
Investigation of the influence of the juvenile hormone...
14
223 Research Article Turk J Zool 2011; 35(2): 223-236 © TÜBİTAK doi:10.3906/zoo-0902-24 Investigation of the influence of the juvenile hormone analogue fenoxycarb on major hemolymph proteins of the silkworm Bombyx mori during the last larval instar Ramazan URANLI*, Ebru GÖNCÜ, Osman PARLAK Department of Biology, Faculty of Science, Ege University, 35100 Bornova, İzmir - TURKEY Received: 25.02.2009 Abstract: Fenoxycarb 0-ethyl N-(2-(4-pheoxyphenoxy)-ethyl) carbamate is the most potent juvenile hormone analogue (JHA) against a variety of insect species including the silkworm Bombyx mori. In this study, we aimed to demonstrate the effects of this juvenile hormone on major hemolymph proteins of Bombyx mori during the fiſth larval instar, by using fenoxycarb and clarifying the interactions between these proteins and the juvenile hormone. Topical applications of fenoxycarb to the fiſth instar larvae of Bombyx mori were performed immediately aſter the fourth ecdysis (on day 0) and at the beginning of the prepupal period (on day 6). Fenoxycarb application on day 0 increased the length of instar in both sexes and induced high total protein content in the hemolymph. When fenoxycarb application was performed on day 6, there was no difference in the length of larval instar and a similar total protein amount was reached with control. Detected hemolymph proteins are grouped as follows by using sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS- PAGE): storage proteins (72 kDa-76 kDa), 30kDa proteins, lipophorin (230 kDa), and vitellogenins (178 kDa and 42 kDa). Our results suggest that major hemolymph proteins have different sensitivities to fenoxycarb and that application days and sex are more important factors to get response. Key words: 30K proteins, Bombyx mori, fenoxycarb, hemolymph proteins, lipophorins, storage proteins, vitellogenins İpekböceği Bombyx mori’de, son larval evre süresince juvenil hormon analoğu fenoksikarb’ın temel hemolenf proteinleri üzerine etkisinin araştırılması Özet: Fenoksikarb 0-ethyl N-(2-(4-pheoxyphenoxy)-ethyl) karbamat, ipekböceği Bombyx mori’yi de içeren farklı böcek türlerinde en etkili juvenil hormon analoğudur. Bu çalışmada, fenoksikarb kullanılarak ipekböceği Bombyx mori’de beşinci larval evre süresince temel hemolenf proteinleri üzerine juvenil hormonun etkisi ve bu proteinlerle juvenil hormon arasındaki bağlantının aydınlatılması amaçlanmıştır. Bombyx mori’de 5. larval evrede topikal fenoksikarb uygulamaları 4. larval deri değişiminden hemen sonra (0.gün) ve pupa öncesi periyodun başlangıcında (6.gün) gerçekleştirilmiştir. 0 günde fenoksikarb uygulaması her iki eşeyde de 5. larval evreyi uzatmış ve total hemolenf protein içeriğinin artışını uyarmıştır. Altıncı gün fenoksikarb uygulamaları sonrasında larval evrenin süresinde herhangi bir farklılık olmamıştır. Elektroforetik olarak belirlenen temel hemolenf proteinleri şu şekilde gruplandırılmıştır: 30K proteinleri, depo proteinleri (72 kDa, 76 kDa), vitellogenin (178 kDa, 42 kDa), lipoforin (230 kDa). Çalışmamızın sonucunda temel hemolenf proteinlerinin fenoksikarb’a hassasiyetlerinin farklı olduğu; uygulama gününün ve eşey faktörünün önemli olduğu sonucuna varılmıştır. Anahtar sözcükler: 30K proteinleri, Bombyx mori, depo proteinleri, fenoksikarb, hemolenf proteinleri, lipoforinler; vitellogenin * E-mail: [email protected]
-
Upload
truongnguyet -
Category
Documents
-
view
217 -
download
4
Transcript of Investigation of the influence of the juvenile hormone...

223
Research Article
Turk J Zool2011; 35(2): 223-236 © TÜBİTAKdoi:10.3906/zoo-0902-24
Investigation of the influence of the juvenile hormone analoguefenoxycarb on major hemolymph proteins of the silkworm
Bombyx mori during the last larval instar
Ramazan URANLI*, Ebru GÖNCÜ, Osman PARLAKDepartment of Biology, Faculty of Science, Ege University, 35100 Bornova, İzmir - TURKEY
Received: 25.02.2009
Abstract: Fenoxycarb 0-ethyl N-(2-(4-pheoxyphenoxy)-ethyl) carbamate is the most potent juvenile hormone analogue(JHA) against a variety of insect species including the silkworm Bombyx mori. In this study, we aimed to demonstratethe effects of this juvenile hormone on major hemolymph proteins of Bombyx mori during the fifth larval instar, by usingfenoxycarb and clarifying the interactions between these proteins and the juvenile hormone. Topical applications offenoxycarb to the fifth instar larvae of Bombyx mori were performed immediately after the fourth ecdysis (on day 0) andat the beginning of the prepupal period (on day 6). Fenoxycarb application on day 0 increased the length of instar in bothsexes and induced high total protein content in the hemolymph. When fenoxycarb application was performed on day 6,there was no difference in the length of larval instar and a similar total protein amount was reached with control. Detectedhemolymph proteins are grouped as follows by using sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE): storage proteins (72 kDa-76 kDa), 30kDa proteins, lipophorin (230 kDa), and vitellogenins (178 kDa and 42kDa). Our results suggest that major hemolymph proteins have different sensitivities to fenoxycarb and that applicationdays and sex are more important factors to get response.
Key words: 30K proteins, Bombyx mori, fenoxycarb, hemolymph proteins, lipophorins, storage proteins, vitellogenins
İpekböceği Bombyx mori’de, son larval evre süresince juvenil hormon analoğufenoksikarb’ın temel hemolenf proteinleri üzerine etkisinin araştırılması
Özet: Fenoksikarb 0-ethyl N-(2-(4-pheoxyphenoxy)-ethyl) karbamat, ipekböceği Bombyx mori’yi de içeren farklı böcektürlerinde en etkili juvenil hormon analoğudur. Bu çalışmada, fenoksikarb kullanılarak ipekböceği Bombyx mori’debeşinci larval evre süresince temel hemolenf proteinleri üzerine juvenil hormonun etkisi ve bu proteinlerle juvenil hormonarasındaki bağlantının aydınlatılması amaçlanmıştır. Bombyx mori’de 5. larval evrede topikal fenoksikarb uygulamaları4. larval deri değişiminden hemen sonra (0.gün) ve pupa öncesi periyodun başlangıcında (6.gün) gerçekleştirilmiştir. 0günde fenoksikarb uygulaması her iki eşeyde de 5. larval evreyi uzatmış ve total hemolenf protein içeriğinin artışınıuyarmıştır. Altıncı gün fenoksikarb uygulamaları sonrasında larval evrenin süresinde herhangi bir farklılık olmamıştır.Elektroforetik olarak belirlenen temel hemolenf proteinleri şu şekilde gruplandırılmıştır: 30K proteinleri, depo proteinleri(72 kDa, 76 kDa), vitellogenin (178 kDa, 42 kDa), lipoforin (230 kDa). Çalışmamızın sonucunda temel hemolenfproteinlerinin fenoksikarb’a hassasiyetlerinin farklı olduğu; uygulama gününün ve eşey faktörünün önemli olduğusonucuna varılmıştır.
Anahtar sözcükler: 30K proteinleri, Bombyx mori, depo proteinleri, fenoksikarb, hemolenf proteinleri, lipoforinler;vitellogenin
* E-mail: [email protected]

IntroductionThe juvenile hormone (JH) and the 20-
hydroxyecdysone (20E) regulate a large number ofprocesses on insects, mostly metamorphosis andreproductive maturation, but also behavior,morphology, diapause, stress resistance, immunefunction, and aging (Nijhout, 1994; Dingle andWinchell, 1997; Flatt et al., 2005; Flatt et al., 2006).Quantities of JH and 20E titers have fundamentalimportance for the development of insects (Dedosand Fugo, 1996). During molting and metamorphosis,JH determines the characteristics of ecdysis: releasingof 20E in the presence of JH triggers the larval-larvalmolts, but in its absence it causes larval-pupal andpupal-adult transformations (Riddiford, 1985).
Fenoxycarb is a potent juvenile hormone analogue(JHA) against a variety of insect species including asilkworm, B. mori (Kamimura and Kiuchi, 1998).Application of the juvenile hormone (JH) or JHA inearly larval instar can induce prolongation of thelarval stage and inhibit the initiation of larval-pupaldifferentiation (Sakurai and Imokowa, 1988). Duringthe development of holometabolous insects, processessuch as the destruction of certain larval tissues,rejuvenation, and remolding of various tissues intoadult ones lead to the synthesis and utilization ofvarious macromolecules (Hyrsl and Simek, 2005)including the proteins. Hemolymph proteins aresynthesized by fat body cells and then secreted intothe hemolymph in a time-dependent situation duringpost embryonic development and metamorphosis(Tojo et al., 1980; Tomino, 1985; Kishimoto et al.,1999). Several classes of abundant insect hemolymphproteins have been identified of which productionsare regulated by hormones (Tojo et al., 1980;Goodman et al., 1985; Cole et al., 1990). Theabundant hemolymph proteins of B. mori arelipophorins, female specific protein vitellogenin,storage proteins, 30K proteins, and someundetermined proteins (Gamo, 1978; Tojo et al., 1980;Tomino, 1985; Hyrsl and Simek, 2005).
30K proteins are major hemolymph proteins thatinclude the storage proteins (Kishimoto et al., 1999).In silkworms, the 30K proteins are barely detecteduntil the later stage of the fourth instar larvae (Gamo,1978; Izumi et al., 1984). At the fifth instar, their
amounts increase sharply and they are concentratedin fat bodies (Ogawa et al., 2005). Physiologicalfunctions of 30K proteins are mostly still unknown.Previous studies suggest that some of them may beinvolved in inhibiting apoptosis (Kim et al., 2003; Parket al., 2003) and/or play a role in self-defense (Ujita etal., 2005). Insect hexamerins called storage proteins,circulate as oligomers of approximately 450 kDa andtheir subunits of about 75-80 kDa (Telfer and Kunkel,1991; Terwilliger, 1999). Silkworms have 2 kinds ofstorage protein (SP): SP1, which is methionine rich,and SP2, which is arylphorin. Storage proteins existin hemolymph from 1st to 5th instar of B. mori larvae(Nagata and Kobayashi, 1990). Just before pupation,storage proteins are taken up by fat body and used toform new structures such as somatic tissues andfemale reproductive products (Nagata and Kobayashi,1990; Terwilliger, 1999). Vitellogenin is a tetramerwith molecular weight of 440 kDa, composed of eachtwo molecules of non-identical subunits termed‘heavy chain’ and ‘light chain’ (Izumi et al., 1980). InB. mori, vitellogenin is synthesized by the fat bodycells of females at larval-pupal ecdysis and releasedinto the hemolymph before being taken up by thedeveloping oocytes (Yano et al., 1994). Lipophorin(Lp) of B. mori has a native molecular weight of 730kDa and consists of ApoLp-I and ApoLp-II withmolecular weights of respectively 250 kDa and 90 kDa(Kim and Kim, 1994).
Hemolymph protein profiles of B. mori (Hyrsland Simek, 2005) and their expression rate in the fatbody have been studied in detail (Izumi et al., 1984;Bosquet et al., 1989; Kishimoto et al., 1999), but thereis no report of the effects of the juvenile hormone onhemolymph proteins. Therefore, in this study, wetopically treated a juvenile hormone analogue (JHA)fenoxycarb just after fourth larval ecdysis (day 0) andafter hemolymph first peak of 20E (day 6) of fifthinstar larvae of silkworm, B. mori. We compared thecontrol and fenoxycarb application groups of majorhemolymph proteins in males and females, usingBradford assay and sodium dodecyl sulfatepolyacrylamide gel electrophoresis (SDS-PAGE). Weinvestigated repressive and stimulant effects offenoxycarb on hemolymph protein profiles in atime-, dose-, and sex-dependent manner.
Investigation of the influence of the juvenile hormone analogue fenoxycarb on major hemolymph proteins of the silkwormBombyx mori during the last larval instar
224

Materials and methodsInsects and fenoxycarb treatmentB. mori larvae were used for this experiment.
Hybrid B. mori (white monovoltin Japan × Chinesehybrid) eggs were provided from Bursa CocoonAgriculture Sales Cooperatives Association(Kozabirlik). Insects were reared at 24 ± 2 °Ctemperature, under 65%-85% relative humidity, andin natural photoperiodic conditions at the silkwormculture laboratory, and fed with fresh mulberry leaves.
Fenoxycarb, supplied by Riedel-de Haën, wasdissolved in acetone and the diluted solutions werestored at −20 °C. Then 1 ng and 10 ng of fenoxycarbdissolved in 10 μL of acetone was treated topicallywith a micropipette on day 0 and day 6 of the fifthinstar larvae. Each experiment group contained 150of larvae.
Hemolymph collectionHemolymphs of insects were collected every 24 h
from day 0 until pupation. Ten larvae were used forhemolymph collection each day. The insects’ first pairprolegs were cut by scissors and the hemolymph wascollected into Eppendorf tubes that were kept in ice.To prevent melanization we added 75 μL of 2 mMphenylthiourea solution to each tube. Samples werecentrifuged at 14,000 × g for 15 min and thesupernatant was stored at -80 °C in a deep freeze.
Bradford protein assayBovine serum albumin (Pierce) was used as
protein standard. Various concentrations of standardand 30 μL of diluted hemolymph were reacted with1000 μL of Bradford reagent (Sigma) in polystyrenecuvettes. Each sample and standard absorbance wasmeasured as 595 nm with a spectrophotometer(Unicam heλios α V 1.30).
ElectrophoresisSodium dodecyl sulfate polyacrylamide gel
electrophoresis (SDS-PAGE) was performed with aBiorad protean II xi Cell electrophoresis apparatus in adiscontinuous buffer system according to Laemmli(1970). All solutions, buffers, and gels were preparedaccording to apparatus instruction manuals. We used10% separating and 4% stacking gel. Five micrograms ofprotein obtained from each sample was placed in eachwell and samples were run at 100 V overnight. Bio-rad
marker was used for calibration. Gels were stained withsilver nitrate. Stained gels were analyzed by using imageJ analysis software from the National Institutes ofHealth, US (http://rsb.info.nih.gov/nih-image/).
Results and discussionIn insects, JH exhibited a regulatory effect on
protein synthesis. JH and 20E can suppress or induceprotein synthesis independently or antagonistically(Hiruma et al., 1999; Zhou and Riddiford, 2002;Dubrovsky, 2005; Fang et al., 2005; Flatt et al., 2005).However, these hormones can also interactsynergistically (Fang et al., 2005) and it can be moreeffective to induce protein synthesis (Flatt et al., 2006).At the fifth instar larvae of Bombyx mori, 20E releaseshemolymph from the prothoracic gland on day 3, day6, and day 9 (Sakurai and Imokowa, 1988). JH issecreted during the fourth ecdysis and thendisappears from the hemolymph during early days ofthe fifth instar. Following this, JH titer increasesgradually from day 5 until pupation (Niimi andSakurai, 1997).
The effect of fenoxycarb during last larval instar inBombyx mori has been studied in detail (Kamimura,1995; Leonardi et al., 1996; Kamimura and Kiuchi,1998; Dedos and Fugo, 1999; Monconduit andMauchamp, 1999). Application of fenoxycarb in thelast larval instar can induce a prolongation of thelarval stage and inhibit larval-pupal differentiation ina dose- and application-time-dependent manner(Kamimura and Kiuchi, 1998). In our study, the final(fifth) larval instar stadium lasts 11 days: 8 days forfeeding, followed by 3 days of spinning to build thecocoon in the control groups. Larvae started theirspinning behavior on day 8 and 97% of larvae showedlarval-pupal ecdysis on day 11. When fenoxycarb wastreated immediately after the fourth larval ecdysis(day 0), spinning and pupation were delayed. Thelength of the stage was prolonged for about 4 days;spinning was seen on day 11 and 92% of larvaeshowed spinning behavior but only 38% pupated. Thelength of the fifth larval instar and cell deathprogression in the day 6 treatment group were similarto those in the controls. Almost all larvae showedspinning behavior and pupation occurred on day 10but when compared with the control groups; far morelarvae died during the larval-pupal ecdysis.
R. URANLI, E. GÖNCÜ, O. PARLAK
225

The B. mori hemolymph contains many differentproteins. They are usually classified according to theirfunctions but their functions have not beencompletely clarified yet. In this study, the majorhemolymph proteins are grouped as follows: storageproteins (72 kDa-76 kDa), 30kDa proteins, lipophorin(230 kDa), and vitellogenins (178 kDa and 42 kDa).Effects of fenoxycarb on total protein content ofhemolymph and relative abundance of major proteinswere analyzed. Day 0 fenoxycarb applications (1 ngand 10 ng) prolonged the larval period and pupationdid not occur in the 10 ng treated group, unlike thecontrols. Monconduit and Mauchamp (1998)reported that fenoxycarb treatments on day 0 suppressthe rise of ecdysteroid titer but Kamimura and Kiuchi(1998) demonstrated that prothoracic glands improvethe ecdysone secretion ability on day 10 of the fifthinstar. Different studies showed similar results; day 0application of JHAs prolonged the larval period ofsilkworm in a dose dependent manner (Kamimuraand Kiuchi, 1998; Monconduit and Mauchamp, 1998;Dedos et al., 2002). In the day 6 application groups (1ng and 10 ng), fenoxycarb did not affect the length ofthe fifth instar unlike day 0 applications. The onlyeffect is that larvae pupated 1 day earlier comparedwith the control groups.
It has been reported that the total protein contentof hemolymph increases during the larvaldevelopment in some insect species. Total proteinconcentrations in Drosophila (Chen, 1956) and Samiacynthia (Laufer, 1960) were low at early stages butincreased just before puparium formation. In ourstudy, control groups showed similar results. Theprotein concentrations in the control hemolymphremained low at the beginning of the final instar andthen increased slowly until day 8 in both sexes. A peakoccurred on day 9, just before pupation,-andfollowing this peak the protein content of hemolymphdecreased gradually (Figure 1a). Endocrineconditions during the fifth larval stage differ fromthose of other larval instars and are important forlarval-pupal metamorphosis. The ecdysone levelsduring the first 3 days of the last instar areundetectable, but juvenile hormone is still found inthe hemolymph (Sakurai et al., 1998). Disappearanceof juvenile hormone on day 3 is required for recoveryof the prothoracic gland activity for secretionecdysone. Both ecdysone and juvenile hormone can
be detected in the hemolymph (Sakurai et al., 1989)after day 5 of the fifth instar. It seems that the lowprotein content of the hemolymph results fromecdysone depletion during first 3 days andcombination of ecdysone and juvenile hormonestimulates the total protein level in the hemolymphafter day 5. The total hemolymph protein amount washigher in females than in males during the fifth larvalinstar. The yolk protein synthesis from the fat body offemales may be the reason for this result (Figure 1a).
When fenoxycarb was applied on day 0, the levelsof protein content of both sexes were observed to bevery similar to those of the controls for the first 4 days,independent of the application dose. After day 5,protein level began to increase gradually on day 0 inthe treatment groups when compared with thecontrols, but the day 9 peak in the controls was notseen in these experiment groups (Figure 1b for male,1c for female). These results may be related to theinhibition effect of the fenoxycarb on the ecdysonerelease from the prothoracic gland. Kamimura andKiuchi (1998) reported that fenoxycarb application onday 0 suppresses 20E secretion until day 12. It hasbeen shown that a juvenile hormone analoguemethoprene application to the fourth instar larvae ofB. mori causes the increase of the hemolymph proteinconcentrations and this result arises due to preventingsequestration of the storage proteins by the fat body(Kajiura and Yamashita, 1992).
Fenoxycarb application (1-10 ng) on males on day 6caused lower protein level compared to the controls, butit began to increase on day 8 and did not decrease afterday 9, unlike in the controls (Figure 1d). The maximumhemolymph protein content was determined on day 9in both the control and day 6 experiment groups.Application of 1 ng of fenoxycarb to female on day 6caused a decline in protein content of hemolymph incontrast to 10 ng application (Figure 1e). When thehemolymph 20E and juvenile hormone levels areconsidered on this day, our results indicate that thesechanges in protein content may result from synergisticeffects of these 2 hormones (Fang et al., 2005).
During this study, 16-24 bands were determinedusing SDS-PAGE with silver staining the hemolymphproteins of control and experiment groups during thefifth instar in B. mori. No qualitative difference wasfound between males and females of the control groups
Investigation of the influence of the juvenile hormone analogue fenoxycarb on major hemolymph proteins of the silkwormBombyx mori during the last larval instar
226

from day 0 to day 10, except for the female specific 42-178 kDa determined molecular weight protein bands,which probably corresponds vitellogenin, determinedon day 11 (Figure 2). Moreover, 30 kDa protein bands,76-72 kDa protein bands, 230 kDa protein band, and42-178 kDa protein bands were mainly estimated in thelarval hemolymph of B. mori.
30K proteins are synthesized and secreted intohemolymph during the fourth instar but theirquantities are low up until the fifth instar (Gamo, 1978;Tojo et al., 1980; Tomino, 1985). At the beginning ofthe fifth instar, 30K proteins accumulate into thehemolymph and reach high amounts after thedisappearance of JH from hemolymph (Izumi et al.,
R. URANLI, E. GÖNCÜ, O. PARLAK
227
a b
c d
e
100
80
60
40
20
0
Control Male
Control Female
Control Female
0 1 2 3 4 5 6 7 8 9 10 11DaysH
emol
ymph
pro
tein
conc
entr
atio
n m
g/m
l
100
80
60
40
20
0
Hem
olym
ph p
rote
in co
ncen
trat
ion
mg/
ml
120
100
80
60
40
20
0
Hem
olym
ph p
rote
in co
ncen
trat
ion
mg/
ml
80
60
40
20
0
Control MaleDay 6-1 ngFenoxycarb maleDay 6-10 ngFenoxycarb male
Day 0-1 ngFenoxycarb femaleDay 0-10 ngFenoxycarb female
Control maleDay 0-1 ngFenoxycarb maleDay 0-10 ngFenoxycarb male
Control FemaleDay 6-1 ngFenoxycarb femaleDay 6-10 ngFenoxycarb male
0 2 4 6 8 10
6 7 8 9 10 11
Days
0 2 4 6 8 10Days Days
6 7 8 9 10 11Days
Hem
olym
ph p
rote
in co
ncen
trat
ion
mg/
ml
80
60
40
20
0
Hem
olym
ph p
rote
in co
ncen
trat
ion
mg/
ml
Figure 1. Hemolymph total protein concentrations of fifth instar larvae (mg/mL). (a) control male and female; (b) 1 ng and 10 ng offenoxycarb subjected male larvae on day 0; (c) 1 ng and 10 ng of fenoxycarb subjected female larvae on day 0; (d) 1 ng and 10ng of fenoxycarb subjected male larvae on day 6; (e) 1 ng and 10 ng of fenoxycarb subjected female larvae on day 6.
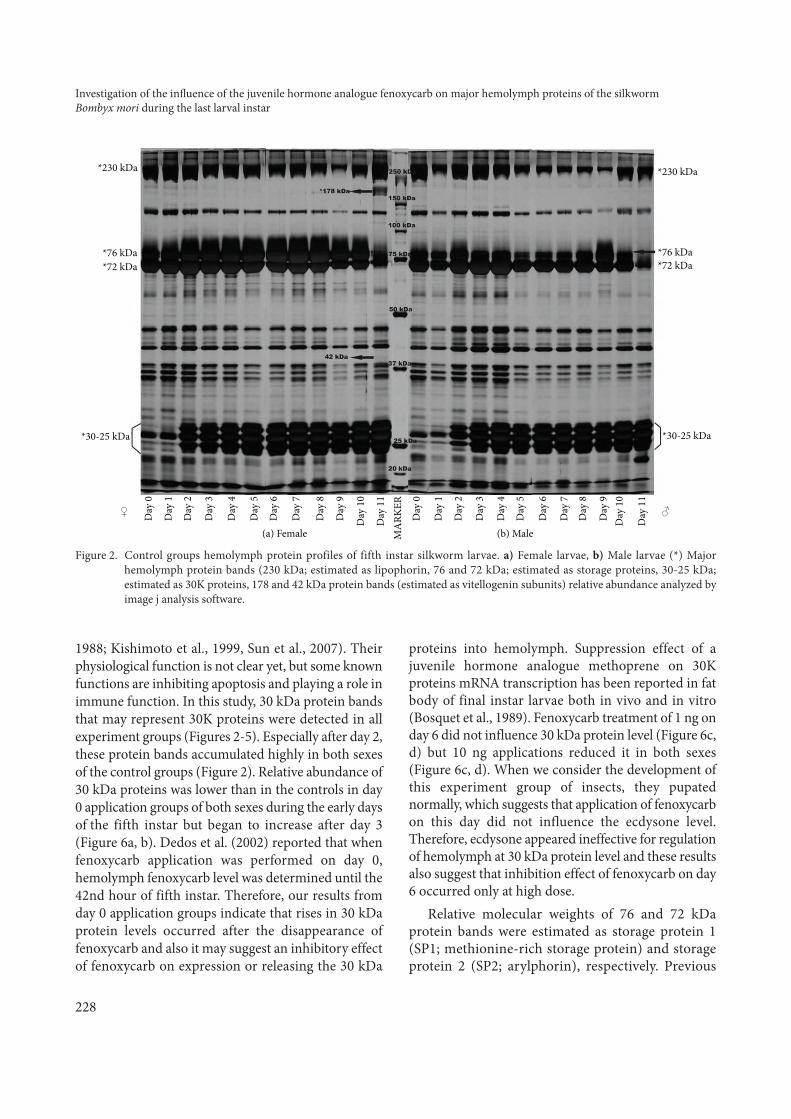
1988; Kishimoto et al., 1999, Sun et al., 2007). Theirphysiological function is not clear yet, but some knownfunctions are inhibiting apoptosis and playing a role inimmune function. In this study, 30 kDa protein bandsthat may represent 30K proteins were detected in allexperiment groups (Figures 2-5). Especially after day 2,these protein bands accumulated highly in both sexesof the control groups (Figure 2). Relative abundance of30 kDa proteins was lower than in the controls in day0 application groups of both sexes during the early daysof the fifth instar but began to increase after day 3(Figure 6a, b). Dedos et al. (2002) reported that whenfenoxycarb application was performed on day 0,hemolymph fenoxycarb level was determined until the42nd hour of fifth instar. Therefore, our results fromday 0 application groups indicate that rises in 30 kDaprotein levels occurred after the disappearance offenoxycarb and also it may suggest an inhibitory effectof fenoxycarb on expression or releasing the 30 kDa
proteins into hemolymph. Suppression effect of ajuvenile hormone analogue methoprene on 30Kproteins mRNA transcription has been reported in fatbody of final instar larvae both in vivo and in vitro(Bosquet et al., 1989). Fenoxycarb treatment of 1 ng onday 6 did not influence 30 kDa protein level (Figure 6c,d) but 10 ng applications reduced it in both sexes(Figure 6c, d). When we consider the development ofthis experiment group of insects, they pupatednormally, which suggests that application of fenoxycarbon this day did not influence the ecdysone level.Therefore, ecdysone appeared ineffective for regulationof hemolymph at 30 kDa protein level and these resultsalso suggest that inhibition effect of fenoxycarb on day6 occurred only at high dose.
Relative molecular weights of 76 and 72 kDaprotein bands were estimated as storage protein 1(SP1; methionine-rich storage protein) and storageprotein 2 (SP2; arylphorin), respectively. Previous
Investigation of the influence of the juvenile hormone analogue fenoxycarb on major hemolymph proteins of the silkwormBombyx mori during the last larval instar
228
*230 kDa
*76 kDa *76 kDa*72 kDa *72 kDa
*30-25 kDa *30-25 kDa
(a) Female (b) Male
♀ ♂Day
0
Day
1
Day
2
Day
3
Day
4
Day
5
Day
6
Day
7
Day
8
Day
9
Day
10
Day
11
MA
RKER
Day
0
Day
1
Day
2
Day
3
Day
4
Day
5
Day
6
Day
7
Day
8
Day
9D
ay 1
0
Day
11
*230 kDa
Figure 2. Control groups hemolymph protein profiles of fifth instar silkworm larvae. a) Female larvae, b) Male larvae (*) Majorhemolymph protein bands (230 kDa; estimated as lipophorin, 76 and 72 kDa; estimated as storage proteins, 30-25 kDa;estimated as 30K proteins, 178 and 42 kDa protein bands (estimated as vitellogenin subunits) relative abundance analyzed byimage j analysis software.

R. URANLI, E. GÖNCÜ, O. PARLAK
229
230 kDa
*76 kDa *76 kDa*72 kDa *72 kDa
*30-25 kDa *30-25 kDa
(a) Female (b) Male
0-1- ♀ 0-1 ♂Day
0
Day
1
Day
2
Day
3
Day
4
Day
5
Day
6
Day
7
Day
8
Day
9
Day
10
Day
11
MA
RKER
Day
0
Day
1
Day
2
Day
3
Day
4
Day
5
Day
6
Day
7
Day
8
Day
9D
ay 1
0
Day
11
*230 kDa
Figure 3. Female and male hemolymph protein profile after 1 ng of fenoxycarb applied on day 0 of the final instar larvae. a) Female, b)Male (*) Major hemolymph protein bands relative abundance analyzed by image j analysis software.
*230 kDa
*76 kDa *76 kDa*72 kDa *72 kDa
*30-25 kDa *30-25 kDa
(a) Female (b) Male
0-10- ♀ 0-10 ♂Day
0
Day
1
Day
2
Day
3
Day
4
Day
5
Day
6
Day
7
Day
8D
ay 9
Day
10
Day
11
MA
RKER
Day
0
Day
1
Day
2
Day
3
Day
4
Day
5
Day
6
Day
7
Day
8
Day
9
Day
10
Day
11
*230 kDa
Figure 4. Female and male hemolymph protein profile after 10 ng of fenoxycarb applied on day 0 of the final instar larvae. a) Female,b) male (*) Major hemolymph protein bands relative abundance analyzed by image j analysis software.
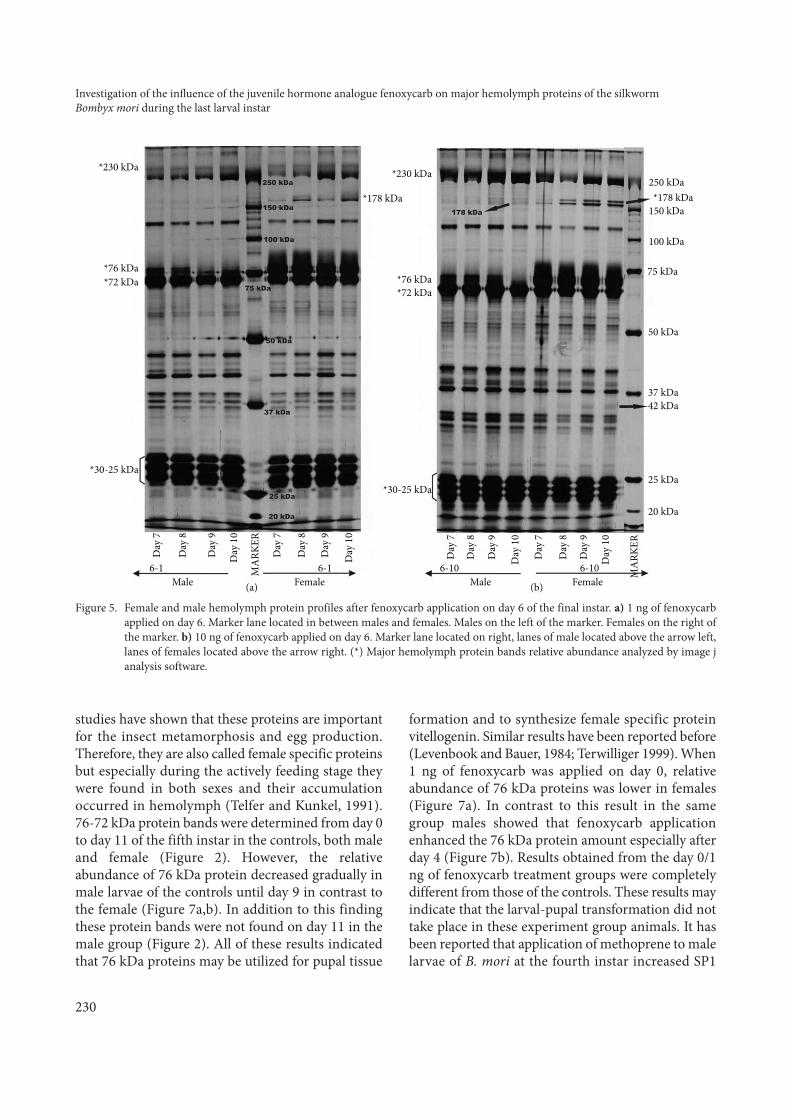
studies have shown that these proteins are importantfor the insect metamorphosis and egg production.Therefore, they are also called female specific proteinsbut especially during the actively feeding stage theywere found in both sexes and their accumulationoccurred in hemolymph (Telfer and Kunkel, 1991).76-72 kDa protein bands were determined from day 0to day 11 of the fifth instar in the controls, both maleand female (Figure 2). However, the relativeabundance of 76 kDa protein decreased gradually inmale larvae of the controls until day 9 in contrast tothe female (Figure 7a,b). In addition to this findingthese protein bands were not found on day 11 in themale group (Figure 2). All of these results indicatedthat 76 kDa proteins may be utilized for pupal tissue
formation and to synthesize female specific proteinvitellogenin. Similar results have been reported before(Levenbook and Bauer, 1984; Terwilliger 1999). When1 ng of fenoxycarb was applied on day 0, relativeabundance of 76 kDa proteins was lower in females(Figure 7a). In contrast to this result in the samegroup males showed that fenoxycarb applicationenhanced the 76 kDa protein amount especially afterday 4 (Figure 7b). Results obtained from the day 0/1ng of fenoxycarb treatment groups were completelydifferent from those of the controls. These results mayindicate that the larval-pupal transformation did nottake place in these experiment group animals. It hasbeen reported that application of methoprene to malelarvae of B. mori at the fourth instar increased SP1
Investigation of the influence of the juvenile hormone analogue fenoxycarb on major hemolymph proteins of the silkwormBombyx mori during the last larval instar
230
*230 kDa
*178 kDa *178 kDa
*76 kDa 75 kDa*72 kDa
50 kDa
37 kDa42 kDa
*76 kDa*72 kDa
*30-25 kDa25 kDa
20 kDa
*30-25 kDa
(a)Male Female (b)Male Female6-1 6-106-1 6-10
Day
7
Day
8
Day
9
Day
10
Day
7
Day
8
Day
9
Day
10
MA
RKER
MA
RKER
Day
7
Day
7
Day
8
Day
8
Day
9
Day
9
Day
10
Day
10
150 kDa
100 kDa
250 kDa*230 kDa
Figure 5. Female and male hemolymph protein profiles after fenoxycarb application on day 6 of the final instar. a) 1 ng of fenoxycarbapplied on day 6. Marker lane located in between males and females. Males on the left of the marker. Females on the right ofthe marker. b) 10 ng of fenoxycarb applied on day 6. Marker lane located on right, lanes of male located above the arrow left,lanes of females located above the arrow right. (*) Major hemolymph protein bands relative abundance analyzed by image janalysis software.

accumulation in the hemolymph (Kajiura andYamashita, 1992). Moreover, 10 ng of fenoxycarbapplication to females on day 0 did not causedifferences in 76 kDa protein level (Figure 7a) butrelatively high protein levels were found in the malegroup (Figure 7b). When fenoxycarb was applied onday 6, there were no differences in femalesindependent of dose (Figure 7c) and only changeswere observed in males on day 9 in which peak wasnot seen, unlike in the controls (Figure 7d).
The biological functions of arylophorins provideamino acids for the synthesis of structural andnutritional substances during metamorphosis ininsects. They also appear to be structural componentsof the sclerotized cuticle in Calliphora vicine and
Manduca sexta (Urich, 1994). Relative abundance of72 kDa in the female control group was high in theearly days of the fifth instar and small decreases weredetermined until day 8 and rising again during theprepupal period (Figure 8a). Similar results werefound in the 1 ng of fenoxycarb application on day 0but, in contrast, 10 ng of fenoxycarb enhanced 72kDaprotein amount in female hemolymph (Figure 8a).The relative abundance of 72kDa protein in the malecontrol group was estimated lower than that infemales and nearly the same level was determineduntil day 11 (Figure 8b). Juvenile hormone analoguefenoxycarb application on day 0 did not affectfluctuation of this protein during the experimentperiod independent of the applied dose.
R. URANLI, E. GÖNCÜ, O. PARLAK
231
a
c d
b
Figure 6. Relative abundance of 30-25 kDa proteins (estimated as 30K proteins) in experiment groups. Relative abundance of 30 kDaproteins in day 0 fenoxycarb application groups were lower in the early days of fifth instar in both a) female and b) male larvaeindependent of dose. c) 1 ng of fenoxycarb application to female larvae on day 6 enhanced 30K protein for the first 2 days, incontrast 10 ng application reduced protein abundance. d) Maximum 30K protein abundance was determined on day 9 in maleexperiment group and it was higher than control and similar to female, 10 ng of dose inhibited 30K protein in hemolymph.

Furthermore, 1 ng of fenoxycarb application onday 6 to females did not influence the 72 kDa proteinabundance when compared with the controls untilday 8, but the protein peak observed on day 9 was notdetermined in this experiment (Figure 8c) and arelatively high dose of fenoxycarb showed asuppressive effect in this female experiment group.Future studies about female reproductive ability ofthese experiment groups will help to clarify the resultsof the reducing effect of fenoxycarb. However,exposure of male larvae to fenoxycarb on day 6 of thefifth instar had no effect on 72 kDa protein abundancein hemolymph independent of the treatment dosewhen considered in the controls (Figure 8d).
Vitellogenins are lipoglycoproteins with about 7%-16% lipid and 1%-14% carbohydrate (Urich, 1994).
They are female specific proteins that have 440 kDamolecular weight (MW) and 2 subunits (MW: 178kDa and 42 kDa) (Izumi et al., 1980). This protein issynthesized in fat body and released in thehemolymph of the pupa (Janarthanan et al., 1998). Inour study, estimated molecular weights of 178 and 42kDa proteins were seen on day 11 of the final instar incontrol females (Figure 2). These protein bands wereconsidered vitellogenin protein subunits. Besides thisresult, 76 and 72 kDa protein bands’ density decreasedwith vitellogenin synthesis on day 11 of the fifth instar(Figure 2). It has been previously reported that, duringvitellogenin synthesis in B. mori, storage proteinslevels fall (Tojo et al., 1980; Ogawa and Tojo, 1981).Day 0 applications inhibited 178 and 42 kDa proteins(Figures 3 and 4). Ohnishi (1987) reported that
Investigation of the influence of the juvenile hormone analogue fenoxycarb on major hemolymph proteins of the silkwormBombyx mori during the last larval instar
232
a
c d
b
0
10
20
30
40
0 1 2 3 4 5 6 7 8 9 10 11
rela
tive
abun
danc
e (%
)
control
day 0 1 ng
day 0 10 ng
Figure 7. Relative abundance of 76 kDa protein (estimated as storage protein, SP1). a) 1 ng of fenoxycarb application to female on day0 caused lower SP but 10 ng of dose did not cause an alteration but both of doses maintained the protein at a high level on day11. b) Fenoxycarb applied males on day 0 showed high SP1 protein levels 4 days after treatment. c) Fenoxycarb application tofemale larvae on day 6 had no effect on SP1 level in contrast, d) notably low SP1 level were determined in male in controls andapplication groups.

vitellogenin expression in B. mori depends onecdysone. Therefore, the inhibitory effect offenoxycarb on vitellogenin synthesis in these groupswas probably related to metamorphic prevention byinhibition of ecdysone secretion from the prothoracicgland. Vitellogenin synthesis is induced or suppressedby juvenile hormone analogues in different insectspecies (Pinto et al., 2000; Comas et al., 2001). Incontrast with this result, 178 and 42 kDa proteinbands were detected in female larvae of day 6experiment groups after treatment, which occurredearlier than in the controls (Figure 9) and fenoxycarbtriggered overexpression of vitellogenin or earlyrelease from the fat body. In addition, weak
R. URANLI, E. GÖNCÜ, O. PARLAK
233
a
c d
b
0
10
20
30
0 1 2 3 4 5 6 7 8 9 10 11
rela
tive
abun
danc
e (%
)
control
day 0 1 ng
day 0 10 ng
Figure 8. Relative abundance of 72 kDa protein (estimated as storage protein 2, SP2). a) Abundance of 72 kDa protein in 1 ng offenoxycarb application on day 0 group females was approximately the same level with the control but 10 ng fenoxycarbapplication induced this protein level in hemolymph, which was higher than controls; b) there were no significant differencesbetween the control and day 0 fenoxycarb applied male larvae c) fenoxycarb application to female on day 6 did not make adifference in hemolymph SP2 protein d) fenoxycarb application to male on day 6 decreased SP2 protein in hemolymph, thebiggest differences between the control and experiment group were seen on day 10 in which larval-pupal ecdysis occurred inthe experiment groups.
Figure 9. Relative abundance of 178 kDa protein (estimatedvitellogenin subunit). Vitellogenin bands were firstlydetermined just before pupation (on day 11) in thecontrol but fenoxycarb application to female on day 6caused early presence of vitellogenin in hemolymph.
vitellogenin bands were also detected in males (Figure5). Fenoxycarb application on day 6 to male larvaecaused augmentation of female specific proteinamount like 76 kDa storage protein and vitellogenindepending on the application dose. Early reports inthe pupal period of Antherae yamamai suggest thatjuvenile hormone exerts a promoting effect onvitellogenin synthesis in males (Gang-yin et al., 2002).We found that the same effect of fenoxycarb in day 6fenoxycarb treated males.
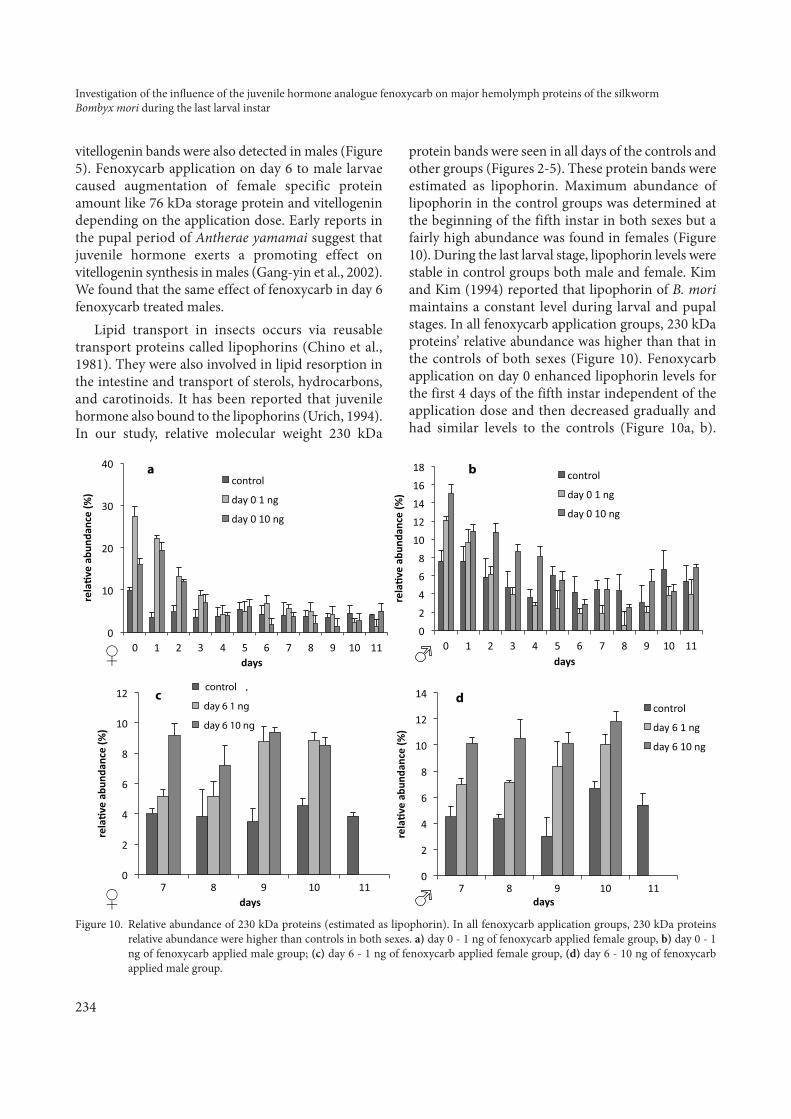
Lipid transport in insects occurs via reusabletransport proteins called lipophorins (Chino et al.,1981). They were also involved in lipid resorption inthe intestine and transport of sterols, hydrocarbons,and carotinoids. It has been reported that juvenilehormone also bound to the lipophorins (Urich, 1994).In our study, relative molecular weight 230 kDa
protein bands were seen in all days of the controls andother groups (Figures 2-5). These protein bands wereestimated as lipophorin. Maximum abundance oflipophorin in the control groups was determined atthe beginning of the fifth instar in both sexes but afairly high abundance was found in females (Figure10). During the last larval stage, lipophorin levels werestable in control groups both male and female. Kimand Kim (1994) reported that lipophorin of B. morimaintains a constant level during larval and pupalstages. In all fenoxycarb application groups, 230 kDaproteins’ relative abundance was higher than that inthe controls of both sexes (Figure 10). Fenoxycarbapplication on day 0 enhanced lipophorin levels forthe first 4 days of the fifth instar independent of theapplication dose and then decreased gradually andhad similar levels to the controls (Figure 10a, b).
Investigation of the influence of the juvenile hormone analogue fenoxycarb on major hemolymph proteins of the silkwormBombyx mori during the last larval instar
234
control
day 6 1 ng
day 6 10 ng
a
c d
b
Figure 10. Relative abundance of 230 kDa proteins (estimated as lipophorin). In all fenoxycarb application groups, 230 kDa proteinsrelative abundance were higher than controls in both sexes. a) day 0 - 1 ng of fenoxycarb applied female group, b) day 0 - 1ng of fenoxycarb applied male group; (c) day 6 - 1 ng of fenoxycarb applied female group, (d) day 6 - 10 ng of fenoxycarbapplied male group.

Moreover, 10 ng of fenoxycarb application on day 0seemed much more effective to induce lipophorinlevel (Figure 10a, b). Our results suggest thatfenoxycarb application stimulated 230 kDa proteinsynthesis after treatment and independent of theapplication days (Figure 10). Juvenile hormone hasbinding ability to the lipophorins and it may beimportant for carrying this hormone in thehemolymph (Urich, 1994). Therefore, lipophorin risesjust after the fenoxycarb treatment may be related tothis binding feature of juvenile hormone.
All of these results suggest that major hemolymphproteins have different sensitivities to the juvenilehormone analogue fenoxycarb. Application day andsex are more important for the response and theseresults may also be related to developmentalprogression of all experiment groups.
AcknowledgmentThe research was supported by Ege University
(2006 Science 047).
R. URANLI, E. GÖNCÜ, O. PARLAK
235
Bosquet, G., Fourche, J. and Guillet, C. 1989. Respective contributionsof juvenile hormone and 20-hydroxyecdysone to the regulation ofmajor haemolymph protein synthesis in Bombyx mori larvae.Journal of Insect Physiology, 35: 1005-1015.
Chen, P.S. 1956. Electrophoretische Bestimmung des Proteingehaltesitn Blut normaler und letaler Larven von Drosophilamelanogaster. Rev. Suisse Zool. 61: 437.
Chino, H., Downer, R.G.H., Wyatt, G.R. and Gilbert, L.I. 1981.Lipophorin, a major class of lipoproteins of insect hemolymph.Insect Biochem. 11: 491.
Cole, K.D., Smith, A.F. and Wells M.A. 1990. The structure of theapolipophorin-III. gene from Manduca sexta. Insect Biochem. 20:381-388.
Comas, D., Piulachs, M.D. and Belles, X. 2001. Induction of vitellogeningene transcription in vitro by juvenile hormone in Blattellagermanica. Molecular and Cellular Endocrinology. 183: 93-100.
Dedos, S.G. and Fugo, H. 1996. Effects of fenoxycarb on the secretoryactivity of the prothoracic glands in the fifth instar of thesilkworm, Bombyx mori. General and ComparativeEndocrinology 104: 213-224.
Dedos, S.G. and Fugo, H. 1999. Induction of dauer larvae by applicationof fenoxycarb early in the 5th instar of the silkworm, Bombyxmori. Journal of Insect Physiology 45: 769-775.
Dedos, S.G., Szurdoki, F., Szekacs, A., Shiotsuki, T., Hammock, B.D.,Shimada, J. and Fugo, H. 2002. Fenoxycarb levels and their effectson general and juvenile hormone esterase activity in thehemolymph of the silkworm, Bombyx mori. PesticideBiochemistry and Physiology 73: 174-187.
Dingle, H. and Winchell, R. 1997. Juvenile hormone as a mediator ofplasticity in insect life histories. Arch. Insect. Biochem. Physiol.35: 359-373.
Dubrovsky, E.B. 2005. Hormonal cross talk in insect development.Trends Endocrinol. Metab. 16: 6-11.
Fang, F., Xu, Y., Jones, D. and Jones, G. 2005. Interactions ofultraspiracle with ecdysone receptor in the transduction ofecdysone and juvenile hormone-signaling. FEBS J. 272: 1577-1589.
Flatt, T., Moroz, L.L., Tatar, M. and Heyland, A. 2006. Comparingthyroid and insect hormone signaling. Integrative andComparative Biology 46: 777–794.
Flatt, T., Tu, M.P. and Tatar, M. 2005. Hormonal pleiotropy and thejuvenile hormone regulation of Drosophila development and lifehistory. Bioassays. 27: 999-1010.
Gamo, T. 1978. Low molecular weight lipoproteins in the hemolymphof the silkworm, Bombyx mori: inheritance, isolation and someproperties. Insect Biochemistry 8: 457-470.
Gang-yin, Y., Cui, H. and He, G. 2002. Hormonal control of thevitellogenesis in the japanese oak silkworm, Antheraea yamamai(Lepidoptera:Saturniidae). Entomologia Sinica 9: 17-25.
Goodman, W.G., Adams, B. and Trost, J.T. 1985. Purification andcharacterization of a biliverdin-associated protein from thehemolymph of Manduca sexta. Biochemistry 24: 1168-1175.
Hiruma, K., Shinoda, T., Malone, F. and Riddiford, L.M. 1999. Juvenilehormone modulates 20-hydroxyecdysone-inducible ecdysonereceptor and ultraspiracle gene expression in the tobaccohornworm, Manduca sexta. Dev. Genes Evol. 209: 18-30.
Hyrsl, P. and Simek, V. 2005. An analysis of hemolymph protein profilesduring the final instar, prepupa and pupa of the silkworm Bombyxmori (Lepidoptera, Bombycidae). Biologia 60: 207-213.
Image, J analysis software from the National Institutes of Health, US(http://rsb.info.nih.gov/nih-image/).
Izumi, S., Tomino, S. and Chino H. 1980. Purification and molecularproperties of vitellin from the silkworm, Bombyx mori. InsectBiochem. 10: 199-208
Izumi, S., Kiguchi, K. and Tomino, S. 1984. Hormonal regulation ofbiosynthesis of major plasma proteins in Bombyx mori. Zool. Sci.1: 223-228.
Izumi, S., Sakurai, H., Fujii, T., Ikeda, W. and Tomino, S. 1988. Cloningof mRNA sequence coding for sex-specific storage protein ofBombyx mori. Biochim. Biophys. Acta. 949: 181-188.
Janarthanan, S., Krishnan, M. and Ignacimuthu, S. 1998. Changes inprotein profiles during pupal development in silkworm Bombyxmori (L.). Indian J. Exp. Biol. 36: 1158-1160.
Kajiura, Z. and Yamashita, O. 1992. Induction of metamorphosis of themethoprene induced dauer larvae of the silkworm, Bombyx mori,by ecdysteroids. Comp. Biochem. Physiol. 101 (A): 277-280.
Kamimura, M. 1995. Effects of juvenile hormone analogue, fenoxycarb,on larval growth of the silkworm, Bombyx mori (Lepidoptera:Bombycidae). Appl. Ent. Zool. 30: 487-489.
References

Investigation of the influence of the juvenile hormone analogue fenoxycarb on major hemolymph proteins of the silkwormBombyx mori during the last larval instar
236
Kamimura, M. and Kiuchi, M. 1998. Effects of a juvenile hormoneanalog, fenoxycarb, on 5th stadium of the larvae of the silkworm,Bombyx mori (Lepidoptera: Bombycidae). Appl. Entomol. Zool.33: 333-338.
Kim, B.S. and Kim, H.R. 1994. Purification and characteristics oflipophorin in Bombyx mori. Animal Physiology andBiochemistry 37: 76-87.
Kim, E.J., Park, H.I. and Park, T.H. 2003. Inhibition of apoptosis byrecombinant 30K protein originating from silkwormhemolymph. Biochemical and Biophysical ResearchCommunications 308: 523-528.
Kishimoto, A., Nakato, H., Izumi, S. and Tomino, S. 1999. Biosynthesisof major plasma proteins in the primary culture of fat body cellsfrom the silkworm, Bombyx mori. Cell Tissue Res. 297: 329-335.
Laemmli, U.K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227: 680-685.
Laufer, H. 1960. Blood proteins in insect development. Ann. N. Y. Acad.Sci. 89: 490-515.
Leonardi, M.G., Cappellozza S., Ianne P., Cappellozza L., Parentis P. andGiordina, B. 1996. Effects of the topical application an insectgrowth regulator (fenoxycarb) on some physiological parametersin the fifth instar larvae of the silkworm Bombyx mori. Comp.Biochem. Physiol. 113B: 361-365.
Levenbook, L. and Bauer, A.C. 1984. The fate of the larval storageprotein calliphorin during adult development of Calliphoravicina. Insect Biochemistry 14: 77-86.
Monconduit, H. and Mauchamp, B. 1998. Effects of ultralow doses offenoxycarb on juvenile hormone-regulated physiologicalparameters in the silkworm, Bombyx mori L. Archives of InsectBiochemistry and Physiology 37: 178-189.
Monconduit, H. and Mauchamp, B. 1999. Fenoxycarb, a potentcontaminant of the silkworm Bombyx mori L., does not influenceits juvenile hormone titer. Archives of Insect Biochemistry andPhysiology 40: 141-149
Nagata, M. and Kobayashi, M. 1990. Quantitative changes in storageproteins during larval development of the silkworm, Bombyxmori. Journal of Sericultural Science of Japan 59: 461-468.
Niimi, S., and Sakurai, S. 1997. Development changes in juvenilehormone and juvenile hormone acid titers in the hemolymph andin vitro juvenile hormone synthesis by corpora allata of silkworm,Bombyx mori. J. Insect Physiol. 43: 875-884.
Nijhout, H.F. 1994. Insect Hormone. Princeton University Press,Princeton. 142-170.
Ogawa, K. and Tojo, S. 1981. Quantitative changes of storage proteinsand vitellogenin during the pupal-adult development in thesilkworm, Bombyx mori (Lepidoptera: Bombycidae). Appl.Entomol. Zool. 16: 288-296.
Ogawa, N., Kishimoto, A., Asano, T. and Izumi, S. 2005. Thehomeodomain protein PBX participates in JH-related suppressiveregulation on the expression of major plasma protein genes in thesilkworm, Bombyx mori. Insect Biochemistry and MolecularBiology 35: 217-229.
Ohnishi, E. 1987. Growth and maturation of ovaries in isolatedabdomens of Bombyx mori: response to ecdysteroids and othersteroids. Zoological Science 4: 315-321.
Park, H.J., Kim, E.J., Koo, T.Y. and Park, T.H. 2003. Purification ofrecombinant 30K protein produced in Escherichia coli and itsanti-apoptotic effect in mammalian and insect cell systems.Enzyme and Microbial Technology 33: 466-471.
Pinto, L.Z., Bitondi, M.M.G. and Simoes, Z.L.P. 2000. Inhibition ofvitellogenin synthesis in Apis mellifera workers by a juvenilehormone analogue, Pyriproxyfen. Journal of Insect Physiology46: 153-160.
Riddiford, L.M. 1985. Hormonal action at the cellular level. In:Comprehensive Insect Physiology Biochemistry andPharmacology (ed. Kerkut G.A. and Gilbert L.I.), Pergamon,Oxford. pp. 37-84.
Sakurai, S. and Imokawa, H. 1988. Developmental arrest induced byjuvenile hormone in larvae of Bombyx mori. Archives of InsectBiochemistry and Physiology 8: 219-228.
Sakurai, S., Kaya, M. and Satake, S. 1998. Hemolymph ecdysteroid titerand ecdysteroid-dependent developmental events in the last-larval stadium of the silkworm, Bombyx mori: role of lowecdysteroid titer in larval-pupal metamorphosis and reappraisalof the head critical period. Journal of Insect Physiology 44: 867-881.
Sakurai, S., Okuda, M. and Ohtaki, T. 1989. Juvenile hormone inhibitsecdysone secretion and responsiveness to protharacicotropichormone in prothoracic glands of Bombyx mori. General andComparative Endocrinology 75: 22-230.
Sun, Q., Zhao, P., Lin, Y., Hou, Y., Xia, Q.Y. and Xiang, Z.H. 2007.Analysis of the structure and expression of the 30K protein genesin silkworm, Bombyx mori. The Authors Insect Science 14: 5-14.
Telfer, W.H. and Kunkel, J.G. 1991. The function and evolution of insectstorage hexamers. Annu. Rev. Entomol. 36: 205-228.
Terwilliger, N.B. 1999. Hemolymph proteins and molting in crustaceansand insects. Amer. Zool. 39: 589-599.
Tojo, S., Nagata, M. and Kobayashi, M. 1980. Storage proteins in thesilkworm, Bombyx mori. Insect Biochem. 10: 289-303.
Tomino, S. 1985. Major plasma proteins of Bombyx mori. Zool. Sci. 2:293-303.
Ujita, M., Katsuno, Y., Kawachi, I., Ueno, Y., Banno, Y., Fujii, H. andHara, A. 2005. Glucan-binding activity of silkworm 30-kDaapolipoprotein and its involvement in defense against fungalinfection. Bioscience Biotechnology and Biochemistry 69: 1178-1185.
Urich, K. 1994. Comparative Animal Biochemistry. Springer Verlag,Berlin.
Yano, K., Sakurai, M. T., Izumi, S. and Tomino, S. 1994. Vitellogeningene of the silkworm, Bombyx mori: structure and sex-dependentexpression. FEBS Letters 356: 207-211.
Zhou, X. and Riddiford, L.M. 2002. Broad specifies pupal developmentand mediates the ‘status quo’ action of juvenile hormone on thepupal-adult transformation in Drosophila and Manduca.Development 129: 2259-69.








![Dpp regulates autophagy-dependent midgut removal and ... · in insects [6]. The hormone-mediated juvenile-to-adult transition requires the remodeling or removal of juvenile structures](https://static.fdocuments.in/doc/165x107/60cd9a3e3cd5a305df023b54/dpp-regulates-autophagy-dependent-midgut-removal-and-in-insects-6-the-hormone-mediated.jpg)









