
Intracellular Shigella flexneri Associated the Locus …INFECTION ANDIMMUNITY, Feb. 1989, p. 477-486...
10
INFECTION AND IMMUNITY, Feb. 1989, p. 477-486 0019-9567/89/020477-10$02.00/0 Copyright © 1989, American Society for Microbiology Intracellular Spread of Shigella flexneri Associated with the kcpA Locus and a 140-Kilodalton Protein TIBOR PAL,lt JOHN W. NEWLAND,' BEN D. TALL,2 SAMUEL B. FORMAL,' AND THOMAS LARRY HALE'* Department of Enteric Infections, Walter Reed Army Institute of Research, Washington, D.C. 20307,1 and Center for Vaccine Development, University of Maryland School of Medicine, Baltimore, Maryland 212012 Received 27 July 1988/Accepted 17 October 1988 Escherichia coli K-12 hybrids carrying both the 220-kilobase plasmid and the purE-linked kcpA locus from Shigella flexneri expressed a 140-kilodalton (kDa) protein which was recognized by convalescent sera from monkeys infected with S. flexneri. These hybrids were tested for the ability to produce plaques in HeLa cell monolayers. Hybrid strains which carried both the 220-kilobase plasmid and the kcpA locus had a plaque-forming efficiency of at least 10-4 PFU/CFU, whereas the plaque-forming efficiency of hybrids that carried only the shigelia invasion plasmid ranged from undetectable to 10-6. Variants were purified from the rare plaques formed by E. coli hybrids that carried only the shigelia invasion plasmid. These plaque-purified strains also expressed the 140-kDa protein, and they had a plaque-forming efficiency of at least 1o-4. Transduction of the purE locus from a plaque-purified hybrid into a non-plaque-forming E. coli K-12 strain did not alter the phenotype of the recipient, but conjugation of the shigelia invasion plasmid into this transductant reconstituted both expression of the 140-kDa protein and the plaque-forming phenotype. Invasive E. coli K-12 hybrids carrying only the shigelia invasion plasmid remained localized within discrete areas of the HeLa cell cytoplasm, whereas hybrids that also carried the S. flexneri kcpA locus grew in a dispersed pattern throughout the host cell cytoplasm. The dispersal of these organisms was inhibited by cytochalasin D, which suggested that host cell microfilaments may play a role in the intracellular spread of enteroinvasive pathogens. Genes carried on a 220-kilobase (kb) Shigella flexneri plasmid encode determinants that are required for epithelial cell invasion (11, 27). Four plasmid-encoded proteins that are associated with the invasive phenotype are the ipa (invasion plasmid antigen) gene products (2, 3, 14). These proteins induce strong antibody responses after shigella infections in monkeys and in humans (14, 15, 22). In addition to the ipa genes, a plasmid locus designated virG is neces- sary for intracellular spread of shigellae (17) and for kerato- conjunctivitis provocation in the guinea pig eye (Sereny test) (17, 30). It has recently been reported that both the ipa locus and virG are positively regulated by a third plasmid locus, designated virF (26). It was also suggested that the protein product of virG is an immunogenic 130-kilodalton (kDa) protein (26). This protein is probably identical to the 140-kDa protein that has previously been associated with the 220-kb plasmid of S. flexneri (4, 22). Other virulence-associated genes are located in the shi- gella chromosome. These genes are linked to the purE locus at 12 min, the his operon at 44 min, the mtl operon at 75 min, and the arg operon at 90 min (27). The His marker is closely linked to genes encoding the Shigella somatic antigen (5), the Arg marker is linked to an ability to elicit fluid accumulation in the rabbit ileal loop model (27), and the Mtl marker is linked to aerobactin genes (8). In contrast to these identifi- able virulence determinants, the virulence-associated gene(s) linked to purE has not been characterized. It is clear that this region of the shigella chromosome is necessary for a positive Sereny test; therefore, the purE-linked virulence marker has been designated kcpA, denoting keratoconjunc- tivitis provocation (6). Since P1 vir transduction of the purE region from the Escherichia coli K-12 chromosome converts * Corresponding author. t Present address: Institute of Microbiology, University Medical School, Pecs, Hungary H-7643. 477 up to 60% of transductants to a Sereny-negative phenotype (6), kcpA is closely linked to purE. In this communication, we present evidence of genetic regulation involving a shigella virulence plasmid gene(s) and chromosomal gene(s) linked to the purE locus. These genetic elements are necessary for expression of the 140-kDa immu- nogenic protein, and they are also associated with the spread of bacteria both within and between epithelial cells. (A preliminary report of these findings has been presented [T. Pail, T. L. Hale, P. J. Sansonetti, B. Tal, and S. B. Formal, Abstr. Annu. Meet. Am. Soc. Microbiol. 1988, B24, p. 33].) MATERIALS AND METHODS Strains. E. coli 395-1, a nalidixic acid-resistant mutant of K-12 AB-1133, was used as a recipient for the S. flexneri plasmid and chromosomal genes listed in Table 1. The construction of these E. coli hybrid strains, which was described previously (27), can be summarized as follows. The 220-kb plasmid (pWR100) of S. flexneri serotype 5 strain M9OT was tagged with TnS, which encodes kanamycin resistance, and this plasmid (pWR110) was mobilized as a cointegrate with R64drd-11, which encodes tetracycline re- sistance, into S. flexneri serotype 1 strain M25-8A. The latter strain served as a donor of shigella virulence plasmid genes to E. coli K-12 in the form of pWR11O-R64drd-11. The kcpA locus of S. flexneri was conjugally transferred in matings by using an isolate of E. coli Hfr H-6 which had previously incorporated the purE region of S. flexneri 2a M4243 by P1 vir transduction (6). Additional markers from the shigella chromosome were conjugally transferred in matings with S. flexneri 2a Hfr strain 256 (5). Plaque-forming strains of E. coli K-12 containing pWR110- R64drd-11 were isolated from rare HeLa cell plaques formed by strain 7262-1-10 (23). To determine whether mutations had occurred in the pWR110 plasmid carried by such plaque- Vol. 57, No. 2 on January 25, 2020 by guest http://iai.asm.org/ Downloaded from
Transcript of Intracellular Shigella flexneri Associated the Locus …INFECTION ANDIMMUNITY, Feb. 1989, p. 477-486...

INFECTION AND IMMUNITY, Feb. 1989, p. 477-4860019-9567/89/020477-10$02.00/0Copyright © 1989, American Society for Microbiology
Intracellular Spread of Shigella flexneri Associated with the kcpALocus and a 140-Kilodalton Protein
TIBOR PAL,lt JOHN W. NEWLAND,' BEN D. TALL,2 SAMUEL B. FORMAL,' AND THOMAS LARRY HALE'*Department of Enteric Infections, Walter Reed Army Institute of Research, Washington, D.C. 20307,1 and Center for
Vaccine Development, University of Maryland School of Medicine, Baltimore, Maryland 212012
Received 27 July 1988/Accepted 17 October 1988
Escherichia coli K-12 hybrids carrying both the 220-kilobase plasmid and the purE-linked kcpA locus fromShigella flexneri expressed a 140-kilodalton (kDa) protein which was recognized by convalescent sera frommonkeys infected with S. flexneri. These hybrids were tested for the ability to produce plaques in HeLa cell
monolayers. Hybrid strains which carried both the 220-kilobase plasmid and the kcpA locus had a
plaque-forming efficiency of at least 10-4 PFU/CFU, whereas the plaque-forming efficiency of hybrids thatcarried only the shigelia invasion plasmid ranged from undetectable to 10-6. Variants were purified from therare plaques formed by E. coli hybrids that carried only the shigelia invasion plasmid. These plaque-purifiedstrains also expressed the 140-kDa protein, and they had a plaque-forming efficiency of at least 1o-4.Transduction of the purE locus from a plaque-purified hybrid into a non-plaque-forming E. coli K-12 strain didnot alter the phenotype of the recipient, but conjugation of the shigelia invasion plasmid into this transductantreconstituted both expression of the 140-kDa protein and the plaque-forming phenotype. Invasive E. coli K-12hybrids carrying only the shigelia invasion plasmid remained localized within discrete areas of the HeLa cellcytoplasm, whereas hybrids that also carried the S. flexneri kcpA locus grew in a dispersed pattern throughoutthe host cell cytoplasm. The dispersal of these organisms was inhibited by cytochalasin D, which suggested thathost cell microfilaments may play a role in the intracellular spread of enteroinvasive pathogens.
Genes carried on a 220-kilobase (kb) Shigella flexneriplasmid encode determinants that are required for epithelialcell invasion (11, 27). Four plasmid-encoded proteins thatare associated with the invasive phenotype are the ipa(invasion plasmid antigen) gene products (2, 3, 14). Theseproteins induce strong antibody responses after shigellainfections in monkeys and in humans (14, 15, 22). In additionto the ipa genes, a plasmid locus designated virG is neces-sary for intracellular spread of shigellae (17) and for kerato-conjunctivitis provocation in the guinea pig eye (Sereny test)(17, 30). It has recently been reported that both the ipa locusand virG are positively regulated by a third plasmid locus,designated virF (26). It was also suggested that the proteinproduct of virG is an immunogenic 130-kilodalton (kDa)protein (26). This protein is probably identical to the 140-kDaprotein that has previously been associated with the 220-kbplasmid of S. flexneri (4, 22).
Other virulence-associated genes are located in the shi-gella chromosome. These genes are linked to the purE locusat 12 min, the his operon at 44 min, the mtl operon at 75 min,and the arg operon at 90 min (27). The His marker is closelylinked to genes encoding the Shigella somatic antigen (5), theArg marker is linked to an ability to elicit fluid accumulationin the rabbit ileal loop model (27), and the Mtl marker islinked to aerobactin genes (8). In contrast to these identifi-able virulence determinants, the virulence-associatedgene(s) linked to purE has not been characterized. It is clearthat this region of the shigella chromosome is necessary fora positive Sereny test; therefore, the purE-linked virulencemarker has been designated kcpA, denoting keratoconjunc-tivitis provocation (6). Since P1 vir transduction of the purEregion from the Escherichia coli K-12 chromosome converts
* Corresponding author.t Present address: Institute of Microbiology, University Medical
School, Pecs, Hungary H-7643.
477
up to 60% of transductants to a Sereny-negative phenotype(6), kcpA is closely linked to purE.
In this communication, we present evidence of geneticregulation involving a shigella virulence plasmid gene(s) andchromosomal gene(s) linked to the purE locus. These geneticelements are necessary for expression of the 140-kDa immu-nogenic protein, and they are also associated with the spreadof bacteria both within and between epithelial cells.(A preliminary report of these findings has been presented
[T. Pail, T. L. Hale, P. J. Sansonetti, B. Tal, and S. B.Formal, Abstr. Annu. Meet. Am. Soc. Microbiol. 1988, B24,p. 33].)
MATERIALS AND METHODS
Strains. E. coli 395-1, a nalidixic acid-resistant mutant ofK-12 AB-1133, was used as a recipient for the S. flexneriplasmid and chromosomal genes listed in Table 1. Theconstruction of these E. coli hybrid strains, which was
described previously (27), can be summarized as follows.The 220-kb plasmid (pWR100) of S. flexneri serotype 5 strainM9OT was tagged with TnS, which encodes kanamycinresistance, and this plasmid (pWR110) was mobilized as a
cointegrate with R64drd-11, which encodes tetracycline re-sistance, into S. flexneri serotype 1 strain M25-8A. The latterstrain served as a donor of shigella virulence plasmid genesto E. coli K-12 in the form of pWR11O-R64drd-11. The kcpAlocus of S. flexneri was conjugally transferred in matings byusing an isolate of E. coli Hfr H-6 which had previouslyincorporated the purE region of S. flexneri 2a M4243 by P1vir transduction (6). Additional markers from the shigellachromosome were conjugally transferred in matings with S.flexneri 2a Hfr strain 256 (5).
Plaque-forming strains of E. coli K-12 containing pWR110-R64drd-11 were isolated from rare HeLa cell plaques formedby strain 7262-1-10 (23). To determine whether mutationshad occurred in the pWR110 plasmid carried by such plaque-
Vol. 57, No. 2
on January 25, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

478 PAL ET AL.
TABLE 1. Bacterial strains and relevant characteristics
Genotype' PhenotypeHybrid strain Plasmid
Mt,purE 140-kDa Plaque Sereny
pWR110 tSm kcpA proteinb formation test
E. coli K-127262-1-10 + - - - - -7290-1-19 + + - - - - -7185-2-2 + + + - - - -7275-1-6 + - - + + + d
7293-2-2 + + - + + + -7194-2-3 + + + + + + +7262-1-24PP2e + - - -f + +7262-4-2 + - - -g + + -
S. flexneri 2aG283A + + + + + + +G283B + + + h (+)i - -
a S. flexneri genes inherited.b Immunogenic outer membrane protein.c Plaque-forming efficiency ranging from undetectable to 10-6 PFU/CFU
was considered negative.d Plaque-forming efficiency greater than 10-4 PFU/CFU was considered
positive.ePlaque-purified isolate from 7262-1-10.f Plaque-purified strains express a KcpA-positive phenotype even though
they have not inherited purE kcpA from S. flexneri.g Transductant of 7262-1-10 carrying the purE region from 7262-1-24PP2
and expressing the KcpA-positive phenotype.h Transductant carrying the purE region from E. coli K-12.Weak expression of the 140-kDa protein.
purified strains, the plasmid was conjugally transferred froma plaque-forming variant (7262-1-1OPP7) into a rifampin-resistant E. coli K-12 395-1 recipient. The latter strain hadbeen isolated by selection on tryptic soy agar (TSA; DifcoLaboratories, Detroit, Mich.) containing 100 ,ug of rifampinper ml. The transconjugant which carried pWR110-R64drd-11 from 7262-1-1OPP7 was then selected on rifampin (100,ug/ml) and tetracycline (20 ,ug/ml).To determine whether the plaque-forming phenotype was
linked to the purE chromosomal region of E. coli K-12, thisregion was transferred from a plaque-purified isolate (7262-1-24PP2) back into the 7262-1-10 parent strain by the follow-ing procedure. A tetracycline-sensitive variant of 7262-1-10(designated 7262-1-24) was selected after growth on fusaricacid (18), and this culture was applied to HeLa cell mono-layers in the plaque assay (23). A plaque-forming variant,designated 7262-1-24PP2, was isolated from a HeLa cellplaque. The purE79::TnJO region of E. coli NK6051 (ob-tained from the E. coli Genetic Stock Center as strain CGSC6189) was transduced by P1 vir into 7262-1-24PP2. Theresulting strain, 7262-1-24PP2purE: :TnJO, was a purine aux-otroph which retained the plaque-forming phenotype. ThepurE region of 7262-1-24PPpurE: :TnJO was then transducedby P1 vir back into 7262-1-24 by selecting for tetracyclineresistance. The resulting transductant was designated 7262-4-2. The purE region of 7262-1-24PP2purE::TnJO was alsobackcrossed into the original 395-1 K-12 strain, and thetransductant was designated 395-3A2. This strain was thenmated with S. flexneri M25-8A, and a kanamycin-resistanttransconjugant (8165-2-2) receiving pWR110-R64drd-11 wasselected.
S. flexneri hybrid strains G283A and G283B were obtainedby P1 vir transduction of the purE region from either M4243or E. coli K-12 into S. flexneri 2a G256-2 (6). The G256-2recipient strain is an adenine auxotroph of S. flexneri 2aM4243 that was obtained by nitrosoguanidine mutagenesis.This mutation did not alter the ability of G256-2 to evoke
keratoconjunctivitis. P1 vir transductants of G256-2 thatacquired adenine prototrophy from M4243 remained uni-formly Sereny positive. In contrast, 12 of 19 adenine-prototrophic transductants from E. coli K-12 were found tobe Sereny negative. Presumably, the shigella kcpA gene wasdisplaced during homologous recombination with the E. coliK-12 purE region in the Sereny-negative hybrids. G283A is aSereny-positive purE transductant from an M4243 P1 virlysate, and strain G283B is a Sereny-negative purE transduc-tant from an E. coli K-12 P1 vir lysate. G108-1-13 is aSereny-negative strain that is similar to G283B except thatthe E. coli K-12 purE region was transferred to a wild-typeM4243 strain by conjugation with E. coli Hfr H (6).
S. flexneri M9OT is a virulent strain of serotype 5, andM9OT-55 is a variant of this strain that was cured of the220-kb plasmid by one subculture at 42°C. M9OT-55 alsocarries a cryptic TnS chromosomal insertion. All strains usedwere maintained in lyophilized ampoules, and a fresh culturewas prepared from a rehydrated lyophile for each experi-ment by overnight culture at 37°C on TSA. Hybrid strainscarrying pWR110 were always cultured in medium contain-ing 20 p.g of kanamycin per ml. Individual colonies were thetransferred from TSA to Penassay broth (Difco), and theculture was grown, with aeration, for approximately 4 h at370C.Immunoblotting (Western blotting) procedure. Bacterial
cultures were grown with shaking at 370C in 10 ml ofPenassay broth until an A750 of 0.8 was reached. Thecultures were centrifuged into a pellet, the cells were lysedby boiling for 3 min in 0.5 ml of 1% sodium dodecyl sulfate(SDS) sample buffer, and 20-pd portions of these lysateswere separated into component parts by SDS-polyacryl-amide gel electrophoresis (PAGE). The gels were electro-blotted onto nitrocellulose and incubated with rhesus mon-key serum, and antibody recognizing shigella antigens wasdetected with alkaline phosphatase-protein A as previouslydescribed (4, 22). Sera used in the immunoblots were drawnfrom monkeys that had been orally challenged with virulentS. flexneri 2a 2 weeks before bleeding (4, 22). Even thoughthe sera varied with respect to recognition of ipa geneproducts, they were selected because they reacted stronglywith a 140-kDa protein.HeLa cell invasion and intracellular growth characteristics.
Assay of the invasive potential of bacterial cultures wascarried out in HeLa cell monolayers as previously described(12, 27). Organisms were centrifuged onto the surface ofnonconfluent HeLa cell monolayers, and infection of indi-vidual host cells occurred within 45 min. Extracellularorganisms were removed by four washings in Earle balancedsalts solution (EBSS; GIBCO Laboratories, Grand Island,N.Y.), and the extent of invasion was determined by micro-scopic examination of Giemsa-stained monolayers. Whenthe intracellular growth characteristics of invasive bacteriawere studied, basal medium Eagle (BME; GIBCO) contain-ing 5% heat-inactivated fetal bovine serum (GIBCO) and 20,ug of gentamicin (GIBCO) per ml was added to the mono-layers after a 45-min infection period. The plates containingthis BME-gentamicin medium were then incubated at 37°C in5% CO2 for up to 4 h. At hourly intervals after the additionof antibiotic-containing medium, plates were removed fromthe incubator, fixed in methanol, stained with Giemsa, andobserved microscopically.When the effect of cytochalasin D (Sigma Chemical Co.,
St. Louis, Mo.) on infected HeLa cells was studied, a stocksolution of 1 mg/ml in dimethyl sulfoxide (Sigma) was dilutedto a concentration of 0.25 ,ug/ml in BME-gentamicin me-
INFECT. IMMUN.
on January 25, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
kcpA AND VIRULENCE IN S. FLEXNERI 479
dium, and 2 ml of this medium was added to the tissueculture dishes after a 45-min infection period. After 2 h oftreatment with cytochalasin D, the plates were washed fourtimes with EBSS, and some were fixed in methanol andstained with Giemsa. In the remaining plates, the mediumwas replaced with BME containing 5% fetal bovine serumand 20 ,ug of gentamicin per ml, and incubation was contin-ued for an additional 2 h. When the effects of Nocodazole{methyl[5(2 - thienylcarbonyl) -1H -benzimidazol - 2 - yl]carba-mate; Sigma} were studied, HeLa cell monolayers werepreincubated with 3 ,ug of the drug per ml for 1 h. Nocoda-zole was also present during the initial infection period andthroughout the subsequent 4-h period of intracellular bacte-rial multiplication.
Quantification of intracellular bacterial growth. Noncon-fluent HeLa cell monolayers were infected for 30 min,washed four times with EBSS, and incubated (37°C, 5%C02) for up to 4 h in BME with 5% fetal bovine serum and20 ,g of gentamicin per ml. At hourly intervals, the culturemedium on a subset of plates was removed, and the cellswere suspended in 0.25% trypsin (GIBCO). The HeLa cellswere lysed by brief sonication, and quantitative plate countsof surviving (i.e., intracellular) bacteria were performed onTSA as previously described (13).
Plaque formation assay. Bacterial cultures for the plaqueassay were monitored for invasive potential in the HeLa cellassay described above so that only invasive cultures wereused. The plaque assay protocol, which is described else-where (23), used 107 CFU of exponential-phase culturessuspended in 0.2-ml volumes of brain heart infusion broth(Difco) for inoculation of the confluent HeLa cell monolay-ers. Three 10-fold dilutions of this inoculum were alsoinoculated onto HeLa cell monolayers because there wereusually too many plaques to count in monolayers exposed tothe higher inoculum of a plaque-forming strain. After 90 minof exposure to the bacteria, the monolayers were overlaidwith 0.5% agarose consisting of Dulbecco modified Eaglemedium (GIBCO) with 4.5 mg of glucose per ml, 5% fetalbovine serum, and 20 ,ug of gentamicin per ml. The plateswere incubated at 37°C in 5% CO2 for up to 7 days andexamined daily for areas of cytopathic effect (plaques)indicative of intercellular spread of invasive organisms. Toenhance visualization of the plaques, a second overlayconsisting of Dulbecco modified Eagle medium, 0.01% neu-tral red (GIBCO), 20 pug of gentamicin per ml, and 0.5%agarose was added.
Plaque-purified strains were isolated by aspiration with asterile Pasteur pipette of HeLa cell debris and the agaroseoverlaying individual plaques. These agarose plugs werestreaked directly into TSA plates and incubated at 37°C. Thestrains isolated from HeLa cell plaques were sensitive to 20,ug of gentamicin per ml.Electron microscopy. Nonconfluent HeLa cell monolayers
were infected for 45 min, washed, and incubated in BMEwith gentamicin for 3 h as described above. At the end of theinfection period or at the end the period of intracellularbacterial growth, the cells were suspended in 0.25% trypsin(GIBCO). The cells were pelleted by centrifugation, washedonce in EBSS, and prefixed in 2% paraformaldehyde-2.5%gluteraldehyde in 0.2 M cacodylate buffer (pH 7.2) for 24 h at4°C. The cells were then washed five times in EBSS,postfixed in 1% OS04 in the same buffer, and dehydratedthrough a graded series of ethanol (30 to 100%). After furtherdehydration in propylene oxide, the specimens were embed-ded in Epon, sectioned, stained with uranyl acetate and lead
1 2 3 4 5 6 7
140 kd
a
b
c
d
a-hv eS e *-- - ----S. ..
iS:. ,d'!.2 = .:iieW .iji;_'
a tWSFSF*: g ':
'.:'.^s ,^^ :.::.' __^ j_._
*X_S __ -F _-. '2lS- S ^-E z s-: -f UlDi.,...., .R,. .,.,..,.,,.F _
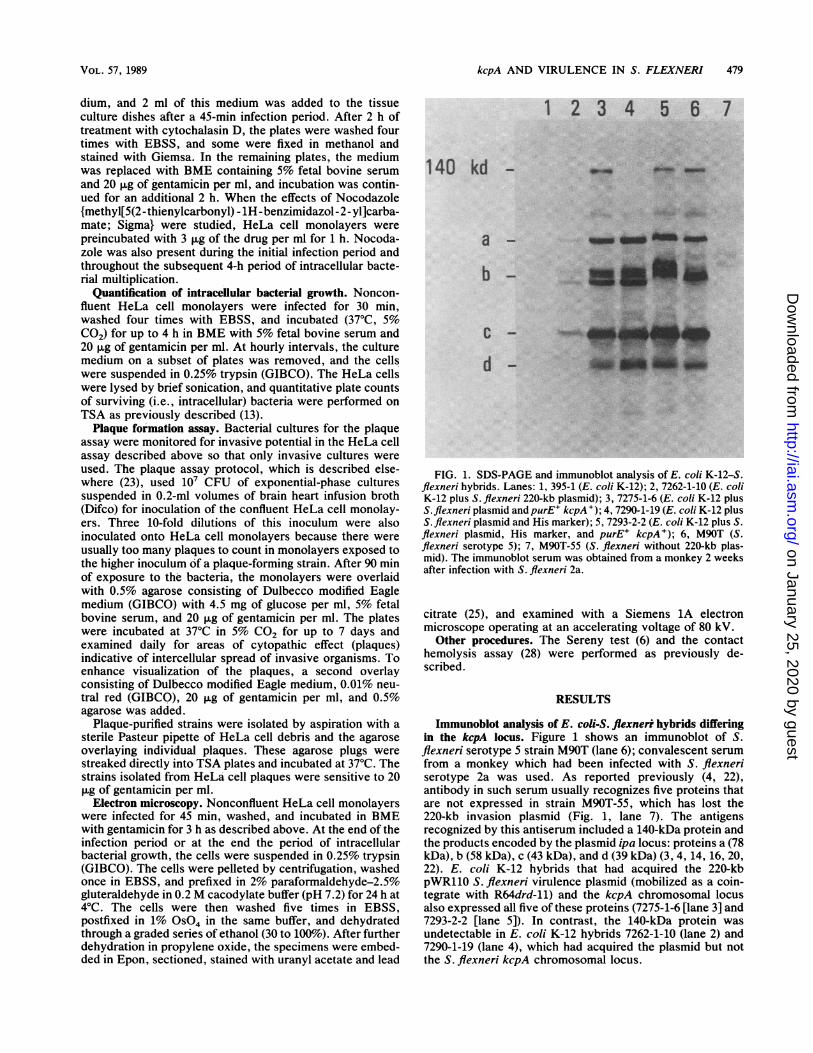
FIG. 1. SDS-PAGE and immunoblot analysis of E. coli K-12-S.flexneri hybrids. Lanes: 1, 395-1 (E. coli K-12); 2, 7262-1-10 (E. coliK-12 plus S. flexneri 220-kb plasmid); 3, 7275-1-6 (E. coli K-12 plusS.flexneri plasmid and purE+ kcpA+); 4, 7290-1-19 (E. coli K-12 plusS.flexneri plasmid and His marker); 5, 7293-2-2 (E. coli K-12 plus S.flexneri plasmid, His marker, and purE+ kcpA+); 6, M90T (S.flexneri serotype 5); 7, M90T-55 (S. flexneri without 220-kb plas-mid). The immunoblot serum was obtained from a monkey 2 weeksafter infection with S. flexneri 2a.
citrate (25), and examined with a Siemens 1A electronmicroscope operating at an accelerating voltage of 80 kV.Other procedures. The Sereny test (6) and the contact
hemolysis assay (28) were performed as previously de-scribed.
RESULTS
Immunoblot analysis of E. coli-S. flexnerr hybrids differingin the kcpA locus. Figure 1 shows an immunoblot of S.flexneri serotype 5 strain M90T (lane 6); convalescent serumfrom a monkey which had been infected with S. flexneriserotype 2a was used. As reported previously (4, 22),antibody in such serum usually recognizes five proteins thatare not expressed in strain M90T-55, which has lost the220-kb invasion plasmid (Fig. 1, lane 7). The antigensrecognized by this antiserum included a 140-kDa protein andthe products encoded by the plasmid ipa locus: proteins a (78kDa), b (58 kDa), c (43 kDa), and d (39 kDa) (3, 4, 14, 16, 20,22). E. coli K-12 hybrids that had acquired the 220-kbpWR110 S. flexneri virulence plasmid (mobilized as a coin-tegrate with R64drd-11) and the kcpA chromosomal locusalso expressed all five of these proteins (7275-1-6 [lane 3] and7293-2-2 [lane 5]). In contrast, the 140-kDa protein wasundetectable in E. coli K-12 hybrids 7262-1-10 (lane 2) and7290-1-19 (lane 4), which had acquired the plasmid but notthe S. flexneri kcpA chromosomal locus.
VOL. 57, 1989
on January 25, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

480 PAL ET AL.
1 2345
a-~~~~~~ai
1 2
1 40 kd
*R .:
OjCd
...*.. W B
FIG. 2. SDS-PAGE and immunoblot analysis of S. flexneri-E.coli K-12 hybrids. Lanes: 1, G283A (S. flexneri 2a purE plus S.flexneri 2apurE+ kcpA+ [transduced]); 2, G283B (S.flexneri 2a purEplus E. coli K-12 purE+ [transduced]); 3, G108-1-13 (S. flexneri 2apurE plus E. coli K-12 purE+ [conjugated]); 4, G256-2 (S. flexneri 2apurE); 5, M4243 (S. flexneri 2a).
Figure 2 shows an immunoblot of S. flexneri hybrids thathad incorporated the purE region of the E. coli K-12 chro-mosome. Strain G108-1-13 (lane 3) had acquired the purEregion by conjugation, whereas strain G283B (lane 2) hadacquired the same chromosomal region by P1 vir transduc-tion (6). Compared with results for strains carrying the S.flexneri purE region (lanes 1, 4, and 5), less antibody wasbound to a 140-kDa protein in transductants carrying thepurE region from E. coli K-12.
Plaque formation and the kepA locus. The ability of E.coli-S. flexneri hybrids and S. flexneri transductants to formplaques in HeLa cell monolayers is summarized in Table 1.The plaque assays were repeated four times with three platesper dilution, and the average number of PFU per CFU in thestrains carrying both the pWR110 S. flexneri plasmid and thekepA locus (7275-1-6, 7293-2-2, 7194-2-3, and G283A) rangedfrom 2.5 x 1O-4 to 5.0 x 10-2. The plaque-forming efficiencywas not influenced by the His and Mtl chromosomal markersfrom S. flexneri. The 7293-2-2 hybrid, which carried both thehis and kepA loci from S. flexneri, was previously reportedto be a plaque-forming strain (23).The PFU per CFU of E. coli K-12 hybrids that did not
carry the shigella kcpA locus (7262-1-10, 7209-1-19, and7185-2-2) ranged from nondetectable to 10-6. Giemsa-stained monolayers indicated that these strains were able toinvade HeLa cells at a frequency which approximated thatobserved in the E. coli hybrids carrying the shigella kcpAlocus. Therefore, the low PFU per CFU of E. coli hybridsthat did not carry the shigella kcpA locus probably reflecteda difference in the intracellular behavior of these organisms.The occurrence of occasional plaques in HeLa cell mono-layers exposed to hybrids that did not carry the kcpA locusallowed the isolation of stable strains with a plaque-forming
FIG. 3. SDS-PAGE and immunoblot analysis of E. coli K-12-S.flexneri hybrids. Lanes: 1, 7262-1-10 (E. coli K-12 plus S. flexneri220-kb plasmid); 2, 7262-1-1OPP7 (plaque-purified variant of 7262-1-10). Positions of proteins a, b, c, and d are indicated.
1 2 3 4 5 6 7
140 kd
FIG. 4. SDS-PAGE and immunoblot analysis of E. ccli K-12-S.flexneri hybrids. Lanes: 1, 395-1 (E. ccli K-12); 2, 7262-1-24 (E. ccliK-12 plus S. flexneri 220-kb plasmid [tetra'cycline sensitive]); 3,7262-1-24PP2 (plaque-purified variant of 7262-1-24); 4, 7262-4-2(7262-1-24 plus 7262-1-24PP2 purE' [transduced from 7262-1-24PP2purE: :TnJO]); 5, 395-3A2 (E. ccli K-12 [rifampin resistant]plus 7262-1-24PP2 purE' [transduced from 7262-1-24PP2purE::Tn 10]); 6, 8165-2-2 (395-3A2 plus S. flexneri 220-kb plasmid); 7,M90T (S. flexneri 5).
INFECT. IMMUN.
on January 25, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
kcpA AND VIRULENCE IN S. FLEXNERI 481
. f ;,a jsr-. .,t* - *~ .
6,s41NsVI
,.82 Xs ; X Xi'..'.-.^t14~~~~~~~~~~~4
ol~~ ~ ~ ~ ~ ~
41,~~~~~~~~~~~~~~~~~~~ ,
I~~~~~~~~~~O
rk~~ ~ ~-~A
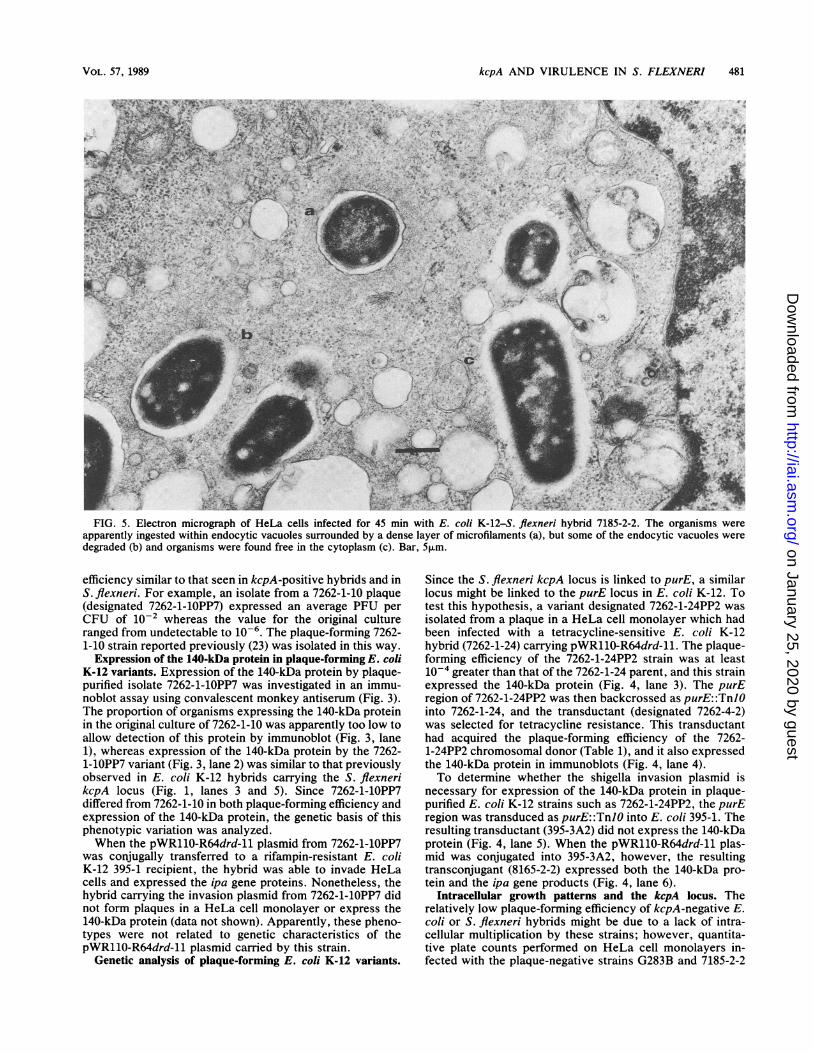
FIG. 5. Electron micrograph of HeLa cells infected for 45 min with E. coli K-12-S. flexneri hybrid 7185-2-2. The organisms wereapparently ingested within endocytic vacuoles surrounded by a dense layer of microfilaments (a), but some of the endocytic vacuoles weredegraded (b) and organisms were found free in the cytoplasm (c). Bar, 5,um.
efficiency similar to that seen in kcpA-positive hybrids and inS. flexneri. For example, an isolate from a 7262-1-10 plaque(designated 7262-1-10PP7) expressed an average PFU perCFU of 10-2 whereas the value for the original cultureranged from undetectable to 10-6. The plaque-forming 7262-1-10 strain reported previously (23) was isolated in this way.
Expression of the 140-kDa protein in plaque-forming E. coliK-12 variants. Expression of the 140-kDa protein by plaque-purified isolate 7262-1-10PP7 was investigated in an immu-noblot assay using convalescent monkey antiserum (Fig. 3).The proportion of organisms expressing the 140-kDa proteinin the original culture of 7262-1-10 was apparently too low toallow detection of this protein by immunoblot (Fig. 3, lane1), whereas expression of the 140-kDa protein by the 7262-1-10PP7 variant (Fig. 3, lane 2) was similar to that previouslyobserved in E. coli K-12 hybrids carrying the S. flexnerikcpA locus (Fig. 1, lanes 3 and 5). Since 7262-1-10PP7differed from 7262-1-10 in both plaque-forming efficiency andexpression of the 140-kDa protein, the genetic basis of thisphenotypic variation was analyzed.When the pWR110-R64drd-11 plasmid from 7262-1-1OPP7
was conjugally transferred to a rifampin-resistant E. coliK-12 395-1 recipient, the hybrid was able to invade HeLacells and expressed the ipa gene proteins. Nonetheless, thehybrid carrying the invasion plasmid from 7262-1-10PP7 didnot form plaques in a HeLa cell monolayer or express the140-kDa protein (data not shown). Apparently, these pheno-types were not related to genetic characteristics of thepWR11O-R64drd-11 plasmid carried by this strain.
Genetic analysis of plaque-forming E. coli K-12 variants.
Since the S. flexneri kcpA locus is linked to purE, a similarlocus might be linked to the purE locus in E. coli K-12. Totest this hypothesis, a variant designated 7262-1-24PP2 wasisolated from a plaque in a HeLa cell monolayer which hadbeen infected with a tetracycline-sensitive E. coli K-12hybrid (7262-1-24) carrying pWR110-R64drd-11. The plaque-forming efficiency of the 7262-1-24PP2 strain was at least10-4 greater than that of the 7262-1-24 parent, and this strainexpressed the 140-kDa protein (Fig. 4, lane 3). The purEregion of 7262-1-24PP2 was then backcrossed as purE: :TnJOinto 7262-1-24, and the transductant (designated 7262-4-2)was selected for tetracycline resistance. This transductanthad acquired the plaque-forming efficiency of the 7262-1-24PP2 chromosomal donor (Table 1), and it also expressedthe 140-kDa protein in immunoblots (Fig. 4, lane 4).To determine whether the shigella invasion plasmid is
necessary for expression of the 140-kDa protein in plaque-purified E. coli K-12 strains such as 7262-1-24PP2, the purEregion was transduced as purE: :TnJO into E. coli 395-1. Theresulting transductant (395-3A2) did not express the 140-kDaprotein (Fig. 4, lane 5). When the pWR110-R64drd-11 plas-mid was conjugated into 395-3A2, however, the resultingtransconjugant (8165-2-2) expressed both the 140-kDa pro-tein and the ipa gene products (Fig. 4, lane 6).
Intracellular growth patterns and the kcpA locus. Therelatively low plaque-forming efficiency of kcpA-negative E.coli or S. flexneri hybrids might be due to a lack of intra-cellular multiplication by these strains; however, quantita-tive plate counts performed on HeLa cell monolayers in-fected with the plaque-negative strains G283B and 7185-2-2
VOL. 57, 1989
on January 25, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

482 PAL ET AL.
.m
il .r::
:: :: :::.: .::
::: ..::
.:.: :.:. ::
*: ilS | -'.
:. 3P: ,,*:: :' # . .. , 5t.'* gs ' :. :! _:.fi£ :.': .::
:.:E.H..:#:: s.:w ^
4: ::f.ij* F
..... lgDr_DMs :::
INFECT. IMMUN.
..ffil**C 0 ct
S~ ~
CA
CZ
*Ni]i 0
A | M-. am,
z~~~~~~~~~~~~~~-.i'I-a
i I I |||| YAiC13~~~~~~~~~~~~~~~~~~2> 4c
Cd,
Xt~~~~~~~~
f's~~ ~ ~ ~ ~ ~ ... , :................
''..'.s' " ':".ss;'' ;
_SX..'.".'..''s ,: e=~~~~~~~~~C_ .Rm =z
_Ms =.~~~~~~~~~~~~~~~~C)
_ _ ,~~~~~~~~~~~~~~~~~~~C
r. . . r n;~~~~~~~~~~~r
CO.(A CZ
C =
...
on January 25, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
kcpA AND VIRULENCE IN S. FLEXNERI 483
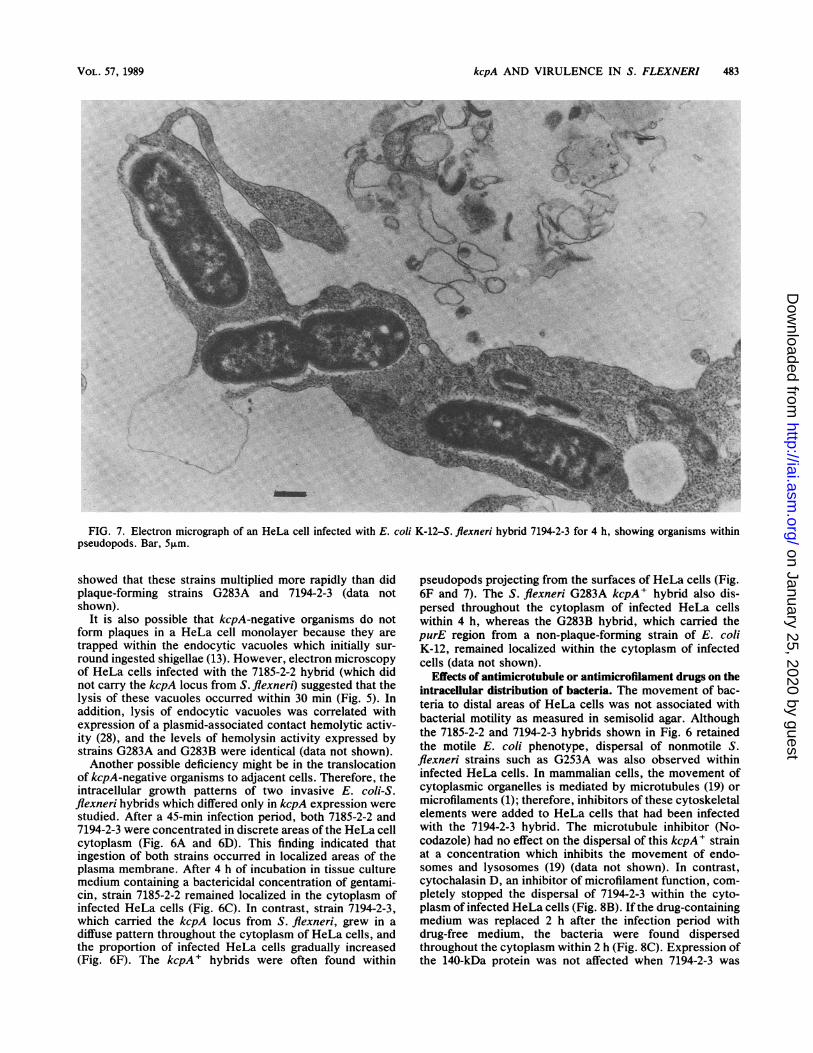
FIG. 7. Electron micrograph of an HeLa cell infected with E. coli K-12-S. flexneri hybrid 7194-2-3 for 4 h, showing organisms withinpseudopods. Bar, 5,m.
showed that these strains multiplied more rapidly than didplaque-forming strains G283A and 7194-2-3 (data notshown).
It is also possible that kcpA-negative organisms do notform plaques in a HeLa cell monolayer because they aretrapped within the endocytic vacuoles which initially sur-round ingested shigellae (13). However, electron microscopyof HeLa cells infected with the 7185-2-2 hybrid (which didnot carry the kcpA locus from S. flexneri) suggested that thelysis of these vacuoles occurred within 30 min (Fig. 5). Inaddition, lysis of endocytic vacuoles was correlated withexpression of a plasmid-associated contact hemolytic activ-ity (28), and the levels of hemolysin activity expressed bystrains G283A and G283B were identical (data not shown).Another possible deficiency might be in the translocation
of kcpA-negative organisms to adjacent cells. Therefore, theintracellular growth patterns of two invasive E. coli-S.flexneri hybrids which differed only in kcpA expression werestudied. After a 45-min infection period, both 7185-2-2 and7194-2-3 were concentrated in discrete areas of the HeLa cellcytoplasm (Fig. 6A and 6D). This finding indicated thatingestion of both strains occurred in localized areas of theplasma membrane. After 4 h of incubation in tissue culturemedium containing a bactericidal concentration of gentami-cin, strain 7185-2-2 remained localized in the cytoplasm ofinfected HeLa cells (Fig. 6C). In contrast, strain 7194-2-3,which carried the kcpA locus from S. flexneri, grew in adiffuse pattern throughout the cytoplasm of HeLa cells, andthe proportion of infected HeLa cells gradually increased(Fig. 6F). The kcpA + hybrids were often found within
pseudopods projecting from the surfaces of HeLa cells (Fig.6F and 7). The S. flexneri G283A kcpA' hybrid also dis-persed throughout the cytoplasm of infected HeLa cellswithin 4 h, whereas the G283B hybrid, which carried thepurE region from a non-plaque-forming strain of E. coliK-12, remained localized within the cytoplasm of infectedcells (data not shown).
Effects of antimicrotubule or antimicrofilament drugs on theintracellular distribution of bacteria. The movement of bac-teria to distal areas of HeLa cells was not associated withbacterial motility as measured in semisolid agar. Althoughthe 7185-2-2 and 7194-2-3 hybrids shown in Fig. 6 retainedthe motile E. coli phenotype, dispersal of nonmotile S.flexneri strains such as G253A was also observed withininfected HeLa cells. In mammalian cells, the movement ofcytoplasmic organelles is mediated by microtubules (19) ormicrofilaments (1); therefore, inhibitors of these cytoskeletalelements were added to HeLa cells that had been infectedwith the 7194-2-3 hybrid. The microtubule inhibitor (No-codazole) had no effect on the dispersal of this kcpA' strainat a concentration which inhibits the movement of endo-somes and lysosomes (19) (data not shown). In contrast,cytochalasin D, an inhibitor of microfilament function, com-pletely stopped the dispersal of 7194-2-3 within the cyto-plasm of infected HeLa cells (Fig. 8B). If the drug-containingmedium was replaced 2 h after the infection period withdrug-free medium, the bacteria were found dispersedthroughout the cytoplasm within 2 h (Fig. 8C). Expression ofthe 140-kDa protein was not affected when 7194-2-3 was
VOL. 57, 1989
on January 25, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

484 PAL ET AL.
.
..
-ad
-
....
!1
4 *:e
...I
fAw~~~~
::go*-k 'A:-
...*s: ..:
44
.x":*
.:.. *.;..jt
Tiu. *41
coCCC
0 0=
_'4 F - s
uco
90.
* O )-
C 0
CaCZ
CoCuCZ
.. _
CZa
* o 2
O~C Cr
o4-'0
a) CoC_a._C
C-.-C*:r~~~~~, a)Q
0..Y
a).SD *_' 0,
h? QQa)O
E--
j '-Co a
C.. - 2
Sf m0~~~~~~~.
1-
Sr *tI--ttqI..4.pw 4s1A!,I
lb
0 ,*
*^s: ..
,w..f ;~
p-
INFECT. IMMUN.
..
on January 25, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

kVpA AND VIRULENCE IN S. FLEXNERI 485
grown in Penassay broth containing 25 ,ug of cytochalasin Dper ml (data not shown).
DISCUSSION
A characteristic pathological feature of bacillary dysen-tery is ulcerative lesions in the colonic mucosa which allowerythrocytes and inflammatory cells to pass into the lumen ofthe bowel. These ulcers form in localized areas of thesuperficial intestinal epithelium as a result of infection byshigellae (9). A simplified model of such shigella infectionsuses confluent monolayers of cultured mammalian cells assurrogates for intestinal epithelium. When shigellae invadeindividual tissue culture cells and spread to contiguous cellsin the monolayer, the resulting plaque is an in vitro approx-imation of the colonic ulcers of bacillary dysentery. Ourstudy shows that the ability to form plaques is associatedwith the purE-linked S. flexneri chromosomal locus desig-nated kcpA and the 220-kb shigella invasion plasmid. Thesegenetic elements are also necessary for the expression of a140-kDa immunogenic bacterial protein. The HeLa cellplaque assay also allowed selection of variants of E. coliK-12 which express some attributes of kcpA' strains, i.e.,plaque formation and synthesis of a 140-kDa immunogenicprotein. Backcrossing the purE chromosomal region from aplaque-purified E. coli donor into a non-plaque-formingrecipient confirmed that these attributes are also associatedwith the purE region of E. coli K-12.The studies of Sasakawa et al. (30) show that TnS inser-
tions into an S. flexneri 2a plasmid gene designated virGresult in a Sereny-negative phenotype. These mutants alsoremain localized within discrete areas of the cytoplasm ofinfected monkey kidney cells. The phenotypic similaritybetween virG mutants and the kcpA-negative strains de-scribed here suggests that the plasmid gene which interactswith the kcpA locus is virG. The recent studies of Sakai et al.(26) have suggested that an immunogenic 130-kDa protein isthe product of virG, and they have shown that the expressionof this protein is positively regulated by another plasmidlocus, designated virF. The 3 to 4-kb cistron size reported forvirG (17) would be consistent with a large protein product. Itshould be noted, however, that Sakai et al. cloned virG intoan S. flexneri background which would presumably containthe kepA chromosomal locus. Therefore, the possibilityremains that both virG and virF regulate a kepA structuralgene for the 140-kDa protein. Nonetheless, it is likely thatthe product of the virG gene is a large immunogenic proteinof 130 or 140 kDa. Expression of virG is probably regulatedby the virF plasmid gene and by genes linked to the purElocus in S. flexneri or in plaque-forming variants of E. coliK-12.
Residual expression of a 140-kDa protein in S. flexnerihybrids carrying the purE region from an E. coli K-12 strainthat was not plaque purified (i.e., strains G283B and G108-1-13 in Fig. 2) suggests that control of this protein is lessstringent in S. flexneri than in E. coli K-12. Nonetheless,these hybrids were unable to form plaques in HeLa cellmonolayers or provoke keratoconjunctivitis in the guinea pigeye. This attenuated phenotype indicates that induction of a140-kDa protein by kcpA is crucial for expression of viru-lence in S. flexneri.
Plaque formation in HeLa cell monolayers was associatedwith intracellular bacterial movement; such polar-oriented,unidirectional movement of S. flexneri within HeLa cellswas first observed by Ogawa et al., who used cinemicro-graphic techniques (24). Since intracellular spread of shigel-
lae is independent of flagellar motility, the organisms prob-ably employ endogenous transport systems of the host cell.Cellular organelles such as lysosomes and endosomes movealong microtubules within the cytoplasm of cultured ratkidney cells, and this activity is inhibited by Nocodazole(19). However, treatment ofHeLa cells with Nocodazole didnot inhibit the intracellular spread of E. coli K-12 hybridscarrying the shigella invasion plasmid and the kcpA locus.Another transport system for cellular organelles is depen-dent on actin filaments (1, 19). Treatment of infected HeLacells with cytochalasin D, a fungal metabolite which causessustained contracture of microfilaments (21), effectivelystopped the intracellular spread of shigellae. These observa-tions suggest that shigellae (or E. coli-S. flexneri hybridscarrying the invasion plasmid and the kcpA locus) interactwith host cell microfilaments in a way that facilitates thedispersal of bacteria within the cytosol.Ogawa et al. (24) reported that intracellular shigellae often
impinged on the inner surface of the plasma membrane andproduced protrusions from the body of the cell. In our study,kcpA-positive E. coli K-12 hybrids were also found withinpseudopod-like protrusions both in photomicrographs and inelectron micrographs of fixed HeLa cells. It is possible thatthese pseudopods are involved in the intercellular spread ofshigellae to contiguous host cells.
Since the movement of shigellae within HeLa cells isslowed by tetracycline, which inhibits protein synthesis inintracellular bacteria (24), bacterial proteins such as the140-kDa species may be directly involved in intracellularmotility. This protein is apparently exposed on the bacterialsurface, because it is not detectable in immunoblots oftrypsinized shigellae (T. L. Hale, unpublished data). Al-though the 140-kDa protein induces an antibody response inthe colonic mucosa of infected monkeys (4), this antibodywould not be expected to neutralize a virulence determinantthat functions exclusively within the cytosol of infectedepithelial cells. Therefore, antibody recognizing the 140-kDaprotein may not protect against the ulcerative colonic lesionswhich characterize shigella infections.
ACKNOWLEDGMENTS
We gratefully thank Philippe J. Sansonetti and Edwin V. Oaks forhelpful suggestions and for critical reading of the manuscript.
T.P. was a National Research Council research associate atWalter Reed Army Institute of Research.
LITERATURE CITED1. Adams, R. J., and T. D. Pollard. 1986. Propulsion of organelles
isolated from Acanthamoeba along actin filaments by myosin-I.Nature (London) 322:754-756.
2. Baudry, B., A. T. Maurelli, P. Clerc, J. C. Sadoff, and P. J.Sansonetti. 1987. Localization of plasmid loci necessary for theentry of Shigella flexneri into HeLa cells, and characterizationof one locus encoding four immunogenic proteins. J. Gen.Microbiol. 133:3403-3413.
3. Buysse, J. M., C. K. Stover, E. V. Oaks, M. Venkatesan, andD. J. Kopecko. 1987. Molecular cloning of invasion plasmidantigen (ipa) genes from Shigella flexneri: analysis of ipa geneproducts and genetic mapping. J. Bacteriol. 169:2561-2569.
4. Dinari, G., T. L. Hale, S. W. Austin, and S. B. Formal. 1987.Local and systemic antibody responses to Shigella infection inrhesus monkeys. J. Infect. Dis. 155:1065-1069.
5. Formal, S. B., P. Gemski, Jr., L. S. Baron, and E. H. LaBrec.1970. Genetic transfer of Shigella flexneri antigens to Esche-richia coli K-12. Infect. Immun. 1:279-287.
6. Formal, S. B., P. Gemski, Jr., L. S. Baron, and E. H. LaBrec.1971. A chromosomal locus which controls the ability of Shi-gella flexneri to evoke keratoconjunctivitis. Infect. Immun.
VOL. 57, 1989
on January 25, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

486 PAL ET AL.
3:73-79.7. Formal, S. B., T. L. Hale, C. Kapfer, J. P. Cogan, P. J. Snoy, R.
Chung, M. E. Wingfield, B. L. Elisberg, and L. S. Baron. 1984.Oral vaccination of monkeys with an invasive Escherichia coliK-12 hybrid expressing Shigella flexneri 2a somatic antigen.Infect. Immun. 46:465-469.
8. Griffiths, E., P. Stevenson, T. L. Hale, and S. B. Formal. 1985.Synthesis of aerobactin and a 76,000-dalton iron-regulated outermembrane protein by Escherichia coli K-12-Shigella flexnerihybrids and by enteroinvasive strains of Escherichia coli. In-fect. Immun. 49:67-71.
9. LaBrec, E. H., H. Schneider, T. J. Magnani, and S. B. Formal.1964. Epithelial cell penetration as an essential step in thepathogenesis of bacillary dysentery. J. Bacteriol. 88:1503-1518.
10. Hale, T. L. 1986. Invasion of epithelial cells by shigellae. Ann.Inst. Pasteur 137A:311-314.
11. Hale, T. L. 1987. Organization and expression of plasmidvirulence genes in Shigella flexneri, p. 253-265. In M. A.Horwitz (ed.), Bacteria-host cell interaction. Alan R. Liss, Inc.,New York.
12. Hale, T. L., and S. B. Formal. 1981. Protein synthesis in HeLaor Henle 407 cells infected with Shigella dysenteriae 1, Shigellaflexneri 2a, or Salmonella typhimurium W118. Infect. Immun.32:137-144.
13. Hale, T. L., R. E. Morris, and P. F. Bonventre. 1979. Shigellainfection of Henle intestinal epithelial cells: role of the host cell.Infect. Immun. 24:887-894.
14. Hale, T. L., E. V. Oaks, and S. B. Formal. 1985. Identificationand antigenic characterization of virulence-associated, plasmid-coded proteins of Shigella spp. and enteroinvasive Escherichiacoli. Infect. Immun. 50:620-629.
15. Hale, T. L., E. V. Oaks, S. B. Formal, G. Dinari, and P.Echeverria. 1986. Immune response to Shigella infections and toShigella vaccines, p. 181-185. In F. Brown, R. M. Chanock,and R. A. Lerner (ed.), Vaccines 86. Cold Spring HarborLaboratory, Cold Spring Harbor, N.Y.
16. Hale, T. L., P. J. Sansonetti, P. A. Schad, S. Austin, and S. B.Formal. 1983. Characterization of virulence plasmids and plas-mid-associated outer membrane proteins in Shigella flexneri,Shigella sonnei, and Escherichia coli. Infect. Immun. 40:340-350.
17. Makino, S., C. Sasakawa, K. Kamata, T. Kurata, and M.Yoshikawa. 1986. A genetic determinant required for continuousreinfection of adjacent cells on large plasmid in S. flexneri 2a.Cell 46:551-555.
18. Maloy, S. R., and W. D. Nunn. 1981. Selection for loss oftetracycline resistance by Escherichia coli. J. Bacteriol. 145:1110-1112.
19. Matteoni, R., and T. E. Kreis. 1987. Translocation and cluster-ing of endosomes and lysosomes depends on microtubules. J.Cell Biol. 105:1253-1265.
20. Maurelli, A. T., B. Baudry, H. d'Hauteville, T. L. Hale, andP. J. Sansonetti. 1985. Cloning of plasmid DNA sequencesinvolved in invasion of HeLa cells by Shigella flexneri. Infect.Immun. 49:164-171.
21. Miranda, A. F., G. C. Godman, A. D. Deitch, and S. W.Tanenbaum. 1974. Action of cytochalasin D on cells of estab-lished lines. J. Cell Biol. 61:481-500.
22. Oaks, E. V., T. L. Hale, and S. B. Formal. 1986. Serum immuneresponse to Shigella protein antigens in rhesus monkeys andhumans infected with Shigella spp. Infect. Immun. 53:57-63.
23. Oaks, E. V., M. E. Wingfield, and S. B. Formal. 1985. Plaqueformation by virulent Shigella flexneri. Infect. Immun. 48:124-129.
24. Ogawa, H., A. Nakamura, and R. Nakaya. 1968. Cinemicro-graphic study of tissue cell cultures infected with Shigellaflexneri. Jpn. J. Med. Sci. Biol. 21:259-273.
25. Reynolds, E. S. 1963. The use of lead citrate at high pH as anelectronopaque stain in electron microscopy. J. Cell Biol.17:298-342.
26. Sakai, T., C. Sasakawa, and M. Yoshikawa. 1988. Expressionfour virulence antigens of Shigella flexneri is positively regu-lated at the transcriptional level by the 30 kilodalton virFprotein. Mol. Microbiol. 2:589-597.
27. Sansonetti, P. J., T. L. Hale, G. J. Dammin, C. Kapfer, H. H.Collins, Jr., and S. B. Formal. 1983. Alterations in the pathoge-nicity of Escherichia coli K-12 after transfer of plasmid andchromosomal genes from Shigella flexneri. Infect. Immun. 39:1392-1402.
28. Sansonetti, P. J., A. Ryter, P. Clerc, A. T. Maurelli, and J.Mounier. 1986. Multiplication of Shigella flexneri within HeLacells: lysis of the phagocytic vacuole and plasmid-mediatedcontact hemolysis. Infect. Immun. 51:461-469.
29. Sasakawa, C., K. Kamata, T. Sakai, S. Makino, M. Yamada, N.Okada, and M. Yoshikawa. 1988. Virulence-associated geneticregions comprising 31 kilobases of the 230-kilobase plasmid inShigella flexneri 2a. J. Bacteriol. 170:2480-2484.
30. Sasakawa, C., S. Makino, K. Kamata, and M. Yoshikawa. 1986.Isolation, characterization, and mapping of Tn5 insertions intothe 140-megadalton invasion plasmid defective in the mouseSereny test in Shigella flexneri 2a. Infect. Immun. 54:32-36.
INFECT. IMMUN.
on January 25, 2020 by guesthttp://iai.asm
.org/D
ownloaded from


















