. E-mail [email protected]
Annals of Botany 96: 965–980, 2005
doi:10.1093/aob/mci250, available online at www.aob.oxfordjournals.org
� The Author 2005. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved.
For Permissions, please email: [email protected]
Dow
nloaded from https://academ
ic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022

Seed coat structure is described using specific terminol-ogies. The seed coat in all bitegmic species consists of atesta (formed from the outer integument of the ovule) and ategmen (formed from the inner integument of the ovule)with the outer epidermis of the testa called the exotesta andthe inner epidermis called the endotesta; similarly, the outerepidermis of the tegmen is called the exotegmen and innerepidermis the endotegmen. The cell layers between theexo- and endotesta are called the mesophyll, mesotesta ormesotegmen (Corner, 1976). The position of the mainmechanical layer within the seed coat determines whetherthe seed is further described as ‘testal’ or ‘tegmic’. In theProteaceae the seed coat is considered to be endotestal witha fibrous tegmen. The endotestal cells are thin-walled (withor without lignification) and strengthened by an internalfibrillar reticulum that fills the central cavity. The exo-and endotegmen both consist of narrow, cylindrical fibreswith the endotegmal fibres lying across the outer longitu-dinal fibres (Corner, 1976).
Investigations of seed structure in Grevillea species havebeen undertaken by Brough (1933), Kausik (1938, 1939)and Venkata Rao (1967). These studies focused on floraldevelopment, fertilization and development of the embryo,and the structure of the mature seed was treated very briefly;histochemical techniques were not used. G. robusta wasstudied by Brough (1933) and Kausik (1938), G. banksiiby Kausik (1939), and Venkata Rao (1967) covered a rangeof species.
Grevillea ovules are orthopterous, hemianatropous ornearly anatropous, bitegmic, crassinucellate, receive a singlevascular bundle and are attached laterally to the marginalplacenta (Kausik, 1938, 1939; Venkata Rao, 1967). Duringenlargement of the embryo sac the nucellar tissue maybecome completely destroyed, or it may persist as a thinlayer pressed against the surface of the inner integument(Kausik, 1938, 1939; Corner, 1976). In some species, thenucellus survives as a small group of intact cells at themicropylar and chalazal ends of the ovule. Prior to fertiliza-tion, the chalazal nucellar remnant divides to form a distinctregion called a ‘hypostase’ that becomes filled with tanninsduring seed maturation (Kausik, 1938, 1939; Venkata Rao,1967). In many species producing bitegmic ovules, thecuticles covering the inner epidermis of the outer integu-ment fuse with the cuticle covering the outer epidermis of theinner integument to form a single, thick cuticle; cuticles alsooccur between the innermost integument and the nucellusand over the surface of the ovule (Maheshwari, 1950).
Olde (1997) divides seeds of Grevillea into threemorphological types: (1) the ‘winged’ seed, in which amembranous outgrowth of the testa forms a ‘wing’ thatcompletely surrounds the seed; (2) the ‘oat’-style seed,which lacks the membranous wing but instead has marginsthat are revolute; and (3) an ellipsoidal/hemispherical seedwith a convex outer surface and a flat inner surface.
In Grevillea species the mature seeds are non-endosper-mic and the embryo completely occupies the embryo saccavity (Ventaka Rao, 1967). The testa is multilayered(owing to multiplication of the mesophyll cell layers),the cells have lost their tannin and the cells of the innerepidermis although thin-walled are radially elongated like a
palisade and contain prominent raphides (Venkata Rao,1967). The tegmen consists of three crushed cell layers(Venkata Rao, 1967) that in the wing-producing speciesG. robusta and G. banksii contributes to a fairly hard,protective covering surrounding the embryo (Kausik,1938, 1939). According to Corner (1976), the cells of thetegmen in Grevillea species are thick-walled, lignifiedfibres with the middle layer crossing the outer and innerlayers of longitudinal fibres.
Most of the studies on seed structure in Grevillea haveconcentrated on the winged seed-type. However, those eastAustralian species in which germination responses havebeen studied (Kenny, 2000; Morris, 2000; Morris et al.,2000) have the ‘oats’-style seed. There is clearly a needfor updated information on the basic structure and histo-chemistry of this style of Grevillea seed to provide a basisfor further investigations on the mechanism of action ofvarious germination treatments.
Grevillea linearifolia, G. buxifolia and G. sericea showgermination behaviour that is broadly similar, but with someimportant differences. Almost all seeds of G. buxifolia donot germinate unless treated with fire-related cues in labor-atory experiments, whereas a variable fraction of G. sericeaand G. linearifolia seeds do germinate without further treat-ment (Kenny, 2000; Morris, 2000). Heat shock and smokecombined synergistically to increase germination ofG. buxifolia, but additively for G. sericea (Kenny, 2000).To understand further the role of the seed coat in imposingdormancy, we have studied the structure of the mature seedof all three species, used histochemical techniques to deter-mine wall composition, and investigated the movement ofwater and high-molecular-weight compounds in intactseeds. This work is a necessary prerequisite for futureexperiments to test hypotheses about the breaking ofdormancy by fire-related germination cues.
MATERIALS AND METHODS
Seed source
Seeds of G. linearifolia (Cav.) Druce, G. sericea (Smith)R. Br and G. buxifolia (Smith) R. Br. were collected bypicking ripe fruits from multiple plants at Narrabeen, NewSouth Wales, in December 2000, returning the fruits to thelaboratory and collecting the seeds after the fruits opened.Seeds were stored in paper bags in the laboratory until used(October–December 2001 for gross dissections and initialhistochemistry; February–May 2004 for later histochem-istry and diffusion experiments).
Gross dissection/photography
Sixteen seeds of each species were placed on moist filterpaper in a Petri dish for 24 h, sectioned with a razor blade,and examined for gross morphology, anatomy and colour ofpigments in walls and vacuoles.
Tissue preparation for sectioning
Seeds of each species were fixed in a 5% glutaraldehydesolution in 2�5% potassium phosphate buffer, pH 6�8, at
966 Briggs et al. — Histochemistry of Seed Coat Structure in Grevillea Species
Dow
nloaded from https://academ
ic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022

room temperature for 4 d. The seeds were dehydratedthrough a graded ethanol series, gradually infiltrated withethanolic LR White resin (London Resin Co., London, UK)into 100% LR White, passed through three changes of100% LR White, stored at 4 �C for several weeks to com-plete infiltration and then embedded in LR White by poly-merization under UV light in a nitrogen-flow cabinet. Thinsections (1�0mm) were cut on a microtome from 4–5 seedsper species and mounted on glass slides prior to staining.
Histology
Semithin sections of LR White-embedded material werestained with 0�025% Toluidine Blue in 0�05M acetate buffer(pH 4�4), 1% Amido Black 10B in 7% acetic acid forproteins, Periodic Acid-Schiff (PAS) reaction (blockedfor 30min with 2,4-dinitrophenyl hydrazine in 15% aceticacid) for carbohydrates with 1,2 glycol groups, and 0�023%(w/v) aqueous (aq.) Calcofluor White M2R (CI 40622,Sigma, St Louis, USA) for b-linked glucans includingcellulose, following the methods of O’Brien and McCully(1981).
Handcut sections of unfixed material made in the trans-verse or medial longitudinal plane were stained with thefollowing dyes: Toluidine Blue, pH 4�4, and CalcofluorWhite, Sudan Black B for total lipids (see above),100mgmL�1 Nile Red for neutral lipids (Oparka andRead, 1994), 0�02% (aq.) Ruthenium Red for pectins andpectic acids, Chlor-Zinc-Iodide solution for cellulose/lignin/suberin, Acidic Phloroglucinol for lignin, and1�0% (w/v) Ferric Chloride in 0�1M HCl for tannins, fol-lowing the methods of Harris et al. (1994). Stained sectionswere mounted in water or glycerol. Freehand sections werealso stained with 0�1% (w/v, aq.) Berberine hemi-sulphate(Sigma, CI 75160) for 1 h then with 0�5% (w/v, aq.) AnilineBlue for 30min and mounted in glycerol–FeCl3 (0�1%, w/v)for suberin (Barnabas, 1994), or were stained only with0�1% (aq.) Berberine solution for 1 h, rinsed in distilledwater and mounted in water or glycerol. For confirmationof the presence of suberin, sections were covered with con-centrated H2SO4 (Barnabas, 1996) for 27 h, rinsed, thenstained with Sudan IV solution for 25min, rinsed brieflywith 70% ethanol, mounted in glycerol and examined.
Movement of water to the embryo
While intact seeds take up water (Morris, 2000), it is notknown whether water crossed the seed coat to the embryo.This was investigated by comparing the water content ofembryos from imbibed and unimbibed seeds of one speciesonly. Four seeds of G. sericea were placed into ten replicatePetri dishes lined with filter paper; five dishes wererandomly selected and water was added. After 24 h, theembryos were dissected out, dried at 80 �C for 24 h andthe water content calculated as a percentage of dry weight.Mean water content of imbibed and control seeds wascompared by using a t-test.
Apoplasmic tracing
Six seeds from each species were immersed in0�1% (w/v, aq.) Lucifer Yellow CH (Molecular Probes
L453 Lot 28C 1-5) for 24 h, rinsed three times in distilledwater, blotted, sectioned with a safety razor blade andexamined unmounted with an Olympus epifluorescentmicroscope using the 4· objective. Additional seeds wereimmersed in MilliQ water for the same amount of time todetermine autofluorescence.
Fluorescence microscopy
Sections stained with Calcofluor White M2R andBerberine hemi-sulphate were examined under UV excita-tion (excitation filter UG.1, dichroic mirror 400 nm, barrierfilter 420 nm). Sections stained with Nile Red and LuciferYellow CHwere examined under blue excitation (excitationfilter 490 nm, dichroic mirror 500 nm, barrier filter 515 nm).Unstained, unfixed sections were examined under UV andblue excitation for autofluorescence.
Fluorescence photomicrography was mainly carried outon an Olympus BHC epifluorescence microscope usingFuji Xtra 400 ASA colour film. Prints were scanned witha Canon Scanner N1240U USB using ArcSoft PhotoStudio2000. All images of the apoplasmic tracing with LuciferYellow were taken via a water-cooled digital camera withexposures of 145, 193 and 269ms. Images were assembledinto plates using Adobe Illustrator 11.0.0.
RESULTS
When seeds of all three species gave similar results, resultsare reported for only one species. Where differences wereapparent, results are reported accordingly.
Gross morphology and anatomy of the mature seed
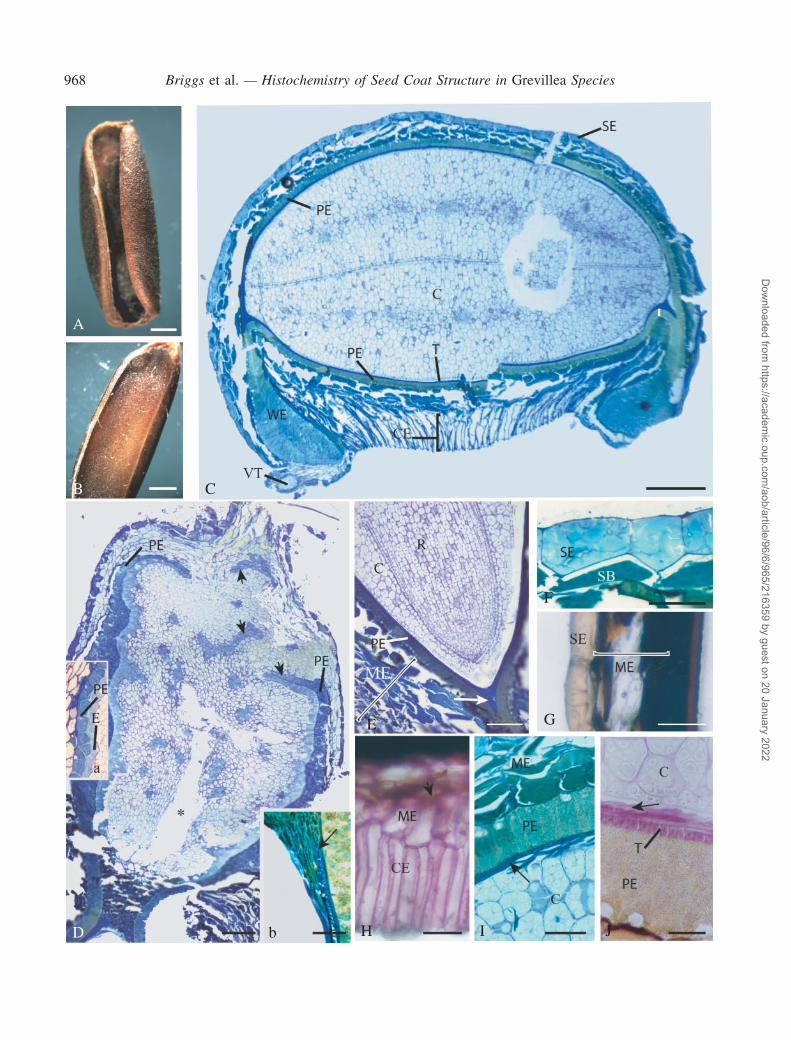
The dry seed was elongated along the main axis, with anelaiosome at the former placental (i.e. chalazal) end andrevolute margins folded under the seed (Fig. 1A). Imbibitionresulted in a generalized swelling of the seed and a confor-mational change, as the margins of the seed coat unfoldedfrom under the seed to a more lateral position (Fig. 1B).The embryo, which consisted of two cotyledons, a plumuleand radicle, completely filled the embryo sac and therewas no endosperm remaining (Fig. 1C). Between theembryo and the elaiosome was a structural discontinuity,the hypostase (Fig. 1D). A single vascular trace was foundthrough the central region of the elaiosome, stoppingjust before the hypostase. Unfortunately, the vasculartrace did not often survive sectioning and dropped out leav-ing an empty space (Fig. 1D). The radicle and plumuleoccupied a small amount of space at the micropylar end(Fig. 1E).
Internal structure of the seed coat
Exotesta. The exotesta covering the lateral regions,upper surface, chalazal and micropylar regions of theseed consisted of small, squat, polygonal-shaped cells(Fig. 1C, D, F and G) but along the lower surface of theseed, between the two revolute lateral margins (wings inFig. 1C), the cells were radially elongate, i.e. columnar inshape (Fig. 1H).
Briggs et al. — Histochemistry of Seed Coat Structure in Grevillea Species 967
Dow
nloaded from https://academ
ic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022
A
B C
D b
E
F
G
IH J
968 Briggs et al. — Histochemistry of Seed Coat Structure in Grevillea Species
Dow
nloaded from https://academ
ic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022

Mesotesta. Immediately beneath the squat exotestal cellsthere were 3–4 cell layers of thin-walled pigmented cells(Fig. 1C, D, F, G and I); the number of cell layers increasedbeneath the columnar exotestal cells (Fig. 1C) and at themicropylar end (Fig. 1E) where they abutted the wing-likeextension of the endotesta.
Endotesta layer. The endotesta surrounded the lateralsides of the embryo sac (Fig. 1C) and continued into theelaiosome as far as the elaiosome apex (Fig. 1D). It con-sisted of thin-walled, palisade-shaped cells with a lace-likefibrous reticular network filling the interior (Fig. 1C, E, Iand J). It was one cell thick around the embryo sac (Fig. 1C)and elaiosome [Figs 1D (inset a) and 2A], but in the ‘wing-like’ regions it was folded back on itself so that it appearedtwo cell layers thick (Figs 1C and 2B) and the radial andinner tangential cell walls were much thicker (Fig. 2B). Thepalisade cells of the endotesta did not extend across theelaiosome apex nor across the hypostase (Fig. 1D). Incross-section, small square-shaped areas occurred close tothe outer tangential wall. These areas represent the locationof crystals.
Tegmen. The tegmen consisted of 2–3 cell layers ofelongated, thick-walled cells (Figs 1I and J. and 2C–F).Although non-lignified (see below under Histology), wehave still termed these cells ‘sclerenchyma’ followingthe broad definition of Esau (1960). In transverse sectionsthrough themiddle of the seed, the long axis of the exotegmicsclerenchyma cells was parallel to the long the axis of theseed, while the second, endotegmic layer was perpendicularto the first, running around the seed. In the recurved marginsof the seed, the tegmenwaswedge-shaped (Fig. 2C, E and F).In longitudinal sections, the tegmen formed another wedge-shaped region around the micropylar end of the embryo sac(Fig. 1E) and at the chalazal end extended just past the end ofthe embryo sac to line the sides of the hypostase, ending there(Fig. 1D, inset b). The sclerenchyma cells were not continu-ous across the hypostase (Fig. 1D, inset b).A cuticle occurredon either side of the tegmen (Fig. 2D).
Hypostase. The hypostase consisted of brown-pigmentedcells that could be grouped into two distinct regions. Theinnermost region abutting the embryo sac consisted of
irregular, tessellated cells and the outer region abuttingthe elaiosome consisted of less distorted cells (Fig. 2G).The number of cells forming the hypostase varied betweenthe three species; G. buxifolia had the greatest number ofcell layers (i.e. �20), with the other two species having�14–16 layers.
Elaiosome. The elaiosome consisted of a central regionof large, variously shaped thin-walled cells bounded by an‘epidermis’-like layer (Figs 1D, inset a, and 2A) thatstarted at the hypostase and extended along the sides ofthe central region with short, bilayered extensions intothe central region (Fig. 1D). As mentioned above, outsidethis central region were the endotesta, mesotesta andexotesta layers.
Crushed cell layer. A glutinous layer of crushed cellsoccurred between the endotegmen and the lateral sides ofthe embryo.
Histochemistry of the seed coat. Fresh and fixed/embedded samples were examined unstained and stainedwith several bright-field and fluorescent dyes to investigatethe composition of the cell walls, vacuolar content and thepresence/absence of cuticles. Although staining intensityand colour depths varied according to the thickness of thesection, similar staining reactions were found in all threespecies for each of the different cell layers and types. Alldescriptions apply to all three species unless indicatedotherwise.
Exotesta. The vacuoles of squat exotestal cells containeda brown substance that appeared sky blue in fixed (Fig. 1F)and fresh (not shown) material when stained with ToluidineBlue at pH 4�4 (indicating phenols), was positive withAmido Black, weakly positive with PAS and slightlycoloured brown with Chlor-Zinc-Iodide (Fig. 1G), but wasnegative for tannins when stained with FeCl3 (Table 1).Irregular lines ran throughout the centre of the cells.These lines stained darker with Toluidine Blue (Fig. 1F)and Chlor-Zinc-Iodide (Fig. 1G). This material was notfound in the columnar exotestal cells.
The walls of the squat exotestal cells (in both fresh andfixed material) exhibited various staining reactions withToluidine Blue at pH 4�4. The outer tangential and radial
F I G . 1. (A) Grevillea buxifolia. External view of a dry seed showing the recurved lateral margin. Elaiosome is at the top end of the figure. Scale bar = 1mm.(B) G. buxifolia. External view of imbibed seed showing the lateral margins unrolled. Elaiosome is at top end of the figure. Scale bar = 2mm. (C) G. sericea.Resin-embedded cross-section (TS) through the mid-transverse plane of a dry seed stained with Toluidine Blue showing squat exotestal cells (SE), columnarexotestal cells (CE), palisade endotesta (PE),wing endotesta (WE), tegmen (T), cotyledon (C) andvascular trace (VT). Scale bar=400mm. (D)G. linearifolia.Resin-embedded longitudinal section (LS) through the elaiosome and hypostase (H). Montage of several abutting photographs shows that the palisadeendotestal layer reached but did not enclose the elaiosome core tissue and that only the abutting elaiosome ‘epidermal-like’ layer extended into the elaiosomecore (arrowheads). The position of the vascular trace corresponds to the empty channel in the centre of the core (*). Inset (a) shows a portion of the palisadeendotesta and elaiosome ‘epidermal’ layer with the thickened cell walls from G. buxifolia. Inset (b) shows the terminus of the tegmen at the edge of thehypostase (arrow). Scale bar = 150mm (60mm in b). (E) G. linearifolia. Resin-embedded LS through the radicle and inner part of the micropyle stained withToluidine Blue. Palisade endotesta (PE), tegmen ‘wedge’ (arrow), mesotesta (ME), radical (R), cotyledon (C). Scale bar = 150mm. (F) G. buxifolia. Resin-embedded TS showing the squat exotestal cells (SE) and adjacent mesotestal cells (ME) stained with Toluidine Blue. The outer tangential and radial wallsstained purple but the inner tangential walls stained blue. The pigment in the vacuoles of the squat exotestal and mesotestal cells stained light blue and darkblue–green, respectively, suggesting phenolic compounds. Scale bar = 40mm. (G) G. linearifolia. Hand-cut TS through seed coat stained with Chlor-Zinc-Iodide. The outer transverse and radial walls stained purple but the inner tangential walls could not be clearly seen. There was no obvious staining reactionfrom the pigment in the vacuolar contents. Squat exotesta (SE),mesotesta (ME). Scale bar = 60mm. (H)G. linearifolia. Hand-cut TS through the lower side ofthe seed stainedwithRutheniumRed.Thewalls of the columnar exotestal (CE) andmesotestal (ME) cells are positive. The vacuolar contents of themesotestalcells were amber in colour (arrowhead). Scale bar = 25mm. (I) G. linearifolia. Resin-embedded TS of inner part of the seed coat stained with Toluidine Blue.Mesotesta (ME), palisade endotesta (PE), tegmen (arrow), cotyledon (C). Scale bar = 50mm. (J) G. sericea. Resin-embedded TS stained with PAS reagents.The cell walls of the cotyledon (C), crushed cell layer (arrow) and tegmen (T) were PAS positive whereas the walls and fibrillar network of the palisade
endotesta (PE) were not. Scale bar = 25mm.
Briggs et al. — Histochemistry of Seed Coat Structure in Grevillea Species 969
Dow
nloaded from https://academ
ic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022

walls stained purple but the inner tangential walls stainedblue (Fig. 1F). The walls stained purple with Chlor-Zinc-Iodide (Fig. 1G), stained positively with Amido Black, PAS,Ruthenium Red, Calcofluor White, Nile Red, Berberine
hemi-sulphate and the Berberine/Aniline Blue combination(Tables 1 –3). They also stained with Sudan IV both beforeand after prolonged reaction with H2SO4, confirming thepresence of suberin.
A B C D
E
H I J K
L M N
F G
970 Briggs et al. — Histochemistry of Seed Coat Structure in Grevillea Species
Dow
nloaded from https://academ
ic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022

With Toluidine Blue at pH 4�4, the outer tangential wallsand outer part of the radial walls of the columnar exotestalcell stained purple, indicating the presence of polyanions,whereas the inner half (or base) of the radial walls wereblue, indicating phenols (not shown). All walls of thecolumnar exotestal cells stained with Amido Black(weakly), Ruthenium Red (Fig. 1H) and PAS reagents(Table 1) and were autofluorescent white under UVillumination (Fig. 2H). In G. linearifolia and G. sericea,the entire wall stained with Calcofluor White, but inG. buxifolia only the tangential walls reacted (Table 2).After staining with Berberine hemi-sulphate, or the Berber-ine/Aniline Blue combination, the walls fluoresced anintense yellow (Fig. 2I), indicating polyanions and phenols.After Nile Red they stained emerald green (Fig. 2J) (Table 3)and after acid/Sudan IV they were red, indicating suberin.The cuticle over the surface of the exotesta reacted for lipid(orange) with Nile Red (Fig. 2J).
Mesotesta. The vacuoles of the mesophyll cells formingthe mesotesta contained an ecru- to honey-coloured pigment(Fig. 2F) that was unstained by Chlor-Zinc-Iodide solution(Fig. 1G). Following staining with Toluidine Blue, the pig-ment was brown/blue (Fig. 1F and I), suggesting phenolicmaterial. There was no reaction with FeCl3 (indicating anabsence of tannins) nor with any other stain tested (Tables 1and 3). The walls of the mesophyll cells in all speciesstained with Amido Black, PAS and Ruthenium Red. InG. linearifolia and G. sericea they stained purple with
Toluidine Blue but gave a blue reaction in G. buxifolia(Table 1). The latter indicates incorporation of phenolsinto the walls, but this was not lignin because walls ofall species did not react with Phloroglucinol–HCl. Follow-ing staining with Berberine hemi-sulphate solution, inG. buxifolia the walls in the outer cell layers of the meso-testa fluoresced gold (Fig. 2I) but the innermost layers abut-ting the endotesta were not fluorescent. In G. sericea thewalls fluoresced a weaker yellow, and in G. linearifolia theywere not fluorescent. With the Berberine/Aniline Blue com-bination there was no reaction from the walls of mesophyllcells beneath squat exotestal cells although there was somefluorescence from cells abutting the columnar exotestalcells in G. sericea, but not in G. linearifolia. Followingstaining with Nile Red, the walls were emerald green(Fig. 2J, Table 2).
Endotesta. Cells of the palisade-shaped endotestastained somewhat differently depending on whether theywere located close to the embryo, in the margins of theseed coat or around the elaiosome. The lace-like fibrillarnetwork of the cells close to the embryo and in the wingregion appeared yellowish outlined by blue–green whenstained with Toluidine Blue (Fig. 1I). The contents of cellsabutting the elaiosome stained blue with Toluidine Blue(Fig. 1D, insert a). The cell contents did not react withany other stains tested except for Amido Black, which out-lined the lace-like fibrillar network (Table 1). The cell wallsalso gave varying staining reactions depending upon cell
F I G . 2. (A) G. linearifolia. Hand-cut cross-section (TS) through the elaiosome and adjacent endotesta stained with Phloroglucinol–HCl. All of the walls ofthe palisade endotesta (PE) and the outer tangential and radial walls of the abutting ‘epidermal-like’ layer (arrows) gave a positive response for lignin. Theelaiosome oil cells (EO) were negative. Scale bar = 100mm. (B) G. linearifolia. Hand-cut TS through the wing-like endotestal region stained withPhloroglucinol–HCl. Strong magenta staining of the radial and inner tangential walls indicates lignin. The adjacent squat exotestal cells (SE) did not stainand were easily detached. Scale bar = 30mm. (C) G. linearifolia. Resin-embedded longitudinal section (LS) through the tegmen wedge (TW) stained withToluidine Blue. The radial walls of the palisade endotesta (PE) layer stained yellow. Scale bar = 50mm. (D)G. linearifolia. Hand-cut TS through the palisadeendotesta (PE) and abutting tegmen (T) stained with Nile Red. The inner cuticle (arrowhead) between the tegmen and the crushed cell layer was thinner anddiscontinuous comparedwith the thicker outer cuticle (arrow) lyingbetween the exotegmen and the endotesta. Scale bar=20mm. (E)G. linearifolia. Hand-cutTS through the tegmen wedge (TW) and palisade endotestal (PE) layers stained with Nile Red and viewed by blue excitation (5-s exposure). The outer of thetwo inner cuticles (arrow) is thick and strongly fluoresced gold. Note the green fluorescence (arrowhead) of the lignifiedwalls of the endotestal cells. Scale bar= 20mm. (F)G. linearifolia. Resin-embedded TS stainedwithAmidoBlack for protein. The cotyledons contained numerous black protein bodies (arrow) andthe crushed cell layer (arrowhead) was also positively stained, but the tegmen (T) and palisade endotesta (PE) layers were unstained. Scale bar = 100mm. (G)G. buxifolia. Hand-cut LS through part of the hypostase stainedwith SudanBlackB.Only thewalls of the inner cell layers of the hypostase (i.e. adjacent to thecotyledons) gave a strong positive response for lipids (arrows), whereas the walls of the outer part of the hypostase (i.e. next to the elaiosome) did not. Thepigment in the hypostase cells was unstained with Sudan Black B. Scale bar = 20mm. (H) G. linearifolia. Hand-cut TS viewed under UV excitation forautofluorescence (6�4-s exposure). The walls of the columnar exotestal cells (CE) were strongly autofluorescent (white). In the palisade endotestal cells (PE),only square-shaped areas (arrowhead) abutting the outer tangential walls of the endotestal cells were autofluorescent (blue); the rest of the walls and contentswere not. The walls of the tegmen (T) were weakly fluorescent (white). The walls and contents of themesotesta (ME)were very weakly autofluorecent. Scalebar = 100mm. (I) G. buxifolia. Hand-cut TS stained with Berberine Sulphate solution and viewed with UV excitation (6�4-s exposure). The yellowfluorescence of the Berberine-stained mesotestal (ME) cell walls can be seen against the strong yellowish-white fluorescence of the columnar exotesta(CE).Note that the inner tangential and lower radialwall regions of the exotestal cells (arrow) fluoresced yellow rather than yellow–white. Scale bar= 100mm.(J) G. linearifolia. Hand-cut TS through the columnar exotestal and adjacent mesotestal cells stained with Nile Red and viewed with blue excitation (6�4-sexposure). The external cuticle (arrow) fluoresced orange/gold. The fluorescence in thewalls of the columnar exotestal (CE) cells wasmore golden due to thecombined Nile Red fluorescence and autofluorescence. Note the faint emerald green fluorescence of the mesotestal (ME) cell walls (arrowhead). Scale bar =100mm. (K) G. buxifolia. Hand-cut TS stained with Berberine Sulphate solution and viewed with UV excitation (2�81-s exposure). The vacuolar contents ofthe squat exotestal cells (SE) strongly fluoresced yellow.Weaker yellowfluorescencewas seen in thewalls of themesotestal (ME) cells and the lignifiedwallsof the endotestal wing (WE) cells. Scale bar = 100mm. (L) G. buxifolia. Hand-cut TS stained with CalcofluorWhiteM2R (12�8-s exposure). The walls of theendotestal wing (WE) cells were strongly fluorescent (bluishwhite) as was the internal fibrillar network.Mesotestal cells (ME) also fluoresced blue–white butnot as strongly as the wing cell walls. Vacuolar contents of the squat exotestal cells (SE) autofluoresced strongly. Scale bar = 100mm. (M) G. linearifolia.Hand-cut TS viewed under UV excitation for autofluorescence (6�4-s exposure). The endotestal wing cells (WE) showed strong autofluorescence (white) ofthe lignified radial walls with noticeable but weaker (bluish) autofluorescence from the cell contents. The walls and contents of the surrounding mesotestalcells (ME) were weakly autofluorescent. Scale bar = 100mm. (N) G. sericea. Hand-cut LS through the hypostase (H) and adjacent palisade endotestal (PE)layer stainedwithNileRed viewed by blue excitation (6�4-s exposure). Any orangefluorescence due to theNileRed binding to neutral lipids in thewalls of theinner layers of the hypostase (HI) was masked by the yellow autofluorescence of the walls. The cells in the adjacent cotyledon (C) were strongly
autofluorescent yellow. Scale bar = 20mm.
Briggs et al. — Histochemistry of Seed Coat Structure in Grevillea Species 971
Dow
nloaded from https://academ
ic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022
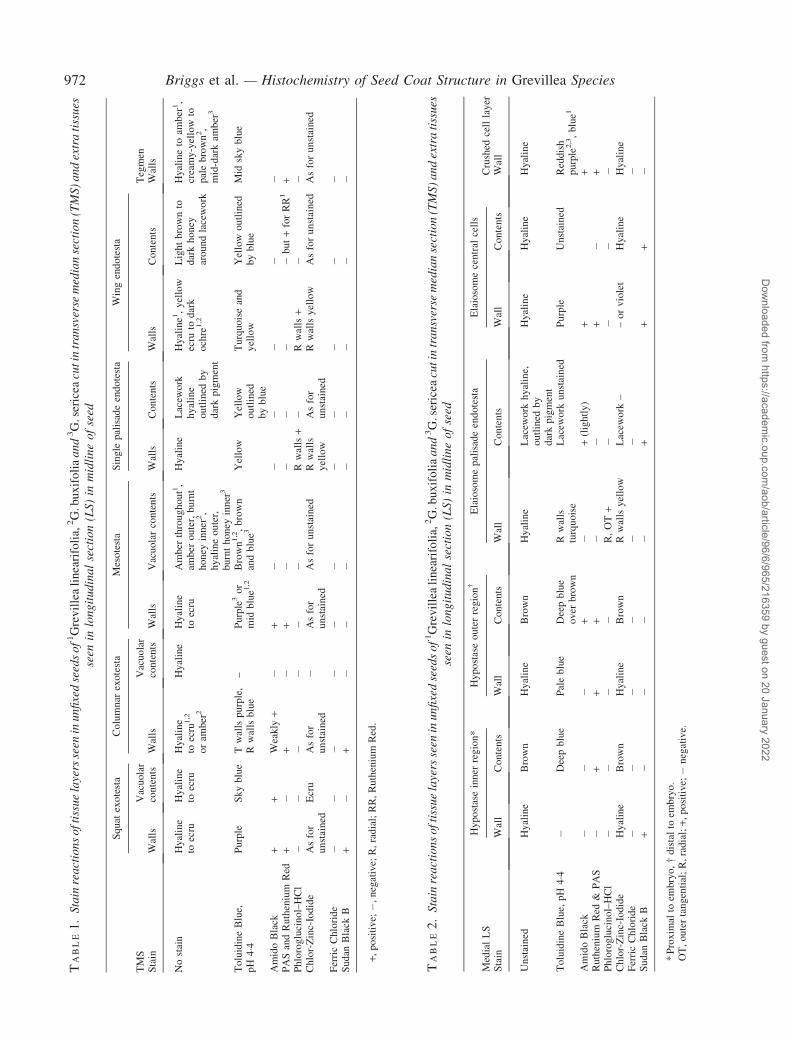
TABLE1.
Sta
inre
act
ion
so
fti
ssu
ela
yers
seen
inu
nfi
xed
seed
so
f1Grevillealinearifolia,
2G.buxifolia
an
d3G.sericea
cutin
tra
nsv
erse
med
ian
sect
ion
(TM
S)
an
dex
tra
tiss
ues
seen
inlo
ng
itu
din
al
sect
ion
(LS
)in
mid
lin
eo
fse
ed
Squat
exotesta
Columnar
exotesta
Mesotesta
Single
palisadeendotesta
Wingendotesta
TMS
Stain
Walls
Vacuolar
contents
Walls
Vacuolar
contents
Walls
Vacuolarcontents
Walls
Contents
Walls
Contents
Tegmen
Walls
Nostain
Hyaline
toecru
Hyaline
toecru
Hyaline
toecru
1,2
oram
ber
2
Hyaline
Hyaline
toecru
Amber
throughout1,
amber
outer,burnt
honey
inner
2,
hyalineouter,
burnthoney
inner
3
Hyaline
Lacew
ork
hyaline
outlined
by
darkpigment
Hyaline1,yellow
ecru
todark
ochre
1,2
Lightbrownto
darkhoney
aroundlacework
Hyalineto
amber
1,
creamy-yellow
topalebrown2,
mid-darkam
ber
3
ToluidineBlue,
pH
4. 4
Purple
Skyblue
Twallspurple,
Rwallsblue
–Purple3or
mid
blue1
,2Brown1,2,brown
andblue3
Yellow
Yellow
outlined
byblue
Turquoiseand
yellow
Yellow
outlined
byblue
Mid
skyblue
AmidoBlack
++
Weakly
+�
+�
��
��
�PASandRuthenium
Red
+�
+�
+�
��
��
but+forRR1
+Phloroglucinol–HCl
��
��
��
Rwalls+
�Rwalls+
��
Chlor-Zinc-Iodide
Asfor
unstained
Ecru
Asfor
unstained
�Asfor
unstained
Asforunstained
Rwalls
yellow
Asfor
unstained
Rwallsyellow
Asforunstained
Asforunstained
FerricChloride
��
��
��
��
��
�Sudan
Black
B+
�+
��
��
��
��
+,positive;�,negative;
R,radial;RR,Ruthenium
Red.
TABLE2.
Sta
inre
act
ion
so
fti
ssu
ela
yers
seen
inu
nfi
xed
seed
so
f1Grevillealinearifolia,
2G.buxifolia
an
d3G.sericea
cutin
tra
nsv
erse
med
ian
sect
ion
(TM
S)
an
dex
tra
tiss
ues
seen
inlo
ng
itu
din
al
sect
ion
(LS
)in
mid
lin
eo
fse
ed
MedialLS
Hypostaseinner
region*
Hypostaseouterregion†
Elaiosomepalisadeendotesta
Elaiosomecentral
cells
Crushed
celllayer
Stain
Wall
Contents
Wall
Contents
Wall
Contents
Wall
Contents
Wall
Unstained
Hyaline
Brown
Hyaline
Brown
Hyaline
Lacew
ork
hyaline,
outlined
by
darkpigment
Hyaline
Hyaline
Hyaline
ToluidineBlue,
pH
4. 4
�Deepblue
Paleblue
Deepblue
over
brown
Rwalls
turquoise
Lacew
ork
unstained
Purple
Unstained
Reddish
purple2,3,blue1
AmidoBlack
��
�+
�+(lightly)
++
Ruthenium
Red
&PAS
�+
++
��
+�
+Phloroglucinol–HCl
��
��
R,OT+
��
��
Chlor-Zinc-Iodide
Hyaline
Brown
Hyaline
Brown
Rwallsyellow
Lacew
ork
––orviolet
Hyaline
Hyaline
FerricChloride
��
��
��
�Sudan
Black
B+
��
��
++
+�
*Proxim
alto
embryo,†distalto
embryo.
OT,outertangential;R,radial;+,
positive;
�negative.
972 Briggs et al. — Histochemistry of Seed Coat Structure in Grevillea Species
Dow
nloaded from https://academ
ic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022

TABLE3.
Flu
ore
scen
tre
act
ion
so
fti
ssu
ela
yers
seen
inse
eds
of1Grevillealinearifolia,2G.buxifolia
an
d3G.sericea
cut
intr
an
sver
sem
edia
nse
ctio
n(T
MS
)a
nd
extr
ati
ssu
esse
enin
lon
git
ud
ina
lse
ctio
n(L
S)
inm
idli
ne
of
seed
Squat
exotesta
Columnar
exotesta
Mesotesta
Single
palisade
endotesta
Wingendotesta
TMS
Stain
Walls
Vacuolar
contents
Walls
Vacuolar
contents
Walls
Vacuolar
contents
Walls
Contents
Walls
Contents
Tegmen
Walls
Autofluorescence
UV
excitation.40·
objectivepositive
response
(White)
Cuticle+
Walls+,
6in
3+(1,2)
but–in
3Cuticle+
Allwalls6
1,3
butmostof
Rwalls++
+in
2
��
��
�Rwallsin
expanded
area
intensely
white1
,2,
butpalebluein
3;
restofcells6
LW
6Cuticleover
tangential
surfaces
+Walls–
Autofluorescence
Blueexcitation.
40·objective
Positiveresponse
(Yellow)
Cuticle+
Walls++
+++
+++
+1,2,6
3�
–exceptnext
tocolumnar
cellsin
1
O-amber
1R walls+1
,3LW
O1,
62,–3
Rwalls+,
other
walls6
or–
LW
white
Cuticleover
surfaceyellow
Walls6
CalcofluorWhite
+–,yellow1
+1,3but
Rwalls–2
�+
�6
LW
portion6
6LW
6Mostly
–but
Rwallsin
wedge
regionwere6
NileRed
Positive
(gold
ororange)
Interm
ittent
cuticleover
surface+
WallsEG
–exceptfor
internal
mem
branes
Cuticleover
surface+
WallsEG
–except
forinternal
mem
branes
EG
–except
forinternal
mem
branes
�LW
EG2,O3,–1
��
�
Berberine
Hem
isulphate
positive(yellow)
++
+andwhite
�OL+,
IL�
�6
+Rwalls+
++
6,weakly
positive;
+,positive;
+++,
intensely
positive;–,negative;
OL,outercelllayers;IL,inner
celllayers;EG,em
eraldgreen;O,orange;
LW,lace
work.
Briggs et al. — Histochemistry of Seed Coat Structure in Grevillea Species 973
Dow
nloaded from https://academ
ic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022

location. Where the endotesta was single layered, the wallsappeared yellow after Toluidine Blue staining (Figs 1I and2C), but in the wing-like regions, the radial and inner tan-gential walls of the cells stained turquoise with ToluidineBlue (not shown). Around the elaiosome, the radial wallswere slightly thicker in G. buxifolia and stained blue–green(Fig 1D, inset a) but in G. linearifolia and G. sericea thesewalls were thinner and stained yellow (not shown). How-ever, the radial walls of all endotestal cells reacted posi-tively with Phloroglucinol–HCl, as did the inner tangentialwalls of cells in the wing-like region (Fig. 2B) and thoseabutting the elaiosome (Fig. 2A). After Chlor-Zinc-Iodide,the radial walls of all endotestal cells were yellow.Following Berberine/Aniline Blue staining the walls inall endotestal cells fluoresced yellow but when stained byBerberine hemi-sulphate alone, only the elaiosome andwing endotestal cells gave a positive reaction (greeny yel-low) (Fig. 2K). The endotestal walls did not react with otherstains (Table 1), except for Calcofluor White, which gavea slight increase in fluorescence (Fig. 2L) compared withautofluorescence in controls (Fig. 2M), and Nile Red,which gave green fluorescence in (Table 3).
Tegmen. The thick cell walls of the tegmen stained deepblue with Toluidine Blue (Fig. 1C and I), pink with PAS(Fig. 1J) and Ruthenium Red, and blue–black with AmidoBlack but were negative with all other bright-field stainsespecially those testing for lignin (Table 1). They were onlypartially positive for b-glucan (probably cellulose), exceptin the wedge-shaped region where a stronger reaction wasfound in the radial walls (Table 3). Following staining withBerberine hemi-sulphate alone, the walls fluoresced goldenamber but stained negatively after the Berberine/AnilineBlue combination.
There was a thick cuticle between the exotegmen andthe endotesta, particularly around the wedge-shaped areas(Fig. 2D and E), and between the endotegmen and thecrushed cell layer there was a thin and sometimes discon-tinuous cuticle. The cuticles were autofluorescent and gavea positive reaction with Nile Red (Fig. 2D and E) and withSudan Black B (not shown).
Crushed cell layer. The amorphous material lyingbetween the endotegmen and the embryo stained blue withToluidine Blue (Fig. 1I) but only gave a positive reactionwith PAS (Fig. 1J), Amido Black (Fig. 2F), Ruthenium Red(Table 1) and Calcofluor White (Table 2, 3).
Hypostase. In the hypostase cells, the brown pigment inthe vacuoles suggested the presence of tannins, but therewas no positive reaction with FeCl3 in any seed. Only thecell walls of the inner 5–7 layers of cells immediately adja-cent to the cotyledons stained positively with Sudan Blackfor total lipids (Fig. 2G). Nile Red-induced fluorescence forneutral lipids (Fig. 2N, Table 4) varied but was impossibleto interpret because the walls of the inner layers of thehypostase were strongly autofluorescent (canary yellow)under blue excitation. When seeds were pre-stained withPhloroglucinol–HCl and examined under UV and blueexcitation, only the radial walls of the endotestal cells
lost their autofluorescence. When counterstained withChlor-Zinc-Iodide solution, there was a diminution of UVautofluorescence in the cell walls of the inner layers of thehypostase, indicating that suberin may be present (notshown). The greatest loss of autofluorescence occurred inG. sericea, followed by G. linearifolia. There was onlyminimal loss of autofluorescence in G. buxifolia. Followingstaining with Berberine hemi-sulphate the walls of theinnermost cell layers fluoresced brightly yellow (Table 4)but the outer five layers (i.e. next to the elaiosome) did not(not shown).
Histochemistry of the central elaiosome cells and embryo
Central elaiosome cells. The thin walls of cells of thecentral region of the elaiosome stained purple withToluidine Blue (Fig. 1D), and gave a positive reactionwith PAS, Calcofluor White and Berberine hemi-sulphate(Tables 1, 4); the cell contents stained with Sudan Black,Nile Red (lipids) and Berberine hemi-sulphate (not shown),and the cytoplasm was positive with Amido Black (notshown).
Embryo. The cells of the cotyledons contained numerousstorage bodies that were negative for starch when stainedby the PAS reaction (Fig. 1J) but were positive for proteins(Fig. 2G) and lipids (not shown).
Movement of water to the embryo
The water content of embryos from seeds of G. sericeathat had imbibed for 24 h (mean 30�46 0�5%) was about sixtimes greater than that of embryos from dry seeds [mean5�3 6 0�02%; comparison of (log-transformed) means,t8d.f. = 100�6, P < 0�001].
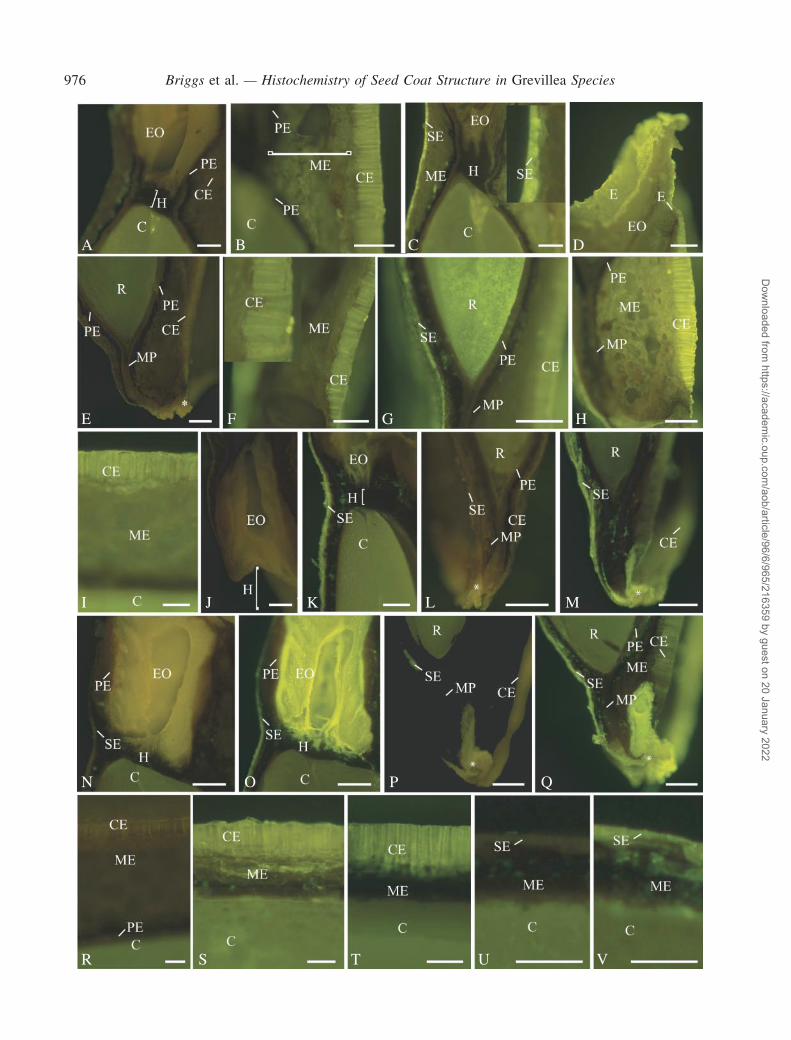
Apoplasmic tracing with Lucifer Yellow CH
Seeds examined after soaking for 24 h in Lucifer Yellow(LY) showed a variable fluorescence that differed fromautofluorescence in the controls. In all species, there wasstrong lemon-yellow fluorescence indicative of LY in theouter tangential and radial walls of the columnar and squatexotestal cells in all regions of the seed (Fig. 3B–D, F–I, K,M, O, Q, S, T and V). This was quite distinct from the dullmore olive-green autofluorescence of these tissues in thecontrols (Fig. 3A, E, J, L, N, P, R and U). In two G. buxifoliaseeds (Fig. 3H and I) and one G. sericea seed, there wasalso LY-induced fluorescence in the adjacent mesophyllcell layers and in one G. buxifolia seed section the radicle(Fig. 3G) and cotyledons (Fig. 3I) also exhibited LY-fluorescence. In other replicates of these species therewas no LY-fluorescence in the embryo or mesophyll cellwalls. Lucifer Yellow consistently accumulated around theoil-filled thin-walled elaiosome cells in G. sericea (compareFig. 3N and O), and in one replicate of G. linearifolia (notshown). Lucifer Yellow also accumulated in certain cells inthe micropylar region (compare Fig. 3P and Q); the origin ofthese cells is unknown. No LY-fluorescence was found inany of the cells of the endotesta or tegmen (Fig. 3G, K, O, S,T and V).
974 Briggs et al. — Histochemistry of Seed Coat Structure in Grevillea Species
Dow
nloaded from https://academ
ic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022

TABLE4.
Flu
ore
scen
tre
act
ion
so
fti
ssu
ela
yers
seen
inse
eds
of1Grevillealinearifolia,
2G.buxifolia
an
d3G.sericea
cutin
tra
nsv
erse
med
ian
sect
ion
(TM
S)
an
dex
tra
tiss
ues
seen
inlo
ng
itu
din
al
sect
ion
(LS
)in
mid
lin
eo
fse
ed
MedialLS
Hypostaseinner
region*
Hypostaseouterregion†
Elaiosomepalisade
Elaiosomecentral
cells
Stain
Wall
Contents
Wall
Contents
Wall
Contents
Wall
Contents
Crushed
celllayer
Autofluorescence
UV
excitation.
Positive:
white=p1
blue–white=p2
yellow–white=p3
+p2.Note
in2,a
yellow
band
occurred
inthe
middle
ofthis
region
��
�+p
11,p22,6p13
–1,p22,–3
Intensely
p31,
p22,3
Intensely
p31,p22,3
�
Autofluorescence
Blueexcitation.
Positiveresponse
Canaryyellow
+++
��
�Rwalls+
+++
+++
++sometim
esalso
yellow–green
CalcofluorWhite.
Positiveresponse
blue/
blue-white
��
+�
�6
+�
+++
NileRed
Orange
Positiveresponse
gold
ororange
No+lineover
the
surfaceofthis
region.Walls
yellow
Mem
branes
+�
�Green
LW
lime-green
1,2
gold
3Yellow–green
Yellow–green
�
Berberine
Hem
isulphate
Positiveyellow
+�
��
++
++
�
*Proxim
alto
embryo,†distalto
embryo.
+,positive;
+++,
intensely
positive;–negative;
LW,lace
work.
Briggs et al. — Histochemistry of Seed Coat Structure in Grevillea Species 975
Dow
nloaded from https://academ
ic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022
A
E
I
N
R S T U V
O P Q
J K L M
F G H
B C DC
976 Briggs et al. — Histochemistry of Seed Coat Structure in Grevillea Species
Dow
nloaded from https://academ
ic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022

DISCUSSION
Seed coat structure
The anatomy of the seed coat of the three Grevillea speciesexamined in this study is consistent with previous descrip-tions, particularly those by Corner (1976), for Grevilleaspecies and the Proteaceae. The presence of the two internalcuticles, one on either side of the sclerenchyma layer, con-firms that the bi-layered sclerenchyma is the tegmen and theinner palisade layer with the fibrillar contents and crystalsrepresents the endotesta. The layer of crushed cells betweenthe tegmen and the embryo may be the remains of thenucellus (Venkata Rao, 1967; Corner, 1976), although astudy of the development of these seeds is needed to confirmthis. The revolute margins of the seeds appear to be formedby the folding of the testa and tegmen, as evidenced by theexpansion and folding of the single-layered palisade-likeendotesta into the double-layered palisade forming the‘wing’; the formation of the sclerenchyma ‘wedge’ indi-cates that the tegmen has been similarly folded. The highlytessellated cells lying at the chalazal or elaiosome end of theseed clearly represent a hypostase, as the sclerenchymatoustegmen extends along the lateral edges, but not across thiszone and the zone lies between the elaiosome and theembryo sac. The columnar exotestal cells and adjacent
mesophyll cells constitute the ‘narrow wing’ of the oat-styleseed.
Our study of the anatomy and histochemistry of the seedcoat has identified a number of potential dormancy mecha-nisms. These will now be considered in turn.
Barriers to diffusion
We observed an exterior cuticle, and two internal cuticleson either side of the tegmen. Cuticles over the surfaces ofleaves, fruits and seeds consist of a cutin membrane overlainwith waxes. The semi-permeable nature of cuticles isthought to permit the inward diffusion of water and oxygen(Bewley and Black, 1982), but prevents the outward flow ofendogenous inhibitors such as polyphenolics (Swain, 1979,cited in Egerton-Warbuton, 1998). A comparison of thewater content of embryos from imbibed and dry seeds ofG. sericea indicated that water crosses the seed coat andreaches the embryo; thus, the internal cuticles are permeableto water.
The cuticles identified in the seed coat of Grevillea arepotential sites for action by smoke (Egerton-Warburton,1998). In the seeds of the chaparral species Emmenanthependuliflora, a post-fire recruiter, Lucifer Yellow CH (LY)penetrated the external cuticle but was stopped by the
F I G . 3. Hand-cut LS through various regions of seeds immersed either in Lucifer Yellow (LY) for 24 h or in distilled water as controls to determineautofluorescence. Blue excitation was used throughout. All images were taken using a 4· objective lens and an exposure time of 145 or 193 ms unless statedotherwise. (A) G. buxifolia. Control: chalazal end of seed (269-ms exposure). Weak yellow or yellow–green autofluorescence occurred in the cotyledons (C)and central elaiosome cells (EO), but not in the hypostase (H) or columnar exotestal cells (CE). Palisade endotesta (PE). Scale bar = 300mm. (B)G. buxifolia.Treated: lower edge of elaiosome showing fluorescence attributed to LY penetration into the columnar exotestal (CE) layer and partly into the adjacentmesotestal region (ME). The probe did not penetrate or pass the palisade endotestal layer (PE) and there was no probe within the cotyledon (C). Scale bar =300mm. (C) G. buxifolia. Treated: upper edge of the elaiosome of seed seen in (B). LY fluorescence occurred between the squat exotestal (SE) cells but notbetween the cells of the adjacent mesotesta (ME). Inset shows enlargement of several squat exotestal cells. The probe was not detected in the elaiosome (EO)tissue, hypostase (H) or cotyledon (C). Scale bar = 300mm. (D) G. buxifolia. Treated: apical end of the elaiosome (EO) of seed in (B) showing the LYfluorescence in the exotesta (E) but not in the fatty elaiosome cells (EO). Scale bar = 300mm. (E) G. buxifolia. Control: micropylar end of the seed showinglimited yellow autofluorescence in the radicle (R) and lower autofluorescence in the palisade endotestal (PE), columnar exotestal cells (CE) and micropylarpalisade endotesta (MP). Note the slightly stronger autofluorescence of the non-sclerenchymatous tegmen cells at the end of the seed (*). Scale bar = 300mm.(F) G. buxifolia. Treated: micropylar end of the seed. LY fluorescence occurred in the columnar exotesta (CE) layer and but not in the adjacent mesotesta(ME). Inset shows enlargement of columnar exotestal cells and the accumulation of LY along the radial walls. Scale bar = 300mm. (G) G. buxifolia. Treated:same seed as in (F). LY fluorescence occurred between the squat exotestal cells and in the radicle (R) tissue. No probe was apparent within the mesotesta,palisade endotesta (PE) ormicropylar palisade endotesta (MP). Scale bar= 300mm. (H)G. buxifolia. Treated:micropylar end of seed depicted in (B)–(D). LYfluorescence occurred in the columnar exotesta (CE), where it was located in thewalls, and themesotesta (ME) cell layers. Themicropylar palisade endotesta(MP) and the palisade endotesta (PE) surrounding the embryo were only autofluorescent. Scale bar = 300mm. (I) G. buxifolia. Treated: lower side of the seedfrom (F) and (G) showing LY fluorescence in the columnar exotesta (CE) and in the mesotesta. The cotyledon (C) also shows LY fluorescence. Scale bar =100mm. (J) G. linearifolia. Control: chalazal end of seed showing autofluorescence of elaiosome cells (EO), hypostase (H) and adjacent tissues. Scale bar =300mm. (K) G. linearifolia. Treated: chalazal end showing LY fluorescence within the squat exotesta (SE). No trace of the probe was found in the elaiosome(EO), hypostase (H) or cotyledons (C). Scale bar = 300mm. (L) G. linearifolia. Control: micropylar end of the seed showing low autofluorescence of theradicle (R), palisade endotesta (PE), columnar exotesta (CE), squat exotesta (SE) and micropylar palisade endotesta (MP). Note the slightly strongerautofluorescence of the cells at the end of the seed (*). Scale bar = 300mm. (M)G. linearifolia. Treated: LY fluorescence occurred between the squat (SE) andcolumnar (CE) exotestal cells and the cells at the tip of the micropyle. There was no probe though in the micropyle or in the radicle end (R) of the embryo orabutting tissues. Scale bar = 300mm. (N) G. sericea. Control: chalazal end of seed. The central oil-filled cells of the elaiosome (EO) were stronglyautofluorescent yellow but the adjacent cell layers were not. The cotyledon tissue (C) had weak green autofluorescence. The squat exotesta (SE), elaiosomepalisade endotesta (PE) and hypostase (H) cells had low or no autofluorescence. Scale bar = 300mm. (O) G. sericea. Treated: LY fluorescence occurredthroughout the elaiosome down to the hypostase. The hypostase (H) and elaiosome palisade endotesta (PE) were negative. Squat exotestal (SE) cells weresurrounded by the probe but not the columnar exotestal cells (not shown). Scale bar = 300mm. (P) G. sericea. Control: micropylar end of the seed.Autofluorescencewas yellowishgreen in the radicle (R) and olive green in the cells in the squat exotesta (SE), columnar exotesta (CE) andmicropylar palisadeendotesta (MP) layers, as well as in cells at the end of themicropyle (*). Scale bar = 300mm. (Q)G. sericea. Treated: LY occurred between the squat exotestal(SE) and columnar exotestal (CE) cells as well as between the autofluorescent cells at themicropylar end of the seed (*). The remaining tissues showed no LYfluorescence. (R) radicle, (P) palisade, (ME)mesotesta. Scale bar = 300mm. (R)G. buxifolia. Control: lower side of seed showing limited autofluorescence ofthe columnar exotestal cells (CE), no autofluorescence of the adjacent mesotestal (ME) or palisade endotestal (PE) cells, and weak green fluorescence in thecotyledon (C). Scale bar = 100mm. (S) G. linearifolia. Treated: lower side of seed. LY occurred between the columnar exotestal (CE) cells and also occurredbetween the adjacent mesotestal cells (ME). The cotyledon (C) was autofluorescent. Scale bar = 100mm. (T) G. sericea. Treated: lower side of seed. LYoccurred between the columnar exotestal (CE) but not between the adjacentmesotestal (ME) cells. The cotyledon (C)wasweakly autofluorescent green. Scalebar=100mm. (U)G. sericea. Control: upper side of seed.No autofluorescencewas detectable in the squat exotesta (SE),mesotesta (ME) or palisade endotesta.Scale bar = 100mm. (V)G. sericea. Treated: upper side of seed. LY occurred between the squat exotestal (SE) and somemesotestal (ME) cells. The cotyledon
(C) showed only olive green autofluorescence. Scale bar = 100mm.
Briggs et al. — Histochemistry of Seed Coat Structure in Grevillea Species 977
Dow
nloaded from https://academ
ic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022

internal cuticle that separated the seed coat fromthe endosperm. The inner cuticle of dormant seeds ofE. penduliflora had pores that allowed movement ofwater into and out of the endosperm, but not of largermolecules such as Lucifer Yellow CH (molecular mass457Da). However, after treatment with smoke, the perme-ability of the inner cuticle increased and LY penetrated intothe endosperm and embryo, apparently due to an increase inthe number and diameter of the cuticular pores (Egerton-Warbuton, 1998). A sucrose ester that inhibited germinationwas subsequently identified from seeds of E. penduliflora;smoke thus broke dormancy by allowing the inhibitor toescape the seed (Egerton-Warburon and Ghisalberti, 2001).
Our study with LY on untreated Grevillea seeds identifiedseveral barriers to diffusion of high-molecular-weight com-pounds. In the three species, although the external cuticlewas no barrier to the movement of LY, there was a majorbarrier located at the base of the exotesta (i.e. the basalportion of the radial walls and the inner tangential walls)that effectively prevented the inward flow of large mole-cules. The barrier effect of the exotestal cells to LY mayhave been due to suberization and/or incorporation of phe-nols into the walls. The presence of suberin in the exotestalcell walls of these three Grevillea species was demonstratedfollowing staining with Berberine hemi-sulphate and con-firmed by acid hydrolysis followed by staining with SudanIV. Berberine hemi-sulphate is an acid salt of the alkaloidberberine, which has been used supposedly to confirm thepresence of suberin in plant roots (Barnabas, 1996). How-ever, as a cationic dye it will also bind to polyanions and sowill bind to phenolic compounds.
Another barrier to diffusion of high-molecular-weightcompounds was identified at the endotesta because althoughLY was able to penetrate through the central elaiosomeregion in G. sericea it did not spread laterally into theabutting endotesta. The endotesta thus acted as a secondbarrier to the inward/outward movement of large moleculessuch as germination inhibitors. This situation was paralleledat the micropylar end, where LY penetrated the short dis-tance through the thin-walled micropylar cells but wasstopped at the endotesta. Hence, if the exotestal layerbecomes disrupted due to wear and tear, the endotestamay still impose dormancy by regulating movement ofgermination inhibitors present in embryo or seed coat.The palisade layer in Glycine seeds has been identifiedas the site of the barrier to diffusion (of water; Ma et al.,2004).
The endotesta layer is not continuous across thehypostase; however, Lucifer Yellow did not cross thehypostase to the embryo, indicating a barrier to diffusionin the hypostase. The suberin identified in the cell wallsof the innermost cell layers of the hypostase may act as abarrier to movement of inhibitors to and from the embryo.
The embryo would thus appear to be completely sur-rounded by an outer and an inner barrier to the diffusionof high-molecular-weight compounds. Whether these bar-riers play a role in dormancy, for example by restricting theexit of germination inhibitors from the embryo, and whethersmoke and/or heat shock modify this as in E. penduliflorastill remain to be tested.
Phenolic compounds—inhibitors of gas exchange
Phenolic compounds have been proposed to inhibit seedgermination (Bewley and Black, 1982; Egerton-Warburton,1998). The presence of phenolic compounds in the radialwalls of the columnar exotestal cells and in the innertangential walls and vacuoles of the mesotestal cells wasindicated by the autofluorescence and blue reaction withToluidine Blue. Phenolic compounds such as coumarin,chlorogenic acid, ferulic acid, and caffeic and sinapicacid (and their derivatives) are esterified to wall carbohy-drates (Pan et al., 2002) and can be found in the seed coatand other seed tissues in many species (Gubler and Ashford,1985). The strongly negative environment in all the walls ofthe squat exotestal cells and the outer half of the columnarexotestal walls is indicated by the metachromatic reaction ofToluidine Blue at pH 4�4, suggesting polyanions (O’Brienand McCully, 1981). (They are also likely to be present inother regions but were masked by the blue reaction forphenols.) These conclusions are supported by the positivereaction for polyanions with Ruthenium Red (Websterand Stone, 1994) and demonstration of acid residues ofthe phenolic compounds by Berberine hemi-sulphate.This would be expected to result in a very hydrated systemfollowing wetting, and it is upon wetting of the seed coatthat phenolic compounds undergo the oxidation that isreported to interfere with gas exchange between the embryoand the environment (Bewley and Black, 1994). The phe-nolic compounds identified in the exotestal cell walls couldthus potentially interfere with gas exchange.
The normal staining response of Toluidine Blue, pH 4�4,given by lignified walls was only found in cells formingthe expanded wing area of the endotesta, not elsewherein the endotesta in these seeds. (Toluidine Blue at pH 4�4produces a turquoise colour when it binds to lignin in thesecondary walls of xylem vessels). A turquoise colour onlyoccurred in some of the walls; most walls were yellow as incontrols of embedded material. However, when acidifiedphloroglucinol was used, the entire endotestal layer gavea positive response for lignin, albeit only in the radial andinner tangential walls; no other cell wall gave a positiveresult with this dye. Staining variability may be due tomasking of lignin by other phenolic compounds, but ismost likely to be caused by variability in access of thedyes to staining sites, a problem encountered particularlywith fresh material.
Mechanical restraint
Seeds of the Proteaceae are endotestal, i.e. the region ofchief mechanical restraint is the lignified (or unlignified)palisade cells of the endotesta and the adjacent fibroustegmen, as was found in the three Grevillea species inves-tigated here. Another potential region of mechanical restraincould be the exotestal cells due to deposition of phenolsand suberin in the cell walls.
The discontinuity of the exotesta and tegmen across thehypostase could be important if the seed coat is acting as amechanical restraint: it gives a point of structural differencein the seed coat, which could potentially be affected bygermination cues such as heat shock.
978 Briggs et al. — Histochemistry of Seed Coat Structure in Grevillea Species
Dow
nloaded from https://academ
ic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022

Unrolling of margins during hydration
The structural reinforcement of the sclerenchymatoustegmen and lignified endotesta also appears to be respon-sible for the unrolling of the recurved margins duringimbibition. When G. sericea sections were compared beforeand after hydration, the region between the recurved mar-gins doubled in width in the direction between the palisadeand the columnar epidermal cells and increased by 72% inthe direction between the margins themselves. No change inthe width of the seed coat was found in the rest of the seed.Expansion of the seed in the columnar exotestal and meso-testal cells would force the rolled margins outward. Thedeposition of phenolic compounds in the radial walls ofthe columnar exotestal and the endotestal cells would resultin a tangential expansion of these cells whilst the absence ofphenolic compounds in the walls of the mesophyll cellswould permit both tangential and radial expansion.
Nile Red staining of cell walls: a novelhistochemical result
Nile Red was used to confirm the presence of neutrallipids such as formed in the cuticles mentioned above.However, an unexpected emission colour, emerald green(500–550 nm), was observed when the walls of theexotestal, mesotestal and elaiosome endotestal cells stainedby Nile Red were excited by blue light. The normalemission range of Nile Red is c. 100 nm, i.e. from red(660–680 nm) to yellow (550–580 nm). The emeraldgreen emission colour represents an extra shift of at least30 nm, towards the blue end of the spectrum. Nile Red is anuncharged heterocyclic planar hydrophobic phenoxazinedye (Fig. 4) (Sackett and Wolf, 1987; Haugland, 2002).It exhibits solvatochromism, i.e. its emission maximumvaries depending on the hydrophobicity of its environment.In polar solvents it appears red but undergoes a blue shiftas the polarity of the solvent decreases. Because of itshydrophobic properties, Nile Red partitions not only intoneutral lipids but also into hydrophobic regions of lipo-proteins (Greenspan and Fowler, 1985), native proteins(Sackett and Wolf, 1987) and alpha-acid glycoprotein(Brown et al., 1995). The observed blue shift seen in thewalls of the exotestal and endotestal cells may be due tohigh hydrophobicity of the wall; it may also be due to anacid–base interaction between the dye’s ketone side groupand wall acids. If the ketone side group of the dye is pro-tonated, there will be a shift in the conjugation of the doublebonds. When the conjugation of double bonds is altered incyclic molecules there is spectral shift in absorbance andemission; for example, when cyclic b-diketones are placedinto alkaline solutions there is a change to the enolate form
with the formation of a single double bond in the ring.The formation of this double bond results in a 30-nm redshift in absorbance (UV illumination) (Silverstein et al.,1991). Therefore, when the double bond is removed orshifted from a conjugated system there should be a blueshift of 30 nm. To determine whether a blue shift occurs inNile Red following acidification, a simple test was carriedout with Nile Red and 10% acetic acid, and between NileRed and a crystal of pure phenol. Both the acid and thephenol caused strong blue shifts (purple with acetic acid anddeep blue with phenol) in the colour of the dye when viewedunder white light. These shifts were large enough to explainthe shift from orange to green observed in the Grevilleawalls and thus to support the acid–base hypothesis.
CONCLUSIONS
In summary, Grevillea linearifolia, G. buxifolia andG. sericea show almost identical staining reactions withthe dyes used. The slight differences may contribute tothe observed differences in dormancy of collected seed,and to differences in germination response to heat/smoketreatments (Kenny, 2000; Morris, 2000). Differencesbetween the three species were most marked in the hypo-stase, which may determine the dormancy and germinationcharacteristics of each species. These three Grevillea spe-cies possess potentially three of the five criteria outlinedby Bewley and Black (1982) for seed-coat imposed dorm-ancy: mechanical restraint, possible interference with gasexchange and possible prevention of the exit of inhibitorsfrom the embryo. One criterion that could not be determinedwas the supply of inhibitors to the embryo. The influenceof the internal cuticles on movement of large molecules, asshown for Emmenanthe penduliflora (Egerton-Warbuton,1998), could not be determined because of the barriersimposed on the diffusion of Lucifer Yellow by theepidermis and the palisade.
ACKNOWLEDGEMENT
We thank Dr Mark Williams, University of WesternSydney, Hawkesbury, for his assistance in explaining thecolour shift shown by Nile Red when applied to theexotestal and endotestal cell walls.
LITERATURE CITED
AuldTD,DenhamAJ. 1999.The role of ants andmammals in dispersal andpost-dispersal seed predation of the shrubs Grevillea (Proteaceae).Plant Ecology 144: 201–213.
Auld TD, Tozer M. 1995. Patterns in emergence of Acacia and Grevilleaseedlings after fire. Proceedings of the Linnean Society of New SouthWales 115: 5–15.
BarnabasAD.1994.Apoplastic andsymplasticpathways in leavesand rootsof the seagrass Halodule uninervis (Forssk.) Aschers. Aquatic Botany47: 155–174.
Barnabas AD. 1996.Casparian band-like structures in the root hypodermisof some aquatic angiosperms. Aquatic Botany 55: 217–225.
Beveridge T, Loubert E, Harrison JE. 2000. Simple measurement ofphenolic esters in plant cell walls. Food Research International 22:775–783.
O(H5C2)2N O
N
F I G . 4. Line diagram of the Nile Red molecule.
Briggs et al. — Histochemistry of Seed Coat Structure in Grevillea Species 979
Dow
nloaded from https://academ
ic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022

BewleyJD,BlackM.1982.Physiology and biochemistry of seeds in relationto germination. Volume 2. Viability, dormancy and environmentalcontrol. Berlin: Springer-Verlag.
Bewley JD,BlackM. 1994.Seeds: physiology of development and germina-tion, 2nd edn. New York: Plenum Press.
BroughP. 1933.The life-history ofGrevillea robusta (Cunn.).Proceedingsof the Linnean Society of New South Wales 58: 33–73.
BrownMB,Miller JN, Seare NJ. 1995.An investigation of the use of NileRed as a long-wavelength fluorescent probe for the study of a1-acidglycoprotein–drug interactions. Journal of Pharmaceutical andBiomedical Analysis 13: 1011–1017.
Corner EJH. 1976. The seeds of Dictyledons, Volume 1. Cambridge, UK:Cambridge University Press.
Edwards W, Whelan R. 1995. The size, distribution and germinationrequirements of the soil-stored seed-bank of Grevillea barklyana(Proteaceae). Australian Journal of Ecology 20: 548–555.
Egerton-Warburton L. 1998. A smoke-induced alteration of the sub-testacuticle in seeds of the post-fire recruiter, Emmenanthe pendulifloraBenth. (Hydrophyllaceae). Journal of Experimental Botany 49:1317–1327.
Egerton-Warburton L, Ghisalberti EL. 2001. Isolation and structuralidentification of a germination inhibitor in fire-recruiters from theCalifornia chaparral. Journal of Chemical Ecology 27: 371–382.
Greenspan P, Fowler SD. 1985. Spectrofluorometric studies of the lipidprobe, Nile Red. Journal of Lipid Research 26: 781–789.
Gubler F, Ashford AE. 1985. Release of ferulic acid esters from barleyaleurone I. Time course of gibberellic-acid-induced release fromisolated layers. Australian Journal of Plant Physiology 12: 297–305.
HarrisN,SpenceJ,OparkaKJ.1994.General and enzymehistochemistry.In: Harris N, Oparka KJ, eds.Plant cell biology: a practical approach.Oxford: IRL Press at Oxford University Press.
Haugland RP. 2002. Handbook of Fluorescent Probes and ResearchProducts, 9th edn. Eugene, OR: Molecular Probes Inc.
Kausik SB. 1938. Studies in the Proteaceae I. Cytology and floral morpho-logy of Grevillea robusta Cunn. Annals of Botany 2: 899–910.
Kausik SB. 1939. Studies in the Proteaceae III. Embryology of Grevilleabanksii R. Br. Annals of Botany 3: 815–824.
Kenny B. 2000. Influence of multiple fire-related germination cues on threeSydney Grevillea (Proteaceae) species. Austral Ecology 25: 664–669.
Lequeu J, Fauconnier M-L, Chammai A, Bronner R, Blee E. 2003.Formation of plant cuticle: evidence for the occurrence of theperoxygenase pathway. The Plant Journal 36: 155–165.
Ma F, Cholewa E, Mohamed T, Peterson CA, GijzenM. 2004. Cracks inthe palisade cuticle of soybean seed coats correlate with theirpermeability to water. Annals of Botany 94: 213–228.
Maheshwari P. 1950. An introduction to the embryology of angiosperms.New York: McGraw-Hill.
Morris EC. 2000.Germination response of seven east Australian Grevilleaspecies (Proteaceae) to smoke, heat exposure and scarification.Australian Journal of Botany 48: 179–189.
Morris EC, Tieu A, Dixon K. 2000. Seed coat dormancy in two species ofGrevillea (Proteaceae). Annals of Botany 86: 771–775.
O’Brien TP, McCully ME. 1981. The study of plant structure. Principlesand selected methods. Melbourne: Termarcarphi Pty Ltd.
Olde P. 1997. The Grevillea book. Australia: Kangaroo Press Ltd.Oparka KJ, Read ND. 1994. The use of fluorescent probes for studies of
living plant cells. In: Harris N, Oparka KJ, eds. Plant cell biology:a practical approach. Oxford: IRL Press at Oxford University Press.
Pan GX, Thomson CI, Leary GJ. 2002. UV–Vis spectroscopic character-istics of ferulic acid and related compounds. Journal of WoodChemistry and Technology 22: 137–146.
Sackett DL, Wolff J. 1987. Nile Red as a polarity-sensitive fluorescentprobe of hydrophobic protein surfaces. Analytical Biochemistry167: 228–234.
Silverstein RM, Clayton Bassler G, Morrill TC. 1991. Spectrometricidentification of organic compounds, 5th edn. New York: John Wileyand Sons.
Venkata Rao C. 1967. Studies in the Proteaceae VIII. Morphology, floralanatomy and embryology of Grevillea R. Br. Proceedings of theNational Institute of Sciences of India, Part B Biological Sciences33: 162–199.
Webster J, Stone BA. 1994. Isolation, histochemistry and monosaccharidecomposition of the walls of root hairs from Heterozostera tasmanica(Martens ex Aschers.) den Hartog. Aquatic Botany 47: 29–37.
980 Briggs et al. — Histochemistry of Seed Coat Structure in Grevillea Species
Dow
nloaded from https://academ
ic.oup.com/aob/article/96/6/965/216359 by guest on 20 January 2022


















