
INTRACELLULAR CENTRIFUGAL SEPARATION OF ORGANELLES...
16
INTRACELLULARCENTRIFUGALSEPARATION OFORGANELLESIN PHYC,OMYCES MARKOZALOKAR FromtheDivisionofBiology,CaliforniaInstituteofTechnology,Pasadena,California91109 .The author'spresentaddressisCentredeGenetiqueMoleculaire,CentreNationaldelaRecherche Scientifique,Gif-sur-Yvette,France ABSTRACT Livesporangiophoresof Phycomycesblakesleeanus werecentrifugedat35,000rpm .Thecell contentssedimentedintodistinctlayers,andeachlayerwasstudiedwithanelectronmicro- scopeandwithcytochemicalmethods .Thefollowinglayerswerefound(theirvolumesand theirdensitiesareshowninFig .3) :1 .polyphosphates ;2 .polyphosphatesandproteincrys- tals ;3 .glycogen ;4 .yellowlayerwithferritin ;5 .ribosomes ;6 .proteincrystals ;7 .mitochon- dria ;8 .mitochondriaandfibrils ;9.nuclei ;10 .endoplasmicreticulum ;11 .vesicles,mem- branes,andreticulum ;12 .vacuole ;13 .lipoproteins,membranes ;14.fatdroplet .Theden- sitiesofthevariouslayersweredeterminedbytheinjectionofdropletsofinertoilsofknown densityintothesporangiosphoresbeforecentrifugation .Sedimentedcellorganellescouldbe isolated .Centrifugednucleiofalycopene-producingmutantwereinjectedintotheintact sporangiophoreofanalbinohostwheretheyinducedcolorformation .Theensuingspores, whenplated,gaveamixtureofwhiteandcoloredcolonies .Itwasconcludedthatcell organelles,sedimentedbycentrifugationoflivingsporangiophores,remainaliveandcanbe usedforbiochemicalstudies .Microspectrophotometricexaminationofthelayersindicated thepresenceofcytochromesandflavinesinthemitochondriaandofcytochromesinthe nuclei .Nopigmentscorrespondingtotheactionspectrumforthelightgrowthresponsewere found . INTRODUCTION Muchoftheprogressofmolecularbiologyisbased onstudiesofcellorganellesisolatedfromhomoge- nizedcellpreparations .Nomatterhow"gentle" thedisruptionofthecell,onecannotexpectthat isolatedparticlesarephysiologicallycompletely equivalenttoorganellesinthelivingcell .Although theisolatedparticlescontinuetoperformmany vitalfunctionsinvitro,theirratesofactivitiesare oftenonlyafractionofthoseoftheorganellesin undisturbedcells .Also,itispossiblethatsomeof theactivitiesarechangedqualitativelybyisola- tion .Thedamagedoneinisolatingparticlesmay beirreversible,andthereisnoproofthatthese particlescanfunctionnormallyifreintroduced intoalivingcell . 494 Ithasbeenoftenshownthatonecouldsediment cellorganellesinsidelivingcellsbyhighcentrifugal forcesandthatthecellscontinuedtofunctionnor- mallyaftertheorganelleswereredistributedinthe cytoplasm(BeamsandKing,1939 ;Zalokar,1959) . Onecan,therefore,expectthatcellorganelles, displacedbythecentrifugationoflivingcells, retainalltheirvitalcapacities,althoughtheir functionsmaybetemporarilymodifiedbecausethe organellesareseparatedfromeachother .Ifone couldisolateandrecombinedifferentcellorgan- elleswithoutactuallykillingthecell,onemight learnsomethingoftheirinteractionsandoftheir dependenceonthenormalstructuralorganization ofthecell.Studiesofsuchorganellescouldalso on August 22, 2006 www.jcb.org Downloaded from
Transcript of INTRACELLULAR CENTRIFUGAL SEPARATION OF ORGANELLES...

INTRACELLULAR CENTRIFUGAL SEPARATION
OF ORGANELLES IN P H Y C, O M Y C E S
MARKO ZALOKAR
From the Division of Biology, California Institute of Technology, Pasadena, California 91109 . Theauthor's present address is Centre de Genetique Moleculaire, Centre National de la RechercheScientifique, Gif-sur-Yvette, France
ABSTRACTLive sporangiophores of Phycomyces blakesleeanus were centrifuged at 35,000 rpm . The cellcontents sedimented into distinct layers, and each layer was studied with an electron micro-scope and with cytochemical methods . The following layers were found (their volumes andtheir densities are shown in Fig . 3) : 1 . polyphosphates ; 2 . polyphosphates and protein crys-tals ; 3 . glycogen ; 4 . yellow layer with ferritin ; 5 . ribosomes ; 6 . protein crystals ; 7 . mitochon-dria ; 8 . mitochondria and fibrils ; 9. nuclei ; 10 . endoplasmic reticulum ; 11 . vesicles, mem-branes, and reticulum ; 12 . vacuole ; 13 . lipoproteins, membranes ; 14. fat droplet . The den-sities of the various layers were determined by the injection of droplets of inert oils of knowndensity into the sporangiosphores before centrifugation . Sedimented cell organelles could beisolated . Centrifuged nuclei of a lycopene-producing mutant were injected into the intactsporangiophore of an albino host where they induced color formation . The ensuing spores,when plated, gave a mixture of white and colored colonies . It was concluded that cellorganelles, sedimented by centrifugation of living sporangiophores, remain alive and can beused for biochemical studies . Microspectrophotometric examination of the layers indicatedthe presence of cytochromes and flavines in the mitochondria and of cytochromes in thenuclei . No pigments corresponding to the action spectrum for the light growth response werefound .
INTRODUCTION
Much of the progress of molecular biology is basedon studies of cell organelles isolated from homoge-nized cell preparations. No matter how "gentle"the disruption of the cell, one cannot expect thatisolated particles are physiologically completelyequivalent to organelles in the living cell . Althoughthe isolated particles continue to perform manyvital functions in vitro, their rates of activities areoften only a fraction of those of the organelles inundisturbed cells . Also, it is possible that some ofthe activities are changed qualitatively by isola-tion . The damage done in isolating particles maybe irreversible, and there is no proof that theseparticles can function normally if reintroducedinto a living cell .
494
It has been often shown that one could sedimentcell organelles inside living cells by high centrifugalforces and that the cells continued to function nor-mally after the organelles were redistributed in thecytoplasm (Beams and King, 1939 ; Zalokar, 1959) .One can, therefore, expect that cell organelles,displaced by the centrifugation of living cells,retain all their vital capacities, although theirfunctions may be temporarily modified because theorganelles are separated from each other . If onecould isolate and recombine different cell organ-elles without actually killing the cell, one mightlearn something of their interactions and of theirdependence on the normal structural organizationof the cell. Studies of such organelles could also
on August 22, 2006
ww
w.jcb.org
Dow
nloaded from

reinforce or modify our knowledge obtained fromconventional methods of molecular biology .
Sporangiophores of Phvcomvces blakesleeanus areparticularly well suited for the centrifugal separa-tion of cell constituents inside the living cell . Theyare large coenocytic cells of cylindrical shape,tapered at the end, and enclosed by a relativelyrigid cell wall, which, when properly mounted, canfunction as a minute centrifuge tube . In this paper,we present experiments which show that cell or-ganelles can be separated by centrifugation withinliving sporangiophores and that they remain func-tional . We will also characterize the organellesmorphologically and biochemically and measuretheir total amounts and their densities . SincePhvcomvces is widely used in the research on photo-tropism,' our particular concern was to identifyparticles which may contain the photoreceptorpigments .
MATERIALS AND METHODSWild type of Phycomyces blakesleeanus was used . Thefungus was grown on potato dextrose agar medium(Zankel et al ., 1967) at 25 éC and sporangiophores(spphs) were harvested when 1-2 cm long . Cultureswere illuminated from above to assure straight growthof the spphs . Stage I spphs (Castle, 1942) were pluckedfrom the mycelium and centrifuged in an invertedposition. Since the cell wall is not strong enough towithstand high centrifugal forces, the spph had to besupported within the centrifuge tube by a liquid thedensity of which was similar to that of the spph. At thecentrifugal end, a denser liquid functioned as a cushionto prevent the spph from being squashed against thebottom of the tube (Fig . 1 A) . High centrifugal forceswere obtained with a Spinco Model A centrifuge andswinging bucket rotor SW 39 . Sporangiophores weremounted in capillary tubes, 1 mm X 4 cm, containing40% Ficoll (Pharmacia Inc ., Uppsala, Sweden) atthe bottom (first 10 mm) and filled with 10c%o Ficoll .The capillary tube was placed in a 1.5-mm-wideaxial well of a plexiglas insert for regular Spinco cen-trifuge tubes. Heavy fluorochemical 3M CompanyKel-F3 (Minnesota Mining and Manufacturing Co .,Minneapolis) was introduced into the well for sup-porting the capillary and preventing breakage athigh speeds .
The cell contents were stratified best when thecentrifugation speed was increased slowly and keptat 25,000 rpm for 10 min and then raised to 35,000
0U
v-
10%
spph
40%
C
b
a
rA
BFIGURE 1 A, Spph mounted in a capillary for cen-trifugation (X 1 .2) . B, Centrifuged spph (X 6) (a),with injected oil droplet (b), mounted in a small capil-lary, and sealed with wax (c) .
rpm for 20 min . Smaller centrifuge forces were notsufficient to separate all the particles, and a longercentrifugation time or higher forces did not yieldsignificant improvement. The centrifuge tempera-ture was 5 éC. At the end of the centrifugation, thecentrifuge was braked to a stop ; the capillary tubeswere removed from the containers and inspectedunder a dissecting microscope . For better observation,the capillaries were broken at an appropriate leveland the specimens were removed and mounted inwater on microscope slides. Coverslips, supported withspacers, prevented crushing the specimen . Centri-fuged layers were sufficiently stable to remain in placeduring a short microscopic observation even when thespph was in a horizontal position .
Cytological and cytochemical studies of spphs arerendered difficult by the low permeability of the cellwall. Added chemicals or fixatives penetrate slowlyand unevenly into the cell, causing displacements andcontraction of the cytoplasm . Centrifuged layers canbe disorganized easily by fixation . The growing tip ofthe spph is more permeable, and chemicals may reachreaction sites in the cytoplasm only in its vicinity, thusfalsely indicating localized reactions . For cytological
' K. Bergman, P. V . Burke, E . Cerda-Olmedo, C . N. staining and some cytochemical reactions, spphs wereDavid, M . Delbriick, D . S. Dennison, K . Foster, fixed with formol-acetic acid-alcohol, or acetic acid-E. W. Goodell, M . Heisenberg, G . Meissner, W. alcohol alone . For electron microscopy, glutaralde-Shropshire, and M . Zalokar, 1969 . Bact. Rev . Data to hyde (5 jo in 0.1 mt phosphate buffer at pH 6 .8) gavebe published.
good results . As good or better results could be ob-
MARKO ZALOKAR Intracellular Centrifugal Separation
49 5
on August 22, 2006
ww
w.jcb.org
Dow
nloaded from

tained with 5% glutaraldehyde and 5% acrolein in0.1 M cacodylate buffer at pH 7 .0 (F . A . Burr, per-sonal communication) . The fixed specimens wereposttreated with 170 Os04, dehydrated in alcohol,transferred to propylene oxide, and embedded inVestopal . The transfer from propylene oxide toVestopal was made by dissolving Vestopal (with ini-tiator and activator added) in propylene oxide andevaporating the solvent slowly in a dry atmosphere.Blocks were properly oriented, and thin sections weremade with an LKB microtome. The sections werestained with uranyl acetate and lead citrate and werestudied with a Phillips (EM 200) electron microscope,operated at 60 kv.
Various substances can be injected into spphs aftercentrifugation . The normal spph loses its turgor andcollapses as soon as an opening is made in its wall,unless special precautions are taken . For protectionduring manipulation, the spph is introduced, tip first,into a capillary filled with 20% Ficoll . This capillaryis closed at one end and has an inside diameter ofabout 200 µ (Fig . 1 B) . All operations are performedunder water. The protruding part of the spph is cutwith a sharp razor. A micro-injection needle, about10 µ in diameter, is introduced through the openinginto the vacuole, and an appropriate amount of mate-rial is injected. The spph is then pushed farther intothe capillary, so that the protruding end is about 200 µlong. The capillary is then taken out of the water,placed in a vertical position, and the open end issealed with a droplet of molten beeswax. The waxseals the spph and keeps it attached to the rim of thecapillary . This capillary is then introduced into a1-mm X 4-cm tube filled with paraffin oil and is cen-trifuged. Spphs prepared in this way can withstandcentrifugal forces up to 25,000 rpm . For most work,the spph was first centrifuged at 35,000 rpm for 20min, then injected according to this procedure, andcentrifuged again at 25,000 rpm for 20 min .
RESULTS
Description of the Layers
When a stage I spph is centrifuged for 10 min at25,000 rpm and for 20 min at 35,000 rpm, its con-tents sediment in several well defined layers (Fig .2) . Typical results for a I-cm spph grown underoptimal conditions are presented in Fig . 3 . Startingfrom the centrifugal end, the following layers wereobserved :
1 . Polyphosphates (Fig . 4) . The heaviest mate-rial settles to the bottom in the form of large drop-lets, confluent at the tip into a colorless mass, notrevealing any structure . In the electron micro-scope, one sees that the droplets are enclosed by asimple membrane, taking up stain avidly . At least
496 THE JOURNAL OF CELL BIOLOGY • VOLUME 41, 1969
FIGURE 2 Photograph of a centrifuged spph . Numberscorresponding to the ones in the text identify each layer .
two types of droplets are present : one is morehomogeneous, with stained material aggregatingin the center ; the other is coarsely reticulated . Theinterstitial material is composed of barely percepti-
on August 22, 2006
ww
w.jcb.org
Dow
nloaded from

A
ble fibrils . Cytochemical tests indicate that thislayer contains polyphosphates. The layer stainsheterochromatically with toluidine blue, turnsblack after reaction with lead acetate and hydro-gen sulfide, and gives a faintly positive molybdatereaction for phosphates. It could not be determinedwhich of the three types of sedimented materialscorrespond to polyphosphates . The droplets giveonly very weak reactions for carbohydrates byperiodic acid-Schiff reaction and for proteins(basophilic cytological stains) .
2 . Polyphosphates and crystals (Fig. 5) . Thefirst layer changes gradually into the dark secondlayer. The droplets described above becomesmaller and are intermixed with rhombohedralprotein crystals . In electron micrographs, these
B total
31 nl
FiouxE 3 Drawing of a centrifuged spph with layers and their volume given at the right and specificgravity determined by oil drops at the left. The arrows indicate the position that oil drops took aftercentrifugation .
crystals display a regular lattice pattern (Fig . 6) .The crystals were first described by Thornton andThimann (1964) .
3 . Glycogen (Figs . 7 and 8) . This layer appearstransparent and structureless in a microscope, andhas a bluish cast in reflected light . It stains reddish-brown with iodine and gives a positive periodicacid-Schiff reaction, both characteristic for fungalglycogen. Electron micrographs show tightlypacked granules about 800 A in diameter (Fig . 7) .It is very difficult to make thin sections of theglycogen layer, since glycogen swells and preventsthe sections from spreading properly . In most sec-tions, the glycogen must have been dissolved, leav-ing the corresponding empty spaces unstained withlead citrate (Fig . 8) .
MARKO ZALOKAR Intracellular Centrifugal Separation
497
on August 22, 2006
ww
w.jcb.org
Dow
nloaded from
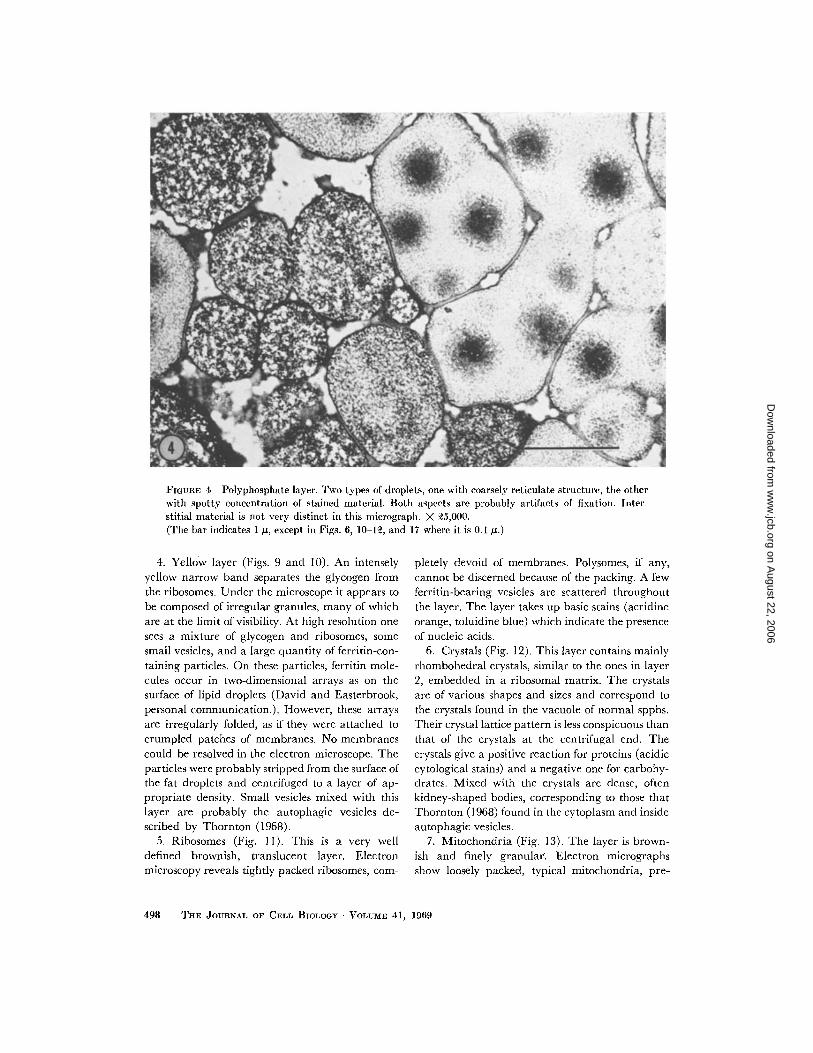
FIGURE 4 Polyphosphate layer. Two types of droplets, one with coarsely reticulate structure, the otherwith spotty concentration of stained material . Both aspects are probably artifacts of fixation . Inter-stitial material is not very distinct in this micrograph . X 25,000.(The bar indicates 1 µ, except in Figs. 6, 10-12, and 17 where it is 0 .1 ,u .)
4. Yellow layer (Figs . 9 and 10) . An intenselyyellow narrow band separates the glycogen fromthe ribosomes . Under the microscope it appears tobe composed of irregular granules, many of whichare at the limit of visibility. At high resolution onesees a mixture of glycogen and ribosomes, somesmall vesicles, and a large quantity of ferritin-con-taining particles . On these particles, ferritin mole-cules occur in two-dimensional arrays as on thesurface of lipid droplets (David and Easterbrook,personal communication.). However, these arraysare irregularly folded, as if they were attached tocrumpled patches of membranes . No membranescould be resolved in the electron microscope . Theparticles were probably stripped from the surface ofthe fat droplets and centrifuged to a layer of ap-propriate density. Small vesicles mixed with thislayer are probably the autophagic vesicles de-scribed by Thornton (1968) .
5 . Ribosomes (Fig. 11). This is a very welldefined brownish, translucent layer. Electronmicroscopy reveals tightly packed ribosomes, com-
498
THE JOURNAL OF CELL BIOLOGY • VOLUME 41, 1969
pletely devoid of membranes. Polysomes, if any,cannot be discerned because of the packing . A fewferritin-bearing vesicles are scattered throughoutthe layer . The layer takes up basic stains (acridineorange, toluidine blue) which indicate the presenceof nucleic acids .
6 . Crystals (Fig. 12), This layer contains mainlyrhombohedral crystals, similar to the ones in layer2, embedded in a ribosomal matrix . The crystalsare of various shapes and sizes and correspond tothe crystals found in the vacuole of normal spphs .Their crystal lattice pattern is less conspicuous thanthat of the crystals at the centrifugal end . Thecrystals give a positive reaction for proteins (acidiccytological stains) and a negative one for carbohy-drates. Mixed with the crystals are dense, oftenkidney-shaped bodies, corresponding to those thatThornton (1968) found in the cytoplasm and insideautophagic vesicles .
7 . Mitochondria (Fig . 13) . The layer is brown-ish and finely granular. Electron micrographsshow loosely packed, typical mitochondria, pre-
on August 22, 2006
ww
w.jcb.org
Dow
nloaded from
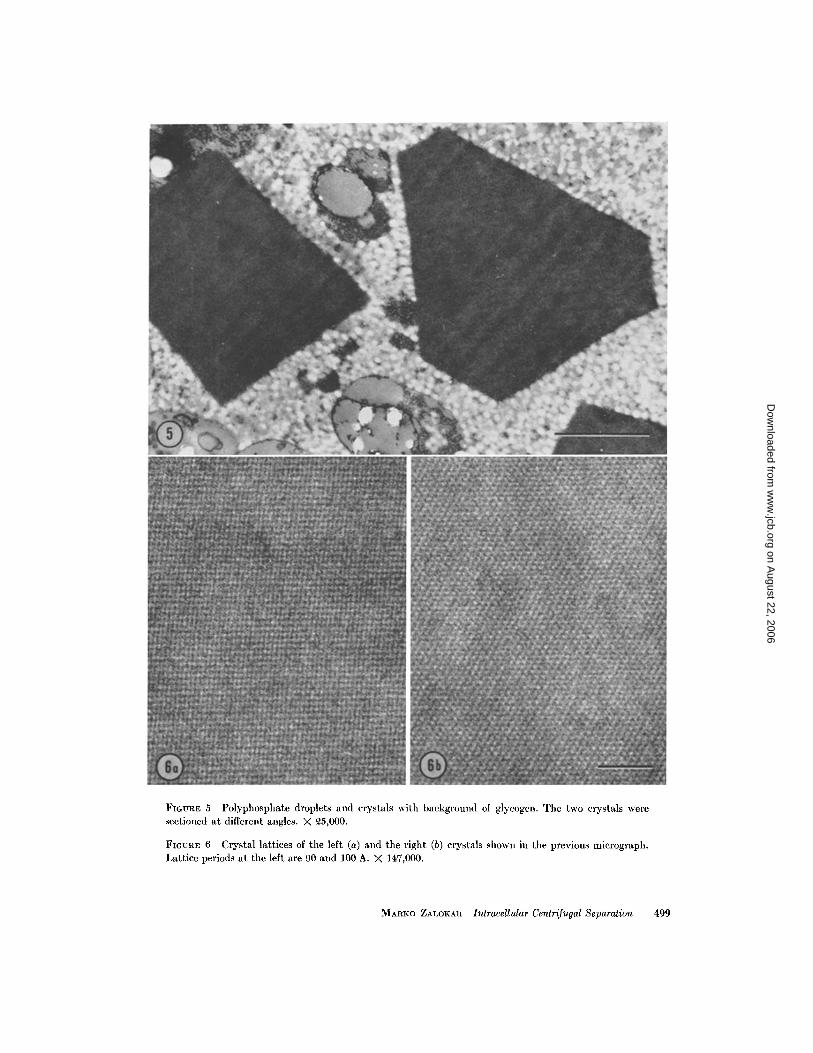
FIGURE 5 Polyphosphate droplets and crystals with background of glycogen . The two crystals weresectioned at different angles . X 25,000.
FIGURE 6 Crystal lattices of the left (a) and the right (b) crystals shown in the previous micrograph .Lattice periods at the left are 90 and 100 A . X 147,000 .
MARKO ZALOKAR Intracellular Centrifugal Separation
499
on August 22, 2006
ww
w.jcb.org
Dow
nloaded from

dominantly circular or oval, with well developedcristae . The spaces between the mitochondria arefilled with granules which, owing to their size andappearance, can be identified as ribosomes . Someferritin-bearing particles are scattered between themitochondria, but there are no other types of par-ticles and no nuclei . The mitochondrial layer givesa positive Nadi reaction and it reduces tetrazoliumsalts, indicating active cytochrome oxidase andsuccinic dehydrogenase systems .
8. Mitochondria and fibrils . The upper portionof the mitochondrial layer is less homogeneous . Itcontains some opaque vesicles and an indication offibrils parallel to the longitudinal axis . Distinctfibrils are not seen in the electron microscope, butthey may have been poorly fixed . There are, how-ever, elongated vesicles, probably correspondingto the cytoplasmic strands which support cyto-plasmic streaming . Some of the irregular vesicles inthis layer resemble the autophagic vesicles in themicrographs published by Thornton (1968) . Theupper end of the layer is a zone of transition to thenext layer .
9. Nuclei (Fig . 14) . This layer contains uni-formly packed nuclei, approximately 2 µ in diam-
500
FIGURE 7 Glycogen granules stained with lead citrate . X 25,000 .
FIGURE 8 Glycogen layer showing empty emplacements of glycogen granules . X 25,000 .
TnE JOURNAL OF CELL BIOLOGY • VOLUME 41, 1969
eter. Electron micrographs show loosely packednuclei with their nucleoli randomly oriented . Onewould expect the denser nucleolus to be at thecentrifugal end of the nucleus, but it is possiblethat the nuclear layer was strongly disturbed bypenetrating fixatives. Interstices between the nucleiare occupied by small vesicles of endoplasmicreticulum. Very few, if any, mitochondria werefound in this layer . The layer gives a strong positiveFeulgen reaction and a negative test for succinicdehydrogenase .
10. Endoplasmic reticulum (Fig . 15) . On top ofthe nuclear layer, a layer of finely granular mate-rial often separates . It exhibits brownian motion .Electron micrographs show vesicles of the samekind as those which fill the interstices between thenuclei, and which are considered to be endo-plasmic reticulum . The layer was not observedunder high enough power to see the fine structureof the membranes enclosing the vesicles .
11 . Membranes and vesicles (Fig . 16). Thislayer is composed of various vesicles and clumps ofcytoplasm, often contsining material stained withcarotenoids, and is difficult to characterize . Elec-tron microscopy reveals many irregular large
on August 22, 2006
ww
w.jcb.org
Dow
nloaded from
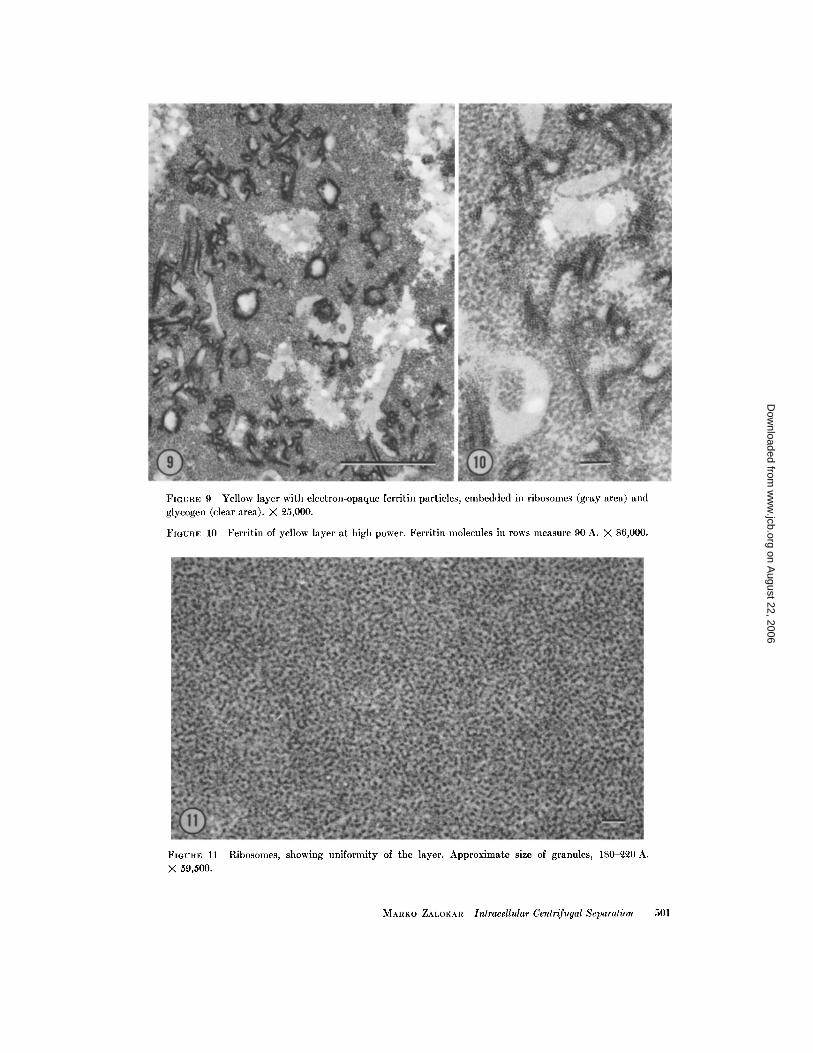
FIGURE 9 Yellow layer with electron-opaque ferritin particles, embedded in ribosomes (gray area) andglycogen (clear area) . X 25,000 .
FIGURE 10 Ferritin of yellow layer at high power . Ferritin molecules in rows measure 90 A. X 86,000 .
FIGURE 11 Ribosomes, showing uniformity of the layer . Approximate size of granules, 180-220 A .X 59,500 .
MARKO ZALOKAR Intracellular Centrifugal Separation
501
on August 22, 2006
ww
w.jcb.org
Dow
nloaded from
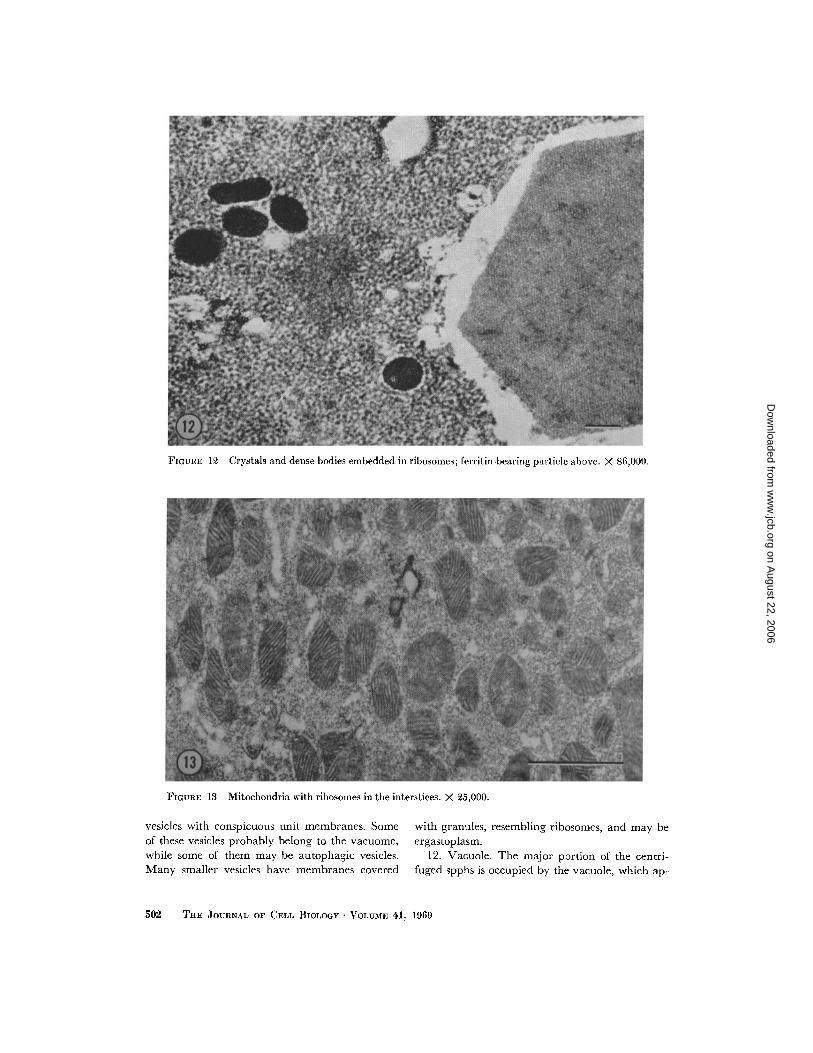
FIGURE 12 Crystals and dense bodies embedded in ribosomes ; ferritin-bearing particle above . X 86,000 .
FIGURE 13 Mitochondria with ribosomes in the interstices . X 25,000 .
vesicles with conspicuous unit membranes . Some with granules, resembling ribosomes, and may beof these vesicles probably belong to the vacuome, ergastoplasm .while some of them may be autophagic vesicles .
12. Vacuole . The major portion of the centri-Many smaller vesicles have membranes covered fuged spphs is occupied by the vacuole, which ap .-
502
THE JOURNAL OF CELL BIOLOGY - VOLUME 41, 1969
on August 22, 2006
ww
w.jcb.org
Dow
nloaded from
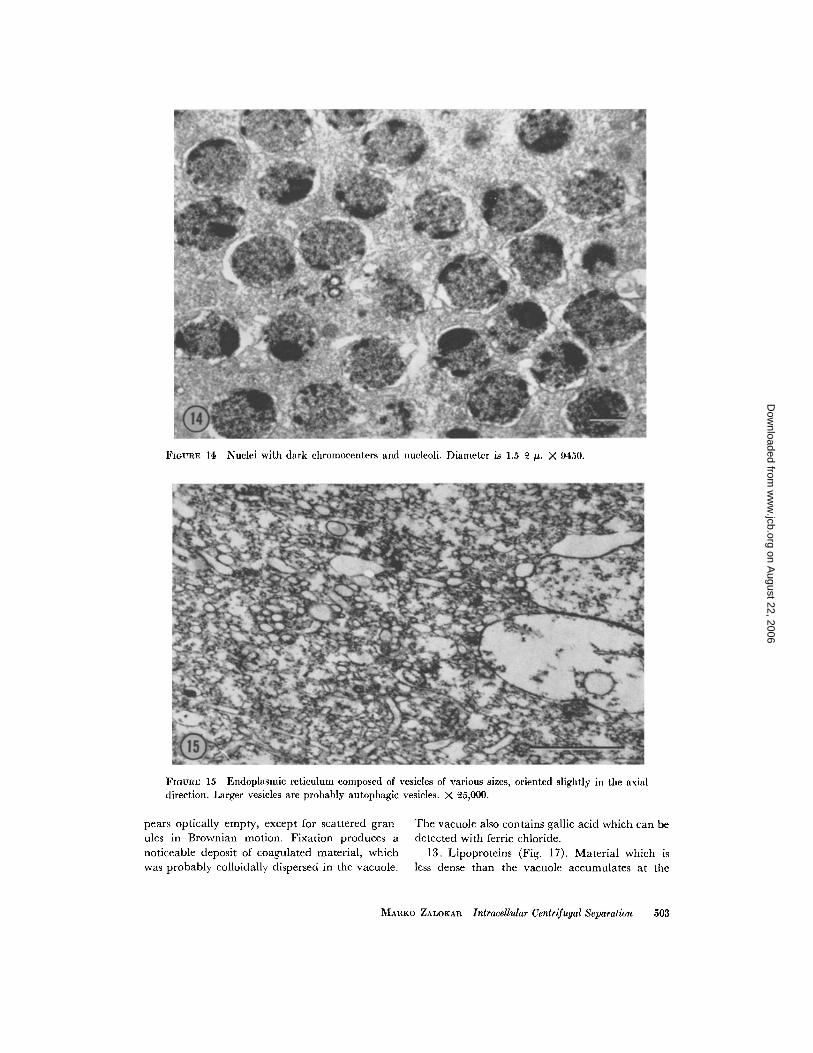
FIGURE 14 Nuclei with dark chromocenters and nucleoli . Diameter is 1 .5-2 µ . X 9450 .
FIGURE 15 Endoplasmic reticulum composed of vesicles of various sizes, oriented slightly in the axialdirection . Larger vesicles are probably autophagic vesicles . X 25,000 .
pears optically empty, except for scattered gran
The vacuole also contains gallic acid which can beules in Brownian motion . Fixation produces a detected with ferric chloride .noticeable deposit of coagulated material, which
13 . Lipoproteins (Fig . 17) . Material which iswas probably colloidally dispersed in the vacuole . less dense than the vacuole accumulates at the
MARKO ZALOKAR Intracellular Centrifugal Separation
503
on August 22, 2006
ww
w.jcb.org
Dow
nloaded from
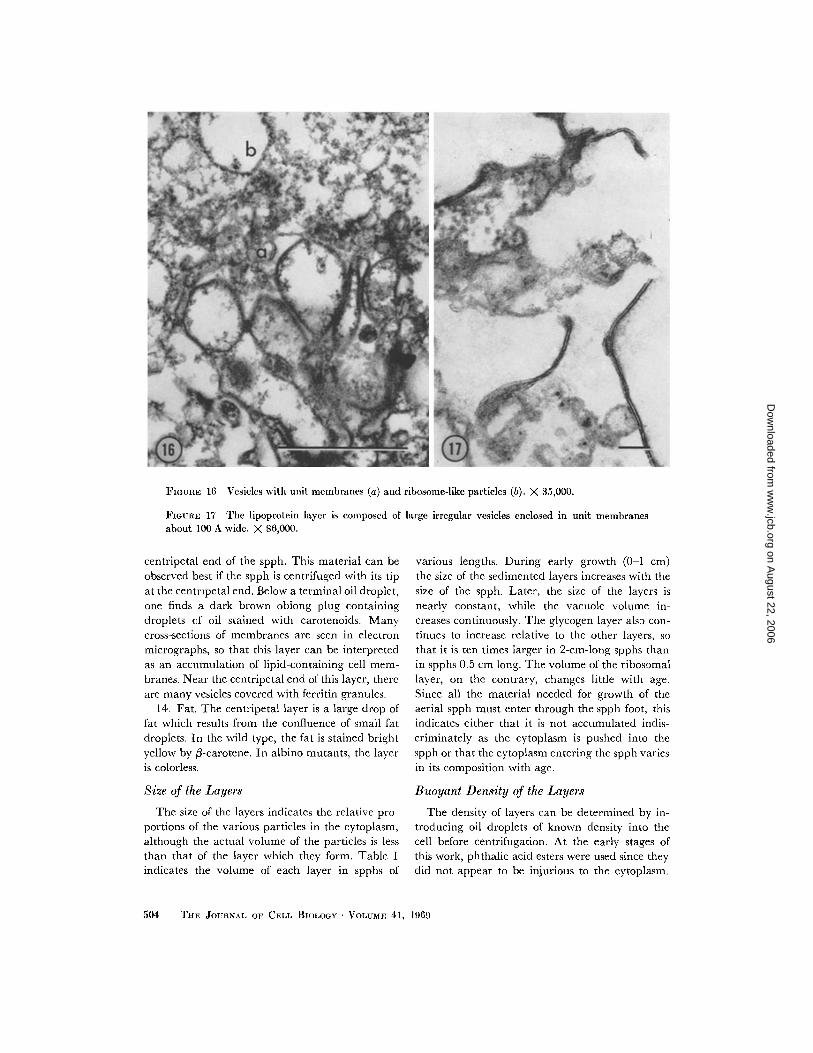
FIGURE 16 Vesicles with unit membranes (a) and ribosome-like particles (b) . X 35,000 .
FIGURE 17 The lipoprotein layer is composed of large irregular vesicles enclosed in unit membranesabout 100 A wide. X 86,000.
centripetal end of the spph . This material can beobserved best if the spph is centrifuged with its tipat the centripetal end . Below a terminal oil droplet,one finds a dark brown oblong plug containingdroplets of oil stained with carotenoids . Manycross-sections of membranes are seen in electronmicrographs, so that this layer can be interpretedas an accumulation of lipid-containing cell mem-branes . Near the centripetal end of this layer, thereare many vesicles covered with ferritin granules .
14. Fat . The centripetal layer is a large drop offat which results from the confluence of small fatdroplets. In the wild type, the fat is stained brightyellow by ß-carotene . In albino mutants, the layeris colorless .
Size of the Layers
The size of the layers indicates the relative pro-portions of the various particles in the cytoplasm,although the actual volume of the particles is lessthan that of the layer which they form . Table Iindicates the volume of each layer in spphs of
5 04
THE JOURNAL OF CELL BIOLOGY . VOLUME 41, 1969
various lengths. During early growth (0-1 cm)the size of the sedimented layers increases with thesize of the spph . Later, the size of the layers isnearly constant, while the vacuole volume in-creases continuously . The glycogen layer also con-tinues to increase relative to the other layers, sothat it is ten times larger in 2-cm-long spphs thanin spphs 0 .5 cm long . The volume of the ribosomallayer, on the contrary, changes little with age .Since all the material needed for growth of theaerial spph must enter through the spph foot, thisindicates either that it is not accumulated indis-criminately as the cytoplasm is pushed into thespph or that the cytoplasm entering the spph variesin its composition with age .
Buoyant Density of the Layers
The density of layers can be determined by in-troducing oil droplets of known density into thecell before centrifugation . At the early stages ofthis work, phthalic acid esters were used since theydid not appear to be injurious to the cytoplasm .
on August 22, 2006
ww
w.jcb.org
Dow
nloaded from

TABLE I
Volume of Different Layers
(Relatively small volumes of crystals andyellow band were divided between neighbor-ing layers .)
Size of spph
Various densities were obtained by mixing diethylphthalate and dibutyl phthalate . Later, mixturesof silicone oil and fluorochemicals were used . DowCorning (Midland, Mich .) silicone oil 706 (den-sity 1 .10 g/cc) is miscible with 3M Company'sfluorochemical Kel-F3 (density 1 .96 at 5 °C) andalso with common paraffin oil (density 0.9) . Thusit is possible to make mixtures of inert oils withdensities between 0 .9 and 1 .96 . Also, these oils canserve to establish density gradients in this range .
The oils were injected into spphs which previ-ously had been centrifuged at 35,000 rpm for 20min. After a second centrifugation at 25,000 rpm,the position of the oil droplets relative to the cyto-plasmic layers was observed (Fig . 3, left ; Fig . 18) .The oil droplets indicate the average density ofthe layers, i .e . particles and suspension fluid, andnot the density of the particles themselves .
Particle density can be determined if particlesof similar size and known density are introduced .Bacteria can be injected into the cell without anyapparent interaction with the cytoplasm . Densities(p) of several bacteria were determined by cen-trifugation in step density gradients made of Ficoll(Pharmacia Inc .) or Urographin (E . R. Squibband Sons, New York) (Tamir and Gilvarg, 1966) .The osmotic action of the latter is great enough toincrease the densities of bacteria by dehydration .
FIGURE 18 Oil droplet (density = 1 .06) inside cen-trifuged spph settles above mitochondria . Numbersidentify each layer.
MARKO ZALOKAR Intracellular Centrifugal Separation
505
Layer 5 mm 10 mm 15 mm
Lipoprotein membrane 5 .0 9 .0 15 .0Vacuole 52 .0 160 .0 400 .0MembranesEndoplasmic reticulum} 6 .5 8 .0 10 .0Nuclei 2 .0 6 .4 8 .0Fibrils, mitochondria)Mitochondria
f 2 .1 9 .0 13 .0CrystalsRibosomes 1 .5 2 .8 3 .4Yellow bandGlycogen 0.8 3 .0 9 .6Polyphosphates, crystals
0 .4 1 .0 0 .8Polyphosphates
fTotal below vacuole 13 .3 30 .2 44 .8
on August 22, 2006
ww
w.jcb.org
Dow
nloaded from
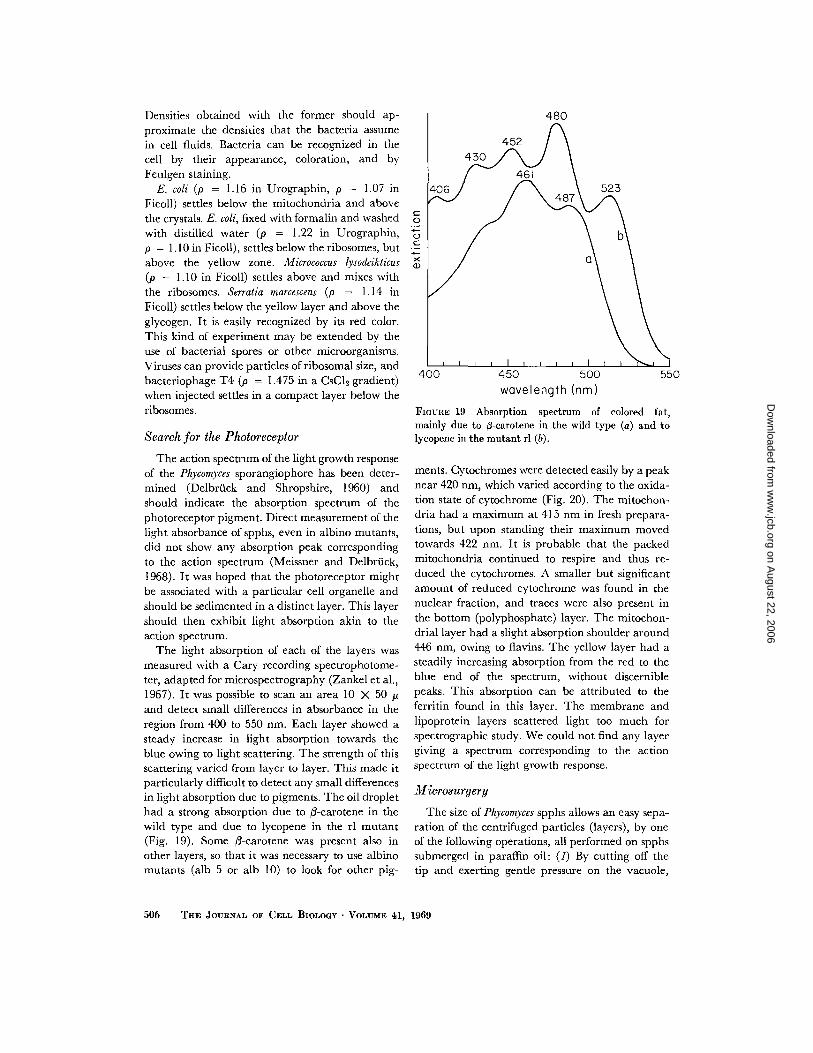
Densities obtained with the former should ap-proximate the densities that the bacteria assumein cell fluids . Bacteria can be recognized in thecell by their appearance, coloration, and byFeulgen staining .E. coli (p = 1 .16 in Urographin, p = 1 .07 in
Ficoll) settles below the mitochondria and abovethe crystals . E . coli, fixed with formalin and washedwith distilled water (p = 1 .22 in Urographin,p = 1 .10 in Ficoll), settles below the ribosomes, butabove the yellow zone . Micrococcus lysodeikticus(p = 1 .10 in Ficoll) settles above and mixes withthe ribosomes . Serratia marcescens (p = 1 .14 inFicoll) settles below the yellow layer and above theglycogen. It is easily recognized by its red color .This kind of experiment may be extended by theuse of bacterial spores or other microorganisms .Viruses can provide particles of ribosomal size, andbacteriophage T4 (p = 1 .475 in a CsCl2 gradient)when injected settles in a compact layer below theribosomes .
Search for the Photoreceptor
The action spectrum of the light growth responseof the Phycomyces sporangiophore has been deter-mined (Delbrück and Shropshire, 1960) andshould indicate the absorption spectrum of thephotoreceptor pigment . Direct measurement of thelight absorbance of spphs, even in albino mutants,did not show any absorption peak correspondingto the action spectrum (Meissner and Delbrück,1968) . It was hoped that the photoreceptor mightbe associated with a particular cell organelle andshould be sedimented in a distinct layer . This layershould then exhibit light absorption akin to theaction spectrum .
The light absorption of each of the layers wasmeasured with a Cary recording spectrophotome-ter, adapted for microspectrography (Zankel et al .,1967) . It was possible to scan an area 10 X 50 pand detect small differences in absorbance in theregion from 400 to 550 nm. Each layer showed asteady increase in light absorption towards theblue owing to light scattering . The strength of thisscattering varied from layer to layer . This made itparticularly difficult to detect any small differencesin light absorption due to pigments . The oil droplethad a strong absorption due to 3-carotene in thewild type and due to lycopene in the rl mutant(Fig. 19) . Some /3-carotene was present also inother layers, so that it was necessary to use albinomutants (alb 5 or alb 10) to look for other pig-
5 06 THE JOURNAL OF CELL BIOLOGY, VOLUME 41, 1969
400
450
500wavelength (nm)
FIGURE 19 Absorption spectrum of colored fat,mainly due to 3-carotene in the wild type (a) and tolycopene in the mutant rl (b) .
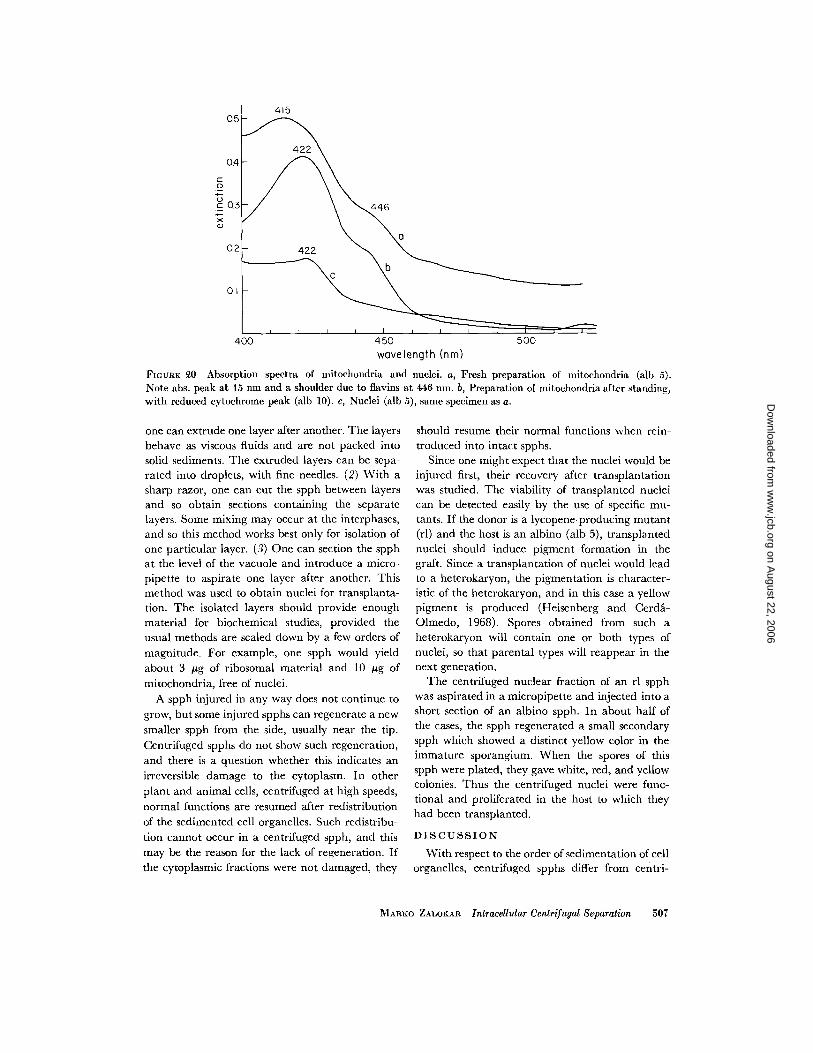
ments. Cytochromes were detected easily by a peaknear 420 nm, which varied according to the oxida-tion state of cytochrome (Fig . 20) . The mitochon-dria had a maximum at 415 nm in fresh prepara-tions, but upon standing their maximum movedtowards 422 nm . It is probable that the packedmitochondria continued to respire and thus re-duced the cytochromes . A smaller but significantamount of reduced cytochrome was found in thenuclear fraction, and traces were also present inthe bottom (polyphosphate) layer. The mitochon-drial layer had a slight absorption shoulder around446 nm, owing to flavins. The yellow layer had asteadily increasing absorption from the red to theblue end of the spectrum, without discerniblepeaks. This absorption can be attributed to theferritin found in this layer . The membrane andlipoprotein layers scattered light too much forspectrographic study . We could not find any layergiving a spectrum corresponding to the actionspectrum of the light growth response .
Microsurgery
The size of Phycomyces spphs allows an easy sepa-ration of the centrifuged particles (layers), by oneof the following operations, all performed on spphssubmerged in paraffin oil : (1) By cutting off thetip and exerting gentle pressure on the vacuole,
550
on August 22, 2006
ww
w.jcb.org
Dow
nloaded from
400 450
500wavelength (nm)
FIGURE 20 Absorption spectra of mitochondria and nuclei. a, Fresh preparation of mitochondria (alb 5) .Note abs . peak at 15 Ion and a shoulder due to flavins at 446 run . b, Preparation of mitochondria after standing,with reduced cytochrome peak (alb 10) . e, Nuclei (alb 5), same specimen as a .
one can extrude one layer after another. The layersbehave as viscous fluids and are not packed intosolid sediments . The extruded layers can be sepa-rated into droplets, with fine needles . (2) With asharp razor, one can cut the spph between layersand so obtain sections containing the separatelayers . Some mixing may occur at the interphases,and so this method works best only for isolation ofone particular layer . (3) One can section the spphat the level of the vacuole and introduce a micro-pipette to aspirate one layer after another . Thismethod was used to obtain nuclei for transplanta-tion . The isolated layers should provide enoughmaterial for biochemical studies, provided theusual methods are scaled down by a few orders ofmagnitude . For example, one spph would yieldabout 3 µg of ribosomal material and 10 µg ofmitochondria, free of nuclei,
A spph injured in any way does not continue togrow, but some injured spphs can regenerate a newsmaller spph from the side, usually near the tip .Centrifuged spphs do not show such regeneration,and there is a question whether this indicates anirreversible damage to the cytoplasm . In otherplant and animal cells, centrifuged at high speeds,normal functions are resumed after redistributionof the sedimented cell organelles. Such redistribu-tion cannot occur in a centrifuged spph, and thismay be the reason for the lack of regeneration . Ifthe cytoplasmic fractions were not damaged, they
should resume their normal functions when rein-troduced into intact spphs .
Since one might expect that the nuclei would beinjured first, their recovery after transplantationwas studied . The viability of transplanted nucleican be detected easily by the use of specific mu-tants . If the donor is a lycopene-producing mutant(rl) and the host is an albino (alb 5), transplantednuclei should induce pigment formation in thegraft. Since a transplantation of nuclei would leadto a heterokaryon, the pigmentation is character-istic of the heterokaryon, and in this case a yellowpigment is produced (Heisenberg and Cerdâ-Olmedo, 1968). Spores obtained from such aheterokaryon will contain one or both types ofnuclei, so that parental types will reappear in thenext generation.
The centrifuged nuclear fraction of an rl spphwas aspirated in a micropipette and injected into ashort section of an albino spph . In about half ofthe cases, the spph regenerated a small secondaryspph which showed a distinct yellow color in theimmature sporangium . When the spores of thisspph were plated, they gave white, red, and yellowcolonies. Thus the centrifuged nuclei were func-tional and proliferated in the host to which theyhad been transplanted .
DISCUSSION
With respect to the order of sedimentation of cellorganelles, centrifuged spphs differ from centri-
MARKO ZALOKAR Intracellular Centrifugal Separation
507
on August 22, 2006
ww
w.jcb.org
Dow
nloaded from

fuged cell homogenates . In cell homogenates, thecell debris and the nuclei sediment at low speeds,the mitochondria at intermediate speeds, and theribosomes only after prolonged centrifugation at100,000 g. This order of sedimentation is deter-mined by the velocity of sedimentation, and thisvelocity, for spherical particles, is proportional toà'-Op (a = radius of the particle, Ap = p p - po =
density difference of particles and fluid) . Thus, theradius of the particle is the dominant feature,larger particles coming out at low speed, smallerparticles at successively higher speeds . In contrast,the layering inside spphs is essentially the result ofequilibrium centrifugation, the order being deter-mined by the density of the layers formed by theparticles .
The density of the layer differs from that of theparticle depending on the volume-packing frac-tion (the fraction of the total volume occupied bythe particles) . For spheres of uniform size, this frac-tion is 0 .739, independent of the size of the sphere,and the layer density is
Plsrer = Po + (pp - p0) 0.739 .
For nonspherical uniform particles, the packingfraction can be anything from unity (for cubes) tonear zero (very spiny particles or inflexible tangledthreads) . Spheres of equal size and different den-sity will form, under equilibrium conditions, sepa-rate layers .
For nonuniform particles, it is useful to considerthe case of a mixture of large and small spheres, ofequal density pa > po . The small spheres fill inter-stices between the larger spheres and thus form amixed layer which has a higher packing fractionthan either of the pure layers . Thus, centrifugationwould produce at the bottom a mixed layer andabove it a pure layer of either larger or smallerparticles, depending upon which particle was inexcess . A more complex situation obtains when theparticles are of different densities, but then also,mixed layers can be formed .
An immiscible fluid will form a layer by itself,whose position is determined solely by its densityrelative to that of the particle layer. Densities ofthe various layers, as found by the position of oildroplets, are given in Fig . 3 . The ribosome layerhas a density 1 .10, and M. lysodeikticus (p = 1 .10)mixes with the ribosomes, as might be expected .The density of the ribosomes must be less than1 .475-since phage T4 settles below the ribosomes
508
THE JOURNAL OF CELL BIOLOGY • VOLUME 41, 1969
-but higher than the density of the layer theyform. The density of the glycogen particles mustbe more than that of the glycogen layer (1 .25) . In acesium chloride gradient, a density of 1 .66 has beenmeasured, and the relatively low density of theglycogen layer may be due to a loose structure ofglycogen particles . The density of the mitochondriamust be close to 1 .07, since this is the averagedensity of the layer in which they are found andsince E . coli (p = 1 .07), which is similar in size tothe mitochondria, sinks below them . The densityof the nuclei is less than 1 .07, since they float ontop of the mitochondrial layer .
These estimates of particle densities are signifi-cantly different from the densities of particlesobtained by centrifugation of cell homogenates ina sucrose density gradient . Thus, mitochondriaobtained from Phycomyces spphs have a density of1 .13-1 .14 in a sucrose gradient, and mitochondriafrom the mycelium a density of 1 .17-1 .19 (E . W .Goodell, personal communication) . Mitochondriaof Neurospora have a density of 1 .16-1 .20 in asucrose gradient (Luck, 1963). The differencesmust be at least partially due to the osmotic effectof the sucrose solution, as discussed by de Duveet al . (1959). These authors corrected the densityof rat liver mitochondria, measured by Thomsonand Mikata (1954) as 1 .22, to 1 .103 for isosmotic0.25 M sucrose . The density of isolated nuclei isgenerally considered to be high, but this may beowing to improper interpretation of the highsedimentation constant. In centrifuged cells, thenuclei sometimes move towards the heavy end,but often prove to be lighter than the mito-chondria, as in Phycomyces and Neurospora (Zalokar,1959) .
The separation of cell organelles in a living statemay open new ways for studying their propertiesby properly refined techniques of molecularbiology. It may be valuable to obtain particleswhich are not contaminated with the debris ofother cellular components, as so often happens incell homogenates . Thus, the ribosome layer ispractically pure, the mitochondria are free ofnuclei, and nuclei are free of mitochondria . How-ever, the separations are not perfect since themitochondria are mixed with the ribosomes andthe nuclei are mixed with the endoplasmic retic-ulum. Various layers may be recombined for astudy of their metabolic interdependence . Theorganelles may be tested for genetic and otherfunctions by their injection into appropriate hosts .
on August 22, 2006
ww
w.jcb.org
Dow
nloaded from

This work was supported by the Gosney Fund and bya National Science Foundation grant GB-4642 . Theauthor wishes to thank Dr. M. Delbrück for helpfulsuggestions and criticism, Mrs . P. V . Burke for help inthe preparation of the manuscript, and Dr . G . Meiss-
REFERENCES
BEAMS, H . W., and R. L. KING. 1939 . Bot . Rev. 5 :132 .CASTLE, E. S . 1942 . Amer. J . Bot . 29:664.DEDuvE, C., J . BERTHET, and H. BEAUFAY. 1959 .
Progr . Biophys . Chem . 91 :325 .DELBRUCK, M., and W . SHROPSHIRE, JR . 1960. Plant
Physiol . 35 :194 .HEISENBERG, M., and E . CERDA-OLMEDO. 1968 . Mol.
Gen . Genet . 102 :187 .LUCK, J . L. 1963 . Proc. Nat . Acad. Sci. U.S.A . 49 :233 .MEISSNER, G., and M . DELBRÜCK. 1968. Plant Physiol.
43 :1279 .
ner for the measurements of light absorption . G .Adam did some early work on centrifugation .
Received for publication 15 October1968, and in revised form30 December 1968 .
TAMIR, H ., and C . GILVARG. 1966 . J. Biol. Chem . 241 :1085 .
THOMSON, J . F., and E . T. MIKATA . 1954 . Arch. Bio-chem . Biophys . 51 :487,
THORNTON, R. M. 1968 . J. Ultrastruct . Res . 21 :269.THORNTON, R. M., and K . V. THIMANN . 1964, J. Cell
Biol . 20 :345 .ZALOKAR, M . 1959 . Exp . Cell Res . 19 :559 .ZANKEL, K. L., P . V, BURKE, and M. DELBRUCK .
1967 . J . Gen . Physiol . 50 :1893,
MARKO ZALOKAR Intracellular Centrifugal Separation
509
on August 22, 2006
ww
w.jcb.org
Dow
nloaded from
![Review Mitochondrial Stat3, the Need for Design Thinking · 2016. 2. 28. · into the mitochondria [45] (Fig. 2). Mitochondria were the one of the first intracellular organelles to](https://static.fdocuments.in/doc/165x107/600275e837e9e64e9746e911/review-mitochondrial-stat3-the-need-for-design-2016-2-28-into-the-mitochondria.jpg)

















