
Intestinal Sodium Glucose Cotransporter 1 Inhibition...
11
1521-0103/354/3/279–289$25.00 http://dx.doi.org/10.1124/jpet.115.225508 THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS J Pharmacol Exp Ther 354:279–289, September 2015 Copyright ª 2015 by The American Society for Pharmacology and Experimental Therapeutics Intestinal Sodium Glucose Cotransporter 1 Inhibition Enhances Glucagon-Like Peptide-1 Secretion in Normal and Diabetic Rodents Takahiro Oguma, Keiko Nakayama, Chiaki Kuriyama, Yasuaki Matsushita, Kumiko Yoshida, Kumiko Hikida, Naoyuki Obokata, Minoru Tsuda-Tsukimoto, Akira Saito, Kenji Arakawa, Kiichiro Ueta, and Masaharu Shiotani Research Division, Mitsubishi Tanabe Pharma Corporation, Toda-shi, Saitama, Japan Received April 27, 2015; accepted June 22, 2015 ABSTRACT The sodium glucose cotransporter (SGLT) 1 plays a major role in glucose absorption and incretin hormone release in the gastrointestinal tract; however, the impact of SGLT1 inhibition on plasma glucagon-like peptide-1 (GLP-1) levels in vivo is controversial. We analyzed the effects of SGLT1 inhibitors on GLP-1 secretion in normoglycemic and hyperglycemic rodents using phloridzin, CGMI [3-(4-cyclopropylphenylmethyl)-1-(b-D- glucopyranosyl)-4-methylindole], and canagliflozin. These com- pounds are SGLT2 inhibitors with moderate SGLT1 inhibitory activity, and their IC 50 values against rat SGLT1 and mouse SGLT1 were 609 and 760 nM for phloridzin, 39.4 and 41.5 nM for CGMI, and 555 and 613 nM for canagliflozin, respectively. Oral administration of these inhibitors markedly enhanced and prolonged the glucose-induced plasma active GLP-1 (aGLP-1) increase in combination treatment with sitagliptin, a dipeptidyl peptidase-4 (DPP4) inhibitor, in normoglycemic mice and rats. CGMI, the most potent SGLT1 inhibitor among them, enhanced glucose-induced, but not fat-induced, plasma aGLP-1 increase at a lower dose compared with canagliflozin. Both CGMI and canagli- flozin delayed intestinal glucose absorption after oral administration in normoglycemic rats. The combined treatment of canagliflozin and a DPP4 inhibitor increased plasma aGLP-1 levels and im- proved glucose tolerance compared with single treatment in both 8- and 13-week-old Zucker diabetic fatty rats. These results suggest that transient inhibition of intestinal SGLT1 promotes GLP-1 secretion by delaying glucose absorption and that concomitant inhibition of intestinal SGLT1 and DPP4 is a novel therapeutic option for glycemic control in type 2 diabetes mellitus. Introduction Long-term hyperglycemia leads to toxic effects in various tissues and organs (Rossetti et al., 1990; Leahy et al., 1992; Nathan, 1993), and therapeutic strategies to treat patients with type 2 diabetes mellitus (T2DM) are currently focused on controlling blood glucose levels (Blonde, 2010; Bennett et al., 2012; Inzucchi et al., 2012; Qaseem et al., 2012). Many classes of oral antihyperglycemic drugs (OADs) have been developed, but it is still difficult to maintain long-term glycemic control in most patients with T2DM because of the limited efficacy and safety issues of drugs (Kahn et al., 2006; Blonde, 2010; Qaseem et al., 2012). Recently, multiple functions of incretins, such as glucagon-like peptide-1 (GLP-1), have been investigated. GLP-1 is secreted from enteroendocrine L cells in the distal part of the small intestine in response to dietary stimulation (Orskov et al., 1993) and exerts antidiabetic effects, such as enhancing glucose-dependent insulin secretion and inhibiting gastric emptying, food intake, and glucagon secretion (Drucker, 2003). Dipeptidyl peptidase-4 (DPP4) inhibitors, a novel class of OADs, delay enzymatic degradation and inactivation of GLP-1, thereby increasing insulin release and decreasing glucagon secretion in a glucose-dependent manner (Drucker, 2003; Fukuda-Tsuru et al., 2012). Sodium glucose cotransporter (SGLT) 2 is predominantly located on the apical side of renal proximal tubules and plays a critical role in glucose reabsorption from glomerular filtrates (Kanai et al., 1994a; Hediger et al., 1995; You et al., 1995; Abdul-Ghani et al., 2013). SGLT2 inhibitors, the newest class of OADs, lower plasma glucose levels independent of insulin action by inhibiting renal glucose reabsorption (Oku et al., 1999, 2000; Arakawa et al., 2001; Ueta et al., 2005). On the other hand, SGLT1 is highly expressed on the brush- border membrane of villus enterocytes in the proximal part of the small intestine and is responsible for dietary glucose absorption (Takata et al., 1992; Freeman et al., 1993; Kanai et al., 1994b, 1995). Glucose stimulates GLP-1 secretion from All authors are employees of Mitsubishi Tanabe Pharma Corporation and have not received financial support from any other institution. T.O. and K.N. contributed equally to this work. dx.doi.org/10.1124/jpet.115.225508. ABBREVIATIONS: aGLP-1, active glucagon-like peptide-1; AMG, a-methyl-D-glucopyranoside; AUC, area under the curve; CGMI, 3-(4- cyclopropylphenylmethyl)-1-(b-D-glucopyranosyl)-4-methylindole; DPP4, dipeptidyl peptidase-4; ELISA, enzyme-linked immunosorbent assay; GI, gastrointestinal; GLP-1, glucagon-like peptide-1; GLUT, glucose transporter; OAD, oral antihyperglycemic drug; OGTT, oral glucose tolerance test; SD, Sprague-Dawley; SGLT, sodium glucose cotransporter; T2DM, type 2 diabetes mellitus; ZDF, Zucker diabetic fatty. 279 at ASPET Journals on April 14, 2020 jpet.aspetjournals.org Downloaded from
Transcript of Intestinal Sodium Glucose Cotransporter 1 Inhibition...

1521-0103/354/3/279–289$25.00 http://dx.doi.org/10.1124/jpet.115.225508THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS J Pharmacol Exp Ther 354:279–289, September 2015Copyright ª 2015 by The American Society for Pharmacology and Experimental Therapeutics
Intestinal Sodium Glucose Cotransporter 1 InhibitionEnhances Glucagon-Like Peptide-1 Secretion inNormal and Diabetic Rodents
Takahiro Oguma, Keiko Nakayama, Chiaki Kuriyama, Yasuaki Matsushita, Kumiko Yoshida,Kumiko Hikida, Naoyuki Obokata, Minoru Tsuda-Tsukimoto, Akira Saito, Kenji Arakawa,Kiichiro Ueta, and Masaharu ShiotaniResearch Division, Mitsubishi Tanabe Pharma Corporation, Toda-shi, Saitama, Japan
Received April 27, 2015; accepted June 22, 2015
ABSTRACTThe sodium glucose cotransporter (SGLT) 1 plays a major rolein glucose absorption and incretin hormone release in thegastrointestinal tract; however, the impact of SGLT1 inhibitionon plasma glucagon-like peptide-1 (GLP-1) levels in vivo iscontroversial. We analyzed the effects of SGLT1 inhibitors onGLP-1 secretion in normoglycemic and hyperglycemic rodentsusing phloridzin, CGMI [3-(4-cyclopropylphenylmethyl)-1-(b-D-glucopyranosyl)-4-methylindole], and canagliflozin. These com-pounds are SGLT2 inhibitors with moderate SGLT1 inhibitoryactivity, and their IC50 values against rat SGLT1 and mouseSGLT1 were 609 and 760 nM for phloridzin, 39.4 and 41.5 nM forCGMI, and 555 and 613 nM for canagliflozin, respectively. Oraladministration of these inhibitors markedly enhanced andprolonged the glucose-induced plasma active GLP-1 (aGLP-1)
increase in combination treatment with sitagliptin, a dipeptidylpeptidase-4 (DPP4) inhibitor, in normoglycemic mice and rats.CGMI, the most potent SGLT1 inhibitor among them, enhancedglucose-induced, but not fat-induced, plasma aGLP-1 increase at alower dose compared with canagliflozin. Both CGMI and canagli-flozin delayed intestinal glucose absorption after oral administrationin normoglycemic rats. The combined treatment of canagliflozinand a DPP4 inhibitor increased plasma aGLP-1 levels and im-proved glucose tolerance compared with single treatment inboth 8- and 13-week-old Zucker diabetic fatty rats. These resultssuggest that transient inhibition of intestinal SGLT1 promotes GLP-1secretion by delaying glucose absorption and that concomitantinhibition of intestinal SGLT1 and DPP4 is a novel therapeuticoption for glycemic control in type 2 diabetes mellitus.
IntroductionLong-term hyperglycemia leads to toxic effects in various
tissues and organs (Rossetti et al., 1990; Leahy et al., 1992;Nathan, 1993), and therapeutic strategies to treat patientswith type 2 diabetes mellitus (T2DM) are currently focused oncontrolling blood glucose levels (Blonde, 2010; Bennett et al.,2012; Inzucchi et al., 2012; Qaseem et al., 2012). Many classesof oral antihyperglycemic drugs (OADs) have been developed,but it is still difficult to maintain long-term glycemic control inmost patients with T2DM because of the limited efficacy andsafety issues of drugs (Kahn et al., 2006; Blonde, 2010; Qaseemet al., 2012). Recently, multiple functions of incretins, such asglucagon-like peptide-1 (GLP-1), have been investigated. GLP-1is secreted from enteroendocrine L cells in the distal part ofthe small intestine in response to dietary stimulation (Orskov
et al., 1993) and exerts antidiabetic effects, such as enhancingglucose-dependent insulin secretion and inhibiting gastricemptying, food intake, and glucagon secretion (Drucker, 2003).Dipeptidyl peptidase-4 (DPP4) inhibitors, a novel class of OADs,delay enzymatic degradation and inactivation of GLP-1, therebyincreasing insulin release and decreasing glucagon secretionin a glucose-dependent manner (Drucker, 2003; Fukuda-Tsuruet al., 2012).Sodium glucose cotransporter (SGLT) 2 is predominantly
located on the apical side of renal proximal tubules and playsa critical role in glucose reabsorption from glomerularfiltrates (Kanai et al., 1994a; Hediger et al., 1995; You et al.,1995; Abdul-Ghani et al., 2013). SGLT2 inhibitors, the newestclass of OADs, lower plasma glucose levels independent ofinsulin action by inhibiting renal glucose reabsorption (Okuet al., 1999, 2000; Arakawa et al., 2001; Ueta et al., 2005). Onthe other hand, SGLT1 is highly expressed on the brush-border membrane of villus enterocytes in the proximal part ofthe small intestine and is responsible for dietary glucoseabsorption (Takata et al., 1992; Freeman et al., 1993; Kanaiet al., 1994b, 1995). Glucose stimulates GLP-1 secretion from
All authors are employees of Mitsubishi Tanabe Pharma Corporation andhave not received financial support from any other institution.
T.O. and K.N. contributed equally to this work.dx.doi.org/10.1124/jpet.115.225508.
ABBREVIATIONS: aGLP-1, active glucagon-like peptide-1; AMG, a-methyl-D-glucopyranoside; AUC, area under the curve; CGMI, 3-(4-cyclopropylphenylmethyl)-1-(b-D-glucopyranosyl)-4-methylindole; DPP4, dipeptidyl peptidase-4; ELISA, enzyme-linked immunosorbent assay; GI,gastrointestinal; GLP-1, glucagon-like peptide-1; GLUT, glucose transporter; OAD, oral antihyperglycemic drug; OGTT, oral glucose tolerance test;SD, Sprague-Dawley; SGLT, sodium glucose cotransporter; T2DM, type 2 diabetes mellitus; ZDF, Zucker diabetic fatty.
279
at ASPE
T Journals on A
pril 14, 2020jpet.aspetjournals.org
Dow
nloaded from

the gastrointestinal (GI) tract, and there is an increasing bodyof evidence that SGLT1 is a sensor linking glucose to incretinrelease (Moriya et al., 2009; Gorboulev et al., 2012). However,the observation that the SGLT1 inhibitor increased portalGLP-1 levels after intestinal glucose infusion contradicts thishypothesis (Shibazaki et al., 2012). The relative contribu-tions of SGLT1 in glucose-induced GLP-1 secretion remaincontroversial.Canagliflozin, a first-in-class SGLT2 inhibitor with potent
antihyperglycemic activity (Nomura et al., 2010; Liang et al.,2012; Cefalu et al., 2013; Kuriyama et al., 2014), has modestSGLT1 inhibitory potency compared with other highly selec-tive SGLT2 inhibitors (Grempler et al., 2012). A recent clinicalstudy of canagliflozin found delayed intestinal glucose ab-sorption accompanied by increased plasma GLP-1 levels inhealthy subjects (Polidori et al., 2013). Furthermore, the com-bined treatment with canagliflozin and teneligliptin, a DPP4inhibitor, increased plasma active GLP-1 (aGLP-1) levels indiabetic rats (Oguma et al., 2015). Because plasma GLP-1 levelswere not potentiated in SGLT2-deficient mice after a mealchallenge (Powell et al., 2013), it has been suggested that GLP-1elevation by canagliflozin is mediated by a mechanism otherthan the inhibition of SGLT2. However, the contribution ofSGLT1 inhibition to increased plasmaGLP-1 levels has not beendirectly determined in either animals or humans.In the present study, we hypothesized that pharmacological
inhibition of intestinal SGLT1 increases GLP-1 secretion and,when combined with DPP4 inhibition, the increase in plasmaaGLP-1 has a therapeutic potential for T2DM. To addressthis hypothesis, we used three SGLT2 inhibitors: phloridzin,CGMI [3-(4-cyclopropylphenylmethyl)-1-(b-D-glucopyranosyl)-4-methylindole], and canagliflozin. A previous report showedthat phloridzin reduced glucose-inducedGLP-1 release (Moriyaet al., 2009). CGMI is reported to have more potent inhibitoryactivity against SGLT1 than canagliflozin (Nomura et al.,2014). We explored the effects of these compounds on plasmaGLP-1 levels and intestinal carbohydrate absorption andevaluated the combined effect of canagliflozin and sitagliptin,a DPP4 inhibitor, in normal and diabetic rodents.
Materials and MethodsReagent and Chemicals
Canagliflozin, CGMI, sitagliptin, and voglibose were synthesized atthe Medicinal Chemistry Laboratory at Mitsubishi Tanabe PharmaCorporation (Toda-shi, Saitama, Japan). Phloridzin was purchasedfrom Sigma-Aldrich (St. Louis, MO). All other chemicals were pur-chased from commercial sources and were of reagent or tissue-culturegrade.
Cell-Based Assays
SGLTs Inhibition Assay. Expression plasmids containing hu-man SGLTs, rat SGLTs, and mouse SGLTs were stably transfectedinto Chinese hamster ovary–K1 cells. Cells were seeded into 24-wellplates at a density of 4 � 105 cells/well and cultured for 2 days. Thecells were washed with assay buffer (50 mM HEPES, 20 mM Trisbase, 5 mM KCl, 1 mM MgCl2, 1 mM CaCl2, and 137 mM NaCl at pH7.4) and were preincubated with the compound solutions for10 minutes. Transport reaction was initiated by adding 0.3 or 0.5 mMAMG (a-methyl-D-glucopyranoside) (Sigma-Aldrich) in the presenceof [14C]AMG (PerkinElmer, Waltham, MA) and was incubated for120 minutes at 37°C. Radioactive counts in cells were assessed using
a liquid scintillation counter (PerkinElmer). Protein concentrationwas determined using the Coomassie Plus protein assay kit (Pierce,Rockford, IL).
Facilitated Glucose Transporter Inhibition Assay. Glucosetransporter (GLUT) 1 activity was assessed in L6 myoblast cells(Health Science Research Resources Bank, Osaka, Japan) (Mitsumotoet al., 1991). Cells were seeded into 24-well plates at a density of3 � 105 cells/well and cultured for 24 hours. Prior to the transportexperiment, the cells were washed with Krebs-Ringer-phosphate-HEPES buffer (150 mM NaCl, 5 mM KCl, 1.25 mM MgSO4, 1.25 mMCaCl2, 10 mM HEPES, and 2.9 mM Na2HPO4 at pH 7.4) and werepreincubated with the compound solution for 5 minutes. Transportreaction was initiated by adding 2-deoxyglucose (Tokyo ChemicalIndustry, Tokyo, Japan) and [3H]2-deoxyglucose (American Radio-labeled Chemicals, St. Louis, MO) and was incubated for 30 minutesat room temperature. Incorporated radioactivity was determinedusing a liquid scintillation counter (PerkinElmer). Protein concentrationwas determined using the Coomassie Plus protein assay kit (Pierce).
DPP4 Inhibition Assay. Inhibitory activities of test compoundsagainst rat DPP4 were measured using serum collected from 7-week-old male Sprague-Dawley (SD) rats (Charles River Japan, Yokohama,Japan). Rat serum and Gly-Pro 4-methylcoumaryl-7-amide were mixedwith the assay buffer (phosphate-buffered saline containing 0.003%Brij-35 solution) to initiate the enzyme reaction, as described previously(Fukuda-Tsuru et al., 2012). The fluorescence intensity of Gly-Pro4-methylcoumaryl-7-amide was measured using a microplate reader(Molecular Devices, Sunnyvale, CA) after 1-hour incubation at 37°C.
In Vivo Studies
Animals and Test Compound Administration. All animalexperimental procedures were approved by the Institutional AnimalCare and Use Committee of Mitsubishi Tanabe Pharma Corporationand met the Japanese Experimental Animal Research Associationstandards, as defined in the Guidelines for Animal Experiments. MaleC57BL/6J mice were purchased from CLEA Japan (Kanagawa, Japan).Male SD rats and Zucker diabetic fatty (ZDF) rats were purchased fromCharles River Japan. During an acclimatization period of at least 1 week,the animals were housed in a temperature- and humidity-controlledenvironmentwith 12-hour light/dark cycles. Theywere providedwater adlibitum and a standard commercial diet. Test compounds for oral gavagewere prepared in 0.5% carboxymethylcellulose containing 0.2% Tween80. The vehicle, instead of the drug, was administered in the controlgroups.
Oral Glucose Tolerance Test in Mice. After overnight fasting,9-week-old C57BL/6J mice were simultaneously treated with testcompounds and glucose solution at 2 g/kg body weight. Blood wascollected from the abdominal aorta into chilled tubes containing EDTA(Dojindo Laboratories, Kumamoto, Japan) and a DPP4 inhibitor(Millipore, Billerica,MA) while under anesthesia with isoflurane (Escain;Mylan, Tokyo, Japan) before and at 5, 30, 60, 120, and 180 minutes afteradministration. After centrifugation, plasma was stored at 280°C untilthe measurement of plasma aGLP-1.
Oral Glucose Tolerance Test and Fat Tolerance Test inRats. After overnight fasting, test compounds and glucose solution(2 g/kg p.o.) were simultaneously administered to 7- to 8-week-old SDrats. Blood was collected from the tail vein before and at 10, 30, 60,120, and 180 minutes after administration. Oral glucose tolerancetests (OGTTs) in 8- or 13-week-old ZDF rats were performed using thesame protocol. In the fat tolerance test, 0.3 mg/kg of CGMI and wateror a fat emulsion, 2 g/kg Intralipos (Otsuka Pharmaceutical Factory,Tokushima, Japan), instead of glucose solution were orally adminis-tered. Blood was collected into chilled tubes containing EDTA anda DPP4 inhibitor, and after centrifugation, plasmawas stored at280°Cuntil the measurement of plasma glucose, insulin, and aGLP-1.
Sucrose Loading Test in Normal Rats. Overnight-fasted7-week-old SD rats were orally treated with CGMI and sucrose solu-tion (2.5 g/kg) simultaneously. Blood was collected from the tail vein at
280 Oguma et al.
at ASPE
T Journals on A
pril 14, 2020jpet.aspetjournals.org
Dow
nloaded from

0, 30, 60, 120, and 180 minutes after administration for plasma glucosemonitoring. In another set of experiments, the rats were anesthetizedwith pentobarbital sodium (Tokyo Chemical Industry) (50 mg/kg i.p.) at1 hour after administration, and the small intestine was removed anddivided into three parts: upper small intestine (lower segment 20 cmfrom the pylorus), middle small intestine (segment between upper andlower intestine), and lower small intestine (upper segment 20 cm fromthe ileocecal junction). The contents of these intestinal segments werecollected with 5ml of ice-cold saline, and the volume of the contents wasmeasured. Carbohydrates in collected samples were hydrolyzed withH2SO4 by boiling and neutralized with NaOH. Glucose contents werethen measured. For calculating sucrose dosage, the sucrose solutionwas hydrolyzed, and the glucose concentration was determined in thesame way.
For evaluating the effects of canagliflozin and voglibose, the ratswere anesthetized 1 or 6 hours after administration and GI tractswere removed and divided into five parts: upper small intestine,middle small intestine, lower small intestine, cecum, and largeintestine. Other procedures were performed in the same way as theexperiment with CGMI. The intraluminal contents of canagliflozin ina portion of the fluid collected from the upper small intestine weredetermined by liquid chromatography–tandem mass spectrometry,API 4000 (AB SCIEX, Framingham, MA) equipped with a CadenzaCD C-18 column, 2.0-mm i.d. � 50 mm, 3 mm (Imtakt Corporation,Kyoto, Japan), after deproteinization with acetonitrile followed bydilution with 0.1 mol/l ammonium acetate.
Determination of Metabolic Parameters. Glucose concentra-tions in plasma and GI contents were determined using a glucoseCII-test Wako kit (Wako Pure Chemical Industries, Osaka, Japan).Insulin concentrations were measured using an ultrasensitive ratinsulin enzyme-linked immunosorbent assay (ELISA) kit (MorinagaInstitute of Biologic Science, Yokohama, Japan). Plasma active GLP-1was determined by an ELISA kit (Millipore or Epitope Diagnostics,San Diego, CA) after solid phase extraction with an Oasis HLBmElution plate (Waters, Milford, MA) (Franz and Li, 2012; Kinoshitaand Kondo, 2015). Plasma total GLP-1 was determined by an ELISAkit (Meso Scale Discovery, Rockville, MD). The baseline values aredefined as plasma GLP-1 concentrations at 0 hours in the vehicle-treated group, and plasma GLP-1 relative levels were normalized andexpressed as the ratio to the baseline in the each figure. aGLP-1baseline values ranged from 0.10 to 1.17 pmol/l in all experiments,and the total GLP-1 baseline value was 0.62 pmol/l.
Statistical Analysis
Data in all figures were presented as the arithmetic means6 S.E.M.for each group, and IC50 values in Tables 1 and 2 were expressed as thegeometric mean of two to three experiments (Fleming et al., 1972;Stórustovu and Ebert, 2006; Kuriyama et al., 2014). The area underthe curve (AUC) and incremental AUC (DAUC, defined as the AUCoccurring above the baseline value) were calculated by the trapezoidalrule. Statistical differences between vehicle and single treatmentgroups or between single and combination treatment groups were
determined by one- or two-way analysis of variance followed byStudent’s t test (Hwee et al., 2015; Wang et al., 2015), unless otherwiseindicated in figure legends. Statistical analyses were performed usinga SAS-based system (SAS Institute, Cary, NC) or Prism software(GraphPad Software, San Diego, CA). Differences assessed by each testwere considered statistically significant at P , 0.05.
ResultsSGLT1 Inhibition and Selectivity. Tables 1 and 2
summarize the effect of the compounds used in this study onthe activities of SGLTs, GLUTs, and DPP4. Similar tophloridzin, CGMI and canagliflozin inhibited human, rat,and mouse SGLT1 (Table 1). These compounds also inhibitedSGLT2 activity, and CGMI was the most potent SGLT1inhibitor among the three compounds. These SGLT inhibitorsdid not inhibit GLUT1 and DPP4 activity, and sitagliptinpotently inhibited DPP4 activity (Table 2).Effects of Phloridzin and Canagliflozin in Combina-
tion with Sitagliptin on Plasma aGLP-1 Levels inNormal Mice and Rats. To determine whether oral treat-ment of phloridzin and canagliflozin affects GLP-1 secretion,we analyzed the time course of plasma GLP-1 levels duringOGTT. For this purpose, the in vivo effects of these inhibitorswere examined in combination with sitagliptin to preventaGLP-1 degradation. In normoglycemic mice and rats, plasmaaGLP-1 levels were not significantly changed after oral glucoseloading. In mice treated with 10 mg/kg sitagliptin, the plasmaaGLP-1 level was increased at 5 minutes after glucose load-ing (Fig. 1A). In contrast, in mice treated with 500 mg/kgphloridzin, the plasma aGLP-1 level was unchanged at5 minutes after glucose loading, but increased at 30 minutesand thereafter. Moreover, combined treatment of phloridzin andsitagliptin increased and prolonged plasma aGLP-1 elevationcompared with either monotherapy at 30 minutes and laterfollowing glucose loading. Similar results were obtained in rats(Fig. 1, B and C).The combined treatment of canagliflozin (0.3–30 mg/kg)
and sitagliptin (10 mg/kg) also increased plasma aGLP-1levels in a dose-dependent manner in normoglycemic miceand rats (Fig. 2). Monotherapy with canagliflozin increasedplasma total GLP-1 at 30 minutes after glucose loading innormoglycemic rats (Fig. 3A), and canagliflozin did not affectplasma DPP4 activity in combination with or withoutsitagliptin at 30 minutes after the oral administration ofcompounds (Fig. 3B). These data suggest that canagliflozindoes not prevent inactivation but enhances GLP-1 secretion.
TABLE 1Inhibitory effects of SGLT inhibitors on SGLT1 and SGLT2All values are expressed as the geometric mean of two to three experiments.
IC50
Human Rat Mouse
SGLT1 SGLT2 SGLT1 SGLT2 SGLT1 SGLT2
nmol/l
Phloridzin 241 23.8 609 49.1 760 65.6CGMI 22.1 1.39 39.4 2.73 41.5 6.31Canagliflozina 663 4.2 555 3.7 613 5.6
aData from Kuriyama et al. (2014).
TABLE 2Inhibitory effects of SGLT inhibitors and sitagliptin on rat GLUT andDPP4All values are expressed as the geometric mean of two to three experiments.
IC50
Rat GLUT Rat DPP4
nmol/l
Phloridzin .10,000 .10,000CGMI .10,000 .10,000Canagliflozin .10,000a .10,000Sitagliptin N.D. 12.5
N.D., not determined.aData from Kuriyama et al. (2014).
Intestinal SGLT1 Inhibition Enhances GLP-1 Secretion In Vivo 281
at ASPE
T Journals on A
pril 14, 2020jpet.aspetjournals.org
Dow
nloaded from

Effects of CGMI and Canagliflozin in Combinationwith Sitagliptin on Plasma aGLP-1 Levels in NormalRats. Subsequently, we analyzed the involvement of SGLT1in GLP-1 secretion in detail using CGMI and canagliflozin,which are potent and weak SGLT1 inhibitors, respectively. Insitagliptin-treated rats, glucose loading increased plasmaaGLP-1 concentrations in a manner dependent on the dose ofCGMI (0.1–1 mg/kg) (Fig. 4A). The efficacy of 0.3 mg/kg CGMIin increasing plasma aGLP-1 levels was almost equivalent to
that of 10 mg/kg canagliflozin (Fig. 4B). This dose ratio ofCGMI and canagliflozin was consistent with the inhibitoryactivities of these compounds against SGLT1.To further investigate whether intestinal SGLT1 inhibition
contributes to elevated plasma aGLP-1, we compared thestimulant for GLP-1 secretion, glucose as SGLT1 substrateand water or fat as a non-SGLT1 substrate. Oral waterloading did not increase plasma aGLP-1 levels in ratstreated with sitagliptin alone and with combination CGMI
Fig. 1. Effects of phloridzin combined with sitagliptin on plasma aGLP-1 levels during OGTT in C57BL/6J mice and SD rats. (A) Time course of plasmaaGLP-1 relative levels with sitagliptin (10 mg/kg) and phloridzin (500 mg/kg) treatment in mice. (B and C) Time course and AUC0–3 hours of plasmaaGLP-1 relative levels with sitagliptin (10 mg/kg) and phloridzin (300mg/kg) treatment in rats. Phloridzin, sitagliptin, and glucose solution (2 g/kg) weresimultaneously administered to 8-week-old C57BL/6J mice or 7-week-old SD rats at time 0 by oral gavage. Blood was collected from the abdominal veinin mice and the tail vein in rats. Data are presented as the mean 6 S.E.M. (N = 5–6). *P , 0.05 and **P , 0.01 for sitagliptin compared with vehicle;|P, 0.05 and ||P, 0.01 for phloridzin compared with vehicle; #P, 0.05 and ##P, 0.01 for sitagliptin + phloridzin compared with sitagliptin; $P, 0.05and $$P , 0.01 for sitagliptin + phloridzin compared with phloridzin group.
Fig. 2. Effects of canagliflozin combined with sitagliptin on plasma GLP-1 levels during OGTT in C57BL/6Jmice and SD rats. (A) Time course of plasmaaGLP-1 relative levels with sitagliptin (10 mg/kg) and canagliflozin (10 mg/kg) treatment in mice. (B and C) Time course and AUC0–2 hours of plasmaaGLP-1 relative levels with sitagliptin (10 mg/kg) and canagliflozin (0.3–30mg/kg) treatment in rats. Canagliflozin, sitagliptin, and glucose solution (2 g/kg)were simultaneously administered to mice and rats at time 0 by oral gavage. Blood was collected from the abdominal vein in mice or the tail vein in rats.Data are presented as the mean6 S.E.M. (N = 4–6). **P, 0.01 for sitagliptin compared with vehicle; ||P, 0.01 for canagliflozin compared with vehicle;##P , 0.01 for sitagliptin + canagliflozin compared with sitagliptin; $P , 0.05 and $$P , 0.01 for sitagliptin + canagliflozin compared with canagliflozingroup; &&P , 0.01 versus sitagliptin group.
282 Oguma et al.
at ASPE
T Journals on A
pril 14, 2020jpet.aspetjournals.org
Dow
nloaded from

and sitagliptin treatment (Fig. 5, A and B). Although oral fatloading also increased plasma aGLP-1 levels in sitagliptin-treated rats, the plasma aGLP-1 increase induced by fat
was not enhanced by combined treatment with CGMI andsitagliptin in contrast to that induced by glucose (Fig. 5, Cand D).
Fig. 3. Effects of canagliflozin on plasma total GLP-1 levels and DPP4 activity during OGTT in SD rats. (A) Plasma total GLP-1 relative levels at 0 and30 minutes after glucose administration in canagliflozin (0.3–30 mg/kg) treated rats. (B) Plasma DPP4 activity at 0 and 30 minutes after glucoseadministration in rats. Canagliflozin, sitagliptin, and glucose solution (2 g/kg) were simultaneously administered to SD rats at time 0 by oral gavage. Bloodwas collected from the tail vein in rats. Data are presented as the mean6 S.E.M. (N = 6). ++P , 0.01 versus vehicle group by one-way analysis of variancewith Dunnett’s post hoc test. In plasmaDPP4 activity, there were no significant differences between the canagliflozin (1–10mg/kg) versus vehicle group andbetween the sitagliptin + canagliflozin (1–10 mg/kg) versus sitagliptin group by one-way analysis of variance with Dunnett’s post hoc test, respectively.
Fig. 4. Effects of CGMI combined with sitagliptin on plasma aGLP-1 levels during OGTT in SD rats. (A and B) Time course and AUC0–3 hours of plasmaaGLP-1 relative levels. CGMI (0.1–1 mg/kg), canagliflozin (10 mg/kg), sitagliptin (10 mg/kg), and glucose solution (2 g/kg) were administered to SD ratsat time 0 by oral gavage, and blood was collected from the tail vein. Data are presented as the mean 6 S.E.M. (N = 6). *P , 0.05 versus vehicle group;##P , 0.01 versus sitagliptin group; and $P , 0.05 and $$P , 0.01 versus sitagliptin group by one-way analysis of variance with Dunnett’s post hoc test.
Intestinal SGLT1 Inhibition Enhances GLP-1 Secretion In Vivo 283
at ASPE
T Journals on A
pril 14, 2020jpet.aspetjournals.org
Dow
nloaded from

Fig. 5. Effects of CGMI combined with sitagliptin on plasma aGLP-1 levels during oral glucose or fat tolerance test in SD rats. (A and B) Time course andAUC0–3 hours of plasma aGLP-1 relative levels after glucose or water loading. (C and D) Time course and AUC0–3 hours of plasma aGLP-1 relative levelsafter glucose or fat loading. CGMI (0.3 mg/kg), sitagliptin (10 mg/kg), and glucose solution (2 g/kg) were administered to SD rats at time 0 by oral gavage,and blood was collected from the tail vein. Data are presented as the mean 6 S.E.M. (N = 6). **P , 0.01 versus vehicle-treated glucose-loading group;##P , 0.01 versus sitagliptin-treated glucose-loading group; $$P , 0.01 versus vehicle-treated fat-loading group.
284 Oguma et al.
at ASPE
T Journals on A
pril 14, 2020jpet.aspetjournals.org
Dow
nloaded from
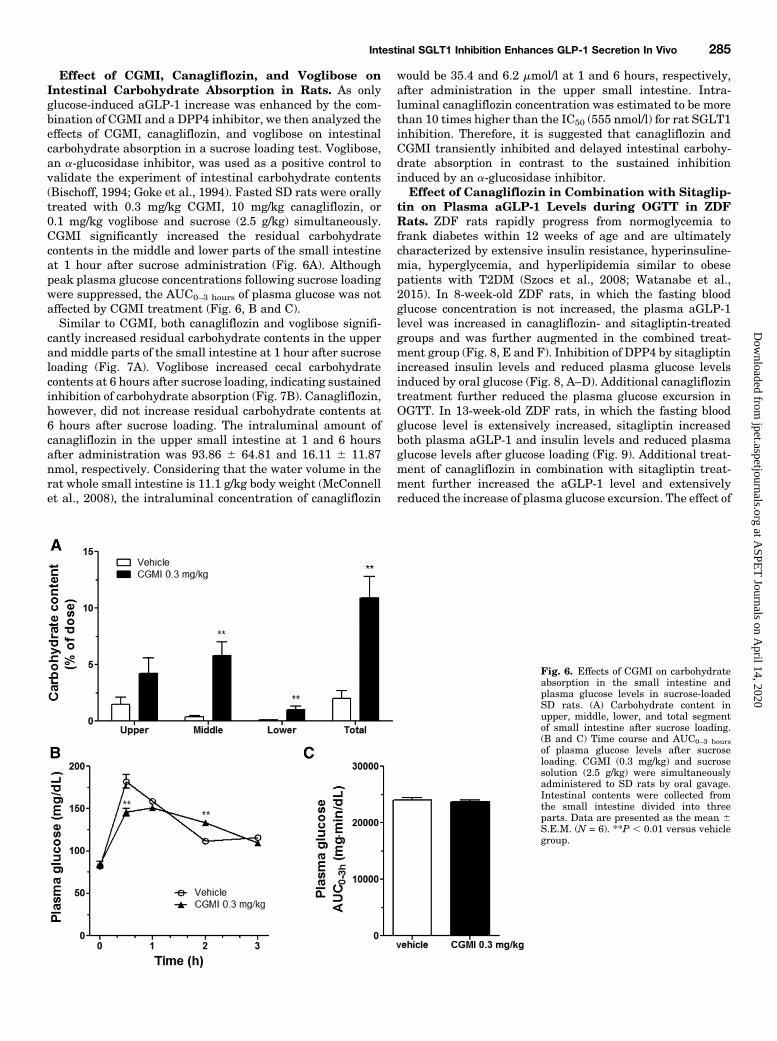
Effect of CGMI, Canagliflozin, and Voglibose onIntestinal Carbohydrate Absorption in Rats. As onlyglucose-induced aGLP-1 increase was enhanced by the com-bination of CGMI and a DPP4 inhibitor, we then analyzed theeffects of CGMI, canagliflozin, and voglibose on intestinalcarbohydrate absorption in a sucrose loading test. Voglibose,an a-glucosidase inhibitor, was used as a positive control tovalidate the experiment of intestinal carbohydrate contents(Bischoff, 1994; Goke et al., 1994). Fasted SD rats were orallytreated with 0.3 mg/kg CGMI, 10 mg/kg canagliflozin, or0.1 mg/kg voglibose and sucrose (2.5 g/kg) simultaneously.CGMI significantly increased the residual carbohydratecontents in the middle and lower parts of the small intestineat 1 hour after sucrose administration (Fig. 6A). Althoughpeak plasma glucose concentrations following sucrose loadingwere suppressed, the AUC0–3 hours of plasma glucose was notaffected by CGMI treatment (Fig. 6, B and C).Similar to CGMI, both canagliflozin and voglibose signifi-
cantly increased residual carbohydrate contents in the upperand middle parts of the small intestine at 1 hour after sucroseloading (Fig. 7A). Voglibose increased cecal carbohydratecontents at 6 hours after sucrose loading, indicating sustainedinhibition of carbohydrate absorption (Fig. 7B). Canagliflozin,however, did not increase residual carbohydrate contents at6 hours after sucrose loading. The intraluminal amount ofcanagliflozin in the upper small intestine at 1 and 6 hoursafter administration was 93.86 6 64.81 and 16.11 6 11.87nmol, respectively. Considering that the water volume in therat whole small intestine is 11.1 g/kg body weight (McConnellet al., 2008), the intraluminal concentration of canagliflozin
would be 35.4 and 6.2 mmol/l at 1 and 6 hours, respectively,after administration in the upper small intestine. Intra-luminal canagliflozin concentration was estimated to be morethan 10 times higher than the IC50 (555 nmol/l) for rat SGLT1inhibition. Therefore, it is suggested that canagliflozin andCGMI transiently inhibited and delayed intestinal carbohy-drate absorption in contrast to the sustained inhibitioninduced by an a-glucosidase inhibitor.Effect of Canagliflozin in Combination with Sitaglip-
tin on Plasma aGLP-1 Levels during OGTT in ZDFRats. ZDF rats rapidly progress from normoglycemia tofrank diabetes within 12 weeks of age and are ultimatelycharacterized by extensive insulin resistance, hyperinsuline-mia, hyperglycemia, and hyperlipidemia similar to obesepatients with T2DM (Szocs et al., 2008; Watanabe et al.,2015). In 8-week-old ZDF rats, in which the fasting bloodglucose concentration is not increased, the plasma aGLP-1level was increased in canagliflozin- and sitagliptin-treatedgroups and was further augmented in the combined treat-ment group (Fig. 8, E and F). Inhibition of DPP4 by sitagliptinincreased insulin levels and reduced plasma glucose levelsinduced by oral glucose (Fig. 8, A–D). Additional canagliflozintreatment further reduced the plasma glucose excursion inOGTT. In 13-week-old ZDF rats, in which the fasting bloodglucose level is extensively increased, sitagliptin increasedboth plasma aGLP-1 and insulin levels and reduced plasmaglucose levels after glucose loading (Fig. 9). Additional treat-ment of canagliflozin in combination with sitagliptin treat-ment further increased the aGLP-1 level and extensivelyreduced the increase of plasma glucose excursion. The effect of
Fig. 6. Effects of CGMI on carbohydrateabsorption in the small intestine andplasma glucose levels in sucrose-loadedSD rats. (A) Carbohydrate content inupper, middle, lower, and total segmentof small intestine after sucrose loading.(B and C) Time course and AUC0–3 hoursof plasma glucose levels after sucroseloading. CGMI (0.3 mg/kg) and sucrosesolution (2.5 g/kg) were simultaneouslyadministered to SD rats by oral gavage.Intestinal contents were collected fromthe small intestine divided into threeparts. Data are presented as the mean 6S.E.M. (N = 6). **P , 0.01 versus vehiclegroup.
Intestinal SGLT1 Inhibition Enhances GLP-1 Secretion In Vivo 285
at ASPE
T Journals on A
pril 14, 2020jpet.aspetjournals.org
Dow
nloaded from

combination treatment on plasma aGLP-1 and glucose levelswas greater than that of the canagliflozin single treatment.
DiscussionAlthough SGLT1 was the first molecule identified within
the SGLT family and its role of dietary glucose absorption hasbeen studied well (Kanai et al., 1994b, 1995), the role ofSGLT1 on incretin secretion in vivo is still controversial. Ithas been demonstrated that SGLT1 is crucial for incretinsecretion because phloridzin blocked the increase in portalaGLP-1 levels immediately after glucose administration intothe small intestine (Moriya et al., 2009). Furthermore, inSglt12/2 mice, plasma GLP-1 levels at 5 minutes after oralglucose challenge are reduced and glucose fails to stimulateincretin secretion from intestinal cultures in vitro (Gorboulevet al., 2012). In these previous studies, however, GLP-1 levelswere only measured in an early phase at 5 minutes afterglucose loading, but not in delayed phases at 30 minutes andlater. In the present study, phloridzin did not suppressplasma aGLP-1 levels at 5minutes, but rather enhanced themat 30 minutes and later after glucose loading in normoglycemicmice, especially in combination with sitagliptin. Similarly,
CGMI and canagliflozin, an SGLT2 inhibitor with a moderateinhibitory activity against SGLT1, enhanced GLP-1 secretionand increased plasma aGLP-1 levels in combination withsitagliptin in glucose-loaded SD rats without affecting basalplasma aGLP-1 levels. Because plasma GLP-1 levels were notpotentiated in SGLT2-deficient mice after a meal challenge(Powell et al., 2013), plasma aGLP-1 increase by phloridzin,CGMI, and canagliflozin would be attributed to SGLT1inhibition.CGMI, a potent SGLT1 inhibitor, increased glucose-
induced, but not fat-induced, plasma aGLP-1 increase ata lower dose compared with canagliflozin. Moreover, theresidual carbohydrate contents after oral sucrose ingestionwere increased in the middle and lower part of small intestineand the peak plasma glucose concentration was delayed inanimals orally treated with CGMI. Total absorption ofcarbohydrates was not inhibited, as the AUC of plasma glu-cose levels following oral sucrose challenge was not reducedby CGMI. Therefore, the results indicate that intestinalSGLT1 inhibition suppresses carbohydrate absorption in theupper segment of the GI tract and increases delivery to thelower segment. Because glucose is a representative stimulantof GLP-1 secretion, it is likely that nonabsorbed glucose
Fig. 7. Effects of canagliflozin on carbo-hydrate absorption in the GI tract insucrose-loaded SD rats. (A and B) Carbo-hydrate content in the upper small in-testine, middle small intestine, lowersmall intestine, cecum, large intestine,and total segment of intestine 1 hour(A) or 6 hours (B) after sucrose loading.Canagliflozin (10 mg/kg) or voglibose(0.1 mg/kg) and sucrose solution (2.5 g/kg)were simultaneously administered to SDrats by oral gavage. Intestinal contentswere collected from the GI tract dividedinto five parts. Data are presented as themean 6 S.E.M. (N = 4–6). *P , 0.05 and**P , 0.01 versus vehicle group.
286 Oguma et al.
at ASPE
T Journals on A
pril 14, 2020jpet.aspetjournals.org
Dow
nloaded from

delivered into the lower segment facilitates the release ofGLP-1 from L cells, which are abundantly located in the lowerpart of the small intestine.CGMI and canagliflozin transiently inhibited carbohydrate
absorption in the upper small intestine after oral adminis-tration. Although SGLT1 is abundantly present in theintestine and plays a critical role in intestinal glucoseabsorption (Gorboulev et al., 2012), SGLT2 is rarely expressedin the intestine (Chen et al., 2010). Therefore, it is suggestedthat the effects of these compounds are mediated mainly byinhibition of SGLT1. A previous study (Kuriyama et al., 2014)has shown that the plasma Cmax level of canagliflozinfollowing oral administration is below the IC50 value forSGLT1; therefore, it is unlikely that canagliflozin inhibitsintestinal SGLT1 as a systemic effect. Although it is difficultto determine the precise concentration of canagliflozin in theupper small intestine from the remaining intraluminalcontent, the estimated concentration markedly exceeded theIC50 value for SGLT1. The concentrations of CGMI andcanagliflozin were transiently high enough to inhibit SGLT1and delay the absorption of glucose in the upper smallintestine after oral administration. It is suggested thattransient inhibition of SGLT1 in the small intestine delayedthe absorption of glucose and the nonabsorbed glucosedelivered to the lower small intestine augmented GLP-1secretion in the L cells.
Sustained inhibition of carbohydrate absorption in thesmall intestine is accompanied by severe intestinal symp-toms, as shown in the phenotype of SGLT1 deficiency(Gorboulev et al., 2012) and adverse effects of a-glucosidaseinhibitors (Goke et al., 1994; Martin and Montgomery, 1996;Scott and Spencer, 2000). Systemic inhibition of SGLT1suppresses glucose absorption in the entire GI tract, andnonabsorbed glucose delivered into the cecum and colonfacilitates microbial fermentation and production of shortchain fatty acids, which in turn activate GLP-1 secretion fromL cells (Gorboulev et al., 2012; Powell et al., 2013). In contrastto the sustained inhibition of carbohydrate absorption in theentire digestive tract by agents, such as the a-glucosidaseinhibitor, transient inhibition of SGLT1 only delayed glucoseabsorption and enhanced GLP-1 secretion without in-creasing residual carbohydrate contents in the cecum andcolon. This suggests that transient and moderate SGLT1inhibition leads to GLP-1 release without adverse effects indigestive systems.Because SGLT1 is abundantly expressed in the duodenum
in patients with T2DM (Freitas et al., 2008; Gerich, 2010) andaGLP-1 increases nutrient-induced insulin release (Naucket al., 1997; Gromada et al., 1998), we determined thepotential of concomitant inhibition of SGLT1 and DPP4in hyperglycemic animals. In ZDF rats, an animal modelexhibiting progressive diabetes similar to human T2DM,
Fig. 8. Effects of canagliflozin combined with sitagliptin on glucose, insulin, and aGLP-1 levels during OGTT in 8-week-old ZDF rats. (A and B) Timecourse and DAUC0–3 hours of plasma glucose levels. (C and D) Time course and DAUC0–3 hours of plasma insulin levels. (E and F) Time course andDAUC0–3 hours of plasma aGLP-1 relative levels. Canagliflozin (10 mg/kg), sitagliptin (10 mg/kg), and glucose solution (2 g/kg) were administered to8-week-old ZDF rats at time 0 by oral gavage, and blood was collected from the tail vein. Data are presented as the mean6 S.E.M. (N = 6). *P, 0.05 and**P , 0.01 versus vehicle group; ||P , 0.01 versus sitagliptin group; $$P , 0.01 versus canagliflozin group.
Intestinal SGLT1 Inhibition Enhances GLP-1 Secretion In Vivo 287
at ASPE
T Journals on A
pril 14, 2020jpet.aspetjournals.org
Dow
nloaded from

insulin resistance develops at 7 weeks of age and plasmaglucose levels increase by 13 weeks of age (Szocs et al., 2008;Watanabe et al., 2015). Plasma aGLP-1 levels in ZDF ratswere higher after combination treatment with canagliflozinand sitagliptin compared with monotherapy of either drug atboth 8 and 13 weeks of age. In sitagliptin-treated ZDF rats atboth early and middle stages of diabetes, canagliflozintreatment enhanced the increase in plasma aGLP-1 levelsand further reduced plasma glucose excursion during OGTT.These antihyperglycemic effects are likely to be mediated bythe enhancement of GLP-1 secretion and the delay of glucoseabsorption because of intestinal SGLT1 inhibition in additionto the suppression of renal glucose reabsorption by SGLT2inhibition. Although glucose response in OGTT was markedlysuppressed, the plasma insulin response in the combinedtreatment group was not significantly different from that inthe sitagliptin-treated group. It is likely that increasedplasma aGLP-1 enhances glucose-induced insulin releasein OGTT in spite of suppressed glucose levels. These resultssuggest that concomitant inhibition of SGLT1 and DPP4increases plasma aGLP-1 levels and improves glucose toler-ance in hyperglycemia.In summary, the present study demonstrates that transient
and moderate inhibition of SGLT1 in the proximal part of theintestine increases the release of GLP-1 through increasing
glucose concentrations in the distal part of the small intestine.Concomitant inhibition of SGLT1 and DPP4 is a noveltherapeutic option for the improvement of glucose toleranceand hyperglycemia in T2DM.
Acknowledgments
Canagliflozin was developed by Mitsubishi Tanabe Pharma Corporationin collaboration with Janssen Research & Development, LLC.
Authorship Contributions
Participated in research design: Oguma, Nakayama, Kuriyama,Matsushita, Hikida, Obokata, Tsuda-Tsukimoto, Arakawa, Ueta,Shiotani.
Conducted experiments: Oguma, Nakayama, Kuriyama, Matsushita,Yoshida, Hikida, Obokata, Tsuda-Tsukimoto, Ueta, Shiotani.
Performed data analysis: Oguma, Nakayama, Hikida.Wrote or contributed to the writing of the manuscript: Oguma,
Saito, Ueta.
References
Abdul-Ghani MA, DeFronzo RA, and Norton L (2013) Novel hypothesis to explainwhy SGLT2 inhibitors inhibit only 30-50% of filtered glucose load in humans.Diabetes 62:3324–3328.
Arakawa K, Ishihara T, Oku A, Nawano M, Ueta K, Kitamura K, Matsumoto M,and Saito A (2001) Improved diabetic syndrome in C57BL/KsJ-db/db mice by oraladministration of the Na(1)-glucose cotransporter inhibitor T-1095. Br J Phar-macol 132:578–586.
Bennett WL, Odelola OA, Wilson LM, Bolen S, Selvaraj S, Robinson KA, Bass EB,and Puhan MA (2012) Evaluation of guideline recommendations on oral
Fig. 9. Effects of canagliflozin combined with sitagliptin on plasma glucose, insulin, and aGLP-1 levels during OGTT in 13-week-old ZDF rats. (A and B)Time course and DAUC0–3 hours of plasma glucose levels. (C and D) Time course and DAUC0–3 hours of plasma insulin levels. (E and F) Time course andDAUC0–3 hours of plasma aGLP-1 relative levels. Canagliflozin (10 mg/kg), sitagliptin (10 mg/kg), and glucose solutions (2 g/kg) were administered to13-week-old ZDF rats at time 0 by oral gavage, and blood was collected from the tail vein. Data are presented as the mean 6 S.E.M. (N = 6). *P , 0.05and **P , 0.01 versus vehicle group; |P , 0.05 and ||P , 0.01 versus sitagliptin group; $P , 0.05 and $$P , 0.01 versus canagliflozin group.
288 Oguma et al.
at ASPE
T Journals on A
pril 14, 2020jpet.aspetjournals.org
Dow
nloaded from

medications for type 2 diabetes mellitus: a systematic review. Ann Intern Med 156:27–36.
Bischoff H (1994) Pharmacology of alpha-glucosidase inhibition. Eur J Clin Invest 24(Suppl 3):3–10.
Blonde L (2010) Current antihyperglycemic treatment guidelines and algorithms forpatients with type 2 diabetes mellitus. Am J Med 123 (Suppl):S12–S18.
Cefalu WT, Leiter LA, Yoon KH, Arias P, Niskanen L, Xie J, Balis DA, CanovatchelW, and Meininger G (2013) Efficacy and safety of canagliflozin versus glimepiridein patients with type 2 diabetes inadequately controlled with metformin(CANTATA-SU): 52 week results from a randomised, double-blind, phase 3 non-inferiority trial. Lancet 382:941–950.
Chen J, Williams S, Ho S, Loraine H, Hagan D, Whaley JM, and Feder JN (2010)Quantitative PCR tissue expression profiling of the human SGLT2 gene and re-lated family members. Diabetes Ther 1:57–92.
Drucker DJ (2003) Enhancing incretin action for the treatment of type 2 diabetes.Diabetes Care 26:2929–2940.
Fleming WW, Westfall DP, De la Lande IS, and Jellett LB (1972) Log-normal dis-tribution of equiefective doses of norepinephrine and acetylcholine in several tis-sues. J Pharmacol Exp Ther 181:339–345.
Franz T and Li X (2012) The OASIS HLB mElution plate as a one-step platform formanual high-throughput in-gel digestion of proteins and peptide desalting. Pro-teomics 12:2487–2492.
Freeman TC, Wood IS, Sirinathsinghji DJ, Beechey RB, Dyer J, and Shirazi-BeecheySP (1993) The expression of the Na1/glucose cotransporter (SGLT1) gene in lambsmall intestine during postnatal development. Biochim Biophys Acta 1146:203–212.
Freitas HS, Anhê GF, Melo KF, Okamoto MM, Oliveira-Souza M, Bordin S,and Machado UF (2008) Na(1) -glucose transporter-2 messenger ribonucleic acidexpression in kidney of diabetic rats correlates with glycemic levels: involvement ofhepatocyte nuclear factor-1alpha expression and activity. Endocrinology 149:717–724.
Fukuda-Tsuru S, Anabuki J, Abe Y, Yoshida K, and Ishii S (2012) A novel, potent,and long-lasting dipeptidyl peptidase-4 inhibitor, teneligliptin, improves post-prandial hyperglycemia and dyslipidemia after single and repeated admin-istrations. Eur J Pharmacol 696:194–202.
Gerich JE (2010) Role of the kidney in normal glucose homeostasis and in thehyperglycaemia of diabetes mellitus: therapeutic implications. Diabet Med 27:136–142.
Göke B, Herrmann C, Göke R, Fehmann HC, Berghöfer P, Richter G, and Arnold R(1994) Intestinal effects of alpha-glucosidase inhibitors: absorption of nutrientsand enterohormonal changes. Eur J Clin Invest 24 (Suppl 3):25–30.
Gorboulev V, Schürmann A, Vallon V, Kipp H, Jaschke A, Klessen D, Friedrich A,Scherneck S, Rieg T, Cunard R, et al. (2012) Na(1)-D-glucose cotransporter SGLT1is pivotal for intestinal glucose absorption and glucose-dependent incretin secre-tion. Diabetes 61:187–196.
Grempler R, Thomas L, Eckhardt M, Himmelsbach F, Sauer A, Sharp DE, BakkerRA, Mark M, Klein T, and Eickelmann P (2012) Empagliflozin, a novel selectivesodium glucose cotransporter-2 (SGLT-2) inhibitor: characterisation and compar-ison with other SGLT-2 inhibitors. Diabetes Obes Metab 14:83–90.
Gromada J, Holst JJ, and Rorsman P (1998) Cellular regulation of islet hormonesecretion by the incretin hormone glucagon-like peptide 1. Pflugers Arch 435:583–594.
Hediger MA, Kanai Y, You G, and Nussberger S (1995) Mammalian ion-coupledsolute transporters. J Physiol 482:7S–17S.
Hwee DT, Kennedy AR, Hartman JJ, Ryans J, Durham N, Malik FI, and Jasper JR(2015) The small-molecule fast skeletal troponin activator, CK-2127107, improvesexercise tolerance in a rat model of heart failure. J Pharmacol Exp Ther 353:159–168.
Inzucchi SE, Bergenstal RM, Buse JB, Diamant M, Ferrannini E, Nauck M, PetersAL, Tsapas A, Wender R, and Matthews DR; American Diabetes Association(ADA); European Association for the Study of Diabetes (EASD) (2012) Manage-ment of hyperglycemia in type 2 diabetes: a patient-centered approach: positionstatement of the American Diabetes Association (ADA) and the European Associ-ation for the Study of Diabetes (EASD). Diabetes Care 35:1364–1379.
Kahn SE, Haffner SM, Heise MA, Herman WH, Holman RR, Jones NP, Kravitz BG,Lachin JM, O’Neill MC, Zinman B, et al.; ADOPT Study Group (2006) Glycemicdurability of rosiglitazone, metformin, or glyburide monotherapy. N Engl J Med355:2427–2443.
Kanai Y, Lee WS, You G, Brown D, and Hediger MA (1994a) The human kidney lowaffinity Na1/glucose cotransporter SGLT2. Delineation of the major renal reab-sorptive mechanism for D-glucose. J Clin Invest 93:397–404.
Kanai Y, Nussberger S, Romero MF, Boron WF, Hebert SC, and Hediger MA (1995)Electrogenic properties of the epithelial and neuronal high affinity glutamatetransporter. J Biol Chem 270:16561–16568.
Kanai Y, Stelzner M, Nussberger S, Khawaja S, Hebert SC, Smith CP, and HedigerMA (1994b) The neuronal and epithelial human high affinity glutamate trans-porter. Insights into structure and mechanism of transport. J Biol Chem 269:20599–20606.
Kinoshita S and Kondo K (2015) Evaluation of pharmacokinetic and pharmacody-namic interactions of canagliflozin and teneligliptin in Japanese healthy malevolunteers. Expert Opin Drug Metab Toxicol 11:7–14.
Kuriyama C, Xu JZ, Lee SP, Qi J, Kimata H, Kakimoto T, Nakayama K, Watanabe Y,Taniuchi N, Hikida K, et al. (2014) Analysis of the effect of canagliflozin on renalglucose reabsorption and progression of hyperglycemia in Zucker diabetic Fattyrats. J Pharmacol Exp Ther 351:423–431.
Leahy JL, Bonner-Weir S, and Weir GC (1992) Beta-cell dysfunction induced bychronic hyperglycemia. Current ideas on mechanism of impaired glucose-inducedinsulin secretion. Diabetes Care 15:442–455.
Liang Y, Arakawa K, Ueta K, Matsushita Y, Kuriyama C, Martin T, Du F, Liu Y, XuJ, Conway B, et al. (2012) Effect of canagliflozin on renal threshold for glucose,
glycemia, and body weight in normal and diabetic animal models. PLoS One 7:e30555.
Martin AE and Montgomery PA (1996) Acarbose: an alpha-glucosidase inhibitor. AmJ Health Syst Pharm 53:2277–2290.
McConnell EL, Basit AW, and Murdan S (2008) Measurements of rat and mousegastrointestinal pH, fluid and lymphoid tissue, and implications for in-vivoexperiments. J Pharm Pharmacol 60:63–70.
Mitsumoto Y, Burdett E, Grant A, and Klip A (1991) Differential expression of theGLUT1 and GLUT4 glucose transporters during differentiation of L6 muscle cells.Biochem Biophys Res Commun 175:652–659.
Moriya R, Shirakura T, Ito J, Mashiko S, and Seo T (2009) Activation of sodium-glucose cotransporter 1 ameliorates hyperglycemia by mediating incretin secretionin mice. Am J Physiol Endocrinol Metab 297:E1358–E1365.
Nathan DM (1993) Long-term complications of diabetes mellitus. N Engl J Med 328:1676–1685.
Nauck MA, Holst JJ, Willms B, and Schmiegel W (1997) Glucagon-like peptide 1(GLP-1) as a new therapeutic approach for type 2-diabetes. Exp Clin EndocrinolDiabetes 105:187–195.
Nomura S, Sakamaki S, Hongu M, Kawanishi E, Koga Y, Sakamoto T, Yamamoto Y,Ueta K, Kimata H, Nakayama K, et al. (2010) Discovery of canagliflozin, a novelC-glucoside with thiophene ring, as sodium-dependent glucose cotransporter 2inhibitor for the treatment of type 2 diabetes mellitus. J Med Chem 53:6355–6360.
Nomura S, Yamamoto Y, Matsumura Y, Ohba K, Sakamaki S, Kimata H, NakayamaK, Kuriyama C, Matsushita Y, Ueta K, et al. (2014) Novel indole-N-glucoside, TA-1887 as a sodium glucose cotransporter 2 inhibitor for treatment of type 2 diabetes.ACS Med Chem Lett 5:51–55.
Oguma T, Kuriyama C, Nakayama K, Matsushita Y, Yoshida K, Kiuchi S, Ikenaga Y,Nakamaru Y, Hikida K, Saito A, et al. (2015) The effect of combined treatment withcanagliflozin and teneligliptin on glucose intolerance in Zucker diabetic fatty rats.J Pharmacol Sci 127:456–461.
Oku A, Ueta K, Arakawa K, Ishihara T, Nawano M, Kuronuma Y, Matsumoto M,Saito A, Tsujihara K, Anai M, et al. (1999) T-1095, an inhibitor of renalNa1-glucose cotransporters, may provide a novel approach to treating diabetes.Diabetes 48:1794–1800.
Oku A, Ueta K, Nawano M, Arakawa K, Kano-Ishihara T, Matsumoto M, Saito A,Tsujihara K, Anai M, and Asano T (2000) Antidiabetic effect of T-1095, an inhibitorof Na(1)-glucose cotransporter, in neonatally streptozotocin-treated rats. Eur JPharmacol 391:183–192.
Orskov C, Wettergren A, and Holst JJ (1993) Biological effects and metabolic rates ofglucagonlike peptide-1 7-36 amide and glucagonlike peptide-1 7-37 in healthysubjects are indistinguishable. Diabetes 42:658–661.
Polidori D, Sha S, Mudaliar S, Ciaraldi TP, Ghosh A, Vaccaro N, Farrell K,Rothenberg P, and Henry RR (2013) Canagliflozin lowers postprandial glucoseand insulin by delaying intestinal glucose absorption in addition to increasingurinary glucose excretion: results of a randomized, placebo-controlled study.Diabetes Care 36:2154–2161.
Powell DR, Smith M, Greer J, Harris A, Zhao S, DaCosta C, Mseeh F, Shadoan MK,Sands A, Zambrowicz B, et al. (2013) LX4211 increases serum glucagon-like pep-tide 1 and peptide YY levels by reducing sodium/glucose cotransporter 1 (SGLT1)-mediated absorption of intestinal glucose. J Pharmacol Exp Ther 345:250–259.
Qaseem A, Humphrey LL, Sweet DE, Starkey M, and Shekelle P; Clinical GuidelinesCommittee of the American College of Physicians (2012) Oral pharmacologictreatment of type 2 diabetes mellitus: a clinical practice guideline from theAmerican College of Physicians. Ann Intern Med 156:218–231.
Rossetti L, Giaccari A, and DeFronzo RA (1990) Glucose toxicity. Diabetes Care 13:610–630.
Scott LJ and Spencer CM (2000) Miglitol: a review of its therapeutic potential in type2 diabetes mellitus. Drugs 59:521–549.
Shibazaki T, Tomae M, Ishikawa-Takemura Y, Fushimi N, Itoh F, Yamada M,and Isaji M (2012) KGA-2727, a novel selective inhibitor of a high-affinity sodiumglucose cotransporter (SGLT1), exhibits antidiabetic efficacy in rodent models. JPharmacol Exp Ther 342:288–296.
Stórustovu SI and Ebert B (2006) Pharmacological characterization of agonists at delta-containing GABAA receptors: Functional selectivity for extrasynaptic receptors isdependent on the absence of gamma2. J Pharmacol Exp Ther 316:1351–1359.
Szöcs Z, Brunmair B, Stadlbauer K, Nowotny P, Bauer L, Luger A, and Fürnsinn C(2008) Age-dependent development of metabolic derangement and effects of in-tervention with pioglitazone in Zucker diabetic fatty rats. J Pharmacol Exp Ther326:323–329.
Takata K, Kasahara T, Kasahara M, Ezaki O, and Hirano H (1992) Immunohisto-chemical localization of Na(1)-dependent glucose transporter in rat jejunum. CellTissue Res 267:3–9.
Ueta K, Ishihara T, Matsumoto Y, Oku A, Nawano M, Fujita T, Saito A, and ArakawaK (2005) Long-term treatment with the Na1-glucose cotransporter inhibitorT-1095 causes sustained improvement in hyperglycemia and prevents diabeticneuropathy in Goto-Kakizaki rats. Life Sci 76:2655–2668.
Wang B, Yu W, Guo J, Jiang X, Lu W, Liu M, and Pang X (2015) The antiparasiticdrug, potassium antimony tartrate, inhibits tumor angiogenesis and tumor growthin nonsmall-cell lung cancer. J Pharmacol Exp Ther 352:129–138.
Watanabe Y, Nakayama K, Taniuchi N, Horai Y, Kuriyama C, Ueta K, Arakawa K,Senbonmatsu T, and Shiotani M (2015) Beneficial effects of canagliflozin in com-bination with pioglitazone on insulin sensitivity in rodent models of obese type 2diabetes. PLoS One 10:e0116851.
You G, Lee WS, Barros EJ, Kanai Y, Huo TL, Khawaja S, Wells RG, Nigam SK,and Hediger MA (1995) Molecular characteristics of Na(1)-coupled glucose trans-porters in adult and embryonic rat kidney. J Biol Chem 270:29365–29371.
Address correspondence to: Takahiro Oguma, 2-2-50 Kawagishi, Toda-shi,Saitama, 335-8505, Japan. E-mail: [email protected]
Intestinal SGLT1 Inhibition Enhances GLP-1 Secretion In Vivo 289
at ASPE
T Journals on A
pril 14, 2020jpet.aspetjournals.org
Dow
nloaded from


![INDEX [jpet.aspetjournals.org]jpet.aspetjournals.org/content/jpet/234/3/local/back...effect, 708 Blockade, reticuloendothelial, enzyme-al-bumin conjugates, chronic adininis-tration](https://static.fdocuments.in/doc/165x107/60757ab7f966210d5e51d2f2/index-jpet-jpet-effect-708-blockade-reticuloendothelial-enzyme-al-bumin.jpg)



![A Cation-Chloride Cotransporter Gene Is … Cation-Chloride Cotransporter Gene Is Required for Cell Elongation and Osmoregulation in Rice1[OPEN] Zhi Chang Chen, Naoki Yamaji, Miho](https://static.fdocuments.in/doc/165x107/5d0f99e188c99345078b4fbd/a-cation-chloride-cotransporter-gene-is-cation-chloride-cotransporter-gene-is-required.jpg)

![RECEIVED EEilERAL ELECTIOH CGMi'^ISSION William B ...eqs.fec.gov/eqsdocsMUR/15044380209.pdf · Growth and Prosperity [sic] - a S01c4 founded by GOP operative Scott Reed - has posted](https://static.fdocuments.in/doc/165x107/5f6fba492be49e29ba780d08/received-eeileral-electioh-cgmiission-william-b-eqsfecgoveqsdocsmur-.jpg)



![INDEX [jpet.aspetjournals.org]jpet.aspetjournals.org/content/jpet/215/3/local/back-matter.pdf · synaptic neuromuscular ef-fects(rats),53 Caldwell, R.W.,seeCook, L.S., 198 Calcium](https://static.fdocuments.in/doc/165x107/6048ac2eb6e98b514e6c33d6/index-jpet-jpet-synaptic-neuromuscular-ef-fectsrats53-caldwell-rwseecook.jpg)


![INDEX [jpet.aspetjournals.org]jpet.aspetjournals.org/content/jpet/220/3/local/back...703 INDEX Volume 220, January-March, 1982 Abernethy, DR.,seeGreenblatt, D.J.,120 Acetaminophen-induced](https://static.fdocuments.in/doc/165x107/5d3598fd88c993f1228bd8ae/index-jpet-jpet-index-volume-220-january-march-1982-abernethy-drseegreenblatt.jpg)


![New INDEX [jpet.aspetjournals.org]jpet.aspetjournals.org/content/jpet/187/3/local/back... · 2005. 12. 3. · hycanthone effects onspermatogonial cells, de-oxyribonucleic acidsynthesis](https://static.fdocuments.in/doc/165x107/6067c6518625ed3f66076f25/new-index-jpet-jpet-2005-12-3-hycanthone-effects-onspermatogonial-cells.jpg)