Intracellular highways in the plants: the role of the cytoskeleton in camv infections
76
Intracellular highways in the plants: the role of the cytoskeleton in CaMV infections. James Schoelz
Transcript of Intracellular highways in the plants: the role of the cytoskeleton in camv infections

Intracellular highways in the plants: the role of the cytoskeleton in CaMV infections.
James Schoelz

42 Division of Plant Sciences Faculty 101 Graduate Students
34 MS67 PhD

Five Program Areas Crop, Soil and Pest ManagementEntomologyHorticulturePlant Stress BiologyPlant Breeding, Genetics and Genomics

Plant Sciences Biological SciencesBiochemistryComputer ScienceForestry ChemistryUSDA-ARS Plant Genetics Unit
The IPG --- who we are…• Established in 1981 to develop a program of excellence in
plant science• 58 faculty teams; recognized for interactive and cooperative
research and education community• 2 National Academy of Science members• 18 Fellows of the American Association for the Advancement
of Science (AAAS)
In 2010, MU was ranked 14th in the world among universities for plant science research (Times Higher Education)

Building on a strong history in plant genetics

INTERDISCIPLINARY PLANT GROUP
U N I V E R S I T Y O F M I S S O U R I
A RABIDOPSISC O L U M B I A W I L D T Y P E
PROF. GEORGE RÉDEI (1921-2008)

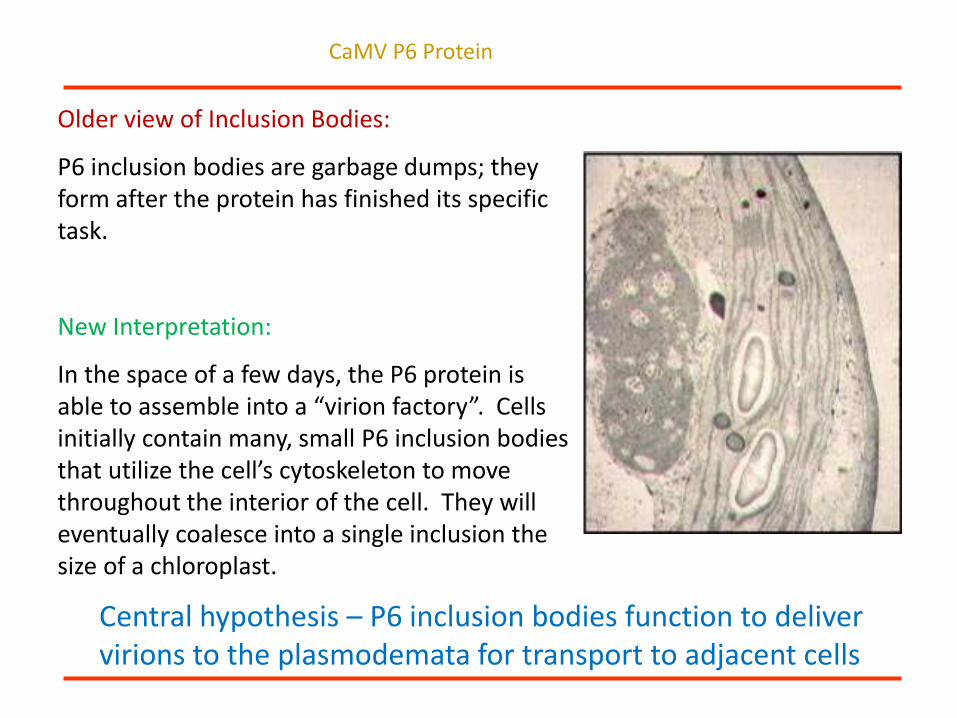
Older view of Inclusion Bodies:
P6 inclusion bodies are garbage dumps; they form after the protein has finished its specific task.
CaMV P6 Protein
Older view of Inclusion Bodies:
P6 inclusion bodies are garbage dumps; they form after the protein has finished its specific task.
New Interpretation:
In the space of a few days, the P6 protein is able to assemble into a “virion factory”. Cells initially contain many, small P6 inclusion bodies that utilize the cell’s cytoskeleton to move throughout the interior of the cell. They will eventually coalesce into a single inclusion the size of a chloroplast.
Central hypothesis – P6 inclusion bodies function to deliver virions to the plasmodemata for transport to adjacent cells
CaMV P6 Protein

What Every Plant Virus Must Do to Survive
R
(-)3'
(+)5'
5 ' (+)
cp cp cp
cp cp
(+ )5'
(- )
(+ )5'R
RR R R R
RR
5' (+)
Replicate Progeny
Genomes
Move to Other Plants
Uncoat the
Viral Genome
Express Proteins
needed for genome
Replication.
Enter the Cell
Produce proteins needed
for Virion formation and
Cell to cell MovementMove through
the plant
Assemble Virions

Cauliflower Mosaic virus
Source of the the 35S promoter
First plant virus genome to be sequenced – 1981
First plant virus to be cloned in infectious form – 1981
First viral avirulence gene i.e. a pathogen gene that triggers a plant defense response) to be characterized by recombinant DNA techniques – P6 - 1984

Genome Structure of Cauliflower Mosaic virus
Gene I - cell-to-cell movement
Gene II - aphid
transmission
Gene III - DNA binding;
aphid transmission
Gene IV - coat protein
V - reverse
transcriptase
VI - Translational
transactivator/Forms
inclusion bodies.
CaMV
8.0 kb
(Modified from Haas et al., 2002)

Turnip leaf epidermal
strip stained with
phloxine B illustrates
CaMV inclusions
Electron micrograph of a
turnip leaf cell infected with
CaMV

Viruses have developed Movement Proteins (MPs) to facilitate transport through plasmodesmata
Class I
Class IIVirions
Major Movement Classes : MP:RNA complexes & Virions

Tubules mediate cell-to-cell movement of virions of some
virusesTubule-like structures (aprox. 35 nm dia) containing a single row of virions are present in plants infected by CPMV.
See Pouwels et al., Mol Plant Path. 3:411 (2002).

Tubules Mediate Cell-to-cell Movement of Virions
Tubule-like structures (aprox. 35 nm dia) containing a single row of virions are present in plants infected by CPMV.
Movement proteins are responsible for tubule formation
See Pouwels et al., Mol Plant Path. 3:411 (2002).

Tubules Mediate Cell-to-cell Movement of Virions
Tubule-like structures (aprox. 35 nm dia) containing a single row of virions are present in plants infected by CPMV.
Movement proteins are responsible for tubule formation
Tubules containing virions have also been observed for:NepovirusesTospovirusesCaulimoviruses
See Pouwels et al., Mol Plant Path. 3:411 (2002).

Where are CaMV P6 inclusion bodies
found within the cell?

The Agroinfiltration Technique
1. Grow Agrobacterium tumefaciens
carrying the desired binary vector in 1.0
ml of broth overnight at 28 C in a
shaker/incubator.
2. Add acetosyringone, a signal produced by wounded plants that mobilizes the
transfer of the T-DNA of the binary vector into plant cells. Incubate overnight.
3. Fill a 5.0 ml syringe with Agrobacterium cells, apply the syringe to the
surface of a leaf, and gently infiltrate the solution into plant tissue.

Agroinfiltration of Gene VI of from different
CaMV strains
W260 Gene VI
D4 Gene VI
pW260VI rbcs35S
rbcs35SpD4VI

P6 inclusions are associated with actin
microfilaments, as well as ER and
microtubules.
P6-GFP
dsRed2-talin (labels thick
actin cables)
Co-agroinfiltrationof P6-GFP and
dsRed2-talin into N. benthamiana
and visualization of P6/actin
association.
Harries et al. (2009) The Cauliflower mosaic virus protein P6 forms motile inclusions that traffic
along actin microfilaments and stabilize microtubules. Plant Physiology 149, 1005-1016.
Phil Harries

P6 inclusion bodies can traffic along actin filaments.
Harries P et al. Plantphysiol 2009;149:1005-1016
©2009 by American Society of Plant Biologists

CaMV P6 inclusion bodies move on microfilaments

The actin inhibitor LatB inhibits CaMV infection.

Virions are assembled in the P6 inclusion bodies and most of the CaMV virions remain in the inclusion bodies.
P6 Inclusion Bodies as “Virion Factories”
Consequently, for CaMV virions to move from cell-to-cell there must be some mechanism for transfer of virions from the P6 inclusion body, the site for viral synthesis and virion assembly, to the plasmodesmata.

Nucleus
Model for P6 motility function
during CaMV infection
35S RNA
P6 protein
Microtubule
Microfilament
Virion
CaMV/P6
Inclusion

What host proteins interact with the P6
protein of CaMV?
What host proteins might potentially
facilitate intracellular movement to
plasmodesmata?

Previous yeast two-hybrid screens focused on
the role of P6 in translational transactivation and
silencing suppression
Eukaryotic translation factor 3 subunit g (eIF3g)(Park et al. 2001)
Ribosomal subunits L13, L18, and L24 (Leh et al., 2000)
RNA silencing protein DRB4 (Haas et al. 2008)

Interactions of P6 with virus and host proteins – Results of a Y2H Screen
Eighteen proteins identified in a total of 85 clones
Ribosomal initiation factor eIF3 ( represented in 17 independent clones)
Cytoskeleton-related
CHUP1 – interacts with actin for movement of chloroplasts on
microfilaments (represented in one clone)
Membrane Associations
AtSRC2.2 – C2 calcium-dependent membrane targeting protein
kinase C (represented in 17 independent clones)

CHUP1 is responsible for chloroplast movement within the cell in response to light intensity
Accumulation Avoidance
Low Light High Light
Redrawn from Wada (2013). Chloroplast movement. Plant Science 210, 177-182

Col-0 T-DNA line
129128C
White Band Assay
Analysis of Arabidopsis thaliana CHUP1 T-
DNA knockouts

Localization of CHUP1-CFP to the outer membrane of chloroplasts in N. benthamiana leaves
CHUP1-eCFP Chloroplasts
Merged
10 µm
Carlos Angel
Angel et al. 2013. The P6 protein of Cauliflower mosaic virus interacts with CHUP1, a plant protein which moves chloroplasts on actin microfilaments. Virology 443, 363-374.

Co-localization of CHUP1 with P6-Venus(YFP)
10 µm
MergedChloroplasts
CHUP1-eCFP P6-Venus(YFP)

Sample
CHUP1-GFP
P6-GFP
P6-RFP
+
-
-
-
+
-
-
-
+
+
-
+
-
+
+
72kDa
95kDa
72kDa
95kDa
TotalBlot: αGFP
TotalBlot: αRFP
72kDa
95kDa
Co-IP: αGFPBlot: αRFP
1 2 3 4 5
Sample
CHUP1-GFP
P6-GFP
P6-RFP
+
-
-
-
+
-
-
-
+
+
-
+
-
+
+
1 2 3 4 5
Co-Immunoprecipitation of P6-RFP with the truncated CHUP1-GFPCo-Immunopreciptation of P6-RFP with P6-GFP

Dominant Negative inhibition of chloroplast movement
Expression of a truncated CHUP1 protein consisting of
the first 500 amino acids, abolished the movement of
chloroplasts in Arabidopsis plants, acting as a
dominant negative inhibitor (Oikawa et al., 2008).
Question:
Since our CHUP1-CFP construct consists of the 436 first
amino acids, can we block the movement of P6 –
Venus YFP inclusion bodies?

Time lapse imaging of P6- Venus YFP inclusion bodies

Time lapse imaging of AtCHUP-eCFP and P6- Venus YFP inclusion bodies
(Angel et al., 2012, submitted)

Question
What effect does the loss of CHUP1 have on CaMV
infections?

Experimental Design for Chup1 VIGS in N. edwardsonii
and inoculation of CaMV virions
CaMV virions inoculation21 days
5-6 week old plants
CaMV HRlocal lesions development
The Hypersensitive Response (HR – programmed cell death) in N. edwardsonii.
Necrotic local lesions induced by CaMV strain W260 in inoculated leaves. This virus is
unable to move systemically because N. edwardsonii mounts an effective defense.
Question:
Will lesion number or size be affected in
N. edwardsonii CHUP1 silenced plants?

Percentage of necrotic lesion development in N. edwardsonii : lesions
develop at a slower rate in CHUP1- silenced plants than in controls
(ANOVA, p=0.01).

The Association of CHUP1 with CaMV P6
Confirmed with Y2H, co-IP and co-
localization in vivo
Silencing of CHUP1 in N. benthamiana
significantly slows the rate of CaMV lesion
development.
Transient expression of a truncated CHUP1
blocks the movement of chloroplasts (Oikawa
et al. 2003) and P6 inclusion bodies (Angel
et al. 2013).

Nucleus
P6 protein
Microtubule
Microfilament
Virion
CaMV/P6
Inclusion
CHUP1 – May explain the interaction of P6 inclusion bodies with microfilaments, and participates in movement of P6 inclusion bodies on microfilaments.
CHUP1
Angel et al. 2013. The P6 protein of Cauliflower mosaic virus interacts with CHUP1, a plant protein which moves chloroplasts on actin microfilaments. Virology 443, 363-374.

Question
Are CaMV P6 inclusion bodies associated with
plasmodesmata?
Andres Rodriguez

Interactions of P6 with virus and host proteins – Results of a Y2H Screen
Eighteen proteins identified in a total of 85 clones
Ribosomal initiation factor eIF3 ( represented in 17 independent clones)
Cytoskeleton-related proteins
CHUP1 – interacts with actin for movement of chloroplasts on
microfilaments (represented in one clone)
Membrane Associations
AtSRC2.2 – C2 calcium-dependent membrane targeting protein
kinase C (represented in 17 independent clones)
SRC2.2 = Soybean Response to Cold

Three experimental techniques indicate an association
of AtSRC2.2 with CaMV P6
Y2H
Co-IP
Co-localization in vivo
Where is AtSRC2.2 found in the cell?

AtSCR2.2-RFP PDLP1-GFP
Cell wall
Plasma membrane
Overlay
Plasmolyzed cell
A portion of AtSRC2.2-RFP co-localizes with the plasmodesmal protein PDLP1
Co-agroinfiltration of PDLP1-GFP with AtSCR2.2-RFP into N. benthamiana leaves

AtSCR2.2-RFPP1-GFP
Overlay
A portion of AtSRC2.2-RFP co-localizes with CaMV P1-GFP
CaMV P1 (the CaMV movement protein) is localized to plasmodesmata
Co-agroinfiltration of P1-GFP with AtSRC2.2-RFP into N. benthamiana leaves

P1-GFP P1-GFP + untagged P1
Formation of tubules is likely carried out by untagged P1. P1-GFP allows for the visualization of the tubules.

Conti G.G., Vegetti G., Bassi M., Favali M.A. 1972. Some Ultrastructural and CytochemicalObservations on Chinese Cabbage Leaves Infected with Cauliflower Mosaic Virus. Virology 47, 697-700.
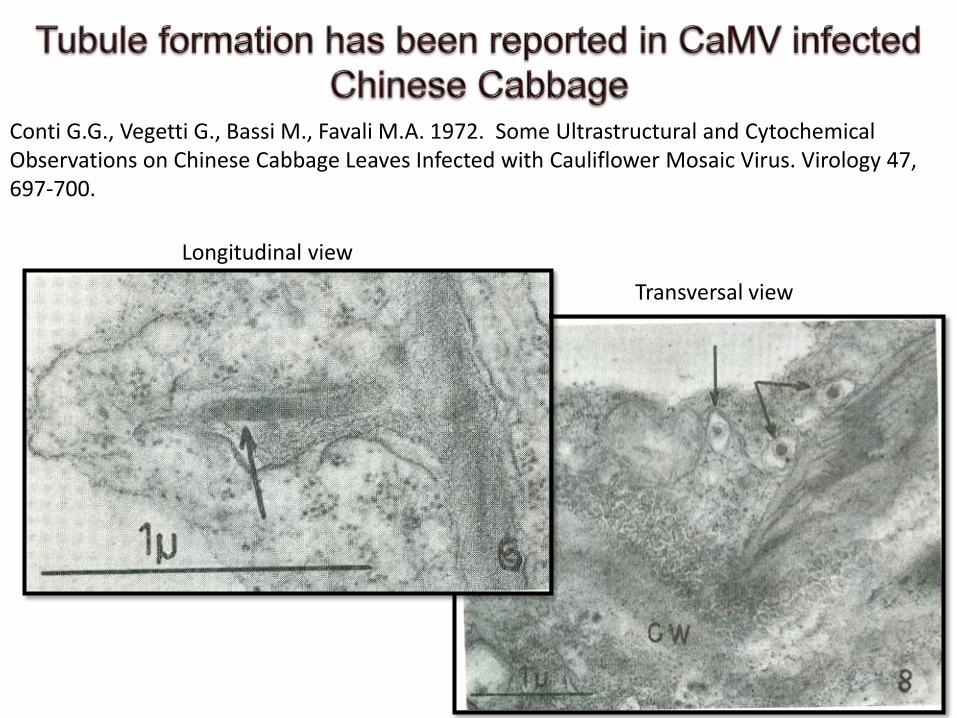
Longitudinal view
Transversal view
Conti G.G., Vegetti G., Bassi M., Favali M.A. 1972. Some Ultrastructural and CytochemicalObservations on Chinese Cabbage Leaves Infected with Cauliflower Mosaic Virus. Virology 47, 697-700.

AtSRC2.2 was co-localized with 43.1% of P1 tubules, with C2CDMT found at the base (80%) or within the tubule (20%).
AtSRC2.2-RFP MergedP1-GFP + Untagged P1

Question
Are P6 inclusion bodies associated with
plasmodesmata?

PDLP1-GFP P6-RFP MERGED
Some P6 inclusion bodies are found adjacent to plasmodesmal markers
Co-agroinfiltration of P6-RFP with PDLP1-GFP into N. benthamiana leaves

PDLP1-GFP P6-RFP MERGED

Aniline Blue P6-RFP MERGED
Thomas et al, 2008. Use of Aniline Blue as plasmodesmal marker
Some P6 inclusion bodies are found adjacent to plasmodesmal markers

P6
P1AtSRC2.2
Y2HCo-localization
in vivoCo-IP
Co-localization in vivo
Y2H (Hapiak et al, 2008)Co-localizationin vivo

Nucleus
P6 protein
Microtubule
Microfilament
Virion
CaMV/P6
Inclusion
CaMV P6 is capable of forming a complex with CaMV P1 protein, AtSRC2.2, PDLP1 at
plasmodesmata
CaMV MP (P1)
AtSRC2.2
PDLP1
Plasmodesma modified
into tubule by CaMV P1
Protein

A new function for CaMV P6: delivery of virions to
plasmodesmata
P6 inclusion bodies are considered the site for virion assembly and
accumulation
P6 inclusion bodies associate with and move on microfilaments
P6 inclusion body movement on microfilaments may be facilitated through an
interaction with CHUP1 and one or more myosins
At least a portion of P6 inclusion bodies are associated with
plasmodesmata
P6 appears to be capable to form a complex at the plasmodesmata with
CaMV P1 (the cell-to-cell movement protein), AtSRC2.2, and PDLP1.

Exactly what is the function of the CaMV P6 protein?
1. Is the matrix protein for the vacuolated, amorphous inclusion bodies: forms “virion
factories”
2. Avirulence gene product and symptom determinant
3. Translational transactivator: facilitates the translation of viral genes on the
polycistronic 35S RNA
4. A shuttle protein, moving between the cytoplasm and nucleus
5. A silencing suppressor
6. Regulates SA-mediated defenses and cell death pathways
7. Delivery of virions to plasmodesmata

Exactly what is the function of the CaMV P6 protein?
The “function” of P6 may be its capacity to interact with a broad range of host proteins.
The capacity to form indiscriminate associations with host proteins might be considered a survival mechanism to take advantage of whatever a host has to offer.
In this hypothesis, the composition of host proteins in the P6 inclusion bodies might be dynamic, constantly changing as the infection within the cell matures.
In

AcknowledgmentsSchoelz Lab (current and former members) Nelson Lab (current and former members)
Dr. Carlos Angel Dr. Philip Harries
Dr. Boovaraghan Balaji Dr. Xiaohua Yang
Dr. Andres Rodriguez Dr. Xin S. Ding
Yu Zhang Dr. Malay Saha
Sandra Valdes Bethany Bishop
Mohammad Fereidouni
Mustafa Adhab
Adam Adair
Dr. Scott M. Leisner and Lindy Lutz. (The University of Toledo, Toledo OH).
Dr. Michael Goodin and Kathleen Martin. (University of Kentucky, Lexington KY)
Dr. Aleksandr Jurkevic. (MU Molecular Cytology and Microscopy Core)
Dr. Howard Berg. (Cytology Core, The Donald W. Danforth Center, St. Louis MO).
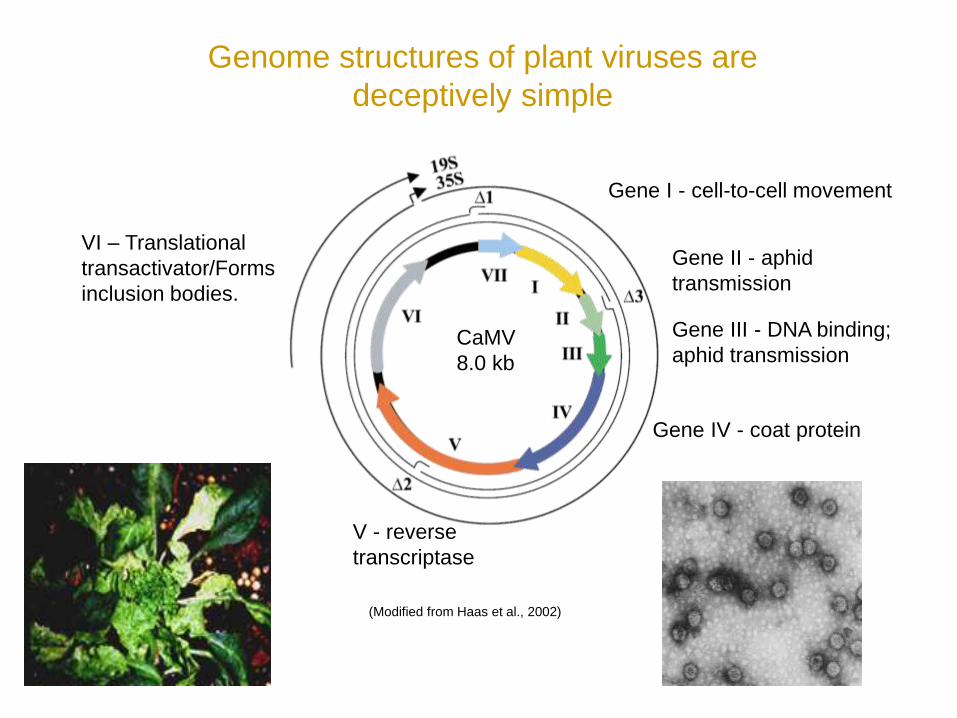
Genome structures of plant viruses are
deceptively simple
Gene I - cell-to-cell movement
Gene II - aphid
transmission
Gene III - DNA binding;
aphid transmission
Gene IV - coat protein
V - reverse
transcriptase
VI – Translational
transactivator/Forms
inclusion bodies.
CaMV
8.0 kb
(Modified from Haas et al., 2002)

D1 D4D3D2
Host Range,
Symptoms & P6
Self AssociationMini TAV
RNA
Binding
Zinc
Finger
P6 Self Association
Chloroplast
Interaction
Leucine
zipper
Proline-rich,
interacts with
Profilin/Actin Leucine zipper
Coiled-coil Actin binding
CaMV P6 binding
1004 aa
CHUP1 structure
P6 structure CHUP1
binding
What domains are responsible for the interaction of
CHUP1 and CaMV P6?

(Modified from Wada and Suetsugu 2004)
P6
Is there any relationship between CaMV P6 and
chloroplast movement?

P6-interacting region10 218
360 aa
C2 Domain
6 112 189
Pro-Rich region
320
Cho & Stahelin, 2006
C2 domain is the second most abundant lipid binding domain.
C2-domain-calcium dependent proteins are involved in signal transduction and membrane trafficking.

P6-GFP C2CDMT-RFP Overlay

P6-GFP - - + +
Initial ExtractBlot: αRFP
Co-IP: αGFPBlot: αRFP
Co-IP: αGFPBlot: αGFP
C2CDMT-RFP - + - +
Initial ExtractBlot: αGFP
70KDa
90KDa
70 kDa
90 kDa
1 2 3 4
Sample
B
A
C
D
C2CDMT-RFP is co-immunoprecipitated with P6-GFP

Three experimental techniques indicate an association
of C2CDMT with CaMV P6
Y2H
Co-IP
Co-localization in vivo
Where is C2CDMT found in the cell?

Question
Do myosins contribute to intracellular movement of
CaMV?
Andres Rodriguez
Carlos Angel
Yu Zhang

(Peremyslov et al., 2011)
Arabidopsis Myosins implicated in plant virus
movement through T-DNA knockouts or silencing

Development of CaMV infections in an A. thaliana chup1 T-
DNA knockout line versus wild type Col-0
Symptoms in the AtCHUP1 T-DNA knockout line were delayed, but CaMV
systemic symptom development was not abolished.
Days Post Inoculation (dpi)P
erc
en
tage o
f P
lan
ts w
ith
Syste
mic
Sym
pto
ms
0
10
20
30
40
50
60
70
80
9 10 11 12 13
Col-0
CHUP1
0
10
20
30
40
50
60
70
80
90
100
13 14 15 16 17 18 19 20 21 22 23 24 25 26 27
Col-0
CHUP
Days Post Inoculation (dpi)
Pe
rcen
tage o
f P
lan
ts w
ith
Lo
ca
l L
esio
ns

0
10
20
30
40
50
60
70
80
90
100
7 8 9 10 11 12 13
Col-0
XI-20
20
40
60
80
100
120
10 11 12 13 14 15 16 17 18
Col
XI-2
Development of CaMV infections in an A. thaliana myosin XI-
2 T-DNA knockout line versus wild type Col-0
Pe
rcen
tage o
f P
lan
ts w
ith
Lo
ca
l L
esio
ns
Days Post Inoculation (dpi)Days Post Inoculation (dpi)P
erc
en
tage o
f P
lan
ts w
ith
Syste
mic
Sym
pto
ms
Primary lesion development in the myosin XI-2 T-DNA knockout line was
consistently delayed by one day, but CaMV symptom development was not
abolished.

Development of CaMV infections in the A. thaliana XI-2/chup1 double T-
DNA knockout line versus wild type Col-0
Pe
rcen
tage o
f P
lan
ts w
ith
Lo
ca
l L
esio
ns
Days Post Inoculation (dpi)Days Post Inoculation (dpi)P
erc
en
tage o
f P
lan
ts w
ith
Syste
mic
Sym
pto
ms
Primary lesion development in the double T-DNA knockout line was consistently
delayed by one day, and CaMV systemic symptom development was delayed by
one to three days.
0
10
20
30
40
50
60
70
80
90
100
9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
Col-0
CHUP/XI-2
0
10
20
30
40
50
60
70
80
90
100
1 2 3 4 5 6 7 8 9 10 11
Col-0
CHUP/XI-2

PDLP1-GFP - - + +
Initial ExtractBlot: αRFP
Co-IP: αGFPBlot: αRFP
Co-IP: αGFPBlot: αGFP
P6-RFP - + - +
Initial ExtractBlot: αGFP
60KDa
90KDa
90 kDa
60 kDa
1 2 3 4
Sample
B
A
C
D
P6-RFP is co-immunoprecipitated with PDLP1-GFP
Heavy chain IgG

Development of CaMV infections in a Atsrc2.2 A. thaliana T-DNA
knockout line and in the triple T-DNA knockout pdlp123
Two tests were completed to compare the susceptibility of a Atsrc2.2 T-
DNA knockout line to wild type Col-0. There was no difference in the
timing of local or systemic symptoms.
By Contrast
A delay in CaMV infections was observed in the triple T-DNA knockout
pdlp123. However, even with the triple knockout line, CaMV
infections were not abolished (Amari et al., 2010).
Amari et al. 2010. A family of plasmodesmal proteins with receptor-like properties for plant viral
movement proteins. PLoS Pathogens 6 (9) e1001119 1-10.
100
A

Percentage of necrotic lesion development in N. edwardsonii : lesions
develop at a slower rate in CHUP1- silenced plants than in controls
(ANOVA, p=0.01).

Development of Local Lesions by CaMV in N. edwardsoniiand in CHUP1-Silenced Plants
Number of HR lesions per leaf was not statistically different (ANOVA, p=0.05)
Lesi
on
Nu
mb
ers
per
Lea
f















![The Actin Cytoskeleton: Functional Arrays forUpdate on the Actin Cytoskeleton The Actin Cytoskeleton: Functional Arrays for Cytoplasmic Organization and Cell Shape Control1[OPEN] Dan](https://static.fdocuments.in/doc/165x107/5f0830197e708231d420c69d/the-actin-cytoskeleton-functional-arrays-update-on-the-actin-cytoskeleton-the-actin.jpg)