
Introduction to Mathematical Methods in Neurobiology: Dynamical Systems Oren Shriki 2009 Modeling...
41
Introduction to Mathematical Methods in Neurobiology: Dynamical Systems Oren Shriki 2009 Modeling Conductance-Based Networks by Rate Models 1
-
Upload
ophelia-bruce -
Category
Documents
-
view
218 -
download
3
Transcript of Introduction to Mathematical Methods in Neurobiology: Dynamical Systems Oren Shriki 2009 Modeling...

Introduction to Mathematical Methods in Neurobiology:
Dynamical Systems
Oren Shriki
2009
Modeling Conductance-Based Networks by Rate Models 1

References:
• Shriki, Hansel, Sompolinsky, Neural Computation 15, 1809–1841 (2003)
• Tuckwell, HC. Introduction to Theoretical Neurobiology, I&II, Cambridge UP, 1988.
2

Conductance-Based Models vs. Simplified Models
• There are two main classes of theoretical approaches to the behavior of neural systems:
– Simulations of detailed biophysical models.
– Analytical (and numerical) solutions of simplified models (e.g. Hopfield models, rate models).
• Simplified models are extremely useful for studying the collective behavior of large neuronal networks. However, it is not always clear when they provide a relevant description of the biological system, and what meaning can be assigned to the quantities and parameters used in them.
3

Conductance-Based Models vs. Simplified Models
• Using mean-field theory we can describe the dynamics of a conductance-based network in terms of firing-rates rather than voltages.
• The analysis will lead to a biophysical interpretation of the parameters that appear in classical rate models.
• The analysis will be divided into two parts:
– A. Steady-state analysis (constant firing rates)
– B. Firing-rate dynamics
4
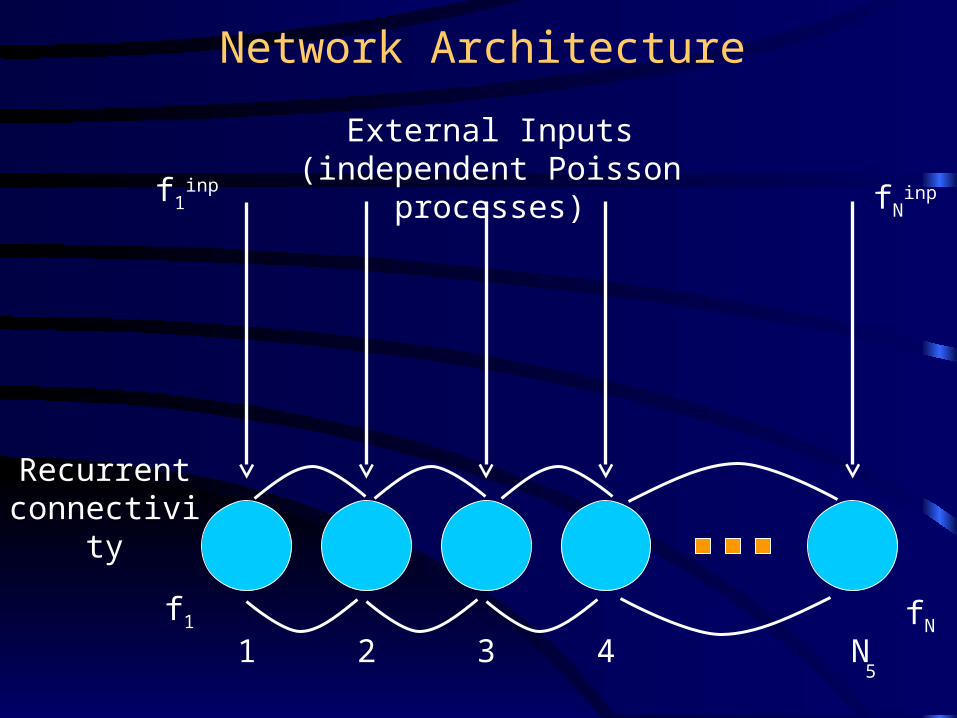
Network Architecture
External Inputs (independent Poisson processes)
Recurrent connectivity
1 2 3 4 N
f1inp
f1
fNinp
fN
5

Voltage Dynamics for A Network of Conductance-Based Point Neurons
We assume that the neurons are point neurons obeying Hodgkin-Huxley type dynamics:
• Iactive – Ionic current involved in the action potential• Iext – External synaptic inputs• Inet – Synaptic inputs from within the network• Iapp – External current applied by the experimentalist
),,1( )( NiIIIIEtVgdt
dVC app
ineti
exti
activeiLiL
im
6

tt_spike
sg t
G
s
A spike at time tspike of the presynaptic cell contributes to the postsynaptic cell a time-depndent conductance , gs(t):
Synaptic Conductances
Peak conductance
7

Synaptic Dynamics: An Example• Synaptic dynamics are usually characterized by
fast rise and slow decay.
• The simplest model assumes instantaneous rise and exponential decay:
)(tGRg
dt
dg ss
spiket
spiketttR )( (Presynaptic rate)
8
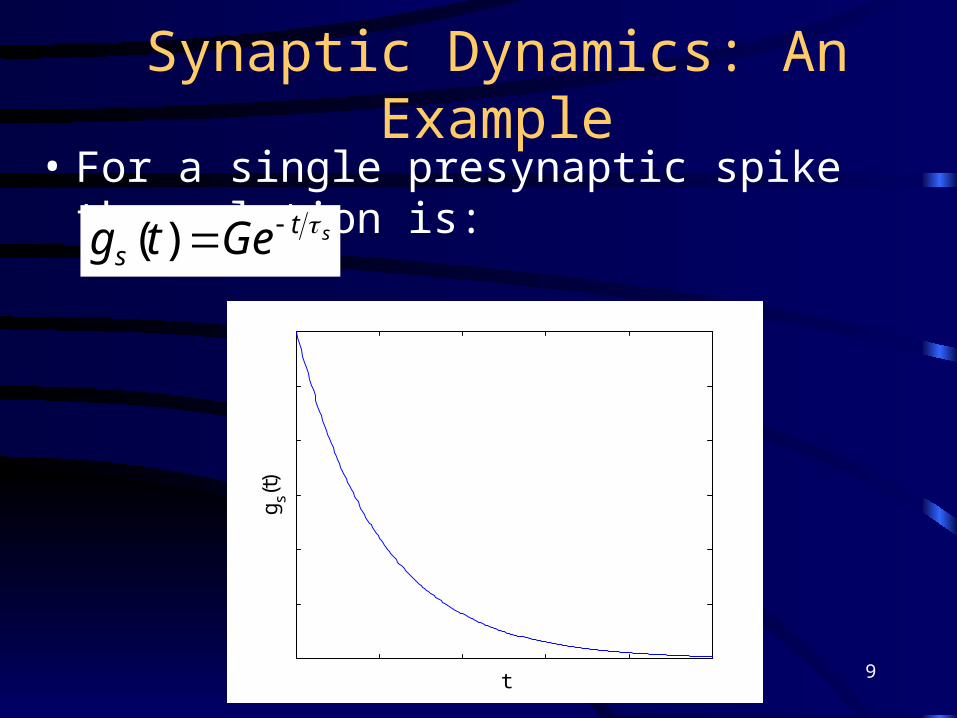
Synaptic Dynamics: An Example• For a single presynaptic spike the solution is:
sts Getg )(
t
g s(t)
9

Synaptic Dynamics: An Example• Implementation in numerical simulations:
• Given the time step dt define the attenuation factor:
• A dimensionless parameter, f, is increased by 1 after each presynaptic spike and multiplied by the attenuation factor in each time step.
• The conductance is the product of f and the peak conductance, G.
sdtdt ee
10

Synaptic Dynamics• For simplicity, we shall write in general:
• K(t) is the time course (dimensionless) function.
• We define:
• For example:
)()( tGKtgs
0
)( dttKs
st
st ss edte
0
0 11

External Synaptic Current
• The explicit expression for the external synaptic current is:
• The peak synaptic conductance is:
• The time constant is:
inpiG
inpi
)()()( tVEtgtI iinpinp
iexti
12

Internal Synaptic Current
• The explicit expression for the internal synaptic current is:
• The peak synaptic conductance is:
• The time constant is:
ijG
ij
N
jijij
neti tVEtgtI
1
)()()(
13

Part A: Steady-State Analysis
The main assumptions are:
• Firing rates of external inputs are constant in time
• Firing rates within the network are constant in time
• The network state is asynchronous
• The network contains many neurons
14

Asynchronous States in Large Networks
• In large asynchronous networks each neuron is bombarded by many synaptic inputs at any moment.
• The fluctuations in the total input synaptic conductance around the mean are relatively small.
• Thus, the total synaptic conductance in the input to each neuron is (approximately) constant.
15

Asynchronous States in Large Networks
• The figure below shows the synaptic conductance of a certain postsynaptic neuron in a simulation of two interacting populations (excitatory and inhibitory):
16

Mean-Field Approximation
• We can substitute the total synaptic conductance by its mean value.
• This approximation is called the “Mean-Field” (MF) Approximation.
• The justification for the MF approximation is the central limit theorem.
17

The Central Limit Theorem
• A random variable ,which is the sum of many independent random variables, has a Gaussian distribution with mean value equal to the sum of the mean values of the individual random variables.
• The ratio between the standard deviation and the mean of the sum satisfies:
where N is the number of individual random variables.
Nmean
std 1
18

Mean-Field Analysis of the Synaptic Inputs
• The total contribution to the i’th neuron from within the network is:
• The contribution to this sum from the j’th neuron is:
N
j tjijij
N
jij
neti
j
ttKGtgtg11
)()(
jt
jijijij ttKGtg )(
19

Mean-Field Analysis of the Synaptic Inputs
• Consider a time window T>>1/fj, where fj is the firing rate of the presynaptic neuron j.
• Schematically, the contribution of the j’th neuron in this time window looks like this:
)(tgij
Tt
ijg
spikes
Tt
20

Mean-Field Analysis of the Synaptic Inputs
• The mean conductance due to the j’th neuron over a long time window is:
• The mean conductance resulting from all neurons in the network is:
jijij
ijijspike
T
ijij
fG
dttKGT
TNdttg
Tg
00
)()(
)(1
N
jjijij
N
jij
neti fGgg
11
21

Mean-Field Analysis of the Synaptic Inputs
• The mean-field approximation is:
• Effectively, we replace a spatial averaging by a temporal averaging .
• Using a similar analysis, the contribution of the external inputs can be replaced by:
N
jjijij
neti
neti fGgtg
1
)(
inpi
inpi
inpi
exti
exti fGgtg )(
22

Mean-Field Analysis of the Synaptic Inputs
• The synaptic currents are not constant in time since the voltage of the neuron varies significantly over time:
• We can decompose the last expression in the following way:
N
jijij
neti tVEtgtI
1
)()()(
N
jijLi
N
jLjij
N
jiLLjij
neti
tgEtVEEtg
tVEEEtgtI
11
1
)()()(
)()()(
23

Mean-Field Analysis of the Synaptic Inputs
• We now use the MF approximation :
• This gives: Lj
N
jjijij
N
jLjij
N
jjijij
neti
N
jij
EEfGEEtg
fGtgtg
11
11
)(
)()(
N
jjijijLijLj
N
jijij
neti fGEtVfEEGtI
11
)()(
A constant applied current
A constant increase in the leak conductance 24

Mean-Field Analysis of the Synaptic Inputs
• Similarly:
• We obtained the following mapping:
inpi
inpi
inpiLi
inpiL
inpi
inpi
inpi
exti fGEtVfEEGtI )()(
inpi
inpi
inpi
N
jjijijLL
inpiL
inpi
inpi
inpijLj
N
jijij
appi
appi
fGfGgg
fEEGfEEGII
1
1
25

Mean-Field Analysis of the Synaptic Inputs
• To sum up:• Asynchronous Synaptic Inputs Produce a Stationary Shift in
the Voltage-Independent Currentand in the Input Passive Conductanceof the Postsynaptic Cell.
• To complete the loop and determine the network’s firing rates we need to know how the firing rate of a single cell is affected by these shifts.
26

Current-Frequency Response Curves of Cortical Neurons are Semi-Linear
Excitatory Neuron (After: Ahmed et. al., Cerebral Cortex 8, 462-476, 1998):
Inhibitory Neurons (After: Azouz et. al., Cerebral Cortex 7, 534-545, 1997) :
27

The Effect of Changing the Input Conductance is Subtractive
Experiment:
f-I curves of a cortical neuron before and after iontophoresis
of baclofen, which opens synaptic conductances.
(Connors B. et. al., Progress in Brain Research, Vol. 90, 1992).
28

A Hodgkin-Huxley Neuron with an A-current
b
n
h
/τ(V)-mbdb/dt
/τ(V)-nndn/dt
/τ(V)-hhdh/dt
)(, tIwVIdt
dVC ion
)()()()(
,,,
LL3
K4
KNa3
Na EVgEVbagEVngEVhmg
nhmVI
KA
ion
29
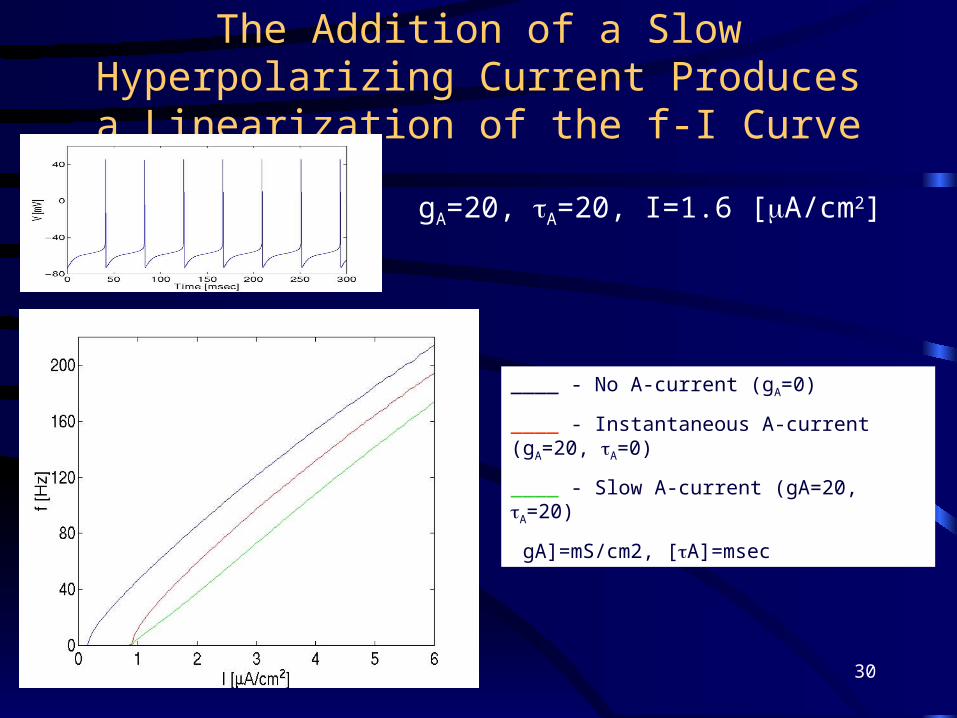
The Addition of a Slow Hyperpolarizing Current Produces a Linearization of the f-I Curve
____ - No A-current (gA=0)
____ - Instantaneous A-current (gA=20, A=0)
____ - Slow A-current (gA=20, A=20)
[gA]=mS/cm2, [A]=msec
gA=20, A=20, I=1.6 [A/cm2]
30

The Effect of Increasing gL is Subtractive
31

Model Equations with Parameters:
Shriki et al., Neural Computation 15, 1809–1841 (2003)
32

The Dependence of the Firing Rate on I and on gL Can Be Described by a Simple Phenomenological Model
LcC
C
gVII
IIf0
[x]+=x if x>0 and 0 otherwise.
We find for the model neuron :
=35.4 [Hz/(A/cm2)]Vc=5.6 [mV]IC
0=0.65 [A/cm2] 33

The Effect of the Synaptic Input On the Firing Rate
Combining the previous results, we find that the steady-state firing rates obey the following equations:
LccjcLj
N
jijij
inpicL
inpi
inpi
inpi
appii
gVIfVEEG
fVEEGIf
0
1
34

The Effect of the Synaptic Input On the Firing Rate
c
N
jjij
inpi
inpi
appii IfJfJIf
1
This can be written us:
Where:
cL
inpi
inpi
inpi
inpi
cLjijijij
VEEGJ
VEEGJ
35

The Units of the Interactions
• The units of J are units of electric charge:
• The quantity Jijfj reflects the mean current due to the j’th synaptic source.
• The strength of the interaction Jij reflects the amount of charge that is transferred with each presynaptic action potential.
CQtIVtgJ
36

E Eij ij ij s L cJ G E E V
s L cE E V
s L cE E V
‘‘excitatoryexcitatory’’
‘‘inhibitoryinhibitory’’
The Sign of the Interaction
• The interaction strength in the rate model has the form:
• The rule for excitation / inhibition is:
• This does not necessarily coincide with the biological definition of excitation/inhibition.
37

The Sign of the Interaction
• The biological definition is:
• A positive J implies that this synaptic source increases the firing rate.
• In general, it may be that a certain synaptic source tends to elicit a spike but increases the conductance in a way that reduces the overall firing rate.
sE
sE
excitatoryexcitatory
inhibitoryinhibitory
38

The Model Parameters
• Neuron:
• β – Slope of frequency-current response
• Vc, Ic0 – Dependence of current threshold on leak
conductance
• EL – Reversal potential of leak conductance
• Synapse:
• Gij – Peak synaptic conductance
• Ej – Synaptic reversal potential
• τij – Synaptic time constant 39

ccinpinp IJfNfJf
cinpinp
c
IfJJN
f
1
Rate Model for a Homogeneous, Highly Connected Excitatory Network:
40

Simulations of the Excitatory Network and Rate Model Prediction
41


















